
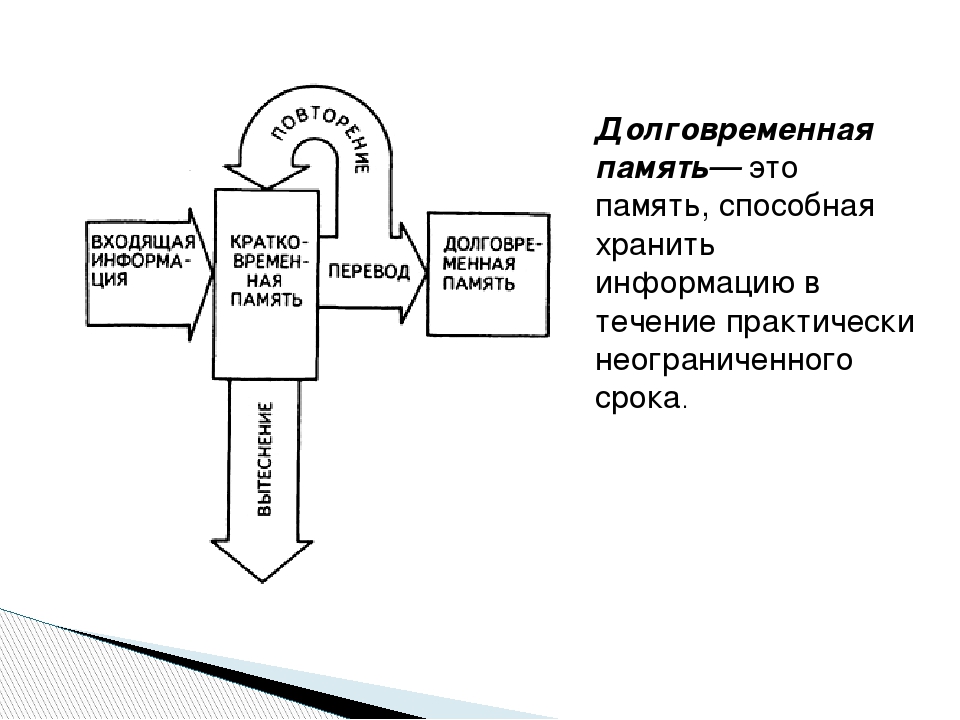
Долговременная память: Кратковременная и долгосрочная память
Военные научились стимулировать долговременную память
Гиппокамп
Иллюстрация: lifesciencedb.jp
Исследователи, работающие по заказу Агентства перспективных оборонных разработок (DARPA) министерства обороны США, на конференции Общества нейробиологов в Чикаго представили доказательства принципиальной возможности создания имплантатов, которые могут стимулировать долговременную память. Об этом пишет Nature. Конференция состоялась 17-21 октября 2015 года. Исследователи заявили, что они научились имитировать сигналы, переводящие кратовременную память в долговременную.
Считается, что за консолидацию памяти, то есть перевод кратковременной памяти в долговременную, отвечает гиппокамп, парная структура головного мозга, расположенная в медиальных височных отделах полушарий. Она же отвечает и за формирование кратковременной памяти на основе сенсорной информации, а также восприятия человеком пространства и времени. Исследователи выяснили, что при консолидации памяти возникает сигнал, проходящий от участка CA3 гиппокампа в CA1.
По мнению исследователей, точная имитация этого сигнала позволит стимулировать долговременную память у людей, включая перенесших травму головы военных и пожилых. Ученые проводили исследования на людях, страдающих эпилепсией с установленными мозговыми электродами для записи электроэнцефалограммы мозга и стимуляции отдельных его зон. В эксперименте участвовали 12 человек. Сперва ученые показывали участникам эксперимента различные изображения, а спустя полторы минуты просили вспомнить, какие из них они видели.
Во время показа изображений ученые вели запись сигналов, проходящих между зонами CA3 и CA1. Затем, на основе полученных данных, они разработали алгоритм, способный предугадывать вид сигнала, который должен прийти из CA3, по активности зоны CA1. В 80 процентов случаев реальный сигнал зоны CA3 был предсказан точно. Ученые полагают, что благодаря этому алгоритму они смогут стимулировать CA1, а значит и консолидировать память, даже если клетки зоны CA3 сильно повреждены.
Затем, на основе полученных данных, они разработали алгоритм, способный предугадывать вид сигнала, который должен прийти из CA3, по активности зоны CA1. В 80 процентов случаев реальный сигнал зоны CA3 был предсказан точно. Ученые полагают, что благодаря этому алгоритму они смогут стимулировать CA1, а значит и консолидировать память, даже если клетки зоны CA3 сильно повреждены.
В пользу этого предположения говорят эксперименты, проводившиеся на обезьянах в начале 2010-х годов. Обезьян учили запоминать различные картинки, а за правильное запоминание их поощряли соком. У некоторых животных во время эксперимента стимулировалась зона CA1 гиппокампа. По итогам эксперимента оказалось, что обезьяны, которым стимулировали CA1, быстрее запоминали изображения, чем животные, не подвергавшиеся стимуляции.
В рамках нового исследования уже был проведен один эксперимент по стимуляции зоны CA1 гиппокампа у женщины, страдающей эпилепсией. Впрочем, по словам ученых, говорить о результатах пока рано. Эксперименты планируется распространить и на других добровольцев с мозговыми электродами. По итогам этих исследований может быть разработан первый образец прибора, стимулирующего зоны гиппокампа, ответственные за консолидацию памяти.
Эксперименты планируется распространить и на других добровольцев с мозговыми электродами. По итогам этих исследований может быть разработан первый образец прибора, стимулирующего зоны гиппокампа, ответственные за консолидацию памяти.
Между тем, ученые полагают, что обнаружение и имитация сигнала, проходящего между CA3 и CA1, может оказаться недостаточным для стимуляции долговременной памяти. Например, в случае повреждения клеток зоны CA1 одной лишь имитации сигнала может оказаться недостаточно. Такими исследованиями по заказу DARPA занимаются ученые Университета Южной Калифорнии в Лос-Анджелесе. Работы ведутся по проекту создания приборов, которые смогут стимулировать память солдат с повреждениями головы.
По заказу DARPA также работает команда исследователей из Университета Пенсильвании. Эти исследователи пришли к выводу, что стимуляция медиальных височных долей мозга может улучшать память. Во время экспериментов на людях с эпилепсией выяснилось, что при ухудшении памяти, стимуляция приводит к улучшению, однако у людей с хорошей памятью при стимуляции наблюдается обратный эффект. В их эксперименте участвовали 28 человек с мозговыми электродами.
В их эксперименте участвовали 28 человек с мозговыми электродами.
Ученые давали участникам эксперимента ознакомиться со списком слов, а затем попытаться вспомнить их в заданном порядке. При этом исследователи вели запись мозговой активности. На основе записи они разработали алгоритм, который при повторении эксперимента смог с высокой точностью предсказывать вероятность вспоминания того или иного слова. При этом, во время чтения людьми слов, для которых была высока вероятность забывания, исследователи проводили стимуляцию; вероятность запоминания возрастала на 140 процентов.
Программа DARPA по созданию имплантатов, стимулирующих долговременную память, стартовала в феврале 2014 года. В ней участвуют две группы исследователей. Университет Южной Калифорнии получил на проведение работ 15 миллионов долларов, а Университет Пенсильвании — 22,5 миллиона долларов.
Василий Сычёв
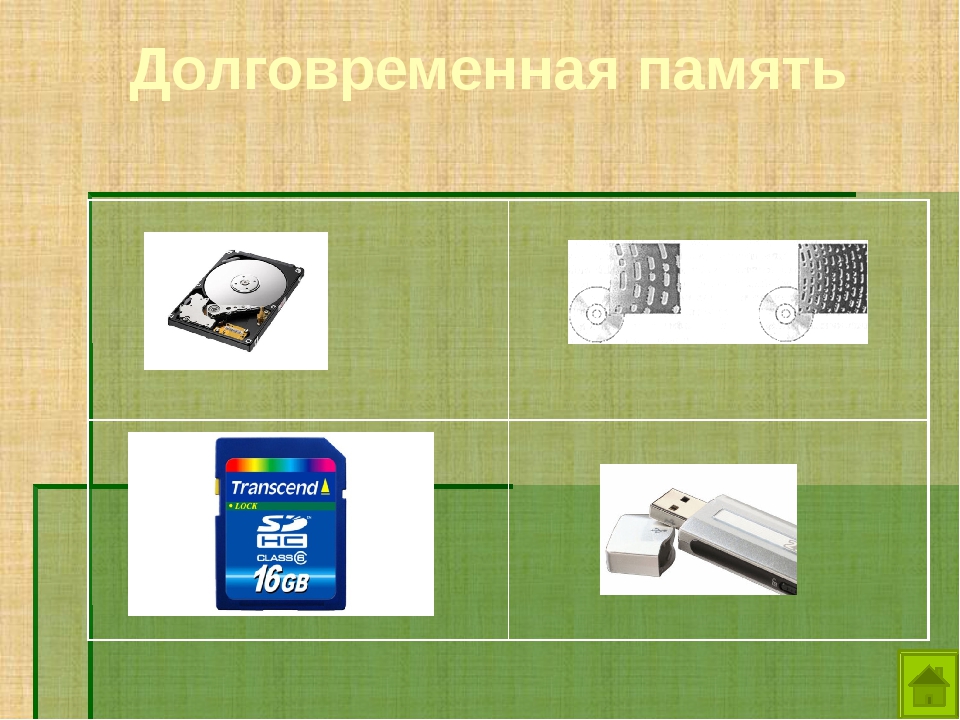
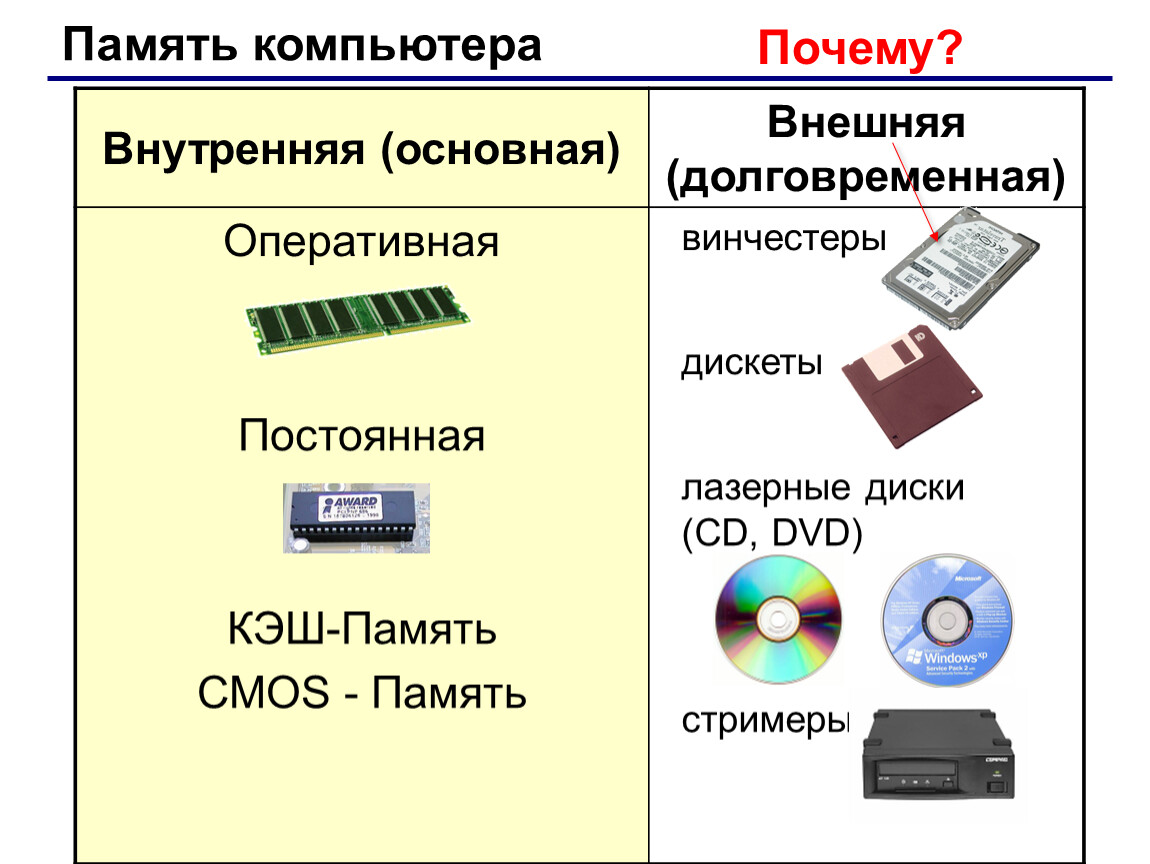
Внешняя (долговременная) память — урок.
 Информатика, 7 класс.
Информатика, 7 класс.Основной функцией внешней памяти компьютера является способность долговременно хранить большой объём информации (программы, документы, аудио- и видеоклипы и пр.).
Устройство, которое обеспечивает запись/считывание информации, называется накопителем, или дисководом, а хранится информация на носителях (например дискетах, дисках).
Магнитный принцип записи и считывания информации
В основу записи информации на жёсткий диск положено намагничивание ферромагнетиков в магнитном поле, хранение информации основывается на сохранении намагниченности, а считывание информации базируется на явлении электромагнитной индукции.
В процессе записи информации на гибкие и жёсткие магнитные диски головка дисковода с сердечником из магнитомягкого материала (малая остаточная намагниченность) перемещается вдоль магнитного слоя магнитожёсткого носителя (большая остаточная намагниченность).
На магнитную головку поступают последовательности электрических импульсов (последовательности логических единиц и нулей), которые создают в головке магнитное поле.
В результате последовательно намагничиваются (логическая единица) или не намагничиваются (логический нуль) элементы поверхности носителя.
При считывании информации посредством движения магнитной головки над поверхностью носителя намагниченные участки носителя вызывают в ней импульсы тока (явление электромагнитной индукции).
Последовательности таких импульсов передаются по магистрали в оперативную память компьютера.
Обрати внимание!
При отсутствии сильных магнитных полей и высоких температур элементы носителя могут сохранять свою намагниченность в течение долгого времени (лет, десятилетий).
Гибкие магнитные диски
Гибкие магнитные диски помещаются в пластмассовый корпус. Такой носитель информации называется дискетой.
В центре дискеты имеется приспособление для захвата и обеспечения вращения диска внутри пластмассового корпуса. Дискета вставляется в дисковод, который вращает диск с постоянной угловой скоростью.
При этом магнитная головка дисковода устанавливается на определённую концентрическую дорожку диска, на которую и производится запись или с которой производится считывание информации.
Обрати внимание!
Информационная ёмкость дискеты невелика и составляет всего \(1,44\) Мбайт. Скорость записи и считывания информации также мала (составляет всего около \(50\) Кбайт/с) из-за медленного вращения диска (\(360\) об/мин).
В целях сохранения информации гибкие магнитные диски необходимо предохранять от воздействия сильных магнитных полей и нагревания, так как такие физические воздействия могут привести к размагничиванию носителя и потере информации.Жёсткие магнитные диски
Жёсткий магнитный диск представляет собой несколько десятков дисков, размещённых на одной оси, заключённых в металлический корпус и вращающихся с большой угловой скоростью.
За счёт гораздо большего количества дорожек на каждой из сторон дисков и большого количества дисков информационная ёмкость жёсткого диска может в сотни тысяч раз превышать информационную ёмкость дискеты и достигать \(500\) Гбайт. Скорость записи и считывания информации с жёстких дисков достаточно велика (может достигать \(300\) Мбайт/с) за счёт быстрого вращения дисков (до \(7200\) об/мин).
Скорость записи и считывания информации с жёстких дисков достаточно велика (может достигать \(300\) Мбайт/с) за счёт быстрого вращения дисков (до \(7200\) об/мин).
В жёстких дисках используются достаточно хрупкие и миниатюрные элементы (пластины носителей, магнитные головки и пр.), поэтому в целях сохранения информации и работоспособности жёсткие диски необходимо оберегать от ударов и резких изменений пространственной ориентации в процессе работы.
Источники:
https://pixabay.com/images/id-160525/
https://pixabay.com/images/id-154463/
Что такое долговременная память, и как ее улучшить? — Блог Викиум
Под определением «долгосрочная память» в психологии подразумевается хранение и воспроизведение нужной информации на протяжении длительного времени. В биологии такая память рассматривается, как функционирование нейронов, благодаря которому информация может храниться продолжительное время. С помощью гиппокампа происходит процесс, когда кратковременная память переходит в долгосрочную.
Разновидности долговременной памяти
Психологи выделяют 2 вида долгосрочной памяти — имплицитная и эксплицитная. Первая отвечает за неосознанное запоминание, вторая — формируется из знаний, полученных человеком ранее.
Развитие долговременной памяти
Нельзя молниеносно натренировать память, так как эта функция обусловлена генетическими моментами. Очень сильно влияет на память образ жизни человека. Однако с помощью специальных упражнений можно более рационально использовать функцию запоминания:
- Используйте ассоциации к любой информации, чтобы она лучше откладывалась у вас в голове. Лучше придумать сразу несколько ассоциаций. Можно также рисовать у себя в голове картинки.
- При запоминании необходимо обязательно концентрироваться на информации, в противном случае она не попадает в мозг, и запоминание становится невозможным.

- Основой запоминания является умение большие блоки информации делить на маленькие части. Так у вас будет автоматически возникать ассоциация.
- Чтобы память не подводила, необходимо хорошо и полноценно отдыхать. Спать взрослому человеку рекомендуется не менее 6-8 часов в день.
- Даже если вы изучили определенную информацию, со временем ее рекомендуется повторить. Таким образом активный метод запоминания поможет вам надолго удержать нужную информацию в голове.
Наилучшим образом новые знания усваиваются, когда изученное применяют на практике. Одним из наиболее популярных способов является представление действия в голове. Подобное упражнение следует применять по отношению к каждому признаку.
Еще одним популярным методом по развитию долговременной памяти является метод Цицерона. В его основе лежит визуальная ассоциация. Так, например, человек может представлять на улице или в квартире определенный предмет. Если вам необходимо купить муку и хлеб, то вы можете представить, как эти продукты лежат на подоконнике или крыше дома.
Если вам необходимо купить муку и хлеб, то вы можете представить, как эти продукты лежат на подоконнике или крыше дома.
Развивать долговременную память с помощью упражнений можно и у ребенка. Например, можно предложить ему из нескольких слов составить небольшую историю. Или же можно предоставить чаду набор слов, которые он должен повторить через час. Даже если вспомнить не получается, не стоит относиться к упражнению с пренебрежением, а рекомендуется немного поразмыслить.
В качестве примеров долговременной памяти можно привести умение человека водить автомобиль, открывать дверь или пользоваться плитой на кухне. Чтобы улучшить свою память, необходимо регулярно упражняться. Например, для этого есть когнитивные тренажеры Викиум.
Тест «Долговременная память»
Цель теста «Долговременная память». Определение объема кратковременной и долговременной памяти.
Контрольные задачи. Определить объем кратковременной и долговременной памяти у каждого студента учебной группы путем определения числа запомнившихся слов после каждого из пяти зачитываний исследователем 20 текстовых слов.
МЕТОДИКА 1. Заучивание 20 текстовых слов
Ход работы: Экспериментатор зачитывает 20 слов. Не должно быть слов, которые обозначают предметы, окружающие испытуемого в данный момент. Испытуемый воспроизводит письменно слова после каждого зачитывания в любом порядке (закрывая результаты предыдущего воспроизведения).
Оценка кратковременной памяти. Для оценки памяти рекомендуется построить график. На оси ординат откладывается число запомнившихся слов, по оси абсцисс – число повторений. Если к 5-му зачитыванию испытуемый запомнил 20 слов – удовлетворительно, если к 3-му – хорошо. Если число слов нарастает и к третьему зачитыванию устанавливается максимум, то это означает, что психической истощаемости не отмечается. Если испытуемый воспроизводит мало слов и после 2 – 4 повторений количество их уменьшается, то это свидетельствует об истощаемости. Вплетение лишних слов расценивается как один из признаков ослабления психической деятельности.
МЕТОДИКА 2. Производится запоминание и воспроизведение текста со смысловой связью.
Проверка долговременной памяти.
В протоколе и на классной доске в виде таблицы записывают результаты исследования у различных испытуемых и делают вывод об индивидуальных различиях памяти и значении ассоциативной (смысловой) памяти у человека.
Анализ результатов проведенной работы. Результаты опыта запишите в виде протокола. Сформулируйте выводы из приведенных лабораторных исследований, сделайте резюме.
ЭКСПЕРИМЕНТАЛЬНЫЙ МАТЕРИАЛ К МЕТОДИКЕ 1. 20 предлагаемых слов и их порядковые номера.
Закройте текст, на листике напишите слова с их порядковыми номерами.
|
|
Продуктивность запоминания вы сможете вычислить по формуле:
количество правильно воспроизведенных слов |
х 100 % |
|
количество предлагаемых слов |
ЭКСПЕРИМЕНТАЛЬНЫЙ МАТЕРИАЛ К МЕТОДИКЕ 2. В течение 60 секунд прочитайте тест. В нем выделены жирным шрифтом и пронумерованы 10 главных мыслей. Попытайтесь воспроизвести их, сохраняя указанную последовательность.
В течение 60 секунд прочитайте тест. В нем выделены жирным шрифтом и пронумерованы 10 главных мыслей. Попытайтесь воспроизвести их, сохраняя указанную последовательность.
В 1912 году в Атлантическом океане произошла катастрофа. Огромный пассажирский пароход «Титаник», шедший первым рейсом из Европы в Америку, столкнулся в тумане с плавающей ледяной горой – айсбергом. 1) Получил пробоину и стал тонуть. 2) «Спустить шлюпки!» — скомандовал капитан. Но шлюпок оказалось недостаточно. 3) Их хватило только на половину пассажиров. Женщины и дети – к сходням, мужчинам надеть спасательные пояса», — раздалась вторая команда. 4) Мужчины молча отошли от борта. Пароход медленно погружался в темную холодную воду. 5) Вот началась посадка в последнюю шлюпку. 6). И вдруг к сходням крича бросился какой-то толстяк с перекошенным от страха лицом. 7) Расталкивая женщин и детей, он пытался вскочить в шлюпку. 8)Послышался щелчок – это капитан выстрелил из пистолета. 9) Трус упал на палубу мертвый. 10) Но никто не оглянулся в его сторону.
7) Расталкивая женщин и детей, он пытался вскочить в шлюпку. 8)Послышался щелчок – это капитан выстрелил из пистолета. 9) Трус упал на палубу мертвый. 10) Но никто не оглянулся в его сторону.
Формула:
количество правильно воспроизведенных мыслей |
х 100 % |
|
количество выделений в тексте |
Сложив цифры, полученные по двум методикам, а затем разделив сумму на два, вы узнаете среднюю продуктивность запоминания. 90–100 –отличный результат; 70-90 – очень хороший результат; 50-69 – хороший; 30-49 – удовлетворительный; 10-29 – плохой; 0-9 – очень плохой.
Роль микроРНК в обучении и долговременной памяти | Гринкевич
1. Ai J., Sun L.H., Che H., Zhang R., Zhang T.Z., Wu W.C., Su X.L., Chen X., Yang G., Li K., Wang N., Ban T., Bao Y.N., Guo F., Niu H.F., Zhu Y.L., Zhu X.Y., Zhao S.G., Yang B.F. MicroRNA195 protects against dementia induced by chronic brain hypoperfusion via its anti-amyloidogenic effect in rats. J. Neurosci. 2013;33(9):3989-4001. DOI 10.1523/JNEUROSCI.1997-12.2013. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6619292.
Ai J., Sun L.H., Che H., Zhang R., Zhang T.Z., Wu W.C., Su X.L., Chen X., Yang G., Li K., Wang N., Ban T., Bao Y.N., Guo F., Niu H.F., Zhu Y.L., Zhu X.Y., Zhao S.G., Yang B.F. MicroRNA195 protects against dementia induced by chronic brain hypoperfusion via its anti-amyloidogenic effect in rats. J. Neurosci. 2013;33(9):3989-4001. DOI 10.1523/JNEUROSCI.1997-12.2013. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6619292.
2. Aksoy-Aksel A., Zampa F., Schratt G. MicroRNAs and synaptic plasticity – a mutual relationship. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2014;369(1652):20130515. DOI 10.1098/rstb.2013.0515. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4142036.
3. Aquino-Jarquin G. Emerging role of CRISPR/Cas9 technology for microRNAs editing in cancer research. Cancer Res. 2017;77(24): 6812-6817. DOI 10.1158/0008-5472.CAN-17-2142. https://cancerres.aacrjournals.org/content/77/24/6812.long.
https://cancerres.aacrjournals.org/content/77/24/6812.long.
4. Aten S., Hansen K.F., Snider K., Wheaton K., Kalidindi A., Garcia A., Alzate-Correa D., Hoyt K.R., Obrietan K. miR-132 couples the circadian clock to daily rhythms of neuronal plasticity and cognition. Learn. Mem. 2018;25(5):214-229. DOI 10.1101/lm.047191.117. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5903403.
5. Baby N., Alagappan N., Dheen S.T., Sajikumar S. MicroRNA-134-5p inhibition rescues long-term plasticity and synaptic tagging/capture in an Aβ(1-42)-induced model of Alzheimer’s disease. Aging Cell. 2020;19(1):e13046. DOI 10.1111/acel.13046. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6974725.
6. Baek S., Hwan C., Kim J. Ebf3-miR218 regulation is involved in the development of dopaminergic neurons. Brain Res. 2014;1587: 23-32. DOI 10.1016/j.brainres.2014.08.059. https://pubmed.ncbi.nlm.nih.gov/25192643.
Brain Res. 2014;1587: 23-32. DOI 10.1016/j.brainres.2014.08.059. https://pubmed.ncbi.nlm.nih.gov/25192643.
7. Banks S.A., Pierce M.L., Soukup G.A. Sensational microRNAs: neurosensory roles of the microRNA-183 family. Mol. Neurobiol. 2020;57(1):358-371. DOI 10.1007/s12035-019-01717-3. https://link.springer.com/article/10.1007%2Fs12035-019-01717-3.
8. Bartel D.P. MicroRNAs: target recognition and regulatory functions. Cell. 2009;136:215-233. DOI 10.1016/j.cell.2009.01.002. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3794896.
9. Benito E., Kerimoglu C., Ramachandran B., Pena-Centeno T., Jain G., Stilling R.M., Islam M.R., Capece V., Zhou Q., Edbauer D., Dean C., Fischer A. RNA-dependent intergenerational inheritance of enhanced synaptic plasticity after environmental enrichment. Cell Rep. 2018;23(2):546-554. DOI 10.1016/j.celrep.2018.03.059. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5912949.
Cell Rep. 2018;23(2):546-554. DOI 10.1016/j.celrep.2018.03.059. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5912949.
10. Berger S.L. The complex language of chromatin regulation during transcription. Nature. 2007;447(7143):407-412. DOI 10.1038/nature 05915. https://pubmed.ncbi.nlm.nih.gov/17522673.
11. Beveridge N.J., Gardiner E., Carroll A.P., Tooney P.A., Cairns M.J. Schizophrenia is associated with an increase in cortical microRNA biogenesis. Mol. Psychiatry. 2010;15(12):1176-1189. DOI 10.1038/mp.2009.84. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2990188.
12. Bicker S., Khudayberdiev S., Weiss K., Zocher K., Baumeister S., Schratt G. The DEAH-box helicase DHX36 mediates dendritic localization of the neuronal precursor-microRNA-134. Genes Dev. 2013;27(9):991-996. DOI 10.1101/gad.211243.112. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3656329.
DOI 10.1101/gad.211243.112. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3656329.
13. Bitetti A., Mallory A.C., Golini E., Carrieri C., Carreño Gutiérrez H., Perlas E., Pérez-Rico Y.A., Tocchini-Valentini G.P., Enright A.J., Norton W.H.J., Mandillo S., O’Carroll D., Shkumatava A. MicroRNA degradation by a conserved target RNA regulates animal behavior. Nat. Struct. Mol. Biol. 2018;25(3):244-251. DOI 10.1038/s41594-018-0032-x. https://pubmed.ncbi.nlm.nih.gov/29483647.
14. Cao T., Zhen X.C. Dysregulation of miRNA and its potential therapeutic application in schizophrenia. CNS Neurosci. Ther. 2018; 24(7):586-597. DOI 10.1111/cns.12840.
15. Chen W., Qin C. General hallmarks of microRNAs in brain evolution and development. RNA Biol. 2015;12(7):701-708. DOI 10. 1080/15476286.2015.1048954. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4615839.
1080/15476286.2015.1048954. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4615839.
16. Cheng Y., Wang Z.M., Tan W., Wang X., Li Y., Bai B., Li Y., Zhang S.F., Yan H.L., Chen Z.L., Liu C.M., Mi T.W., Xia S., Zhou Z., Liu A., Tang G.B., Liu C., Dai Z.J., Wang Y.Y., Wang H., Wang X., Kang Y., Lin L., Chen Z., Xie N., Sun Q., Xie W., Peng J., Chen D., Teng Z.Q., Jin P. Partial loss of psychiatric risk gene miR137 in mice causes repetitive behavior and impairs sociability and learning via increased Pde10a. Nat. Neurosci. 2018;21(12):1689-1703. DOI 10.1038/s41593-018-0261-7. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6261680.
17. Chmielarz P., Konovalova J., Najam S.S., Alter H., Piepponen T.P., Erfle H., Sonntag K.C., Schütz G., Vinnikov I.A., Domanskyi A. Dicer and microRNAs protect adult dopamine neurons. Cell Death Dis. 2017;8(5):e2813. DOI 10.1038/cddis.2017.214. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5520729.
DOI 10.1038/cddis.2017.214. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5520729.
18. Danka Mohammed C.P., Park J.S., Nam H.G., Kim K. MicroRNAs in brain aging. Mech. Ageing Dev. 2017;168:3-9. DOI 10.1016/j.mad.2017.01.007. https://pubmed.ncbi.nlm.nih.gov/28119001.
19. Dias B.G., Goodman J.V., Ahluwalia R., Easton A.E., Andero R., Ressler K.J. Amygdala-dependent fear memory consolidation via miR-34a and notch signaling. Neuron. 2014;83(4):906-918. DOI 10.1016/j.neuron.2014.07.019. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4172484.
20. Dimmeler S., Nicotera P. MicroRNAs in age-related diseases. EMBO Mol. Med. 2013;5(2):180-190. DOI 10.1002/emmm.201201986. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3569636.
21. Fiorenza A., Barco A. Role of Dicer and the miRNA system in neuronal plasticity and brain function. Neurobiol. Learn. Mem. 2016; 135:3-12. DOI 10.1016/j.nlm.2016.05.001. https://pubmed.ncbi.nlm.nih.gov/27163737.
Fiorenza A., Barco A. Role of Dicer and the miRNA system in neuronal plasticity and brain function. Neurobiol. Learn. Mem. 2016; 135:3-12. DOI 10.1016/j.nlm.2016.05.001. https://pubmed.ncbi.nlm.nih.gov/27163737.
22. Fiorenza A., Lopez-Atalaya J.P., Rovira V., Scandaglia M., GeijoBarrientos E., Barco A. Blocking miRNA biogenesis in adult forebrain neurons enhances seizure susceptibility, fear memory, and food intake by increasing neuronal responsiveness. Cereb. Cortex. 2016;26:1619-1633. DOI 10.1093/cercor/bhu332. https://pubmed.ncbi.nlm.nih.gov/25595182.
23. Fischer A. Epigenetic memory: the Lamarckian brain. EMBO J. 2014;33(9):945-967. DOI 10.1002/embj.201387637. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4193930.
24. Gaine M.E., Chatterjee S., Abel T. Sleep deprivation and the epigenome. Front. Neural Circuits. 2018;12:14. DOI 10.3389/fncir.2018.00014. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5835037.
Sleep deprivation and the epigenome. Front. Neural Circuits. 2018;12:14. DOI 10.3389/fncir.2018.00014. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5835037.
25. Gantier M.P., McCoy C.E., Rusinova I., Saulep D., Wang D., Xu D., Irving A.T., Behlke M.A., Hertzog P.J., Mackay F., Williams B.R. Analysis of microRNA turnover in mammalian cells following Dicer1 ablation. Nucleic Acids Res. 2011;39(13):5692-5703. DOI 10.1093/nar/gkr148. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3141258.
26. Gao J., Wang W.Y., Mao Y.W., Gräff J., Guan J.S., Pan L., Mak G., Kim D., Su S.C., Tsai L.H. Anovel pathway regulates memory and plasticity via SIRT1 and miR-134. Nature. 2010;466(7310):1105- 1109. DOI 10.1038/nature09271. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2928875.
27. Griggs E.M., Young E.J., Rumbaugh G., Miller C.A. MicroRNA-182 regulates amygdala-dependent memory formation. Version 2. J. Neurosci. 2013;33(4):1734-1740. DOI 10.1523/JNEUROSCI.2873-12.2013. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3711533.
Griggs E.M., Young E.J., Rumbaugh G., Miller C.A. MicroRNA-182 regulates amygdala-dependent memory formation. Version 2. J. Neurosci. 2013;33(4):1734-1740. DOI 10.1523/JNEUROSCI.2873-12.2013. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3711533.
28. Grinkevich L.N. Epigenetics and long-term memory formation. Rossiyskiy Fiziologicheskiy Zhurnal im. I.M. Sechenova = I.M. Sechenov Physiological Journal. 2012;98(5):553-574. https://pubmed.ncbi.nlm.nih.gov/22838191/ (in Russian)
29. Grinkevich L.N. Influence of PLL treatment on the long-term memory formation in Helix mollusk. Meditsynskiy Akademicheskiy Zhurnal = Medical Academic Journal. 2019;19(4):87-92. DOI 10.17816/MAJ19080. https://journals.eco-vector.com/MAJ/article/view/19080. (in Russian)
30. Gu Q. H., Yu D., Hu Z., Liu X., Yang Y., Luo Y., Zhu J., Li Z. miR-26a and miR-384-5p are required for LTP maintenance and spine enlargement. Nat. Commun. 2015;6:6789. DOI 10.1038/ncomms7789. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4403380.
H., Yu D., Hu Z., Liu X., Yang Y., Luo Y., Zhu J., Li Z. miR-26a and miR-384-5p are required for LTP maintenance and spine enlargement. Nat. Commun. 2015;6:6789. DOI 10.1038/ncomms7789. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4403380.
31. Gu X., Xu Y., Xue W.Z., Wu Y., Ye Z., Xiao G., Wang H.L. Interplay of miR-137 and EZh3 contributes to the genome-wide redistribution of h4K27me3 underlying the Pb-induced memory impairment. Cell Death Dis. 2019;10(9):671. DOI 10.1038/s41419-019-1912. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6739382.
32. Hansen K.F., Sakamoto K., Wayman G.A., Impey S., Obrietan K. Transgenic miR132 alters neuronal spine density and impairs novel object recognition memory. PLoS One. 2010;5(11):e15497. DOI 10.1371/journal.pone.0015497. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2993964.
33. Havekes R., Abel T. The tired hippocampus: the molecular impact of sleep deprivation on hippocampal function. Curr. Opin. Neu¬ robiol. 2017;44:13-19. DOI 10.1016/j.conb.2017.02.005. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5511071.
Havekes R., Abel T. The tired hippocampus: the molecular impact of sleep deprivation on hippocampal function. Curr. Opin. Neu¬ robiol. 2017;44:13-19. DOI 10.1016/j.conb.2017.02.005. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5511071.
34. He L., Hannon G.J. MicroRNAs: small RNAs with a big role in gene regulation. Nat. Rev. Genet. 2004;5(7):522-531. DOI 10.1038/nrg1379. https://pubmed.ncbi.nlm.nih.gov/15211354.
35. Hébert S.S., Papadopoulou A.S., Smith P., Galas M.C., Planel E., Silahtaroglu A.N., Sergeant N., Buée L., De Strooper B. Genetic ablation of Dicer in adult forebrain neurons results in abnormal tau hyperphosphorylation and neurodegeneration. Hum. Mol. Genet. 2010;19(20):3959-3969. DOI 10.1093/hmg/ddq311. https://pubmed.ncbi.nlm.nih.gov/20660113.
36. Hirosawa M., Fujita Y., Parr C.J.C., Hayashi K., Kashida S., Hotta A., Woltjen K., Saito H. Cell-type-specific genome editing with a microRNA-responsive CRISPR-Cas9 switch. Nucleic Acids Res. 2017;45(13):e118. DOI 10.1093/nar/gkx309. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5570128.
Hirosawa M., Fujita Y., Parr C.J.C., Hayashi K., Kashida S., Hotta A., Woltjen K., Saito H. Cell-type-specific genome editing with a microRNA-responsive CRISPR-Cas9 switch. Nucleic Acids Res. 2017;45(13):e118. DOI 10.1093/nar/gkx309. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5570128.
37. Hoffmann M.D., Aschenbrenner S., Grosse S., Rapti K., Domenger C., Fakhiri J., Mastel M., Börner K., Eils R., Grimm D., Niopek D. Cell-specific CRISPR-Cas9 activation by microRNA-dependent expression of anti-CRISPR protein. Nucleic Acids Res. 2019;47(13):e75. DOI 10.1093/nar/gkz271. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6648350.
38. Hu T., Zhou F.J., Chang Y.F., Li Y.S., Liu G.C., Hong Y., Chen H.L., Xiyang Y.B., Bao T.H. miR21 is associated with the cognitive improvement following voluntary running wheel exercise in TBI mice. J. Mol. Neurosci. 2015;57(1):114-122. DOI 10.1007/s12031-015-0584-8. https://pubmed.ncbi.nlm.nih.gov/26018937.
2015;57(1):114-122. DOI 10.1007/s12031-015-0584-8. https://pubmed.ncbi.nlm.nih.gov/26018937.
39. Hu Z., Li Z. miRNAs in synapse development and synaptic plasticity. Curr. Opin. Neurobiol. 2017;45:24-31. DOI 10.1016/j.conb.2017.02.014. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5554733.
40. Hu Z., Yu D., Gu Q.H., Yang Y., Tu K., Zhu J., Li Z. miR-191 and miR-135 are required for long-lasting spine remodelling associated with synaptic long-term depression. Nat. Commun. 2014;5: 3263. DOI 10.1038/ncomms4263. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3951436.
41. Hu Z., Zhao J., Hu T., Luo Y., Zhu J., Li Z. miR-501-3p mediates the activity-dependent regulation of the expression of AMPA receptor subunit GluA1. J. Cell Biol. 2015;208(7):949-959. DOI 10. 1083/jcb.201404092. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4384731.
1083/jcb.201404092. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4384731.
42. Inukai S., de Lencastre A., Turner M., Slack F. Novel microRNAs differentially expressed during aging in the mouse brain. PLoS One. 2012;7:e40028. DOI 10.1371/journal.pone.0040028. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3402511.
43. Jawaid A., Woldemichael B.T., Kremer E.A., Laferriere F., Gaur N., Afroz T., Polymenidou M., Mansuy I.M. Memory decline and its reversal in aging and neurodegeneration involve miR-183/96/182 biogenesis. Mol. Neurobiol. 2019;56(5):3451-3462. DOI 10.1007/s12035-018-1314-3. https://pubmed.ncbi.nlm.nih.gov/30128653.
44. Jessop P., Toledo-Rodriguez M. Hippocampal TET1 and TET2 expression and DNA hydroxymethylation are affected by physical exercise in aged mice. Front. Cell Dev. Biol. 2018;6:45. DOI 10.3389/fcell.2018.00045. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5922180.
Front. Cell Dev. Biol. 2018;6:45. DOI 10.3389/fcell.2018.00045. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5922180.
45. John B., Enright A.J., Aravin A., Tuschl T., Sander C., Marks D.S. Human microRNA targets. PLoS Biol. 2004;2(11):e363. DOI 10.1371/journal.pbio.0020363. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC521178.
46. Jovasevic V., Corcoran K.A., Leaderbrand K., Yamawaki N., Guedea A.L., Chen H.J., Shepherd G.M., Radulovic J. GABAergic mechanisms regulated by miR-33 encode state-dependent fear. Nat. Neurosci. 2015;18(9):1265-1271. DOI 10.1038/nn.4084. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4880671.
47. Kandel E. Small neuron systems. In: The Brain. Scientific American, 1979.
48. Kandel E. The molecular biology of memory: cAMP, PKA, CRE, CREB-1, CREB-2, and CPEB. Mol. Brain. 2012;5(14):1-12. DOI 10.1186/1756-6606-5-1426. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3514210.
Kandel E. The molecular biology of memory: cAMP, PKA, CRE, CREB-1, CREB-2, and CPEB. Mol. Brain. 2012;5(14):1-12. DOI 10.1186/1756-6606-5-1426. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3514210.
49. Karabulut S., Korkmaz Bayramov K., Bayramov R., Ozdemir F., Topaloglu T., Ergen E., Yazgan K., Taskiran A.S., Golgeli A. Effects of post-learning REM sleep deprivation on hippocampal plasticity-related genes and microRNA in mice. Behav. Brain Res. 2019;361:7-13. DOI 10.1016/j.bbr.2018.12.045. https://pubmed.ncbi.nlm.nih.gov/30594545.
50. Kim S., Kaang B.K. Epigenetic regulation and chromatin remodeling in learning and memory. Exp. Mol. Med. 2017;49(1):e281. DOI 10.1038/emm.2016.140. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5291841.
51. Konopka W., Kiryk A. , Novak M., Herwerth M., Parkitna J.R., Wawrzyniak M., Kowarsch A., Michaluk P., Dzwonek J., Arnsperger T., Wilczynski G., Merkenschlager M., Theis F.J., Köhr G., Kaczmarek L., Schütz G. MicroRNA loss enhances learning and memory in mice. J. Neurosci. 2010;30(44):14835-14842. DOI 10.1523/JNEUROSCI.3030-10.2010. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6633640.
, Novak M., Herwerth M., Parkitna J.R., Wawrzyniak M., Kowarsch A., Michaluk P., Dzwonek J., Arnsperger T., Wilczynski G., Merkenschlager M., Theis F.J., Köhr G., Kaczmarek L., Schütz G. MicroRNA loss enhances learning and memory in mice. J. Neurosci. 2010;30(44):14835-14842. DOI 10.1523/JNEUROSCI.3030-10.2010. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6633640.
52. Korneev S.A., Vavoulis D.V., Naskar S., Dyakonova V.E., Kemenes I., Kemenes G. A CREB2-targeting microRNA is required for long-term memory after single-trial learning. Sci. Rep. 2018; 8(1):3950. DOI 10.1038/s41598-018-22278-w. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5834643.
53. Lee R.C., Feinbaum R.L., Ambros V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell. 1993;75(5):843-854. DOI 10.1016/0092-8674(93)90529-y. https://pubmed.ncbi.nlm.nih.gov/8252621.
https://pubmed.ncbi.nlm.nih.gov/8252621.
54. Lee S.T., Chu K., Jung K.H., Kim J.H., Huh J.Y., Yoon H., Park D.K., Lim J.Y., Kim J.M., Jeon D., Ryu H., Lee S.K., Kim M., Roh J.K. miR-206 regulates brain-derived neurotrophic factor in Alzheimer disease model. Ann. Neurol. 2012;72:269-277. DOI 10.1002/ana.23588. https://pubmed.ncbi.nlm.nih.gov/22926857.
55. Lesseur C., Paquette A.G., Marsit C.J. Epigenetic regulation of infant neurobehavioral outcomes. Med. Epigenet. 2014;2(2):71-79. DOI 10.1159/000361026. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4116357.
56. Leung A.K.L. The whereabouts of microRNA actions: cytoplasm and beyond. Trends Cell Biol. 2015;25(10):601-610. DOI 10.1016/j.tcb.2015.07.005. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4610250.
57. Lewis B.P., Shih I.-H., Jones-Rhoades M.W., Bartel D.P., Burge C.B. Prediction of mammalian microRNA targets. Cell. 2003;115(7): 787-798. DOI 10.1016/s0092-8674(03)01018-3. https://pubmed.ncbi.nlm.nih.gov/14697198.
58. Lin Q., Ponnusamy R., Widagdo J., Choi J.A., Ge W., Probst C., Buckley T., Lou M., Bredy T.W., Fanselow M.S., Ye K., Sun Y.E. MicroRNA-mediated disruption of dendritogenesis during a critical period of development influences cognitive capacity later in life. Proc. Natl. Acad. Sci. USA. 2017;114(34):9188-9193. DOI 10.1073/pnas.1706069114. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5576812.
59. Liu E.Y., Cali C.P., Lee E.B. RNA metabolism in neurodegenerative disease. Dis. Model. Mech. 2017;10(5):509-518. DOI 10.1242/dmm.028613. https://www. ncbi.nlm.nih.gov/pmc/articles/PMC5451173.
ncbi.nlm.nih.gov/pmc/articles/PMC5451173.
60. Lugli G., Larson J., Martone M.E., Jones Y., Smalheiser N.R. Dicer and eIF2c are enriched at postsynaptic densities in adult mouse brain and are modified by neuronal activity in a calpain-dependent manner. J. Neurochem. 2005;94(4):896-905. DOI 10.1111/j.1471-4159.2005.03224.x. https://pubmed.ncbi.nlm.nih.gov/16092937.
61. Malmevik J., Petri R., Knauff P., Brattas P.L., Akerblom M., Jakobsson J. Distinct cognitive effects and underlying transcriptome changes upon inhibition of individual miRNAs in hippocampal neurons. Sci. Rep. 2016;6:19879. DOI 10.1038/srep19879. https:// www.ncbi.nlm.nih.gov/pmc/articles/PMC4728481.
62. Mathew R.S., Tatarakis A., Rudenko A., Johnson-Venkatesh E.M., Yang Y.J., Murphy E.A., Todd T.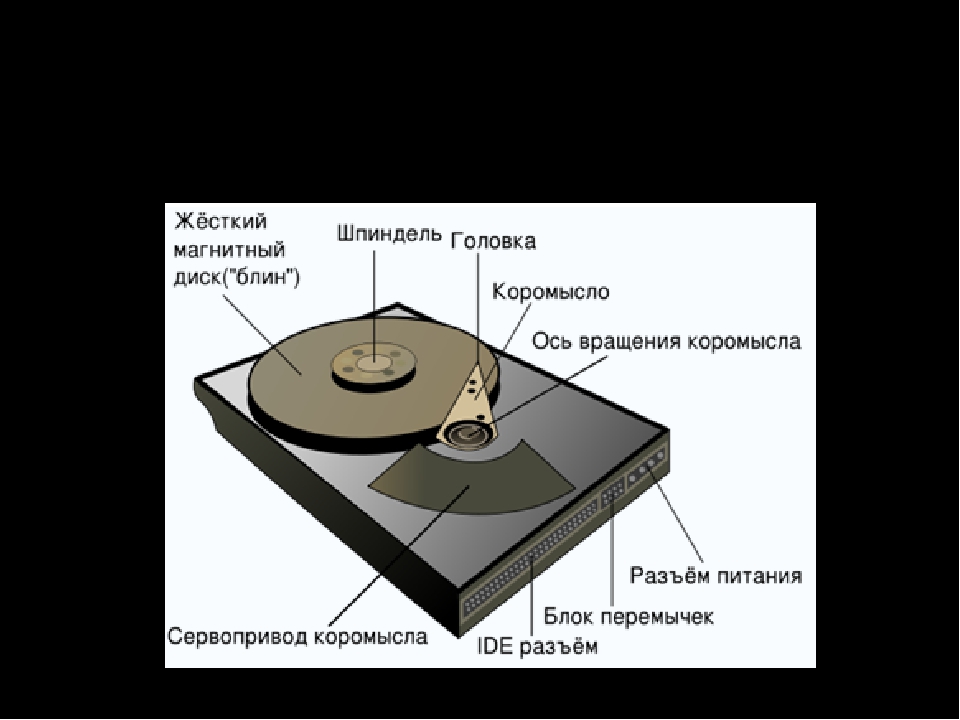 P., Schepers S.T., Siuti N., Martorell A.J., Falls W.A., Hammack S.E., Walsh C.A., Tsai L.H., Umemori H., Bouton M.E., Moazed D.A. microRNA negative feedback loop downregulates vesicle transport and inhibits fear memory. eLife. 2016;5:e22467. DOI 10.7554/eLife.22467. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5293492.
P., Schepers S.T., Siuti N., Martorell A.J., Falls W.A., Hammack S.E., Walsh C.A., Tsai L.H., Umemori H., Bouton M.E., Moazed D.A. microRNA negative feedback loop downregulates vesicle transport and inhibits fear memory. eLife. 2016;5:e22467. DOI 10.7554/eLife.22467. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5293492.
63. McNeill E., Van Vactor D. MicroRNAs shape the neuronal landscape. Neuron. 2012;75(3):363-379. DOI 10.1016/j.neuron.2012.07.005. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3441179.
64. Murphy C.P., Singewald N. Potential of microRNAs as novel targets in the alleviation of pathological fear. Genes Brain Behav. 2018; 17(3):e12427. DOI 10.1111/gbb.12427. https://onlinelibrary.wiley.com/doi/full/10.1111/gbb.12427.
65. Nilsson E.K., Boström A. E., Mwinyi J., Schiöth H.B. Epigenomics of total acute sleep deprivation in relation to genome-wide DNA methylation profiles and RNA expression. OMICS. 2016;20(6): 334-342. DOI 10.1089/omi.2016.0041. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4926204.
E., Mwinyi J., Schiöth H.B. Epigenomics of total acute sleep deprivation in relation to genome-wide DNA methylation profiles and RNA expression. OMICS. 2016;20(6): 334-342. DOI 10.1089/omi.2016.0041. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4926204.
66. Nudelman A.S., DiRocco D.P., Lambert T.J., Garelick M.G., Le J., Nathanson N.M., Storm D.R. Neuronal activity rapidly induces transcription of the CREB-regulated microRNA-132, in vivo. Hippocampus. 2010;20(4):492-498. DOI 10.1002/hipo.20646. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2847008.
67. Paul S., Reyes P.R., Garza B.S., Sharma A. MicroRNAs and child neuropsychiatric disorders: a brief review. Neurochem. Res. 2020;45(2):232-240. DOI 10.1007/s11064-019-02917-y. https://pubmed.ncbi.nlm.nih.gov/31773374.
68. Rajasethupathy P., Fiumara F., Sheridan R., Betel D., Puthanveettil S.V., Russo J.J., Sander C., Tuschl T., Kandel E. Characterization of small RNAs in Aplysia reveals a role for miR-124 in constraining synaptic plasticity through CREB. Neuron. 2009;63(6): 803-817. DOI 10.1016/j.neuron.2009.05.029. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2875683.
Rajasethupathy P., Fiumara F., Sheridan R., Betel D., Puthanveettil S.V., Russo J.J., Sander C., Tuschl T., Kandel E. Characterization of small RNAs in Aplysia reveals a role for miR-124 in constraining synaptic plasticity through CREB. Neuron. 2009;63(6): 803-817. DOI 10.1016/j.neuron.2009.05.029. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2875683.
69. Ramakrishna S., Muddashetty R.S. Emerging role of microRNAs in dementia. J. Mol. Biol. 2019;431(9):1743-1762. DOI 10.1016/j.jmb.2019.01.046. https://pubmed.ncbi.nlm.nih.gov/30738891.
70. Reinhart B.J., Slack F.J., Basson M., Pasquinelli A.E., Bettinger J.C., Rougvie A.E., Horvitz H.R., Ruvkun G. The 21-nucleotide let-7 RNA regulates developmental timing in Caenorhabditis elegans. Nature. 2000;403(6772):901-906. DOI 10.1038/35002607. https://pubmed.ncbi.nlm.nih.gov/10706289.
71. Saus E., Soria V., Escaramis G., Vivarelli F., Crespo J.M., Kagerbauer B., Menchón J.M., Urretavizcaya M., Gratacòs M., Estivill X. Genetic variants and abnormal processing of pre-miR182, a circadian clock modulator, in major depression patients with late insomnia. Hum. Mol. Genet. 2010;19(20):4017-4025. DOI 10.1093/hmg/ddq316. https://pubmed.ncbi.nlm.nih.gov/20656788.
Saus E., Soria V., Escaramis G., Vivarelli F., Crespo J.M., Kagerbauer B., Menchón J.M., Urretavizcaya M., Gratacòs M., Estivill X. Genetic variants and abnormal processing of pre-miR182, a circadian clock modulator, in major depression patients with late insomnia. Hum. Mol. Genet. 2010;19(20):4017-4025. DOI 10.1093/hmg/ddq316. https://pubmed.ncbi.nlm.nih.gov/20656788.
72. Selbach M., Schwanhäusser B., Thierfelder N., Fang Z., Khanin R., Rajewsky N. Widespread changes in protein synthesis induced by microRNAs. Nature. 2008;455(7209):58-63. DOI 10.1038/nature07228. https://pubmed.ncbi.nlm.nih.gov/18668040.
73. Shen J., Li Y., Qu C., Xu L., Sun H., Zhang J. The enriched environment ameliorates chronic unpredictable mild stress-induced depressive-like behaviors and cognitive impairment by activating the SIRT1/miR-134 signaling pathway in hippocampus. J. Affect Disord. 2019;248:81-90. DOI 10.1016/j.jad.2019.01.031. https://pubmed.ncbi.nlm.nih.gov/30716615.
J. Affect Disord. 2019;248:81-90. DOI 10.1016/j.jad.2019.01.031. https://pubmed.ncbi.nlm.nih.gov/30716615.
74. Siegert S., Seo J., Kwon E.J., RudenkoA., Cho S., Wang W., Flood Z., Martorell A.J., Ericsson M., Mungenast A.E., Tsai L.H. The schizophrenia risk gene product miR-137 alters presynaptic plasticity. Nat. Neurosci. 2015;18(7):1008-1016. DOI 10.1038/nn.4023. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4506960.
75. Sim S.E., Lim C.S., Kim J.I., Seo D., Chun H., Yu N.K., Lee J., Kang S.J., Ko H.G., Choi J.H., Kim T., Jang E.H., Han J., Bak M.S., Park J.E., Jang D.J., Baek D., Lee Y.S., Kaang B.K. The brain-enriched microRNA miR-9-3p regulates synaptic plasticity and memory. J. Neurosci. 2016;36(33):8641-8652. DOI 10.1523/JNEUROSCI.0630-16.2016. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6601897.
76. Smalheiser N.R. The RNA-centred view of the synapse: non-coding RNAs and synaptic plasticity. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2014;369(1652):20130504. DOI 10.1098/rstb.2013.0504. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4142025.
Smalheiser N.R. The RNA-centred view of the synapse: non-coding RNAs and synaptic plasticity. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2014;369(1652):20130504. DOI 10.1098/rstb.2013.0504. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4142025.
77. Smith A.C.W., Kenny P.J. MicroRNAs regulate synaptic plasticity underlying drug addiction. Genes Brain Behav. 2018;17(3): e12424. DOI 10.1111/gbb.12424. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5837931.
78. Sweatt J.D. Neural plasticity and behavior – sixty years of conceptual advances. J. Neurochem. 2016;139(Suppl.2):179-199. DOI 10.1111/jnc.13580. https://pubmed.ncbi.nlm.nih.gov/26875778.
79. Vetere G., Barbato C., Pezzola S., Frisone P., Aceti M., Ciotti M., Cogoni C., Ammassari-Teule M., Ruberti F. Selective inhibition of miR-92 in hippocampal neurons alters contextual fear memory. Hippocampus. 2014;24(12):1458-1465. DOI 10.1002/hipo.22326. https://pubmed.ncbi.nlm.nih.gov/24990518.
80. Wang C.N., Wang Y.J., Wang H., Song L., Chen Y., Wang J.L., Ye Y., Jiang B. The anti-dementia effects of Donepezil involve miR-206- 3p in the hippocampus and cortex. Biol. Pharm. Bull. 2017;40(4): 465-472. DOI 10.1248/bpb.b16-00898. https://pubmed.ncbi.nlm.nih.gov/28123152.
81. Wang X., Liu D., Huang H.Z., Wang Z.H., Hou T.Y., Yang X., Pang P., Wei N., Zhou Y.F., Dupras M.J., Calon F., Wang Y.T., Man H.Y., Chen J.G., Wang J.Z., Hébert S.S., Lu Y., Zhu L.Q. A novel microRNA-124/PTPN1 signal pathway mediates synaptic and memory deficits in Alzheimer’s disease. Biol. Psychiatry. 2018;83(5):395-405. DOI 10.1016/j.biopsych.2017.07.023. https://pubmed.ncbi.nlm.nih.gov/28965984.
82. Wingo T.S., Yang J., Fan W., Min Canon S., Gerasimov E.S., Lori A., Logsdon B., Yao B., Seyfried N.T., Lah J.J., LeveyA.I., Boyle P.A., Schneider J.A., De Jager P.L., Bennett D.A., Wingo A.P. Brain microRNAs associated with late-life depressive symptoms are also associated with cognitive trajectory and dementia. NPJ Genom. Med. 2020;5:6. DOI 10.1038/s41525-019-0113-8. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7004995.
83. Woldemichael B.T., Jawaid A., Kremer E.A., Gaur N., Krol J., Marchais A., Mansuy I.M. The microRNA cluster miR-183/96/182 contributes to long-term memory in a protein phosphatase 1-dependent manner. Nat. Commun. 2016;7:12594. DOI 10.1038/ncomms12594. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5007330.
84. Wu Y.Y., Kuo H.C. Functional roles and networks of non-coding RNAs in the pathogenesis of neurodegenerative diseases. J. Biomed. Sci. 2020;27(1):49. DOI 10.1186/s12929-020-00636-z. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7140545.
85. Yan H.L., Sun X.W., Wang Z.M., Liu P.P., Mi T.W., Liu C., Wang Y.Y., He X.C., Du H.Z., Liu C.M., Teng Z.Q. MiR-137 deficiency causes anxiety-like behaviors in mice. Front. Mol. Neuro¬ sci. 2019;12:260. DOI 10.3389/fnmol.2019.00260. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6831983.
86. Yang Y., Shu X., Liu D., Shang Y., Wu Y., Pei L., Xu X., Tian Q., Zhang J., Qian K., Wang Y.X., Petralia R.S., Tu W., Zhu L.Q., Wang J.Z., Lu Y. EPAC null mutation impairs learning and social interactions via aberrant regulation of miR-124 and Zif 268 translation. Neuron. 2012;73(4):774-788. DOI 10.1016/j.neuron.2012.02.003. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3307595.
87. You Y.H., Qin Z.Q., Zhang H.L., Yuan Z.H., Yu X. MicroRNA-153 promotes brain-derived neurotrophic factor and hippocampal neuron proliferation to alleviate autism symptoms through inhibition of JAK-STAT pathway by LEPR. Biosci. Rep. 2019;39(6): BSR20181904. DOI 10.1042/BSR20181904. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6591574.
88. Zovoilis A., Agbemenyah H.Y., Agis-Balboa R.C., Stilling R.M., Edbauer D., Rao P., Farinelli L., Delalle I., Schmitt A., Falkai P., Bahari-Javan S., Burkhardt S., Sananbenesi F., Fischer A. MicroRNA-34c is a novel target to treat dementias. EMBO J. 2011;30: 4299-4308. DOI 10.1038/emboj.2011.327. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3199394.
Долговременная память — ИнфоКомпУниверсал
Для долговременного хранения информации используется долговременная (внешняя) память. Устройство, которое обеспечивает запись и считывание информации, называется накопителем, или дисководом, а хранится информация на носителях информации. Информация на носителях хранится в двоичном компьютерном коде, т. е. в форме последовательностей нулей и единиц.
Дискеты.
Внутри пластмассового корпуса дискеты размещается гибкий магнитный диск.Информация на диске хранится на концентрических дорожках.
Считывание информации с дискеты
Жесткие магнитные диски.
Жесткий магнитный диск представляет собой несколько тонких металлических дисков, очень быстро вращающихся на одной оси и заключенных в металлический корпус.
Сверхминиатюрные магнитные головки могут записывать или считывать информацию с сотен тысяч концентрических дорожек, поэтому информационная емкость жестких дисков очень велика и может достигать 400 Гбайт.
Оптические дисководы и диски.
В оптических дисководах используется оптический принцип записи и считывания информации. Информация на оптическом диске хранится на одной спиралевидной дорожке (похожей по форме на раковину улитки), идущей от центра диска к периферии и содержащей чередующиеся участки с хорошей и плохой отражающей способностью.
Существуют два типа оптических дисков:
- CD-диски (CD — Compact Disk, компакт-диск), на которые может быть записано до 700 Мбайт информации;
- DVD-диски (DVD — Digital Versatile Disk, цифровой универсальный диск), которые имеют значительно большую информационную емкость (4,7 Гбайт и более), так как оптические дорожки на них имеют меньшую толщину и размещены более плотно
Энергонезависимая память.
Энергонезависимая память (карты памяти и flash-диски) применяется для долговременного хранения информации и не требует, в отличие от оперативной памяти, подключения источника электрического напряжения (например, батарейки). Такая память не имеет движущихся частей и поэтому обеспечивает высокую сохранность данных при использовании в мобильных устройствах (портативных компьютерах, цифровых камерах и т. д.). Информационная емкость flash-памяти может достигать 1 Гбайт и более.
Для предотвращения потери информации на носителях и их выхода из строя необходимо:
- модули оперативной памяти оберегать от электростатических зарядов при установке;
- дискеты оберегать от нагревания и сильных магнитных полей, которые могут изменить намагниченность участков поверхности диска;
- жесткие диски оберегать от ударов при установке, которые могут привести к поломке механизма перемещения магнитных головок и повреждению поверхности магнитных дисков;
- оптические диски оберегать от загрязнений и царапин, которые могут привести к изменению отражающей способности отдельных участков поверхности;
- flash-память оберегать от неправильного отключения от компьютера.
flash-диск
Карты энергонезависимой памяти и адаптер
Развитие долговременной памяти у детей
Долговременная памятьДолговременная память существенно улучшается в одном из своих проявлений и незначительно — в другом. Данное заявление может показаться противоречивым, пока мы не проанализируем то, какие требования к долговременной памяти предъявляет решение разных задач.
В любом возрасте узнавать («Какие из этих предметов вы видели прежде?») проще, чем воспроизводить («Опишите, какие предметы вы видели»), однако у маленьких детей эта разница достигает максимума. Иными словами, с возрастом способность к воспроизведению улучшается в большей степени, чем способность к узнаванию. Десятилетний ребенок после предъявления 12 картинок скорее всего сможет описать 8 из них и узнать все 12. Четырехлетний ребенок также узнает все 12 картинок, однако описать может только 2 или 3.
Существует несколько причин, по которым с возрастом улучшается способность актуализировать информацию, хранящуюся в долговременной памяти. Одна из них заключается в том, что происходит совершенствование базы знаний, используемых для кодирования и актуализации информации. Дети расширяют свой кругозор, приобретают систематизированные знания, усваивают более сложные понятия. Но в некоторых областях познания ребенка могут оказаться более глубокими, чем у взрослого. В этом случае можно было бы ожидать, что в данной области память ребенка будет функционировать лучше, чем у взрослого. По-видимому, это утверждение справедливо. Было обнаружено, что дети, участвовавшие в турнире по шахматам, лучше воспроизводили положение фигур на шахматной доске, чем взрослые, поверхностно знакомые с этой игрой (Chi, 1978).
Вторая причина улучшения способности к воспроизведению заключается в развитии стратегий запоминания. Предположим, воспитательница детского сада говорит, обращаясь к группе 4-летних детей: «Я прочитаю вам рассказ. Я хочу, чтобы вы внимательно его послушали, потому что потом я задам о нем вопросы». По всей вероятности, эти дети запомнят рассказ ничуть не лучше, чем те, кому не давали указания запоминать. Но к 8-10-летнему возрасту выполнение задания значительно улучшается при наличии предварительной инструкции. Одна из причин заключается в том, что дети старшего возраста овладевают стратегиями запоминания, т. е. более эффективными стратегиями перекодирования, повторения и удержания в памяти.
Давайте более тщательно проанализируем одну из этих стратегий. Она заключается в том, чтобы основное время отводить на кодирование и анализ наиболее важной информации. С возрастом у детей все лучше получается выделять для запоминания существенную информацию, т. е. «отделять зерна от плевел». Например, в одном из исследований, посвященном этому вопросу, детям показывали два ряда небольших дверок по 6 в каждом ряду (см. рис. 9.4).
Рис. 9.4. Тест на оценку мнемонических стратегий детей. За дверцами скрыты изображения животных (за дверцами с клеткой) и предметов домашнего обихода (за дверцами с домиками). Дети использовали разные стратегии, если их просили сравнить два ряда или запомнить все предметы домашнего обихода. Источник: Р. Н. Miller, V. F. Haynes, D. DeMa-rie-Dreblow, & Woody-Ramsey, 1986, Child Development, 57, p. 1429-1439.
На половине из них были нарисованы клетки, а за ними скрывались изображения животных; на других 6 дверцах были дома, а за ними скрывались картинки с изображениями предметов домашнего обихода. Детям давали одно из двух заданий:
1) решить, совпадают или не совпадают картинки верхнего и нижнего рядов;
2) запомнить всех животных (предметы домашнего обихода).
Перед каждым испытанием дети получали инструкцию открывать дверцы по одной и рассматривать картинки. Наиболее эффективная стратегия сопоставления — открывать одну дверцу наверху, а затем — одну внизу, сравнивая пары картинок. Для задания на воспроизведение более эффективным являлось открывание дверцы только той категории, которую нужно запомнить. Дети лет 8-10 использовали эти стратегии более последовательно, чем 6-летние. Младшие дети были склонны беспорядочно открывать все дверцы, несмотря на то что их просили запомнить картинки только одной категории (Р. Н. Miller, Haynes, DeMarie-Dreblow, & Woody-Ramsey, 1986).
В использовании стратегий существуют и индивидуальные различия между детьми одной возрастной группы. Например, когда детям 5,6 и 7-х классов давали дополнительное время для подготовки к контрольной работе, большинство из них просто перелистывали свои учебники. Тем не менее были и такие, кто подчеркивал и выписывал существенные моменты. Эти дети не только отбирали для кодирования наиболее важную информацию, но и активно ее реструктурировали и группировали удобным для себя образом. Неудивительно, что они получили более высокие оценки за контрольную работу (A. L. Brown, Bransford, Ferrara, & Campione,1983).
Важный практический вопрос, касающийся детской памяти, заключается в том, можно ли доверять свидетельствам детей, данным в суде. Сегодня этот вопрос особо волнует широкую общественность в связи с делами, в которых предъявляются обвинения в сексуальном насилии над детьми. Вывод, сделанный на основе обзора литературы по данному вопросу (Ceci & Bruck, 1993), состоит в том, что хотя дошкольники и могут помнить гораздо больше, чем принято думать, они более подвержены эффектам внушения, чем старшие дети и взрослые. (Во вставке 9.2 представлен рассказ о воспоминаниях раннего детства и оценка их точности.)
Ньюкомб Н. Развитие личности ребенка. — СПб.: Питер, 2003. — 640 с. С. 295-297.
В чем разница между долговременной, краткосрочной и рабочей памятью?
Prog Brain Res. Авторская рукопись; доступно в PMC 2009 18 марта.
Опубликован в окончательной редакции как:
PMCID: PMC2657600
NIHMSID: NIHMS84208
Нельсон Коуэн
Департамент психологических наук, Университет Миссури, 18 Columbialester Hall 65211, США
Нельсон Коуэн, Департамент психологических наук, Университет Миссури, 18 Макалестер Холл, Колумбия, Миссури 65211, США;
* Автор, ответственный за переписку.Тел .: +1 573-882-4232; Факс: +1 573-882-7710; E-mail: ude.iruossim@NnawoC Окончательная отредактированная версия этой статьи издателем доступна на Prog Brain Res. См. Другие статьи в PMC, в которых цитируется опубликованная статья.Abstract
В недавней литературе существует значительная путаница в отношении трех типов памяти: долговременной, кратковременной и рабочей памяти. В этой главе делается попытка уменьшить эту путаницу и сделать современные оценки этих типов памяти. Долговременная и кратковременная память могут различаться двумя фундаментальными способами: только кратковременная память демонстрирует (1) временное затухание и (2) ограничения емкости фрагментов.Оба свойства кратковременной памяти все еще спорны, но текущая литература довольно обнадеживает в отношении существования как распада, так и пределов емкости. Рабочая память была задумана и определена тремя разными, слегка противоречащими друг другу способами: как краткосрочная память, применяемая к когнитивным задачам, как многокомпонентная система, которая хранит и управляет информацией в кратковременной памяти, и как использование внимания для управления краткосрочная память. Независимо от определения, есть некоторые показатели краткосрочной памяти, которые кажутся рутинными и плохо коррелируют с когнитивными способностями и другими показателями (обычно определяемыми термином «рабочая память»), которые кажутся требующими большего внимания и хорошо коррелируют. с этими способностями.Доказательства оцениваются и помещаются в теоретические рамки, изображенные в.
Ключевые слова: внимание, емкость рабочей памяти, контроль внимания, распад кратковременной памяти, фокус внимания, долговременная память, кратковременная память, рабочая память
Исторические корни основного научного вопроса
Сколько фаз в памяти? С наивной точки зрения на память, это могло быть все из одной ткани. У некоторых людей есть хорошая способность фиксировать факты и события в памяти, тогда как у других такая способность меньше.Тем не менее, задолго до того, как появились настоящие психологические лаборатории, более тщательное наблюдение должно было показать, что существуют отдельные аспекты памяти. Можно было бы увидеть пожилого учителя, рассказывающего старые уроки так же живо, как и прежде, и все же может быть очевидно, что его способность фиксировать имена новых учеников или вспоминать, какой ученик какой комментарий делал в продолжающемся разговоре, уменьшилась за время годы.
Научное изучение памяти обычно восходит к Герману Эббингаузу (перевод 1885/1913), который исследовал собственное получение и забывание новой информации в виде серий бессмысленных слогов, проверенных в различные периоды до 31 дня.Среди многих важных наблюдений Эббингаус заметил, что у него часто было «первое мимолетное представление… серии в моменты особой концентрации» (стр. 33), но это непосредственное воспоминание не гарантирует, что серия была запомнена таким образом, чтобы позвольте его вспомнить позже. Устойчивое запоминание иногда требовало повторения ряда. Вскоре после этого Джеймс (1890) предложил различать первичную память, небольшой объем информации, удерживаемой в качестве задней границы сознательного настоящего, и вторичную память, обширный массив знаний, хранимых на протяжении всей жизни.Первичное воспоминание о Джеймсе похоже на первое мимолетное взятие Эббингауза.
Промышленная революция предъявила новые требования к тому, что Джеймс (1890) назвал первичной памятью. В 1850-х годах телеграфистам приходилось запоминать и интерпретировать быстрые серии точек и тире, передаваемых акустически. В 1876 году был изобретен телефон. Три года спустя операторы в Лоуэлле, штат Массачусетс, начали использовать телефонные номера для более чем 200 абонентов, чтобы было легче обучить заменяющих операторов, если бы четыре штатных оператора города стали жертвой бушующей эпидемии кори.Такое использование телефонных номеров, дополненных префиксом слова, конечно же, распространилось. (Телефонный номер автора в 1957 году был Уайтхолл 2–6742; номер все еще присваивается, хотя и в виде семизначного числа.) Еще до книги Эббингауза Нифер (1878) сообщил о кривой порядкового положения, полученной среди цифр в логарифмы, которые он пытался вспомнить. Можно заметить, что бессмысленные слоги, изобретенные Эббингаузом в качестве инструмента, приобрели большую экологическую ценность в индустриальную эпоху с растущими требованиями к информации, что, возможно, подчеркивает практическую важность первичной памяти в повседневной жизни.Первичная память кажется обременительной, поскольку человека просят помнить об аспектах незнакомой ситуации, таких как имена, места, вещи и идеи, с которыми он раньше не сталкивался.
Тем не менее, субъективное переживание разницы между первичной и вторичной памятью автоматически не гарантирует, что эти типы памяти по отдельности способствуют развитию науки о запоминании. Исследователи с другой точки зрения давно надеялись, что они смогут написать единое уравнение или, по крайней мере, единый набор принципов, которые охватили бы всю память, от самой непосредственной до очень долгосрочной.МакГеоч (1932) проиллюстрировал, что забывание с течением времени было не просто вопросом неизбежного распада памяти, а скорее результатом вмешательства во время интервала сохранения; можно было найти ситуации, в которых память со временем улучшалась, а не уменьшалась. С этой точки зрения можно было бы рассматривать то, что казалось забвением из первичной памяти, как глубокий эффект вмешательства со стороны других элементов в память для любого элемента, при этом эффекты вмешательства продолжаются вечно, но не полностью разрушают данное воспоминание.Эта точка зрения поддерживалась и развивалась на протяжении многих лет непрерывной линией исследователей, верящих в единство памяти, включая, среди прочего, Мелтона (1963), Бьорка и Уиттена (1974), Викельгрена (1974), Краудера (1982, 1993). ), Гленберг и Свансон (1986), Браун и др. (2000), Nairne (2002), Neath and Surprenant (2003) и Lewandowsky et al. (2004).
Описание трех видов памяти
В этой главе я оценим силу доказательств для трех типов памяти: долговременной памяти, кратковременной памяти и рабочей памяти. Долговременная память — это обширная база знаний и запись предыдущих событий, и она существует согласно всем теоретическим представлениям; Было бы трудно отрицать, что каждый нормальный человек имеет в своем распоряжении богатый, хотя и не безупречный или полный, набор долговременных воспоминаний.
Кратковременная память относится к первичной памяти Джеймса (1890) и представляет собой термин, который Бродбент (1958), Аткинсон и Шиффрин (1968) использовали несколько иначе. Как Аткинсон и Шиффрин, я считаю, что это отражает способности человеческого разума, который может временно удерживать ограниченный объем информации в очень доступном состоянии.Одно различие между термином «кратковременная память» и термином «первичная память» состоит в том, что последний может рассматриваться как более ограниченный. Возможно, что не каждая временно доступная идея находится или даже находилась в сознательном осознании. Например, согласно этой концепции, если вы разговариваете с человеком с иностранным акцентом и непреднамеренно изменяете свою речь, чтобы она соответствовала акценту иностранного говорящего, на вас влияет то, что до этого момента было бессознательным (и, следовательно, неконтролируемым) аспектом вашей короткой речи. -срочная память.Можно связать кратковременную память с паттерном нейронного возбуждения, который представляет конкретную идею, и можно было бы считать, что идея находится в кратковременной памяти только тогда, когда активен паттерн возбуждения или сборка клеток (Hebb, 1949). Человек может осознавать или не осознавать идею в течение этого периода активации.
Рабочая память не полностью отличается от кратковременной памяти. Это термин, который использовали Миллер и др. (1960) для обозначения памяти, поскольку она используется для планирования и выполнения поведения.Можно полагаться на рабочую память, чтобы сохранить частичные результаты при решении арифметической задачи без бумаги, чтобы объединить предпосылки в длинном риторическом споре или испечь торт, не допустив досадной ошибки добавления одного и того же ингредиента дважды. (Ваша рабочая память была бы более загружена при чтении предыдущего предложения, если бы я сохранил фразу «один полагается на рабочую память» до конца предложения, что я сделал в моем первом черновике этого предложения; рабочая память, таким образом, влияет на хорошее письмо.Термин «рабочая память» стал гораздо более доминирующим в этой области после того, как Баддели и Хитч (1974) продемонстрировали, что один модуль не может учитывать все виды временной памяти. Их мышление привело к влиятельной модели (Baddeley, 1986), в которой вербально-фонологические и визуально-пространственные репрезентации проводились отдельно, а управление ими и манипулирование ими осуществлялись с помощью процессов, связанных с вниманием, называемых центральным исполнителем. В статье 1974 года у этого центрального руководителя, возможно, была своя собственная память, которая пересекала области репрезентации.К 1986 году эта общая память была исключена из модели, но она была снова добавлена Баддели (2000) в форме эпизодического буфера . Это казалось необходимым для объяснения кратковременной памяти функций, которые не совпадали с другими хранилищами (особенно семантической информации в памяти), и объяснения междоменных ассоциаций в рабочей памяти, таких как сохранение связей между именами и лицами. Благодаря работе Baddeley et al. (1975) рабочая память обычно рассматривается как комбинация нескольких компонентов, работающих вместе.Некоторые даже включают в этот набор значительный вклад долговременной памяти, которая снижает нагрузку на рабочую память за счет организации и группировки информации в рабочей памяти в меньшее количество единиц (Miller, 1956; Ericsson and Kintsch, 1995). Например, буквенную серию IRSCIAFBI гораздо легче запомнить как серию сокращений для трех федеральных агентств Соединенных Штатов Америки: налоговой службы (IRS), Центрального разведывательного управления (ЦРУ) и Федерального бюро разведки. Расследование (ФБР).Однако этот фактор не был подчеркнут в известной модели Баддели (1986).
Из моего определения ясно, что рабочая память включает в себя кратковременную память и другие механизмы обработки, которые помогают использовать кратковременную память. Это определение отличается от того, которое использовали некоторые другие исследователи (например, Engle, 2002), которые хотели бы зарезервировать термин рабочая память для обозначения только связанных с вниманием аспектов кратковременной памяти. Это, однако, не столько дискуссия по существу, сколько немного сбивающее с толку несоответствие в использовании терминов.
Одна из причин использовать термин рабочая память состоит в том, что показатели рабочей памяти, как было установлено, коррелируют с интеллектуальными способностями (и особенно подвижным интеллектом) лучше, чем показатели кратковременной памяти, и, фактически, возможно, лучше, чем показатели любых других конкретный психологический процесс (например, Данеман и Карпентер, 1980; Киллонен и Кристал, 1990; Данеман и Мерикл, 1996; Энгл и др., 1999; Конвей и др., 2005). Считалось, что это отражает использование мер, которые включают не только хранение, но и обработку, при этом предполагается, что и хранение, и обработка должны выполняться одновременно для оценки емкости рабочей памяти способом, связанным с когнитивными способностями.Совсем недавно Engle et al. (1999) представили понятие, что и способности, и рабочая память зависят от способности контролировать внимание или применять контроль внимания к управлению как первичной, так и вторичной памятью (Unsworth and Engle, 2007). Однако необходимы дополнительные исследования того, что мы узнаем из высокой корреляции между рабочей памятью и интеллектуальными способностями, и этот вопрос будет обсуждаться далее после того, как будет решен более фундаментальный вопрос различия краткосрочной и долгосрочной памяти.
Между тем, может быть полезно резюмировать теоретические основы (Cowan, 1988, 1995, 1999, 2001, 2005), основанные на прошлых исследованиях. Эта структура, проиллюстрированная в, помогает учесть взаимосвязь между механизмами долгосрочной, краткосрочной и рабочей памяти и объясняет то, что я вижу как взаимосвязь между ними. В этой структуре кратковременная память получается из временно активированного подмножества информации в долговременной памяти. Это активированное подмножество может распадаться со временем, если оно не обновляется, хотя свидетельства распада в лучшем случае являются предварительными.В фокусе внимания находится подмножество активированной информации, которое, по-видимому, ограничено по емкости блока (сколько отдельных элементов может быть включено одновременно). Новые ассоциации между активированными элементами могут стать центром внимания. Теперь мы обсудим доказательства, связанные с этой структурой моделирования.
Различие между кратковременной памятью и долговременной памятью
Если есть разница между кратковременной и долговременной памятью, есть два возможных способа, которыми эти хранилища могут отличаться: длительностью и вместимость .Разница в продолжительности означает, что предметы, находящиеся на краткосрочном хранении, со временем распадаются из-за такого хранения. Разница в емкости означает, что существует ограничение на количество предметов, которые можно хранить на кратковременном хранении. Если есть только ограничение по емкости, количество элементов, меньшее, чем ограничение по емкости, может оставаться в краткосрочном хранилище до тех пор, пока они не будут заменены другими элементами. Оба типа ограничения спорны. Следовательно, чтобы оценить полезность концепции краткосрочного хранения, поочередно будут оцениваться ограничения продолжительности и емкости.
Пределы продолжительности
Концепция кратковременной памяти, ограничиваемой распадом с течением времени, присутствовала даже на заре когнитивной психологии, например, в работе Бродбента (1958). Если бы распад был единственным принципом, влияющим на производительность в эксперименте с непосредственной памятью, возможно, его было бы легко обнаружить. Однако даже в работе Бродбента загрязняющие переменные были признаны. Чтобы оценить распад, нужно принять во внимание или преодолеть загрязняющие эффекты репетиции, длительного поиска и временной различимости, которые будут обсуждаться по отдельности вместе с доказательствами за и против распада.
Преодоление загрязнения от репетиции
По мнению различных исследователей, существует процесс, с помощью которого можно представить себе, как произносятся слова в списке, не произнося их вслух, и этот процесс называется скрытой словесной репетицией. С практикой этот процесс происходит с минимумом внимания. Гуттентаг (1984) использовал второстепенное задание, чтобы показать, что репетиция списка, который нужно вспомнить, требует усилий у маленьких детей, но не у взрослых. Если в конкретной экспериментальной процедуре не наблюдается потери кратковременной памяти, можно приписать этот образец реакции репетиции.Поэтому были предприняты шаги по устранению репетиций посредством процесса, называемого артикуляционным подавлением, в котором простое высказывание, такое как слово «the», многократно произносится участником в течение части или всего задания на краткосрочную память (например, Baddeley et al. др., 1975). Все еще есть возможное возражение, что любое высказывание, используемое для подавления репетиции, к сожалению, вызывает помехи, которые могут быть истинной причиной потери памяти с течением времени, а не распада.
Эта проблема интерференции может показаться спорной в свете выводов Левандовски и др.(2004). Они представили списки писем, которые нужно было вспомнить, и различали, сколько времени должно было потребоваться участнику, чтобы вспомнить каждый элемент в списке. В некоторых условиях они добавляли артикуляционное подавление, чтобы предотвратить репетицию. Несмотря на это подавление, они не наблюдали никакой разницы в производительности, когда время между элементами в ответе варьировалось от 400 до 1600 мс (или между условиями, в которых слово «супер» произносилось один, два или три раза между последовательными элементами в ответе. ).Они не обнаружили никаких свидетельств разрушения памяти.
Ограничение этого открытия состоит в том, что скрытая словесная репетиция может быть не единственным типом репетиции, который могут использовать участники. Возможно, есть виды, которым артикуляторное подавление не предотвращает. В частности, Коуэн (Cowan, 1992) предположил, что процесс мысленного внимания к словам или поиска по списку, требующий внимания процесс, может служить для повторной активации элементов, которые нужно вспомнить, аналогично скрытой вербальной репетиции.Ключевое отличие состоит в том, что нельзя было ожидать, что подавление артикуляций может помешать репетициям такого типа. Вместо этого, чтобы предотвратить репетиции такого типа, нужно было бы использовать задачу, требующую внимания.
Barrouillet et al. (2004, 2007) есть результаты, которые, кажется, предполагают, что существует другой, более требующий внимания тип репетиции. В них вставлены материалы между вызываемыми элементами, которые требуют выбора; это могут быть числа для чтения вслух или время реакции с множественным выбором.Было обнаружено, что они мешают удержанию в степени, соизмеримой с долей интервала между пунктами, израсходованного на отвлекающие элементы. По мере того, как количество отвлекающих элементов увеличивается, вызывается меньше элементов, которые нужно отозвать. Идея состоит в том, что, когда отвлекающая задача не требует внимания, высвободившееся внимание позволяет вспомнить основанную на внимании репетицию предметов. Когда вставленная задача более автоматическая и не требует такого внимания (например,g., задача подавления артикуляции) гораздо меньше влияние скорости этих вставленных элементов.
На основе этой логики можно представить себе версию задачи Левандовски, в которой не артикуляционное подавление, а требующие внимания вербальные стимулы помещаются между элементами в ответе, и в которой продолжительность этого заполненного времени между элементами в ответе варьируется от от суда к делу. Вербальные, требующие внимания стимулы должны препятствовать как репетициям, основанным на внимании, так и репетициям на основе артикуляции.Если есть спад, то производительность по последовательным позициям должна снизиться сильнее, когда между элементами в ответе будут помещены более длинные заполненные интервалы. К сожалению, такие результаты могут быть объяснены альтернативно как результат вмешательства отвлекающих стимулов, без необходимости вызывать затухание.
В таком случае, кажется, необходима процедура для предотвращения репетиций, основанных как на артикуляции, так и на внимании, без создания помех. Коуэн и Обушон (в печати) опробовали один тип процедуры, с помощью которой можно добиться этого.Они представили списки из семи печатных цифр, в которых время между пунктами в списке варьировалось. В дополнение к некоторым спискам заполнителей, составленных случайным образом, было четыре критических типа испытаний, в которых все шесть интервалов между цифрами были короткими (0,5 с после каждого элемента) или все длинными (2 с после каждого элемента) или состояли из трех коротких а затем три длинных интервала или три длинных, а затем три коротких интервала. Более того, было две реплики для ответа на пост-лист. Согласно одной из подсказок, участник должен был вспомнить список с пунктами в представленном порядке, но в любом случае они хотели.Согласно другой реплике, список должен был быть отозван в то же время, в которое он был представлен. Ожидалось, что необходимость запомнить время в последнем условии ответа предотвратит репетицию любого типа. Как следствие, производительность должна снижаться в испытаниях, в которых первые три интервала ответа длинные, потому что в этих испытаниях больше времени для того, чтобы забыть большинство пунктов списка. Как и предполагалось, было существенное взаимодействие между сигналом ответа и длиной первой половины интервалов ответа.Когда участники могли свободно вспоминать задания в своем собственном темпе, результативность в короткой первой половине ( M = 0,71) была не лучше, чем в длинной первой половине ( M = 0,74). Небольшая выгода от длинной первой половины в этой ситуации могла быть получена, потому что она позволяла отрепетировать список на ранней стадии ответа. Напротив, когда время отзыва должно было соответствовать времени представления списка, производительность была лучше с короткой первой половиной ( M = 0,70), чем с длинной первой половиной ( M =.67). Таким образом, это предполагает, что краткосрочная память может ухудшиться.
Преодоление загрязнения из-за длительного поиска
Если существует более одного типа хранилища памяти, то все еще остается проблема, какое хранилище предоставило информацию, лежащую в основе ответа. Нет никакой гарантии, что только потому, что процедура считается тестом на краткосрочное хранение, долгосрочное хранение не будет использоваться. Например, в простой задаче с диапазоном цифр представлена серия цифр, которую необходимо повторить сразу после этого из памяти.Если эта серия окажется лишь немного отличной от телефонного номера участника, участник может быстро запомнить новый номер и повторить его из долговременной памяти. Теории памяти с двойным хранилищем допускают это. Хотя Бродбент (1958), Аткинсон и Шиффрин (1968) изобразили свои модели обработки информации как серию прямоугольников, представляющих различные хранилища памяти, причем долговременная память следует за кратковременной памятью, эти прямоугольники не подразумевают, что память находится исключительно в одном коробка или другое; их лучше интерпретировать как относительное время первого ввода информации от стимула в одно хранилище, а затем в следующее.Остается вопрос, как определить, исходит ли реакция из кратковременной памяти.
Во и Норман (1965) разработали математическую модель для этого. Модель работала с предположением, что долговременная память имеет место для всего списка, включая плато в середине списка. Напротив, к моменту припоминания кратковременная память остается только в конце списка. Эта модель предполагает, что для любой конкретной серийной позиции в списке вероятность успешного краткосрочного хранения (S) и долгосрочного хранения (L) независимы, так что вероятность отзыва элемента равна S + L-SL. .
Несколько иное предположение состоит в том, что краткосрочные и долгосрочные магазины не являются независимыми, а используются во взаимодополняемости. Наличие кратковременной памяти элемента может позволить переместить ресурсы, необходимые для долговременного запоминания, в другое место в списке. Данные кажутся более согласующимися с этим предположением. В нескольких исследованиях списки, которые следует вспомнить, были представлены пациентам с амнезией Корсакова и нормальным участникам контрольной группы (Baddeley and Warrington, 1970; Carlesimo et al., 1995). Эти исследования показывают, что при немедленном воспроизведении показатели у пациентов с амнезией сохраняются на последних нескольких порядковых позициях в списке. Как если бы производительность в этих последовательных положениях основывалась в основном или полностью на кратковременном хранении, и у пациентов с амнезией не было уменьшения такого рода хранения. При отсроченном воспоминании пациенты с амнезией демонстрируют дефицит во всех последовательных положениях, как и следовало ожидать, если кратковременная память на конец списка теряется в зависимости от заполненного периода задержки (Glanzer and Cunitz, 1966).
Преодоление загрязнения из-за временной различимости
Наконец, утверждалось, что потеря памяти с течением времени не обязательно является результатом распада. Напротив, это может быть вызвано временными различиями при поиске. Такая теория предполагает, что временной контекст элемента служит сигналом для извлечения этого элемента даже при свободном вызове. Предмет, отделенный во времени от всех других предметов, относительно отличен и его легко вспомнить, тогда как предмет, который относительно близок к другим предметам, вспомнить труднее, потому что он разделяет их временные сигналы для извлечения.Вскоре после того, как список представлен, самые свежие элементы становятся наиболее отчетливыми во времени (во многом как отчетливость телефонного столба, которого вы практически касаетесь, по сравнению с столбами, идущими дальше по дороге). По прошествии интервала хранения относительная различимость самых последних элементов уменьшается (так же, как если бы они стояли далеко от последнего полюса в серии).
Хотя есть данные, которые можно интерпретировать в соответствии с различимостью, есть также то, что выглядит как диссоциация между эффектами различимости и подлинным эффектом кратковременной памяти.Это можно увидеть, например, в классической процедуре Петерсона и Петерсона (1959), в которой буквенные триграммы следует вызывать сразу или только после отвлекающей задачи, считая в обратном порядке от начального числа на три в течение периода до 18 с. Петерсон и Петерсон обнаружили серьезную потерю памяти для буквенной триграммы при увеличении заполненной задержки. Однако впоследствии скептики утверждали, что потеря памяти произошла из-за того, что временная различимость текущей буквенной триграммы уменьшалась по мере увеличения заполненной задержки.В частности, было сказано, что этот эффект задержки возникает из-за увеличения задержек между тестами из-за проактивных помех от предыдущих испытаний. В первых нескольких испытаниях задержка не имеет значения (Keppel and Underwood, 1962), и никакого вредного воздействия задержки не наблюдается, если задержки в 5, 10, 15 и 20 с тестируются в отдельных пробных блоках (Turvey et al., 1970; Грин, 1996).
Тем не менее, при более коротких интервалах испытаний может наблюдаться настоящий эффект распада. Баддели и Скотт (1971) установили трейлер в торговом центре, чтобы они могли протестировать большое количество участников по одному испытанию, чтобы избежать упреждающего вмешательства.Они обнаружили эффект задержки теста в течение первых 5 с, но не при более длительных задержках. Тем не менее, кажется, что концепция распада еще не имеет прочной основы и требует дальнейшего изучения. Может случиться так, что распад на самом деле отражает не постепенное ухудшение качества записи кратковременной памяти, а внезапный коллапс в точке, которая меняется от испытания к испытанию. С контролем временной различимости Cowan et al. (1997a) обнаружили, что может быть внезапный коллапс в представлении памяти для тона с задержками от 5 до 10 с.
Пределы емкости блоков
В истории когнитивной психологии концепция пределов емкости поднималась несколько раз. Миллер (1956), как известно, обсуждал «магическое число семь плюс-минус два» как константу в краткосрочной обработке, включая отзыв списка, абсолютное суждение и эксперименты с численной оценкой. Однако его автобиографическое эссе (Miller, 1989) показывает, что он никогда не относился серьезно к числу семь; это был риторический прием, который он использовал, чтобы связать воедино не связанные друг с другом направления своего исследования для выступления.Хотя верно, что объем памяти у взрослых составляет примерно семь элементов, нет гарантии, что каждый элемент является отдельным объектом. Возможно, наиболее важным моментом статьи Миллера (1956) было то, что несколько элементов можно объединить в более крупную значимую единицу. Более поздние исследования показали, что предел мощности, как правило, составляет всего три или четыре единицы (Broadbent, 1975; Cowan, 2001). Этот вывод был основан на попытке принять во внимание стратегии, которые часто повышают эффективность использования ограниченной емкости или позволяют хранить дополнительную информацию отдельно от этой ограниченной емкости.Чтобы понять эти методы обсуждения пределов емкости, я еще раз упомяну три типа загрязнения. Это происходит из-за разбивки на части и использования долговременной памяти, из-за репетиций и из-за типов хранения без ограничений по емкости.
Преодоление загрязнения от фрагментов и использования долговременной памяти
Реакция участника на задачу немедленной памяти зависит от того, как информация, которую нужно вызвать, сгруппирована для формирования фрагментов из нескольких элементов (Miller, 1956). Поскольку обычно неясно, какие фрагменты использовались при отзыве, неясно, сколько фрагментов можно сохранить и действительно ли это количество фиксировано.Бродбент (Broadbent, 1975) предложил некоторые ситуации, в которых формирование блока из нескольких элементов не было фактором, и предположил на основе результатов таких процедур, что истинный предел емкости составляет три элемента (каждый из которых служит блоком из одного элемента). Например, хотя объем памяти часто составляет около семи элементов, ошибки делаются со списками из семи элементов, а предел безошибочности обычно составляет три элемента. Когда люди должны вспомнить элементы из категории долговременной памяти, например, штаты США, они делают это рывками, в среднем около трех элементов.Это как если бы ведро кратковременной памяти наполнялось из колодца долговременной памяти и должно быть освобождено, прежде чем оно будет заполнено заново. Коуэн (2001) отметил другие подобные ситуации, в которых невозможно сформировать блоки из нескольких пунктов. Например, в рабочем диапазоне памяти длинный список элементов представлен с непредсказуемой конечной точкой, что делает невозможным группирование. Когда список заканчивается, участник должен вспомнить определенное количество пунктов из конца списка. Обычно люди могут вспомнить три или четыре пункта из конца списка, хотя точное количество зависит от требований задачи (Bunting et al., 2006). Индивидуумы различаются по способностям, которые варьируются от двух до шести пунктов у взрослых (и меньше у детей), и индивидуальный предел способностей является сильным коррелятом когнитивных способностей.
Другой способ учесть роль формирования многоэлементных блоков — это настроить задачу таким образом, чтобы можно было наблюдать за блоками. Талвинг и Паткау (1962) изучали свободное запоминание списков слов с различными уровнями структуры, от случайных слов до хорошо сформированных английских предложений, с несколькими различными уровнями согласованности между ними.Фрагмент был определен как серия слов, воспроизводимых участником в том же порядке, в котором они были представлены. Было подсчитано, что при всех условиях участники запоминали в среднем от четырех до шести фрагментов. Cowan et al. (2004) попытались усовершенствовать этот метод, протестировав последовательное запоминание списков из восьми слов, которые состояли из четырех пар слов, которые ранее были связаны с различными уровнями обучения (0, 1, 2 или 4 предыдущих пары слово-слово ). Каждое слово, использованное в списке, было представлено равное количество раз (четыре, за исключением неисследованного контрольного условия), но различалось, сколько из этих представлений было в виде одиночных и сколько было в виде последовательной пары.Количество парных предшествующих экспозиций оставалось постоянным для четырех пар в списке. Математическая модель использовалась для оценки доли вызванных пар, которые можно отнести к усвоенной ассоциации (то есть к фрагменту из двух слов), в отличие от раздельного вспоминания двух слов в паре. Эта модель предполагала, что предел емкости составлял около 3,5 фрагментов в каждом условии обучения, но что отношение фрагментов из двух слов к фрагментам из одного слова увеличивалось в зависимости от количества предыдущих воздействий на пары в списке.
Преодоление загрязнения от репетиции
Проблема репетиции не полностью отделена от вопроса формирования фрагментов. В традиционной концепции репетиции (например, Baddeley, 1986) можно представить, что элементы скрыто артикулируются в представленном порядке в равномерном темпе. Однако есть еще одна возможность: репетиция предполагает использование артикуляционных процессов для того, чтобы разбить предметы на группы. Фактически, Cowan et al. (2006a) спросили участников эксперимента с размахом цифр, как они выполняли задание, и, безусловно, наиболее распространенным ответом среди взрослых было то, что они сгруппировали элементы; участники редко упоминали, что говорили сами себе.Тем не менее, очевидно, что подавление репетиции влияет на производительность.
Предположительно, ситуации, в которых задания нельзя отрепетировать, по большей части совпадают с ситуациями, в которых задания не могут быть сгруппированы. Например, Cowan et al. (2005) полагались на текущую процедуру запоминания, в которой элементы представлялись с быстрой скоростью 4 раза в секунду. При такой скорости репетировать предметы в том виде, в каком они представлены, невозможно. Вместо этого задача, вероятно, решается путем сохранения пассивного хранилища (сенсорной или фонологической памяти) и последующего переноса последних нескольких элементов из этого хранилища в хранилище, более ориентированное на внимание, во время отзыва.На самом деле, при высокой скорости представления в беговом диапазоне инструкции по репетиции предметов вредны, а не полезны для выступления (Hockey, 1973). Другой пример — память для списков, которые игнорировались во время их представления (Cowan et al., 1999). В этих случаях предел вместимости близок к трем или четырем пунктам, предложенным Бродбентом (1975) и Коуэном (2001).
Вполне возможно, что существует механизм краткосрочного хранения на основе речи, который в целом не зависит от механизма на основе фрагментов.С точки зрения популярной модели Баддели (2000), первая является фонологической петлей, а вторая — эпизодическим буфером. В терминах Коуэна (1988, 1995, 1999, 2005) первая является частью активированной памяти, которая может иметь ограничение по времени из-за распада, а вторая является центром внимания, который, как предполагается, имеет предел емкости блока. .
Чен и Коуэн (2005) показали, что ограничение по времени и предел емкости блока в краткосрочной памяти разделены. Они повторили процедуру Cowan et al.(2004), в которых пары слов иногда предъявлялись на тренировке, предшествующей тесту на запоминание списка. Они объединили списки, составленные из пар, как в этом исследовании. Однако теперь использовались как бесплатные, так и серийные задачи отзыва, а длина списка варьировалась. Для длинных списков и бесплатного отзыва ограничение объема блока определяло отзыв. Например, были вызваны списки из шести хорошо выученных пар, а также списки из шести непарных одиночных слов (т. Е. Были вызваны с одинаковыми пропорциями правильных слов). Для более коротких списков и серийных отзывов с жесткой оценкой, отзыв регулируется ограничением времени.Например, списки из четырех хорошо усвоенных пар не были вызваны почти так же хорошо, как списки из четырех непарных синглтонов, а только так же, как списки восьми непарных синглтонов. Для промежуточных условий казалось, что пределы емкости блока и ограничения времени действуют вместе, чтобы управлять отзывом. Возможно, механизм с ограниченным объемом хранит предметы, а механизм репетиции сохраняет некоторую память последовательного порядка для этих удерживаемых предметов. Пока не ясно, как эти ограничения работают вместе.
Преодоление загрязнения от типов хранилищ без ограничения емкости
Трудно продемонстрировать истинный предел емкости, связанный с вниманием, если, как я полагаю, существуют другие типы механизмов краткосрочной памяти, которые усложняют результаты.Общая емкость должна включать в себя блоки информации всех видов: например, информацию, полученную как от акустических, так и от визуальных стимулов, а также от вербальных и невербальных стимулов. В этом случае должна быть перекрестная интерференция между одним типом загрузки памяти и другим. Тем не менее, в литературе часто показано, что между схожими типами меморандумов, например, двумя визуальными массивами объектов или двумя акустически представленными списками слов, существует гораздо больше взаимовлияния, чем между двумя разнородными типами, такими как один визуальный массив и один вербальный список. .Cocchini et al. (2002) предположили, что разнородные списки практически не интерферируют. Если так, то это может служить аргументом против наличия общего междоменного хранилища краткосрочной памяти.
Мори и Коуэн (2004, 2005) подвергли этот вывод сомнению. Они представили визуальный набор цветных пятен для сравнения со вторым набором, который соответствовал первому или отличался от него цветом одного пятна. Перед первым массивом или сразу после него участники иногда слышали список цифр, которые затем должны были быть произнесены между двумя массивами.В условиях низкой загрузки список представлял собой их собственный семизначный телефонный номер, тогда как в условиях высокой загрузки это был случайный семизначный номер. Только последнее условие мешало производительности сравнения массивов, и то только в том случае, если список должен был читаться вслух между массивами. Это говорит о том, что получение семи случайных цифр способом, который также задействует репетиционные процессы, основывается на каком-то механизме краткосрочной памяти, который также необходим для визуальных массивов. Этот общий механизм может оказаться в центре внимания с его ограниченными возможностями.Однако, по-видимому, если список велся молча, а не читался вслух, это тихое обслуживание происходило без особого использования общего механизма хранения, основанного на внимании, поэтому производительность визуального массива не сильно пострадала.
Типы кратковременной памяти, вклад которой в напоминание может скрывать предел емкости, могут включать любые типы активированной памяти, выходящие за рамки фокуса внимания. В структуре моделирования, изображенной на, это может включать функции сенсорной памяти, а также семантические функции.Сперлинг (1960) классно проиллюстрировал разницу между неограниченной сенсорной памятью и категориальной памятью с ограниченными возможностями. Если за массивом символов последовала частичная реплика отчета вскоре после массива, можно было бы вызвать большинство символов в указанной строке. Если сигнал был задержан примерно на 1 с, большая часть сенсорной информации распадалась, и производительность была ограничена примерно четырьмя символами, независимо от размера массива. Основываясь на этом исследовании, ограничение в четыре символа можно рассматривать либо как ограничение емкости кратковременной памяти, либо как ограничение скорости, с которой информация может быть передана из сенсорной памяти в категориальную форму до того, как она распадется.Однако Darwin et al. (1972) провели аналогичный слуховой эксперимент и обнаружили предел в четыре пункта, хотя наблюдаемый период спада сенсорной памяти составлял около 4 секунд. Учитывая разительные различия между Сперлингом и Дарвином и соавт. в период времени, доступный для передачи информации в категориальную форму, общий предел из четырех элементов лучше всего рассматривать как ограничение емкости, а не как ограничение скорости.
Саулс и Коуэн (Saults and Cowan, 2007) протестировали эту концептуальную основу в серии экспериментов, в которых массивы были представлены в двух модальностях сразу или, в другой процедуре, один за другим.Визуальный массив цветных пятен был дополнен набором произносимых цифр в четырех отдельных громкоговорителях, каждый из которых последовательно назначен разному голосу для облегчения восприятия. В некоторых испытаниях участники знали, что они несут ответственность за обе модальности одновременно, тогда как в других испытаниях участники знали, что они несут ответственность только за визуальные или только за акустические стимулы. Они получили массив зондов, который был таким же, как предыдущий массив (или такой же, как одна модальность в этом предыдущем массиве), или отличался от предыдущего массива идентичностью одного стимула.Задача заключалась в том, чтобы определить, было ли изменение. Использование кросс-модальности хранилища с ограниченным объемом позволяет прогнозировать конкретный образец результатов. Он предсказывает, что производительность в любой модальности должна снизиться в условиях двойной модальности по сравнению с унимодальными условиями из-за нагрузки на хранилище кросс-модальности. Так получились результаты. Более того, если кросс-модальность, хранилище с ограниченной емкостью было единственным используемым типом хранилища, тогда сумма зрительных и слуховых возможностей в состоянии двойной модальности не должна быть больше, чем большая из двух унимодальных возможностей (что случилось с быть зрительной способностью).Причина в том, что магазин ограниченной емкости может содержать одинаковое количество единиц независимо от того, были ли они все из одной модальности или из двух вместе взятых. Это предсказание подтвердилось, но только в том случае, если в обеих модальностях сразу после массива, который нужно запомнить, существовала пост-перцептивная маска. Пост-перцептивная маска включала разноцветное пятно в каждом местоположении визуального объекта и звук, состоящий из всех возможных цифр, наложенных из каждого громкоговорителя. Он был представлен достаточно долго после массивов, чтобы их можно было вспомнить, чтобы их восприятие было полным (например,г., через 1 с; ср. Vogel et al., 2006). Предположительно, маска была способна перезаписывать различные типы сенсорных функций в активированной памяти, оставляя после себя только более общую, категориальную информацию, присутствующую в фокусе внимания, которая предположительно защищена от маскирующего вмешательства процессом внимания. Снова было показано, что предел фокуса внимания составляет от трех до четырех пунктов для одномодальных зрительных или бимодальных стимулов.
Даже без использования маскирующих стимулов можно найти фазу процесса кратковременной памяти, которая является общей для разных областей.Cowan и Morey (2007) представили два набора стимулов, которые следует вспомнить (или, в контрольных условиях, только один набор). Два набора стимулов могут включать в себя два речевых списка цифр, два пространственных массива цветных пятен или по одному каждого в любом порядке. После этой презентации сигнал показал, что участник будет нести ответственность только за первый массив, только за второй массив или за оба массива. До зондирования следовало три секунды. Эффект от загрузки памяти можно сравнить двумя способами. Эффективность в тех испытаниях, в которых были представлены два набора стимулов и оба были запрошены для удержания, можно было сравнить либо с испытаниями, в которых был представлен только один набор, либо его можно было сравнить с испытаниями, в которых были представлены оба набора стимулов, но сигнал позже указывался. что нужно было сохранить только один набор.Часть рабочей памяти, предшествующая сигналу, показывала специфичные для модальности эффекты двойной задачи: кодирование набора стимулов одного типа было вреднее, если также кодировать другой набор, если оба набора были в одной и той же модальности. Однако сохранение информации после сигнала показало эффекты двойной задачи, не зависящие от модальности. Когда были представлены два набора, сохранение их обоих было вредным по сравнению с сохранением только одного набора (как указано в сигнале удержания после стимула, чтобы сохранить один набор по сравнению с обоими наборами), и этот эффект двойной задачи был одинаковым по величине независимо от того, наборы были в одинаковых или разных модальностях.Таким образом, после первоначального кодирования хранение рабочей памяти в течение нескольких секунд может происходить абстрактно, в фокусе внимания.
Другое свидетельство в пользу отдельного краткосрочного хранилища
Наконец, есть другие свидетельства, которые напрямую не подтверждают ни временное затухание, ни ограничение емкости, но подразумевают, что существует тот или иной из этих ограничений. Бьорк и Уиттен (1974) и Ценг (1973) выдвинули аргументы временной различимости на основе того, что называется непрерывным воспроизведением списка отвлекающих факторов, при котором эффект новизны сохраняется даже тогда, когда за списком следует заполненная отвлекающими факторами задержка перед отзывом.Заполненная задержка должна была разрушить кратковременную память, но эффект новизны все равно имеет место, при условии, что элементы в списке также разделены задержками, заполненными отвлекающими факторами, чтобы увеличить их различимость друг от друга. В пользу краткосрочного запоминания, однако, другие исследования показали диссоциацию между тем, что обнаруживается при обычном немедленном вспоминании и постоянном отвлекающем воспоминании (например, эффекты длины слова, обращенные вспять в постоянном отвлекающем воспоминании: Cowan et al., 1997b; проактивное вмешательство в самые последние позиции в списках, которые постоянно вспоминаются только дистракторами: Craik & Birtwistle, 1971; Davelaar et al., 2005).
Есть также дополнительные данные нейровизуализации для краткосрочного хранения. Talmi et al. (2005) обнаружили, что распознавание более ранних частей списка, но не нескольких последних элементов, активировало области в системе гиппокампа, что обычно связано с долгосрочным извлечением памяти. Это согласуется с упомянутым ранее выводом о том, что память для нескольких последних пунктов списка сохраняется при амнезии Корсакова (Baddeley and Warrington, 1970; Carlesimo et al., 1995). В этих исследованиях часть эффекта новизны, основанная на кратковременной памяти, может отражать короткий промежуток времени между презентацией и воспроизведением нескольких последних элементов или может отражать отсутствие интерференции между презентацией и воспроизведением нескольких последних элементов. .Таким образом, мы можем сказать, что кратковременная память существует, но часто без особой ясности относительно того, является ли ограничение ограничением по времени или пределом емкости блока.
Различие между кратковременной памятью и рабочей памятью
Различие между кратковременной памятью и рабочей памятью затуманено некоторой путаницей, но это в значительной степени результат того, что разные исследователи использовали разные определения. Miller et al. (1960) использовали термин «рабочая память» для обозначения временной памяти с функциональной точки зрения, поэтому с их точки зрения нет четкого различия между кратковременной и рабочей памятью.Баддели и Хитч (1974) вполне соответствовали этому определению, но наложили некоторые описания на термины, которые их отличали. Они рассматривали кратковременную память как единое место хранения, как это описано, например, Аткинсоном и Шиффрином (1968). Когда они поняли, что доказательства на самом деле соответствуют многокомпонентной системе, которую нельзя свести к унитарному краткосрочному хранилищу, они использовали термин рабочая память для описания всей системы. Коуэн (1988) придерживался многокомпонентного взгляда, как Бэдделли и Хитч, но не уделял точного внимания их компонентам; вместо этого, основными подразделениями рабочей памяти были названы компоненты краткосрочного хранения (активированная память вместе с фокусом внимания внутри нее, как показано на рисунке) и центральные исполнительные процессы, которые манипулируют хранимой информацией.По мнению Коуэна, фонологическая петля и зрительно-пространственный блокнот Баддели (1986) могут рассматриваться как всего лишь два из многих аспектов активированной памяти, которые подвержены помехам в степени, которая зависит от сходства между характеристиками активированных и мешающих источников информации. Эпизодический буфер Баддели (2000), возможно, совпадает с информацией, хранящейся в фокусе внимания Коуэна, или, по крайней мере, представляет собой очень похожую концепцию.
Произошел некоторый сдвиг в определении или описании рабочей памяти наряду с сдвигом в объяснении того, почему новые задачи рабочей памяти коррелируют с интеллектом и мерами способностей намного выше, чем простые, традиционные задачи краткосрочной памяти. например, серийный отзыв.Данеман и Карпентер (1980) предположили, что критически важно использовать задачи рабочей памяти, которые включают в себя как компоненты хранения, так и обработки, чтобы задействовать все части рабочей памяти, как описано, например, Баддели и Хитч (1974). . Вместо этого Энгл и др. (1999) и Кейн и др. (2001) предположили, что критичным является то, является ли задача рабочей памяти сложной с точки зрения контроля внимания. Например, Kane et al. обнаружили, что задачи по хранению и обработке рабочей памяти хорошо коррелируют со способностью подавлять естественную тенденцию смотреть на внезапно появляющийся стимул и вместо этого смотреть в другую сторону, задача антисаккада.Аналогичным образом Conway et al. (2001) обнаружили, что люди, получившие высокие баллы по тестам на хранение и обработку рабочей памяти, замечают свои имена в канале, который следует игнорировать при дихотическом слушании, гораздо чаще, чем реже , чем люди с малым интервалом; Люди с большим размахом, по-видимому, лучше способны сделать выполнение своей основной задачи менее уязвимым для отвлечения внимания, но это происходит за счет того, что они немного не обращают внимания на несущественные аспекты своего окружения. В ответ на такое исследование Энгл и его коллеги иногда использовали термин рабочая память для обозначения только процессов, связанных с контролем внимания.Таким образом, их определение рабочей памяти, кажется, расходится с предыдущими определениями, но это новое определение допускает простое утверждение, что рабочая память сильно коррелирует со способностями, тогда как краткосрочная память (переопределенная, чтобы включать только аспекты памяти, не связанные с вниманием. хранение) не так сильно коррелирует со способностями.
Cowan et al. (2006b), придерживаясь более традиционного определения рабочей памяти, сделали утверждение о рабочей памяти, подобное утверждению Энгла и его коллег, но немного более сложное.Они предположили, на основе некоторых данных о развитии и корреляции, что множественные функции внимания имеют отношение к индивидуальным различиям в способностях. Контроль внимания имеет значение, но есть независимый вклад от количества элементов, которые можно удерживать во внимании, или его объема. Согласно этой точке зрения, что может быть необходимо для того, чтобы процедура рабочей памяти хорошо коррелировала с когнитивными способностями, так это то, что задача должна предотвращать скрытые словесные репетиции, так что участник должен полагаться на более требовательную к вниманию обработку и / или память для выполнения задачи. .Cowan et al. (2005) обнаружили, что задача может быть намного проще, чем процедуры хранения и обработки. Например, в версии текущего теста объема памяти цифры отображаются очень быстро, и последовательность останавливается в непредсказуемой точке, после чего участник должен вызвать как можно больше элементов из конца списка. Репетиция невозможна, и, когда список заканчивается, информация, по-видимому, должна быть извлечена из активированных сенсорных или фонологических функций в центр внимания.Этот тип задач коррелировал со способностями, как и некоторые другие меры объема внимания (Cowan et al., 2005, 2006b). У детей, слишком маленьких для того, чтобы использовать скрытую словесную репетицию (в отличие от детей старшего возраста и взрослых), даже простая задача по размаху цифр служила отличным коррелятом со способностями.
Другое исследование подтверждает эту идею о том, что тест на рабочую память будет хорошо коррелировать с когнитивными способностями в той степени, в которой он требует, чтобы внимание использовалось для хранения и / или обработки.Гавенс и Барруйе (2004) провели исследование развития, в котором они контролировали сложность и продолжительность задачи обработки, которая возникала между элементами, которые нужно было вспомнить. По-прежнему существовала разница в продолжительности развития, которую они приписывали развитию основных способностей, что могло отражать увеличение объема внимания в процессе развития (см. Cowan et al., 2005). Lépine et al. (2005) показали, что для того, чтобы связанная задача типа хранения и обработки хорошо коррелировала со способностями, было то, чтобы компонент обработки задачи (в данном случае чтение букв вслух) выполнялся достаточно быстро, чтобы предотвратить различные типы репетиция, чтобы прокрасться между ними (см. также Conlin et al., 2005).
В нескольких статьях хранение и обработка (возможно, объем или контроль внимания?) Противопоставлялись друг другу, чтобы понять, что более важно для учета индивидуальных различий. Vogel et al. (2005) использовали задачу визуального массива, модифицированную для использования с компонентом связанных с событием потенциалов, который указывает хранение в визуальной рабочей памяти, называемой контралатеральной задерживающей активностью (CDA). Было обнаружено, что это действие зависит не только от количества соответствующих объектов на дисплее (например,g., красные полосы под разными углами, которые нужно запомнить), но иногда также количество нерелевантных объектов, которые следует игнорировать (например, синие полосы). Для людей с большим размахом было обнаружено, что АКД для двух релевантных объектов одинакова независимо от того, присутствовали ли также два нерелевантных объекта на дисплее. Однако для людей с малым охватом CDA для двух релевантных объектов в сочетании с двумя нерелевантными объектами был аналогичен CDA для дисплеев только с четырьмя релевантными объектами, как если бы нерелевантные объекты нельзя было исключить из рабочей памяти.Одним из ограничений исследования является то, что разделение участников на высокий и низкий диапазон также основывалось на CDA, и задача, используемая для измерения CDA, неизбежно требовала выборочного внимания (к половине дисплея) в каждом испытании, независимо от того, в него входили предметы неактуального цвета.
Gold et al. (2006) исследовали аналогичные проблемы в поведенческом дизайне и проверяли разницу между пациентами с шизофренией и нормальными участниками контрольной группы. Каждое испытание начиналось с того, что нужно было уделить внимание одной части демонстрации за счет другой (например,g., полосы одного актуального цвета, но не другого, нерелевантного цвета). Отображение датчика представляло собой набор, который соответствовал значению в большинстве испытаний (в некоторых экспериментах, 75%), тогда как иногда отображение датчика было набором, на который не подавалось указание. Это позволило по отдельности измерить контроль внимания (преимущество для элементов с указанием очереди по сравнению с элементами без привязки) и емкость рабочей памяти (среднее количество элементов, отозванных из каждого массива, сложение по наборам с отправкой и без привязки). В отличие от первоначальных ожиданий, очевидный результат заключался в том, что разница между группами заключалась в способности, а не в контроле внимания.Было бы интересно узнать, можно ли получить один и тот же тип результата для нормальных людей с высоким или низким размахом, или же это сравнение вместо этого покажет разницу в контроле внимания между этими группами, как Vogel et al. (2005) должен предсказывать. Friedman et al. (2006) обнаружили, что не все центральные исполнительные функции коррелируют со способностями; обновление рабочей памяти имело место, но торможение и переключение внимания — нет. С другой стороны, напомним, что Cowan et al. (2006b) обнаружили, что задача контроля внимания связана со способностями.
В общем, вопрос о том, различаются ли кратковременная память и рабочая память, может быть вопросом семантики. Есть очевидные различия между простыми задачами последовательного воспроизведения, которые не очень хорошо коррелируют с тестами на способности у взрослых, и другими задачами, требующими памяти и обработки или памяти без возможности репетиции, которые гораздо лучше коррелируют со способностями. Использовать ли термин «рабочая память» для последнего набора задач или зарезервировать этот термин для всей системы сохранения и управления кратковременной памятью — дело вкуса.Более важный и существенный вопрос может заключаться в том, почему одни задачи гораздо лучше коррелируют со способностями, чем другие.
Заключение
Различие между долговременной и кратковременной памятью зависит от того, можно ли продемонстрировать наличие свойств, специфичных для кратковременной памяти; основные кандидаты включают временное затухание и ограничение емкости блока. Вопрос о распаде по-прежнему остается открытым для обсуждения, в то время как ограничение емкости блоков данных получает все большую поддержку. Эти ограничения обсуждались в рамках, показанных в.
Различие между кратковременной памятью и рабочей памятью зависит от принятого определения. Тем не менее, главный вопрос заключается в том, почему одни тесты памяти на короткий срок служат одними из лучших коррелятов когнитивных способностей, а другие — нет. Ответ, кажется, указывает на важность системы внимания, используемой как для обработки, так и для хранения. Эффективность этой системы и ее использование в рабочей памяти, по-видимому, существенно различаются у разных людей (например,г., Conway et al., 2002; Кейн и др., 2004; Cowan et al., 2005, 2006b), а также улучшается по мере развития в детстве (Cowan et al., 2005, 2006b) и снижается в старости (Naveh-Benjamin et al., 2007; Stoltzfus et al., 1996; Cowan et al., 2006c).
Благодарность
Эта работа была завершена при поддержке NIH Grant R01 HD-21338.
Ссылки
- Аткинсон Р.К., Шиффрин Р.М. Память человека: предлагаемая система и процессы управления ею. В: Спенс К.В., Спенс Дж. Т., редакторы.Психология обучения и мотивации: достижения в области исследований и теории. Vol. 2. Нью-Йорк: Academic Press; 1968. С. 89–195. [Google Scholar]
- Баддели А. Эпизодический буфер: новый компонент рабочей памяти? Trends Cogn. Sci. 2000. 4: 417–423. [PubMed] [Google Scholar]
- Baddeley AD. Oxford Psychology Series No. 11. Оксфорд: Clarendon Press; 1986. Рабочая память. [Google Scholar]
- Баддели А.Д., Хитч Г. Рабочая память. В: Бауэр Г.Х., редактор. Психология обучения и мотивации.Vol. 8. Нью-Йорк: Academic Press; 1974. С. 47–89. [Google Scholar]
- Баддели А.Д., Скотт Д. Кратковременное забывание при отсутствии упреждающего торможения. Q. J. Exp. Psychol. 1971; 23: 275–283. [Google Scholar]
- Баддели А.Д., Томсон Н., Бьюкенен М. Длина слова и структура кратковременной памяти. J. Словесное обучение. Вербальное поведение. 1975. 14: 575–589. [Google Scholar]
- Baddeley AD, Warrington EK. Амнезия и различие между долговременной и кратковременной памятью. J. Словесное обучение.Вербальное поведение. 1970; 9: 176–189. [Google Scholar]
- Барруйе П., Бернардин С., Камос В. Временные ограничения и совместное использование ресурсов в пределах рабочей памяти взрослых. J. Exp. Psychol .: Gen. 2004; 133: 83–100. [PubMed] [Google Scholar]
- Барруйе П., Бернардин С., Портрат С., Вергаув Э., Камос В. Время и когнитивная нагрузка на рабочую память. J. Exp. Psychol. Учить. Mem. Cogn. 2007; 33: 570–585. [PubMed] [Google Scholar]
- Бьорк Р.А., Уиттен ВБ. Чувствительные к давности процессы поиска в долгосрочном бесплатном отзыве.Cogn. Psychol. 1974. 6: 173–189. [Google Scholar]
- Broadbent DE. Восприятие и общение. Нью-Йорк: Pergamon Press; 1958. [Google Scholar]
- Broadbent DE. Магическое число семь через пятнадцать лет. В: Кеннеди А., Уилкс А., редакторы. Исследования долговременной памяти. Оксфорд, Англия: Wiley; 1975. С. 3–18. [Google Scholar]
- Brown GDA, Preece T, Hulme C. Память на основе осциллятора для последовательного заказа. Psychol. Rev.2000; 107: 127–181. [PubMed] [Google Scholar]
- Бантинг М.Ф., Коуэн Н., Саултс Дж. С.Как работает рабочий диапазон памяти? Q. J. Exp. Psychol. 2006; 59: 1691–1700. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Карлезимо Г.А., Саббадини М., Фадда Л., Кальтаджироне С. Различные компоненты словесного забвения чистой амнезии, дегенеративного слабоумия и здоровых субъектов. Cortex. 1995; 31: 735–745. [PubMed] [Google Scholar]
- Чен З, Коуэн Н. Пределы чанка и ограничения длины при немедленном отзыве: согласование. J. Exp. Psychol. Учить. Mem. Cogn. 2005; 31: 1235–1249. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Cocchini G, Logie RH, Della Sala S, MacPherson SE, Baddeley AD.Одновременное выполнение двух задач памяти: свидетельство для систем рабочей памяти, специфичных для предметной области. Mem. Cogn. 2002; 30: 1086–1095. [PubMed] [Google Scholar]
- Конлин Дж. А., Gathercole SE, Адамс Дж. У. Детская рабочая память: исследование ограничений производительности при выполнении сложных задач. J. Exp. Child Psychol. 2005; 90: 303–317. [PubMed] [Google Scholar]
- Конвей А.Р., Коуэн Н., Бантинг М.Ф. Вернемся к феномену коктейльной вечеринки: важность объема рабочей памяти. Психон. Бык.Ред. 2001; 8: 331–335. [PubMed] [Google Scholar]
- Конвей А.Р., Коуэн Н., Бантинг М.Ф., Террио Д.Д., Минкофф С.Р. Скрытый переменный анализ объема рабочей памяти, объема краткосрочной памяти, скорости обработки и общего гибкого интеллекта. Интеллект. 2002. 30: 163–183. [Google Scholar]
- Конвей ARA, Кейн MJ, Бантинг MF, Hambrick DZ, Wilhelm O, Engle RW. Задачи по объему рабочей памяти: методический обзор и руководство пользователя. Психон. Бык. Ред. 2005; 12: 769–786. [PubMed] [Google Scholar]
- Cowan N.Развитие представлений о хранении в памяти, избирательном внимании и их взаимных ограничениях в системе обработки информации человеком. Psychol. Бык. 1988. 104: 163–191. [PubMed] [Google Scholar]
- Коуэн Н. Объем вербальной памяти и время речевого отзыва. J. Mem. Lang. 1992; 31: 668–684. [Google Scholar]
- Cowan N. Oxford Psychology Series No. 26. Нью-Йорк: Oxford University Press; 1995. Внимание и память: интегрированные рамки. [Google Scholar]
- Cowan N.Модель встроенных процессов рабочей памяти. В: Мияке А., Шах П., редакторы. Модели рабочей памяти: механизмы активного обслуживания и исполнительного контроля. Кембридж, Великобритания: Издательство Кембриджского университета; 1999. С. 62–101. [Google Scholar]
- Коуэн Н. Магическое число 4 в кратковременной памяти: переосмысление способности умственной памяти. Behav. Brain Sci. 2001. 24: 87–185. [PubMed] [Google Scholar]
- Коуэн Н. Объем оперативной памяти. Хоув, Восточный Суссекс, Великобритания: Psychology Press; 2005 г.[Google Scholar]
- Cowan N, Aubuchon AM. Психон. Бык. Rev. Кратковременная потеря памяти с течением времени без вмешательства ретроактивных стимулов. (в печати) [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Cowan N, Chen Z, Rouder JN. Постоянная способность к немедленному выполнению задачи последовательного отзыва: логическое продолжение книги Миллера (1956) Psychol. Sci. 2004. 15: 634–640. [PubMed] [Google Scholar]
- Cowan N, Elliott EM, Saults JS, Morey CC, Mattox S, Hismjatullina A, Conway ARA. О способности внимания: его оценка и его роль в рабочей памяти и когнитивных способностях.Cogn. Psychol. 2005; 51: 42–100. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Cowan N, Elliott EM, Saults JS, Nugent LD, Bomb P, Hismjatullina A. Переосмысление скоростных теорий когнитивного развития: увеличение скорости запоминания без снижения точности. Psychol. Sci. 2006a; 17: 67–73. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Cowan N, Fristoe NM, Elliott EM, Brunner RP, Saults JS. Объем внимания, контроль внимания и интеллект у детей и взрослых. Mem. Cogn.2006b; 34: 1754–1768. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Cowan N, Morey CC. Как можно исследовать пределы удержания оперативной памяти при выполнении двух задач? Psychol. Sci. 2007. 18: 686–688. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Cowan N, Naveh-Benjamin M, Kilb A, Saults JS. Развитие визуальной рабочей памяти на протяжении всей жизни: когда сложно привязать функции? Dev. Psychol. 2006c; 42: 1089–1102. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Cowan N, Nugent LD, Elliott EM, Ponomarev I, Saults JS.Роль внимания в развитии кратковременной памяти: возрастные различия вербальной продолжительности восприятия. Child Dev. 1999; 70: 1082–1097. [PubMed] [Google Scholar]
- Cowan N, Saults JS, Nugent LD. Роль абсолютного и относительного количества времени в забывании в пределах непосредственной памяти: случай сравнения высоты тона. Психон. Бык. Ред. 1997a; 4: 393–397. [Google Scholar]
- Коуэн Н., Вуд Н.Л., Ньюджент Л.Д., Трейсман М. В словесной кратковременной памяти есть два эффекта длины слова: противоположные эффекты длительности и сложности.Psychol. Sci. 1997b; 8: 290–295. [Google Scholar]
- Craik FIM, Birtwistle J. Упреждающее торможение при свободном отзыве. J. Exp. Psychol. 1971; 91: 120–123. [Google Scholar]
- Crowder RG. Исчезновение кратковременной памяти. Acta Psychol. 1982; 50: 291–323. [PubMed] [Google Scholar]
- Crowder RG. Кратковременная память: где мы находимся? Mem. Cogn. 1993; 21: 142–145. [PubMed] [Google Scholar]
- Daneman M, Carpenter PA. Индивидуальные различия в рабочей памяти и чтении. J словесное обучение.Вербальное поведение. 1980; 19: 450–466. [Google Scholar]
- Daneman M, Merikle PM. Рабочая память и понимание языка: метаанализ. Психон. Бык. Ред. 1996; 3: 422–433. [PubMed] [Google Scholar]
- Дарвин С.Дж., Терви М.Т., Краудер Р.Г. Слуховой аналог процедуры частичного отчета Сперлинга: свидетельство для краткого слухового хранения. Cogn. Psychol. 1972; 3: 255–267. [Google Scholar]
- Давелаар Э.Дж., Гошен-Готтштейн Ю., Ашкенази А., Хаарман Х.Дж., Ашер М. Возвращение к исчезновению кратковременной памяти: эмпирические и вычислительные исследования эффектов недавности.Psychol. Ред. 2005; 112: 3–42. [PubMed] [Google Scholar]
- Эббингаус Х. Перевод Х.А. Ругера и К.Е. Буссениуса. Нью-Йорк: педагогический колледж Колумбийского университета; 18851913. Память: вклад в экспериментальную психологию. (Первоначально на немецком языке: Ueber das gedächtnis: Untersuchen zur Experimentellen Psychologie) [Google Scholar]
- Engle RW. Объем рабочей памяти как исполнительное внимание. Curr. Реж. Psychol. Sci. 2002; 11: 19–23. [Google Scholar]
- Engle RW, Tuholski SW, Laughlin JE, Conway ARA.Рабочая память, кратковременная память и общий гибкий интеллект: подход с латентной переменной. J. Exp. Psychol. Gen.1999; 128: 309–331. [PubMed] [Google Scholar]
- Эрикссон К.А., Кинч В. Долговременная рабочая память. Psychol. Ред. 1995; 102: 211–245. [PubMed] [Google Scholar]
- Фридман Н. П., Мияке А., Корли Р. П., Янг С. Е., Де Фрис Дж. К., Хьюитт Дж. К. Не все исполнительные функции связаны с интеллектом. Psychol. Sci. 2006. 17: 172–179. [PubMed] [Google Scholar]
- Гавенс Н., Барруйе П.Задержки удержания, эффективности обработки и ресурсов внимания при развитии рабочей памяти. J. Mem. Lang. 2004. 51: 644–657. [Google Scholar]
- Glanzer M, Cunitz AR. Два механизма хранения в свободном отзыве. J. Словесное обучение. Вербальное поведение. 1966; 5: 351–360. [Google Scholar]
- Glenberg AM, Swanson NC. Теория временной различимости эффектов новизны и модальности. J. Exp. Psychol. Учить. Mem. Cogn. 1986; 12: 3–15. [PubMed] [Google Scholar]
- Gold JM, Fuller RL, Robinson BM, McMahon RP, Braun EL, Luck SJ.Неповрежденный контроль внимания за кодированием рабочей памяти при шизофрении. J. Abnorm. Psychol. 2006. 115: 658–673. [PubMed] [Google Scholar]
- Грин Р.Л. Влияние экспериментального дизайна: пример парадигмы Брауна-Петерсона. Может. J. Exp. Psychol. 1996. 50: 240–242. [Google Scholar]
- Guttentag RE. Требование умственных усилий кумулятивной репетиции: исследование развития. J. Exp. Child Psychol. 1984. 37: 92–106. [Google Scholar]
- Hebb DO. Организация поведения.Нью-Йорк: Уайли; 1949. [Google Scholar]
- Hockey R. Скорость презентации в оперативной памяти и прямое управление стратегиями обработки ввода. Q. J. Exp. Psychol. А. 1973; 25: 104–111. [Google Scholar]
- Джеймс У. Принципы психологии. Нью-Йорк: Генри Холт; 1890. [Google Scholar]
- Кейн MJ, Bleckley MK, Conway ARA, Engle RW. Просмотр объема рабочей памяти с контролируемым вниманием. J. Exp. Psychol. Gen. 2001; 130: 169–183. [PubMed] [Google Scholar]
- Кейн MJ, Hambrick DZ, Tuholski SW, Wilhelm O, Payne TW, Engle RE.Общая емкость рабочей памяти: латентно-переменный подход к вербальной и зрительно-пространственной памяти и рассуждениям. J. Exp. Psychol. Gen. 2004; 133: 189–217. [PubMed] [Google Scholar]
- Кеппель Дж., Андервуд Б.Дж. Упреждающее запрещение краткосрочного хранения отдельных предметов. J. Словесное обучение. Вербальное поведение. 1962; 1: 153–161. [Google Scholar]
- Kyllonen PC, Christal RE. Разумная способность — это (чуть больше) объем рабочей памяти? Интеллект. 1990; 14: 389–433. [Google Scholar]
- Lépine R, Barrouillet P, Camos V.Что делает диапазон рабочей памяти таким предсказательным для познания высокого уровня? Психон. Бык. Ред. 2005; 12: 165–170. [PubMed] [Google Scholar]
- Левандовски С., Дункан М., Браун GDA. Время не вызывает забвения в краткосрочных серийных воспоминаниях. Психон. Бык. Ред. 2004; 11: 771–790. [PubMed] [Google Scholar]
- McGeoch JA. Забвение и закон неиспользования. Psychol. Rev.1932; 39: 352–370. [Google Scholar]
- Melton AW. Значение кратковременной памяти для общей теории памяти.J. Словесное обучение. Вербальное поведение. 1963; 2: 1-21. [Google Scholar]
- Miller GA. Магическое число семь, плюс-минус два: некоторые ограничения нашей способности обрабатывать информацию. Psychol. Rev.1956; 63: 81–97. [PubMed] [Google Scholar]
- Миллер GA. Джордж А. Миллер. В: Линдзей Г., редактор. История психологии в автобиографии. Vol. VIII. Стэнфорд, Калифорния: Издательство Стэнфордского университета; 1989. С. 391–418. [Google Scholar]
- Миллер Г.А., Галантер Э., Прибрам К.Х. Планы и структура поведения.Нью-Йорк: Холт, Райнхарт и Уинстон, Инк; 1960. [Google Scholar]
- Мори С.К., Коуэн Н. Когда визуальная и вербальная память конкурируют: свидетельство междоменных ограничений в рабочей памяти. Психон. Бык. Ред. 2004; 11: 296–301. [PubMed] [Google Scholar]
- Мори С.К., Коуэн Н. Когда возникают противоречия между визуальными и вербальными воспоминаниями? Важность загрузки и извлечения рабочей памяти. J. Exp. Psychol. Учить. Mem. Cogn. 2005. 31: 703–713. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Nairne JS.Кратковременные воспоминания: аргументы против стандартной модели. Анну. Rev. Psychol. 2002; 53: 53–81. [PubMed] [Google Scholar]
- Навех-Бенджамин М., Коуэн Н., Килб А., Чен З. Возрастные различия в немедленном серийном воспроизведении: формирование диссоциированных фрагментов и емкость. Mem. Cognit. 2007. 35: 724–737. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Neath I, Surprenant A. Человеческая память. 2-е изд. Бельмонт, Калифорния: Уодсворт; 2003. [Google Scholar]
- Nipher FE. О распределении ошибок по числам, записанным по памяти.Пер. Акад. Sci. Святой Луи. 1878; 3: ccx – ccxi. [Google Scholar]
- Петерсон Л.Р., Петерсон М.Дж. Кратковременное удержание отдельных словесных заданий. J. Exp. Psychol. 1959; 58: 193–198. [PubMed] [Google Scholar]
- Саултс Дж. С., Коуэн Н. Центральное ограничение емкости для одновременного хранения визуальных и слуховых массивов в рабочей памяти. J. Exp. Psychol. 2007. 136: 663–684. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Sperling G. Информация доступна в виде кратких наглядных презентаций.Psychol. Monogr. 1960; 74 (Целый № 498) [Google Scholar]
- Штольцфус Э. Р., Хашер Л., Закс РТ. Рабочая память и поиск: подход к ресурсам торможения. В: Richardson JTE, Engle RW, Hasher L, Logie RH, Stoltzfus ER, Zacks RT, редакторы. Рабочая память и человеческое познание. Нью-Йорк: издательство Оксфордского университета; 1996. С. 66–88. [Google Scholar]
- Talmi D, Grady CL, Goshen-Gottstein Y, Moscovitch M. Нейровизуализация кривой последовательного положения: тест моделей с одним магазином по сравнению с моделями с двумя магазинами.Psychol. Sci. 2005; 16: 716–723. [PubMed] [Google Scholar]
- Tulving E, Patkau JE. Сопутствующие эффекты контекстного ограничения и частоты слов на немедленное запоминание и усвоение вербального материала. Может. J. Psychol. 1962; 16: 83–95. [PubMed] [Google Scholar]
- Турви М.Т., Брик П., Осборн Дж. Упреждающее вмешательство в кратковременную память в зависимости от интервала сохранения предыдущих элементов. Q. J. Exp. Psychol. 1970; 22: 142–147. [Google Scholar]
- Tzeng OJL. Положительный эффект новизны при отложенном бесплатном отзыве.J. Словесное обучение. Вербальное поведение. 1973; 12: 436–439. [Google Scholar]
- Ансуорт Н., Энгл Р. У. Характер индивидуальных различий в объеме рабочей памяти: активное ведение в первичной памяти и управляемый поиск из вторичной памяти. Psychol. Ред. 2007; 114: 104–132. [PubMed] [Google Scholar]
- Фогель Е.К., Макколлоу А.В., Мачизава М.Г. Нейронные измерения выявляют индивидуальные различия в управлении доступом к рабочей памяти. Природа. 2005; 438: 500–503. [PubMed] [Google Scholar]
- Фогель Е.К., Вудман Г.Ф., Luck SJ.Временной ход закрепления в зрительной рабочей памяти. J. Exp. Psychol. Гм. Восприятие. Выполнять. 2006; 32: 1436–1451. [PubMed] [Google Scholar]
- Во, Северная Каролина, Норман Д.А. Первичная память. Psychol. Rev.1965; 72: 89–104. [PubMed] [Google Scholar]
- Викельгрен, Вашингтон. Теория однократной хрупкости динамики памяти. Mem. Cogn. 1974; 2: 775–780. [PubMed] [Google Scholar]
Долговременная память — BrainHQ от Posit Science
Долговременная память — это все, что вы помните, что произошло более нескольких минут назад.Долговременные воспоминания могут длиться всего несколько дней или многие годы.
Долговременные воспоминания не одинаковы по силе. Более сильные воспоминания позволяют вам вспомнить событие, процедуру или факт по запросу — например, что Париж — столица Франции. Более слабые воспоминания часто приходят на ум только через подсказки или напоминания.
Долговременная память тоже не статична. Вы не запечатлеваете память и не оставляете ее как бы нетронутой. Вместо этого вы часто пересматриваете воспоминание с течением времени — возможно, объединяя его с другим воспоминанием или объединяя то, что другие говорят вам о воспоминании.В результате ваши воспоминания не являются строго постоянными и не всегда надежными.
Есть много разных форм долговременной памяти. Эти воспоминания не формируются и не сохраняются в одной части мозга; вместо этого процесс создания и хранения долговременных воспоминаний распространяется на несколько регионов. Два основных подразделения — это явная память и неявная память. Явные воспоминания — это воспоминания, которые вы помните сознательно, например, какое-то событие из вашей жизни или конкретный факт.Неявные воспоминания — это те воспоминания, о которых вы не задумываетесь, например, езда на велосипеде — вы когда-то научились и запомнили, как, но теперь делаете это без сознательной мысли. Хотя понимание этих различий в типах памяти, которые мы храним в долгосрочной перспективе, полезно, разделения плавны: разные формы памяти часто смешиваются и смешиваются. Чтобы узнать больше о различных типах долговременной памяти, просмотрите наши страницы явной (декларативной) и неявной (недекларативной) памяти.
Насколько хорошо вы что-то запоминаете, отчасти зависит от того, насколько быстро и четко ваши чувства воспринимают происходящее.Если ваш мозг с идеальной точностью записывает то, что вы видите, слышите, ощущаете, вкус и запах, он может лучше вспомнить их позже. У многих людей с плохой памятью основная проблема заключается в способности мозга четко записывать сенсорную информацию, а не в его способности «запоминать».
Вот почему, когда дело доходит до улучшения памяти, важно ускорить и отточить способность мозга обрабатывать то, что вы воспринимаете через органы чувств. Видение и слушание являются наиболее важными, поскольку во многих воспоминаниях то, что вы видите и слышите, составляет большую часть памяти.Некоторые упражнения в BrainHQ специально разработаны для улучшения слуховой и визуальной обработки, чтобы ваш мозг мог воспринимать то, что вы видите и слышите, со скоростью и точностью, необходимой для формирования сильной памяти. Если вы хотите улучшить свою память, улучшив работу мозга, попробуйте BrainHQ.
Улучшите вашу долговременную память с CogniFit.
Что такое долговременная память?
Долговременная память может быть определена как механизм мозга , который позволяет кодировать и сохранять почти неограниченный объем информации в течение длительного периода времени .Воспоминания, хранящиеся в долговременной памяти, могут сохраняться до нескольких лет.
Долговременная память — ключевой элемент для успешного самостоятельного выполнения повседневных дел. Этот тип памяти относится к способности мозга сохранять опыт, события, концепции или навыки и вспоминать их позже. Долговременная память — это сложный навык, который имеет много разных аспектов и использует разные части мозга. Вот почему он чувствителен к повреждению головного мозга. К счастью, тренировка мозга и практика могут помочь улучшить эту ключевую когнитивную функцию.
CogniFit, лидер программы по тренировке мозга, позволяет активировать и укрепить память и другие важные когнитивные способности. Наши игры для мозга были разработаны для стимулирования определенных паттернов нейронной активации, и повторение этих паттернов может помочь укрепить и улучшить нейронные связи, используемые в памяти. Это может помочь создать новые синапсы, способные реорганизовать и / или восстановить поврежденные или ослабленные когнитивные функции.
Типы долговременной памяти
Говоря о памяти, вы можете разбить ее на время, хранящееся в системах памяти, различая сенсорную память, кратковременную память, рабочую память и долговременную память:
- Декларативная память : Информация, хранящаяся в наших системах памяти, которая может быть объяснена и вызвана добровольно и сознательно.Системы мозга, связанные с этой системой памяти, — это медиальная височная доля, промежуточный мозг и неокортекс, и они разделены на две части.
- Semantic emory : Относится к набору имеющейся у нас информации об окружающем мире. Эта информация не связана с тем, как и когда она была изучена, и включает словарный запас, академические концепции или все, что мы знаем об определенном предмете. Например, вы знаете, что яблоко — это фрукт, который вы можете есть, что есть яблоки разного цвета и что оно происходит от яблони, но вы, вероятно, не помните, когда вы узнали эту информацию .
- Эпизодическая память : Включает в себя конкретный опыт, который мы пережили, и имеет очень тесную связь с тем, как и когда усваивается информация. Например, вспомнить, что вы ели на ужин прошлой ночью, где припарковали машину, когда вы впервые посетили определенный город, с кем вы пошли на определенную вечеринку или когда вы встретили этого человека.
- Недекларативная память или неявная память : Эта память хранится в ваших системах памяти, но о ней нельзя говорить.Обычно он приобретается или внедряется посредством неявного обучения (вы можете не осознавать, что изучаете его). Этот тип памяти довольно устойчив к повреждению мозга, что обычно оставляет его менее уязвимым, чем другие системы памяти. Этот тип памяти использует разные части мозга, такие как неокортекс, миндалевидное тело, мозжечок и базальные ганглии. В него также входят другие подразделения.
- Процедурная память : Состоит из информации о мышечных движениях, которые мы научились автоматизировать на практике, например о привычках и других навыках.Например, катание на велосипеде, бросание мяча или перемещение компьютерной мыши.
- Priming : Относится к легкости, с которой мы активируем и запоминаем определенную концепцию в нашем сознании. Например, вы, вероятно, быстрее вспомните слово «седан», если бы говорили о «легковых автомобилях», «грузовиках» или «кабриолетах».
- Classic Conditioning : Относится к связи между условным стимулом и реакцией, которая ранее была связана с безусловным стимулом.Например, если вы слышите звон колокольчика (условный раздражитель) перед тем, как подуть воздух в глаза (безусловный раздражитель), этого звука будет достаточно, чтобы вы моргнули (условная реакция). Эти отношения образуют часть недекларативной или неявной памяти
Хорошая память необходима для автоматического и беспроблемного выполнения нашей повседневной деятельности. Вот почему так важно знать и понимать, насколько хорошо работает ваша память. CogniFit предлагает серию тестов для измерения памяти (например, кратковременную слуховую память, контекстную память, кратковременную память, невербальную память, кратковременную зрительную память, рабочую память и распознавание) на основе Continuous Performance Test (CPT) , прямой и косвенный цифровой тест по шкале памяти Векслера (WMS), NEPSY (по Коркману, Кирку и Кемпу), тест переменных внимания (TOVA), тест на нарушение памяти (TOMM), Tower of Лондонский тест (TOL) и задача по визуальной организации Хупера (VOT).Эти тесты, помимо измерения памяти, также оценивают время отклика, скорость обработки, наименование, визуальное восприятие, когнитивное обновление, планирование, визуальное сканирование и пространственное восприятие.
- Тест секвенирования WOM-ASM: на экране появится серия фигур с цифрами. Пользователь должен будет запомнить серию чисел, чтобы потом читать ее. Эта последовательность начинается с двух чисел, но будет увеличиваться по мере продвижения пользователя, пока не сделает ошибку. Последовательность будет повторяться после каждой презентации.
- Запрос-тест REST-COM: Объекты на экране появятся на короткое время. Позже пользователю нужно будет как можно быстрее выбрать слова, соответствующие представленным изображениям.
- Идентификационный тест COM-NAM: Объекты будут представлены либо с изображениями, либо со звуками. Пользователь должен будет вспомнить, как (изображение или звук) объект был представлен в последний раз, или если он не был ранее представлен вообще.
- Тест на концентрацию VISMEM-PLAN: стимулы будут случайным образом появляться на экране.Стимулы загорятся вместе со звуком в определенном порядке. Пользователь должен обратить внимание на порядок, в котором активируются стимулы, чтобы позже правильно повторить порядок, когда придет их очередь.
- Тест на распознавание WOM-REST: на экране появятся три объекта. Пользователь должен как можно быстрее запомнить порядок, в котором объекты были представлены. Затем появятся четыре серии из трех объектов, и пользователь должен будет определить серию, которая совпадает с предыдущей.
- Test de Recuperación VISMEM: Aparecerán imágenes en la pantalla durante aproximadamente cinco o seis segundos. Durante ese tiempo, hay que intentar recordar la mayor cantidad de objetos que aparezcan en la imageen. Agotado ese tiempo, la imagen desaparece y se ofrecen diferentes opciones, entre las que el usuario debe detectar la correa.
- Recovery Test VISMEM: изображения будут появляться на экране на пять или шесть секунд. За это время пользователь должен будет запомнить как можно больше объектов, которые появляются на изображении.Когда время истечет, появятся разные варианты, и пользователю нужно будет выбрать правильный.
Примеры долговременной памяти
- Большинство концепций, которые мы изучаем в академической среде, хранятся в нашей семантической памяти. Из-за этого, когда вы изучаете или запоминаете географию своей страны, анатомию, химию, математику или любой другой предмет, вы используете свою долговременную память.
- Если вы работаете в ресторане и должны помнить еду, которую заказал каждый клиент, вы будете использовать эпизодическую память.То же самое происходит, например, когда вы вспоминаете своих обычных клиентов.
- Когда впервые учишься ездить на велосипеде, то падение вниз — это нормально, пока ты не освоишься с ним. Это потому, что мышцы, используемые при езде на велосипеде, не научились правильно двигаться. Однако, как только вы освоите езду на велосипеде, ваша процедурная память возьмет верх и автоматически начнет контролировать необходимые двигательные навыки. Это позволяет нам нормально ездить на велосипеде, не падая. Похожий процесс происходит при обучении вождению автомобиля.
- Чтобы запомнить, где вы оставили машину на стоянке, где находится зарядное устройство для телефона, какая столица вашей страны, или любую другую информацию, которую вам нужно запоминать изо дня в день, вы будете использовать свой длинный -срочная память.
Патологии и нарушения, связанные с проблемами долговременной памяти.
Забывчивость — это не проблема памяти. Фактически, наши системы памяти фактически удаляют малоиспользуемую или ненужную информацию, чтобы освободить место для более важных воспоминаний, и это особенно часто встречается с возрастом.Однако патологическая забывчивость действительно существует и будет характеризоваться неспособностью включать новые воспоминания (антероградная амнезия) и / или неспособностью вспоминать прошлые воспоминания (ретроградная амнезия). Существует также гипермнезия или непроизвольный доступ к ярким и подробным воспоминаниям, как в случае с посттравматическим стрессовым расстройством (посттравматическим стрессовым расстройством). Кроме того, воспоминания могут быть изменены или изменены при некоторых расстройствах, таких как синдром Корсакова, когда человек невольно изобретает воспоминания, которые он не может правильно вспомнить.
Самая известная проблема с памятью — это болезнь Альцгеймера (которая затрагивает в основном эпизодическую память), но проблемы с памятью также присутствуют при других типах деменции, как в случае с семантической деменцией (где затронутой системой памяти является семантическая память). или при болезни Паркинсона (когда нарушается процедурная память). В этих случаях обычно наблюдаются как ретроградная, так и антероградная амнезия. В случае черепно-мозговой травмы (ЧМТ) и повреждения мозга в результате инсульта также часто встречается антеоградная амнезия (учитывая, что она встречается чаще, чем ретроградная амнезия).Во всех этих случаях люди нередко создают истории, чтобы дополнить недостающую информацию (конфабуляции). Употребление определенных лекарств или веществ также может вызвать временную или постоянную потерю памяти.
Как нейроны формируют долговременную память — Harvard Gazette
Поздним летним днем 1953 года молодой человек, которого вскоре стали называть пациентом Х. перенесла экспериментальную операцию. Пытаясь вылечить его изнуряющие приступы, хирург удалил части его мозга, в том числе часть структуры, называемой гиппокампом.Приступы прекратились.
К сожалению, для пациента Х.М. то же самое случилось и со временем. Проснувшись после операции, он больше не мог формировать новые долговременные воспоминания, несмотря на то, что сохранил нормальные когнитивные способности, язык и кратковременную рабочую память. Состояние пациента Х.М. в конечном итоге показало, что способность мозга создавать долговременные воспоминания — это отдельный процесс, который зависит от гиппокампа.
Ученые обнаружили, где рождаются воспоминания. А вот , как они сделаны, осталось неизвестным.
Теперь нейробиологи Гарвардской медицинской школы (HMS) сделали решающий шаг в стремлении понять биологию долговременной памяти и найти способы вмешательства, когда дефицит памяти возникает с возрастом или болезнью.
В сообщениив журнале Nature от 9 декабря они описывают недавно идентифицированный механизм, который нейроны в гиппокампе взрослых мышей используют для регулирования сигналов, которые они получают от других нейронов, в процессе, который кажется критически важным для консолидации памяти и отзыва.
Исследование проводили Линн Яп, аспирант HMS в области нейробиологии, и Майкл Гринберг, заведующий кафедрой нейробиологии в Институте Блаватника при HMS.
«Если мы сможем лучше понять этот процесс, у нас будут новые дескрипторы памяти и способы вмешательства, когда что-то пойдет не так,…»
— Майкл Гринберг, Институт Блаватника в HMS
«Память необходима для всех аспектов человеческого существования. Вопрос о том, как мы кодируем воспоминания, которые сохраняются на всю жизнь, является фундаментальным, и наше исследование затрагивает самую суть этого феномена », — сказал Гринберг, профессор нейробиологии HMS Натана Марша Пьюзи и автор исследования.
Исследователи заметили, что новый опыт активирует редкие популяции нейронов в гиппокампе, которые экспрессируют два гена, Fos и Scg2. Эти гены позволяют нейронам точно настраивать сигналы от так называемых тормозных интернейронов, клеток, которые ослабляют нервное возбуждение. Таким образом, небольшие группы разрозненных нейронов могут образовывать устойчивые сети с скоординированной активностью в ответ на опыт.
«Этот механизм, вероятно, позволяет нейронам лучше общаться друг с другом, так что в следующий раз, когда потребуется вызвать воспоминание, нейроны сработают более синхронно», — сказал Яп.«Мы думаем, что одновременная активация этого Fos-опосредованного контура потенциально является необходимой функцией для консолидации памяти, например, во время сна, а также для восстановления памяти в мозгу».
Схемная оркестровка
Чтобы сформировать воспоминания, мозг должен каким-то образом связать опыт с нейронами, чтобы при реактивации этих нейронов можно было вспомнить первоначальный опыт. В своем исследовании Гринберг, Яп и его команда решили изучить этот процесс, изучив ген Fos.
Впервые описанный в нейронных клетках Гринбергом и его коллегами в 1986 году, Fos экспрессируется в течение нескольких минут после активации нейрона. Ученые воспользовались этим свойством, используя Fos в качестве маркера недавней нейронной активности, чтобы идентифицировать клетки мозга, которые регулируют жажду, оцепенение и многие другие формы поведения.
Ученые выдвинули гипотезу, что Fos может играть решающую роль в обучении и памяти, но на протяжении десятилетий точная функция гена оставалась загадкой.
Для исследования исследователи подвергли мышей воздействию новой среды и изучили пирамидные нейроны, основные клетки гиппокампа.Они обнаружили, что относительно редкие популяции нейронов экспрессируют Fos после контакта с новым опытом. Затем они предотвратили экспрессию Fos в этих нейронах с помощью инструмента на основе вируса, доставленного в определенную область гиппокампа, что не затронуло другие клетки.
Мыши, у которых был заблокирован Fos таким образом, показали значительный дефицит памяти при оценке в лабиринте, который требовал от них запоминания пространственных деталей, что указывает на то, что ген играет решающую роль в формировании памяти.
Исследователи изучили различия между нейронами, экспрессирующими Fos, и нейронами, которые не экспрессировали.Используя оптогенетику для включения или выключения входов от различных соседних нейронов, они обнаружили, что на активность Fos-экспрессирующих нейронов наиболее сильно влияют два типа интернейронов.
Было обнаружено, что нейроны, экспрессирующие Fos, получают повышенные подавляющие активность или подавляющие сигналы от одного определенного типа интернейрона и сниженные ингибирующие сигналы от другого типа. Эти сигнальные паттерны исчезли в нейронах с заблокированной экспрессией Fos.
«Что критично в этих интернейронах, так это то, что они могут регулировать, когда и сколько активируются отдельные Fos-активированные нейроны, а также когда они активируются относительно других нейронов в цепи», — сказал Яп.«Мы думаем, что наконец-то у нас есть представление о том, как Fos может на самом деле поддерживать процессы памяти, в частности, оркеструя этот тип пластичности схемы в гиппокампе».
Представьте себе день
Далее исследователи исследовали функцию Fos, который кодирует белок фактора транскрипции, регулирующий другие гены. Они использовали секвенирование отдельных клеток и дополнительные геномные скрининги для идентификации генов, активируемых Fos, и обнаружили, что один ген, в частности, Scg2, играет критическую роль в регуляции тормозных сигналов.
У мышей с экспериментально подавленным Scg2 активированные Fos нейроны в гиппокампе обнаруживают дефект передачи сигналов от обоих типов интернейронов. У этих мышей также были дефекты тета- и гамма-ритмов, свойств мозга, которые считались критическими характеристиками обучения и памяти.
Предыдущие исследования показали, что Scg2 кодирует нейропептидный белок, который может расщепляться на четыре различные формы, которые затем секретируются. В текущем исследовании Яп и его коллеги обнаружили, что нейроны, по-видимому, используют эти нейропептиды для точной настройки сигналов, которые они получают от интернейронов.
В совокупности эксперименты команды предполагают, что после нового опыта небольшая группа нейронов одновременно экспрессирует Fos, активируя Scg2 и производные от него нейропептиды, чтобы создать скоординированную сеть, активность которой регулируется интернейронами.
«Когда нейроны активируются в гиппокампе после нового опыта, они не обязательно заранее связаны друг с другом каким-либо определенным образом», — сказал Гринберг. «Но интернейроны имеют очень широкие аксональные ветви, что означает, что они могут связываться со многими клетками и передавать им сигналы одновременно.Это может быть то, как редкая группа нейронов может быть связана вместе, чтобы в конечном итоге закодировать память ».
Результаты исследования представляют возможный механизм долговременной памяти на молекулярном и схемном уровне. Они проливают новый свет на фундаментальную биологию формирования памяти и имеют широкое значение для заболеваний, связанных с дисфункцией памяти.
Исследователи отмечают, однако, что, хотя результаты являются важным шагом в нашем понимании внутренней работы памяти, остаются без ответа многочисленные вопросы о вновь идентифицированных механизмах.
«Мы еще не совсем пришли к ответу, но теперь мы видим многие из следующих шагов, которые необходимо предпринять», — сказал Гринберг. «Если мы сможем лучше понять этот процесс, у нас будут новые ручки памяти и способы вмешательства, когда что-то пойдет не так, будь то потеря памяти, связанная с возрастом, или нейродегенеративные расстройства, такие как болезнь Альцгеймера».
Результаты также представляют собой кульминацию десятилетий исследований, даже несмотря на то, что они открывают новые направления исследований, на изучение которых, вероятно, уйдут еще десятилетия, добавил Гринберг.
«Я прибыл в Гарвард в 1986 году, когда была опубликована моя статья, описывающая открытие, что активность нейронов может включать гены», — сказал он. «С того времени я представлял себе день, когда мы выясним, как гены, подобные Fos, могут способствовать долговременной памяти».
Дополнительные авторы: Ноа Петтит, Кристофер Дэвис, М. Аурел Надь, Дэвид Хармин, Эмили Голден, Онур Даглиян, Синди Лин, Стефани Рудольф, Нихил Шарма, Эрик Гриффит и Кристофер Харви.
Исследование было поддержано Национальным институтом здравоохранения (гранты R01NS028829, R01NS115965, R01NS089521, T32NS007473 и F32NS112455), Стюарт Х.Q. и Виктория Куан, аспирант кафедры нейробиологии Гарвардского университета, Фонд Арамонта.
Восстановление долговременной памяти после удаления ее поведенческой и синаптической экспрессии в Aplysia
1) Назначение памяти для изменения в ядре не является необоснованным, но все же это полностью спекулятивно. Это особенно верно, когда очевидное деацетилирование гистонов, вызванное хелеритрином, не нарушает скрытую постоянную память.Логически это означает, что остаточная память, которая сохраняется после ингибирования PKC и которая может быть полностью восстановлена (или демаскирована) с помощью еще 3 тренировочных испытаний, не может быть опосредована ацетилированием гистонов. Это важная путаница в авторском анализе. Хотя они хотят определить роль ацетилирования гистонов в «независимом от синапсов» механизме памяти, их данные фактически указывают на то, что существует дополнительный механизм памяти помимо PKC-зависимой памяти, стирание которой блокируется TSA.Этот вопрос следует более объективно и критически обсудить .
Мы согласны с этим важным моментом и благодарим рецензентов за указание на нашу ошибку. В нашем обсуждении потенциальной роли эпигенетических механизмов мы теперь признаем, что результаты хелеритрина опровергают идею о том, что механизм хранения LTM включает ацетилирование гистонов. Теперь мы предлагаем альтернативный эпигенетический механизм хранения LTM, метилирование ДНК. Как мы обсуждаем, недавние данные лаборатории Дэвида Свитта показали, что изменения метилирования ДНК, вызванные обучением или ранней травмой, могут сохраняться на протяжении всей жизни животного.Ремоделирование хроматина после двухцепочечных разрывов ДНК (Suberbielle et al., 2013) является еще одним потенциальным ядерным механизмом для хранения LTM.
2) Прайминг против оккультной памяти. Авторы должны признать, что возможно, что исходная последовательность воздействий 5-HT придает синапсам пластичность даже после стирания памяти. Существует значительное количество экспериментальных результатов, предполагающих праймирование как на синаптическом, так и на поведенческом уровнях (например, Hegde et al, 1993 , 1997 ).Результаты Barco et al (2002, , , рис. 5, , ) с конститутивно активным CREB предполагают, что изменения в экспрессии генов могут впоследствии взаимодействовать с локальными синаптическими сигналами, чтобы инициировать синаптическую пластичность. Даже Guan et al. Результат с ингибитором HDAC, который позволяет одиночному импульсу 5-HT инициировать долгосрочное облегчение, предполагает, что синапсы могут быть подготовлены для долгосрочной пластичности. Таким образом, ранняя тренировка может вызвать молекулярные изменения (например, транскрипцию ИЭГ или транслокацию в ядро), но может потребоваться последующая тренировка, чтобы вызвать изменение синаптической силы или поведения.Эти прайминговые механизмы считаются молекулярным объяснением важности повторных тренировок. Если бы феномен Чена-Гланцмана работал аналогичным образом, могло бы произойти постоянное изменение, которое заставляет клетки реагировать на последующие тренировочные испытания. Одним из таких изменений может быть ацетилирование гистонов, хотя данные TSA не согласуются с этой возможностью. Изменение протеасомы убиквитинирования, киназы или фосфатазы — примеры негеномных стойких изменений, которые могут вносить свой вклад.С этой точки зрения синаптические изменения будут частью памяти, но есть изменения в масштабах всей клетки, которые будут более стойкими. Это менее парадоксальное предложение, но оно может быть не менее интересным. Его преимущество заключается в сохранении возможности специфической пластичности синапсов, способствующей обучению. Авторы должны признать и обсудить эти возможности .
Благодарим рецензентов за содержательное обсуждение. Мы переписали рукопись, чтобы учесть возможность того, что наши результаты отражают устойчивость механизма подготовки, необходимого для восстановления нарушенного LTM, а не самого LTM.Кроме того, мы явно сравниваем первичное объяснение наших результатов с идеей о том, что LTM не был стерт в результате реконсолидационной блокады / лечения хелеритрином.
3) В этих экспериментах стабильность как базовой, так и долгосрочной облегченной синаптической силы в течение нескольких дней предполагает гомеостатический механизм, который определяет чистую синаптическую силу, а не ограничивает силу отдельных синаптических связей. Похоже, существует заданная точка для чистой синаптической силы, хотя эта заданная точка может сдвигаться во время долгосрочной синаптической пластичности (во многом, как описали Ху и его коллеги.) То, что общая сила синаптических связей может оставаться стабильной, пока отдельные синаптические связи формируются или сокращаются, не обязательно означает, что синапсы не «хранят память». Вместо этого, это предполагает возможность существования механизма для установки глобальной синаптической силы, во многом как это было предложено многочисленными исследованиями гомеостатического синаптического масштабирования. Следует более подробно обсудить концепцию уставки, которая смещается с сенсибилизацией .
Мы добавили подробное обсуждение понятия, что общая сила синаптических связей регулируется гомеостатическим механизмом, который переключается между облегченным и нефасилитированным состоянием, как определено опытом.Однако наш аргумент против идеи, что синапсы хранят память, не основывается просто на идее о том, что общая сила сенсомоторной связи остается стабильной, в то время как отдельные синапсы растут и сокращаются. Более веские доказательства против концепции синаптического хранилища прибывают из нашего открытия, что когда гомеостатический механизм сбрасывает общее количество синаптических связей до значения до обучения, результирующая морфология сильно отличается от исходной, и что память может быть восстановлена у животного. после антимнемонических процедур, которые стирают синаптический рост, связанный с обучением.
4) С молекулярной точки зрения, хотя заманчиво возложить основную ответственность за представление памяти на один белок или ген и сосредоточить внимание на этом отдельном игроке (по крайней мере, в рамках данного исследования), это явно не верный способ. чтобы понять биологию памяти. Все молекулярные участники хранилища памяти взаимодействуют с широким спектром других молекул в сложных сетях. Мы знаем из таких исследований, как Casadio et al. (1999) и Barco et al. (2002), что изменения транскрипции взаимодействуют с локальными синаптическими «метками», чтобы инициировать долгосрочные изменения синаптической силы.Действительно, на протяжении почти двух десятилетий напрашивается вывод, что ни транскрипционных изменений, ни локальных синаптических изменений недостаточно для представления долговременной памяти. Авторы предполагают, что синапсы просто выражают память, которая «хранится» на уровне генома. Однако они не представляют доказательств того, что точка гомеостаза задается исключительно в геноме (или даже в транскриптоме), а не возникает в результате динамического процесса, в котором синаптические сайты интерактивно участвуют в хранении памяти.Если рассматривать грубую аналогию, это немного похоже на утверждение, что сила мышц определяется ядрами мышечных волокон, а не посредством сложных динамических взаимодействий между экспрессией генов, элементами цитоскелета, сигнальными молекулами и внеклеточным матриксом, а также немышечными клетками просто потому, что отдельные миозиновые или актиновые филаменты переворачиваются .
Обсуждение могло бы быть лучше воспринято, если бы роль синапса в управлении памятью, специфичной для синапса, по крайней мере, была признана и интегрирована (например, см. Пункт 5 ниже) .
Индукция LTM может оказаться бесполезной моделью для хранения LTM. В частности, хотя результаты Barco et al. и другие показывают, что CREB-зависимые изменения в экспрессии генов могут взаимодействовать с локальными синаптическими тегами, чтобы инициировать LTF, как утверждает автор обзора, наши результаты не предоставляют никаких доказательств для синаптического тегирования. Это связано с тем, что ретракция варикозного расширения сенсорных нейронов после блокады реконсолидации или лечения хелеритрином казалась произвольной. Идея синаптического тегирования во время хранения LTM трудно согласовать с этим очевидно произвольным устранением варикозного расширения вен, как исходного, так и вызванного 5HT.Кроме того, нам кажется, что гомеостатический механизм, каким бы он ни был, находится либо в ядре, либо в теле клетки; Идея автора обзора о том, что синаптический гомеостаз является результатом динамического взаимодействия между некоторым механизмом прайминга и мечеными синаптическими сайтами, кажется жизнеспособной только в контексте синаптического тегирования. Возможно, что синапсы, которые остаются после блокады реконсолидации или лечения хелеритрином, содержат индуцированный 5HT молекулярный сигнал (метку), поэтому они выживают при антимнемоническом лечении; эта метка может взаимодействовать с остаточным механизмом прайминга в ядре или теле клетки для восстановления LTM.
5) Авторы должны обсудить возможность того, что синапсы просто переключаются между базовым состоянием и облегченным состоянием (с двумя амплитудами связей), так что кажущееся восстановление скрытой памяти на самом деле является просто переключением обратно к облегченному состоянию, что происходит легче после грунтования. Могут быть веские доказательства того, что это не так, но это следует прямо учитывать .
Как указано в ответе на Комментарий 3 выше, мы согласны с рецензентами, что в наших данных подразумевается возможность того, что «синапсы просто переключаются между базальным состоянием и облегченным состоянием».(Обратите внимание, что также должно быть третье состояние, включающее доказательства того, что сенсомоторный синапс может проявлять долгосрочную депрессию.) Но идея зависимого от опыта переключения между базальным и облегченным состояниями в конечном итоге согласуется либо с постоянным механизмом прайминга, либо с скрытая память.
6) С точки зрения размещения настоящего исследования в более широком контексте, авторам было бы полезно обсудить исследования взаимодействий между транскрипцией и локальными сигналами, такие как упомянутые выше, а также недавние исследования ЦНС млекопитающих, которые демонстрируют замечательные результаты. морфологическая стабильность отдельных дендритных шипов в течение недель или месяцев, e.грамм. И Цзо и его коллеги .
В измененную рукопись теперь включено обсуждение этого важного вопроса. Вкратце, существуют значительные разногласия относительно степени устойчивости шипов в головном мозге млекопитающих в течение длительных периодов времени. Кроме того, даже в тех исследованиях, в которых сообщалось, что позвоночник в основном стабилен после обучения, наблюдается значительный поворот позвоночника в первые несколько дней после тренировки, когда были сделаны наши измерения. Таким образом, наши результаты не обязательно противоречат результатам исследований изображений млекопитающих in vivo.
7) Одним из самых новых результатов в этой рукописи является открытие на последнем рисунке, что обращение долговременной памяти путем временного ингибирования атипичной протеинкиназы C (гомологичной PKM zeta в головном мозге млекопитающих) требует активности гистондеактилазы. Этот результат требует переоценки давно аргументированной модели, согласно которой постоянная активность PKM необходима для памяти, поскольку она поддерживает повышенную синаптическую силу в отдельных локальных синаптических сайтах. Даже с этим одним набором данных эти результаты могут иметь большее влияние в отдельной короткой публикации, а не в конце настоящего исследования.Эта точка зрения частично подтверждается выводами авторов, которые довольно упрощают этот важный вывод. (Например, в аннотации сказано: «мы предоставляем доказательства того, что LTM в Aplysia сохраняется за счет эпигенетических изменений», и почти идентичные сводные утверждения появляются в конце введения и в обсуждении.) Мы знаем более чем один 10 лет как в аплизии, так и в ЦНС млекопитающих ацетилирование гистонов имеет решающее значение для долговременной синаптической пластичности и долговременной памяти.Новым здесь является то, что атипичная активность PKC, по-видимому, необходима для сохранения эффекта, зависимого от ацетилирования гистонов, возможно, за счет подавления активности деактилазы гистонов .
Мы согласны с рецензентами в том, что это открытие является новым и важным, и, конечно же, не хотим преувеличивать его, добавляя сюда. Мы считаем, что это открытие правильно относится к статье, потому что оно поддерживает нашу общую позицию о том, что ядерные изменения имеют центральное значение для сохранения долговременной памяти (хотя мы согласны с рецензентом в том, что ацетилирование гистонов не может объяснить восстановление памяти после лечения хелеритрином).Тот факт, что TSA блокирует разрушительный эффект хелеритрина на LTM, также перекликается с другим выводом на рисунке 9, а именно, что TSA может преобразовывать промежуточную сенсибилизацию, вызванную тремя приступами ударов хвостом, в LTS. Пересмотренное обсуждение включает анализ роли активности PKC в эпигенетической регуляции индукции и поддержания LTM у Aplysia . Кроме того, мы переписали Аннотация, Введение и Обсуждение, чтобы исключить повторяющиеся утверждения, цитируемые рецензентами.
8) Важное предсказание гипотезы о том, что синаптический рост является механизмом экспрессии LTM, а не сам LTM, что в совместных культурах человек должен иметь возможность повторно экспрессировать больший синаптический рост после 5X HT напоминания-анизомицин 3X 5HT, чем в наивном 3X 5HT. Можно ли это проверить и количественно оценить? (Возможно, эти данные уже есть? )
Хотя наши результаты действительно предсказывают этот результат, его тестирование потребует значительных усилий. Обратите внимание, что для убедительности морфологических экспериментов, предложенных рецензентом, должны предшествовать электрофизиологические эксперименты, подтверждающие, что лечение 5X5HT-1X5HT-Aniso-3X5HT производит LTF, а, как и следовало ожидать, только 3X5HT не дает.Эти эксперименты еще не проводились. Кроме того, поскольку эксперименты потребуют внутриклеточной электрофизиологической записи пре- и постсинаптических нейронов сокультуры в течение трех дней подряд, их успех будет низким. Я полагаю, что электрофизиологические и морфологические эксперименты, необходимые для проверки предсказания, займут у моей лаборатории большую часть года. Поэтому я надеюсь, что рецензенты согласятся разрешить нам публиковать без этих данных.
9) Хотя бумага, как правило, написана четко, читать ее очень утомительно из-за сложной природы протоколов.Этого может быть просто невозможно избежать, но авторы могут еще больше постараться, чтобы текст был потоком. Кроме того, работа в значительной степени зависит от соответствующих физиологических исследований в совместных культурах, и авторам следует приложить все усилия, чтобы указать на детали этих исследований (например, Cai D, Pearce K, Chen S, Glanzman DL. 2011. Protein kinase М. поддерживает долгосрочную сенсибилизацию и долгосрочную фасилитацию при аплизии. J. Neurosci. 31: 6421-31. Цай Д., Пирс К., Чен С., Гланцман Д.Л .. 2012. Реконсолидация долговременной памяти при аплизии.Curr. Биол. 22: 1783-88) в соответствующих областях текста .
Я работал над упрощением написания статьи, но экспериментальные протоколы, по общему признанию, непростые. Что касается другого пункта рецензента, результаты наших предыдущих электрофизиологических исследований подробно описаны в соответствующих разделах текста, чтобы читатель мог точно увидеть, как наши нынешние морфологические результаты соответствуют предшествующим синаптическим физиологическим результатам.
10) Особенно любопытно, что обращение памяти восстановило количество пресинаптических варикозных расширений точно до того числа, которое было до формирования памяти.Этот момент можно интерпретировать более энергично, поскольку он соответствует предыдущей модели, в которой пластичность синапсов лежит в основе памяти и может стать основой для интеллектуальной переинтерпретации авторов (эта память существует, хотя она невидима при потере дополнительных синапсов) .
Мы согласны с рецензентами в том, что это интригующий результат, и подчеркнули его в нескольких местах рукописи. Результат подробно обсуждается во втором параграфе Обсуждения, где мы указываем, что восстановление количества пресинаптических варикозных расширений до предтренировочного уровня после реконсолидационной блокады / лечения хелеритрином проводится параллельно с результатами наших предыдущих электрофизиологических исследований (Cai et al ., 2011, 2012). Здесь также упоминается параллель между нашими результатами и результатами исследований гомеостатической синаптической пластичности в других системах.
11) Статистика. Хотя авторы выполняют ANOVA на своих данных, результаты ANOVA не представлены. Простое включение тестов posthoc нецелесообразно, поскольку общий результат ANOVA так же важен, как и тест posthoc (возможно, даже более). Значения F и точные значения p должны быть включены для каждого ANOVA .
Результаты всех дисперсионных анализов в нашем исследовании, включая точные значения F и p для каждого теста, представлены в соответствующих подписях к рисункам.
12) Даже если авторы хотят объединить группу 5×5-HT без напоминания 5-HT с группой 5×5-HT без напоминания 5-HT, но с поздним анизомицином в одну группу для целей статистического сравнения с другими группами лечения , нам должны быть показаны данные для двух обработок отдельно, по крайней мере, на начальном рисунке ( Рисунок 1C ) .
На рис. 1C теперь представлены данные для групп 5X5HT и 5X5HT-Aniso, построенные отдельно по запросу.
13) Резюме: «Кроме того, мы показываем, что LTS может быть восстановлен после его очевидного устранения посредством блокады реконсолидации и ингибирования PKM, лечения, которое стирает LTS-связанный синаптический рост». Это предложение не передает, что память тайно сохраняется после очевидного устранения. Восстановление предполагает только то, что возможно повторное обучение .
Аннотация переписана. Теперь в нем говорится: «Кроме того, мы находим доказательства того, что LTM для сенсибилизации сохраняется тайно после его очевидного устранения антимнемоническими препаратами, которые стирают связанный с обучением синаптический рост.”
14) Термин «+ группа напоминания» для синапсов, которые получают напоминание о 5-HT плюс анизомицин, является плохим термином, поскольку нет никаких предположений об использовании ингибитора синтеза белка. На рис. 7 эта же группа называется 5xTrained-Reminder-Aniso. В легенде Рис. 2 говорится: «Сокультура, в которой была нарушена синаптическая реконсолидация (+ Напоминание сокультуры)», что несколько сбивает с толку. Это не напоминание 5-HT нарушает память, а напоминание + ингибирование синтеза белка .
Рецензенты правы; первоначальные групповые ярлыки вводили в заблуждение. Мы заменили их новыми терминами, которые, как мы надеемся, будут менее запутанными и более точными. Таким образом, бывшая группа «- Напоминание» теперь была разделена на две экспериментальные группы для графика на Рисунке 1C, группы «5X5HT» и 5X5HT-Aniso ». Бывшая группа «+ Напоминание» теперь называется группой «5X5HT-1X5HT-Aniso». Новые метки групп на Рисунке 1 теперь более похожи на те, что на Рисунке 7. Кроме того, использование новых меток устраняет неточность, указанную рецензентом для легенды на Рисунке 2.
Обратите внимание, что мы также использовали новые групповые метки для рисунка 3. Прежняя группа «–Напоминание» (которая включает объединенные данные для групп 5X5HT, 5X5HT-1X5HT и 5X5HT-Aniso) теперь более точно помечена как «Без повторной консолидации / Никакой блокады ». Бывшая группа «+ Напоминание» теперь помечена как группа «5X5HT-1X-5HT-Aniso», как показано на Рисунке 1.
15) Из введения предложение «Эти морфологические результаты подразумевают, что LTM не зависит от синаптических изменений в аплизии» кажется внутренне противоречивым.Очевидно, авторы полагают, что память включает в себя синаптическую модификацию, поскольку они количественно определяют количество синаптических контактов, считывая для памяти .
Это предложение было переписано следующим образом: «Эти результаты означают, что постоянство памяти не требует стабильности определенных синаптических связей».
16) Результаты. Аннулирование морфологических изменений с помощью анизомицина после 5-HT «поддерживает идею о том, что напоминание запускает реконсолидацию синаптического роста, который опосредует LTF.«Реконсолидация — это то, что происходит только с напоминанием (например, только 5-HT). Обработка анисомицином блокирует обратное уплотнение. Об этом следует говорить более четко, особенно в журнале с широкой читательской аудиторией. Это можно объяснить во Введении при первом упоминании реконсолидации .
Предложение было изменено и теперь гласит: «Этот структурный результат… обеспечивает дополнительную поддержку идеи, что 1X5HT повторно активировал синаптическую память, вызванную тренировкой 5X5HT».Кроме того, введение теперь включает объяснение феномена реконсолидации памяти.
17) Результаты: «В течение этого периода также наблюдалось значительное уменьшение исходного варикозного расширения вен в обеих группах обученных сенсомоторной сокультуры, как у тех, кто получил стимул напоминания, так и у тех, кто этого не сделал». В описании не упоминается, что ретракция ранее существовавшего варикозного расширения вен была значительно более выраженной в группе «Напоминание + анизомицин». Это заслуживает внимания .
Мы добавили к результатам следующее утверждение: «Варикозное расширение вен в группе 5X5HT-1X5HT-Aniso — группе обученных сокультур, подвергнутых блокаде реконсолидации — продемонстрировало аналогичную картину ретракции вызванного 5HT и исходного варикоза, но количество втягивание было значительно больше, чем наблюдаемое в сокультурах без реконсолидации / без блокады. Обратите внимание, что «5X5HT-1X5HT-Aniso» — это новый ярлык для бывшей группы «+ Напоминание», а «Без повторной консолидации / Без блокады» — новый ярлык для бывшей группы «–Напоминание» (см. Ответ на комментарий 14 выше) .
18) Результаты: «ретракция наблюдалась ранее в группе 5-HT». Вероятно, авторы имеют в виду группу, которая получала 5-HT-напоминание плюс анизомицин (большинство групп получали 5-HT).
Приносим извинения за это запутанное предложение. Мы пересмотрели его, чтобы прояснить наш смысл: «Мониторинг судьбы отдельных варикозов в группе 5X5HT выявил ту же модель структурного роста и ретракции, которая наблюдалась ранее в сокультурах, обученных 5X5HT, не подвергшихся блокаде реконсолидации.”
19) Результаты: «Если эта идея верна, то должна быть возможность отделить LTM в Aplysia от синаптического содействия». Это следует уточнить .
Это предложение исключено. Введение к этому разделу статьи было переписано, чтобы прояснить наш смысл. Первый абзац теперь гласит:
.«Настоящие морфологические результаты ставят под сомнение представление о том, что стойкость сенсибилизационной памяти у Aplysia зависит от стойкости определенных облегченных синапсов.Для дальнейшего изучения этой идеи мы проверили, можно ли восстановить LTM для поведенческой сенсибилизации в Aplysia после блокады реконсолидации и ингибирования PKM, двух методов лечения, ранее показанных для устранения LTF, синаптической основы долгосрочной сенсибилизации… »
20) Результаты: «как указано при сравнении …… без тренировки с напоминанием» Это подробное описание сравниваемых групп не требуется в результатах. Предложение, приведенное до этого момента, поясняло, что три дополнительных экспозиции 5-HT восстанавливают стертую память .
Мы удалили это подробное описание.
21) Результаты: «три приступа ударов хвостом могут восстановить LTM после подавления его поведенческого выражения». Это предложение не подходит для результатов, так как оно несколько повторяется. Авторы могут логически предположить, что ингибирование атипичной PKC подавляет выражение долговременной памяти, а не стирает ее, только потому, что последующие удары хвостом восстанавливают (или восстанавливают) память. Конечно, свидетельства в этой рукописи согласуются с скрытой памятью, сохраняющейся после блокады реконсолидации и ингибирования ПКМ.Однако этот вывод о подавлении, а не стирании памяти должен быть рассмотрен в Обсуждении .
Это заявление было удалено.
22) Результаты: «синаптический механизм опосредует» должен иметь «это» перед «опосредует» . ”
Предложение, содержащее эту ошибку, удалено.
23) Обсуждение: «Может ли несинаптический, эпигенетический механизм хранения, такой как тот, который мы предлагаем для аплизии, опосредовать поддержание ассоциативных воспоминаний, особенно тех, которые индуцируются в сложных нейронных цепях в головном мозге млекопитающих, где данный нейрон может иметь тысячи или десятки тысяч синаптических партнеров? Это может показаться маловероятным.«Неужели авторы действительно хотят предположить, что механизм, предлагаемый их данными, актуален только для более простых нервных систем беспозвоночных ?
Благодарим за вопрос рецензентов. Нет, мы не считаем, что наши данные относятся только к нервной системе беспозвоночных. Мы исключили эти утверждения в Обсуждении.
24) L-15, Обсуждение: Была ли эта среда дополнена солями ?
Теперь в Материалах и методах указано, что L-15 был дополнен соответствующими солями.
25) Обсуждение: «основное тело овальной формы». Это странное описание варикозного расширения вен, поскольку оно предполагает наличие участков варикозного расширения за пределами «основного тела ». ”
Описательный термин «основной корпус овальной формы» заменен на «корпус овальной формы».
26) Обсуждение: «непарный t-критерий Стьюдента» Когда использовался t-критерий вместо ANOVA ?
В сводке наших статистических методов теперь говорится: «Непарный тест Стьюдента t использовался для определения статистической значимости различий, когда в наборе данных было только две группы, такие как данные, представленные на рисунках 3, 5, 6, 8 и 9D.”
27) В экспериментах с , рисунки 1 и 7 , этому рецензенту кажется очень важным иметь контроль над стимулом напоминания (после тренировки — 5X 5HT) без анисомицина, то есть 5X Trained Reminder. Это кажется даже более важным, чем наивный Контроль, потому что мы хотим знать, что само напоминание не имеет никакого эффекта. Я здесь не прав? Могут ли авторы объяснить, почему этот контроль не важен? Легенда для Рисунок 1 фактически предполагает, что этот контроль был проведен («Напоминание о 5-HT (одна синяя полоса) было доставлено в сокультуры + напоминания до анизомицина / носителя») .
Мы действительно включили этот контроль в наши морфологические эксперименты; она упоминается как группа «5X5HT-1X5HT». Дополнительный график к рисунку 1 (рисунок 1 — приложение к рисунку 1) включен в исправленную MS; он представляет собой среднее общее число варикозов, нормализованное к количеству варикозных заболеваний, присутствующих в 0 час, для каждой из трех групп обученных сокультив, не подвергшихся блокаде реконсолидации.
Однофакторный дисперсионный анализ, проведенный на данных варикозного расширения вен для 24-часового и 48-часового тестов, подтвердил, что различия между группами были незначительными.Обратите внимание, что группы 5X5HT и 5X5HT-1X5HT были объединены в одну группу (обозначенную как «5X5HT») для графика на Рисунке 1C. Кроме того, три группы, показанные на рисунке 1 — приложение к рисунку 1, группы 5X5HT, 5X5HT-1X5HT и 5X5HT-Aniso, были объединены в группу без реконсолидации / без блокады на рисунке 3.
Мы не включали этот контроль в поведенческие эксперименты.
28) Рисунок 3 статистическое сравнение. Поскольку авторы выполнили ANOVA, можно сравнить степень ретракции вновь образовавшихся варикозных вен с ретракцией исходных варикозных вен (например,грамм. Рисунки 3 и 5 ). Эти результаты должны быть включены .
Эти результаты включены в пересмотренную MS в виде рисунка 3 — добавление к рисунку 1 и рисунка 5 — приложение к рисунку 1. Интересный результат, показанный на графиках на этих новых рисунках, заключается в том, что, в то время как антимнемонические методы лечения — блокада реконсолидации и лечение хелеритрином — привели к При примерно равном сокращении варикозного расширения вен, вызванного 5-HT, и исходного варикоза, в группах, обученных 5X5HT, не подвергавшихся ни одной из антимнемонических терапий, наблюдалось значительно большее уменьшение варикозного расширения вен, вызванного 5-HT, по сравнению с исходным варикозным расширением.
https://doi.org/10.7554/eLife.03896.017Как улучшить кратковременную память
Обучение и память тесно связаны. В то время как обучение предполагает приобретение навыков и знаний, память является выражением того, что было приобретено.
Эти две концепции также можно различать в зависимости от скорости, с которой происходят оба процесса. Если приобретение новых навыков или знаний происходит медленно, это классифицируется как обучение. Однако, если это происходит мгновенно, он становится частью памяти.
Чтобы разобраться в памяти и ее свойствах, ее можно разделить на три категории: долговременная, краткосрочная и рабочая память. Что касается обучения — особенно для обучения в профессиональной среде — основное внимание уделяется внедрению концепций обучения в долговременную память, что является общей проблемой для предприятий. Не многие системы управления обучением (LMS) содержат встроенную функцию, разработанную специально для содействия сохранению знаний посредством распределенной практики, но платформа микрообучения EdApp поддерживает.Фактически, команда EdApp пошла дальше, создав функцию на основе алгоритмов для автоматического создания интерактивных уроков на основе основного, ранее изученного контента с помощью приложения для интервальных повторений Brain Boost.
Процесс памяти
Чтобы понять сохранение памяти и знаний в любом контексте, мы должны понимать принципы, лежащие в основе этого. Память — это активный, рефлексивный и субъективный процесс предыдущего опыта. Формирование памяти обычно подразделяется на три процесса:
Кодирование : этот процесс участвует в формировании памяти и зависит от трех факторов:
1.Контент. На тип кодируемого контента могут влиять структура, размер и тематика контента, а также степень осведомленности учащегося.
2. Окружающая среда : Это условия, которые могут либо тормозить, либо стимулировать процесс запоминания. (Пример: температура, влажность, социально-экономические условия и т. Д.).
3. Субъективный : Эти внешние переменные факторы, влияющие на процесс кодирования. (Пример: усталость, мотивация, интерес и т. Д.)
Сохранение : это второй процесс, в котором закодированная информация сохраняется для будущего использования.В этом процессе информация сохраняется, трансформируется и реорганизуется с новыми ориентирами. Информация хранится на трех уровнях памяти:
1. Долговременная память: эта память используется для хранения и вызова информации на более позднем этапе. Эмоции играют важную роль в процессе воспоминаний. Чем сильнее эмоции, вызванные памятью, тем легче вспомнить событие.
2. Кратковременная память: этот тип памяти длится всего 20-30 секунд, прежде чем они начнут быстро разрушаться.Кратковременную память можно улучшить, многократно репетируя содержание.
3. Рабочая память: рабочую память часто путают с кратковременной памятью. Эта память является «посредником» между краткосрочными и долгосрочными воспоминаниями. Эта память используется для запоминания информации и использования ее в дальнейшем для выполнения задач.
Получение : Этот процесс включает доступ к сохраненной информации посредством отзыва или распознавания. Напоминание происходит путем обнаружения информации из памяти, которая может быть фактом, объектом или событием.
Функция интервального повторения вEdApp, Brain Boost, основана на процессе обработки памяти и использует кривую забывания Эббингауза и широко известный интервальный алгоритм Supermemo SM-2. Благодаря этим двум ресурсам, подкрепленным всем, что мы знаем о памяти, Brain Boost работает за кулисами, чтобы гарантировать, что все ваши ученики учатся и, что более важно, сохраняют весь ваш контент.
Чтобы активировать функцию Brain Boost в EdApp, просто перейдите к выбранному вами курсу и выберите «Brain Boost» в меню параметров (ниже).
Оттуда просто переключите тумблер:
Какие 3 этапа памяти?
Запоминание — это трехэтапный процесс: кодирование, хранение, извлечение.
Кодировка
Воспоминание начинается с набора сенсорных входов — того, что мы видим, обоняем, ощущаем на вкус, слышим и прикасаемся. Этот ввод необходимо преобразовать в один из трех форматов хранения мозга: изображение, звук или значение. Например, аудиосигналы, связанные с визуальными символами, становятся «словами» с определениями — комбинацией форматов хранения звука и значений.
Хранилище
Закодированные элементы нужно где-то хранить. У мозга есть три варианта: кратковременная память, рабочая память и долговременная память.
Извлечение
Целью процесса памяти является извлечение или «запоминание». Другими словами, при необходимости один или несколько закодированных форматов «отображаются» в нашем воображении.
Кратковременная память
Где находится кратковременная память?
Кратковременная память представлена простыми паттернами нейронной коммуникации, присутствующими в этих областях мозга:
- Префронтальная кора
- Лобная доля
- Теменная доля.
Гиппокамп — это область мозга, отвечающая за перемещение информации от краткосрочной к долгосрочной, однако он не хранит саму информацию.
Фото: IFL Science
Где находится долговременная память?
Долговременная память создается изменениями нейронных связей гиппокампа, которые распространяются по всему мозгу. Части мозга, связанные с хранением долговременных воспоминаний:
- Префронтальная кора
- Головной мозг
- Лобная доля
- Медиальная височная доля.
Хотя постоянное хранение долговременных воспоминаний зависит от глубины связей между нейронами. Другими словами, воспоминания с более сильными связями — это те воспоминания, которые мы глубоко переработали.
Как долго длится кратковременная память?
Согласно Аткинсону и Шиффрину (1971), кратковременная память длится от 15 до 30 секунд. Однако эта цифра может варьироваться от секунд до одной минуты, в зависимости от репетиции или активного обслуживания.
Стратегия активной репетиции означает выделение момента для быстрой мысленной заметки, и это может быть так же просто, как повторение новой информации вслух или повторение мысленно от трех до пяти раз. Простое чтение или прослушивание информации без активной репетиции означает, что информация будет сохраняться в течение нескольких секунд. С другой стороны, повторение информации и даже создание мысленных ассоциаций с большей вероятностью сохранится в краткосрочной памяти на минуту (или даже дольше).
Это также зависит от умственных способностей, то есть мозг может сохранять новую информацию, если она еще не «заполнена».Это одна из причин, почему микрообучение является наиболее эффективной формой обучения, поскольку оно делится новым содержанием небольшими пакетами, что дает учащемуся больше шансов понять концепцию и, что более важно, сохранить ее. Чтобы продвинуться дальше минутной отметки, можно ввести другие практики, такие как интервальное повторение в EdApp или функция Brain Boost, чтобы еще больше пересмотреть и закрепить концепцию не только в краткосрочной, но и в долгосрочной памяти.
Что такое потеря кратковременной памяти?
По сути, кратковременная потеря памяти — это невозможность восстановить закодированные форматы недавних эпизодических событий.Другими словами, вы не можете вспомнить то, что видели, делали или слышали совсем недавно. Примеры включают: невозможность ответить вечером о том, что вы ели на обед несколькими часами ранее; пропущенные дозы нового лекарства из-за того, что вы забыли его принять, и выход из торгового центра, не помня, где вы припарковали машину. Хотя время от времени мы все можем забыть, особенно о машине, именно частота является решающим фактором.
Кратковременная потеря памяти может быть нормальным результатом старения.С другой стороны, это может указывать на основную проблему, такую как травма головного мозга или неврологическое (головное) заболевание.
Что влияет на кратковременную память?
Во-первых, мозг каждого человека при рождении разный. Некоторые из них вызваны генетикой. Другие могут быть вызваны внутриутробными состояниями (когда еще не родившийся плод находится внутри матери), такими как курение матери, употребление наркотиков или другие нездоровые привычки.
После рождения ребенка мозг подвергается целому ряду потенциальных опасностей.Слабые места на стенках артерий головного мозга, называемые аневризмами, могут разорваться (лопнуть). Когда они это сделают, кровь может хлынуть в область вокруг мозга, увеличивая черепное (головное) давление и вызывая повреждение клеток мозга. Если эти клетки связаны с кратковременной памятью, это повлияет на нее. Кроме того, травмы головы, инфекции головного мозга и аномальные новообразования, такие как опухоли, могут вызвать изменения в том, сколько человек может запомнить в краткосрочной перспективе. Кроме того, лечение заболеваний может нарушать кратковременную память.
Долговременная память
Является ли долговременная память постоянной?
Ответ на этот вопрос немного сложен. Чтобы лучше понять это, представьте себе долговременную память как набор маленьких коробочек, таких как стена из коробок, которую часто можно увидеть за стойкой администратора в фильмах. Вы знаете те, у которых портье берет ключ от комнаты (а в прежние времена — любые телефонные сообщения, которые человек получал, пока его не было).
Когда все ключи успешно помещены во все ящики, это похоже на то, что все наши воспоминания успешно закодированы в нашу долговременную память.Пока все хорошо, правда?
При обычном и обычном режиме делопроизводитель может в любое время получить (получить) любой ключ, который захочет, а затем заменить его, когда он не нужен. Точно так же мы можем «вспомнить» то, что нам нужно, в любое время, хотя наши воспоминания не входят и не выходят из долговременной памяти (ключевая аналогия может идти только до сих пор).
Однако многие вещи могут помешать служащим отеля получить ключи. Например, кто-то мог запечатать некоторые из ящиков, чтобы сотрудники не могли сунуть руки внутрь.Ключи есть, но клерки не могут получить к ним доступ. Или, возможно, какое-то злое существо зарезало лазером некоторые из ящиков, сжег и ключи, и сами ящики. Оба случая происходят в человеческом мозгу. Заболевания и другие проблемы с мозгом могут помешать нам получить доступ к закодированной информации, хранящейся в долговременной памяти. Широкий спектр повреждений мозга может разрушить клетки долговременной памяти и, следовательно, закодированную информацию, которую они содержат.
Как длится долговременная память?
Способ обеспечить долговременную память — через интервальное повторение, чтобы противодействовать эффектам кривой забывания Эббингауза.Кривую можно избежать, если информация постоянно встраивается в кратковременную память, пока она не запечатлевается в долгосрочной.
Каковы основные характеристики долговременной памяти?
Основные характеристики долговременной памяти включают воспоминания, которые обычно измеряются в месяцах или десятилетиях. Долгосрочные воспоминания или более далекие воспоминания могут включать важные вехи из прошлого, например, ранний день рождения или день свадьбы.
Вот 3 основных характеристики долговременной памяти:
- Сознательная память, которую исследователи также называют явной или декларативной памятью. Явная память включает в себя информацию, которую вам нужно запомнить, например математическую формулу или некоторые важные даты в истории.
- Бессознательная память также называется имплицитной или процедурной памятью. Эти типы воспоминаний включают в себя привычки, навыки и простые формы кондиционирования, такие как умение завязать шнурки или запоминание слов песни после первых нескольких нот.
- Взаимосвязь между обеими системами памяти. По словам исследователей, две системы зависят от гиппокампа и связанных с ним областей мозга, которые работают вместе, чтобы классифицировать и осмыслить сознательную память, бессознательную память и взаимосвязь между ними.
Какие типы долговременной памяти?
Саму долговременную память можно разделить на типы.
3 отдела
Эндел Тулвинг в 1972 году предложил три типа долговременной памяти: эпизодическую , процедурную и семантическую .Эпизодические воспоминания — это сознательные мысли о том, что мы считаем правдой. Например, закодированный элемент, напоминающий нам, что мы выключили плиту перед тем, как выйти из дома. Процедурные воспоминания — это, как звучит их название, закодированная информация о том, как делать такие вещи, как замена шины или чистка зубов. Наконец, семантические воспоминания — это закодированные элементы фактической информации, включая знание мира и язык.
2 подразделения
Некоторые исследователи говорят, что эпизодическая и семантическая память — это два типа декларативной памяти.Другими словами, то, что мы «знаем» (или верим) в истину. В результате они подразделяют долговременную память на два подразделения: декларативную и недекларативную. Pr
Еще один способ назвать эти две категории — явная (декларативная) и неявная (недекларативная) память.
Краткосрочная и долговременная память
В следующей таблице показаны основные физические различия между кратковременной и долговременной памятью.
Характеристика |
Кратковременная память |
Долговременная память |
Продолжительность |
20-30 сек.до начала распада |
«Навсегда», пока не помешает старение или плохое здоровье |
|
Место хранения в головном мозге |
|
|
Эти различия логически соответствуют цели каждого типа памяти.
Кратковременная память начинает процесс запоминания. Его функция — интерпретировать новую информацию. В результате он располагается в областях мозга, отвечающих за речь (чтение и письмо), интерпретацию языка и интерпретацию сенсорных сигналов (зрение, слух, осязание, звук).
Долговременная память — это наш склад памяти. Его функция — хранить наши прошлые интерпретации для извлечения по мере необходимости. Таким образом, он занимает некоторые из тех же областей мозга, что и кратковременная память (чтобы понять язык и сохраненное сенсорное кодирование), но также занимает еще одну важную область — медиальную височную долю.Помимо прочего, эта область мозга отвечает за последовательность и упорядочение. Это имеет смысл, поскольку поиск в памяти должен быть упорядоченным, чтобы информация отображалась в правильном порядке или в правильной последовательности.
Слабая память и обучаемость
Если вы иногда удивляетесь, почему вы так быстро забываете что-то, вы не одиноки. Слабая рабочая память может повлиять на обучение. Людям со слабой рабочей памятью будет сложно организовать и интегрировать новые навыки или знания.
В результате им будет сложно следовать указаниям, многоступенчатым процедурам обучения, организовывать мысли или обрабатывать информацию. Проблемы с долговременной памятью могут затруднять вспоминание информации. Человек может запутаться, дезорганизоваться или дезориентироваться, когда ему будет представлен новый учебный материал.
Память и синапсы
Консолидация памяти зависит от того, как синапсы работают в головном мозге. Синапсы подобны электрической системе, проводящей ток.
Синапсы передают сигналы между нейронами с помощью нейромедиаторов. Когда два нейрона многократно срабатывают вместе, они становятся сенсибилизированными, что дает им возможность работать в будущем. Благодаря новому опыту, воспоминаниям и информации в мозгу устанавливается несколько похожих связей, а старые отсеиваются.
От краткосрочной к долговременной памяти — консолидация
Повторное повторение и повторение информации приводит к усилению синаптического пути.
Повторное срабатывание одних и тех же нейронов повысит вероятность их одновременного срабатывания и в будущем.Это поможет запоминать и вспоминать информацию с большей точностью и легкостью.
Влияние на консолидацию памяти
Воспоминания часто неправильно понимают как картотеки, в которых хранится информация или определенные воспоминания в отдельных файлах. Однако воспоминания распределены по разным частям мозга.
Одна из основных теорий сна показала, что сон может помочь в обработке и консолидации воспоминаний, которые были приобретены, когда люди бодрствовали.Утраченные воспоминания можно восстановить после того, как они будут восстановлены. Вызов и повторное объединение памяти помогает сохранять и укреплять информацию, хранящуюся в долговременной памяти.
Как мне улучшить краткосрочную память?
Процесс консолидации памяти может быть ускорен при изучении новых навыков или информации. Репетиция и запоминание с использованием мнемонических приемов могут улучшить процесс запоминания.
Другой метод улучшения консолидации информации в долговременной памяти — многократное повторение информации через интервалы (интервальное повторение).Этот метод можно найти прямо в платформе микрообучения: EdApp. Это обеспечивает лучшее запоминание, и его следует предпочитать технике зубрежки информации в ночь перед экзаменом.
Можно принять стратегии запоминания, поняв процесс консолидации памяти. Это гарантирует, что студенты смогут более эффективно и рационально использовать свое учебное время.
Если вы заинтересованы в использовании полностью бесплатной отмеченной наградами платформы EdApp, которая включает встроенные функции для автоматического улучшения долговременной памяти посредством обучения? Нажмите ниже, чтобы начать обучение бесплатно.Неограниченное количество пользователей. Кредитная карта не требуется.
Источники:
Память и мозг
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4355270/
Долговременная память, хранящаяся в коре головного мозга — ScienceDaily
«Где» и «как» воспоминания закодированы в нервной системе — один из самых сложных вопросов в биологических исследованиях. Формирование и вспоминание ассоциативных воспоминаний необходимо для независимой жизни.Гиппокамп долгое время считался центром мозга для длительного хранения пространственных ассоциаций. Теперь Мазахир Т. Хасан из Института медицинских исследований Макса Планка и Хосе Мария Дельгадо-Гарсиа из Университета Пабло де Олавиде в Севилье, Испания, смогли предоставить первые экспериментальные доказательства того, что особая форма ассоциаций памяти закодирована в мозговом мозге. кора головного мозга и не локализуется в гиппокампе, как описано в большинстве учебников по нейробиологии. Новое исследование меняет правила игры, поскольку оно настоятельно предполагает, что в качестве хранилища памяти используются моторные корковые цепи, а не гиппокамп.
Генри Молайсон, широко известный как Х.М., широко известен в области исследований памяти. Большие части гиппокампа американца — область мозга, которая является основным элементом процессов обучения и памяти — были удалены в 1950-х годах в попытке вылечить его эпилептические припадки. Впоследствии у него были серьезные провалы в памяти, и он больше не мог вспомнить практически ничего нового, что узнал. Таким образом, большинство ученых пришли к выводу, что гиппокамп — это место долговременной памяти.
Однако степень распространения H.Повреждение мозга М. было явно недооценено, потому что другие области помимо гиппокампа также были удалены или повреждены во время хирургической процедуры. Поэтому исследователи из Гейдельберга и Севильи исследовали обучающее поведение генетически модифицированных мышей, у которых рецепторы NMDA отключаются только в моторной коре головного мозга. Рецепторы NMDA связывают глутамат нейромедиатора с синапсами и становятся активными, когда несколько сигналов поступают в один синапс одновременно. Они являются центральными молекулярными элементами процессов обучения, участвуя в увеличении или уменьшении передачи сигналов в синапсы.
Как показывает новое исследование, в моторной коре эта так называемая синаптическая пластичность больше не функционирует без рецепторов NMDA. Таким образом, ученые смогли исключить гиппокамп или другие области как причину своих наблюдений. Согласно новым открытиям, именно кора головного мозга, а не гиппокамп, является местом хранения некоторых форм памяти.
В поведенческих тестах, так называемое кондиционирование морганием, животные с рецепторами NMDA и без них в первичной моторной коре должны были научиться связывать тон с последующим электрическим раздражителем века.Эта ассоциация двух сенсорных входов включает мозжечок, который координирует необходимые движения, а также гиппокамп и кору головного мозга, которые являются важными центрами обучения и памяти. «После фазы обучения рефлекс животных заключается в том, чтобы закрыть глаза, когда они слышат только звук. Без рецепторов NMDA в первичной моторной коре головного мозга генетически модифицированные мыши, с другой стороны, не могут вспомнить связь между звуком и электрическим стимулом. , и поэтому они держат глаза открытыми, несмотря на тон «, — объясняет Мазахир Т.Хасан из Института медицинских исследований Макса Планка.
Таким образом, исследователи дополнили выводы своих коллег из Гейдельберга о том, что гиппокамп не является местом памяти. В июле 2012 года Рольф Шпренгель и Петер Зеебург из Института медицинских исследований Макса Планка обнаружили, что мыши без рецепторов NMDA в гиппокампе все еще вполне способны к обучению. «Теперь мы думаем, что гиппокамп предоставляет необходимые экологические сигналы, которые передаются в кору головного мозга, где происходят ассоциации, зависящие от обучения.Таким образом, воспоминания надолго сохраняются в различных участках коры головного мозга », — объясняет Хасан.
Таким образом, открытия Хасана и Дельгадо-Гарсиа представляют собой смену парадигмы в исследованиях памяти, поскольку они ясно показывают, что кора головного мозга — это область мозга, в которой связаны и сохраняются ассоциации памяти, а не гиппокамп.