
Толерантность презентация для 1 класса: Презентация ко Дню толерантности «Будьте добрыми» (1 класс)
Все мы разные | Классный час (1 класс) на тему:
Классный час о толерантности
«Все мы разные»
Цель:
Ввести и закрепить определение термина «толерантность”;
Отрабатывать умение видеть индивидуальное в каждом человеке;
Показать, что такое толерантное отношение к различиям;
Сформировать представление о толерантном отношении.
Оборудование: аудиозапись песни «Я,ты,он,она…», мультфильм, «листочки» фломастеры
Дети рассаживаются в классе по кругу звучит песня «Я,ты,он,она…»Я__ТЫ__ОН__ОНА____вместе_целая_страна.mp3
Ход занятия:
1.Просмотр мультфильма. мультфильм о толерантности.exe
О чем этот мультфильм? (о том, что не следует смеяться над другими, если они не такие как ты)
2.Игра «Чем мы похожи?»
Дети стоят в кругу. Ведущий приглашает в круг одного из участников на основе какого-либо реального или воображаемого сходства с собой. Например: «Света, выйди, пожалуйста, ко мне, потому что у нас с тобой одинаковый цвет волос (или мы похожи тем, что мы жители Земли, или мы одного роста и т. д.)». Света выходит в круг и приглашает следующего участника, не забыв назвать его по имени. Игра продолжается до тех пор, пока все не окажутся в кругу.
д.)». Света выходит в круг и приглашает следующего участника, не забыв назвать его по имени. Игра продолжается до тех пор, пока все не окажутся в кругу.
Видите дети, мы все разные, но у нас есть что-то общее. Нам интересно дружить друг с другом, потому что мы не похожи, но имеем общие черты характера, внешности.
3.Знакомство с понятием «толерантность»
Я хочу познакомить вас с древней китайской притчей «Ладная семья»
Жила-была на свете семья. Не простая семья. Более 100 человек насчитывалось в ней. Мало ли таких семей? Да, не мало. Но эта семья была особая. Ни ссор, ни ругани, ни драк, ни раздоров. Дошел слух об этой семье до самого владыки. И решил он проверить, правду ли говорят люди. Прибыл он в село, и душа его порадовалась: чистота и порядок, красота и мир. Хорошо детям, спокойно старикам. Удивился владыка и решил узнать, как добилась всего этого семья. Пришел он к старейшине. «Расскажи»,- говорит. Долго писал что-то на бумаге старейшина. А, когда написал, протянул владыке. Всего 3 слова были написаны на бумаге: «ЛЮБОВЬ, ПРОЩЕНИЕ, ТЕРПЕНИЕ» А в конце листа: « СТО РАЗ ЛЮБОВЬ, СТО РАЗ ПРОЩЕНИЕ, СТО РАЗ ТЕРПЕНИЕ».
Всего 3 слова были написаны на бумаге: «ЛЮБОВЬ, ПРОЩЕНИЕ, ТЕРПЕНИЕ» А в конце листа: « СТО РАЗ ЛЮБОВЬ, СТО РАЗ ПРОЩЕНИЕ, СТО РАЗ ТЕРПЕНИЕ».
-И все?
-Да, — отвечал старик, — это и есть основа жизни всякой хорошей семьи.- И подумав, добавил: «И мира тоже».
Ребята согласны ли вы с тем, что написал на листе бумаги глава семьи? Как вы думаете, о чем я хочу сегодня с вами поговорить? (о дружеских взаимоотношениях в классе…и т.д.)У этого определения есть понятие, которое имеет свою историю:
На рубеже XVIII-XIX веков во Франции жил некто Талейран Перигор, князь Беневентский. презентация.pptx (слайд 2.1). Он отличился тем, что при разных правительствах (и при революционном, и при Наполеоне, и при короле Людовике XVII) оставался неизменно министром иностранных дел. Это был человек, талантливый во многих областях, но, несомненно, более всего — в умении учитывать настроения окружающих, уважительно к ним относиться, искать решение проблем способом, наименее ущемляющим интересы других людей. И при этом сохранять свои собственные принципы, стремиться к тому, чтобы управлять ситуацией, а не слепо подчиняться обстоятельствам. От имени этого мудрого министра появилось слово ТОЛЕРАНТНОСТЬ, презентация.pptx(слайд2.2) что в переводе с английского означает терпение.
И при этом сохранять свои собственные принципы, стремиться к тому, чтобы управлять ситуацией, а не слепо подчиняться обстоятельствам. От имени этого мудрого министра появилось слово ТОЛЕРАНТНОСТЬ, презентация.pptx(слайд2.2) что в переводе с английского означает терпение.
4. Чтение-рассуждение сказки «Дедушкин совет»
Егорка был неплохим мальчиком, но как только встречался с Колей, он сразу же раздражался. Из всех знакомых Колька был самым неприятным, всюду хотел быть первым, но, самое главное, он любил насмехаться и дразнилки обидные придумывал. Однажды Егорка подрался с Колей и пришёл домой весь грязный. Дома его встретил дедушка, и Егорка рассказал ему обо всем, даже о том, как уже целый год, встречаясь с Колей и вспоминая о нём, он сильно сердится и злится.
— Надо быть, Егорка, доброжелательным ко всем людям, даже к тем, кто тебе не нравится,- сказал ему дедушка.
— Как же это можно желать добра тем, кто не нравится и кто плохой?- удивился Егорка.
— Может быть, только ты считаешь Колю плохим, а другие наоборот — считают его хорошим.
— Нет, я точно знаю, что он плохой.
— Тогда скажи, Егорка, мама Коли любит своего сына?
— Наверное, любит, — ответил Егорка.
— А друзья у него в школе или во дворе есть?
— Есть.
— Вот видишь, другие к нему хорошо относятся, а ты не можешь. Значит всё дело в тебе самом, а не в Коле. Просто не хочешь ты быть доброжелательным, а хочешь и дальше на него злым взглядом смотреть.
— Что же, дедушка, мне делать, если он мне не нравится?
— Ты можешь не играть и не дружить с ним, но всегда можешь тихо, про себя, пожелать ему чего-нибудь хорошего, чтобы он стал лучше и добрее. А продолжая злиться, ты будешь только сам себе вредить и душу свою пачкать.
— А просто молчать нельзя?- спросил Егорка.
— Нет, ты слишком долго злостью и раздражением душу свою отравлял, теперь только добрые мысли ее очистить могут. Ты только не забывай вовремя плохие мысли на добрые менять.
Стал Егорка делать так, как дедушка его научил. Увидит Колю, отвернётся в сторону и шёпотом пожелает ему лучше и добрее стать. А если вспомнит о нём, то плохие мысли в голову не пускает, а сразу добрыми их вытесняет.
Через месяц идёт дедушка домой, смотрит, а внук его Егорка с Колей играет и громко смеётся. Улыбнулся дедушка, глядя на сдружившихся ребят, и тут же, шёпотом что-то доброе им пожелал.
— Не мог или не хотел Егорка быть доброжелательным к Коле?
— Какой совет дал дедушка Егорке? (желать людям добра, менять плохие мысли на хорошие)
Понравилась сказка? А вы доброжелательно относитесь к людям? (ответы детей)
Доброжелательность – редкое качество в человеке. Каждому человеку,
даже плохому, очень легко желать добра своим знакомым, друзьям и просто приятным людям («И если любите любящих вас, какая вам за это благодарность?»). Но быть доброжелательным к людям чужим, неприятным, плохим очень трудно. Желать добра только тем, кто нравится, или тому, кто вас любит,- это только половина доброжелательности («Любите врагов ваших»). Вы не хотите наполовину радоваться, наполовину дружить и любить, наполовину быть смелым. Настоящая доброжелательность тоже не должна быть наполовину…
Вы не хотите наполовину радоваться, наполовину дружить и любить, наполовину быть смелым. Настоящая доброжелательность тоже не должна быть наполовину…
5. Изучение правил толерантного общения презентация.pptx (слайд 3)
6. Работа по представлению «Давайте пофантазируем…»
Представьте, что наступит время, когда все люди на Земле станут совершенно одинаковыми(рост, цвет волос и глаза, одежда, сумма знаний и т. д.)
1. Каким будет этот мир? Как люди в нем будут жить?
2. Хорошо или плохо, что мы все разные?
3. Как жить в мире, где столько разных людей?
ВЫВОД: Различия дополняют и обогащают общество. Чтобы жить в мире, людям надо научиться жить, решая проблемы и задачи на основе сотрудничества. Для этого существует правило «Трех П» презентация.pptx (слайд 4)
7. Творческая работа «Дерево толерантности».
Ученики на листочках бумаги в форме листа какого-либо дерева пишут, что надо сделать, чтобы класс стал «Пространством толерантности», листочки наклеиваются на символический рисунок дерева без листьев.
8.Рефлексия
Какие качества присущи толерантной личности?
Трудно ли поставить себя на место другого человека и понять его переживание?
Наш класс – это маленькая семья. И хотелось бы, чтобы в нашей семье всегда царили доброта, уважение, взаимопонимание, не было ни ссор, ни конфликтов. презентация.pptx (слайд5)
Толерантность в 1 классе доклад, проект
- Главная
- Разное
- Образование
- Спорт
- Естествознание
- Природоведение
- Религиоведение
- Французский язык
- Черчение
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Геометрия
- Детские презентации
- Информатика
- История
- Литература
- Математика
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, фоны, картинки для презентаций
- Экономика
Презентация на тему Толерантность в 1 классе, предмет презентации: Детские презентации. Этот материал в формате pptx (PowerPoint) содержит 19 слайдов, для просмотра воспользуйтесь проигрывателем. Презентацию на заданную тему можно скачать внизу страницы, поделившись ссылкой в социальных сетях! Презентации взяты из открытого доступа или загружены их авторами, администрация сайта не отвечает за достоверность информации в них, все права принадлежат авторам презентаций и могут быть удалены по их требованию.
Этот материал в формате pptx (PowerPoint) содержит 19 слайдов, для просмотра воспользуйтесь проигрывателем. Презентацию на заданную тему можно скачать внизу страницы, поделившись ссылкой в социальных сетях! Презентации взяты из открытого доступа или загружены их авторами, администрация сайта не отвечает за достоверность информации в них, все права принадлежат авторам презентаций и могут быть удалены по их требованию.
Толерантность спасет мир!
Презентацию подготовили ученики
1 «В» класса под руководством классного руководителя
Полужаевой О.А.
Цель проекта
формирование у учащихся навыков толерантных отношений
Задачи проекта
Познакомить детей с понятиями «толерантность», «толерантный человек»;
Выявить основные черты толерантности;
Определить качества толерантного человека;
Сформировать правильное представление о толерантном поведении;
Продолжить формирование толерантного отношения учащихся между собой;
Развивать коммуникативные навыки учеников, учить их сотрудничеству и партнерству в совместных делах;
Воспитывать чувство уважения к окружающим людям, к их мнению, образу жизни и поведению;
Воспитывать умение детей слаженно и дружно работать в группах.
Гипотеза проекта
Создание благоприятных (со стороны толерантности) условий образовательной среды приведет к формированию толерантных качеств личности выпускника лицея, начиная с первого класса
А каким быть легче – добрым или злым?
Быть легче добрым или злым?
Наверно легче злым.
Быть добрым – значит отдавать
Свое тепло другим.
Быть добрым – значит понимать
И близких, и чужих,
И радости порой не знать
Заботясь о других.
Конечно доброму трудней
Но все же посмотри:
Как много у него друзей,
А злой всегда один.
Арабские эмираты
Толерантность — снисхождение, милосердие, всепрощение, умение принимать других такими, какие они сеть и прощать.
Китай
Толерантность — принимать других такими, какие они есть, быть великодушными по отношению к другим.
Испания
Толерантность — способность принимать решения или мнения, отличные от собственных.
Россия
Толерантность — умение терпеть (мириться с чем-либо), принимать (признавать) существование кого-либо, примирять, приводить в соответствие с самим собой по отношению к кому-либо
Посмотрите на картинку: вороны черные и только одна белая, но им хорошо вместе.
Они весело танцуют и поют, и совсем неважно кто и какого цвета.
Толерантность – это способность признавать отличные от своего собственного мнения. Мы допускаем, что кто-то может думать иначе, или действовать иначе, нежели ты сам .
Посмотрите на картинку: мы разные – но мы дружим!», хотя мальчик ходит на руках, а девочка как обычно, так как мы привыкли.
Толерантность – это благосклонность и уважение к другому.
Посмотрите: какой большой слон и какая маленькая мышка, они общаются, дружат.
Большой слон не обижает маленькую мышку. Они равны, не смотря на разницу в размерах.
Проведя анкетирование мы пришли к затруднению в следующих вопросах:
— Злит ли тебя, если кто-то из твоей школы одевается или выглядит не так как все?
— Злит ли тебя, если кто-то из твоих одноклассников не соглашается с твоим мнением?
-Есть ли в классе ребята, которые подшучивают над тобой?
— Бывает ли так, что ты сам подшучиваешь над своими одноклассниками?
Чтобы помочь ребятам разрешить эти вопросы, мы создали буклет, в котором поместили самые важные правила толерантности. А еще сделали дерево толерантности.
А еще сделали дерево толерантности.
Мы разные, но мы вместе!
По итогам проекта мы пришли к следующим выводам:
Ценить такие базовые ценности, как добро, терпение, родина, семья, мир, настоящий друг, справедливость, понимать позицию другого и национальность.
Если каждый будет к друг другу терпим,
Мы сделаем вместе толерантным наш мир!
Давайте жить дружно !
Скачать презентацию
Это сайт презентаций, где можно хранить и обмениваться своими презентациями, докладами, проектами, шаблонами в формате PowerPoint с другими пользователями. Мы помогаем школьникам, студентам, учителям, преподавателям хранить и обмениваться учебными материалами.
Для правообладателей
Обратная связь
Email: Нажмите что бы посмотреть
Толерантность 11 класс презентация, доклад, проект
Добродетель в том, чтобы делать ближним добро и не причинять зла. А делать ближним добро — это делать то, чего желал бы самому себе
Древнеиндийское изречение.
Не для того мы рассуждаем, чтобы знать, что такое добродетель, а для того, чтобы быть хорошими людьми.
Аристотель.
Заглянем в прошлое
На рубеже XVIII-XIX веков во Франции жил некто Талейран-Перигор, князь Беневентский. Он отличился тем, что при разных правительствах (и при революционном, и при Наполеоне, и при короле Людовике XVII) оставался неизменно министром иностранных дел.
Заглянем в прошлое
Это был человек, талантливый во многих областях, но, несомненно, более всего — в умении учитывать настроения окружающих, уважительно к ним относиться, искать решение проблем способом, наименее ущемляющим интересы других людей. И при этом сохранять свои собственные принципы, стремиться к тому, чтобы управлять ситуацией, а не слепо подчиняться обстоятельствам.
И при этом сохранять свои собственные принципы, стремиться к тому, чтобы управлять ситуацией, а не слепо подчиняться обстоятельствам.
tolerancia (испанский) – способность признавать отличные
от своих собственных идеи или мнения;
tolerance (французский) – отношение, при котором допускает
ся, что другие могут думать или действовать иначе, нежели
ты сам;
tolerance (английский) – готовность быть терпимым, снисходи
тельным;
kuan rong (китайский) – позволять, принимать, быть по отношению
к другим великодушным;
tasamul’ (арабский) – прощение, снисходительность, мягкость, милосердие, сострадание, благосклонность, терпение, расположенность к другим;
терпимость (русский) – способность терпеть что-то или кого-то, быть выдержанным, выносливым, стойким, уметь мириться с существованием чего-либо, кого-либо, считаться с мнением других, быть снисходительным.
Толерантность на разных языка мира
(от лат. tolerantia — терпение) —
терпимость к иного рода взглядам, нравам, привычкам;
2) способность организма переносить неблагоприятное влияние того или иного фактора среды.
способность понять другого человека, непохожего на тебя;
взаимопонимание
Современный культурный человек
это не только образованный человек, но человек, обладающий чувством самоуважения и уважаемый окружающими.
Тест: Проверь свою толерантность
Выберите ответ, который считаете правильным. Запишите его
1. Для того, чтобы не было войны…
Нельзя ничего сделать, поскольку войны будут всегда!
Нужно понимать, почему они происходят.
Тест: Проверь свою толерантность
Выберите ответ, который считаете правильным. Запишите его
2. В школе говорят о героях, проявивших толерантность…
Это тебя не интересует.
Ты хочешь узнать об этих героях.
Тест: Проверь свою толерантность
Выберите ответ, который считаете правильным. Запишите его
3. Ты противостоишь насилию…
Насилием.
Ты присоединяешься к другим людям, чтобы сказать НЕТ.
Тест: Проверь свою толерантность
Выберите ответ, который считаете правильным. Запишите его
4. Один товарищ тебя обидел…
Ты мстишь ему.
Ты пытаешься объясниться с ним.
Тест: Проверь свою толерантность
Выберите ответ, который считаете правильным. Запишите его
Запишите его
5. Когда говорят о детях, пострадавших от войны …
Ты об этом услышал и забыл.
Ты ищешь возможность для того, чтобы проявить с ними солидарность..
Тест: Проверь свою толерантность
Выберите ответ, который считаете правильным. Запишите его
6. Ты не согласен с кем-то…
Ты не даёшь ему говорить.
Ты всё-таки слушаешь его..
Тест: Проверь свою толерантность
Выберите ответ, который считаете правильным. Запишите его
7. В классе ты уже ответил…
Тебе хочется отвечать ещё, ты снова отвечаешь.
Ты предоставляешь возможность другим ответить.
Тест: Проверь свою толерантность
Если у тебя одни «b»: Прекрасно! Ты проявляешь большую толерантность. Ты являешься будущим гражданином мира, ответственным и солидарным, активным проводником мира. Объясни своим друзьям, как тебе это удаётся.
Объясни своим друзьям, как тебе это удаётся.
Если у тебя от 3 до 7 «b»: Да! Ты не очень толерантен. Ты слишком стремишься навязать свои идеи, но проявляешь любознательность и у тебя хорошее воображение. Используй эти свои качества для борьбы с нетолерантностью.
Если у тебя менее 3 «b»: Ай-ай-ай! Ты совсем не толерантен! Однако если бы ты был настроен более оптимистично и тебе нравилось дискутировать, ты мог бы стать более счастливым! Давай, приложи ещё немного усилий.
Синквейн
Синквейн – это пятистишие
1 строка – существительное.
2 – два прилагательных к нему.
3 – три глагола.
4 – фраза (крылатое выражение, поговорка), выражающие суть существительного.
5 – существительное (выражение), подводящее итог.
Синквейн
Доброта
Отзывчивый душевный
Поддерживает помогает облагораживает
Доброта спасёт мир
Душевность
Толерантность
Терпимый снисходительный
Принимает располагает прощает
Толерантность способствует уважению
Терпение
Учимся ценить индивидуальность
Напишите о каких-то трех признаках, которые отличают вас от других. Это может быть признание своих достоинств или талантов, жизненных принципов. (Информация должна носить позитивный характер). Записи зачитаем вслух, а остальным нужно будет отгадывать, кто автор утверждений.
Это может быть признание своих достоинств или талантов, жизненных принципов. (Информация должна носить позитивный характер). Записи зачитаем вслух, а остальным нужно будет отгадывать, кто автор утверждений.
Учимся ценить индивидуальность
Сотрудничество
Прощение
Цветок толерантности
Только тогда станешь человеком, когда научишься видеть человека в другом
А.Н.Радищев
1
Первый слайд презентации: Толерантность как культурная норма
Горячих П.В. Д-102
Изображение слайда
2
Слайд 2: Понятие толерантности
В своем широком смысле слово «толерантность» означает терпимость к чужим мнениям и поступкам, способность относиться к ним без раздражения. В этом смысле толерантность является редкой чертой характера. Толерантный человек уважает убеждения других, не стараясь доказать свою исключительную правоту. В более узком смысле понятие «толерантность» используется в медицине. Здесь толерантность – это способность организма без потерь переносить негативные влияния внешней среды.
Толерантность – (от лат. toleratia – терпение) – 1) терпимость к чужому образу жизни, поведению, обычаям, чувствам, мнениям, идеям, верованиям. Толерантность является признаком уверенности в себе и сознания надежности своих собственных позиций, признаком открытого для всех взгляда ни мир, который не боится сравнения с другими точками зрения и не избегает духовной конкуренции; 2) способность организма переносить неблагоприятное влияние того или иного фактора среды.
В отличие от «терпимости» (терпеть — «не противодействуя, не жалуясь, безропотно переносить, сносить что-нибудь бедственное, тяжелое, неприятное»), толерантность — готовность благосклонно признавать, принимать поведение, убеждения и взгляды других людей, которые отличаются от собственных.
Изображение слайда
3
Слайд 3: Понятие культурной нормы
Культурная норма – это общепризнанное правило, образец поведения и действия человека или группы людей в тех или иных сферах общественной жизни, а также инструмент согласования, упорядочивания взаимодействия самих этих сфер в рамках целостного общественного организма.
В связи с этим культурные нормы подразделяют:
особые нормы отдельных сфер человеческой деятельности (производственные и технологические нормы, нормы социального поведения, нормы религиозные, нормы художественной деятельности, нормы познавательной деятельности) или
нормы особых социальных групп (семьи, производственных коллективов, профессиональных групп, классов, этнических групп), а также
общесоциальные нормы, которые регулируют поведение и действия людей в любой сфере человеческой деятельности и в любой общественной группе – нравственные нормы, нормы права и политические нормы.
Изображение слайда
4
Слайд 4: История понятия в России
В русский язык слово «толерантность» вошло сравнительно недавно и в основном использовалось как полный синоним «терпимости». Первые свидетельства об использовании понятия русскими писателями относятся к середине 19 в. Однако ни в «Толковом словаре живого великорусского языка» Владимира Даля (изд. 1863-1866 гг.), ни в «Энциклопедическом словаре Брокгауза и Евфрона » (1901 г.) нет упоминаний о толерантности. Впервые это слово появилось в «Малом энциклопедическом словаре» тех же издателей (1903 г.). Толерантность объяснялась как терпимость к иным религиозным воззрениям.
В Словарь иностранных слов понятие «толерантность» было включено в 1937 г. В Толковом словаре Ушакова за 1940 г. указано производное прилагательное «толерантный», которое синонимично прилагательному «терпимый».
 )
)
Изображение слайда
5
Слайд 5: Основные характеристики толерантности как культурной нормы
Предметные формы субъективности образуют культуру. Предметные формы – идеальные образования, и благодаря своим значениям они участвуют в развитии человеческого сознания. Однако не сами идеальные предметные формы, а именно люди обладатели и создатели этих форм востребуют и задействуют их в осознанной деятельности. Именно люди создают и воспроизводят для своего совместного выживания идеальные средства: язык, обычай, письменность, запрет, культ. Идеалы бесконечны в своем разнообразии, потому что именно они являются средством регулирования и воздействия на общественное и субъективное сознание, способствуя их развитию, преумножая свое разнообразие. Применяя это абстрактное знание на термин «толерантность», она выступает утверждением значимости действия людей. Именно в своем предметном выражении, применимом в различных сферах человеческого регулирования, тем самым являясь идеальным образованием, толерантность выступает как культурная норма и моральная ценность. Ее значения и функции, изменяют свое актуальность в силу исторических событий.
Именно в своем предметном выражении, применимом в различных сферах человеческого регулирования, тем самым являясь идеальным образованием, толерантность выступает как культурная норма и моральная ценность. Ее значения и функции, изменяют свое актуальность в силу исторических событий.
Изображение слайда
6
Слайд 6: Виды и свойства толерантности
Этническая (межнациональная) толерантность – формирование и утверждение принципов толерантности в сфере межэтнических отношений, уважительное отношение к представителям всех наций, способность не переносить недостатки и негативные действия отдельных представителей национальности на всю эту национальность, на весь народ, относиться к любому человеку с позиций «презумпции национальной невиновности».
Расовая толерантность – уважительное отношение к представителям другой расы, отсутствие предубеждений, предрассудков, Политическая толерантность – распространение принципов толерантности на отношение к деятельности различных, в том числе и оппозиционных, политических партий и объединений, а также на взгляды людей, придерживающихся иных политических взглядов
Религиозная толерантность – распространение принципов толерантности на сферу межрелигиозных, межконфессиональных и внутриконфессиональных отношений, уважительное отношение к любой религиозной традиции, признание права любой религии на существование, равенство религий перед законом. Гендерная толерантность – распространение принципов толерантности на отношение к представителям другого пола, непредвзятое отношение к ним, отсутствие чувства превосходства одного пола над другим, равенство в социальных позициях женщин и мужчин, отсутствие дискриминации по гендерным признакам.
Возрастная толерантность – отсутствие неравной оценки качеств и способностей отдельных возрастных групп, непредвзятое отношение к априорным «недостаткам» человека, связанных с возрастом последнего (обвинения молодежи в отсутствии опыта, невоспитанности или неспособность понимать молодых людьми старшего поколения, отсталости последних и т.д.). Одной из форм возрастной толерантности является уважение к людям преклонного возраста.
Гендерная толерантность – распространение принципов толерантности на отношение к представителям другого пола, непредвзятое отношение к ним, отсутствие чувства превосходства одного пола над другим, равенство в социальных позициях женщин и мужчин, отсутствие дискриминации по гендерным признакам.
Возрастная толерантность – отсутствие неравной оценки качеств и способностей отдельных возрастных групп, непредвзятое отношение к априорным «недостаткам» человека, связанных с возрастом последнего (обвинения молодежи в отсутствии опыта, невоспитанности или неспособность понимать молодых людьми старшего поколения, отсталости последних и т.д.). Одной из форм возрастной толерантности является уважение к людям преклонного возраста.
Изображение слайда
7
Слайд 7
Организмическая, или физиологическая толерантность – толерантное отношение к людям с врожденными или приобретенными индивидуальными недостатками. Географическая толерантность – непредвзятое отношение к жителям провинции, глубинки. Отсутствие насмешек по поводу манеры их одеваться, речи и т.д.
Социальная (межклассовая) толерантность – терпимое отношение к социальному расслоению в обществе, терпимое отношение к представителям разных социальных слоев – богатых к бедным и наоборот.
Образовательная толерантность – уважительное отношение друг к другу лиц с разным уровнем образования.
Сексуально-ориентационная толерантность – непредвзятое, спокойное отношение к лицам с нетрадиционной сексуальной ориентацией.
Маргинальная толерантность — терпимость по отношению к людям, относимых к маргинальной прослойке общества (нищие, заключенные, бомжи и т.д.)
Географическая толерантность – непредвзятое отношение к жителям провинции, глубинки. Отсутствие насмешек по поводу манеры их одеваться, речи и т.д.
Социальная (межклассовая) толерантность – терпимое отношение к социальному расслоению в обществе, терпимое отношение к представителям разных социальных слоев – богатых к бедным и наоборот.
Образовательная толерантность – уважительное отношение друг к другу лиц с разным уровнем образования.
Сексуально-ориентационная толерантность – непредвзятое, спокойное отношение к лицам с нетрадиционной сексуальной ориентацией.
Маргинальная толерантность — терпимость по отношению к людям, относимых к маргинальной прослойке общества (нищие, заключенные, бомжи и т.д.)
Изображение слайда
8
Слайд 8
Всем разновидностям толерантности присущи одни и те же критерии. Проявления толерантности в этом случае включают:
равноправие ;
взаимоуважение. доброжелательность и терпимое отношение к представителям различных групп, группам в целом, другим идеям и т.д.
сохранение и развитие культурной самобытности и языков национальных меньшинств
возможность следовать своим традициям
свобода совести и вероисповедания.
сотрудничество и солидарность в решении общих вопросов
позитивная лексика в наиболее острых вопросах межэтнических отношений, в отношениях между полами
доброжелательность и терпимое отношение к представителям различных групп, группам в целом, другим идеям и т.д.
сохранение и развитие культурной самобытности и языков национальных меньшинств
возможность следовать своим традициям
свобода совести и вероисповедания.
сотрудничество и солидарность в решении общих вопросов
позитивная лексика в наиболее острых вопросах межэтнических отношений, в отношениях между полами
Изображение слайда
9
Слайд 9
Сущность толерантности более ярко раскрывается через понимание проявлений ее противоположности – интолерантности. В отличие от термина толерантность, который по своему содержанию шире понятия «терпимость», то возражений против перевода « интолерантности » как «нетерпимости» в литературе обычно не встречается. Интолерантность – это антитеза, антипод толерантности, который основывается на убеждениях человека или какой-либо социальной группы, общности, что только его (их) система взглядов является истинной, что только его (их) образ жизни является правильным и возможным, что та группа, к которой он (они) принадлежат, стоят выше, чем все остальные. По существу это неприятие другого (других) лишь по одной причине: он выглядит, думает, поступает иначе, он живет по-другому. Нетерпимость отличается крайне консервативностью, стремится подавить все, что выходит за привычные установленные в данном социуме рамки. Диапозон ее весьма широк и включает в себя и пренебрежительное отношение к окружающим, и умышленное унижение людей с другим цветом кожи и т.д. К сожалению, даже не все сознают, что их можно отнести к нетолерантному типу личности.
В психологии рассмотрели основные четры и отличия:
По существу это неприятие другого (других) лишь по одной причине: он выглядит, думает, поступает иначе, он живет по-другому. Нетерпимость отличается крайне консервативностью, стремится подавить все, что выходит за привычные установленные в данном социуме рамки. Диапозон ее весьма широк и включает в себя и пренебрежительное отношение к окружающим, и умышленное унижение людей с другим цветом кожи и т.д. К сожалению, даже не все сознают, что их можно отнести к нетолерантному типу личности.
В психологии рассмотрели основные четры и отличия:
Изображение слайда
10
Слайд 10
1. Знание самого себя.
Толерантные люди
Относятся к себе критически, сами стараются разобраться в своих проблемах в своих достоинствах и недостатках
Интолерантные люди.
Замечают у себя только достоинства, обвиняют других.
2. Защищенность.
Толерантный человек уверен в себе; убежден, что справится со всем. Это важное условие для формирования толерантной личности.
Опасается своего социального окружения и самого себя: во всем видит угрозу.
3. Ответственность.
Толерантный человек не перекладывает ответственность на других, сам отвечает за свои за свои поступки.
Интолерантный человек считает, что происходящие события от него не зависят, следовательно снимает с себя ответственность за происходящее вокруг. Он считает, что ни он причиняет зло, а ему вредят.
4. Способность к эмпатии
Толерантный человек может правильно оценить и себя и интолерантного человека.
Интолерантный человек оценивает окружающих по своему образу и подобию.
5. Авторитаризм
Толерантный человек предпочитает жить в демократическом, свободном обществе.
Интолерантный человек предпочитает жить в авторитарном обществе с жесткой властью.
Это важное условие для формирования толерантной личности.
Опасается своего социального окружения и самого себя: во всем видит угрозу.
3. Ответственность.
Толерантный человек не перекладывает ответственность на других, сам отвечает за свои за свои поступки.
Интолерантный человек считает, что происходящие события от него не зависят, следовательно снимает с себя ответственность за происходящее вокруг. Он считает, что ни он причиняет зло, а ему вредят.
4. Способность к эмпатии
Толерантный человек может правильно оценить и себя и интолерантного человека.
Интолерантный человек оценивает окружающих по своему образу и подобию.
5. Авторитаризм
Толерантный человек предпочитает жить в демократическом, свободном обществе.
Интолерантный человек предпочитает жить в авторитарном обществе с жесткой властью.
Изображение слайда
11
Слайд 11: Педагогическая толерантность
Толерантность может рассматриваться, с одной стороны, как средство достижения поставленных воспитательных и образовательных задач, с другой, — как одна из целей процесса воспитания. Необходимым условием успешности педагогической деятельности является принятие ребенка таким, какой он есть. Толерантный педагог, благодаря особой тактике построения своего поведения по отношению к детям, добивается большей результативности.
Анализируя особенности толерантности учителя в педагогическом процессе, можно определить два вида толерантности педагога: социальная (или социально-психологическая) и психологическая (или психофизиологическая). Наличие социальной толерантности позволяет учителю эффективно взаимодействовать со всеми участниками образовательного процесса, а сформированность психологической толерантности обеспечивает высокую устойчивость учителя к многочисленным профессиональным стрессам и способствует эффективному построению профессиональной карьеры.
Следовательно, под педагогической толерантностью надо понимать владение умениями и навыками толерантного взаимодействия со всеми субъектами образовательного процесса; установку на толерантность как активную позицию по формированию толерантности своей личности, личности учащихся и их родителей; как качество личности; как норма своего поведения, представляющая собой одну из составляющих педагогической этики.
Необходимым условием успешности педагогической деятельности является принятие ребенка таким, какой он есть. Толерантный педагог, благодаря особой тактике построения своего поведения по отношению к детям, добивается большей результативности.
Анализируя особенности толерантности учителя в педагогическом процессе, можно определить два вида толерантности педагога: социальная (или социально-психологическая) и психологическая (или психофизиологическая). Наличие социальной толерантности позволяет учителю эффективно взаимодействовать со всеми участниками образовательного процесса, а сформированность психологической толерантности обеспечивает высокую устойчивость учителя к многочисленным профессиональным стрессам и способствует эффективному построению профессиональной карьеры.
Следовательно, под педагогической толерантностью надо понимать владение умениями и навыками толерантного взаимодействия со всеми субъектами образовательного процесса; установку на толерантность как активную позицию по формированию толерантности своей личности, личности учащихся и их родителей; как качество личности; как норма своего поведения, представляющая собой одну из составляющих педагогической этики. Педагогическая толерантность является социальной категорией и проявляется в установке на принятие другого человека, на эмпатическое понимание, на открытое и доверительное общение.
Педагогическая толерантность является социальной категорией и проявляется в установке на принятие другого человека, на эмпатическое понимание, на открытое и доверительное общение.
Изображение слайда
12
Слайд 12: Заключение
Воспитание толерантной личности является в настоящее время одной из важнейших общественных проблем. Трудности взаимопонимания, которые закономерно возникают у людей вследствие расовых, национальных, возрастных, половых и других различий в ситуации их постоянного интенсивного взаимодействия, ведут к росту психологической напряженности, культурной нетерпимости, межэтнической агрессии, религиозного экстремизма, которые невозможно преодолеть без решающего вклада со стороны психологической науки. Необходимо воспитывать толерантность у детей с малых лет.
На данном промежутке историчекого момента сложно привить толерантность прошлым поколениям ввиду политической позиции прошлого и воспитания. Но время не стоит на месте, так же, как и общество. Необходимо мыслить намного шире, чем привыкли многие из людей ввиду менталитета России. Но, как говорится: «Москва не сразу строилась». Тогда может и менталитет нашей страны может стать намного лучше, если перестанет воспринимать чужое мнение и выбор в штыки, будь то даже элементарно сексуальная ориентация, цвет волос, татуировки. Если общество это все же усвоит, то будет намного меньше конфликтов на почве непонимания.
Но время не стоит на месте, так же, как и общество. Необходимо мыслить намного шире, чем привыкли многие из людей ввиду менталитета России. Но, как говорится: «Москва не сразу строилась». Тогда может и менталитет нашей страны может стать намного лучше, если перестанет воспринимать чужое мнение и выбор в штыки, будь то даже элементарно сексуальная ориентация, цвет волос, татуировки. Если общество это все же усвоит, то будет намного меньше конфликтов на почве непонимания.
Изображение слайда
13
Последний слайд презентации: Толерантность как культурная норма: Список литературы
Ананьев Б.Г. Человек как предмет познания. — СПб.: Питер, 2002. — 288 с.
Асмолов А.Г. Толерантность от утопии к реальности. // На пути к толерантному сознанию. — М., 2000. —с. 5—7.
Асмолов А.Г. Толерантность в общественном сознании России. — М. : ИЭАРАЮ, 1998.
Бондырев С.К., Колесов Д.В. Толерантность. Введение в проблему. — М.: издательство Московского психолого-социального института; Воронеж: издательство НПО «МОДЭК», 2003. —240 с.
Декларация принципов толерантности // Век толерантности, 2001. № 1. — с. 62—68.
Зимняя И.А. Педагогическая психология. Учебное пособие. — Ростов н/Д.: Феникс, 1997. —480 с.
Золотухин В.М. Две концепции толерантности. — Кемерово: Кузбасс. гос. техн. университет. 1999. —63 с.
Кристалл Г. Аффективная толерантность // Журнал практической психологии и психоанализа, 2001. —№ 3.
Крутецкий В.А., Балбасова Е.Т. Педагогические способности и структура, диагностика, условия формирования и развития. — М.: Прометей, 1991. —112 с.
Поваренков Ю.П. Психологическая характеристика профессиональной толерантности учителя // Вопросы психологии внимания: сб. науч. трудов / под ред. проф. В.И. Страхова. — Саратов: изд-во Сарат. ун-та, 2003. — Вып. 21. —256 с.
Шалин В.В. Толерантность (культурная норма и политическая необходимость).
: ИЭАРАЮ, 1998.
Бондырев С.К., Колесов Д.В. Толерантность. Введение в проблему. — М.: издательство Московского психолого-социального института; Воронеж: издательство НПО «МОДЭК», 2003. —240 с.
Декларация принципов толерантности // Век толерантности, 2001. № 1. — с. 62—68.
Зимняя И.А. Педагогическая психология. Учебное пособие. — Ростов н/Д.: Феникс, 1997. —480 с.
Золотухин В.М. Две концепции толерантности. — Кемерово: Кузбасс. гос. техн. университет. 1999. —63 с.
Кристалл Г. Аффективная толерантность // Журнал практической психологии и психоанализа, 2001. —№ 3.
Крутецкий В.А., Балбасова Е.Т. Педагогические способности и структура, диагностика, условия формирования и развития. — М.: Прометей, 1991. —112 с.
Поваренков Ю.П. Психологическая характеристика профессиональной толерантности учителя // Вопросы психологии внимания: сб. науч. трудов / под ред. проф. В.И. Страхова. — Саратов: изд-во Сарат. ун-та, 2003. — Вып. 21. —256 с.
Шалин В.В. Толерантность (культурная норма и политическая необходимость). — Краснодар: Периодика Кубани, 2000. —256 с.
— Краснодар: Периодика Кубани, 2000. —256 с.
Изображение слайда
Page not found — Сайт учителя начальных классов Труновой М. А.
Unfortunately the page you’re looking doesn’t exist (anymore) or there was an error in the link you followed or typed. This way to the home page.
- Главная
- Нормативно- правовая база
- Давайте познакомимся
- Немного о себе
- Мои достижения
- В помощь учителю начальных классов
- Полезные сайты
- Классному руководителю
- Проектная деятельность
- Творческие работы учащихся
- Достижения учащихся
- Страничка для родителей
- Тесты для родителей
- Для Вас, ученики
- словари
- Электронные библиотеки
- тренажёры и сайты для детей
- сайты детям
- Повторяем правила
- УМК «Школа России»
- Технологические карты уроков УМК «Школа России»
- Перевёрнутый класс
- Функциональная грамотность
- Онлайн тесты
- Готовимся к ВПР
- Внеурочная деятельеность
- Достижения учащихся
- Борьба с иноземными захватчиками
- Безопасность прежде всего
- обуч грам звук м и буква М
- песня о мире для заучивания
- обуч гр Знакомство со звуком и буквой Ч
- песня к 8 марта
- nfytw 8 vfhnf
- матем сложение вида +5
- русский Твердые и мягкие согласные звуки.
 Буквы для обозначения твердых и мягких согласных звуков. Смыслоразличительная роль согласных звуков.
Буквы для обозначения твердых и мягких согласных звуков. Смыслоразличительная роль согласных звуков. - чтен В. Осеева «Собака яростно лаяла». И. Токмакова «Купите собаку». Пересказ текста.
- ложки лагерь
- окр мир живая и неживая природа
- чтен Бальмонт Поспевает брусника
- матем Единицы времени. Час. Минута. Определение времени по часам.
- русск корень слова.однокор слова
- русск Как определить гласные звуки?
- матем проверка сложения
- чтен К.И.Чуковский «Путаница»
- русск яз Буквосочетания ЖИ –ШИ, ЧА – ЩА, ЧУ – ЩУ.
- чтен Федорино горе К.И. Чуковский
- матем Письменный приём сложения вида 45+23
- русск Проверь себя. Буквосочетания жи-ши, ча-ща, чу-щу»
- муз Русские народные инструменты
- матем Письменный приём вычитания вида 57-26
- русск сочинение по картине А.С.Степанова «Лоси»
- ТЕХНОЛ Как изготовить несколько одинаковых прямоугольников? Практическая работа: Аппликация с переплетением.

- русск Как отличить звонкие согласные от глухих?
- чтен С.В.Михалков творчество писателя
- чтен С.В.Михалков «Сила воли»
- рисов О чём говорит искусство. Выражение характера изображаемых животных
- окр мир Строение тела человека.
- русск Правописание слов с парным по глухости-звонкости согласным звуком на конце слова или перед согласными
- матем Прямой угол
- чтен С.В.Михалков «Мой щенок»
- музыка Фольклор — на¬родная мудрость
- русск Правописание парных согласных в корне слова.
- ОКР МИР Если хочешь быть здоров.
- матем Письменный приём сложения вида 37+48
- родн русск Составляем развернутое толкование значения слова.
- чтен Творчество А.Л. Барто.Верёвочка
- русск яз Распознавание проверяемых и проверочных слов. Проверка парных согласных.
- матем Письменный приём сложения вида 37+53
- русск Проверка парных согласных. Изложение повествовательного текста.
- окр мир Берегись автомобиля!
- рисов Изображения характера человека: мужской образ
- матем Прямоугольник
- чтен А.
 Л.Барто «Мы не заметили жука»
Л.Барто «Мы не заметили жука» - музыка Обряды и празд¬ники русского народа
- чтен биография Н.Носов
- окр мир Домашние опасности.
- матем Письменный приём сложения вида 87+13.
- родн русск Устанавливаем связь предложений в тексте.
- технолог Какой секрет у подвижных игрушек? Практическая работа: Игрушка качалка.
- Н.Носов «Затейники»
- окр мир Пожар
- чтение Н.Носов «Живая шляпа»
- рисование Изображение характера человека: женский образ
- матем Письменный приём вида 32+8; 40-8
- матем Письменный приём вычитания вида 50-24
- матем Письменный приём вычитания вида 52-24
- русский яз Правописание слов с разделительным мягким знаком.
- чтен Н.Носов «На горке»
- чтен Внеклассное чтение. Н.Носов «Мишкина каша», «Заплатка»
- окр мир На воде и в лесу.
- окр мир Опасные незнакомцы.
- рисов Образ человека и его характер, выраженный в объеме
- музыка Детский музы¬кальный театр.
 Опера
Опера - родной русский Создаем тексты- инстукции и тексты- повествания.
- чтен Ю.Ермолаев «Два пирожных»
- русск Что такое части речи?
- матем Письменный приём вычитания вида 52-24
- окр мир Наша дружная семья.
- русск Что такое имя существительное?
- матем Свойство противоположных сторон прямоугольника
- окр мир В школе.
- чтен В.Осеева «Волшебное слово»
- русск Собственные и нарицательные имена существительные. Заглавная буква в именах, отчествах и фамилиях людей.
- матем Квадрат
- чтен В.Осеева «Хорошее»
- русск Заглавная буква в написаниях кличек животных.
- окр мир Ты и твои друзья.
- матем Конкретный смысл действия умножения.
- матем Приём умножения с помощью сложения.
- чтен В.Лунин. Стихотворение «Я и Вовка»
- русск Единственное и множественное число имён существительных.
- музыка Театр оперы и балета.Волшебная палочка
- матем Периметр прямоугольника
- чтен Осеева «Синие листья»
- матем Умножение на 1 и на 0
- окр мир Посмотри вокруг.

- рисв Выражение характера человека через украшение.
- матем Название компонентов умножения
- чтен Ф.Тютчев «Зима недаром злится…» Ф.Тютчев «Весенние воды»
- муз Опера «Рус-лан и Люд-мила» М. И. Глинки
- окр мир Ориентирование на местности.
- русск Что такое глагол?
- матем Переместительное свойство умножения
- русский Единственное и множественное число глаголов.
- чтен И.Бунин «Матери» (в сокращении)
- музыка Симфониче-ская сказка С. С. Проко-фьева «Петя и волк».
- матем Конкретный смысл деления
- русский Правописание частицы НЕ с глаголами.
- чтен А.Плещеев «В бурю»
- окр мир Формы земной поверхности.
- чтен Э.Мошковская «Я маму мою обидел…»,Е.Благинина «Посидим в тишине»
- матем Названия компонентов деления
- русск яз Что такое текст-повествование?
- окр мир Водные богатства.
- чтен Б.Заходер. «Товарищам детям»
- русск Что такое имя прилагательное?
- чтен А.
 Введенский «Учёный Петя»
Введенский «Учёный Петя» - чтен Д.Хармс «Врун»
- окр мир В гости к весне
- русск Связь имени прилагательного с именем существительным.
- матем Приёмы умножения и деления на 10
- русск яз Прилагательные близкие и противоположные по значению.
- чтен И.Токмакова «Плим», «В чудной стране»
- музыка Сюита М. П. Му¬соргского «Кар¬тинки с вы¬ставки»
- чтен Б.Заходер «Песенки Винни-Пуха»
- матем Задачи с величинами: цена, количество, стоимость.
- русск яз Единственное и множественное число имён прилагательных.
- русск Что такое текст – описание?
- чтен Э.Успенский «Чебурашка» (из сказки «Крокодил Гена и его друзья»)
- чтен Э.Успенский «Если был бы я девчонкой…», «Над нашей квартирой»
- окр мир Россия на карте.
- рисован Линия как средство выражения: ритм линий
- русск Общее понятие о предлоге.
- музыка Звучит не-ста¬реющий Мо¬царт!
- чтен В.Драгунский «Тайное становится явным»
- окр мир Путешествие по Москве
- матем Умножение числа 2.
 Умножение на 2.
Умножение на 2. - технолог Какие бывают нитки? Как они используются? Практическая работа: Птичка из помпона. Цыплёнок из 2 помпонов.
- чтен Г.Остер «Будем знакомы»
- окр мир Московский кремль и Красная площадь
- чтен Внеклассное чтение. Н.Носов «Фантазёры»
- музыка Волшебный цветик семи-цветик. «И все это — И. С. Бах»
- русск яз что такое местоимение?
- матем деление на 2
- окр мир Город на Неве.
- чтен Английские народные песенки «Перчатки», «Храбрецы» (перевод К.Чуковского), «Храбрецы» (перевод С.Маршака)
- русск Что такое местоимение?
- чтен Ш.Перро «Кот в сапогах»
- окр. мир Путешествие по Оке .Проект «Города России»
- русск яз Что такое текст – рассуждение?
- окр мир Путешествие по планете.
- Чтение Ш.Перро «Красная Шапочка»
- чтение Андерсен «Огниво»
- русск яз «Текст» обобщение-повторение
- окр мир Путешествие по материкам
- рисован Ритм пятен как средство выражения.
 Пропорции выражают характер
Пропорции выражают характер - чтен Андерсен «Принцесса на горошине»
- Страны мира.Проект
- внекл чтение Андерсен «Дюймовочка»
- рисов Ритм линий и пятен, цвет, пропорции – средства выразительности
- 3 класс
- музыка Природа и музыка.Романс
- чтение Сказка Сивка-Бурка
- чтение Тютчев «Весенняя гроза», «Листья»
- музыка «Виват, Россия!» Наша слава – русская держава.
- ИЗО Посуда у тебя дома .Изображение праздничного сервиза
- Технология Знакомимся с компьютером. Компьютер – твой помощник.
Исследование МНС-пептидной матрицы центральной толерантности в тимусе человека. антигенпрезентирующих клеток, иммунологи размышляли о природе этих пептидов, особенно у людей. Здесь, чтобы пролить свет на до сих пор неизвестный репертуар пептидов тимуса человека, мы анализируем пептиды, элюированные из изолированных дендритных клеток тимуса, антигенпрезентирующих клеток с обедненными дендритными клетками и целого тимуса.
 Биоинформатический анализ 842 идентифицированных природных лигандов главного комплекса гистосовместимости I и II выявляет значительные перекрестные помехи между путями главного комплекса гистосовместимости класса I и II и различиями в представлении исходного белка между людьми, а также различными антигенпрезентирующими клетками. Кроме того, в здоровом тимусе представлены несколько аутоиммунных и связанных с опухолью пептидов, например, из енолазы и виментина. 302 пептиды получены непосредственно из негативной селекции дендритных клеток, что обеспечивает первое глобальное представление о пептидной матрице в тимусе человека, которая налагает самотолерантность in vivo .
Биоинформатический анализ 842 идентифицированных природных лигандов главного комплекса гистосовместимости I и II выявляет значительные перекрестные помехи между путями главного комплекса гистосовместимости класса I и II и различиями в представлении исходного белка между людьми, а также различными антигенпрезентирующими клетками. Кроме того, в здоровом тимусе представлены несколько аутоиммунных и связанных с опухолью пептидов, например, из енолазы и виментина. 302 пептиды получены непосредственно из негативной селекции дендритных клеток, что обеспечивает первое глобальное представление о пептидной матрице в тимусе человека, которая налагает самотолерантность in vivo .
Введение
Функциональная иммунная система должна быть способна эффективно распознавать патогены и бороться с ними, сохраняя при этом толерантность к собственным компонентам организма. Это достигается за счет центральных и периферических механизмов толерантности. Центральная толерантность к себе навязывается Т-клеткам во время их развития в тимусе, где их обучают быть самореферентными, распознающими пептиды, связанные с молекулами главного комплекса гистосовместимости (MHC), а также самотолерантными, что означает, что они остаются невосприимчивыми к собственным пептидам, представленным на тех же самых молекулах MHC 1,2 .
Чтобы соответствовать обоим критериям, αβ Т-клетки в своем развитии должны пройти две основные контрольные точки, которые контролируются антигенпрезентирующими клетками (APC) в различных микроокружениях тимуса. Эти APC включают корковые (cTEC) и медуллярные эпителиальные клетки тимуса (mTEC), а также как резидентные, так и иммигрирующие дендритные клетки (DC). Во время развития тимоциты должны перестроить свои гены, чтобы экспрессировать функциональный Т-клеточный рецептор (TCR), который может успешно взаимодействовать с собственными лигандами пептид-MHC на cTEC, от которого они затем получат сигнал выживания (положительный отбор). Положительно отобранные тимоциты мигрируют в мозговое вещество тимуса, где они взаимодействуют с комплексами аутопептид-MHC, отображаемыми mTECs и DCs. Т-клетки, экспрессирующие TCR с высоким сродством к этим комплексам, будут подвергаться отрицательному отбору и подвергаться апоптозу. Только Т-клетки с низкой/промежуточной аффинностью TCR к аутопептидам смогут покинуть тимус в качестве наивных или регуляторных Т-клеток 1,2 . Если Т-клетка, которая хорошо распознает аутопептиды, не может быть удалена, например, из-за того, что соответствующий аутобелок не экспрессируется в тимусе или эпитоп разрушается во время процессинга антигена, аутореактивные Т-клетки могут уйти на периферию, где они может активироваться и вызывать аутоиммунное заболевание 3 . Следовательно, взаимодействие между TCR развивающихся тимоцитов и пептидным лигандомом MHC различных APC тимуса имеет решающее значение для формирования репертуара Т-клеток и центральной толерантности.
Если Т-клетка, которая хорошо распознает аутопептиды, не может быть удалена, например, из-за того, что соответствующий аутобелок не экспрессируется в тимусе или эпитоп разрушается во время процессинга антигена, аутореактивные Т-клетки могут уйти на периферию, где они может активироваться и вызывать аутоиммунное заболевание 3 . Следовательно, взаимодействие между TCR развивающихся тимоцитов и пептидным лигандомом MHC различных APC тимуса имеет решающее значение для формирования репертуара Т-клеток и центральной толерантности.
Миелоидные ДК (мДК) в тимусе были первыми клетками, ответственными за удаление аутореактивных Т-клеток в тимусе, и имеют решающее значение для поддержания толерантности к себе 4,5 . Хотя, в отличие от mTEC, они не специализируются на экспрессии тканеспецифических антигенов, таких как проинсулин, они могут эффективно захватывать их из mTEC и представлять их развивающимся Т-клеткам, вызывая их делецию, процесс, который включает экспрессию аутоиммунного регулятора (AIRE). в мТЭК 6,7 . Роль плазмоцитоидных DCs (pDCs) в отрицательном отборе была описана совсем недавно 8 и В-клетки тимуса могут также вносить вклад в отрицательный отбор 9 . Хотя до сих пор нет прямых доказательств того, что разные подмножества APC тимуса представляют разные наборы пептидов, есть два убедительных косвенных доказательства, подтверждающих это представление. Во-первых, разные APC экспрессируют разные наборы собственных белков. В дополнение к конститутивно экспрессируемым белкам домашнего хозяйства и белкам, специфичным для клеточного типа, mTEC экспрессируют антигены, ограниченные тканями, такие как инсулин, под контролем фактора транскрипции AIRE 9.0011 10,11 . Кроме того, происходящие из крови mDCs и pDCs могут импортировать периферические антигены в тимус, где они удаляют аутореактивные тимоциты 8,12 . Во-вторых, как MHC I, так и II механизм процессинга антигенов различаются между разными APC тимуса, что должно влиять на генерируемые эпитопы 13,14,15,16 .
в мТЭК 6,7 . Роль плазмоцитоидных DCs (pDCs) в отрицательном отборе была описана совсем недавно 8 и В-клетки тимуса могут также вносить вклад в отрицательный отбор 9 . Хотя до сих пор нет прямых доказательств того, что разные подмножества APC тимуса представляют разные наборы пептидов, есть два убедительных косвенных доказательства, подтверждающих это представление. Во-первых, разные APC экспрессируют разные наборы собственных белков. В дополнение к конститутивно экспрессируемым белкам домашнего хозяйства и белкам, специфичным для клеточного типа, mTEC экспрессируют антигены, ограниченные тканями, такие как инсулин, под контролем фактора транскрипции AIRE 9.0011 10,11 . Кроме того, происходящие из крови mDCs и pDCs могут импортировать периферические антигены в тимус, где они удаляют аутореактивные тимоциты 8,12 . Во-вторых, как MHC I, так и II механизм процессинга антигенов различаются между разными APC тимуса, что должно влиять на генерируемые эпитопы 13,14,15,16 . Таким образом, различные АПК вносят вклад в разнообразие «иммунологической самости», проявляемой в тимусе, формируя характеристики репертуара зрелых Т-клеток каждого человека.
Таким образом, различные АПК вносят вклад в разнообразие «иммунологической самости», проявляемой в тимусе, формируя характеристики репертуара зрелых Т-клеток каждого человека.
Наши текущие знания о репертуаре собственных пептидов в тимусе получены в основном из исследований на моделях трансгенных мышей TCR, где экспрессия определенного антигена является предпосылкой для положительного/отрицательного отбора. Однако ни одно из этих исследований не дает доказательств того, что соответствующие пептиды действительно способствуют отрицательному отбору в физиологических условиях. В начале девяностых Marrack et al. 17 успешно идентифицировано ~20 пептидов, связанных с MHC II, в объединенных тимусах нескольких сотен мышей, и известно несколько естественных лигандов для положительной селекции клеток OT-I мыши 18,19 . Совсем недавно другая группа элюировала пептиды из цельного тимуса мыши, используя биоинформатику, чтобы сузить круг вероятных лигандов MHC I 20 . У людей сообщалось о представлении эпитопа проинсулина на ДК тимуса с использованием моноклональных антител 21 , но помимо этого репертуар пептидов тимуса человека неизвестен.
У людей сообщалось о представлении эпитопа проинсулина на ДК тимуса с использованием моноклональных антител 21 , но помимо этого репертуар пептидов тимуса человека неизвестен.
В этой работе мы решили исследовать лигандом МНС тимуса человека. Мы описываем подробный анализ пептидного репертуара, представленного молекулами MHC класса I и класса II в здоровом тимусе человека, как для всего тимуса, так и для изолированных mDC тимуса. Таким образом, здесь мы представляем первые данные о встречающихся в природе пептидах, по которым Т-клетки человека отбирают в физиологических условиях, включая пептиды из ДК, которые ответственны за делецию аутореактивных Т-клеток.
Результаты
Идентификация пептидов, презентируемых АПК тимуса
Чтобы ответить на вопрос, какие пептиды естественным образом презентируются развивающимся тимоцитам для индукции негативной селекции в физиологических условиях у людей, мы проанализировали пептиды, элюированные из обоих MHC I (пан-класс I человека). лейкоцитарного антигена; HLA) и II (HLA-DR) комплексов из изолированных мДК тимуса методом масс-спектрометрии и сравнили их с репертуаром MHC-связанных пептидов, элюированных из клеток APC-обогащенных, mDC-обедненных (содержащих pDCs, TECs и другие АПК тимуса, то есть лиганды как для положительного, так и для отрицательного отбора), а также фракции цельного тимуса (рис. 1).
лейкоцитарного антигена; HLA) и II (HLA-DR) комплексов из изолированных мДК тимуса методом масс-спектрометрии и сравнили их с репертуаром MHC-связанных пептидов, элюированных из клеток APC-обогащенных, mDC-обедненных (содержащих pDCs, TECs и другие АПК тимуса, то есть лиганды как для положительного, так и для отрицательного отбора), а также фракции цельного тимуса (рис. 1).
Изображение в натуральную величину
Одной из основных проблем анализа пептидов, представленных АПК тимуса, на основе МС является их низкая частота встречаемости и их очистка в количествах, достаточных для проведения лигандомного анализа. Как правило, количество клеток, необходимых для таких экспериментов, составляет порядка 10 900 11 8 900 12 , что ограничивает возможности анализа первично очищенных типов клеток. В наших экспериментах мы идентифицировали в общей сложности 842 пептида из различных клеточных препаратов восьми исследованных отдельных тимусов, из которых 621 был элюирован из молекул MHC I и 221 из молекул MHC II. В отдельных препаратах mDC мы успешно идентифицировали до 90 пептидных последовательностей, элюированных из молекул MHC I и до 48 из MHC II всего от 30 × 10 6 клеток от четырех разных доноров (таблица 1). Полные списки пептидных последовательностей, полученных из различных образцов, и их исходных белков приведены в дополнительных данных 1.
В отдельных препаратах mDC мы успешно идентифицировали до 90 пептидных последовательностей, элюированных из молекул MHC I и до 48 из MHC II всего от 30 × 10 6 клеток от четырех разных доноров (таблица 1). Полные списки пептидных последовательностей, полученных из различных образцов, и их исходных белков приведены в дополнительных данных 1.
Полноразмерная таблица
Перекрестные помехи между путями MHC I и MHC II
Пептиды, представленные MHC I и II, происходят из разных клеточных компартментов. Как правило, ядерные и цитозольные белки представлены молекулами MHC I, в то время как белки, которые присутствуют в везикулярных компартментах или клеточной мембране или поглощаются из внеклеточной среды, представлены молекулами MHC II. Однако, особенно в профессиональных APC, между этими двумя путями 22 существует значительная перекрестная связь. Механизмы, с помощью которых цитозольные/ядерные белки получают доступ к пути MHC II, включают аутофагию и поглощение апоптотических клеток 23,24 . И наоборот , белки или пептиды могут перемещаться из лизосомы в цитозоль и, таким образом, получать доступ к пути MHC I 22 . В дополнение к этому внутриклеточному взаимодействию, DCs могут поглощать антигены из других клеток, таких как mTECs, дополнительно расширяя их пептидный репертуар 25,26 .
Механизмы, с помощью которых цитозольные/ядерные белки получают доступ к пути MHC II, включают аутофагию и поглощение апоптотических клеток 23,24 . И наоборот , белки или пептиды могут перемещаться из лизосомы в цитозоль и, таким образом, получать доступ к пути MHC I 22 . В дополнение к этому внутриклеточному взаимодействию, DCs могут поглощать антигены из других клеток, таких как mTECs, дополнительно расширяя их пептидный репертуар 25,26 .
Чтобы получить представление о происхождении представленных пептидов, а также о динамике антигенных перекрестных помех между путями представления MHC в mDC тимуса и не-mDC APC, мы провели автоматизированный анализ базы данных, дополненный ручной аннотацией исходных белков и их субклеточная локализация. В зависимости от их топологического происхождения белки были отнесены либо к классическому пути МНС класса I (цитозольный, ядерный), либо к пути МНС класса II (внеклеточный, секреторный, эндоцитарный и клеточная мембрана), митохондриальному и компартменту ЭР (рис. 2а). . Как и ожидалось, большинство лигандов MHC I, представленных как ДК, так и не-ДК АПК, происходят из цитозольных/ядерных белков, которые в норме не пересекаются с эндоцитарным компартментом, таких как гистоны, белки цитоскелета (актин, тубулин и фасцин), метаболические ферменты. (протеинфосфатаза 1, гидроксипируватредуктаза) и цитозольные шапероны (белки теплового шока). Пептиды, связанные с МНС класса II, происходят из белков, присутствующих в различных клеточных компартментах, включая внеклеточную среду, плазматическую мембрану и везикулярный компартмент, при этом мембранные белки являются наиболее распространенными генераторами пептидов (в основном из белков МНС класса II) в соответствии с предыдущими исследованиями. другие типы клеток 24 . Значительное количество пептидов, представленных MHC II, происходит из белков крови или сыворотки, таких как сывороточный альбумин, гемоглобин и аполипопротеин A2, что является прямым доказательством того, что ДК тимуса пробуют кровоток.
2а). . Как и ожидалось, большинство лигандов MHC I, представленных как ДК, так и не-ДК АПК, происходят из цитозольных/ядерных белков, которые в норме не пересекаются с эндоцитарным компартментом, таких как гистоны, белки цитоскелета (актин, тубулин и фасцин), метаболические ферменты. (протеинфосфатаза 1, гидроксипируватредуктаза) и цитозольные шапероны (белки теплового шока). Пептиды, связанные с МНС класса II, происходят из белков, присутствующих в различных клеточных компартментах, включая внеклеточную среду, плазматическую мембрану и везикулярный компартмент, при этом мембранные белки являются наиболее распространенными генераторами пептидов (в основном из белков МНС класса II) в соответствии с предыдущими исследованиями. другие типы клеток 24 . Значительное количество пептидов, представленных MHC II, происходит из белков крови или сыворотки, таких как сывороточный альбумин, гемоглобин и аполипопротеин A2, что является прямым доказательством того, что ДК тимуса пробуют кровоток.
Клеточная локализация ( a ) и биологические процессы ( b ) категорий белков, представленных графически в процентах, основаны на n = количество отдельных белков, идентифицированных в каждом препарате АРС тимуса.
Увеличить
Интересно, что значительная часть лигандов MHC II происходит из исходных белков, локализованных в цитозоле или ядре (35,4% в ДК и 21,8% в не-ДК), которые топологически отделены от МНС класса II. путь. Точно так же 22,4% лигандов MHC I на ДК происходят из везикулярного / внеклеточного компартмента (по сравнению с 15,5% во фракции, обедненной ДК, и 11,4% в полных образцах тимуса). Периферические ДК очень эффективны при перекрестной презентации эндоцитированных антигенов на MHC I и цитозольных/ядерных антигенов на MHC II. Следовательно, крайне важно индуцировать центральную толерантность также и к перекрестно представленным собственным белкам. Высокая степень перекрестного взаимодействия между двумя путями, которые мы наблюдаем в ДК тимуса, а также, в меньшей степени, во фракции, обедненной ДК, отражает эту необходимость избегать периферических аутоиммунных ответов путем делеции Т-клеток, которые распознают перекрестно представленные собственные клетки. -белки.
Высокая степень перекрестного взаимодействия между двумя путями, которые мы наблюдаем в ДК тимуса, а также, в меньшей степени, во фракции, обедненной ДК, отражает эту необходимость избегать периферических аутоиммунных ответов путем делеции Т-клеток, которые распознают перекрестно представленные собственные клетки. -белки.
Недавнее исследование подчеркнуло важность аутофагии для презентации MHC II во время индукции центральной толерантности. У мышей генетическая интерференция с аутофагией приводит к измененному отбору определенных специфичностей CD4 + Т-клеток, которые вызывают репертуар аутореактивных Т-клеток 27 . В большинстве исследований до сих пор рассматривалась значимость аутофагии in vivo , главным образом, в TEC, а не в тимических DC, но было показано, что в периферических DCs аутофагосомы сливаются с загрузочным отсеком MHC II 23 . Наши данные о пептидах показывают значительную презентацию пептидов, полученных из ядерных / цитозольных, а также некоторых митохондриальных белков на MHC класса II mDC тимуса, что указывает на роль аутофагии в отборе проб лигандов MHC II в этих клетках. Альтернативно, эти белки могут входить в путь MHC II путем поглощения из других клеток, таких как апоптотические тимоциты или живые/умирающие mTEC 7,28,29 , или путем переноса комплексов пептид-MHC из mTEC 30 .
Альтернативно, эти белки могут входить в путь MHC II путем поглощения из других клеток, таких как апоптотические тимоциты или живые/умирающие mTEC 7,28,29 , или путем переноса комплексов пептид-MHC из mTEC 30 .
Природа DC-представленных пептидов
Анализ биологических процессов, связанных с исходными белками пептидов, выявил заметную долю белков, участвующих в «иммунном ответе» и «процессинге/презентации антигена», биологических функциях, отражающих природу тимуса как иммунного органа (рис. 2b). ). Наиболее широко представленным классом белков в этих группах были молекулы HLA, и неудивительно, что их присутствие было более выраженным в образцах DC и DC-истощенных (обогащенных другими APC тимуса) по сравнению с образцами цельного тимуса. Большинство оставшихся природных лигандов класса I и II происходят из белков «домашнего хозяйства» (передача сигналов, трансляция, метаболизм, клеточный цикл и пролиферация). Интересно, что пептиды, происходящие из белков, связанных с метаболическими процессами, преимущественно представлены на молекулах МНС класса II, а пептиды, полученные из белков, участвующих в клеточном цикле, росте, пролиферации и дифференцировке, более обильно представлены на молекулах МНС класса I.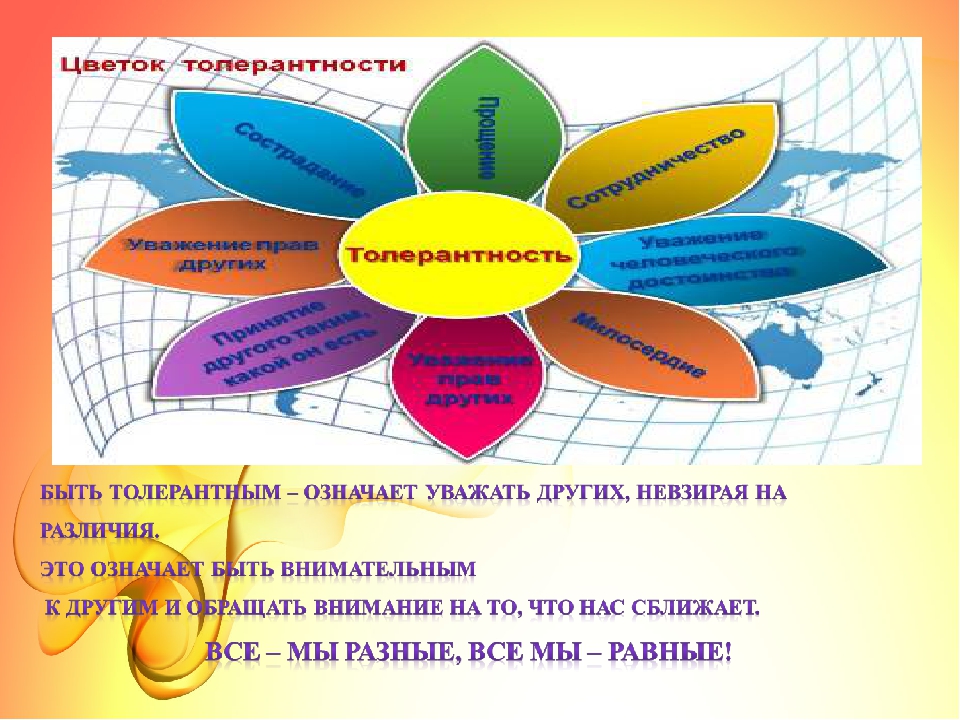 Объяснение предпочтительной презентации пептидов, полученных из белков, участвующих в этих процессах, MHC I заключается в том, что в этих группах большинство пептидов происходят из ядерных и цитозольных белков, участвующих в поддержании клеточной структуры (например, цитоскелета) и транскрипции. Они обычно расщепляются протеасомой и представлены классическим путем MHC I 22 . Более пристальный взгляд на белки, связанные с метаболическими процессами, показал, что многие из идентифицированных пептидов, связанных с MHC II, в этой группе происходят из цитозольных белков. Источниками этих пептидов является значительное количество долгоживущих конститутивных метаболических ферментов, таких как глицеральдегид-3-фосфатдегидрогеназа и фосфоглицераткиназа 1. Интересно, что предполагается, что долгоживущие белки утилизируются путем аутофагической, а не протеасомной деградации белков 31 . Было показано, что аутофагическая деградация клеточного материала вносит значительный вклад в пептидный пул, представленный MHC II (refs 23, 24).
Объяснение предпочтительной презентации пептидов, полученных из белков, участвующих в этих процессах, MHC I заключается в том, что в этих группах большинство пептидов происходят из ядерных и цитозольных белков, участвующих в поддержании клеточной структуры (например, цитоскелета) и транскрипции. Они обычно расщепляются протеасомой и представлены классическим путем MHC I 22 . Более пристальный взгляд на белки, связанные с метаболическими процессами, показал, что многие из идентифицированных пептидов, связанных с MHC II, в этой группе происходят из цитозольных белков. Источниками этих пептидов является значительное количество долгоживущих конститутивных метаболических ферментов, таких как глицеральдегид-3-фосфатдегидрогеназа и фосфоглицераткиназа 1. Интересно, что предполагается, что долгоживущие белки утилизируются путем аутофагической, а не протеасомной деградации белков 31 . Было показано, что аутофагическая деградация клеточного материала вносит значительный вклад в пептидный пул, представленный MHC II (refs 23, 24). В соответствии с нашими результатами и в поддержку этой идеи, глицеральдегид-3-фосфатдегидрогеназа ранее была идентифицирована как источник природных MHC класса II, но не лигандов класса I 24,32 . Следовательно, аутофагическая деградация долгоживущих метаболических белков может объяснить, почему эти цитозольные белки представлены MHC класса II, а не класса I. Наблюдаемое широкое клеточное функциональное распределение белков отражает широкий спектр белков, которые служат матрицей для собственных белков. представление в тимусе.
В соответствии с нашими результатами и в поддержку этой идеи, глицеральдегид-3-фосфатдегидрогеназа ранее была идентифицирована как источник природных MHC класса II, но не лигандов класса I 24,32 . Следовательно, аутофагическая деградация долгоживущих метаболических белков может объяснить, почему эти цитозольные белки представлены MHC класса II, а не класса I. Наблюдаемое широкое клеточное функциональное распределение белков отражает широкий спектр белков, которые служат матрицей для собственных белков. представление в тимусе.
Пептиды различаются между отдельными людьми и APC
Было высказано предположение, что разные типы APC тимуса представляют собой разные неперекрывающиеся наборы собственных белков, несмотря на существование путей, обеспечивающих перенос белков/пептидов между клетками, и несмотря на то, что многие Основные клеточные компоненты, такие как метаболические ферменты и структурные белки, должны экспрессироваться на одинаковых уровнях в большинстве клеток. Одной из причин этого могут быть различия в механизмах процессинга антигенов между разными типами клеток 9.0011 13,14,15,16 . Чтобы ответить на этот вопрос, мы сравнили исходные белки пептидов, представленных на DC, с белками DC-истощенной фракции (обогащенной для других APC тимуса, состав см. В дополнительной таблице S1 и дополнительном рисунке S1). Всего мы идентифицировали 302 пептида, происходящих из 187 различных белков на ДК, и 265 пептидов из 184 различных белков во фракции, истощенной ДК, 71 из которых присутствовал в обеих фракциях (рис. 3а). К ним относятся белки, такие как молекулы HLA, гистоны, ядерный антиген пролиферирующих клеток, актин, пируваткиназа, сывороточный альбумин и β2-микроглобулин. Хотя около 1/3 белкового пула было общим для DC и не-DC клеток, большинство идентифицированных белков были уникальными для каждой группы, что подтверждает идею о том, что разные APC тимуса представляют разные пептиды.
Одной из причин этого могут быть различия в механизмах процессинга антигенов между разными типами клеток 9.0011 13,14,15,16 . Чтобы ответить на этот вопрос, мы сравнили исходные белки пептидов, представленных на DC, с белками DC-истощенной фракции (обогащенной для других APC тимуса, состав см. В дополнительной таблице S1 и дополнительном рисунке S1). Всего мы идентифицировали 302 пептида, происходящих из 187 различных белков на ДК, и 265 пептидов из 184 различных белков во фракции, истощенной ДК, 71 из которых присутствовал в обеих фракциях (рис. 3а). К ним относятся белки, такие как молекулы HLA, гистоны, ядерный антиген пролиферирующих клеток, актин, пируваткиназа, сывороточный альбумин и β2-микроглобулин. Хотя около 1/3 белкового пула было общим для DC и не-DC клеток, большинство идентифицированных белков были уникальными для каждой группы, что подтверждает идею о том, что разные APC тимуса представляют разные пептиды.

На диаграммах показано количество различных исходных белков, идентифицированных в различных клеточных препаратах, а также перекрытие между образцами. ( a ) Перекрытие идентифицированных белков между mDC и mDC-истощенными (обогащенными для других APC) фракциями (отдельные доноры вместе взятые). Индивидуальные вариации белков, представленные на ( b ) ДК четырех разных доноров и ( c ) обедненные ДК фракции трех индивидуумов соответственно.
Изображение в натуральную величину
Данные по целому тимусу указывают на большую степень изменчивости исходных белков, из которых пептиды были представлены у разных доноров (дополнительная рис. S2), вероятно, частично из-за различий в предпочтениях связывания HLA и экспрессии белков. Количественные различия в уровнях экспрессии отдельных комплексов MHC-пептид также могут играть роль, поскольку наш экспериментальный подход преимущественно обнаруживает пептиды с высоким содержанием, особенно когда исходный материал ограничен. Чтобы исключить возможность того, что наблюдаемое ограниченное перекрытие было вызвано проблемами технической воспроизводимости, а не bona fide биологическая изменчивость, мы провели обширные проверочные эксперименты, которые показали, что техническая воспроизводимость составляет порядка 85%, в то время как перекрытие между образцами, полученными от разных доноров, было намного ниже (дополнительный рисунок S3), демонстрируя, что наблюдаемые различия были действительно из-за биологической, а не технической изменчивости.
Чтобы исключить возможность того, что наблюдаемое ограниченное перекрытие было вызвано проблемами технической воспроизводимости, а не bona fide биологическая изменчивость, мы провели обширные проверочные эксперименты, которые показали, что техническая воспроизводимость составляет порядка 85%, в то время как перекрытие между образцами, полученными от разных доноров, было намного ниже (дополнительный рисунок S3), демонстрируя, что наблюдаемые различия были действительно из-за биологической, а не технической изменчивости.
Чтобы выяснить, наблюдается ли эта изменчивость и для собственных белков, непосредственно участвующих в негативной селекции Т-клеток, мы сравнили исходные белки пептидов, элюированных из ДК тимуса от разных доноров (рис. 3б). Из 187 белков, чьи пептиды были представлены mDC, только 27 были обнаружены более чем у одного донора, и только один (HLA-DRB1) был общим для четырех отдельных образцов тимических DC. Пептиды из семи белков (HLA-DRA, сывороточный альбумин, цепь тубулина β 2 A, серин/треонин-протеинфосфатаза PP1, астроцитарный фосфопротеин РЕА-15, ингибитор bax 1, преобразователь сигнала и активатор транскрипции 1-α/β) могут быть обнаружены в трех из четырех образцов ДК, но пептиды из подавляющего большинства белков (160) были обнаружены только в одном образце, что отражает высокую межиндивидуальную вариабельность пептидной матрицы для отрицательного отбора.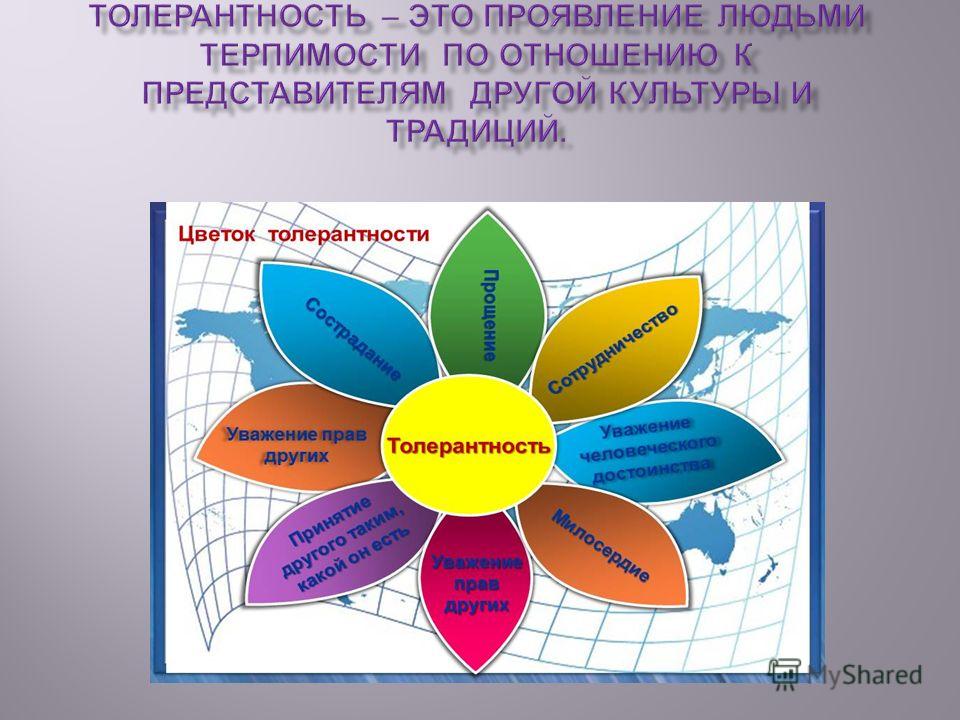 Аналогичная картина наблюдается в пуле пептидов, полученных из ДК-истощенных (рис. 3в).
Аналогичная картина наблюдается в пуле пептидов, полученных из ДК-истощенных (рис. 3в).
Для дальнейшего изучения различий между HLA-лигандомом ДК и не-ДК мы напрямую сравнили исходные белки (рис. 4а) и белковые пептиды (рис. 4б), которые присутствуют в соответствующих фракциях из одного и того же доноров, исключая вклад HLA-связывания в вариабельность пептидов между фракциями. Интересно, что только около пятой части обнаруженных белков были представлены как ДК, так и не-ДК АПК, а для донора ТТ259 перекрытие было еще меньше. Таким образом, мы демонстрируем высокую межиндивидуальную вариабельность лигандома MHC, и, кроме того, наши результаты согласуются с представлением о том, что разные, но перекрывающиеся спектры собственных белков на разных АПК тимуса приводят к частично неповторяющимся вкладам разных АПК в отрицательные выбор.
Рисунок 4: Сравнение фракций mDC и non-mDC у каждого донора. Перекрытие ( a ) исходных белков и ( b ) представленных пептидов между mDC и mDC-истощенными клетками в трех отдельных тимусах.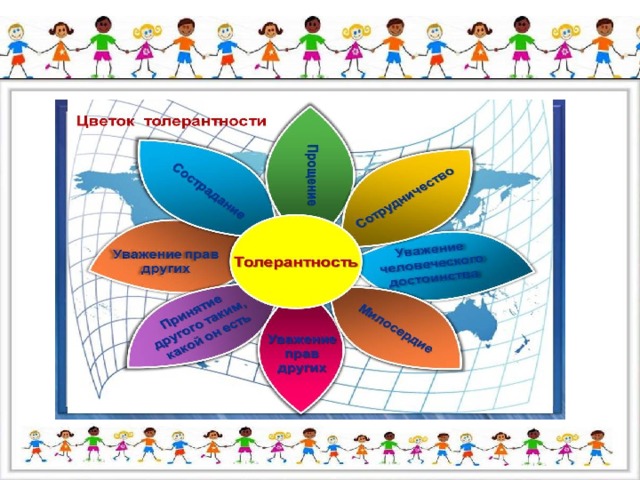
Изображение в полный размер
Т-клеточные эпитопы, связанные с аутоиммунитетом
Неполное представление полного спектра собственных белков («иммунологических собственных») на молекулах МНС мТЭК и ДК приведет к отсутствию центральной толерантности к этим эпитопы, которые не отображаются, увеличивая склонность к аутореактивности среди зрелых Т-клеток на периферии 1,33 .
Поэтому нас особенно интересовало, сможем ли мы идентифицировать пептиды, соответствующие известным Т-клеточным эпитопам, которые были описаны как мишени при аутоиммунных заболеваниях человека. Поиск в литературе дал ряд аутоантигенов Т-клеток, которые также можно было обнаружить в наших образцах (таблица 2). Таблица включает только Т-клеточные аутоантигены, реактивность которых была экспериментально подтверждена у людей, такие как виментин (ревматоидный артрит, саркоидоз) 34,35 , АТФ-синтаза (саркоидоз) 35 , энолаза 2 (рассеянный склероз) 36 и белок теплового шока 70 (сахарный диабет 1 типа) 37 . Для тех аутоантигенов, для которых сообщалось о реактивности против определенных эпитопов (а не всего белка), эпитопы были идентичны или содержались в пептидных последовательностях, которые мы идентифицировали в наших образцах, в основном на DC. Это ясно демонстрирует представление этих эпитопов в нормальном тимусе человека, что должно приводить к делеции аутореактивных Т-клеток. Поэтому мы предполагаем, что у пациентов с аутоиммунным заболеванием презентация этих эпитопов может быть нарушена, что приводит к нарушению центральной толерантности к этим эпитопам.
Для тех аутоантигенов, для которых сообщалось о реактивности против определенных эпитопов (а не всего белка), эпитопы были идентичны или содержались в пептидных последовательностях, которые мы идентифицировали в наших образцах, в основном на DC. Это ясно демонстрирует представление этих эпитопов в нормальном тимусе человека, что должно приводить к делеции аутореактивных Т-клеток. Поэтому мы предполагаем, что у пациентов с аутоиммунным заболеванием презентация этих эпитопов может быть нарушена, что приводит к нарушению центральной толерантности к этим эпитопам.
Полноразмерная таблица
Опухолеассоциированные антигены в тимусе
Центральная толерантность — палка о двух концах. Хотя он защищает нас от аутоиммунных заболеваний, он также может препятствовать противоопухолевому иммунитету. Вполне вероятно, что презентация собственных белков, которые делают репертуар Т-клеток толерантными к собственным антигенам, чтобы избежать аутоиммунитета, также может негативно влиять на способность Т-клеток опосредовать опухолевый иммунитет. Этот прогноз был подтвержден исследованиями, демонстрирующими экспрессию опухолеассоциированных антигенов, включая антигены дифференцировки и антигены зародышевой линии рака, в mTEC человека 38 , и, кроме того, экспрессия человеческого карциноэмбрионального антигена (CEA) в mTEC трансгенных мышей индуцирует центральную толерантность 39 . Тщательный литературный поиск природных репертуаров лигандов MHC в опухолях показал, что значительное количество MHC-связанных пептидов, обнаруженных в наших экспериментах, также (избыточно) представлено в почечно-клеточных карциномах 40,41 или других тканях заболевания (дополнительное Данные 2), включая ядерный онкоген SET и APOL1. Наши результаты дают объяснение, почему на периферии присутствуют только Т-клетки с низкой авидностью против многих антигенов, ассоциированных с раком 42 , так как презентация антигенов, ассоциированных с опухолью, путем отрицательного отбора клеток тимуса должна приводить к делеции Т-клеток с высокой авидностью.
Этот прогноз был подтвержден исследованиями, демонстрирующими экспрессию опухолеассоциированных антигенов, включая антигены дифференцировки и антигены зародышевой линии рака, в mTEC человека 38 , и, кроме того, экспрессия человеческого карциноэмбрионального антигена (CEA) в mTEC трансгенных мышей индуцирует центральную толерантность 39 . Тщательный литературный поиск природных репертуаров лигандов MHC в опухолях показал, что значительное количество MHC-связанных пептидов, обнаруженных в наших экспериментах, также (избыточно) представлено в почечно-клеточных карциномах 40,41 или других тканях заболевания (дополнительное Данные 2), включая ядерный онкоген SET и APOL1. Наши результаты дают объяснение, почему на периферии присутствуют только Т-клетки с низкой авидностью против многих антигенов, ассоциированных с раком 42 , так как презентация антигенов, ассоциированных с опухолью, путем отрицательного отбора клеток тимуса должна приводить к делеции Т-клеток с высокой авидностью. Следовательно, даже если обнаружится, что они избыточно экспрессируются в опухолевых тканях, они не могут быть первым выбором в качестве мишени для иммунотерапии. Примером может служить p53, супрессор опухоли, играющий центральную роль в развитии опухоли. Здесь мы обнаружили пептид, производный от p53, представленный на MHC I на DC (дополнительные данные 1 и 2), что должно привести к делеции тимуса p53-специфического CD8 9.0011 + Т-клеток. Это согласуется с недавним исследованием рака яичников, описывающим отсутствие спонтанного p53-специфического CD8 + , но не CD4 + -специфического Т-клеточного ответа у здоровых доноров и пациентов с высокими титрами антител против p53 (ссылка 43). ).
Следовательно, даже если обнаружится, что они избыточно экспрессируются в опухолевых тканях, они не могут быть первым выбором в качестве мишени для иммунотерапии. Примером может служить p53, супрессор опухоли, играющий центральную роль в развитии опухоли. Здесь мы обнаружили пептид, производный от p53, представленный на MHC I на DC (дополнительные данные 1 и 2), что должно привести к делеции тимуса p53-специфического CD8 9.0011 + Т-клеток. Это согласуется с недавним исследованием рака яичников, описывающим отсутствие спонтанного p53-специфического CD8 + , но не CD4 + -специфического Т-клеточного ответа у здоровых доноров и пациентов с высокими титрами антител против p53 (ссылка 43). ).
Обсуждение
Таким образом, мы приводим здесь первую характеристику пептидного репертуара, представленного молекулами MHC в тимусе здорового человека in vivo , с акцентом на те эпитопы, представленные mDCs, которые важны для отрицательного отбора. Интересно, что были обнаружены как известные аутоантигены, так и антигены, ассоциированные с опухолью. Мы наблюдали высокую степень межиндивидуальной изменчивости между донорами, которая частично может быть объяснена различиями в типе HLA, но также может быть частично связана с различиями в экспрессии белка. Более того, mDCs и non-mDC APC представляют разные, лишь частично перекрывающиеся наборы пептидов, вероятно, из-за различий как в экспрессии собственных белков, так и в их механизмах процессинга антигенов 13,15 .
Интересно, что были обнаружены как известные аутоантигены, так и антигены, ассоциированные с опухолью. Мы наблюдали высокую степень межиндивидуальной изменчивости между донорами, которая частично может быть объяснена различиями в типе HLA, но также может быть частично связана с различиями в экспрессии белка. Более того, mDCs и non-mDC APC представляют разные, лишь частично перекрывающиеся наборы пептидов, вероятно, из-за различий как в экспрессии собственных белков, так и в их механизмах процессинга антигенов 13,15 .
Хотя наши данные помогают ответить на давний вопрос о том, какие пептиды физиологически участвуют в отборе Т-клеток в тимусе человека, многие вопросы еще предстоит решить, прежде чем мы полностью поймем процесс центральной толерантности. Например, анализ лигандов, полученных из изолированных mTEC или cTEC, а также лигандов с низким содержанием, представленных DC, в настоящее время ограничен доступной технологией. По этой причине представленные здесь данные относятся только к mDC и не включают лиганды из изолированных mTEC, которые также способствуют центральной толерантности. Другой вопрос, который остается нерешенным, заключается в том, значительно ли изменяется лигандом тимуса в течение жизни человека. По очевидным причинам будет трудно ответить на этот вопрос экспериментально на людях, но некоторые предыдущие исследования предполагают, что вариации, вероятно, будут иметь место. ДК и мТЭК тимуса имеют период полураспада около 2–3 недель, и, по крайней мере, для мТЭК описаны значительные различия в экспрессии тканевого антигена между разными клетками 44 . Наши и другие данные подтверждают идею о том, что ДК тимуса пробуют кровоток на белки, а периферические ДК могут поглощать антигены, мигрировать в тимус и удалять соответствующие Т-клетки 8,12 . Следовательно, можно ожидать, что изменения на периферии повлияют на пептиды, представленные в тимусе.
Другой вопрос, который остается нерешенным, заключается в том, значительно ли изменяется лигандом тимуса в течение жизни человека. По очевидным причинам будет трудно ответить на этот вопрос экспериментально на людях, но некоторые предыдущие исследования предполагают, что вариации, вероятно, будут иметь место. ДК и мТЭК тимуса имеют период полураспада около 2–3 недель, и, по крайней мере, для мТЭК описаны значительные различия в экспрессии тканевого антигена между разными клетками 44 . Наши и другие данные подтверждают идею о том, что ДК тимуса пробуют кровоток на белки, а периферические ДК могут поглощать антигены, мигрировать в тимус и удалять соответствующие Т-клетки 8,12 . Следовательно, можно ожидать, что изменения на периферии повлияют на пептиды, представленные в тимусе.
Наши данные должны проложить путь к всестороннему пониманию центральной толерантности и однажды могут помочь улучшить дизайн терапевтических вмешательств, таких как противораковые вакцины. Экспрессия лигандов, полученных из антигенов, ассоциированных с опухолью, на APC тимуса подчеркивает необходимость более систематического анализа потенциальных кандидатов на противораковые вакцины с использованием наших данных для предварительного отбора или исключения антигенов из растущего списка кандидатов в будущих испытаниях вакцин.
Экспрессия лигандов, полученных из антигенов, ассоциированных с опухолью, на APC тимуса подчеркивает необходимость более систематического анализа потенциальных кандидатов на противораковые вакцины с использованием наших данных для предварительного отбора или исключения антигенов из растущего списка кандидатов в будущих испытаниях вакцин.
Методы
Образцы тимуса
Нормальные образцы тимуса человека были получены от детей, перенесших корректирующую операцию на сердце (кафедра кардиохирургии, Тюбингенский университет, Германия) после информированного согласия родителей и одобрения Институциональным наблюдательным советом Медицинского факультета Университет Тюбингена. Для получения подробной информации о донорах см. Таблицу 3.
Таблица 3 Возраст, пол и HLA-типирование доноров ткани тимуса, включенных в данное исследование.Полноразмерный стол
Генотипирование тканей тимуса
ДНК для генотипирования HLA выделяли из ткани тимуса с использованием набора QIAamp DNA Blood Mini (Qiagen). HLA-типирование с низким разрешением было выполнено Институтом клинической и экспериментальной трансфузионной медицины (IKET) Университета Тюбингена, Германия.
HLA-типирование с низким разрешением было выполнено Институтом клинической и экспериментальной трансфузионной медицины (IKET) Университета Тюбингена, Германия.
Выделение ДК тимуса
мДК выделены из ткани тимуса после механического и ферментативного разрушения (коллагеназа (1 мг мл –1 ) и ДНКаза (50 мкг мл –1 , оба от Roche), при 37 °C, два-три раза по 30–45 мин) 14 . АПК обогащали из полученной суспензии одиночных клеток с помощью центрифугирования в градиенте Перколла (плотность: 1,07 г мл -1 , 30 мин, 3500 г, 4°С), а ДК выделяли с помощью магнитных шариков (CD11c-PE и анти-PE шарики, Miltenyi Biotec) из фракции низкой плотности согласно инструкции производителя. Были собраны как положительные (DC), так и отрицательные (DC-истощенные) фракции (состав см. в дополнительной таблице S1 и дополнительном рисунке S1). Краткое описание рабочего процесса представлено на рис. 1.
Анализ FACS
Суспензии одиночных клеток из цельного тимуса, обогащенная АПК фракция после перколла, ДК и обедненная ДК фракции окрашивались антителами против HLA–DR (PerCP, 307628, 1:50), CD19 (FITC, 302205, 1:20), CD14 (APC, 301807, 1:50), CD45RA (FITC, 304105, 1:10), CD123 (PE/Cy7, 306009, 1:10), (все от Biolegend), CD45 ( PB, PB986, 1:10, DAKO), BDCA-4 (PE, 130-090-533, 1:10), CD11c (PE, 130-092-411, 1:10), EpCAM (APC, 130-091 -254, 1:10, от Miltenyi Biotec), CDR2 (1:50, Bruno Kyewski, DKFZ, Heidelberg; помечен Alexa488 с использованием набора для мечения белков Alexa Fluor 488 от Molecular Probes в соответствии с инструкциями производителя). mDC были закрыты как CD11c + , В-клетки строяли как CD19 + , PDCS AS CD45RA + BDCA-4 + , моноциты AS CD14 + , MTEC и CTEC AS CD45 LO-NEG EPCAM HI HI HI HI HI HI или CTEC AS CD45 LO-NEG HI или CTEC AS CD45 LO-NEG HI или CTEC AS CD45 LO-NEG HI или CTEC AS CD45 LO-NEG . ло-отрицательный EpCAM ло CDR2 + соответственно.
mDC были закрыты как CD11c + , В-клетки строяли как CD19 + , PDCS AS CD45RA + BDCA-4 + , моноциты AS CD14 + , MTEC и CTEC AS CD45 LO-NEG EPCAM HI HI HI HI HI HI или CTEC AS CD45 LO-NEG HI или CTEC AS CD45 LO-NEG HI или CTEC AS CD45 LO-NEG HI или CTEC AS CD45 LO-NEG . ло-отрицательный EpCAM ло CDR2 + соответственно.
Элюирование лигандов MHC
Пептиды, связанные с MHC, выделяли с использованием антител L234 (HLA–DR) и W6/32 (pan-HLA класса I) 45,46 . Мы также опубликовали подробный пошаговый протокол этой процедуры 45 . Осадок замороженных клеток или кусочки ткани лизировали 0,6% CHAPS с ингибиторами протеазы в PBS, перемешивая в течение 2 ч при 4°С. Полученный лизат обрабатывали ультразвуком, а дебрис осаждали центрифугированием (4000 g в течение 20 мин с последующим ультрацентрифугированием при 40 000 об/мин в течение 1 ч). Супернатант очищали ультрафильтрацией (размер пор 0,2 мкм, Sartolab P plus, Sartorius) и комплексы пептид-ГКГС выделяли из супернатанта аффинной хроматографией (антитела L234 (ГКГС II) или W6/32 (ГКГС I), связанные с CNBr- Сефароза, ночь, 4 °C). Связанные комплексы МНС-пептид элюировали 0,1% ТФУ. Элюированные пептиды отделяли от белков MHC центрифугированием через фильтр с порогом молекулярной массы 10 кДа и анализировали с помощью ЖХ-МС/МС.
Супернатант очищали ультрафильтрацией (размер пор 0,2 мкм, Sartolab P plus, Sartorius) и комплексы пептид-ГКГС выделяли из супернатанта аффинной хроматографией (антитела L234 (ГКГС II) или W6/32 (ГКГС I), связанные с CNBr- Сефароза, ночь, 4 °C). Связанные комплексы МНС-пептид элюировали 0,1% ТФУ. Элюированные пептиды отделяли от белков MHC центрифугированием через фильтр с порогом молекулярной массы 10 кДа и анализировали с помощью ЖХ-МС/МС.
Идентификация лигандов MHC
Наномасштабную жидкостную хроматографию лигандов MHC проводили с помощью системы УЭЖХ Waters NanoAcquity, оснащенной обращенно-фазовой колонкой BEH C18 75 мкм × 150 мм и петлей для образцов PEEKSIL объемом 2,6 мкл (SGE, Дармштадт, Германия). ). Водная подвижная фаза (подвижная фаза А) представляла собой Н 2 О с 0,1% муравьиной кислоты. Органическая подвижная фаза (подвижная фаза В) представляла собой 0,1% муравьиную кислоту в ацетонитриле. Образцы (ввод 2,6 мкл) наносили на колонку в режиме прямого ввода с 3% подвижной фазы Б в течение 15 мин при 400 нл мин -1 , с последующей дополнительной 10-минутной промывкой (3% В) в течение 10 минут при 300 нл мин -1 . Пептиды элюировали из колонки с градиентом от 3 до 35 % подвижной фазы В в течение 120 мин при 300 нл мин -1 с последующей 20-минутной промывкой 80 % подвижной фазы В. Колонку немедленно повторно уравновешивали при начальные условия (3% подвижной фазы B) в течение 20 мин. [Glu 1 ] фибринопептид использовали в качестве лок-массы в количестве 500 фмоль мкл -1 . Раствор Lockmass подавали вспомогательным насосом системы NanoAcquity со скоростью 400 нл мин −1 на эталонный распылитель источника NanoLockSpray. МС/МС-анализ пептидов выполняли с использованием масс-спектрометра Waters Q-Tof Premier, работающего в V-режиме с типичной разрешающей способностью R =10,000 в режиме сбора данных в зависимости от данных, выбирая три наиболее интенсивных иона-предшественника для фракционирования в ячейка столкновения. Фрагментация исходных ионов осуществлялась при столкновении с атомами аргона. Энергия столкновения варьировалась от 15 до 35 эВ в зависимости от массы и заряда иона-предшественника.
Пептиды элюировали из колонки с градиентом от 3 до 35 % подвижной фазы В в течение 120 мин при 300 нл мин -1 с последующей 20-минутной промывкой 80 % подвижной фазы В. Колонку немедленно повторно уравновешивали при начальные условия (3% подвижной фазы B) в течение 20 мин. [Glu 1 ] фибринопептид использовали в качестве лок-массы в количестве 500 фмоль мкл -1 . Раствор Lockmass подавали вспомогательным насосом системы NanoAcquity со скоростью 400 нл мин −1 на эталонный распылитель источника NanoLockSpray. МС/МС-анализ пептидов выполняли с использованием масс-спектрометра Waters Q-Tof Premier, работающего в V-режиме с типичной разрешающей способностью R =10,000 в режиме сбора данных в зависимости от данных, выбирая три наиболее интенсивных иона-предшественника для фракционирования в ячейка столкновения. Фрагментация исходных ионов осуществлялась при столкновении с атомами аргона. Энергия столкновения варьировалась от 15 до 35 эВ в зависимости от массы и заряда иона-предшественника. Время интегрирования для TOF-анализатора составляло 1 с с задержкой между сканированиями 0,1 с.
Время интегрирования для TOF-анализатора составляло 1 с с задержкой между сканированиями 0,1 с.
Интерпретацию спектров фрагментации МС/МС проводили вручную с использованием программного обеспечения MassLynx 4.0 47 . Для ручной идентификации пептидов поиск меток последовательности был выполнен с использованием программного обеспечения Mascot 2.0 (допуск пептидов и МС/МС, 0,2 Da; неизбыточная база данных Национального центра биотехнологической информации (NCBInr), обновляемая ежемесячно и ограниченная для поиска записями человека), и соответствующие попадания оценивались вручную, а не по оценке талисмана. Критериями ручной идентификации были: разумная интерпретация не менее 95% пиков всех фрагментов, полное покрытие последовательности фрагментами МС/МС и интенсивность сигналов пиков ионов фрагментов, которые соответствуют вероятности разрыва соответствующей последовательности.
Полученные пептидные последовательности были отнесены к исходным белкам с использованием алгоритма BLAST и проведен дальнейший биоинформатический анализ 48,49 . Лиганды MHC класса I и MHC класса II в различных препаратах клеток тимуса были отнесены к субклеточным компартментам или функциональным категориям в соответствии с аннотациями Gene Ontology (www.geneontology.org). В тех случаях, когда этот поиск не давал информации или был неполным, аннотация выполнялась вручную на основе информации из WoLF PSORT 9.0011 48 (wolfpsort.org), SubLoc 49 (www.bioinfo.tsinghua.edu.cn/SubLoc/), SwissProt и опубликованная литература. Затем исходные белки были сгруппированы по классическому пути MHC I (цитозольный, ядерный) или пути MHC II (внеклеточный, везикулярный и клеточная мембрана), митохондриальному или ER-компартменту. Чтобы облегчить интерпретацию данных о биологической функции, связанные подкатегории были сгруппированы вместе, например, «передача сигналов», «передача сигналов между клетками» и «передача сигнала» были сгруппированы вместе как «передача сигналов». Все пептиды были отнесены к определенному аллотипу HLA с учетом типа HLA пациента, а также известных мотивов связывания различных аллотипов 50 .
Лиганды MHC класса I и MHC класса II в различных препаратах клеток тимуса были отнесены к субклеточным компартментам или функциональным категориям в соответствии с аннотациями Gene Ontology (www.geneontology.org). В тех случаях, когда этот поиск не давал информации или был неполным, аннотация выполнялась вручную на основе информации из WoLF PSORT 9.0011 48 (wolfpsort.org), SubLoc 49 (www.bioinfo.tsinghua.edu.cn/SubLoc/), SwissProt и опубликованная литература. Затем исходные белки были сгруппированы по классическому пути MHC I (цитозольный, ядерный) или пути MHC II (внеклеточный, везикулярный и клеточная мембрана), митохондриальному или ER-компартменту. Чтобы облегчить интерпретацию данных о биологической функции, связанные подкатегории были сгруппированы вместе, например, «передача сигналов», «передача сигналов между клетками» и «передача сигнала» были сгруппированы вместе как «передача сигналов». Все пептиды были отнесены к определенному аллотипу HLA с учетом типа HLA пациента, а также известных мотивов связывания различных аллотипов 50 . Пептиды были отнесены к представленной молекуле HLA путем сравнения их якорных аминокислот с известными пептидными мотивами, специфичными для аллеля HLA. Для того чтобы выполнить автоматическое воспроизводимое отнесение к правильному аллотипу HLA, все пептидные последовательности подвергали предсказанию эпитопов с использованием базы данных SYFPEITHI (ссылка 50) (www.syfpeithi.de), оценивая предпочтительное связывание с одним из HLA. аллотипы, определяемые HLA-типированием. SYFPEITHI рассчитывает баллы в соответствии с предпочтительными аминокислотами в каждой позиции последовательности. Пептиды, получившие более половины максимальной оценки для данной молекулы HLA, были отнесены к соответствующей молекуле. Следует отметить, что лиганды разных молекул HLA могут достигать различных максимальных показателей. Максимальные баллы: 44 для HLA-A*03, 44 для HLA-A*26, 29.для HLA-A*31, 37 для HLA-A*32, 33 для HLA-B*07, 30 для HLA-B*38 и 38 для HLA-B*51. Если веб-версия SYFPEITHI не позволяла прогнозировать эпитопы, использовалась собственная версия с расширенными возможностями.
Пептиды были отнесены к представленной молекуле HLA путем сравнения их якорных аминокислот с известными пептидными мотивами, специфичными для аллеля HLA. Для того чтобы выполнить автоматическое воспроизводимое отнесение к правильному аллотипу HLA, все пептидные последовательности подвергали предсказанию эпитопов с использованием базы данных SYFPEITHI (ссылка 50) (www.syfpeithi.de), оценивая предпочтительное связывание с одним из HLA. аллотипы, определяемые HLA-типированием. SYFPEITHI рассчитывает баллы в соответствии с предпочтительными аминокислотами в каждой позиции последовательности. Пептиды, получившие более половины максимальной оценки для данной молекулы HLA, были отнесены к соответствующей молекуле. Следует отметить, что лиганды разных молекул HLA могут достигать различных максимальных показателей. Максимальные баллы: 44 для HLA-A*03, 44 для HLA-A*26, 29.для HLA-A*31, 37 для HLA-A*32, 33 для HLA-B*07, 30 для HLA-B*38 и 38 для HLA-B*51. Если веб-версия SYFPEITHI не позволяла прогнозировать эпитопы, использовалась собственная версия с расширенными возможностями. Это имело место для предсказаний лигандов HLA-A*31 и HLA-A*32 и для редко встречающихся длин пептидов (8 мер, 11 мер) других аллотипов HLA.
Это имело место для предсказаний лигандов HLA-A*31 и HLA-A*32 и для редко встречающихся длин пептидов (8 мер, 11 мер) других аллотипов HLA.
Анализ воспроизводимости
Для валидации метода и анализа воспроизводимости были выделены пептиды MHC I из РВМС здоровых добровольцев и выполнены технические повторы. Для статистического анализа внутривыборочной (т.е. технической) воспроизводимости были рассчитаны перекрытия каждой мыслимой пары технических повторностей для одной случайно выбранной выборки. Для оценки межиндивидуальной изменчивости мы проанализировали перекрытия исходных протеомов лиганда HLA пяти образцов РВМС здоровых добровольцев. Мы провели комплексный анализ перекрытия всех пяти образцов. Для статистического анализа межиндивидуальной изменчивости был проведен парный анализ перекрытия всех 10 возможных комбинаций исходных протеомов. Две популяции (технические повторяющиеся перекрытия по сравнению с индивидуальными перекрытиями) сравнивались с использованием двусторонней непарной шкалы Стьюдента. 0005 t — тест, не предполагающий равных стандартных отклонений (дополнительный рисунок S3).
0005 t — тест, не предполагающий равных стандартных отклонений (дополнительный рисунок S3).
Дополнительная информация
Как цитировать эту статью: Adamopoulou, E. et al . Изучение матрикса MHC-пептида центральной толерантности в тимусе человека. Нац. коммун. 4:2039 doi: 10.1038/ncomms3039 (2013).
Ссылки
-
Kyewski, B. & Klein, L. Центральная роль центральной толерантности. год. Преподобный Иммунол. 24 , 571–606 (2006).
КАС Статья пабмед Google ученый
-
Кляйн Л., Хинтербергер М., Вирнсбергер Г. и Кьювски Б. Презентация антигена в тимусе для положительного отбора и индукции центральной толерантности. Нац. Преподобный Иммунол. 9 , 833–844 (2009).
КАС Статья пабмед Google ученый
-
Liu, G.
 Y. et al. Распознавание собственного антигена Т-клетками с низкой авидностью позволяет избежать центральной толерантности. Иммунитет 3 , 407–415 (1995).
Y. et al. Распознавание собственного антигена Т-клетками с низкой авидностью позволяет избежать центральной толерантности. Иммунитет 3 , 407–415 (1995). КАС Статья пабмед Google ученый
-
Mazda, O., Watanabe, Y., Gyotoku, J. & Katsura, Y. Потребность в дендритных клетках и В-клетках при клональной делеции Mls-реактивных Т-клеток в тимусе. Дж. Экспл. Мед. 173 , 539–547 (1991).
КАС Статья пабмед Google ученый
-
Brocker, T., Riedinger, M. & Karjalainen, K. Целенаправленная экспрессия молекул главного комплекса гистосовместимости (MHC) класса II демонстрирует, что дендритные клетки могут индуцировать отрицательную, но не положительную селекцию тимоцитов in vivo . Дж. Экспл. Мед. 185 , 541–550 (1997).
КАС Статья пабмед ПабМед Центральный Google ученый
-
Hubert, F.
 X. et al. Aire регулирует перенос антигена от mTEC к дендритным клеткам для индукции толерантности тимуса. Кровь 118 , 2462–2472 (2011).
X. et al. Aire регулирует перенос антигена от mTEC к дендритным клеткам для индукции толерантности тимуса. Кровь 118 , 2462–2472 (2011). КАС Статья пабмед Google ученый
-
Гальегос, А. М. и Беван, М. Дж. Центральная толерантность к тканеспецифическим антигенам, опосредованная прямой и непрямой презентацией антигена. Дж. Экспл. Мед. 200 , 1039–1049 (2004).
КАС Статья пабмед ПабМед Центральный Google ученый
-
Хадейба, Х. и др. Плазмацитоидные дендритные клетки транспортируют периферические антигены в тимус, способствуя центральной толерантности. Иммунитет 36 , 438–450 (2012).
КАС Статья пабмед ПабМед Центральный Google ученый
-
Кляйндинст П., Кретьен И.
 , Винклер Т. и Брокер Т. Функциональное сравнение В-клеток тимуса и дендритных клеток in vivo . Кровь 95 , 2610–2616 (2000).
, Винклер Т. и Брокер Т. Функциональное сравнение В-клеток тимуса и дендритных клеток in vivo . Кровь 95 , 2610–2616 (2000). КАС пабмед Google ученый
-
Abramson, J., Giraud, M., Benoist, C. & Mathis, D. Партнеры Aire в молекулярном контроле иммунологической толерантности. Cell 140 , 123–135 (2010).
КАС Статья пабмед Google ученый
-
Derbinski, J., Schulte, A., Kyewski, B. & Klein, L. Беспорядочная экспрессия генов в медуллярных эпителиальных клетках тимуса отражает периферическую самость. Нац. Иммунол. 2 , 1032–1039 (2001).
КАС Статья пабмед Google ученый
-
Bonasio, R. et al. Клональная делеция тимоцитов циркулирующими дендритными клетками, направляющимися в тимус.
 Нац. Иммунол. 7 , 1092–1100 (2006).
Нац. Иммунол. 7 , 1092–1100 (2006). КАС Статья пабмед Google ученый
-
Мурата, С. и др. Регуляция развития CD8+ Т-клеток специфическими для тимуса протеасомами. Наука 316 , 1349–1353 (2007).
ОБЪЯВЛЕНИЕ КАС Статья пабмед Google ученый
-
Nakagawa, T. et al. Катепсин L: решающая роль в деградации Ii и отборе Т-клеток CD4 в тимусе. Наука 280 , 450–453 (1998).
ОБЪЯВЛЕНИЕ КАС Статья пабмед Google ученый
-
Stoeckle, C. et al. Катепсин S доминирует в процессинге аутоантигена в дендритных клетках тимуса человека. J. Аутоиммун. 38 , 332–343 (2012).
КАС Статья пабмед Google ученый
-
Толоса, Э.
 и др. Катепсин V участвует в деградации инвариантной цепи в тимусе человека и сверхэкспрессируется при миастении. Дж. Клин. Инвестировать. 112 , 517–526 (2003).
и др. Катепсин V участвует в деградации инвариантной цепи в тимусе человека и сверхэкспрессируется при миастении. Дж. Клин. Инвестировать. 112 , 517–526 (2003). КАС Статья пабмед ПабМед Центральный Google ученый
-
Маррак П., Игнатович Л., Капплер Дж. В., Боймел Дж. и Фрид Дж. Х. Сравнение пептидов, связанных с селезенкой и тимусом класса II. Дж. Экспл. Мед. 178 , 2173–2183 (1993).
КАС Статья пабмед Google ученый
-
Сантори, Ф. Р. и др. Редкие структурно гомологичные аутопептиды способствуют положительному отбору тимоцитов. Иммунитет 17 , 131–142 (2002).
КАС Статья пабмед Google ученый
-
Hogquist, K.A. et al. Идентификация встречающегося в природе лиганда для положительного отбора тимуса.
 Иммунитет 6 , 389–399 (1997).
Иммунитет 6 , 389–399 (1997). КАС Статья пабмед Google ученый
-
Fortier, M.H. et al. Репертуар пептидов MHC класса I формируется транскриптомом. Дж. Экспл. Мед. 205 , 595–610 (2008).
КАС Статья пабмед ПабМед Центральный Google ученый
-
Garcia, C.A. et al. Дендритные клетки в тимусе и периферии человека демонстрируют проинсулиновый эпитоп зависимым от транскрипции и независимым от захвата способом. Дж. Иммунол. 175 , 2111–2122 (2005 г.).
КАС Статья пабмед Google ученый
-
Villadangos, J. A. & Schnorrer, P Внутренние и кооперативные антигенпрезентирующие функции субпопуляций дендритных клеток in vivo . Нац. Преподобный Иммунол. 7 , 543–555 (2007).

КАС Статья пабмед Google ученый
-
Schmid, D., Pypaert, M. & Munz, C. Компартменты, нагруженные антигеном, для молекул главного комплекса гистосовместимости класса II постоянно получают входные данные от аутофагосом. Иммунитет 26 , 79–92 (2007).
КАС Статья пабмед Google ученый
-
Dengjel, J. et al. Аутофагия способствует презентации пептидов MHC класса II из внутриклеточных исходных белков. Проц. Натл акад. науч. США 102 , 7922–7927 (2005 г.).
ОБЪЯВЛЕНИЕ КАС Статья пабмед ПабМед Центральный Google ученый
-
Koble, C. & Kyewski, B. Продолговатый мозг тимуса: уникальная микросреда для межклеточного переноса собственных антигенов. Дж. Экспл. Мед. 206 , 1505–1513 (2009).

КАС Статья пабмед ПабМед Центральный Google ученый
-
Кляйн, Л., Хинтербергер, М., фон Роршайдт, Дж. и Айхингер, М. Автономный и зависящий от дендритных клеток вклад медуллярных эпителиальных клеток тимуса в центральную толерантность. Тенденции Иммунол. 32 , 188–193 (2011).
КАС Статья пабмед Google ученый
-
Nedjic, J., Aichinger, M., Emmerich, J., Mizushima, N. & Klein, L. Аутофагия в эпителии тимуса формирует репертуар Т-клеток и необходима для толерантности. Природа 455 , 396–400 (2008).
ОБЪЯВЛЕНИЕ КАС Статья пабмед Google ученый
-
Gray, D., Abramson, J., Benoist, C. & Mathis, D. Задержка пролиферации и быстрое обновление эпителиальных клеток тимуса, экспрессирующих Aire.
 Дж. Экспл. Мед. 204 , 2521–2528 (2007 г.).
Дж. Экспл. Мед. 204 , 2521–2528 (2007 г.). КАС Статья пабмед ПабМед Центральный Google ученый
-
Thery, C. et al. Непрямая активация наивных CD4+ Т-клеток экзосомами, происходящими из дендритных клеток. Нац. Иммунол. 3 , 1156–1162 (2002).
КАС Статья пабмед Google ученый
-
Millet, V., Naquet, P. & Guinamar, R.R. Межклеточный перенос MHC между эпителиальными и дендритными клетками тимуса. евро. Дж. Иммунол. 38 , 1257–1263 (2008).
КАС Статья пабмед Google ученый
-
Мунц, К. Повышение иммунитета посредством аутофагии. год. Преподобный Иммунол. 27 , 423–449 (2009).
КАС Статья пабмед Google ученый
-
Донгре, А.
 Р. и др. In vivo Презентация цитозольных белков MHC класса II, выявленная с помощью быстрой автоматизированной тандемной масс-спектрометрии и функционального анализа. евро. Дж. Иммунол. 31 , 1485–1494 (2001).
Р. и др. In vivo Презентация цитозольных белков MHC класса II, выявленная с помощью быстрой автоматизированной тандемной масс-спектрометрии и функционального анализа. евро. Дж. Иммунол. 31 , 1485–1494 (2001). КАС Статья пабмед Google ученый
-
Хогквист, К. А., Болдуин, Т. А. и Джеймсон, С. К. Центральная толерантность: обучение самоконтролю в тимусе. Нац. Преподобный Иммунол. 5 , 772–782 (2005).
КАС Статья пабмед Google ученый
-
Feitsma, A.L. et al. Идентификация цитруллинированных пептидов виментина в качестве эпитопов Т-клеток у HLA-DR4-положительных пациентов с ревматоидным артритом. Ревматоидный артрит. 62 , 117–125 (2010).
КАС Статья пабмед Google ученый
-
Уолстром, Дж.
 и др. Идентификация HLA-DR-связанных пептидов, представленных клетками бронхоальвеолярного лаважа человека при саркоидозе. Дж. Клин. Инвестировать. 117 , 3576–3582 (2007).
и др. Идентификация HLA-DR-связанных пептидов, представленных клетками бронхоальвеолярного лаважа человека при саркоидозе. Дж. Клин. Инвестировать. 117 , 3576–3582 (2007). Артикул пабмед ПабМед Центральный Google ученый
-
Forooghian, F., Cheung, R.K., Smith, W.C., O’Connor, P. & Dosch, H.M. Энолаза и аррестин являются новыми немиелиновыми аутоантигенами при рассеянном склерозе. Дж. Клин. Иммунол. 27 , 388–396 (2007).
КАС Статья пабмед ПабМед Центральный Google ученый
-
Абулафия-Лапид Р., Гиллис Д., Йосеф О., Атлан Х. и Коэн И. Р. Т-клетки и аутоантитела к HSP70 человека при диабете 1 типа у детей. J. Аутоиммун. 20 , 313–321 (2003).
КАС Статья пабмед Google ученый
-
Gotter, J.
 , Brors, B., Hergenhahn, M. & Kyewski, B. Медуллярные эпителиальные клетки тимуса человека экспрессируют очень разнообразный набор тканеспецифических генов, совместно локализованных в хромосомных кластерах. Дж. Экспл. Мед. 199 , 155–166 (2004).
, Brors, B., Hergenhahn, M. & Kyewski, B. Медуллярные эпителиальные клетки тимуса человека экспрессируют очень разнообразный набор тканеспецифических генов, совместно локализованных в хромосомных кластерах. Дж. Экспл. Мед. 199 , 155–166 (2004). КАС Статья пабмед ПабМед Центральный Google ученый
-
Бос, Р. и др. Экспрессия естественного опухолевого антигена эпителиальными клетками тимуса нарушает репертуар опухолезащитных CD4+ T-клеток. Рак рез. 65 , 6443–6449 (2005).
КАС Статья пабмед Google ученый
-
Kruger, T. et al. Уроки, которые следует извлечь из первичных почечно-клеточных карцином: новые опухолевые антигены и лиганды HLA для иммунотерапии. Рак Иммунол. Иммунотер. 54 , 826–836 (2005).
Артикул пабмед Google ученый
-
Stickel, J.
 S. et al. Профили лиганда HLA первичной почечно-клеточной карциномы сохраняются в метастазах. Рак Иммунол. Иммунотер. 58 , 1407–1417 (2009).
S. et al. Профили лиганда HLA первичной почечно-клеточной карциномы сохраняются в метастазах. Рак Иммунол. Иммунотер. 58 , 1407–1417 (2009). КАС Статья пабмед Google ученый
-
Эрнандес Дж., Ли П.П., Дэвис М.М. и Шерман Л.А. Использование тетрамеров пептидов HLA A2.1/p53 для визуализации влияния самотолерантности на репертуар TCR. Дж. Иммунол. 164 , 596–602 (2000).
КАС Статья пабмед Google ученый
-
Tsuji, T. et al. Расщепленная толерантность Т-клеток к собственному/опухолевому антигену: спонтанные ответы CD4+, но не CD8+ Т-клеток против p53 у онкологических больных и здоровых доноров. PLoS One 6 , e23651 (2011 г.).
ОБЪЯВЛЕНИЕ КАС Статья пабмед ПабМед Центральный Google ученый
-
Taubert, R.
 , Schwendemann, J. & Kyewski, B. Сильно вариабельная экспрессия аутоантигенов, ограниченных тканями, в тимусе человека: последствия для самопереносимости и аутоиммунитета. евро. Дж. Иммунол. 37 , 838–848 (2007).
, Schwendemann, J. & Kyewski, B. Сильно вариабельная экспрессия аутоантигенов, ограниченных тканями, в тимусе человека: последствия для самопереносимости и аутоиммунитета. евро. Дж. Иммунол. 37 , 838–848 (2007). КАС Статья пабмед Google ученый
-
Ковалевски Д. Дж. и Стеванович С. Биохимическая крупномасштабная идентификация лигандов MHC класса I. Методы Мол. биол. 960 , 145–157 (2013).
КАС Статья пабмед Google ученый
-
Falk, K., Rotzschke, O., Stevanovic, S., Jung, G. & Rammensee, HG. Аллель-специфические мотивы, выявленные путем секвенирования собственных пептидов, элюированных из молекул MHC. Природа 351 , 290–296 (1991).
ОБЪЯВЛЕНИЕ КАС Статья пабмед Google ученый
-
Weinzierl, A.
 O. et al. Искаженное соотношение между числом копий мРНК и соответствующей плотностью лиганда главного комплекса гистосовместимости на поверхности клетки. мол. Клеточная протеомика 6 , 102–113 (2007).
O. et al. Искаженное соотношение между числом копий мРНК и соответствующей плотностью лиганда главного комплекса гистосовместимости на поверхности клетки. мол. Клеточная протеомика 6 , 102–113 (2007). КАС Статья пабмед Google ученый
-
Horton, P. et al. WoLF PSORT: предиктор локализации белка. Рез. нуклеиновых кислот. 35 , W585–W587 (2007 г.).
Артикул пабмед ПабМед Центральный Google ученый
-
Chen, H., Huang, N. & Sun, Z. SubLoc: набор серверов/клиентов для субклеточной локализации белков на основе SOAP. Биоинформатика 22 , 376–377 (2006).
Артикул пабмед Google ученый
-
Rammensee, H., Bachmann, J., Emmerich, N.P., Bachor, O.A. & Stevanovic, S. SYFPEITHI: база данных лигандов MHC и пептидных мотивов.
 Immunogenetics 50 , 213–219 (1999).
Immunogenetics 50 , 213–219 (1999). КАС Статья пабмед Google ученый
-
Уолстром, Дж. и др. Аутоиммунные Т-клеточные ответы на антигенные пептиды, представленные молекулами HLA-DR клеток бронхоальвеолярного лаважа при саркоидозе. клин. Иммунол. 133 , 353–363 (2009).
Артикул пабмед Google ученый
-
Johnston, A., Gudjonsson, J.E., Sigmundsdottir, H., Love, T.J. & Valdimarsson, H. Реакции Т-клеток периферической крови на кератиновые пептиды, которые имеют общие последовательности со стрептококковыми М-белками, в значительной степени ограничены CD8(+), проникающими в кожу. Т-клетки. клин. Эксп. Иммунол. 138 , 83–93 (2004).
КАС Статья пабмед ПабМед Центральный Google ученый
Скачать ссылки
Благодарности
Мы благодарим юных пациентов и их родителей за пожертвование образцов тимуса, профессора Герхарда Цимера, профессора Германа Эберта, доктора Фолькера Штегера и доктора Вильке Шнайдер (Тюбинген, Германия) за предоставление нам образцов, профессора Дороти Вернет (Тюбинген, Германия) за типирование HLA, Симону Пёшель за техническую помощь, Даниэля Ковалевски (Тюбинген, Германия) за методологическую поддержку и профессора Бруно Кьюски (Гейдельберг, Германия) за антитело к CDR2. Эта работа была поддержана Deutsche Forschungsgemeinschaft (SFB 685, Высшая школа 79).4, INST 371/23-1 FUGG), Hertie Foundation, BMBF (e:Bio Express2Present, 0316179C) и Исследовательского центра иммунологии Майнцского университета имени Иоганна Гутенберга.
Эта работа была поддержана Deutsche Forschungsgemeinschaft (SFB 685, Высшая школа 79).4, INST 371/23-1 FUGG), Hertie Foundation, BMBF (e:Bio Express2Present, 0316179C) и Исследовательского центра иммунологии Майнцского университета имени Иоганна Гутенберга.
Информация об авторе
Примечания автора
-
Arthur Melms
Текущий адрес: Текущий адрес: Кафедра неврологии, Университет Эрланген-Нюрнберг, Эрланген, Германия,
-
Christina Stoeckle
Текущий адрес: Текущий адрес: Институт фармакологии Бернского университета, Берн, Швейцария,
Авторы и филиалы
-
Отделение общей неврологии, Институт клинических исследований мозга им. Херти, 72076 Тюбинген, Германия,
Элени Адамопулу, Паула Клуг, Иоанна А. Рота, Сильвия Титц, Мадлен Гебхардт, Артур Мелмс и Кристина Стокл
-
Институт иммунологии, Университетский медицинский центр Майнцского университета им.
 Иоганна Гутенберга, 55131 Майнц, Германия,
Иоганна Гутенберга, 55131 Майнц, Германия,
Stefan Tenzer & Hansjörg Schild
-
Кафедра иммунологии, Институт клеточной биологии, Тюбингенский университет, 72076 Тюбинген, Германия,
Нина Хиллен и Стефан Стеванович
-
Институт иммунологии, Университетский медицинский центр Гамбург-Эппендорф, 20246 Гамбург, Германия,
Ева Толоса
Авторы
- Элени Адамопулу
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
- Stefan Tenzer
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Nina Hillen
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Paula Klug
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Академия
- Иоанна А.
 Рота
Рота Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Silvia Tietz
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Madlen Gebhardt
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Stefan Stevanovic
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
- Hansjörg Schild
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
- Eva Tolosa
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Артур Мелмс
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
- Christina Stoeckle
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
Contributions
C. S. разработал проект при поддержке E.T. и скоординированные исследования. К.С., Э.А. и NH спроектировали и провели эксперименты и анализ данных секвенирования de novo с P.K., S.T., I.R. и М.Г. оказание помощи. ЖХ-МС/МС и биоинформатический анализ был проведен С.Т., С.С. оказал ценную помощь в назначении пептида HLA и методологии, Е.Т., С.Т. и Х.С. способствовал научной дискуссии. ЯВЛЯЮСЬ. координировал сбор образцов тимуса и способствовал направлению исследования. Э.А. и CS оценили данные и написали рукопись, которая была обсуждена, критически отредактирована и одобрена всеми соавторами. К.С. и А.М. поделиться старшим авторством.
S. разработал проект при поддержке E.T. и скоординированные исследования. К.С., Э.А. и NH спроектировали и провели эксперименты и анализ данных секвенирования de novo с P.K., S.T., I.R. и М.Г. оказание помощи. ЖХ-МС/МС и биоинформатический анализ был проведен С.Т., С.С. оказал ценную помощь в назначении пептида HLA и методологии, Е.Т., С.Т. и Х.С. способствовал научной дискуссии. ЯВЛЯЮСЬ. координировал сбор образцов тимуса и способствовал направлению исследования. Э.А. и CS оценили данные и написали рукопись, которая была обсуждена, критически отредактирована и одобрена всеми соавторами. К.С. и А.М. поделиться старшим авторством.
Автор, ответственный за переписку
Кристина Стокл.
Заявление об этике
Конкурирующие интересы
Авторы не заявляют об отсутствии конкурирующих финансовых интересов.
Дополнительная информация
Дополнительные рисунки и таблица
Дополнительные рисунки S1-S3 и дополнительная таблица (PDF 450 kb)
Дополнительные данные 1 приготовления.
 (XLS 176 КБ)
(XLS 176 КБ) Дополнительные данные 2
Пептиды, полученные из тимуса, чрезмерно представлены в а) опухолевых и б) других патологических тканях. (XLS, 46 КБ)
Права и разрешения
Эта работа находится под лицензией Creative Commons Attribution-NonCommercial-NoDerivs 3.0 Unported License. Чтобы просмотреть копию этой лицензии, посетите http://creativecommons.org/licenses/by-nc-nd/3.0/
Перепечатки и разрешения
Об этой статье
Дополнительная литература
-
Широко распространенная и тканеспецифичная экспрессия эндогенных ретроэлементов в соматических тканях человека
- Жан-Давид Ларуш
- Ася Трофимова
- Клод Перро
Геномная медицина (2020)
-
Иммуноортогональные ортологи капсидов AAV и Cas9 обходят иммунный ответ на введение генной терапии
- Ана М.
 Морено
Морено - Натан Палмер
- Прашант Мали
Природа Биомедицинская инженерия (2019)
- Ана М.
-
Процессинг антигена MHC класса II при иммунной толерантности и воспалении
- Молли М. Юревич
- Лоуренс Дж. Стерн
Иммуногенетика (2019)
-
Белки теплового шока в терапии аутоиммунных заболеваний: слишком просто, чтобы быть правдой?
- Стефан Тукай
- Мацей Камински
Клеточный стресс и шапероны (2019)
-
Тканевая схема иммунопептидома мышиного MHC класса I.

- Хайко Шустер
- Венгуан Шао
- Этьен Карон
Научные данные (2018)
Комментарии
Отправляя комментарий, вы соглашаетесь соблюдать наши Условия и Правила сообщества. Если вы обнаружите что-то оскорбительное или не соответствующее нашим условиям или правилам, отметьте это как неприемлемое.
Анализ стека допусков: наихудший случай и RSS
Допуск Анализ стека — это инструмент проектирования, используемый для анализа и оптимизации конструкции изделия для сборки. Он используется для расчета кумулятивных эффектов допусков деталей в сборке. Таким образом, анализ набора допусков обеспечивает плавную сборку деталей на производственном участке. В этой статье мы обсудим наихудший случай и методы RSS для решения проблем с накоплением допусков.
Он используется для расчета кумулятивных эффектов допусков деталей в сборке. Таким образом, анализ набора допусков обеспечивает плавную сборку деталей на производственном участке. В этой статье мы обсудим наихудший случай и методы RSS для решения проблем с накоплением допусков.
Для лучшего понимания допусков сложите. Мы предлагаем вам сначала прочитать эту статью о том, что такое предел, посадка и допуск?
Содержание
- Зачем нужен анализ толерантности?
- Типы допусков Анализ стека
- Анализ допусков для наихудшего случая
- Статистический анализ допусков
- Чем отличаются результаты анализа допусков для наихудшего случая и RSS?
- Калькулятор сложения допусков
Зачем нужен анализ допусков?
Изготовление близко сопрягаемых деталей без допусков экономически нецелесообразно. Жесткие допуски могут привести к увеличению процента брака и производственных затрат. Принимая во внимание, что свободные допуски могут повлиять на функционирование продукта. Поэтому всегда рекомендуется определять оптимизированные допуски. Оптимизация допуска детали увеличивает диапазон допуска детали. Это также помогает снизить затраты на изготовление деталей.
Поэтому всегда рекомендуется определять оптимизированные допуски. Оптимизация допуска детали увеличивает диапазон допуска детали. Это также помогает снизить затраты на изготовление деталей.
Анализ набора допусков используется для расчета оптимизированных допусков деталей. Это можно сделать вручную или с помощью программного обеспечения для анализа стека.
Типы анализа наложения допусков
Различные методы анализа наложения допусков используются для расчета оптимизированных значений допусков. Каждый метод имеет свои преимущества, ограничения и области применения. Из этих наихудших случаев и методов статистического анализа допусков более популярны. Давайте обсудим эти методы подробно.
Анализ допусков для наихудшего случая
Метод наложения допусков для наихудшего случая использует простые арифметические операции (сложение и вычитание) для расчета оптимизированных допусков. В этом методе все размеры принимаются за крайние пределы. Для небольших объемов производства рекомендуется анализ допустимых отклонений для наихудшего случая.
Для небольших объемов производства рекомендуется анализ допустимых отклонений для наихудшего случая.
Преимущества анализа стопки в наихудшем случае
- Анализ стопки в наихудшем случае обеспечивает 100% сборку деталей на производственном участке.
- Нулевой коэффициент отказа.
Недостатки наихудшего анализа стека
- Наихудший анализ допуска требует очень жестких допусков отдельных компонентов. Это увеличивает общие затраты на производство и контроль.
Пример анализа стопки с допуском для наихудшего случая
Рассмотрим пример пяти дисков разного размера с разными допусками, уложенных друг на друга. Мы рассчитаем общую максимальную и минимальную общую высоту диска в стопке, используя метод анализа стопки наихудшего случая.
Анализ допусков для наихудшего случая для данной проблемы можно выполнить, выполнив следующие шаги:
Шаг 1. Создание цепочки размеров
Первым шагом в анализе набора допусков является создание цепочки размеров. Он используется для определения направления допуска. В приведенном выше примере все размеры имеют положительное направление.
Он используется для определения направления допуска. В приведенном выше примере все размеры имеют положительное направление.
Шаг 2 Расчет общей номинальной толщины
Общая номинальная толщина диска рассчитывается путем сложения номинальной толщины всех дисков. Поэтому
Общая номинальная Толщина = (15+10+15+12+15) = 67 мм
Шаг 3: Рассчитать общий допуск
Следующим шагом является сложение общих верхних и нижних допусков.
Общий верхний допуск = (0,5+0,1+0,2+0,3+0,4) = +1,5 мм
Общий нижний допуск = (0,5+0,1+0,2+0,3+0,4) = -1,5 мм
Шаг -4 Расчет верхнего и нижнего пределов
В этом примере максимальная и минимальная высота укладки диска вычисляются путем сложения номинальных размеров с верхним и нижним допусками соответственно.
Макс. Высота = Верхний предел = Номинальный размер + Допуск = 67 + 1,5 = 68,5 мм
Мин. Высота = Нижний предел = Номинальный размер – Допуск = 67 – 1,5 = 65,5 мм
Высота = Нижний предел = Номинальный размер – Допуск = 67 – 1,5 = 65,5 мм
Заключение
В приведенном выше примере общая высота/толщина уложенных друг на друга дисков будет варьироваться от 65,5 мм до 68,5 мм. Мы предлагаем вам также использовать этот калькулятор для расчета общей высоты диска с использованием анализа допусков наихудшего случая.
Статистический анализ допусков
Статистический допуск Анализ не фокусируется на предельных размерах. Потому что изменение размеров изготовленных деталей не является линейным. Каждое измерение имеет уникальное распределение, основанное на процессе изготовления детали, оборудовании и других параметрах.
Статистический анализ допуска основан на кривой Белла и нормальном распределении. Например, при изготовлении тысячи дисков одного размера. Толщина диска будет варьироваться от верхнего предела к нижнему пределу. Но все диски не будут иметь одинаковые размеры, потому что их толщина будет распределена.
Следующие два типа статистических допусков широко используются на рынке.
- Анализ стека допусков RSS
- Моделирование Монте-Карло для анализа стека допусков
В этой статье мы сосредоточимся на анализе стека допусков RSS.
Преимущества статистического анализа допусков
- Статистический анализ допусков позволяет увеличить пределы допусков компонентов. В худшем случае допуски деталей становятся очень жесткими, что увеличивает стоимость детали.
Недостатки статистического анализа допусков
- Когда детали проектируются с использованием статистического анализа допусков. Размеры изготовленных деталей могут выйти за допустимые пределы, а детали могут быть забракованы. Количество забракованных деталей зависит от производственных возможностей детали (3 σ, 4 σ, 6 σ )
- Статистический анализ допуска не дает 100% гарантии сборки детали.
Корень суммы квадратов (RSS) Анализ стека допусков Пример:
Анализ стека допусков по сумме квадратов (RSS) работает на основе статистического подхода. Предполагается, что большая часть деталей попадает в середину поля допуска.
Предполагается, что большая часть деталей попадает в середину поля допуска.
Рассмотрим пример пяти дисков разного размера с разными допусками, уложенных друг на друга. Мы рассчитаем общую максимальную и минимальную общую высоту диска в стопке, используя метод анализа стека RSS-допусков. Анализ стека допусков RSS для данной проблемы можно выполнить, выполнив следующие шаги:
Предположения
Для этого расчета мы будем считать, что производственный процесс 3σ способен, а значение Cpk равно 1 .
Шаг 1 Создание размерной цепочки
Подобно анализу допусков в наихудшем случае, первым шагом в анализе стека допусков RSS является создание размерной цепочки. Он используется для определения направления допуска. В приведенном выше примере все размеры имеют положительное направление.
Шаг 2 Расчет общей номинальной толщины
Общая номинальная толщина диска рассчитывается аналогично тому, как это делается при анализе наихудшего случая наложения, путем сложения номинальной толщины всех дисков. Таким образом,
Таким образом,
Общая номинальная толщина = (15+10+15+12+15) = 67 мм
Шаг 3 Рассчитайте стандартное отклонение для каждого допуска
Стандартное отклонение рассчитывается с учетом того, что производственный процесс допускает 3σ. Математически стандартное отклонение равно удвоенной способности процесса, потому что процесс распределен с обеих сторон. Следовательно, стандартное отклонение для процесса 3σ определяется как:
Стандартное отклонение = Общий допуск / (2 X 3)
σ1 = (0,5+0,5) / (2X3) = 0,166
σ2 = (0,1+0,1) / (2X3) = 0,033
σ0,2+0,2) / (2X3) = 0,066
σ4 = (0,3+0,3) / (2X3) = 0,1
σ5 = (0,4+0,4) / (2X3) = 0,133
Шаг 3 Рассчитать стандартное отклонение для Сборка
Стандартное отклонение для сборки равно корню из суммы квадратов стандартного отклонения отдельного размера.
σ (сборка) = √ [(σ1)² + (σ2)² + (σ3)² + (σ4)² + (σ5)²]
= √ [(0,166)² + (0,033)² + (0,066 ² + (0,1) ² + (0,133) ²]
σ (Сборка) = 0,2472
САК САКОЙ СЛЕДУЮЩИЙ СЛУЧА требуемой производительности процесса и общего стандартного отклонения для сборки.
 Чем ниже значение возможности процесса, тем узкой будет зона допуска и выше будет отбраковка.
Чем ниже значение возможности процесса, тем узкой будет зона допуска и выше будет отбраковка. Зона допуска (Для требуемой возможности процесса = 3σ) = σ (сборка) X 3 = 0,74162
Зона допуска (Для требуемой возможности процесса = 6σ) = σ4 1 X σ4 (сборка)
Шаг 5: Рассчитать верхний и нижний пределы
В этом примере максимальная и минимальная высота стопки дисков вычисляются путем сложения номинального размера с верхним и нижним допусками, соответственно, с учетом требуемого значения производительности процесса, равного 3σ.
Макс. Высота = Верхний предел = Номинальный размер + Допуск = 67 + 0,74162 = 67,74162 мм
Мин. Высота = Нижний предел = Номинальный размер – Допуск = 67 – 0,74162 = 66,2584 мм
Заключение
В приведенном выше примере общая высота/толщина диска в стопке будет варьироваться от 66,258 мм до 67,74 мм. Мы предлагаем вам также использовать этот калькулятор для расчета общей высоты диска с использованием анализа допусков наихудшего случая.
Мы предлагаем вам также использовать этот калькулятор для расчета общей высоты диска с использованием анализа допусков наихудшего случая.
Чем отличаются результаты анализа наихудшего случая и допуска RSS?
В соответствии с анализом наихудшего случая допуска на стопку высота стопки дисков может варьироваться от 65,5 мм до 68,5 мм. Тогда как по методу РСС высота диска может варьироваться от 66,2584 мм до 67,74162 мм.
Если вы посмотрите на результаты наихудшего случая и метода RSS. В методе RSS диапазон допусков уменьшается. Поэтому дизайнеры могут предоставить производителям больше гибкости.
Калькулятор суммирования допусков
Вы также можете использовать наш калькулятор суммирования допусков для наихудшего случая и метода RSS для расчета суммирования допусков. Здесь вы можете скачать таблицу расчета допустимых отклонений.
Подводя итог, можно сказать, что анализ набора допусков является очень важной частью разработки продукта. Это также помогает снизить стоимость деталей. Доступны различные методы анализа наложения допусков.
Это также помогает снизить стоимость деталей. Доступны различные методы анализа наложения допусков.
Есть вопросы? Мы будем рады помочь.
Если вы думаете, что мы что-то пропустили? Вы можете дополнить эту статью, отправив сообщение в поле для комментариев. Мы постараемся добавить его в этот пост.
Хронический миелогенный лейкоз (ХМЛ) Клиническая картина: анамнез, физикальное обследование
-
Редакционный совет по лечению взрослых PDQ. Лечение хронического миелогенного лейкоза (PDQ®): Версия для медицинских работников. 5 марта 2021 г. [Ссылка на QxMD MEDLINE]. [Полный текст].
-
Сойерс CL. Хронический миелоидный лейкоз. N Английский J Med . 1999 г., 29 апреля. 340(17):1330-40. [Ссылка QxMD MEDLINE].
-
Друкер Б.Дж., Сойерс К.Л., Кантарджян Х. и др. Активность специфического ингибитора тирозинкиназы BCR-ABL при бластном кризе хронического миелолейкоза и острого лимфобластного лейкоза с филадельфийской хромосомой.
 N Engl J Med . 2001 г., 5 апреля. 344(14):1038-42. [Ссылка QxMD MEDLINE]. [Полный текст].
N Engl J Med . 2001 г., 5 апреля. 344(14):1038-42. [Ссылка QxMD MEDLINE]. [Полный текст]. -
Kantarjian H, Sawyers C, Hochhaus A и др. для Международной исследовательской группы STI571 CML. Гематологические и цитогенетические ответы на мезилат иматиниба при хроническом миелогенном лейкозе. N Английский J Med . 2002 г., 28 февраля. 346(9):645-52. [Ссылка QxMD MEDLINE]. [Полный текст].
-
Merx K, Muller MC, Kreil S, et al. Раннее снижение уровней транскриптов мРНК BCR-ABL предсказывает цитогенетический ответ у пациентов с ХМЛ в хронической фазе, получавших иматиниб, после неэффективности интерферона альфа. Лейкемия . 2002 г., 16 сентября (9): 1579-83. [Ссылка QxMD MEDLINE]. [Полный текст].
-
Talpaz M, Silver RT, Druker BJ и др. Иматиниб индуцирует стойкий гематологический и цитогенетический ответ у пациентов с хроническим миелоидным лейкозом в фазе акселерации: результаты исследования фазы 2.
 Кровь . 2002 г., 15 марта. 99(6):1928-37. [Ссылка QxMD MEDLINE]. [Полный текст].
Кровь . 2002 г., 15 марта. 99(6):1928-37. [Ссылка QxMD MEDLINE]. [Полный текст]. -
Kantarjian HM, Cortes JE, O’Brien S, et al. Терапия мезилатом иматиниба у впервые диагностированных пациентов с хроническим миелогенным лейкозом с филадельфийской хромосомой: высокая частота ранних полных и основных цитогенетических ответов. Кровь . 2003 1 января. 101 (1): 97-100. [Ссылка QxMD MEDLINE]. [Полный текст].
-
Шах Н.П., Тран С., Ли Ф.Ю. и др. Преодоление устойчивости к иматинибу с помощью нового ингибитора киназы ABL. Наука . 2004 г., 16 июля. 305 (5682): 399-401. [Ссылка QxMD MEDLINE].
-
Volpe G, Panuzzo C, Ulisciani S, Cilloni D. Резистентность к иматинибу при ХМЛ. Рак Летт . 2009 8 февраля. 274(1):1-9. [Ссылка QxMD MEDLINE].
-
 webmd.com»> Фадерл С., Талпаз М., Эстров З., О’Брайен С., Курцрок Р., Кантарджян Х.М. Биология хронического миелоидного лейкоза. N Engl J Med . 1999 г., 15 июля. 341(3):164-72. [Ссылка QxMD MEDLINE].
webmd.com»> Фадерл С., Талпаз М., Эстров З., О’Брайен С., Курцрок Р., Кантарджян Х.М. Биология хронического миелоидного лейкоза. N Engl J Med . 1999 г., 15 июля. 341(3):164-72. [Ссылка QxMD MEDLINE]. -
Факты и цифры о раке, 2022 г. Американское онкологическое общество. Доступно по адресу https://www.cancer.org/content/dam/cancer-org/research/cancer-facts-and-statistics/annual-cancer-facts-and-figures/2022/2022-cancer-facts-and- рисунки.pdf. Доступ: 27 мая 2022 г.
-
Факты статистики рака: Лейкемия — Хронический миелоидный лейкоз (ХМЛ). Программа наблюдения, эпидемиологии и конечных результатов Национального института рака. Доступно по адресу https://seer.cancer.gov/statfacts/html/cmyl.html. Доступ: 27 мая 2022 г.
-
Дыбко Ю., Язвец Б., Хаус О., Урбаняк-Куйда Д., Капелько-Словик К., Врубель Т. и др.
 Шкала Хасфорда может предсказывать молекулярный ответ у пациентов с хроническим миелоидным лейкозом: опыт одного учреждения. Маркеры Dis . 2016. 2016:7531472. [Ссылка QxMD MEDLINE]. [Полный текст].
Шкала Хасфорда может предсказывать молекулярный ответ у пациентов с хроническим миелоидным лейкозом: опыт одного учреждения. Маркеры Dis . 2016. 2016:7531472. [Ссылка QxMD MEDLINE]. [Полный текст]. -
Gambacorti-Passerini C, Antolini L, Mahon FX, Guilhot F, Deininger M al et. Многоцентровая независимая оценка исходов у пациентов с хроническим миелоидным лейкозом, получавших иматиниб. J Natl Cancer Inst . 2011 6 апреля. 103(7):553-61. [Ссылка QxMD MEDLINE].
-
Hochhaus A, Larson RA, Guilhot F, Radich JP, Branford S, Hughes TP, et al. Долгосрочные результаты лечения иматинибом хронического миелоидного лейкоза. N Английский J Med . 2017 9 марта. 376 (10): 917-927. [Ссылка QxMD MEDLINE].
-
Wang W, Cortes JE, Tang G, Khoury JD, Wang S, Bueso-Ramos CE, et al. Стратификация риска хромосомных аномалий при хроническом миелолейкозе в эпоху терапии ингибиторами тирозинкиназы.
 Кровь . 2016 марта 22. [Ссылка QxMD MEDLINE].
Кровь . 2016 марта 22. [Ссылка QxMD MEDLINE]. -
«Иматиниб изменил все»: будущее стало более оптимистичным. Медицинские новости Medscape. Доступно на http://www.medscape.com/viewarticle/876942. 9 марта 2017 г.; Доступ: 10 марта 2017 г.
-
Исламагич Э., Хасич А., Куртович С. и др. Эффективность непатентованного иматиниба в качестве терапии первой и второй линии: 3-летнее наблюдение за пациентами с хроническим миелоидным лейкозом. Clin Лимфома Миелома Лейк . 2017 Feb 16. [Полный текст].
-
Saussele S, et al; Следователи ЕВРО-СКИ. Прекращение терапии ингибиторами тирозинкиназы при хроническом миелоидном лейкозе (EURO-SKI): предварительный промежуточный анализ проспективного, многоцентрового, нерандомизированного исследования. Ланцет Онкол . 2018 19 июня (6): 747-757. [Ссылка QxMD MEDLINE].

-
[рекомендации] Клинические рекомендации NCCN по онкологии (рекомендации NCCN): хронический миелоидный лейкоз. Национальная комплексная онкологическая сеть. Доступно по адресу https://www.nccn.org/professionals/physician_gls/pdf/cml.pdf. Версия 3.2022 — 27 января 2022 г.; Доступ: 27 мая 2022 г.
-
[Рекомендации] Hochhaus A, Saussele S, Rosti G, Mahon FX, Janssen JJWM, Hjorth-Hansen H, et al. Хронический миелоидный лейкоз: Клинические практические рекомендации ESMO по диагностике, лечению и последующему наблюдению. Энн Онкол . 2018 1 окт. 29 (Приложение 4): iv261. [Ссылка QxMD MEDLINE]. [Полный текст].
-
[Руководство] Баккарани М., Дейнингер М.В., Рости Г. и соавт. Европейские рекомендации LeukemiaNet по лечению хронического миелоидного лейкоза: 2013 г. Кровь . 2013 8 августа. 122 (6): 872-84.
 [Ссылка QxMD MEDLINE]. [Полный текст].
[Ссылка QxMD MEDLINE]. [Полный текст]. -
Барретт А.Дж., Ито С. Роль трансплантации стволовых клеток при хроническом миелогенном лейкозе в 21 веке. Кровь . 2015 21 мая. 125 (21): 3230-5. [Ссылка QxMD MEDLINE].
-
Друкер Б.Дж., Талпаз М., Реста Д.Дж. и др. Эффективность и безопасность специфического ингибитора тирозинкиназы BCR-ABL при хроническом миелоидном лейкозе. N Английский J Med . 2001 г., 5 апреля. 344(14):1031-7. [Ссылка QxMD MEDLINE]. [Полный текст].
-
Джаббур Э., Кантарджян Х. Хронический миелоидный лейкоз: обновленная информация о диагностике, терапии и мониторинге за 2020 г. Ам Дж Гематол . 2020 июнь 95 (6): 691-709. [Ссылка QxMD MEDLINE].
-
Сантос Ф.П., Альварадо Ю., Кантарджян Х., Верма Д., О’Брайен С.
 , Маттиуцци Г. и др. Долгосрочное прогностическое влияние применения стимуляторов эритропоэза у пациентов с хроническим миелоидным лейкозом в хронической фазе, получающих иматиниб. Рак . 2011 1 марта. 117 (5): 982-91. [Ссылка QxMD MEDLINE].
, Маттиуцци Г. и др. Долгосрочное прогностическое влияние применения стимуляторов эритропоэза у пациентов с хроническим миелоидным лейкозом в хронической фазе, получающих иматиниб. Рак . 2011 1 марта. 117 (5): 982-91. [Ссылка QxMD MEDLINE]. -
Sawyers CL, Hochhaus A, Feldman E, et al. Иматиниб индуцирует гематологические и цитогенетические ответы у пациентов с хроническим миелогенным лейкозом при миелоидном бластном кризе: результаты исследования II фазы. Кровь . 2002 г., 15 мая. 99(10):3530-9. [Ссылка QxMD MEDLINE].
-
Тан М., Гонен М., Квинтас-Кардама А. и др. Динамика ответа хронического миелоидного лейкоза на длительную таргетную терапию позволяет выявить влияние лечения на лейкемические стволовые клетки. Кровь . 2011 11 августа. 118(6):1622-31. [Ссылка QxMD MEDLINE].
-
Ибрагим А.
 Р., Элиассон Л., Апперли Дж.Ф., Милойкович Д., Буа М., Шидло Р. и др. Плохая приверженность является основной причиной потери CCyR и неэффективности иматиниба у пациентов с хроническим миелоидным лейкозом при длительной терапии. Кровь . 7 апреля 2011 г. 117 (14): 3733-6. [Ссылка QxMD MEDLINE].
Р., Элиассон Л., Апперли Дж.Ф., Милойкович Д., Буа М., Шидло Р. и др. Плохая приверженность является основной причиной потери CCyR и неэффективности иматиниба у пациентов с хроническим миелоидным лейкозом при длительной терапии. Кровь . 7 апреля 2011 г. 117 (14): 3733-6. [Ссылка QxMD MEDLINE]. -
Hochhaus A, Kreil S, Corbin AS, La Rosée P, Müller MC, Lahaye T, et al. Молекулярные и хромосомные механизмы устойчивости к терапии иматинибом (STI571). Лейкемия . 2002 16 ноября (11): 2190-6. [Ссылка QxMD MEDLINE].
-
Марколино М.С., Боэрсма Э., Клементино Н.К. и др. Продолжительность лечения иматинибом связана со снижением предполагаемой скорости клубочковой фильтрации у пациентов с хроническим миелоидным лейкозом. Энн Онкол . 22 сентября 2011 г. (9): 2073-9. [Ссылка QxMD MEDLINE].
-
Дазатиниб.
Национальный институт рака. Доступно по адресу https://www.cancer.gov/about-cancer/treatment/drugs/dasatinib. 5 июня 2019 г.; Доступ: 27 мая 2022 г. -
Нилотиниб. Национальный институт рака. Доступно по адресу https://www.cancer.gov/about-cancer/treatment/drugs/nilotinib. 30 декабря 2020 г .; Доступ: 27 мая 2022 г.
-
Босулиф (бозутиниб) [вкладыш в упаковку]. Нью-Йорк, штат Нью-Йорк: Pfizer, Inc., июнь 2020 г. Доступно на [Полный текст].
-
Джаббур Э., Кантарджян Х., О’Брайен С. и др. Достижение раннего полного цитогенетического ответа является основным определяющим фактором исхода у пациентов с ранней хронической фазой хронического миелоидного лейкоза, получающих лечение ингибиторами тирозинкиназы. Кровь . 2011, 27 октября. 118(17):4541-6; викторина 4759. [Ссылка QxMD MEDLINE].
-
webmd.com»> Hughes TP, Hochhaus A, Branford S, Müller MC, Kaeda JS, Foroni L, et al. Долгосрочное прогностическое значение раннего молекулярного ответа на иматиниб при недавно диагностированном хроническом миелоидном лейкозе: анализ Международного рандомизированного исследования интерферона и ИППП571 (IRIS). Кровь . 2010 11 ноября. 116(19):3758-65. [Ссылка QxMD MEDLINE].
-
Управление по санитарному надзору за качеством пищевых продуктов и медикаментов. Sprycel (дазатиниб): Сообщение о безопасности лекарственных средств – риск легочной артериальной гипертензии. Управление по санитарному надзору за качеством пищевых продуктов и медикаментов США. Доступно по адресу https://wayback.archive-it.org/79.93/20170112165154/http://www.fda.gov/Safety/MedWatch/SafetyInformation/SafetyAlertsforHumanMedicalProducts/ucm275176.htm. 11 октября 2011 г.; Доступ: 24 августа 2020 г.
-
com»> Сойерс CL. Еще лучше ингибиторы киназы при хроническом миелоидном лейкозе. N Английский J Med . 2010 17 июня. 362(24):2314-5. [Ссылка QxMD MEDLINE].
-
Джаббур Э., Кантарджян Х., О’Брайен С., Шан Дж., Гарсия-Манеро Г., Вирда В. и др. Прогностические факторы для исхода и ответа у пациентов, получавших ингибиторы тирозинкиназы второго поколения при хроническом миелоидном лейкозе в хронической фазе после неудачи иматиниба. Кровь . 2010, 28 октября. [Ссылка на MEDLINE QxMD].
-
Верма Д., Кантарджян Х., Стром С.С. и др. Злокачественные новообразования, возникающие во время терапии ингибиторами тирозинкиназы (ИТК) при хроническом миелоидном лейкозе (ХМЛ) и других гематологических злокачественных новообразованиях. Кровь . 2011 20 октября. 118 (16): 4353-8. [Ссылка QxMD MEDLINE].
-
Кантарджян Х.
, Шах Н.П., Хоххаус А., Кортес Дж., Шах С., Айяла М. и др. Дазатиниб по сравнению с иматинибом при впервые диагностированной хронической фазе хронического миелоидного лейкоза. N Engl J Med . 2010 17 июня. 362(24):2260-70. [Ссылка QxMD MEDLINE]. -
Кортес Дж. Э., Джонс Д., О’Брайен С., Джаббур Э., Раванди Ф., Коллер С. и др. Результаты терапии дазатинибом у больных с ранней хронической фазой хронического миелоидного лейкоза. J Клин Онкол . 2010 20 января. 28(3):398-404. [Ссылка QxMD MEDLINE]. [Полный текст].
-
Mulcahy N. FDA обновляет этикетку дазатиниба для лечения хронического миелоидного лейкоза. Медицинские новости Medscape . 26 июня 2013 г. [Полный текст].
-
Джаббур Э., Кантарджян Х.М., Сальо Г., Стигманн Д.Л., Шах Н.П., Боке К. и др. Ранний ответ на дазатиниб или иматиниб при хроническом миелоидном лейкозе: 3-летнее наблюдение в рандомизированном исследовании фазы 3 (DASISION).
Кровь . 2014 23 января. 123 (4): 494-500. [Ссылка QxMD MEDLINE]. [Полный текст]. -
Shah NP, Guilhot F, Cortes JE, Schiffer CA, le Coutre P, Brümmendorf TH, et al. Долгосрочные результаты дазатиниба после неэффективности иматиниба при хронической фазе хронического миелоидного лейкоза: последующее наблюдение за исследованием фазы 3. Кровь . 2014, 25 февраля. [Ссылка на MEDLINE QxMD].
-
Saglio G, Kim DW, Issaragrisil S, le Coutre P, Etienne G, Lobo C, et al. Нилотиниб в сравнении с иматинибом при недавно диагностированном хроническом миелоидном лейкозе. N Английский J Med . 2010 17 июня. 362(24):2251-9. [Ссылка QxMD MEDLINE].
-
Kantarjian HM, Hochhaus A, Saglio G, et al. Нилотиниб по сравнению с иматинибом для лечения пациентов с недавно диагностированной хронической фазой хронического миелоидного лейкоза с положительной филадельфийской хромосомой: минимальное 24-месячное наблюдение за рандомизированным исследованием ENESTnd фазы 3.
Ланцет Онкол . 2011 Сентябрь 12 (9): 841-51. [Ссылка QxMD MEDLINE]. -
Tasigna (нилотиниб) [вкладыш в упаковку]. Восточный Ганновер, Нью-Джерси 07936: Novartis Pharmaceuticals Corporation. Июнь 2020 г. Доступно на [Полный текст].
-
Хури Х.Дж., Кортес Дж.Э., Кантарджян Х.М., Гамбакорти-Пассерини С., Баккарани М., Ким Д.В. и др. Бозутиниб активен при хронической фазе хронического миелоидного лейкоза после неэффективности терапии иматинибом и дазатинибом и/или нилотинибом. Кровь . 2012 12 апреля. 119(15):3403-12. [Ссылка QxMD MEDLINE].
-
Cortes JE, Gambacorti-Passerini C, Deininger MW, Mauro MJ, Chuah C, Kim DW, et al. Бозутиниб в сравнении с иматинибом при недавно диагностированном хроническом миелоидном лейкозе: результаты рандомизированного исследования BFORE. J Клин Онкол .
1 ноября 2017 г. JCO2017747162. [Ссылка QxMD MEDLINE]. [Полный текст]. -
Тим Х. Брюммендорф, Хорхе Э. Кортес, Драгана Милойкович, Карло Гамбакорти-Пассерини и др. . Бозутиниб (ЛС) по сравнению с иматинибом для недавно диагностированной хронической фазы (ХП) хронического миелоидного лейкоза (ХМЛ): окончательные 5-летние результаты исследования Bfore. Кровь . 2020. 136; (Приложение 1):41-42. [Полный текст].
-
Chustecka Z. FDA одобрило понатиниб для лечения редких лейкозов. Медицинские новости Medscape. 14 декабря 2012 г. [Полный текст].
-
Cortes JE, Kantarjian H, Shah NP, Bixby D, Mauro MJ, Flinn I, et al. Понатиниб при рефрактерных лейкозах с филадельфийской хромосомой. N Английский J Med . 2012 29 ноября. 367(22):2075-88. [Ссылка QxMD MEDLINE].
-
webmd.com»> Cortes JE, Kim DW, Pinilla-Ibarz J, le Coutre P, Paquette R, et al. Испытание фазы 2 понатиниба при лейкозах с положительными хромосомами в Филадельфии. N Engl J Med . 2013 7 ноября. 369 (19): 1783-96. [Ссылка QxMD MEDLINE].
-
Сообщение FDA о безопасности лекарственных средств. FDA просит производителя лекарства от лейкемии Iclusig (понатиниб) приостановить маркетинг и продажи. Управление по санитарному надзору за качеством пищевых продуктов и медикаментов США. Доступно по адресу https://wayback.archive-it.org/7993/20170722185758/https://www.fda.gov/Drugs/DrugSafety/ucm373040.htm. 5 ноября 2013 г .; Доступ: 27 мая 2022 г.
-
Сообщение FDA о безопасности лекарственных средств: FDA требует несколько новых мер безопасности для препарата Iclusig от лейкемии; компания, как ожидается, возобновит маркетинг. Управление по санитарному надзору за качеством пищевых продуктов и медикаментов США.
Доступно по адресу https://wayback.archive-it.org/79.93/20170722185750/https://www.fda.gov/Drugs/DrugSafety/ucm379554.htm. 20 декабря 2013 г.; Доступ: 27 мая 2022 г. -
Сосселе С., Хаверкамп В., Ланг Ф., Кошмидер С., Киани А., Дженч-Ульрих К. и др. Понатиниб в лечении хронического миелоидного лейкоза и острого лейкоза с положительной филадельфийской хромосомой: рекомендации консенсусной группы немецких экспертов с упором на сердечно-сосудистую систему. Акта Гематол . 2020. 143 (3): 217-231. [Ссылка QxMD MEDLINE]. [Полный текст].
-
Scemblix (ациминиб) [вкладыш в упаковку]. Восточный Ганновер, Нью-Джерси: Novartis Pharmaceuticals, Corp., октябрь 2021 г. Доступно на [Полный текст].
-
Gotlib J, Kiladjian JJ, Vannucchi A, Rambaldi A, Reiter A, Shomali W, et al. Исследование фазы 2 пемигатиниба (FIGHT-203; INCB054828) у пациентов с миелоидными/лимфоидными новообразованиями (MLN) с перестройкой рецептора 1 фактора роста фибробластов (FGFR1) (MLN FGFR1).
Кровь. 2021 г., 23 ноября (138 (дополнение 1): 385. Представлено на 64-м виртуальном собрании Американского общества гематологов в 2021 г. [Полный текст]. -
Synribo (омацетаксин) [вкладыш]. Северный Уэльс, Пенсильвания: Teva Pharmaceuticals USA, Inc., октябрь 2012 г. Доступно по ссылке [Полный текст].
-
Симонссон Б., Гедде-Даль Т., Маркеварн Б. и др. Комбинация пегилированного IFN-a2b с иматинибом увеличивает частоту молекулярного ответа у пациентов с хроническим миелоидным лейкозом низкого или среднего риска. Кровь . 2011 22 сентября. 118(12):3228-35. [Ссылка QxMD MEDLINE].
-
Мореб Дж., Джонсон Т., Кубилис П., Майерс Л., Облон Д., Миллер А. и др. Повышение выживаемости больных хроническим миелогенным лейкозом, перенесших аллогенную трансплантацию костного мозга. Ам J Гематол . 1995 г.
, декабрь 50 (4): 304-6. [Ссылка QxMD MEDLINE]. -
МакГлэйв П.Б., Битти П., Эш Р., Хоус Дж.М. Терапия хронического миелогенного лейкоза с трансплантацией костного мозга от неродственного донора: результаты 102 случаев. Кровь . 1990 г., 15 апреля. 75(8):1728-32. [Ссылка QxMD MEDLINE].
-
Deininger M, Schleuning M, Greinix H, Sayer HG, Fischer T, Martinez J, et al. Влияние предшествующего воздействия иматиниба на смертность, связанную с трансплантацией. Гематологический . 2006 г., апрель 91 (4): 452-9. [Ссылка QxMD MEDLINE].
-
Лима Л., Бернал-Мизрачи Л., Сакс Д., Манн К.П., Тигиуарт М., Арельяно М. и др. Мониторинг периферической крови при хроническом миелоидном лейкозе во время лечения иматинибом, препаратами второй линии и в дальнейшем. Рак . 2011 15 марта. 117(6):1245-52. [Ссылка QxMD MEDLINE].
-
Baccarani M, Saglio G, Goldman J, Hochhaus A, Simonsson B, Appelbaum F, et al. Развитие концепций лечения хронического миелоидного лейкоза: рекомендации группы экспертов от имени European LeukemiaNet. Кровь . 2006 15 сентября. 108 (6): 1809-20. [Ссылка QxMD MEDLINE]. [Полный текст].
-
Брэнфорд С., Рудзки З., Харпер А., Григг А., Тейлор К., Даррант С. и др. Иматиниб дает значительно лучшие молекулярные ответы по сравнению с интерфероном альфа плюс цитарабин у пациентов с впервые диагностированным хроническим миелоидным лейкозом в хронической фазе. Лейкемия . 2003 г., 17 (12): 2401-9. [Ссылка QxMD MEDLINE].
-
Кортес Дж., Талпаз М., О’Брайен С., Джонс Д., Лутра Р., Шан Дж. и др. Молекулярные реакции у пациентов с хроническим миелогенным лейкозом в хронической фазе, получавших мезилат иматиниба.
Clin Cancer Res . 2005 1 мая. 11 (9): 3425-32. [Ссылка QxMD MEDLINE]. [Полный текст]. -
Брэнфорд С., Рудзки З., Уолш С., Паркинсон И., Григг А., Шер Дж. и др. Обнаружение мутаций BCR-ABL у пациентов с ХМЛ, получающих иматиниб, практически всегда сопровождается клинической резистентностью, а мутации в АТФ-фосфатсвязывающей петле (P-петле) связаны с неблагоприятным прогнозом. Кровь . 2003 г., 1 июля. 102 (1): 276-83. [Ссылка QxMD MEDLINE]. [Полный текст].
-
Press RD, Galderisi C, Yang R, Rempfer C, Willis SG, Mauro MJ, et al. Полулогарифмическое увеличение РНК BCR-ABL предсказывает более высокий риск рецидива у пациентов с хроническим миелоидным лейкозом с полным цитогенетическим ответом, индуцированным иматинибом. Clin Cancer Res . 2007 15 октября. 13 (20): 6136-43. [Ссылка QxMD MEDLINE]. [Полный текст].
-
webmd.com»> Сальо Г., Шарф Г., Алмейда А., Богданович А., Бомбачи Ф., Чугурович Дж. и др. Соображения относительно ремиссии без лечения у пациентов с хроническим миелоидным лейкозом: совместная точка зрения пациента и врача. Клин Лимфома Миелома Лейк . 2018 18 июня (6): 375-379. [Ссылка QxMD MEDLINE]. [Полный текст].
-
Sweet K, Oehler V. Прекращение приема ингибиторов тирозинкиназы при хроническом миелоидном лейкозе: когда это безопасный вариант? Гематология Am Soc Hematol Educ Program . 2013. 2013:184-8. [Ссылка QxMD MEDLINE]. [Полный текст].
-
Мулахи Н. Лекарство от лейкемии понатиниб (иклусиг) снято с продажи. Медицинские новости Medscape. Доступно на http://www.medscape.com/viewarticle/813531. 31 октября 2013 г.; Доступ: 24 августа 2020 г.
-
Нельсон Р. Понатиниб (Иклюзиг) возвращается на рынок с некоторыми оговорками.
Медицинские новости Medscape. Доступно на http://www.medscape.com/viewarticle/818183?src=wnl_edit_specol&uac=72886PK. 20 декабря 2013 г.; Доступ: 24 августа 2020 г.
Аортальный стеноз: диагностика и лечение
БРАЙАН Х. ГРИМАРД, MD, И ДЖАН М. ЛАРСОН, MD
Аортальный стеноз является наиболее распространенным заболеванием клапанов сердца в развитых странах, от него страдают 3 процента лиц старше 65 лет. Хотя выживаемость у бессимптомных пациентов с аортальным стенозом сравнима с таковой у контрольных пациентов того же возраста и пола, средняя общая выживаемость у пациентов с симптомами без протезирования аортального клапана составляет два-три года. В течение бессимптомного латентного периода гипертрофия левого желудочка и увеличение преднагрузки на предсердия компенсируют увеличение постнагрузки, вызванное аортальным стенозом. По мере ухудшения заболевания эти компенсаторные механизмы становятся неадекватными, что приводит к симптомам сердечной недостаточности, стенокардии или обморокам. Замена аортального клапана должна быть рекомендована большинству пациентов с любым из этих симптомов, сопровождающихся признаками значительного аортального стеноза при эхокардиографии. Выжидательная тактика рекомендуется для большинства бессимптомных пациентов, в том числе с гемодинамически значимым аортальным стенозом. Пациенты должны быть информированы о симптомах и важности незамедлительного сообщения о них своим врачам. Серийную допплеровскую эхокардиографию рекомендуется проводить ежегодно при тяжелом аортальном стенозе, каждые 1–2 года при умеренном стенозе и каждые 3–5 лет при легком аортальном стенозе. Направление к кардиологу рекомендуется для всех пациентов с симптоматическим аортальным стенозом, с тяжелым аортальным стенозом без явных симптомов и с дисфункцией левого желудочка. Многие пациенты с бессимптомным аортальным стенозом имеют сопутствующие заболевания сердца, такие как артериальная гипертензия, мерцательная аритмия и ишемическая болезнь сердца, которые также следует тщательно контролировать.
Стеноз аортального клапана поражает 3% лиц старше 65 лет и приводит к большей заболеваемости и смертности, чем другие заболевания сердечного клапана. 1 Патология аортального стеноза включает процессы, сходные с таковыми при атеросклерозе, включая накопление липидов, воспаление и кальцификацию. 2 Развитие значительного аортального стеноза, как правило, происходит раньше у пациентов с врожденными двустворчатыми аортальными клапанами. 3 Хотя выживаемость бессимптомных пациентов сравнима с выживаемостью контрольных пациентов того же возраста и пола, выживаемость заметно ухудшается после появления симптомов.
| Clinical recommendation | Evidence rating | References |
|---|---|---|
| Echocardiography is recommended in patients with classic symptoms of aortic stenosis accompanied by a systolic murmur and in asymptomatic patients with a grade 3/ 6 и более громкий систолический шум. | C | 19 |
| Замена аортального клапана является единственным эффективным методом лечения пациентов с тяжелым симптоматическим аортальным стенозом. | B | 25 , 26 |
| Выжидательная тактика рекомендуется большинству пациентов с бессимптомным аортальным стенозом, в том числе с тяжелым течением заболевания. | B | 28 , 30 , 31 |
| Серийная допплеровская эхокардиография рекомендуется ежегодно при тяжелом аортальном стенозе, каждые пять-три года при умеренном стенозе аорты и каждые 1-3 года при умеренном стенозе. | C | 19 |
| Профилактическое назначение аспирина следует рассматривать у взрослых с 10-летним риском сердечно-сосудистых заболеваний 6 процентов или выше, что характерно для аортального стеноза. | A | 40 |
| Интенсивная гиполипидемическая терапия не останавливает прогрессирование кальцифицирующего аортального стеноза и не рекомендуется при отсутствии других показаний к терапии статинами. | С | 44 |
Патофизиология и естественное течение
Естественное течение аортального стеноза включает длительный латентный период, в течение которого прогрессирующее ухудшение обструкции оттока левого желудочка (ЛЖ) приводит к гипертрофическим изменениям в левом желудочке. 4,5 Когда площадь аортального клапана становится менее половины своего нормального размера от 3 до 4 см 2 (размером примерно с пятицентовую монету), может быть обнаружен измеряемый градиент давления между левым желудочком и восходящей аортой на эхокардиография или путем прямого измерения при катетеризации сердца. 6 Это изменение отражает компенсаторное повышение давления в ЛЖ, которое способствует поддержанию адекватного системного давления. Одним из последствий является гипертрофия ЛЖ с последующей диастолической дисфункцией и повышенным сопротивлением наполнению ЛЖ. 7,8 Таким образом, для обеспечения достаточного диастолического наполнения ЛЖ и адекватного ударного объема необходимо сильное сокращение левого предсердия. 9 Повышение общей сократимости миокарда и увеличение преднагрузки с усилением предсердного толчка сохраняет систолическую функцию ЛЖ, как правило, пока пациент остается бессимптомным.
По мере ухудшения аортального стеноза с уменьшением площади аортального клапана до 1 см 2 или меньше (примерно размером с головку мяча для гольфа), 10 изменения функции ЛЖ могут быть недостаточными для преодоления оттока обструкции и поддерживать систолическую функцию, даже когда она дополняется увеличением преднагрузки. Возникающее в результате нарушение систолической функции, само по себе или в сочетании с диастолической дисфункцией, может привести к клинической сердечной недостаточности. Прогрессирующая гипертрофия ЛЖ вследствие аортального стеноза также приводит к увеличению потребности миокарда в кислороде 11 ; одновременно гипертрофия миокарда может сдавливать интрамуральные коронарные артерии, поскольку они несут кровь к эндокарду. Эти изменения наряду со снижением диастолического наполнения коронарных артерий могут привести к классической стенокардии даже при отсутствии ишемической болезни сердца (ИБС). 12 Кроме того, когда аортальный стеноз становится тяжелым, сердечный выброс больше не увеличивается при физической нагрузке. 13 В этих условиях падение системного сосудистого сопротивления, которое обычно происходит при физической нагрузке, может привести к гипотензии и обмороку. 14–16
На рисунке 1 показаны эхокардиограммы нормального аортального клапана, а на рисунке показаны эхокардиограммы тяжелого аортального стеноза.
Диагностика
ПРИЗНАКИ И СИМПТОМЫ
Классические симптомы аортального стеноза включают одышку и другие симптомы сердечной недостаточности, стенокардию и обмороки. Появление этих классических симптомов указывает на гемодинамически значимый аортальный стеноз и является критическим моментом для принятия управленческих решений. Также важно понимать, что презентации могут различаться. Некоторые пациенты с тяжелым аортальным стенозом, особенно пожилые пациенты, могут проявляться более тонко и первоначально испытывать снижение толерантности к физической нагрузке без распознавания классических симптомов. У других может быть более острое течение, иногда с симптомами, вызванными сопутствующими заболеваниями. Например, впервые возникшая мерцательная аритмия с последующим снижением наполнения предсердий может привести к симптомам сердечной недостаточности, а назначение сосудорасширяющих препаратов может вызвать обмороки.
Классическим физикальным признаком аортального стеноза является грубый систолический шум, нарастающий-декрещендо, который наиболее громок во втором межреберье справа и иррадиирует в сонные артерии. Это может сопровождаться медленным и замедленным подъемом сонной артерии, устойчивым верхушечным импульсом и отсутствием или ослаблением второго тона аорты. Однако у пожилых людей шум может быть менее интенсивным и часто иррадиирует на верхушку, а не на сонные артерии. Также при повышенной заболеваемости атеросклерозом и артериальной гипертензией у пожилых людей могут маскироваться классические изменения пульса сонных артерий. Трудно исключить аортальный стеноз только на основании результатов физикального обследования. Надежные аускультативные признаки отсутствия аортального стеноза включают систолический шум 1/6 степени или мягче, отсутствие иррадиирующего систолического шума, выслушиваемого над головкой правой ключицы, и наличие расщепленного второго тона сердца. 17,18
Крайне важно, чтобы врачи первичной медико-санитарной помощи рассматривали аортальный стеноз у взрослых с любым из классических симптомов, сопровождающихся систолическим шумом. Пожилые пациенты с нечеткими жалобами, такими как утомляемость или снижение толерантности к физической нагрузке, а также бессимптомные пациенты, проходящие предоперационное медицинское обследование, должны пройти дальнейшее обследование, если при физикальном обследовании выявляется систолический шум силой 2/6 или выше. Практические рекомендации Американского колледжа кардиологов/Американской кардиологической ассоциации (ACC/AHA) рекомендуют эхокардиографию бессимптомным пациентам с уровнем систолического шума 3/6 или выше. 19
ДИАГНОСТИЧЕСКОЕ ОБСЛЕДОВАНИЕ
Допплер-эхокардиография рекомендуется для пациентов с классическими симптомами аортального стеноза. 19 Это полезно для оценки площади аортального клапана, пикового и среднего трансклапанных градиентов и максимальной скорости аорты. Это первичные меры для оценки тяжести заболевания, и они были хорошо подтверждены по сравнению с измерениями, полученными при катетеризации сердца. 20
Эхокардиография также предоставляет полезную информацию о функции ЛЖ, давлении наполнения левого желудочка и сопутствующих аномалиях других клапанов. Рекомендации по соотнесению этих гемодинамических показателей с тяжестью аортального стеноза представлены в Таблица 1 . 19 Изолированный аортальный стеноз редко проявляется симптомами до тех пор, пока площадь аортального клапана не станет менее 1 см 2 , средний градиент не превысит 40 мм рт. ст. или скорость струи аорты не превысит 4 м в секунду.
| Тяжесть | Скорость аортальной струи (м в секунду) | Средний градиент (мм рт.ст.) | Площадь аортального клапана0006 | ||||
|---|---|---|---|---|---|---|---|
| Normal | < 2.5 | — | 3 to 4 | ||||
| Mild | 2.5 to 2.9 | < 25 | 1.5 to 2 | ||||
| Moderate | 3 to 4 | от 25 до 40 | от 1 до 1,5 | ||||
Тяжелая
Лечение Замена аортального клапана является единственным эффективным методом лечения гемодинамически значимого аортального стеноза. Хирургия имеет средний уровень периоперационной смертности 4 процента 21–23 и риск отказа протеза клапана примерно 1 процент в год. СИМПТОМНЫЕ ПАЦИЕНТЫ Смертность резко возрастает после того, как аортальный стеноз становится симптоматическим. Средняя общая выживаемость составляет два-три года у симптоматических пациентов без хирургического лечения. Сообщается, что среди пожилых членов этой группы населения одно- и трехлетняя смертность составляет 44 и 75 процентов соответственно. 24 Хотя проспективных рандомизированных исследований, сравнивающих замену аортального клапана с медикаментозным лечением, не проводилось, ретроспективные обсервационные исследования показали, что хирургическое вмешательство приводит к значительному улучшению выживаемости, обычно сопровождающемуся улучшением симптомов. 25,26 10-летняя выживаемость пациентов возраста Medicare после протезирования аортального клапана практически идентична таковой у лиц того же возраста и пола, у которых нет аортального стеноза. У симптоматических пациентов с эхокардиографическими показателями, соответствующими тяжелому аортальному стенозу, следует предположить, что симптомы являются результатом аортального стеноза, даже если присутствуют другие потенциально причинные состояния, такие как ИБС. Однако, если данные эхокардиографии предполагают только умеренный аортальный стеноз, могут потребоваться дополнительные диагностические исследования (например, коронароангиография, исследование функции легких, оценка аритмии). БЕССИМПТОМНЫЕ ПАЦИЕНТЫ Замена аортального клапана также рекомендуется бессимптомным пациентам с тяжелым аортальным стенозом, сопровождающимся систолической дисфункцией ЛЖ (т. Однако большинству бессимптомных пациентов с аортальным стенозом, в том числе с тяжелым заболеванием, рекомендуется выжидательная тактика. Выживаемость у пациентов с аортальным стенозом, которым проводят выжидательную тактику, сравнима с таковой у пациентов без аортального стеноза. Кроме того, хирургический риск превышает примерно 1% ежегодный риск внезапной смерти у бессимптомных пациентов с аортальным стенозом. Были предприняты попытки выявить тех, у кого с большей вероятностью будут неблагоприятные исходы без «ранней» замены аортального клапана. Пациенты с очень тяжелым аортальным стенозом (площадь аортального клапана 0,6 см 2 или меньше, или скорость струи аорты 5 м в секунду или более), более быстрое увеличение скорости струи аорты с течением времени (0,3 м в секунду или более в год), или тяжелая кальцификация клапана имеют высокий риск развития симптомов и необходимости замены аортального клапана в течение следующих одного-двух лет. Важно отличать пациентов, которые действительно бессимптомны, от тех, у которых уровень обычной активности снизился до уровня ниже порога симптомов. Это особенно важно для пожилых пациентов, которые могут приписывать свои симптомы нормальному старению или сопутствующим заболеваниям. Когда статус симптомов неясен, рекомендации ACC/AHA рекомендуют проводить тесты с физической нагрузкой под непосредственным наблюдением опытного кардиолога. 19 Замену аортального клапана следует рассматривать, если симптомы возникают, когда частота сердечных сокращений пациента составляет менее 80 процентов от прогнозируемой максимальной частоты сердечных сокращений, или если у пациента наблюдается ненормальная реакция артериального давления. НАБЛЮДЕНИЕ И НАПРАВЛЕНИЕ Выжидательная тактика до выявления симптомов уместна у большинства пациентов с бессимптомным аортальным стенозом. Рекомендации ACC/AHA рекомендуют проводить серийную допплер-эхокардиографию ежегодно при тяжелом аортальном стенозе, каждые один-два года при умеренном аортальном стенозе и каждые три-пять лет при легком аортальном стенозе. 19 Поскольку симптомы являются лучшим индикатором гемодинамической значимости, следует регулярно собирать тщательный анамнез пациента. Самое главное, пациенты должны быть осведомлены о симптомах и важности своевременного сообщения о них своим врачам. Медикаментозное лечение во время выжидательной тактикиНе было доказано, что медикаментозное лечение замедляет прогрессирование заболевания аортального клапана или улучшает выживаемость. Однако многие пациенты с бессимптомным аортальным стенозом имеют сопутствующие сердечные заболевания, включая гипертензию, мерцательную аритмию и ИБС; эти условия следует тщательно контролировать. 19,28 ГИПЕРТЕНЗИЯ Приблизительно у 40% пациентов с аортальным стенозом также имеется гипертензия. Препараты, снижающие артериальное давление, следует начинать с низких доз и постепенно титровать до достижения желаемого эффекта. Ингибиторы ангиотензинпревращающего фермента (АПФ) хорошо переносятся, улучшают толерантность к нагрузке и уменьшают одышку у симптоматических пациентов с тяжелым аортальным стенозом. 38 Однако в настоящее время нельзя рекомендовать рутинное использование ингибиторов АПФ у пациентов с бессимптомным аортальным стенозом. ФИБРИЛЛЯЦИЯ ПРЕДСЕРДИЙ Фибрилляция предсердий возникает у 5 процентов пациентов с аортальным стенозом. Его присутствие может затруднить эхокардиографическую оценку аортального стеноза. Впервые возникшая фибрилляция предсердий может спровоцировать сердечную недостаточность у ранее бессимптомного пациента со значительным аортальным стенозом. Контроль сердечного ритма важен, чтобы дать время для оптимального диастолического наполнения. Тем не менее, врачи должны знать о возможных гемодинамических эффектах некоторых препаратов, контролирующих ЧСС, у пациентов с аортальным стенозом. СНИЖЕНИЕ КОРОНАРНОГО РИСКАСопутствующая ИБС часто встречается у пациентов с аортальным стенозом. Рекомендации ACC/AHA рекомендуют оценку и модификацию факторов сердечного риска у таких пациентов. 19 Это включает прекращение употребления табака, начало профилактического приема аспирина у взрослых пациентов с 10-летним риском сердечно-сосудистых заболеваний 6 процентов или выше, а также участие в регулярных физических упражнениях. 40 Пациентам с легким аортальным стенозом не следует ограничивать физическую активность. Бессимптомным пациентам с умеренным или тяжелым аортальным стенозом следует избегать соревновательной или энергичной деятельности, связанной с высокими динамическими и статическими мышечными нагрузками, хотя другие формы упражнений безопасны. 19,41 Хотя первоначальные обсервационные исследования показали значительно более низкую скорость прогрессирования аортального стеноза у пациентов, получавших статины, 42,43 более позднее рандомизированное проспективное исследование не показало статистических различий в гемодинамическом прогрессировании аортального стеноза между пациентами, получавшими агрессивная терапия аторвастатином (Липитор) и получавшие плацебо. Наконец, антимикробная профилактика бактериального эндокардита рекомендуется пациентам, перенесшим замену аортального клапана, но больше не рекомендуется пациентам с аортальным стенозом или другими приобретенными пороками клапана. 46 Презентация аллоантигена и реакция «трансплантат против хозяина»: топливо для огня | КровьПропустить пункт назначения ОБЗОРНАЯ СТАТЬЯ| 16 июня 2016 г. Мотоко Кояма, Джеффри Р. Хилл Кровь (2016) 127 (24): 2963–2970. https://doi.org/10.1182/blood-2016-02-6
История статьи Сдано: 1 февраля 2016 г. Принято: 5 марта 2016 г. Первое издание: 30 марта 2016 г.
Citation Мотоко Кояма, Джеффри Р.
Скачать файл цитирования:
Аллогенная трансплантация стволовых клеток (ТСК) — уникальная процедура, в первую очередь у пациентов со злокачественными новообразованиями кроветворной системы, включающая химиолучевую терапию с последующим введением донорских гемопоэтических и иммунных клеток в воспаленную и лимфопенную среду. Прерывание процесса презентации реципиентного аллоантигена донорским Т-клеткам для возникновения реакции «трансплантат против хозяина» (РТПХ) представляет собой привлекательную терапевтическую стратегию для предотвращения заболеваемости и смертности после ТСК и все больше изучается в последние 15 лет. Темы: Обзорные статьи, Трансплантация Темы: доноров, болезнь «трансплантат против хозяина», т-лимфоциты, трансплантация, антигены Аллогенная трансплантация гемопоэтических стволовых клеток (ТСК) остается важным методом лечения гематологических злокачественных новообразований. Терапевтический эффект «трансплантат против лейкемии» (GVL) носит иммунологический характер и опосредован донорскими Т-клетками и естественными клетками-киллерами, распознающими аллогенные, гемопоэтические или специфические для лейкемии антигены реципиента. 1 Таким образом, GVL связан с патогенными иммунными реакциями на некроветворные ткани, которые проявляются в виде реакции «трансплантат против хозяина» (GVHD), что является основным процедурным ограничением. РТПХ проявляется главным образом в виде острых и хронических заболеваний, характеризующихся апоптозом и фиброзом тканей соответственно. Очевидно, что мишенями РТПХ являются MHC и пептиды, представленные в них (у доноров и реципиентов с несовпадением MHC), и/или минорные антигены гистосовместимости (mHA) как в парах MHC-несовместимых, так и в MHC-совместимых донор-реципиент. Однако было меньше ясности в отношении природы презентации аллоантигенов, ответственной за инициацию иммунных ответов в донорских Т-клетках, которые приводят к РТПХ, и, в частности, задействованных клеток и молекулярных путей. В этом обзоре излагаются последние разработки в этой области, которые улучшают наше понимание презентации аллоантигенов, инициирующих патологический и защитный иммунитет после ТСК, подчеркивая сложный, но в значительной степени модифицируемый процесс. При рассмотрении аллогенной ТСК, при которой донор и реципиент совпадают по MHC, аллоантигены, вызывающие РТПХ, определяются как mHA. В широком смысле эти mHA представляют собой пептиды, генерируемые полиморфными генами, которые различаются между донором и хозяином, которые, в свою очередь, могут быть представлены MHC. Большинство mHA, определенных на сегодняшний день, представлены классом HLA I, 6,7 , преимущественно из-за молекулярных методов, используемых для их идентификации. В последнее время растет энтузиазм по поводу идентификации mHA, ограниченных гемопоэзом, поскольку эти антигены привлекательны для использования в испытаниях иммунотерапии для усиления GVL 9.0011 8 и предотвратить рецидив (рассмотрено в Bleakley et al 9 ). Интересно, что продолжающаяся экспрессия mHA на некроветворных клетках, по-видимому, приводит к истощению аллореактивных Т-клеток и нарушению эффектов GVL. 10,11 Таким образом, идентификация mHA, ограниченных гемопоэзом, крайне желательна, и было идентифицировано несколько таких mHA, включая HA-1, HA-2, LRH-1 и ACC-2 (обзор в Bleakley et al. Распознавание аллоантигена донорскими Т-клетками Истощение донорских Т-клеток приводит к низкой частоте острой и хронической РТПХ, хотя и за счет рецидива лейкемии. 12,13 Аналогично, аутологичная ТСК с Т-клетками связана с высокой частотой рецидивов по сравнению с аллогенной ТСК, что позволяет предположить, что стимуляция Т-клеток аллоантигеном имеет решающее значение как для эффектов РТПХ, так и для эффектов РТПХ. 14 РТПХ может быть концептуализирована как зависимая от MHC класса I и/или MHC класса II (т. е. CD8 + и CD4 + Т-клеток соответственно). Несоответствие HLA класса I и II является значительным фактором риска тяжелой РТПХ и смертности, связанной с трансплантацией 3 ; таким образом, вовлечены как CD8 + Т-клетки, так и CD4 + Т-клетки, и действительно, истощение любой подгруппы Т-клеток недостаточно для предотвращения РТПХ. В MHC-совместимых SCT аллоантигены, представленные в MHC класса I, имеют преимущественно эндогенное происхождение (т. е. антиген присущ клетке), 17 и донорские Т-клетки распознают через Т-клеточный рецептор (TCR), полиморфные пептиды реципиента, представленные в MHC, который является общим как для донора, так и для реципиента (рис. 1). Что касается процесса презентации, собственные или вирусные цитозольные белки процессируются внутри протеосомы, а затем транспортируются в эндоплазматический ретикулум транспортером, связанным с процессингом антигена (TAP). Рисунок 1 Просмотреть в большом формате Загрузить PPT Презентация и распознавание аллоантигенов при трансплантации, совместимой с MHC и несовместимой с MHC. Рисунок 1 Просмотреть в большом формате Загрузить PPT Презентация и распознавание аллоантигенов при трансплантации, совместимой с MHC и несовместимой с MHC. Распознавание пептида в трансплантате, совместимом с MHC (слева). Поскольку донорские Т-клетки отбираются посредством положительного и отрицательного отбора в тимусе донора, TCR донора распознает происходящие от хозяина (не собственные) mHA (зеленый) в MHC (красный), который является общим для донора и хозяина. Близкий модальный В отличие от MHC класса I, аллоантигены, представленные в MHC класса II, преимущественно экзогенного происхождения, и донорские Т-клетки снова распознают через TCR полиморфные реципиентные пептиды, представленные в MHC, которые являются общими для обоих донор и реципиент. Таким образом, аллоантиген может быть представлен в МНС класса II реципиентными или донорскими антигенпрезентирующими клетками (АРС). Экзогенный антиген приобретается АПК посредством фагоцитоза мертвых или некротических клеток, эндоцитоза или макропиноцитоза. Первые 2 процесса опосредованы рецепторами (например, клатрин), и эти пути существуют с разной степенью эффективности во всех профессиональных АПК (т. Недавно также был описан третий «полупрямой» путь презентации антигена. Здесь донорские клетки могут приобретать мембрану клеточной поверхности реципиентной клетки, включая MHC класса I и класса II, посредством либо трогоцитоза, либо поглощения экзосом. 19,21 В этом процессе молекулы MHC, нагруженные аллоантигеном, переносятся из соседних клеток клеточным контактом или зависимым от секреции экзосом образом, что впоследствии может активировать донорские Т-клетки. 22 Сообщается, что этот процесс способствует активации или подавлению Т-клеток 22,23 и, вероятно, важен как средство приобретения антигена и, потенциально, презентации. 19,21 В условиях трансплантации, где присутствуют несоответствия MHC, донорские Т-клетки реагируют на APC реципиента с очень высокой частотой (т. Потребность в АПК хозяина и/или донора для инициации и поддержания РТПХ была предметом значительного изучения. Исторически это называлось прямой или непрямой презентацией антигена. При прямой презентации антигена донорские Т-клетки распознают комплексы MHC-пептид на APC хозяина; при непрямой презентации антигена донорские Т-клетки распознают пептиды реципиента, загруженные в MHC донорских APC. 32,33 Основополагающее исследование, проведенное Shlomchik et al., продемонстрировало, что APC хозяина необходимы и достаточны для инициации летальной острой РТПХ в пределах MHC класса I. 17 Последующие исследования подтвердили то же самое для MHC класса II. 34,35 Однако природа требуемых APC, по-видимому, различается для MHC класса I и II. В MHC класса I гемопоэтические АПК реципиента, по-видимому, являются критическими, 17,36 , хотя негематопоэтические АПК могут играть незначительную роль. 36 В MHC класса II как гемопоэтические, так и негематопоэтические АПК могут инициировать летальную острую РТПХ, а негематопоэтические АПК кажутся доминирующими. Как упоминалось ранее, APC хозяина играют важную роль в индукции РТПХ. Впоследствии во многих исследованиях были предприняты попытки идентифицировать вовлеченные критические подмножества APC, основываясь на представлении о том, что делеция может предотвратить РТПХ. Давняя догма считала, что DC-реципиенты являются APC, ответственными за GVHD, на основании их способности инициировать заболевание при переносе реципиентам с дефицитом MHC 38 и их преходящее сохранение и активация после тотального облучения тела (ЧМТ). 39 В аналогичных исследованиях плазмоцитоидные ДК хозяина также могли инициировать РТПХ у смертельно облученных реципиентов, тогда как В-клетки хозяина не могли. 40 Что касается последствий ЧМТ и миелоаблативного кондиционирования, частота острой РТПХ все еще высока, хотя и с задержкой, после кондиционирования пониженной интенсивности в клинике. Что касается ДК и РТПХ, доклинические исследования установили способность ДК реципиента вызывать РТПХ у реципиентов, в противном случае лишенных всех функциональных АПК, но не их важность у реципиентов, у которых все АПК компетентны. Недавно появилось новое поколение трансгенных мышей, у которых CD11c 9Клетки 0011 + могут быть условно удалены, что перевернуло эту концепцию с ног на голову, продемонстрировав, что реципиентные ДК не требуются для инициации MHC-класса II-зависимой РТПХ и могут фактически регулировать заболевание с помощью индукции апоптоза Т-клеток. 35,43,44 Кроме того, реципиентные плазмацитоидные ДК, В-клетки, макрофаги и клетки Лангерганса также, по-видимому, являются избыточными изолированно для инициации летальной острой РТПХ и способны регулировать заболевание. 35,43,45-48 Пути, участвующие в регуляции этих профессиональных субпопуляций АПК, включают интерлейкин-10 (ИЛ-10) для В-клеток, 49 CD47 и поглощение донорских Т-клеток для макрофагов, 45 и клональная делеция для ДК. Таким образом, негемопоэтические АПК реципиента, активированные с помощью кондиционирования химиолучевой терапией, стали высокоэффективными в инициации MHC класса II-зависимой РТПХ. 35,36 Действительно, TBI индуцирует экспрессию молекул, необходимых для костимуляции (CD40, CD80 и CD86) на некроветворных клетках MHC класса II + . | |||||||
| Цель . | Коммерческие образцы . | Предполагаемые эффекты . | |||||
| Таргетные терапевтические средства | |||||||
| DAMP/PAMP inhibition | RAGE inhibitor | PF-044
| Inhibit donor DC activation and expansion in the GI tract 42 | ||||
| TLR4 inhibitor | Eritoran | Inhibit donor DC activation and expansion in the GI tract 42 | |||||
| Ингибирование хемокинов | Анти-CCR7 Ab | В разработке | Предотвращение миграции ДК донорской ткани, несущих аллоантиген, в лимфатические узлы 42 | ||||
| Anti-α4β7 integrin Ab | Vedolizumab | Prevent migration of donor T cells with integrin signatures imprinted by donor DCs 42 | |||||
| Cytokine inhibition | Anti-IL-12/23p40 Ab | Устекинумаб | Ингибирует размножение Т-клеток и дифференцировку Th2/Th27 в ответ на APC-производный IL-12/23 42 | ||||
| Антитело к рецептору IL-6 или IL-62602 Siltuximab tocilizumab | Ингибируйте расширение Т-клеток и дифференцировку Th27 в ответ на APC-полученное IL-6 42 | ||||||
| Ингибирование костмуляционной молекулы | .|||||||
| CTLA-4-Fc | Абатацепт | Ингибирует CD80/86 и APC-опосредованную костимуляцию 1 9 8 18405 | DC depletion | CD83 Ab | In development | Depletion of activated DCs 98 | | |
| Cell signaling inhibition | Janus kinase inhibitors | Ruxolitinib | Impaired DC activation and migration 1 | ||||
| Currently использованные иммунодепрессанты | Ингибирование кальциневрина | Циклоспорин такролимус | Ингибирование презентации антигена в ДК 100 | ||||
| MTOR Ингибирование | Рапамицин | Продвижение толерогенного DC фенотипа | |||||
| APTRANSPLANT CYCLOPOPHOSPAR | |||||||
| APTRANSPLANT CYCLOPOPHOSPAR | |||||||
| APTRANSPLANTAPLANT. |
| Путь
. | Цель . | Коммерческие образцы . | Предполагаемые эффекты . | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Targeted therapeutic agents | ||||||||||
| DAMP/PAMP inhibition | RAGE inhibitor | PF-044
| Inhibit donor DC activation and expansion in the GI tract 42 | |||||||
| Ингибитор TLR4 | Эриторан | Ингибирует активацию и размножение донорских DC в желудочно-кишечном тракте 42 | ||||||||
| 1845 | Anti-CCR7 AB | В развитии | Предотвращение миграции аллоантигенов доноров ткани в лимфатические узлы 42 | |||||||
| Anti-α4β7 AB | 7 | Anti-α4β7 AB | 7 | Anti-α4β7 AB | 7. Донором DCS 42 | |||||
| Ингибирование цитокинов | Анти-IL-12/23P40 AB | Ustekinumab | Ингибируйте T-Cell. 0011 42 | |||||||
| Anti–IL-6 or IL-6 receptor Ab | Siltuximab Tocilizumab | Inhibit T-cell expansion and Th27 differentiation in response to APC-derived IL-6 42 | ||||||||
| Costimulatory molecule Ингибирование | антагонистический CD40- AB | Lucatumumab | Ингибируйте активацию CD40 и APC 2 | |||||||
| 2 CTLA-41827 | 2 CTLA-4184449 | 2 CTLA-4-FC | 499 | 5 | 2 CTLA-418427 | 2 | ||||
| 2 | ||||||||||
| 2.0011 1 | ||||||||||
| DC depletion | CD83 Ab | In development | Depletion of activated DCs 98 | |||||||
| Cell signaling inhibition | Janus kinase inhibitors | Ruxolitinib | Impaired DC activation and migration 1 | |||||||
| Используемые в настоящее время иммунодепрессанты | Ингибирование кальциневрина | Циклоспорин такролимус | Ингибирование презентации антигена в ДК 100 | |||||||
| MTOR Ингибирование | Рапамицин | Продвижение толерогенного DC фенотипа | ||||||||
| APTRANSPLANT CYCLOPOPHOSPAR | ||||||||||
| APTRANSPLANT CYCLOPOPHOSPAR | ||||||||||
| APTRANSPLANTAPLANT. |
Ат, антитело.
Увеличить
Авторы благодарят Мадлен Флинн из QIMR Berghofer за создание графики. Авторы приносят свои извинения авторам многих важных статей, на которые нельзя было сослаться из-за нехватки места.
Добавил: М.К. и Г.Р.Х. написал рукопись.
Раскрытие информации о конфликте интересов: G.R.H. получил финансирование от Roche для клинических исследований ингибирования IL-6. М.К. заявляет об отсутствии конкурирующих финансовых интересов.
Для переписки: Джеффри Р. Хилл, Лаборатория трансплантации костного мозга, Медицинский научно-исследовательский институт Бергхофера QIMR, 300 Herston Rd, Brisbane, QLD 4006, Australia; электронная почта: [email protected].
1
Блазар
BR
,
Мерфи
WJ
,
Абеди
7
M
Достижения в области биологии и терапии реакции «трансплантат против хозяина».
,
Nat Rev Immunol
,
2012
, vol.
12
6
(стр.
443
—
458
)
2
Anasetti
)0007
C
,
Logan
BR
,
Lee
SJ
, et al.
Сеть клинических испытаний по пересадке крови и костного мозга
Сравнение стволовых клеток периферической крови и костного мозга от неродственных доноров.
,
N Engl J Med
,
2012
, том.
367
16
(стр.
1487
—
1496
)
3
Arora
M
,
Weisdorf
DJ
,
Spellman
SR
, et al.
HLA-идентичный родственный трансплантат по сравнению с 8/8 совместимыми и несовместимыми трансплантатами неродственного донора костного мозга при хронической фазе хронического миелоидного лейкоза.
,
Дж. Клин Онкол
,
2009
, том.
27
10
(pg.
1644
—
1652
)
4
Kekre
N
,
Antin
JH
.
Источники доноров для трансплантации гемопоэтических стволовых клеток в 21 веке: выбор идеального донора, когда идеальной пары не существует.
,
Кровь
,
2014
, том.
124
3
(pg.
334
—
343
)
5
Soiffer
RJ
,
Lerademacher
J
,
и др.
Влияние иммуномодуляции анти-Т-клеточными антителами на исход аллогенной трансплантации гемопоэтических стволовых клеток пониженной интенсивности при гематологических злокачественных новообразованиях.
,
Кровь
,
2011
, том.
117
25
(стр.
6963
—
6970
)
6
Spierings
9000 7000468
.
Малые антигены гистосовместимости: прошлое, настоящее и будущее.
,
Тканевые антигены
,
2014
, vol.
84
4
(стр.
374
—
60
)
7
Petersdorf
EW
.
Главный комплекс гистосовместимости: модель для понимания реакции «трансплантат против хозяина».
,
Кровь
,
2013
, том.
122
11
(стр.
1863
—
1872
)
8
,
Matte-Martone
C
,
Zheng
H
, et.
Т-клетки памяти от доноров, вакцинированных антигеном с минорной гистосовместимостью, и доноров, иммунных к вирусу, улучшают GVL и восстановление иммунитета.
,
Blood
,
2011
9
Bleakley
M
,
Ридделл
SR
.
Использование Т-клеток, специфичных к антигенам минорной гистосовместимости человека, для терапии лейкемии.
,
Immunol Cell Biol
,
2011
, vol.
89
3
(pg.
396
—
407
)
10
Asakura
S
,
Hashimoto
D
,
Такашима
S
и др.
Экспрессия аллоантигена на некроветворных клетках снижает эффекты реакции «трансплантат против лейкемии» у мышей.
,
J Clin Invest
,
2010
, том.
120
7
(стр.
2370
—
2378
)
11
Fontaine
)0868P
,
Roy-Proulx
G
,
Knafo
L
,
Baron
C
,
Roy
DC
,
Perreault
C
.
Адаптивный перенос антиген-специфических Т-лимфоцитов минорной степени гистосовместимости уничтожает лейкозные клетки, не вызывая реакции «трансплантат против хозяина».
,
Nat Med
,
2001
, том.
7
7
(pg.
789
—
794
)
12
Marmont
AM
,
Horowitz
MM
,
Гейл
RP
, и др.
Истощение Т-клеток HLA-идентичных трансплантатов при лейкемии.
,
Кровь
,
1991
, том.
78
8
(pg.
2120
—
2130
)
13
Horowitz
MM
,
Gale
RP
,
Sondel
PM
, и др.
Реакции «трансплантат против лейкоза» после трансплантации костного мозга.
,
Кровь
,
1990
, том.
75
3
(pg.
555
—
562
)
14
Weiden
PL
,
Flournoy
N
,
Томас
ED
и др.
Противолейкемический эффект реакции «трансплантат против хозяина» у людей, реципиентов аллогенных трансплантатов костного мозга.
,
N Engl J Med
,
1979
, том.
300
19
(pg.
1068
—
1073
)
15
Herrera
C
,
Torres
A
,
García-Castellano
JM
, и др.
Профилактика реакции «трансплантат против хозяина» у пациентов с высоким риском путем истощения CD4+ и снижения CD8+ лимфоцитов в трансплантате костного мозга.
,
Пересадка костного мозга
,
1999
, том.
23
5
(pg.
443
—
450
)
16
Nimer
SD
,
Giorgi
J
,
Гаевский
JL
и др.
Селективное истощение клеток CD8+ для профилактики реакции «трансплантат против хозяина» после трансплантации костного мозга. Рандомизированное контролируемое исследование.
,
Трансплантация
,
1994
, том.
57
1
(pg.
82
—
87
)
17
Shlomchik
WD
,
Couzens
MS
,
Тан
CB
и др.
Профилактика реакции «трансплантат против хозяина» путем инактивации антигенпрезентирующих клеток хозяина.
,
Наука
,
1999
, том.
285
5426
(pg.
412
—
415
)
18
Blum
JS
,
Wearsch
PA
,
Крессуэлл
P
.
Пути процессинга антигена.
,
Annu Rev Immunol
,
2013
, том.
31
(pg.
443
—
473
)
19
Wang
X
,
,
Matte-Martone
C
, и другие.
Механизмы презентации антигена Т-лимфоцитам при реакции «трансплантат против хозяина» у мышей: перекрестная презентация и появление перекрестной презентации.
,
Кровь
,
2011
, том.
118
24
(pg.
6426
—
6437
)
20
Hubbard-Lucey
VM
,
Shono
Y
,
Maurer
K
, и др.
Ген аутофагии Atg16L1 предотвращает летальную аллореактивность Т-клеток, опосредованную дендритными клетками.
,
Иммунитет
,
2014
, том.
41
4
(pg.
579
—
591
)
21
Markey
KA
,
Koyama
M
,
Gartlan
KH
и др.
Кроссдрессинг донорскими дендритными клетками после аллогенной трансплантации костного мозга способствует формированию иммунологического синапса и максимизирует ответы на косвенно представленный антиген.
,
J Иммунол
,
2014
, том.
192
11
(стр.
5426
—
5433
)
22
. Nakayama
4 MI
967.
Презентация антигена клетками, покрытыми MHC.
,
Фронт Иммунол
,
2015
, том.
5
стр.
672
23
Dubrot
J
,
Duraes
FV
,
Potin
L
, et al.
Стромальные клетки лимфатических узлов приобретают комплексы пептид-MHCII из дендритных клеток и индуцируют антиген-специфическую толерантность CD4⁺ Т-клеток.
,
J Exp Med
,
2014
, том.
211
6
(pg.
1153
—
1166
)
24
Colf
LA
,
Bankovich
AJ
,
Hanick
NA
, и др.
Как один рецептор Т-клетки распознает как собственный, так и чужеродный MHC.
,
Сотовый
,
2007
, том.
129
1
(pg.
135
—
146
)
25
Speir
JA
,
Garcia
KC
,
Brunmark
A
, и др.
Структурная основа распознавания 2C TCR пептидных комплексов H-2Ld.
,
Иммунитет
,
1998
, том.
8
5
(pg.
553
—
562
)
26
Reiser
JB
,
Darnault
C
,
Гимезан
A
, и др.
Кристаллическая структура Т-клеточного рецептора, связанного с аллогенной молекулой MHC.
,
Нат Иммунол
,
2000
, том.
1
4
(pg.
291
—
297
)
27
Macdonald
WA
,
Chen
Z
,
Gras
S
, и др.
Распознавание Т-клеток посредством молекулярной мимикрии.
,
Иммунитет
,
2009
, том.
31
6
(pg.
897
—
908
)
28
Gras
S
,
Kjer-Nielsen
L
,
Chen
Z
,
Rossjohn
J
,
McCluskey
,
Дж
.
Структурные основы прямого аллоузнавания Т-клеток: последствия отторжения трансплантата, опосредованного Т-клетками.
,
Immunol Cell Biol
,
2011
, vol.
89
3
(pg.
388
—
395
)
29
Petersdorf
EW
,
Gooley
T
,
Малкки
M
,
Горовиц
7 .
Международная рабочая группа по гистосовместимости при трансплантации гемопоэтических клеток
Клиническое значение соответствия HLA донор-реципиент для выживаемости после миелоаблативной трансплантации гемопоэтических клеток от неродственных доноров.
,
Тканевые антигены
,
2007
, том.
69
suppl 1
(pg.
25
—
30
)
30
Weisdorf
D
,
Spellman
S
,
Haagenson
M
, и др.
Классификация соответствия HLA для ретроспективного анализа трансплантации от неродственного донора: пересмотренные определения для прогнозирования выживаемости.
,
Трансплантация костного мозга Биол
,
2008
, том.
14
7
(pg.
748
—
758
)
31
Sprent
J
,
Schaefer
M
,
,
Корнгольд
Р
.
Свойства очищенных субпопуляций Т-клеток. II. In vivo ответы на различия класса I и класса II H-2.
,
J Exp Med
,
1986
, том.
163
4
(pg.
998
—
1011
)
32
Rogers
NJ
,
Lechler
RI
.
Всезнание.
,
Am J Transplant
,
2001
, vol.
1
2
(pg.
97
—
102
)
33
Chakraverty
R
,
Sykes
M
.
Роль антигенпрезентирующих клеток в инициировании реакции «трансплантат против хозяина» и лейкемии «трансплантат против хозяина».
,
Кровь
,
2007
, том.
110
1
(pg.
9
—
17
)
34
Teshima
T
,
Ordemann
R
,
Reddy
P
и др.
Острая реакция «трансплантат против хозяина» не требует экспрессии аллоантигена на эпителии хозяина.
,
Нат Мед
,
2002
, том.
8
6
(pg.
575
—
581
)
35
Koyama
M
,
Kuns
RD
,
Olver
SD
и др.
Реципиентных негемопоэтических антигенпрезентирующих клеток достаточно, чтобы вызвать летальную острую реакцию «трансплантат против хозяина».
,
Nat Med
,
2011
, том.
18
1
(pg.
135
—
142
)
36
Toubai
T
,
Tawara
I
,
и др.
Индукция острой РТПХ несовместимыми по полу антигенами H-Y в отсутствие функциональных радиочувствительных гемопоэтических антигенпрезентирующих клеток хозяина.
,
Кровь
,
2012
, том.
119
16
(pg.
3844
—
3853
)
37
Matte
CC
,
,
Cormier
J
и др.
Донорские АПК необходимы для максимальной РТПХ, но не для ГВЛ.
,
Nat Med
,
2004
, том.
10
9
(pg.
987
—
992
)
38
Duffner
UA
,
Maeda
Y
,
Cooke
KR
, и др.
Одних только дендритных клеток хозяина достаточно, чтобы инициировать острую реакцию «трансплантат против хозяина».
,
J Иммунол
,
2004
, том.
172
12
(pg.
7393
—
7398
)
39
Zhang
Y
,
Louboutin
JP
,
,
Ривера
AJ
,
Эмерсон
СГ
.
Претерминальные дендритные клетки хозяина у облученных мышей инициируют опосредованную CD8+ Т-клетками острую реакцию «трансплантат против хозяина».
,
J Clin Invest
,
2002
, том.
109
10
(pg.
1335
—
1344
)
40
Koyama
M
,
Hashimoto
D
,
Аояма
K
, и др.
Плазмацитоидные дендритные клетки запускают аллореактивные Т-клетки для опосредования реакции «трансплантат против хозяина» в качестве антигенпрезентирующих клеток.
,
Кровь
,
2009
, том.
113
(стр.
2088
—
2095
)
41
Haniffa
)0868M
,
Ginhoux
F
,
Wang
XN
, и др.
Дифференциальные скорости замещения дендритных клеток дермы человека и макрофагов при трансплантации гемопоэтических стволовых клеток.
,
J Exp Med
,
2009
, том.
206
2
(стр.
371
—
385
)
42
Koyama
M
,
9000 9000
8. 8. 8. 8. 67. 8968 67. 67. 8 67. 67. 67. CD103+ дендритные клетки толстой кишки донора определяют тяжесть острой реакции «трансплантат против хозяина».
,
J Exp Med
,
2015
, том.
212
8
(pg.
1303
—
1321
)
43
,
Demetris
AJ
,
McNiff
J
, et al.
Глубокое истощение обычных дендритных клеток хозяина, плазмоцитоидных дендритных клеток и В-клеток не предотвращает индукцию реакции «трансплантат против хозяина».
,
Дж Иммунол
,
2012
, том.
188
8
(pg.
3804
—
3811
)
44
Weber
M
,
Rudolph
B
,
Stein
P
, и др.
CD8⁺ дендритные клетки хозяина защищают от острой реакции «трансплантат против хозяина» после экспериментальной аллогенной трансплантации костного мозга.
,
Трансплантация костного мозга Биол
,
2014
, том.
20
11
(pg.
1696
—
1704
)
45
Hashimoto
D
,
Chow
A
,
Гретер
M
, и др.
Предтрансплантационная терапия CSF-1 увеличивает количество макрофагов реципиента и улучшает РТПХ после аллогенной трансплантации гемопоэтических клеток.
,
J Exp Med
,
2011
, том.
208
5
(pg.
1069
—
1082
)
46
MacDonald
KP
,
Palmer
JS
,
Кронау
S
и др.
Антитело против рецептора колониестимулирующего фактора 1 истощает резидентную субпопуляцию моноцитов и ассоциированных с тканью и опухолью макрофагов, но не ингибирует воспаление.
,
Кровь
,
2010
, том.
116
19
(стр.
3955
—
3963
).
C
и др.
Клетки Лангерганса не требуются для реакции «трансплантат против хозяина».
,
Кровь
,
2011
, том.
117
2
(pg.
697
—
707
)
48
Matte-Martone
C
,
Wang
X
,
Андерсон
B
, и др.
В-клетки-реципиенты не требуются для индукции реакции «трансплантат против хозяина».
,
Трансплантация костного мозга Биол
,
2010
, том.
16
9
(pg.
1222
—
1230
)
49
Rowe
V
,
Banovic
T
,
MacDonald
KP
, и др.
В-клетки-хозяева продуцируют IL-10 после ЧМТ и ослабляют острую РТПХ после аллогенной трансплантации костного мозга.
,
Кровь
,
2006
, том.
108
7
(pg.
2485
—
2492
)
50
Saada
JI
,
Pinchuk
IV
,
Баррера
CA
и др.
Субэпителиальные миофибробласты являются новыми непрофессиональными АПК в слизистой оболочке толстой кишки человека.
,
J Иммунол
,
2006
, том.
177
9
(pg.
5968
—
5979
)
51
Kündig
TM
,
Bachmann
MF
,
DiPaolo
C
и др.
Фибробласты как эффективные антигенпрезентирующие клетки в лимфоидных органах.
,
Наука
,
1995
, том.
268
5215
(pg.
1343
—
1347
)
52
Powell
DW
,
Pinchuk
IV
,
Саада
JI
,
Чен
X
,
Миффлин
RC
.
Мезенхимальные клетки собственной пластинки кишечника.
,
Annu Rev Physiol
,
2011
, том.
73
(pg.
213
—
237
)
53
Hershberg
RM
,
Framson
PE
,
Cho
DH
и др.
Эпителиальные клетки кишечника используют два различных пути процессинга антигена HLA класса II.
,
J Clin Invest
,
1997
, том.
100
1
(pg.
204
—
215
)
54
Hershberg
RM
,
Cho
DH
,
Юаким
A
, и др.
Процессинг и презентация высокополяризованного антигена HLA класса II эпителиальными клетками кишечника человека.
,
J Clin Invest
,
1998
, том.
102
4
(стр.
792
—
803
)
55
ОБС0007
C
,
Burich
A
,
Kleijmeer
MJ
,
Griffith
JM
,
Wong
P
,
Руденский
АУ
.
Катепсин S контролирует презентацию антигена, опосредованную МНС класса II, эпителиальными клетками in vivo.
,
Дж Иммунол
,
2005
, том.
174
3
(pg.
1205
—
1212
)
56
Jones
SC
,
Murphy
GF
,
Фридман
ТМ
,
Korngold
R
.
Важность незначительной экспрессии антигена гистосовместимости негематопоэтическими тканями в модели CD4+ Т-клеточной болезни «трансплантат против хозяина».
,
J Clin Invest
,
2003
, том.
112
12
(pg.
1880
—
1886
)
57
Stevanovic
S
,
van Bergen
CA
,
ван Люксембург-Хейс
SA
, и др.
Активация HLA класса II во время вирусной инфекции приводит к HLA-DP-направленной реакции «трансплантат против хозяина» после инфузии CD4+ донорских лимфоцитов.
,
Кровь
,
2013
, том.
122
11
(pg.
1963
—
1973
)
58
Petersdorf
EW
,
Malkki
M
,
O’hUigin
C
, и др.
Высокая экспрессия HLA-DP и реакция «трансплантат против хозяина».
,
N Engl J Med
,
2015
, vol.
373
7
(pg.
599
—
609
)
59
Hill
GR
,
Ferrara
JLM
.
Приоритет желудочно-кишечного тракта как органа-мишени при острой реакции «трансплантат против хозяина»: обоснование использования цитокиновых щитов при аллогенной трансплантации костного мозга.
,
Кровь
,
2000
, том.
95
9
(pg.
2754
—
2759
)
60
Jenq
RR
,
Ubeda
C
,
Телец
Y
, и др.
Регуляция микробиотой воспаления кишечника после аллогенной трансплантации костного мозга.
,
J Exp Med
,
2012
, том.
209
5
(pg.
903
—
911
)
61
Storb
R
,
Prentice
RL
,
Бакнер
CD
и др.
Реакция «трансплантат против хозяина» и выживаемость у пациентов с апластической анемией, получавших трансплантацию костного мозга от HLA-идентичных братьев и сестер. Благотворное влияние защитной среды.
,
N Engl J Med
,
1983
, том.
308
6
(стр.
302
—
307
)
.
Müller
KD
,
Hirche
H
,
Шефер
UW
.
Влияние кишечной бактериальной деконтаминации с использованием метронидазола и ципрофлоксацина или только ципрофлоксацина на развитие острой реакции «трансплантат против хозяина» после трансплантации костного мозга у пациентов с гемобластозами: окончательные результаты и долгосрочное наблюдение открытого проспективного рандомизированного исследования пробный.
,
Кровь
,
1999
, том.
93
10
(pg.
3267
—
3275
)
63
Jenq
RR
,
Taur
Y
,
Девлин
SM
и др.
Кишечная блаутия связана с уменьшением смертности от реакции «трансплантат против хозяина».
,
Трансплантация костного мозга крови
,
2015
, том.
21
8
(стр.
1373
—
1383
)
64
978 7 78 7 76 78 7 77978 7 7 79767 67.
Като
K
и др.
α-маннан индуцирует Th27-опосредованную болезнь легких «трансплантат против хозяина» у мышей.
,
Кровь
,
2015
, том.
125
19
(pg.
3014
—
3023
)
65
,
Matte-Martone
C
,
Tan
HS
и др.
Болезнь «трансплантат против хозяина» не зависит от врожденных сигнальных путей, запускаемых патогенами в гемопоэтических клетках хозяина.
,
J Иммунол
,
2011
, том.
186
1
(pg.
230
—
241
)
66
Cooke
KR
,
Hill
GR
,
Кроуфорд
JM
и др.
Продукция TNFa при стимуляции ЛПС донорскими клетками предсказывает тяжесть экспериментальной острой реакции «трансплантат против хозяина».
,
J Clin Invest
,
1998
, том.
102
(pg.
1882
—
1891
)
67
Saliba
RM
,
de Lima
M
,
Giralt
S
и др.
Сверхострая РТПХ: факторы риска, исходы и клинические проявления.
,
Кровь
,
2007
, том.
109
7
(pg.
2751
—
2758
)
68
Lee
DW
,
Gardner
R
,
Porter
DL
, и др.
Современные концепции диагностики и лечения синдрома высвобождения цитокинов.
,
Кровь
,
2014
, том.
124
2
(pg.
188
—
195
)
69
Kennedy
GA
,
Varelias
A
,
Вукович
S
и др.
Добавление ингибирования интерлейкина-6 с помощью тоцилизумаба к стандартной профилактике реакции «трансплантат против хозяина» после аллогенной трансплантации стволовых клеток: исследование фазы 1/2.
,
Ланцет Онкол
,
2014
, том.
15
13
(pg.
1451
—
1459
)
70
Filipovich
AH
,
Weisdorf
D
,
Павлетич
S
, и др.
Проект разработки консенсуса Национального института здравоохранения по критериям клинических испытаний хронической реакции «трансплантат против хозяина»: I. Отчет рабочей группы по диагностике и стадированию.
,
Трансплантация костного мозга Биол
,
2005
, том.
11
12
(pg.
945
—
956
)
71
Reddy
P
,
Negrin
R
,
Холм
GR
.
Мышиные модели трансплантации костного мозга.
,
Трансплантация костного мозга Биол
,
2008
, том.
14
1 suppl 1
(pg.
129
—
135
)
72
Sakoda
Y
,
Hashimoto
D
,
Асакура
S
, и др.
Зависимые от тимуса Т-клетки донорского происхождения вызывают хроническую реакцию «трансплантат против хозяина».
,
Кровь
,
2007
, том.
109
4
(pg.
1756
—
1764
)
73
Dudakov
JA
,
Hanash
AM
,
Jenq
RR
, и др.
Интерлейкин-22 стимулирует эндогенную регенерацию тимуса у мышей.
,
Наука
,
2012
, том.
336
6077
(pg.
91
—
95
)
74
Matsuoka
K
,
Kim
HT
,
McDonough
S
и др.
Изменение регуляторного гомеостаза Т-клеток у пациентов с лимфопенией CD4+ после аллогенной трансплантации гемопоэтических стволовых клеток.
,
J Clin Invest
,
2010
, том.
120
5
(pg.
1479
—
1493
)
75
Teshima
T
,
Reddy
P
,
,
Уильямс
D
,
Кук
KR
,
Феррара
JL
.
Нарушенная негативная селекция тимуса вызывает аутоиммунную реакцию «трансплантат против хозяина».
,
Кровь
,
2003
, том.
102
2
(стр.
429
—
435
)
76
Palmer
7 .
Негативная селекция — удаление плохих яблок из репертуара Т-клеток.
,
Nat Rev Immunol
,
2003
, том.
3
5
(pg.
383
—
391
)
77
Williams
JA
,
Sharrow
SO
,
Адамс
AJ
,
Ходс
RJ
.
Лиганд CD40 функционирует внеклеточно автономно, способствуя удалению аутореактивных тимоцитов.
,
Дж Иммунол
,
2002
, том.
168
6
(pg.
2759
—
2765
)
78
Anderson
BE
,
McNiff
JM
,
Джейн
D
,
Blazar
BR
,
SHLOMCHIK
WD
,
9798 98 98 98 98 98 98 98 98 98 98 98 967 967 967 967 98 967 967 98 967 98 967 98 967 98 967 968 968 968 968 968 968 . Различная роль антигенпрезентирующих клеток донора и хозяина и костимулирующих молекул при хронической реакции «трансплантат против хозяина» у мышей: требования зависят от органа-мишени.
,
Кровь
,
2005
, том.
105
5
(pg.
2227
—
2234
)
79
Coghill
JM
,
Fowler
KA
,
Запад
ML
и др.
CC Хемокиновый рецептор 8 повышает выживаемость донорских Treg и имеет решающее значение для предотвращения мышиной реакции «трансплантат против хозяина».
,
Кровь
,
2013
, том.
122
5
(pg.
825
—
836
)
80
Matsuoka
K
,
Koreth
J
,
Ким
HT
и др.
Терапия низкими дозами интерлейкина-2 восстанавливает регуляторный гомеостаз Т-клеток у пациентов с хронической реакцией «трансплантат против хозяина».
,
Sci Transl Med
,
2013
, том.
5
179
стр.
179Ra43
81
Корет
J
,
Matsuoka
K
,
76768 967 77977.
Интерлейкин-2 и регуляторные Т-клетки при реакции «трансплантат против хозяина».
,
N Engl J Med
,
2011
, vol.
365
22
(pg.
2055
—
2066
)
82
Gavin
MA
,
Clarke
SR
,
Негру
E
,
Гальегос
A
,
Руденский
А
.
Гомеостаз и анергия CD4(+)CD25(+) Т-клеток-супрессоров in vivo.
,
Нат Иммунол
,
2002
, том.
3
1
(pg.
33
—
41
)
83
Suffner
J
,
Hochweller
K
,
Kühnle
MC
, и др.
Дендритные клетки поддерживают гомеостатическую экспансию регуляторных Т-клеток Foxp3+ у мышей Foxp3.LuciDTR.
,
Дж Иммунол
,
2010
, том.
184
4
(стр.
1810
—
1820
)
84
Shimabukuro -Vonhagen
0007
A
,
Hallek
MJ
,
Storb
RF
,
von Bergwelt-Baildon
MS
.
Роль В-клеток в патогенезе реакции «трансплантат против хозяина».
,
Кровь
,
2009
, том.
114
24
(стр.
4919
—
4927
)
85
Zhang
C
,
Todorov
I
,
Zhang
Z
, et al.
Донорские CD4+ Т- и В-клетки в трансплантатах вызывают хроническую реакцию «трансплантат против хозяина» с аутоиммунными проявлениями.
,
Кровь
,
2006
, том.
107
7
(pg.
2993
—
3001
)
86
Alexander
KA
,
Flynn
R
,
Lineburg
KE
, и др.
CSF-1-зависимые макрофаги донорского происхождения опосредуют хроническую реакцию «трансплантат против хозяина».
,
Дж. Клин Инвест
,
2014
, том.
124
10
(pg.
4266
—
4280
)
87
Srinivasan
M
,
Flynn
R
,
Цена
A
, и др.
Отложение аллоантител донорских В-клеток и формирование зародышевого центра необходимы для развития у мышей хронической РТПХ и облитерирующего бронхиолита.
,
Кровь
,
2012
, том.
119
6
(pg.
1570
—
1580
)
88
Markey
KA
,
Koyama
M
,
Кунс
RD
и др.
Иммунная недостаточность при РТПХ обусловлена дефектной презентацией антигена в субпопуляциях дендритных клеток.
,
Кровь
,
2012
, том.
119
24
(pg.
5918
—
5930
)
89
Wikstrom
ME
,
Fleming
P
,
Кунс
RD
и др.
Острая РТПХ приводит к тяжелому дефекту ДК, который предотвращает примирование Т-клеток и приводит к молниеносной цитомегаловирусной болезни у мышей.
,
Кровь
,
2015
, том.
126
12
(pg.
1503
—
1514
)
90
Elmaagacli
AH
,
Steckel
NK
,
Koldehoff
M
, и др.
Ранняя репликация цитомегаловируса человека после трансплантации связана со снижением риска рецидива: доказательства предполагаемого эффекта вируса против лейкемии у пациентов с острым миелоидным лейкозом.
,
Кровь
,
2011
, том.
118
5
(pg.
1402
—
1412
)
91
Hermouet
S
,
Sutton
CA
,
Роза
ТМ
, и др.
Качественный и количественный анализ вирусов герпеса человека при хроническом и остром В-клеточном лимфолейкозе и множественной миеломе.
,
Лейкемия
,
2003
, том.
17
1
(pg.
185
—
195
)
92
Reddy
P
,
Maeda
Y
,
,
Крияновски
ОИ
,
Корнгольд
Р
,
Феррара
JL
.
Важнейшая роль антигенпрезентирующих клеток и экспрессии аллоантигенов в реакции «трансплантат против лейкемии».
,
Nat Med
,
2005
, том.
11
11
(стр.
1244
—
1249
)
93
Toubai
9000
Toubai
9000
0868 ,
,
Luker
G
, и др.
CD8+ дендритные клетки хозяина необходимы для индукции оптимального ответа «трансплантат против опухоли» после экспериментальной аллогенной трансплантации костного мозга.
,
Кровь
,
2013
, том.
121
20
(стр.
4231
—
4241
7
) 84
MATTE-Martone
C
,
,
Jain
D
,
9000 8
8 8 8 8 8 8 8 8 8 8 8 8 8 98 98 98 98 98 98 98 98 98 98 98 98 8. CD8+, но не CD4+ T-клетки требуют родственных взаимодействий с тканями-мишенями для опосредования GVHD только через минорные H-антигены, тогда как CD4+ и CD8+ T-клетки требуют прямого лейкемического контакта для опосредования GVL.
,
Кровь
,
2008
, том.
111
7
(стр.
3884
—
3892
).
,
Желто-коричневый
HS
, и др.
Трансплантат против лейкемии (GVL) против хронического миелогенного лейкоза мыши с бластным кризисом (BC-CML) и хронической фазы хронического миелогенного лейкоза (CP-CML): общие механизмы уничтожения Т-клеток, но лиганды запрограммированной смерти вызывают CP- CML и не BC-CML устойчивы к GVL.
,
J Иммунол
,
2011
, том.
187
4
(pg.
1653
—
1663
)
96
Rezvani
K
,
Yong
AS
,
Савани
БН
и др.
Эффекты реакции «трансплантат против лейкемии», связанные с обнаружением специфичных для опухоли Вильмса-1 Т-лимфоцитов после аллогенной трансплантации стволовых клеток при остром лимфобластном лейкозе.
,
Кровь
,
2007
, том.
110
6
(pg.
1924
—
1932
)
97
Molldrem
JJ
,
Lee
PP
,
Ван
C
, и др.
Доказательства того, что специфические Т-лимфоциты могут участвовать в элиминации хронического миелогенного лейкоза.
,
Nat Med
,
2000
, том.
6
9
(pg.
1018
—
1023
)
98
Seldon
TA
,
Pryor
R
,
Палькова
А
, и др.
Иммунодепрессивное человеческое моноклональное антитело против CD83 истощение активированных дендритных клеток при трансплантации.
,
Leukemia
,
2016
99
Heidegger
S
,
van den Brink
MR
,
Haas
T
,
Поек
H
.
Роль рецепторов распознавания образов в реакции «трансплантат против хозяина» и «трансплантат против лейкемии» после аллогенной трансплантации стволовых клеток.
,
Фронт Иммунол
,
2014
, том.
5
стр.
337
100
Stenger
EO
,
Turnquist
HR
,
Mapara
MY
,
Thomson
AW
.
Дендритные клетки и регуляция реакции «трансплантат против хозяина» и активности «трансплантат против лейкемии».
,
Кровь
,
2012
, том.
119
22
(стр.
5088
—
5103
) Подпишитесь в учреждении
(Уровень 1, 2, 3, 4)
Уровни центра обработки данных являются решающим фактором при выборе объекта для размещения ваших данных. Рейтинг уровня показывает, что может предложить центр обработки данных с точки зрения надежности и производительности, поэтому неспособность определить правильный уровень может привести к простоям или ненужным расходам.
В этой статье объясняется, как работают уровни центра обработки данных и как эти рейтинги позволяют оценить качество центра обработки данных. Читайте дальше, чтобы узнать, как использовать стандарт многоуровневого обслуживания, чтобы найти объект, соответствующий потребностям и бюджету вашего бизнеса.
Что такое уровни центра обработки данных?
Уровни центра обработки данных — это стандартизированная система ранжирования, которая указывает надежность инфраструктуры центра обработки данных. Эта классификация ранжирует объекты от 1 до 4, где 1 — худший уровень, а 4 — лучший уровень.
Центр обработки данных получает этот международный рейтинг от Uptime Institute, независимой организации, которая определяет уровень объекта в первую очередь на основе:
- Гарантия безотказной работы.
- Отказоустойчивость (способность обрабатывать как запланированные, так и незапланированные сбои).
- Стоимость услуги.
Эта беспристрастная многоуровневая система дает объективное представление о том, как работает конкретный центр обработки данных. Однако иметь рейтинг необязательно, поэтому не всем центрам обработки данных назначен уровень. Однако большинство крупных объектов предпочитают запрашивать оценку у Uptime Institute, поскольку официальный рейтинг помогает:
- Повысьте доверие к сервису.
- Продавайте возможности объекта.
- Завоевать доверие и привлечь потенциальных клиентов.
- План будущих обновлений и расширений объекта.
Сотрудникам центров обработки данных необходимо отправить планы и чертежи объекта ( Уровень сертификации проектной документации ) в Uptime Institute для получения официального рейтинга. Затем представители Uptime Institute посещают центр на месте, чтобы проверить операции и присвоить рейтинг.
Определение уровня центра обработки данных, необходимого вашему бизнесу, — это только первый шаг к поиску подходящего партнера. Наша статья о выборе центра обработки данных поможет вам в остальной части процесса.
Какие факторы учитываются при классификации уровней центра обработки данных
Uptime Institute держит в секрете точный метод присвоения уровней, но большинство ключевых показателей общеизвестны. При оценке объекта Институт учитывает следующие факторы:
- Гарантия доступности и времени безотказной работы сервиса.
- Уровни резервирования (процесс дублирования критически важных компонентов и их хранения в качестве резервных и отказоустойчивых на случай запланированных или незапланированных сбоев).
- Состояние холодильной и энергетической инфраструктуры.
- Квалификация персонала и протоколы обслуживания (в частности, способность обеспечивать параллельную ремонтопригодность).
- Стоимость услуги.
- Операционная устойчивость и способность центра достигать долгосрочных бизнес-целей.
- Время, необходимое учреждению для настройки нового клиента.
- Уровни безопасности центра обработки данных.
- Нейтралитет авианосца.
Многоуровневая система не требует использования каких-либо конкретных технологий или вариантов дизайна . Свобода выбора между решениями обеспечивает гибкость при достижении целей уровня, поэтому каждый центр обработки данных может выбрать наилучший способ соответствия стандартам и достижения желаемого рейтинга.
Объяснение уровней центров обработки данных
Четыре уровня центра обработки данных, сертифицированные Uptime Institute:
-
Уровень 1: Центр обработки данных с единым каналом питания и охлаждения и без резервных компонентов. Ожидаемое время безотказной работы этого уровня составляет 99,671% в год.
- Уровень 2: Центр обработки данных с единым каналом питания и охлаждения, а также некоторыми резервными и резервными компонентами. Этот уровень предлагает ожидаемое время безотказной работы 99,741% в год.
- Уровень 3: Центр обработки данных с несколькими путями питания и охлаждения, а также резервными системами, которые позволяют персоналу работать над установкой, не отключая ее от сети. Этот уровень имеет ожидаемое время безотказной работы 99,982% в год.
- Уровень 4: Полностью отказоустойчивый центр обработки данных с резервированием каждого компонента. Ожидаемое время безотказной работы этого уровня составляет 99,995% в год.
Четыре уровня центра обработки данных являются прогрессивными. Центры обработки данных могут перемещаться вверх и вниз по рейтингу, и каждый уровень включает в себя требования более низкого рейтинга.
Хотя надежность повышается с повышением уровня, уровень 4 не всегда лучше, чем центр обработки данных с более низким рейтингом. Каждый уровень соответствует различным потребностям бизнеса, поэтому уровни 3 или 4 (самые дорогие варианты) часто являются чрезмерными инвестициями.
Сравнение уровней центра обработки данных
В таблице ниже представлен общий обзор четырех уровней центра обработки данных и показано, что различные модели предоставляют клиентам.
| ПАРАМЕТРЫ | УРОВЕНЬ 1 | УРОВЕНЬ 2 | УРОВЕНЬ 3 | УРОВЕНЬ 4 |
| Гарантия безотказной работы | 99,671% | 99,741% | 99,982% | г.99,995% |
| Время простоя в год | <28,8 часов | <22 часов | <1,6 часа | <26,3 минуты |
| Резервирование компонентов | Нет | Частичное резервирование питания и охлаждения (частичное N+1) | Полный N+1 | Отказоустойчивый (2N или 2N+1) |
| Одновременно обслуживаемый | № | № | Частично | Да |
| Цена | $ | $$ | $$$ | $$$$ |
| Компартментализация | № | № | № | Да |
| Кадровое обеспечение | Нет | 1 смена | 1+ смена | 24/7/365 |
| Типичный покупатель | Небольшие компании и стартапы с простыми требованиями | малого и среднего бизнеса | Растущий и крупный бизнес | Государственные учреждения и крупные предприятия |
| Основная причина, по которой компании выбирают этот уровень | Самый доступный уровень ЦОД | Хорошее соотношение цены и качества | Тонкая грань между высокой производительностью и доступностью | Отказоустойчивое средство, идеально подходящее для стабильно высоких уровней трафика или требований к обработке | г.
У вас уже есть собственное оборудование для хостинга? Совместный хостинг позволяет вам размещать свои серверы в стороннем центре обработки данных и пользоваться всеми преимуществами производительности и резервирования лучшего объекта.
Центр обработки данных уровня 1
Инфраструктура уровня 1 обеспечивает питание и охлаждение для поддержки полной ИТ-нагрузки. Эти объекты имеют единый путь для питания и охлаждения, и нет резервирования для какой-либо критической системы. Персонал должен полностью прекратить работу для регулярного технического обслуживания или аварийного ремонта.
Требования к объекту уровня 1:
- Источник бесперебойного питания (ИБП) на случай скачков напряжения и перебоев в подаче электроэнергии.
- Специальное место для ИТ-систем.
- Двигатель-генератор.
- Специальное охлаждающее оборудование, работающее в нерабочее время.
Для центров обработки данных уровня 1 также требуются системы, протоколы и оборудование, обеспечивающие работоспособность центра обработки данных в нерабочее время (ночью и в выходные дни).
Из-за отсутствия резервных копий клиенты центра обработки данных уровня 1 могут рассчитывать на до 28,8 часов простоя в год .
Центр обработки данных уровня 2
Инфраструктура уровня 2 обладает всеми функциями центра обработки данных уровня 1, но с дополнительными возможностями резервного копирования. Эти центры обработки данных обеспечивают лучшую защиту от сбоев благодаря:
- Дополнительным двигателям-генераторам.
- Аккумулятор энергии.
- Чиллеры.
- Фальшполы. Модули ИБП
- .
- Насосы.
- Оборудование для отвода тепла.
- Топливные баки и ячейки.
- Дополнительные холодильные агрегаты.
Как и центры уровня 1, центры уровня 2 полагаются на единый путь распределения питания и охлаждения, поэтому эти объекты по-прежнему уязвимы для неожиданных сбоев. Время безотказной работы лучше, чем в центре обработки данных с более низким рейтингом, поэтому клиенты уровня 2 испытывают до 22 часов простоя в год .
Центр обработки данных уровня 3
Центр обработки данных уровня 3 — это одновременно обслуживаемый объект с несколькими путями распределения питания и охлаждения. В отличие от центров обработки данных уровня 1 и 2, объект уровня 3 не требует полной остановки во время технического обслуживания или замены оборудования.
Для объекта уровня 3 требуются все компоненты, присутствующие в центре обработки данных уровня 2, но эти объекты также должны иметь доступность N+1 :
- «N» относится к необходимой мощности для поддержки полной ИТ-нагрузки.
- «+1» означает дополнительный компонент для целей резервного копирования.
Резервирование по схеме N+1 гарантирует, что дополнительный компонент начнет работать, если основной элемент выйдет из строя или персонал уберет компонент для планового обслуживания.
Для центров обработки данных уровня 3 также требуется решение для резервного копирования, которое может поддерживать работу в случае локального или регионального отключения электроэнергии. Предприятие должно гарантировать, что оборудование может продолжать работать в течение не менее 72 часов после отключения.
Установки уровня 3 имеют значительный скачок в доступности по сравнению с более низкими рейтингами. Клиенты, использующие центры обработки данных уровня 3, испытывают до 1,6 часов простоя в год .
Центр обработки данных уровня 4
Центры обработки данных уровня 4 добавляют механизмы отказоустойчивости к списку требований уровня 3. Эти центры обработки данных имеют несколько физически изолированных систем, которые действуют как резервные компоненты и пути распространения. Помимо всех условий уровня 3, объект уровня 4 должен обеспечивать:
- Все компоненты поддерживают два генератора, две системы ИБП и две системы охлаждения.
- Каждый путь распространения является независимым, поэтому единичный сбой в одном из них не вызывает эффект домино для других компонентов.
- Операции продолжаются в течение как минимум 96 часов после локального или регионального отключения электроэнергии.
- Источник питания не подключен ни к какому внешнему источнику.
Разделение между резервными компонентами жизненно важно для центра обработки данных уровня 4. Физическое разделение не позволяет локальному событию поставить под угрозу обе системы.
Центры обработки данных уровня 4 имеют резервирование 2N или 2N+1 :
- Резервирование 2N (или N+N) означает, что объект имеет полностью зеркальную независимую систему в режиме ожидания. Если что-то случится с основным компонентом, заработает идентичная резервная копия, чтобы обеспечить непрерывную работу.
- Модель 2N+1 обеспечивает вдвое большую оперативную емкость (2N) и дополнительный резервный компонент (+1) на случай сбоя, когда резервная система активна.
Учреждение уровня 4 гарантирует, что клиенты не будут иметь простоев более 26,3 минут в год . Причина, по которой соглашения об уровне обслуживания (SLA) уровня 4 не гарантируют 100% времени безотказной работы, заключается в небольшой вероятности того, что компонент может столкнуться с проблемой во время обслуживания его дублирующих аналогов.
Ищете более гибкий вариант хостинга? Виртуальный частный центр обработки данных обеспечивает преимущества локального центра обработки данных без высоких первоначальных затрат на масштабируемость. Узнайте больше о программно-определяемых настройках в нашем введении в виртуальные центры обработки данных (VDC).
Какой уровень центра обработки данных вам подходит?
Хотя более высокие уровни предлагают более надежный сервис, выбор уровня 3 или 4 не всегда является правильным выбором. Лица, принимающие решения, должны выбрать уровень центра обработки данных, который наилучшим образом соответствует потребностям их бизнеса.
Определенные типы компаний обычно тяготеют к определенным уровням. Вот разбивка обычных клиентов каждого уровня:
-
Уровень 1: Эти центры обработки данных лучше всего подходят для малых предприятий и стартапов, которые ищут наиболее доступный вариант хостинга. Небольшие фирмы без сложных ИТ-требований или круглосуточной работы могут терпеть более частые простои.
- Уровень 2: Эти объекты являются оптимальным вариантом для малого и среднего бизнеса, которым нужен экономичный и более надежный вариант, чем партнер уровня 1. Малые и средние фирмы обычно используют средства уровня 2, часто для размещения резервных копий данных или баз данных, не являющихся критически важными.
- Уровень 3: Эти типы центров обработки данных идеально подходят для крупных компаний с ИТ-операциями, которым требуется дополнительная отказоустойчивость. Предприятия, которые размещают обширные наборы данных (особенно данные о клиентах), являются первыми кандидатами на этот уровень.
-
Уровень 4: Эти центры обработки данных подходят предприятиям без бюджетных ограничений, которым требуется непрерывная доступность. Государственные организации и крупные предприятия с критически важными серверами и интенсивными требованиями клиентов или бизнеса являются типичными пользователями объекта уровня 4.
Обычно при выборе уровня учитываются два основных фактора: стоимость и время безотказной работы . Платить за центр обработки данных уровня 3, в то время как более дешевое оборудование справится с этой задачей, — пустая трата денег. Аналогичным образом установка на объекте уровня 2, когда вам требуется более длительное время безотказной работы, может повлиять на ваш доход, производительность, удовлетворенность клиентов и репутацию.
Bare Metal Cloud от PhoenixNAP — это простой в развертывании выделенный сервер с длительным временем безотказной работы и моделью оплаты по мере использования, которая гарантирует, что вы никогда не переплачиваете за свои потребности в хостинге.
Выберите правильный уровень и избегайте проблем с простоями (или ненужной переплатой)
Важно, где вы храните свои данные, поэтому очень важно знать различия между объектами, на которых хранится ваша информация. Используйте эту статью, чтобы ознакомиться с различными вариантами на рынке и принять обоснованное решение, которое гарантирует, что ваш бизнес не переплатит и не столкнется с проблемами простоя.