
День толерантности презентация 4 класс: Презентация Классный час 4 класс на тему: «Международный день толерантности»
Презентация Толерантность для учащихся 4 класса доклад, проект
- Главная
- Разное
- Образование
- Спорт
- Естествознание
- Природоведение
- Религиоведение
- Французский язык
- Черчение
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Геометрия
- Детские презентации
- Информатика
- История
- Литература
- Математика
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, фоны, картинки для презентаций
- Экономика
Презентация на тему Презентация Толерантность для учащихся 4 класса, предмет презентации: Детские презентации. Этот материал в формате pptx (PowerPoint) содержит 13 слайдов, для просмотра воспользуйтесь проигрывателем. Презентацию на заданную тему можно скачать внизу страницы, поделившись ссылкой в социальных сетях! Презентации взяты из открытого доступа или загружены их авторами, администрация сайта не отвечает за достоверность информации в них, все права принадлежат авторам презентаций и могут быть удалены по их требованию.
Этот материал в формате pptx (PowerPoint) содержит 13 слайдов, для просмотра воспользуйтесь проигрывателем. Презентацию на заданную тему можно скачать внизу страницы, поделившись ссылкой в социальных сетях! Презентации взяты из открытого доступа или загружены их авторами, администрация сайта не отвечает за достоверность информации в них, все права принадлежат авторам презентаций и могут быть удалены по их требованию.
Что такое толерантность?
для учащихся 4 класса.
16 ноября – Международный день толерантности.
Сегодня мы поговорим, что такое
ТОЛЕРАНТНОСТЬ.
Посмотрите на картинку: вороны черные и только одна белая, но им хорошо вместе.
Они весело танцуют и поют, и совсем неважно кто и какого цвета.
Толерантность – это способность признавать отличные от своего собственного мнения. Мы допускаем, что кто-то может думать иначе, или действовать иначе, нежели ты сам.
Мы допускаем, что кто-то может думать иначе, или действовать иначе, нежели ты сам.
Толерантность – это уважение, принятие и понимание многообразия мира.
Мы разные, но мы — вместе!
Толерантность
– это благосклонность и уважение к другому.
Мы все разные: кто-то любит читать, кто-то заниматься спортом.
Кто-то играет с собачкой, а кто-то разводит цветы, но нам хорошо вместе и мы приглашаем тебя.
Ты нам нужен!
Сотрудничество
Прощение
Цветок толерантности
Люди на свет рождаются разными: непохожими, своеобразными.
Чтобы других ты смог понимать, нужно терпенье в себе воспитать.
Вы поняли, что такое толерантность? Давайте поиграем и проверим: «Толерантны ли вы?»
Ситуации:
1. Какой вы дома? Например: Младший братишка сломал твою игрушку:
Какой вы дома? Например: Младший братишка сломал твою игрушку:
— Ты его прощаешь, он сделал это ненарочно…
— Ты ударишь его… Как вы поступите?
2. Ты поссорился со своей сестрой…
— Ты пытаешься объясниться с ней…
3.Как вы поступите в такой ситуации:
— Ты не согласен с кем-нибудь..
— Ты все-таки слушаешь его…
— Ты не даешь ему говорить…
Как вы поступите?
4.В классе ты уже ответил…
— Тебе хочется ответить еще…
— Ты предоставишь другим возможность ответить….
Как вы поступите?
Наша земля – это место, где мы можем любить друг друга, соблюдать традиции и продолжать историю.
Уважайте друг друга, будьте толерантны…
Будете толерантны.
Скачать презентацию
Это сайт презентаций, где можно хранить и обмениваться своими презентациями, докладами, проектами, шаблонами в формате PowerPoint с другими пользователями. Мы помогаем школьникам, студентам, учителям, преподавателям хранить и обмениваться учебными материалами.
Для правообладателей
Обратная связь
Email: Нажмите что бы посмотреть
Внеклассное мероприятие для младших школьников (3-4 классы) День толерантности презентация, доклад
ВНЕКЛАССНОЕ МЕРОПРИЯТИЕ
ДЛЯ МЛАДШИХ ШКОЛЬНИКОВ (3-4 КЛАССЫ)
«ДЕНЬ ТОЛЕРАНТНОСТИ»
Выполнил: студент 3 курса
направления «Педагогическое образование»
Егорова К.В.
Цель мероприятия по поликультурному воспитанию:
знакомство детей с понятием «толерантность, толерантное отношение»;
воспитание уважения и доброго отношения к другим людям;
формирование человека, способного к эффективной жизнедеятельности в многонациональной и поликультурной среде, обладающего обостренным чувством понимания и уважения других культур, умениями жить в мире и согласии с людьми разных национальностей, рас, верований.
Задачи поликультурного воспитания:
— освоение культуры собственного народа;
— воспитание положительного отношения к культурным различиям;
— воспитание в духе мира и сотрудничества;
— развитие поведенческих навыков общения с представителями иных культур и этносов.
План мероприятия
Введение (5 мин.).
«Ребята, сегодня у нас «День толерантности». Давайте разберемся, что это такое – толерантность?»
это – принятие иного мировоззрения, образа жизни, поведения и обычаев.
«Какими чертами должен обладать толерантный человек?»
он должен быть внимательным и добрым, уметь общаться с людьми, хорошо излагать свои мысли. Люди отличаются друг от друга национальностью, привычками, одеждой, но живут они вместе, и надо уважать молодых и старых, здоровых и больных, бедных и богатых.
«А как вы думаете, вы сами — люди толерантные?»
а вот мне кажется, что вы все добрые, понимающие, толерантные.
И сейчас я вам это покажу, а вы мне поможете.
2. Первое упражнение «Комплименты» (10 мин.).
«Каждому из нас приятно, когда его хвалят, говорят о нем хорошие, добрые слова, особенно когда заслуженно.
Любой человек хочет, чтобы его любили и уважали, ценили и понимали, и очень часто нам просто необходимо слышать от окружающих людей добрые слова и пожелания. Ведь от этого улучшается настроение и мы готовы делиться радостью и помогать другим.
Сейчас каждый из вас, кто сядет на волшебный стул, узнает о себе много хорошего, все остальные ребята расскажут про положительные качества этого человека».
После того, как каждый ребенок побывал на «волшебном стуле», необходимо обсудить с классом, что дети почувствовали во время упражнения. Задаем следующие вопросы:
Вам понравилось это упражнение?
Почему?
Что нового вы узнали друг о друге?
Были комплименты одинаковыми или разными?
Почему разные?
«Действительно, ребята, каждый человек — это индивидуальность. Личность, обладающая своими собственными особенностями. Если бы мы были все одинаковыми — жить на свете было бы просто не интересно».
Личность, обладающая своими собственными особенностями. Если бы мы были все одинаковыми — жить на свете было бы просто не интересно».
3. Второе упражнение «Волшебный микрофон» (10 мин.).
садимся в круг вместе с детьми.
«Часто ребята, которые учатся в одном классе несколько лет, очень мало знают друг о друге — кто чем увлекается, кто что любит, а чего не любит. Когда знаешь человека, то можешь проявить к нему внимание, сравнить его интересы со своими. Попытаться понять совершенно не похожего на тебя человека, проявить к нему интерес, а если надо то и терпимость, толерантность не просто, потому что надо и себя хорошо знать, иметь представление о себе как о личности. Надо учиться жить, не ущемляя как своих интересов так и интересов других людей».
Каждый ребенок, получая «волшебный микрофон», отвечает на 2 вопроса (выбираем любые, чередуя):
Ваше увлечение (чем вы любите заниматься?)
О чем вы мечтаете?
Узнали ли вы о своих друзьях что–то новое?
Что нового вы узнали?
Что вас удивило, что понравилось?
«Теперь вы можете поделиться с друзьями своими интересами, умениями, мечтами не только не занятиях».
4. Третье упражнение «Нарисуй себя» (10 мин.).
Все дети получают нарисованный на бумаге шаблон человека (силуэт). На основе этого шаблона они рисуют себя, добавляя свои индивидуальные особенности. Затем следует обсуждение, вопросы:
Что общего у всех человечков? (голова, руки, ноги….)
Чем они отличаются? (одеждой, цветом волос, прической, цветом глаз.)
«Да, ребята, все мы очень разные, у каждого есть свои особенности, не говоря уже о внутреннем мире каждого человека. Мы должны уважать индивидуальность каждого человека, потому что мы все люди, у нас есть различия, но каждый из нас неповторим и нужен всем остальным».
4. Четвертое упражнение «Изгой» (10 мин.).
Сообщаем каждому ребенку «на ушко» название одного животного (кошка, собака, корова, лягушка). Важно поделить детей на 4 примерно одинаковые группы. Одному, как правило, самому озорному, сообщается слово «ворона».
«Не говоря того слова что я вам сказала, а произнося лишь звуки, которые издают эти животные, найдите таких же, как и вы, и соберитесь в группы».
Задаем вопросы всем:
Как вы узнали, где ваша группа?
Легко ли было найти «своих»?
Задаем вопросы, обращаясь к «вороне»:
Как ты себя чувствовал?
Возвращаемся ко всем детям:
Ребята, почему вы не обращали на него внимания?
Хотели ли вы быть на месте вороны?
Тяжело ли быть не похожим на всех?
Как вы в следующий раз отнесетесь к такому человеку?
4. Пятое упражнение «Звали, зовут, будут звать…» (10 мин.)
ученики рассаживаются в круг и по очереди произносят:
«Когда я был совсем маленький, меня звали Вовочка, сейчас в школе меня зовут Володя, а когда я вырасту, меня будут звать Владимир Николаевич».
Для многих детей их имя и отчество, произносимые вслух, звучат непривычно, но при этом они повышают уважение ребенка к себе и родителям, дают ориентир на будущее, на взросление.
«Нам хочется, чтобы к нам относились уважительно, называли ласково по имени, но для этого надо и самим понимать людей, чувствовать их настроение, стараться никого не обижать».
5. Творческая работа «Толерантный мир» (15 мин.).
работа в общей группе.
Требуемое оснащение: фломастеры, клеевые карандаши, ватман, вырезки из газет и рисунки детей с улыбающимися лицами людей (желательно разных рас и национальностей).
Музыкальное сопровождение: детские песни о дружбе – «От улыбки станет всем светлей», «Вместе весело шагать», «Солнечный круг» и т.п.
Детям предлагается сделать коллаж из вырезок и рисунков.
После оформления коллажа-стенгазеты – она вывешивается в классе или рекреации.
6. Итог внеклассного мероприятия (5 мин.).
задаем детям вопросы:
Ребята, я сегодня очень много нового узнала о вас, а вы друг о друге узнали что–то новое?
Зачем нам надо стремиться узнавать новое друг о друге?
Как вы считаете, наш «День толерантности» удался?
«К сожалению, еще не всегда люди бывают толерантны по отношению друг к другу, но я надеюсь, что придет время и «День толерантности» будет каждый день: все люди будут уважительно относиться друг к другу, никто никого обижать не будет. Каждый человек будет иметь собственное мировоззрение, будет отстаивать свои убеждения, и все люди научатся договариваться, а не станут воевать, ругаться, конфликтовать.
Каждый человек будет иметь собственное мировоззрение, будет отстаивать свои убеждения, и все люди научатся договариваться, а не станут воевать, ругаться, конфликтовать.
понимание и уважение иных народов, культур, цивилизаций, жизненных ценностей, включая культуру быта;
осознание необходимости взаимопонимания между людьми и народами;
способность общаться;
понимание необходимости активной межнациональной солидарности и сотрудничества;
готовность участвовать в решении проблем другого сообщества и этноса.
Результатами мероприятия должны быть:
Классный час 4 класс. Тема: Все мы разные или поговорим о толерантности.
Классный час 4 класс. Тема: Все мы разные или поговорим о толерантности.
Цели: познакомить учащихся с понятием “толерантность”, его происхождением, значением и актуальностью его формирования как нравственного качества, с основными чертами толерантной личности; способствовать воспитанию терпимого отношения к окружающим людям; воспитывать у детей человеколюбие, доброту и сопереживание, внимание к окружающим: близким людям, соседям по дому, по парте.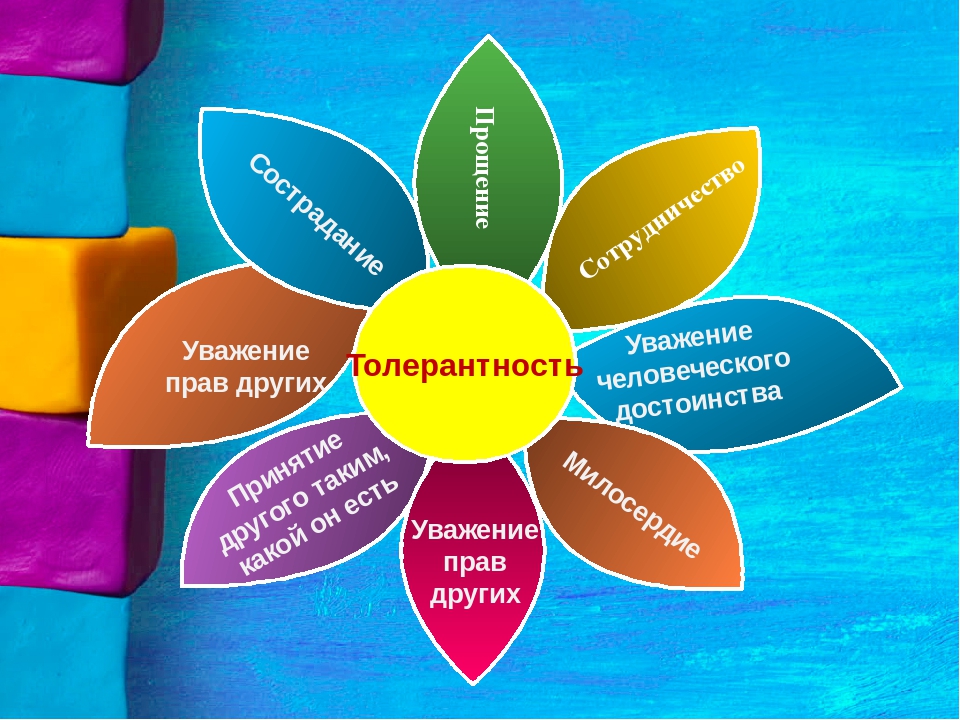 Предоставить учащимся возможность узнать своих одноклассников, развить умение наблюдать и правильно оценивать свои поступки и поступки друг друга.
Предоставить учащимся возможность узнать своих одноклассников, развить умение наблюдать и правильно оценивать свои поступки и поступки друг друга.
Задачи:
— формировать у детей понятие о толерантности;
— развивать представления о других людях на основе сопоставления себя с ними, выделения сходства и различий;
-воспитывать чувство уважения друг к другу;
— учить понимать проблемы других людей, воспитывать гуманное к ним отношение.
Оборудование и наглядность:
— Презентация к классному часу «Все мы разные или поговорим о толерантности»
— Мультипликационный фильм: «Ежик должен быть колючим?»
Ход классного часа
Организационный момент. Дети входят в класс под песню «Улыбка».
Вступительная беседа.
— Под какую песню вошли в класс? («Улыбка»).
— Что такое улыбка? ( Выражение на лице, мимика).
— А когда человек улыбается? (когда хорошее настроение, ему весело, когда человек добрый…)
— Верно, улыбка всегда располагает к общению, к уважению, к вниманию, к доброте. А если человек обладает всеми этими качествами, то говорят, что человек толерантный.
А если человек обладает всеми этими качествами, то говорят, что человек толерантный.
— Необычное слово? А вам интересно узнать больше об этом слове? (Да).
— Очень часто при чтении журналов, газет можно встретиться со словами, вошедшими в русский язык из других языков мира. Издавна люди общались с соседними народами. Налаживали с ними торговые и культурные связи. При общении в речь проникали иноязычные слова. Толерантность – имеет латинское происхождение и означает терпение, терпимость.
(Просмотр м/ф «Ежик должен быть колючим?»)
А теперь давайте посмотрим мультфильм о необычном ежике.
(Просмотр м/ф «Ежик должен быть колючим?»)
— Ребята, скажите, виноват ли ежик в том, что он родился с кудряшками вместо колючек? (Нет)
— А можно ли относиться к человеку плохо только потому, что он выглядит не так, как все вокруг? (Нет)
— А знаете, как называют людей, которые не похожи на других, которые выделяются в своем коллективе? (Нет)
— Таких людей называют «Белая ворона». А как вы думаете, почему их так называют? (Ответы детей.)
— Да, вы правы. Белая ворона – это человек, который сильно выделяется в коллективе (толпе).
— Ребята, а как вы думаете, хорошо, что все мы разные и не похожи друг на друга? (Да).
В нашем классе сегодня очень многолюдно. У нас в гостях человечки, которых вы нарисовали. Сразу вспомнилось замечательное стихотворение Алексея Моргуна.
Я рисую человечка –
Вышел он прямой, как свечка.
На кого же он похож?
Посмотрите, как хорош!
А у Нади и Аркаши
Человек объелся каши –
Круглый, добрый и веселый,
Очень сильный и здоровый.
Петин мальчик — великан.
У Алеши-атаман.
У Сергея же худой,
С ярко-рыжей головой.
А у Славика девчонка,
Словно Быхова Аленка,
Паша негра рисовал –
Сам от краски черным стал.
Люди разные на свете,
Словно человечки эти.
— Действительно, все человечки получились разные. Как вы думаете, почему? (Ответы детей.)
— А хорошо это или плохо, что все мы разные? (Ответы детей.)
— Если бы мы все стали одинаковыми, что тогда было бы?
Мы все разные — и это замечательно! У каждого из нас неповторимая внешность и свой уникальный внутренний мир.
— Ребята, на какой планете мы живем? ( Земля).
— Сколько материков на Земле? (6).
-Как называется наша страна?
— А какой народ живет в России?
— Ребята, посмотрите на иллюстрации. Кто изображен? ( Люди).
— Похожи ли они между собой? (Нет, есть молодые, пожилые, русские и нет, с разным цветом кожи, есть дети…).
— Совершенно верно, все люди разные, живут в разных уголках нашей планеты.
«Что такое толерантность?»
Толерантность это:
терпимость к чужому мнению;
понимание и уважение к личности другого человека независимо от каких-либо отличий;
доброжелательность в отношении к окружающим;
умение считаться с интересами и вкусами других людей.

Представьте себе такую ситуацию: два друга, Саша и Коля, разговаривая о своих домашних животных, начинают спорить и ругаться, потому что Саше нравятся собаки, и он считает их самыми умными созданиями, а Коле нравятся кошки, он считает их лучшими животными в мире. Мальчики поссорились и больше не разговаривают.
— Как вы думаете, из-за чего возник спор? (Из-за непонимания.)
— Кто из мальчиков прав? (Никто.)
— Чему мальчики должны научиться? (Принимать чужое мнение.)
Ребята, все мы разные люди, у нас у всех разные вкусы, привычки, разные таланты. Кто-то любит животных, кому-то нравятся машины, кто-то любит читать, а кто-то отлично играет в футбол, кому-то нравится рисовать, а кому-то — танцевать и т. д. И это не значит, что кто-то из нас хуже или лучше, умнее или глупее. Каждый из нас уникален и неповторим, поэтому мы должны ценить и уважать друг друга, то есть быть толерантными.
Понятие «толерантность» более широкое. Оно означает способность, умение терпеть, мириться с чужим мнением, быть снисходительным к поступкам других людей.
— Как вы понимаете выражение «уметь терпеть, мириться с чужим мнением»? (Выслушивать и уважать мнение других людей.)
Правильно, быть толерантным — значит признавать, что у каждого человека есть право на собственное мнение.
Физкультминутка. «Аплодисменты»
Необходимо хлопнуть в ладоши, если относишь себя к следующей категории:
Хлопают все присутствующие;
Только девчонки;
Только те, кто любит вкусно поесть;
Только те, кто опаздывает на уроки;
Только те, кто не любит каникулы;
Только те, кто любит сладкое;
Только те, кому нравится играть в футбол;
Только те, у кого прекрасное настроение.
Мой сосед по парте.
— Много времени мы с вами проводим в школе, в классе, за одной партой с соседом или соседкой. И порой не замечаем, друг друга, обижаем, говорим в их адрес плохие, обидные слова. А ведь вы все разные, непохожие друг на друга, но все по – своему интересны и неповторимы. Поэтому мы с вами сейчас поговорим о ваших соседях по парте.
— Когда у него день рождения?
— Есть ли у него братья, сёстры? Как их зовут?
— Где работают его мама и папа?
— Какие домашние животные живут у них дома?
— Чем увлекается в свободное время?
— В какие игры любит играть? Книги читать? Телепередачи смотреть?
— Я думаю, что теперь вы обратите внимание на своих одноклассников, соседа или соседку по парте, постараетесь узнать о нем (о ней) как можно больше. Ведь из этого слагается большая дружба.
Толерантный человек
Толерантное отношение друг к другу подразумевает умение людей договариваться между собой. Сейчас мы проверим, как вы умеете это делать.
(Каждая группа получает конверт с карточками, на которых написаны качества. Ребята должны выбрать пять качеств, которыми, по их общему мнению, обладает толерантный человек.)
— Перед вами набор слов. Пожалуйста, распределите эти слова на положительные качества, которые помогли бы построить доброе соседство, и отрицательные черты, на которые следует обратить внимание и постараться искоренить их.
Доброжелательность, равнодушие, отзывчивость, сочувствие, бездушие, злость, забота, зависть, внимание, высокомерие, ласка, жадность, сострадание, жестокость, человеколюбие, безжалостность, благотворительность.
| + | — |
| Доброжелательность, отзывчивость, сочувствие, забота, внимание, ласка, сострадание, человеколюбие, благотворительность. | Равнодушие, бездушие, злость, зависть, высокомерие, жадность, жестокость, безжалостность. |
Итог обсуждения:
Вежливо и доброжелательно обращаться с соседями;
Помогать, если возникнут проблемы;
Нужно уважают ценности и взгляды каждого, даже если чувствуешь несогласие с ним.
Итак, сочувствие, доброжелательность, терпение — все эти качества очень важны, ведь без них мы не можем жить в мире и согласии. Каждый из нас должен развивать в себе эти качества.
— Терпимость к чужому мнению, понимание и уважение к личности другого человека независимо от каких-либо отличий, доброжелательность в отношении к окружающим, умение считаться с интересами и вкусами других людей называется толерантность.
В первую очередь толерантность проявляется дома, в школе. Все знают, что нужно жить дружно, но иногда трудно сдержаться, когда мы видим недостатки других. Иногда у нас возникает ощущение, что к нам придираются, стремясь быть сильными, мы становимся нетолерантными и остаемся в одиночестве. Как нам стать толерантными? Прежде всего, надо остаться самим собой, видеть свои ошибки.
Давайте поиграем. Я называю ситуацию, а вы подумайте, как бы вы поступили?
Младший брат сломал твою игрушку.
1. Ты его прощаешь.
2. Ты ударяешь его.
Ты поссорился со своей сестрой.
1. Ты попытаешься объясниться с ней.
2. Ты обижаешься и мстишь.
С тобой поступают жестоко.
1. Ты отвечаешь тем же.
2. Ты говоришь «нет» и стремишься заручиться помощью.
Ты недоволен собой.
1. Ты говоришь, что людей без недостатков не бывает.
2. Ты все сваливаешь на других.
Тебе не хочется идти на прогулку со своими близкими.
Ты устраиваешь истерику.
2. Ты идешь с ними гулять.
В школе, как и везде, все разные: есть маленькие, большие, худые, полные. Почему мы иногда смеемся над ними? Быть толерантным – означает уважать других, невзирая на различия. Это означает быть внимательным к другим и обращать внимание на то, что нас сближает. Давайте попробуем проверить себя, проявляем ли мы толерантность.
1. Саша плохо одет…
— Это неважно.
— Ты подсмеиваешься над ним.
2. Пожилая женщина медленно идет.
— Ты отталкиваешь ее, чтобы обогнать.
— Ты помогаешь ей и придерживаешь дверь
3. На твоих глазах на кого-то нападают…
— Ты пытаешься защитить его.
— Ты делаешь вид, что ничего не замечаешь.
4. Цвет кожи у Джо отличается от твоего…
— Ты стремишься лучше узнать его.
— Ты говоришь: «Все люди твоего цвета кожи – это нули».
5. К тебе подходит ребенок-инвалид…
— Ты естественным образом разговариваешь с ним.
— Ты отходишь от него и не знаешь, что сказать.
Проявлять толерантность – это значит бережно относиться ко всему живому на нашей планете, вместе бороться с насилием, понимать друг друга, чтобы строить мирное будущее.
Толерантное отношение
(Работа в группах)
В разных странах существуют разные выражения, обозначающие толерантное отношение людей друг к другу. Давайте обратимся к ним. Ваша задача — объяснить, о чем эти выражения и какой общий смысл заложен в них. (карточки с крылатыми выражениями народов мира:
— Не делай другому того, от чего больно тебе.
— Во всем, как хотите, чтобы с вами поступали люди, так поступайте и вы с ними.
— Считай успех соседа своим успехом, а потерю соседа — своей потерей.
— В счастье и страдании, в радости и в горе мы должны относиться ко всем так, как относимся к себе.
(карточки с недописанными предложениями)
— Если у меня будет друг другой национальности, который не соглашается со мной в чем-то, то я. ..
— Если я увижу больного престарелого человека, то…
— Когда ко мне обратятся с просьбой дать совет в трудную минуту, то я…
— Если бы я был волшебником, то…
Заключительное задание “Дерево толерантности”
— Возьмите каждый по листочку и напишите на них, что, по-вашему, надо сделать, чтобы школа стала пространством толерантности, то есть, чтобы отношения в ней стали как можно более толерантными. (Ученики на листочках бумаги в форме листа какого-либо дерева пишут, что надо сделать, чтобы школа стала “Пространством толерантности”, листочки наклеиваются на символический рисунок дерева без листьев, и он вывешивается в классе.)
Подведение итогов
Учитель: Итак, давайте сделаем вывод: как стать толерантным человеком?
Уважайте людей со всеми их различиями физического или религиозного характера, различиями в образе жизни или мыслей.
Умейте чувствовать рядом с собой человека.
Каждый человек имеет право на индивидуальность, нужно уважать это право.
Помните, что, сделав добро, человек сам становится лучше, чище, светлее.
— Я бы хотела, чтобы мы добавили ещё одно правило:
Учитель: Вы скажете – это сложно. Нет, начните с малого и двигайтесь потихоньку. Если проявляем толерантность по отношению к самому себе, то мы становимся толерантными по отношению к другим и тогда нам легче принимать их такими, какие они есть. Конечно, мы не можем в одночасье сделать толерантным свое поведение, поведение других людей, но важно, что вы задумаетесь об этом.
– Так что значит быть толерантным? (Это значит бережно относиться ко всем, понимать друг друга.)
В конце мне хочется сказать, что класс – это маленькая семья. И хочется, чтобы в этой семье всегда царили доброта, уважение, взаимопонимание, не было бы ни ругани, ни ссор
Учитель: Пусть всегда в ваших сердцах живут любовь, терпение, прощение, гуманное отношение к другим людям, и это поможет вам научиться жить на земле, как люди.
(Дополнительно)
Учитель: Ребята, сегодня особо мне хотелось поговорить с вами о терпимом, гуманном отношении к людям с ограниченными возможностями здоровья, т.е. инвалидам. 3 декабря в календаре отмечен как международный день инвалида. Поэтому мы не можем не затронуть эту тему. Эти люди нуждаются в особой заботе. Ведь они лишены многого. Зачастую люди с ограниченными возможностями здоровья остаются один на один со своими проблемами. Современное общество не готово принять таких людей, считать их равными себе. А вот как вы поступите в следующих ситуациях.
4. Обсуждение ситуаций.
К тебе подходит ребёнок-инвалид…
Ты познакомился(-ась) с девочкой, с ней очень интересно общаться, но она – инвалид, передвигаться может только с помощью инвалидной коляски. Вокруг много любопытных глаз…
(Ответы детей.)
Учитель: Такие люди нуждаются в защите, поддержке, психологической, человеческой. Им нужно наше признание, принятие их такими, какие они есть. А некоторые из них достойны нашего уважения и восхищения, потому что судьба наградила удивительными талантами. Я хочу, чтобы сегодня по-новому взглянули на детей с ограниченными возможностями и узнали, какие замечательные они слагают стихи, не видя света, как они танцуют, не слыша музыки, какую проявляют силу воли, занимаясь спортом. А какие картины они рисуют, держа карандаш ногами и зубами! Как они, не умеющие ни говорить, ни читать, ни писать, тонко чувствуют природу, цвет, музыку, какие шедевры наивного искусства выходят из-под их неловких рук. А те, кто не могут ничего, проникновенно смотрят своими прекрасными глазами прямо в нашу душу. Эти дети пришли в мир точно так же, как и все другие люди. Они родились у обыкновенных родителей. Их ждали, им радовались. Но судьба распорядилась по-иному. Перспективы жизни таких детей и их семей стали иными — размытыми и пугающими. Но дети с инвалидностью при этом так и остались детьми, остались людьми. Нет сомнения, для них очень важно наше уважение, понимание, любовь, доверие. Они ждут от нас этого. Так давайте будем внимательны и милосердны к таким детям, взрослым, будем оказывать им всяческую поддержку.
Ученики читают стихотворения:
1) Не стой в стороне равнодушно,
Когда у кого-то беда.
Рвануться на выручку нужно
В любую минуту, всегда.
И если когда-то кому-то
Поможет твоя доброта,
Улыбка твоя,
Ты счастлив, что день
Не напрасно был прожит,
Что годы живешь ты не зря
2) Ах, как нам добрые слова нужны!
Не раз мы в этом убедились сами,
А может не слова – дела важны?
Дела – делами, а слова – словами.
Они живут у каждого из нас,
На дне души до времени хранимы,
Чтоб их произнести в тот самый час,
Когда они другим необходимы.
Международный День толерантности. Виртуальная выставка «Толерантность спасет мир»
Вы можете изучить и скачать доклад-презентацию на
тему Международный День толерантности. Виртуальная выставка «Толерантность спасет мир».
Презентация на заданную тему содержит 12 слайдов. Для просмотра воспользуйтесь
проигрывателем,
если материал оказался полезным для Вас — поделитесь им с друзьями с
помощью социальных кнопок и добавьте наш сайт презентаций в закладки!
Презентации» Литература» Международный День толерантности. Виртуальная выставка «Толерантность спасет мир»
Слайды и текст этой презентации
Слайд 1
Описание слайда:
Дорогие друзья! 16 ноября – Международный День толерантности.
Слайд 2
Описание слайда:
Бабаевская детская библиотека предлагает познакомиться с книгами виртуальной выставки «Толерантность спасет мир»
Слайд 3
Описание слайда:
Слайд 4
Описание слайда:
Виртуальная книжная выставка
«Толерантность спасет мир»
Бичер-Стоу Г.
Хижина дяди Тома. – М. : Дет. лит., 1987. – 556 с. : ил..
Роман американской писательницы Гарриет Бичер-Стоу, написанный в тот момент, когда борьба против рабства стала в центре внимания, приобрёл мировую известность. Он проникнут страстным негодованием против безнравственной сущности рабовладения.
Слайд 5
Описание слайда:
Слайд 6
Описание слайда:
Слайд 7
Описание слайда:
Троепольский, Г.Н.
Белый Бим Черное ухо: Повесть,- М.:ООО «Издательство АСТ»,2004.
Встретились и подружились одинокий пожилой человек и ласковый щенок. «Теплая дружба и преданность становились счастьем, потому что каждый понимал каждого и каждый не требовал от другого больше того, что он может дать». Но случилось горе – и собака осталась одна, хотя и среди людей, и отправилась на поиски любимого хозяина…
Слайд 8
Описание слайда:
Виртуальная книжная выставка «Толерантность спасет мир» Винник Л. Я не виноват! – М. : ЭКСМО, 2008. – 79 с. : (Детский проект Людмилы Улицкой). – (Другой, другие, о других) Как надо воспитывать ребёнка, почему в Японии детям до семи лет всё дозволено и как жилось маленьким спартанцам? Как устроена тюрьма, чем колокол заслужил порку и что такое инквизиция? В книге Ларисы Винник «Я не виноват!» вы найдете ответы на эти вопросы.
Слайд 9
Описание слайда:
Виртуальная книжная выставка
«Толерантность спасет мир»
Ли Х.
Убить пересмешника… – М. : АСТ : Хранитель, 2006. – 396 с..
Главный герой романа адвокат Аттикус Финч — носитель самых высоких человеческих качеств. Он справедлив, благороден, во всём и всегда верен своим принципам, по-настоящему смел. Взаимоотношения Аттикуса Финча со своими детьми, отношения доброго, умного, внимательного взрослого к детям – одна из сильнейших воспитательных сторон этой книги.
Слайд 10
Описание слайда:
Нанетти, Анджела.
Нанетти, Анджела.
«Мой дедушка был вишней».- М.: Самокат,2010.
Старик Оттавиано посадил вишневое дерево, которое стало лучшим другом его внуку, местом познания и приключений, символом непрерывности жизни. Главный герой этой книги — дедушка, который со своей энергией, чувством свободы и способностью слышать дыхание дерева, кажется немного «странным». И так же легко, как дышит дерево, автор книги касается таких тем, как страдание и смерть близких, и искренне верит, что «человек не умирает, пока вишневые деревья продолжают жить для него».
Книга «Мой дедушка был вишней» вошла в список выдающихся книг для детей «Белые вороны», была отмечена премиями в Италии, Германии и Франции .
Слайд 11
Описание слайда:
Виртуальная книжная выставка
«Толерантность спасет мир»
Мурашова, Е.В.
Класс коррекции. – М. : Самокат, 2007. – 191 с..
Повесть «Класс коррекции» сильно выделяется в общем потоке современной подростковой литературы. Тема детей-отбросов общества, зачастую умственно неполноценных, инвалидов, социально запущенных, слишком неудобна и некрасива, трудно решиться говорить об этом. Но у автора получается жизнелюбивое, оптимистическое произведение там, где, кажется, ни о каком оптимизме и речи быть не может. Е. Мурашова не развлекает читателя. Она призывает читающего подростка к совместной душевной и нравственной работе, помогает через соучастие, сочувствие героям книги осознавать себя как человека, личность, гражданина.
Слайд 12
Описание слайда:
Виртуальная книжная выставка «Толерантность спасет мир» Патерсон К. Великолепная Гилли Хопкинс – М. : Центр «Нарния», 2007. -224 с. : ил. Повесть о трудной девочке, которая пытается обрести чувство дома, несмотря на то, что её постоянно отправляют то к одним, то к другим приёмным родителям из-за якобы плохого поведения. Гилли начинает ненавидеть окружающих её людей и втайне мечтает вернуться к родной матери, хотя та бросила девочку. Кэтрин Патерсон – американская писательница, лауреат премии Х.К. Андерсена 1998 года.
Tags Международный День толерантности. Виртуальная выставка «Толерантность спасет мир»
Похожие презентации
Презентация успешно отправлена!
Ошибка! Введите корректный Email!
Память о Холокосте — путь к толерантности презентация к уроку (1, 2, 3, 4 класс) презентация
Слайд 1 Холокост и нацистский оккупационный режим на
территории СССР
Слайд 2 Холокост (с греч.
яз. Holocaust «всесожжение» ) — обозначение массовых убийств евреев, цыган, славян в 1933 — 1945 гг. фашистами.
«Холокост — это наконечник на выструганной веками стреле антисемитизма…» (Л. Коваль).
Слайд 3 Зарождение антисемитизма в Германии.
«Антисемитизм —
это международный язык фашистов» (И. Эренбург).
Слайд 4 Юдофобия — дословно «страх перед евреем»
— нетерпимость к евреям, синоним антисемитизма.
Слайд 5 Геноцид – уничтожение или преследование людей
по признаку определенной общности или происхождению.
Слайд 6 Геноцид —
уничтожение или преследование людей по
признаку определенной общности или происхождению. Но это не обязательно предполагает немедленное уничтожение народа или этнической группы, хотя может проявляться и в массовых убийствах. Чаще всего геноцид выступает как ряд спланированных акций, следующих одна за другой или происходящих одновременно; их цель — уничтожение тех черт народа, которые определяют его своеобразие. Этническая группа лишается возможности вести нормальную жизнь; происходит ликвидация общественных и политических институтов этноса, организуется дискриминация национальной культуры, языка, религии. Ликвидируются условия полноценного экономического существования; представителей народа лишают свободы, здоровья, унижают их человеческое достоинство.
Слайд 7 Национализм
– идеология и политика, основа
которых – идеи национальной исключительности, национального превосходства, трактовка нации как высшей формы общности
Слайд 8 Фашизм
— форма тоталитаризма, открыто террористическая
диктатура, уничтожающая демократические права и свободы, опирающаяся на прямое насилие, расизм.
Слайд 9 В 1932 году к власти
пришла
Национал
Социалистическая
Рабочая Партия Германии (нацисты),
а в январе 1933 года
Гитлер был назначен
канцлером
Германии.
Слайд 11 Закон о защите немецкой крови
и немецкой расы от 15 сентября 1935г.
1.1. Браки между евреями и государственными подданными немецкой или родственной крови запрещены.
2. Половая связь между евреями и государственными подданными немецкой или родственной крови запрещена.
3. Евреям запрещается нанимать на работу у себя в дом женщин — государственных подданных немецкой или родственной крови, не достигших 45 лет.
4.2. Евреям разрешено ношение одежды еврейских цветов. Это право охраняется государством.
ПЕРВОЕ ОБЪЯСНЕНИЕ К ЗАКОНУ О
ГРАЖДАНСТВЕ РЕЙХА
ОТ 14 НОЯБРЯ 1935 ГОДА
4. 1. Еврей не может быть гражданином Рейха. Он не имеет права голоса в политических вопросах, не может занимать должностей в государственных учреждениях.
4.2. Чиновники-евреи будут уволены до 31 декабря 1935 года.
5.1. Евреем считается лицо, среди предков которого во втором поколении (бабки и деды) имелось не менее трех лиц еврейской расы…
5.2. «Митлинг», который является государственным подданным, также считается евреем, если он происходит от двух еврейских предков.
Слайд 13 В 1937 году нацисты начали
отправлять евреев в концентрационные лагеря.
Слайд 14 В ночь на 4 ноября 1938
г. произошел еврейский погром, вошедший в историю как «Хрустальная ночь» (ночь битых стекол).
Слайд 15 Вторая мировая война и Холокост
«Гетто» -охраняемые
немецкими нацистами или поддерживающими их режим местными националистами жилые зоны, находящиеся на оккупированных территориях, куда насильственно перемещались евреи для компактного проживания.
Слайд 16 Вторжение гитлеровской армии в СССР
22 июня 1941г. — новый этап антиеврейской политики фашистов. На оккупированные территории Советского Союза вступали мобильные подразделения – айнзацгруппы.
Слайд 17 29-30 сентября 1941 г. в
Бабьем Яре, на окраине Киева, были расстреляны десятки тысяч евреев.
Слайд 18 В 1942 г. было решено создать
в Польше шесть лагерей смерти, куда депортировать еврейское население Европы (Треблинка, Хелм, Собибор, Майданек, Освенцим и Белжец).
Слайд 19 Помимо евреев в концлагеря попадали солдаты
Красной армии и гражданские лица СССР, плененные фашистами.
Слайд 20 27 января 1945 г.
солдаты Красной
Армии освободили узников Освенцима.
Эту дату принято считать окончанием холокоста.
Слайд 21 За период холокоста (1933 –
1945 гг.) было уничтожено
16 000 000 военных и гражданских лиц.
Слайд 22 Итоги Холокоста
6 000 000 евреев.
5 000
000 русских
3 000 000 украинцев
1 500 000 белорусов.
Слайд 23 Генеральная Ассамблея ООН провозгласила 27 января,
день освобождения Освенцима, Международным днём памяти жертв холокоста.
Планы уроков и мероприятия, посвященные Дню памяти жертв Холокоста
24 января 2022 г.
Эти ресурсы были обновлены 24 января 2022 г.
Хашоа. Следующая коллекция предназначена для того, чтобы предоставить учителям богатые и содержательные ресурсы о Холокосте, увлекательные планы уроков и информацию, чтобы помочь учащимся двигаться вперед, не забывая о прошлом.
1. Рекомендуемые интернет-учреждения
Мемориальный музей Холокоста США вдохновляет граждан и лидеров всего мира на борьбу с ненавистью, предотвращение геноцида и защиту человеческого достоинства. Музей продвигает ответственное преподавание Холокоста с помощью различных ресурсов и программ, чтобы помочь национальным педагогам расширить свои знания об истории Холокоста и реализовать эффективные стратегии обучения.
Являясь живым мемориалом Холокоста, Яд Вашем хранит память о прошлом и передает его значение будущим поколениям. Основан в 1953, как всемирный центр документации, исследований, образования и памяти о Холокосте, Яд Вашем сегодня является динамичным и жизненно важным местом межпоколенческих и международных встреч.
IWitness представляет учителям и их ученикам истории от первого лица о выживших и свидетелях геноцида из архива Института визуальной истории и образования USC Shoah Foundation с помощью мультимедийных учебных мероприятий, которые поощряют критическое мышление и самоанализ. и помочь учащимся понять, какое сильное влияние их слова и действия могут оказать на других.
2. Анкета о Холокосте
Этот полезный тест из 10 вопросов дает учащимся возможность решить, «что бы они сделали» на месте человека, преследуемого за веру. Используйте это как быстрое занятие или как разминку для более глубокого разговора.
3. Хронология событий
Используйте эту хронологию, заполненную изображениями и видео из Мемориального музея Холокоста США , чтобы помочь учащимся получить контекст и точку зрения на события, которые произошли, начиная с незадолго до 1933 к событиям, произошедшим после 1945 года.
4. Документальный | Путь к нацистскому геноциду
Этот 38-минутный фильм из Мемориального музея Холокоста США исследует подъем нацистов и укрепление власти в Германии. Используя редкие кадры, фильм исследует их идеологию, пропаганду и преследование евреев и других жертв. Он также описывает путь, по которому нацисты и их пособники вели государство к войне и убийству миллионов людей.
1. Холокост: оценка ответственности и совести
Этот план урока помогает учащимся проанализировать различные роли, сыгранные участниками Холокоста, и дает учащимся возможность осознать индивидуальное и общее влияние их действия.
2. Лидице и Лежаки: их истории через марки
Этот урок раскрывает захватывающую историю городов Лидице и Лежаки, которые были стерты с лица земли нацистами, рассказанную через памятные марки. Студенты изучат реальные изображения марок, чтобы раскрыть историю этих двух мест опустошения.
3. Праведники народов мира: спасатели Холокоста
Холокост был одним из самых мрачных периодов в истории человечества, но были и акты мужества и сопротивления, которые казались лучом надежды в море отчаяния и отчаяния. горе. И евреи, и неевреи рисковали своими жизнями, чтобы спасти других, и в этом уроке рассказывается о тех, кто получил звание «Праведник народов мира» за их храбрость и самопожертвование. Этот урок, основанный на исследованиях, поможет учащимся определить и установить связь с теми, кто решил не быть сторонним наблюдателем, а вместо этого стать героем.
3. План урока и фильм | Один выживший помнит: антисемитизм
Этот план урока, созданный Преподавание толерантности, знакомит учащихся с антисемитизмом, исследует использование пропаганды и стереотипов, знакомит учащихся с отрицанием Холокоста и связывает их с современным антисемитизмом, расизм, предрассудки и фанатизм. Этот урок был сделан в дополнение к фильму « Один выживший вспоминает », который был доступен для онлайн-трансляции в сотрудничестве с Мемориальным музеем Холокоста США и HBO. (Для просмотра необходимо создать учетную запись.)
1. Ресурс | Противостоять антисемитизму
Используйте этот увлекательный ресурс Мемориального музея Холокоста США , чтобы привлечь внимание учащихся к постоянной проблеме антисемитизма, с которой сталкиваются евреи во всем мире, и к тому, как они могут помочь стать частью решение, чтобы положить этому конец.
2. Документальный фильм | Хуже, чем война
Используйте этот документальный фильм Дэниела Гольдхагена и PBS, чтобы помочь учащимся понять геноцид, чтобы они, вырастая, не становились сторонними наблюдателями. Гольденхаген использует личные интервью, чтобы осветить историю геноцида в 20 веке. «Лицом к истории и самим себе» дает богатую пособие для преподавателей , которое прилагается к фильму.
Осложненные инфекции мочевыводящих путей у взрослых
1. McGowan JE Jr, Chesney PJ, Crossley KB, LaForce FM. Руководство по применению системных глюкокортикостероидов при лечении отдельных инфекций. Рабочая группа по использованию стероидов, Комитет по противомикробным препаратам, Американское общество инфекционных заболеваний. J Infect Dis 1992; 165:1-13. [PubMed] [Google Scholar]
2. Рубин Р.Х., Шапиро Э.Д., Андриоле В.Т., Дэвис Р.Дж., Штамм В.Е. Оценка новых противоинфекционных препаратов для лечения инфекций мочевыводящих путей. Американское общество инфекционистов и Управление по санитарному надзору за качеством пищевых продуктов и медикаментов. клин заразить дис 1992;15(Приложение 1):S216-27. [PubMed] [Google Scholar]
3. Николь Л.Е. Практический подход к лечению осложненной инфекции мочевыводящих путей. Наркотики и старение 2001;18:243-54. [PubMed] [Google Scholar]
4. Донлан Р.М., Костертон Дж.В. Биопленки: механизмы выживания клинически значимых микроорганизмов. Clin Microbiol Rev 2002; 15:167-93. [Бесплатная статья PMC] [PubMed] [Google Scholar]
5. Николь Л.Э. Хронический постоянный катетер и инфекция мочевыводящих путей у жителей учреждения длительного ухода. Infect Control Hosp Epidemiol 2001;22:316-21. [PubMed] [Академия Google]
6. Липский Б.А. Инфекции мочевыводящих путей у мужчин. Эпидемиология, патофизиология, диагностика и лечение. Энн Интерн Мед 1989; 110:138-50. [PubMed] [Google Scholar]
7. Raz R, Gennesin Y, Wasser J, et al. Рецидивирующие инфекции мочевыводящих путей у женщин в постменопаузе. Clin Infect Dis 2000;30:152-6. [PubMed] [Google Scholar]
8. Cardenas DD, Hooton TM. Инфекция мочевыводящих путей у лиц с травмой спинного мозга. Arch Phys Med Rehabil 1995; 76: 272-80. [PubMed] [Академия Google]
9. Николь Л.Е. Бессимптомная бактериурия у пожилых людей. Infect Dis Clin North Am 1997; 11: 647-67. [PubMed] [Google Scholar]
10. Тенни Дж. Х., Уоррен Дж. В. Бактериурия у женщин с долгосрочными катетерами: парное сравнение постоянных и сменных катетеров. J Infect Dis 1988; 157:199-202. (Ошибки в 1988 г.; 157:1112). [PubMed] [Google Scholar]
11. Уэйтс К.Б., Канупп К.С., ДеВиво М.Дж. Эффективность и переносимость норфлоксацина при лечении осложненной инфекции мочевыводящих путей у амбулаторных больных с нейрогенным мочевым пузырем, вторичным по отношению к травме спинного мозга. Урология 1991;38:589-96. [PubMed] [Google Scholar]
12. Николь Л.Э., Луи Т.Дж., Дюбуа Дж., Мартель А., Хардинг Г.К., Синав С.П. Лечение осложненных инфекций мочевыводящих путей ломефлоксацином по сравнению с триметоприм-сульфаметоксазолом. Противомикробные агенты Chemother 1994;38:1368-73. [Бесплатная статья PMC] [PubMed] [Google Scholar]
13. Cox CE, Holloway WJ, Geckler RW. Многоцентровое сравнительное исследование меропенема и имипенема/циластатина в лечении осложненных инфекций мочевыводящих путей у госпитализированных пациентов. клин заразить дис 1995;21:86-92. [PubMed] [Google Scholar]
14. Johnson JR, Roberts PL, Olsen RJ, Moyer KA, Stamm WE. Профилактика катетер-ассоциированной инфекции мочевыводящих путей с помощью мочевого катетера, покрытого оксидом серебра: клинические и микробиологические корреляты. J Infect Dis 1990;162:1145-50. [PubMed] [Google Scholar]
15. Николь Л.Э., Бьорнсон Дж., Хардинг Дж.К., Макдонелл Дж.А. Бактериурия у пожилых мужчин в лечебных учреждениях. N Engl J Med 1983;309:1420-5. [PubMed] [Google Scholar]
16. Bakke A, Digranes A. Бактериурия у пациентов, получавших чистую прерывистую катетеризацию. Scand J Infect Dis 1991;23:577-82. [PubMed] [Google Scholar]
17. Bennett CJ, Young MN, Darrington H. Различия в инфекциях мочевыводящих путей у мужчин и женщин с травмами спинного мозга при периодической катетеризации. Параплегия 1995; 33:69-72. [PubMed] [Google Scholar]
18. Николь Л.Е. Возбудители мочевыводящих путей при осложненной инфекции и у лиц пожилого возраста. J Infect Dis 2001;183(Приложение 1):S5-8. [PubMed] [Google Scholar]
19. Tambyah PA, Maki DG. Взаимосвязь между пиурией и инфекцией у пациентов с постоянным мочевым катетером: проспективное исследование 761 пациента. Arch Intern Med 2000; 160: 673-82. [PubMed] [Академия Google]
20. Уэйтс К.Б., Канупп К.С., ДеВиво М.Дж. Эпидемиология и факторы риска инфекции мочевыводящих путей после травмы спинного мозга. Arch Phys Med Rehabil 1993; 74: 691-5. [PubMed] [Google Scholar]
21. Raz R, Schiller D, Nicolle LE. Хроническая замена постоянного катетера перед антимикробной терапией при симптоматической инфекции мочевыводящих путей. Дж. Урол 2000; 164:1254-8. [PubMed] [Google Scholar]
22. Райт С.В., Ренн К.Д., Хейнс М.Л. Резистентность к триметоприму-сульфаметоксазолу среди изолятов кишечной палочки в моче. J Gen Intern Med 1999;14:606-9. [Бесплатная статья PMC] [PubMed] [Google Scholar]
23. Дембри Л.М., Андриоле В.Т. Почечные и периренальные абсцессы. Infect Dis Clin North Am 1997; 11: 663-80. [PubMed] [Google Scholar]
24. Паттерсон Дж. Э., Андриоле В. Т. Бактериальные инфекции мочевыводящих путей при сахарном диабете. Infect Dis Clin North Am 1997; 11: 735-50. [PubMed] [Google Scholar]
25. Trop CS, Bennett CJ. Вегетативная дисрефлексия и ее урологические последствия: обзор. Дж. Урол 1991; 146:1461-9. [PubMed] [Академия Google]
26. Рапп Н.С., Гилрой Дж., Лернер А.М. Роль бактериальной инфекции в обострении рассеянного склероза. Am J Phys Med Rehabil 1995; 74: 415-8. [PubMed] [Google Scholar]
27. Николь Л.Е. Последствия бессимптомной бактериурии у пожилых. Int J Antimicrob Agents 1994;4:107-11. [PubMed] [Google Scholar]
28. Orr P, Nicolle LE, Duckworth H, et al. Фебрильная инфекция мочевыводящих путей у пожилых людей, находящихся в специализированных учреждениях. Am J Med 1996; 100:71-7. [PubMed] [Академия Google]
29. Гриббл М.Дж., МакКаллум Н.М., Шехтер М.Т. Оценка диагностических критериев бактериурии у пациентов с острым повреждением спинного мозга, перенесших периодическую катетеризацию. Diagn Microbiol Infect Dis 1988;9:197-206. [PubMed] [Google Scholar]
30. Старк Р.П., Маки Д.Г. Бактериурия у катетеризированного больного. Какой количественный уровень бактериурии имеет значение? N Engl J Med 1984; 311:560-4. [PubMed] [Google Scholar]
31. Bakke A, Digranes A. Бактериурия у пациентов, получавших чистую прерывистую катетеризацию. Scand J Infect Dis 1991;23:577-82. [PubMed] [Google Scholar]
32. Stamm WS. Катетер-ассоциированные инфекции мочевыводящих путей. Эпидемиология, патогенез и профилактика. Am J Med 1991; 91 (Приложение B): 65S-71S. [PubMed] [Google Scholar]
33. Riedl CR, Plas E, Hubner WA, Zimmerl H, Ulrich W, Pfluger H. Бактериальная колонизация мочеточниковых стентов. Евр Урол 1999;36:53-9. [PubMed] [Google Scholar]
34. Monane M, Gurwitz JH, Lipsitz LA, Glynn RJ, Choodnovskiy I, Avorn J. Эпидемиологические и диагностические аспекты бактериурии: продольное исследование пожилых женщин. J Am Geriatr Soc 1995;43:618-22. [PubMed] [Google Scholar]
35. Ouslander JG, Schapira M, Fingold S, Schnelle J. Точность экспресс-тестов мочи среди жителей домов престарелых с недержанием мочи и бессимптомной бактериурией. J Am Geriatr Soc 1995;43:772-5. [PubMed] [Google Scholar]
36. Hooton TM, Stamm WE. Диагностика и лечение неосложненной инфекции мочевыводящих путей. Infect Dis Clin North Am 1997; 11: 551-81. [PubMed] [Google Scholar]
37. Николь Л.Э., Мэйхью Дж.В., Брайан Л. Проспективное рандомизированное сравнение терапии и отсутствия терапии бессимптомной бактериурии у женщин в лечебных учреждениях. Am J Med 1987;83:27-33. [PubMed] [Google Scholar]
38. Abrutyn E, Mossey J, Berlin JA, et al. Является ли бессимптомная бактериурия предиктором смертности и снижает ли антимикробное лечение смертность у пожилых амбулаторных женщин? Энн Интерн Мед 1994;120:827-33. (Ошибки в 1994 г.; 121:901). [PubMed] [Google Scholar]
39. Ouslander JG, Shapira M, Schnelle JF, et al. Влияет ли эрадикация бактериурии на тяжесть хронического недержания мочи у проживающих в домах престарелых? Энн Интерн Мед 1995; 122: 749-54. [PubMed] [Google Scholar]
40. Boscia JA, Kobasa WD, Knight RA, Abrutyn E, Levison ME, Kaye D. Терапия по сравнению с отсутствием терапии бактериурии у пожилых амбулаторных негоспитализированных женщин. ЯМА 1987; 257:1062-71. [PubMed] [Google Scholar]
41. Mohler JL, Cowen DL, Flanigan RC. Подавление и лечение инфекции мочевыводящих путей у пациентов с прерывистой катетеризацией нейрогенного мочевого пузыря. Дж. Урол 1987; 138:336-40. [PubMed] [Google Scholar]
42. Warren JW, Anthony WC, Hoopes JM, Muncie HL Jr. Цефалексин при чувствительной бактериурии у пациентов с длительной катетеризацией без лихорадки. ДЖАМА 1982;248:454-8. [PubMed] [Google Scholar]
43. Harding GK, Nicolle LE, Ronald AR, et al. Как долго следует лечить катетерную инфекцию мочевыводящих путей у женщин? Рандомизированное контролируемое исследование. Энн Интерн Мед 1991;114:713-9. [PubMed] [Google Scholar]
44. Harding GKM, Zhanel GG, Nicolle LE, Cheang M; Группа по изучению диабетической мочевой инфекции Манитобы. Антимикробная терапия у женщин с сахарным диабетом и бессимптомной бактериурией. N Engl J Med 2002;347:1576-83. [PubMed] [Академия Google]
45. Syndman DR. Посттрансплантационное микробиологическое наблюдение. Clin Infect Dis 2001; 33 (Приложение 1): S22-5. [PubMed] [Google Scholar]
46. Fox BC, Sollinger HW, Belzer FO, Maki DG. Проспективное рандомизированное двойное слепое исследование триметоприма-сульфаметоксазола для профилактики инфекции при трансплантации почки: клиническая эффективность, абсорбция триметоприм-сульфаметоксазола, влияние на микрофлору и соотношение затрат и результатов профилактики. Am J Med 1990; 89: 255-74. [PubMed] [Академия Google]
47. Фишман Ю.А., Рубин Р.Х. Инфекция у реципиентов трансплантированных органов. N Engl J Med 1998;338:1741-51. [PubMed] [Google Scholar]
48. Takai K, Tollemar J, Wilczek HE, Groth CG. Инфекции мочевыводящих путей после трансплантации почки. Clin Transplant 1998; 12:19-23. [PubMed] [Google Scholar]
49. Лёрова Л., Лача Дж., Скибова Дж., Теплан В., Витко С., Шук О. Инфекция мочевыводящих путей у пациентов с урологическими осложнениями после трансплантации почки в отношении долгосрочной функции и выживаемости аллотрансплантата . Энн Трансплант 2001; 6:19-20. [PubMed] [Google Scholar]
50. Гасемян С.М., Гулерия А.С., Хаванд Н.Ю., Лайт Дж.А. Диагностика и лечение урологических осложнений трансплантации почки. Клин Трансплант 1996; 10:218-23. [PubMed] [Google Scholar]
51. Олсон Э.С., Куксон Б.Д. Играют ли противомикробные препараты роль в предотвращении септицемии после инструментальной обработки мочевыводящих путей? J Hosp Infect 2000;45:85-97. [PubMed] [Google Scholar]
52. Cafferkey MT, Falkiner FR, Gillespie WA, Murphy DM. Антибиотики для профилактики септицемии в урологии. J Антимикроб Chemother 1982;9:471-7. [PubMed] [Google Scholar]
53. Grabe M. Периоперационная антибиотикопрофилактика в урологии. Курр Опин Урол 2001;11:81-5. [PubMed] [Google Scholar]
54. Jewes LA, Gillespie WA, Leadbetter A, et al. Бактериурия и бактериемия у пациентов с длительно установленными постоянными катетерами — домашнее исследование. J Med Microbiol 1988; 26:61-5. [PubMed] [Google Scholar]
55. Bregenzer T, Frei R, Widmer AF и др. Низкий риск бактериемии при замене катетера у пациентов с длительно установленными мочевыми катетерами. Арх стажер Мед 1997;157:521-5. [PubMed] [Google Scholar]
56. Cronan JJ, Horn DL, Marcello A, et al. Антибиотики и уход за нефростомической трубкой: предварительные наблюдения. Часть II. Бактериемия. Радиология 1989;172:1043-5. [PubMed] [Google Scholar]
57. Carmignani G, De Rose AF, Olivieri L, Salvatori E, Rosignoli MT, Dionisio P. Прулифлоксацин по сравнению с ципрофлоксацином при лечении взрослых с осложненными инфекциями мочевыводящих путей. Urol Int 2005;74:326-31 [PubMed] [Google Scholar]
58. Talan DA, Klimberg IW, Nicolle LE, Song J, Kowalsky SF, Church DA. Ципрофлоксацин пролонгированного действия один раз в сутки при осложненных инфекциях мочевыводящих путей и остром неосложненном пиелонефрите. Дж. Урол 2004; 171: 734-9. [PubMed] [Google Scholar]
59. Naber KG, Bartnicki A, Bischoff W, et al. Гатифлоксацин в дозе 200 мг или 400 мг один раз в сутки столь же эффективен, как и ципрофлоксацин в дозе 500 мг два раза в сутки при лечении пациентов с острым пиелонефритом или осложненными инфекциями мочевыводящих путей. Int J Antimicrob Agents 2004; 23 (Приложение 1): S41-S53. [PubMed] [Google Scholar]
60. Уллерид П., Сандберг Т. Ципрофлоксацин в течение 2 или 4 недель при лечении фебрильной инфекции мочевыводящих путей у мужчин: рандомизированное исследование с последующим наблюдением в течение 1 года. Scand J Infect Dis 2003;35:34-9. [PubMed] [Google Scholar]
61. Jimenez-Cruz F, Josovich A, Cajigas J, et al. Проспективное, многоцентровое, рандомизированное, двойное слепое исследование, в котором сравнивали эртапенем и цефтриаксон с последующей соответствующей пероральной терапией при осложненных инфекциях мочевыводящих путей у взрослых. Урология 2002;60:16-22. [PubMed] [Google Scholar]
62. Cox CE, Marbury TC, Pittman WG, et al. Рандомизированное двойное слепое многоцентровое сравнение гатифлоксацина и ципрофлоксацина при лечении осложненной инфекции мочевыводящих путей и пиелонефрита. Клин Тер 2002; 24: 223-36. [PubMed] [Академия Google]
63. Tomera KM, Burdmann EA, Pamo Reyna OG, et al. Эртапенем по сравнению с цефтриаксоном с последующей соответствующей пероральной терапией для лечения осложненных инфекций мочевыводящих путей у взрослых: результаты проспективного рандомизированного двойного слепого многоцентрового исследования. Противомикробные агенты Chemother 2002;46:2895-900. [Бесплатная статья PMC] [PubMed] [Google Scholar]
64. Набер К.Г., Савов О., Салмен Х.К. Пиперациллин 2 г/тазобактам 0,5 г столь же эффективен, как имипенем 0,5 г/циластатин 0,5 г при лечении острого неосложненного пиелонефрита и осложненных инфекций мочевыводящих путей. Int J Антимикробные агенты 2002; 19: 95-103. [PubMed] [Google Scholar]
65. Raz R, Naber NG, Raizenberg C, et al. Ципрофлоксацин 250 мг два раза в день по сравнению с офлоксацином 200 мг два раза в день при лечении осложненных инфекций мочевыводящих путей у женщин. Eur J Clin Microbiol Infect Dis 2000;19:327-31. [PubMed] [Google Scholar]
66. Кркмери С., Набер Н.Г. Ципрофлоксацин один или два раза в день при лечении осложненных инфекций мочевыводящих путей. Немецкая исследовательская группа по ципрофлоксацину ИМП. Int J Antimicrob Agents 1999;11:133-8. [PubMed] [Академия Google]
67. Климберг И.В., Кокс С.Е. II, Фаулер С.Л., Кинг В., Ким С.С., Каллери-Д’Амико С. Контролируемое исследование левофлоксацина и ломефлоксацина при лечении осложненной инфекции мочевыводящих путей. Урология 1998;51:610-5. [PubMed] [Google Scholar]
68. Франкеншмидт А., Набер К.Г., Бишофф В., Кульманн К. Флероксацин один раз в день против ципрофлоксацина два раза в день при лечении осложненных инфекций мочевыводящих путей. Дж. Урол 1997; 158:1494-9. [PubMed] [Google Scholar]
69. Pisani E, Bartoletti R, Trinchieri A, Rizzo M. Ломефлоксацин против ципрофлоксацина при лечении осложненных инфекций мочевыводящих путей: многоцентровое исследование. Джей Чемотер 1996;8:210-3. [PubMed] [Google Scholar]
70. Naber KG, di Silverio F, Geddes A, Guibert J. Сравнительная эффективность спарфлоксацина и ципрофлоксацина при лечении осложненной инфекции мочевыводящих путей. J Antimicrob Chemother 1996;37(Приложение A):135-44. [PubMed] [Google Scholar]
71. Cox CE, Holloway WJ, Geckler RW. Многоцентровое сравнительное исследование меропенема и имипенема/циластатина в лечении осложненных инфекций мочевыводящих путей у госпитализированных пациентов. клин заразить дис 1995;21:86-92. [PubMed] [Google Scholar]
72. Gottlieb PL. Сравнение эноксацина с триметоприм-сульфаметоксазолом при лечении пациентов с осложненной инфекцией мочевыводящих путей. Клин Тер 1995; 17:493-502. [PubMed] [Google Scholar]
73. Николь Л.Э., Луи Т.Дж., Дюбуа Дж., Мартель А., Хардинг Г. К., Синав С.П. Лечение осложненных инфекций мочевыводящих путей ломефлоксацином по сравнению с лечением триметопримом-сульфаметоксазолом. Противомикробные агенты Chemother 1994;38:1368-73. [Бесплатная статья PMC] [PubMed] [Google Scholar]
74. Кокс СЕ. Сравнение внутривенного введения флероксацина с цефтазидимом при лечении осложненных инфекций мочевыводящих путей. Am J Med 1993; 94:118S-25S. [PubMed] [Google Scholar]
75. Pummer K. Сравнение флероксацина и норфлоксацина при лечении инфекций мочевыводящих путей: многоцентровое двойное слепое проспективное рандомизированное сравнительное исследование. Am J Med 1993; 94(3A):108S-13S. [PubMed] [Google Scholar]
76. Чайлдс С.Дж. Флероксацин по сравнению с норфлоксацином для перорального лечения серьезных инфекций мочевыводящих путей. Am J Med 1993;94(3А):105S-7S. [PubMed] [Google Scholar]
77. Pittman W, Moon JO, Hamrick LC Jr и соавт. Рандомизированное двойное слепое исследование высоких и низких доз флероксацина по сравнению с норфлоксацином при осложненной инфекции мочевыводящих путей. Am J Med 1993:94(3A):101S-4S. [PubMed] [Google Scholar]
78. Цефпиром против цефтазидима при лечении инфекций мочевыводящих путей. J Antimicrob Chemother 1992; 29 (Приложение А): 95-104. [PubMed] [Google Scholar]
79. Fang GD, Brennen C, Wagener M, et al. Использование ципрофлоксацина по сравнению с использованием аминогликозидов для лечения осложненной инфекции мочевыводящих путей: проспективное рандомизированное клиническое и фармакокинетическое исследование. Противомикробные агенты Chemother 1991;35:1849-55. [Бесплатная статья PMC] [PubMed] [Google Scholar]
80. Cox CE. Последовательное внутривенное и пероральное введение ципрофлоксацина по сравнению с внутривенным введением цефтазидима при лечении осложненных инфекций мочевыводящих путей. Am J Med 1989:87(5A):157S-9S. [PubMed] [Google Scholar]
81. Cox CE, Drylie DM, Klimberg I, et al. Многоцентровое двойное слепое контролируемое исследование применения эноксацина триметоприм-сульфаметоксазолом при лечении пациентов с осложненными инфекциями мочевыводящих путей. Дж Урол 1989;141:575-8. [PubMed] [Google Scholar]
82. Horowitz EA, Preheim LC, Safranek TJ, Pugsley MP, Sanders CC, Bittner MJ. Рандомизированное двойное слепое сравнение цефтазидима и моксалактама при осложненных инфекциях мочевыводящих путей. Противомикробные агенты Chemother 1985; 28:299-301. [Бесплатная статья PMC] [PubMed] [Google Scholar]
83. Bailey RR, Peddie BA, Lynn KL, Swainson CP. Сравнение нетилмицина с цефоперазоном при лечении тяжелых или осложненных инфекций мочевыводящих путей. Aust N Z J Med 1985;15:22-6. [PubMed] [Google Scholar]
84. Nishiura T. Клиническое сравнение цефоперазона при осложненных инфекциях мочевыводящих путей двойным слепым методом. Клин Тер 1980; 3:190-205. [PubMed] [Google Scholar]
85. Mombelli G, Pezzoli R, Pinoja-Lutz G, Monotti R, Marcone C, Franciolli M. Пероральное и внутривенное введение ципрофлоксацина в начальной эмпирической терапии тяжелого пиелонефрита или осложненных инфекций мочевыводящих путей: A проспективное рандомизированное клиническое исследование. Арх стажер Мед 1999;159:53-8. [PubMed] [Google Scholar]
86. Madsen PO, Baumueller A, Frimodt-Moller N, et al. Лечение нетилмицином осложненных инфекций мочевыводящих путей. Scand J Infect Dis 1980; (Приложение 23): 128-31. [PubMed] [Google Scholar]
87. De Gier R, Karperien A, Bouter K, et al. Последовательное исследование внутривенного и перорального введения флероксацина в течение 7 или 14 дней при лечении осложненных инфекций мочевыводящих путей. Int J Antimicrob Agents 1995; 6:27-30. [PubMed] [Google Scholar]
88. Sifuenles-Osornio J, Jakob E, Clara L, et al. Пиперациллин/тазобактам в лечении госпитализированных пациентов с инфекциями мочевыводящих путей: открытое несравнительное и многоцентровое исследование. Джей Чемотер 1996;8:122-9. [PubMed] [Google Scholar]
89. Nowe P. Пиперациллин/тазобактам при осложненных инфекциях мочевыводящих путей. Intensive Care Med 1994; 20 (Приложение 3): S39-S42. [PubMed] [Google Scholar]
90. Dow G, Rao P, Harding G, et al. Проспективное рандомизированное исследование 3 или 14 дней лечения ципрофлоксацином острой инфекции мочевыводящих путей у пациентов с травмой спинного мозга. Clin Infect Dis 2004;39:658-65. [PubMed] [Google Scholar]
91. Николь Л.Э., Мэйхью Дж.В., Брайан Л. Результат антимикробной терапии бессимптомной бактериурии у пожилых женщин, проживающих в лечебном учреждении. Возраст Старение 1988;17:187-92. [PubMed] [Google Scholar]
92. Чинн Р. Х., Маскелл Р., Мид Дж. А., Полак А. Камни в почках и мочевая инфекция: исследование лечения антибиотиками. Br Med J 1976; 2:1411-3. [Бесплатная статья PMC] [PubMed] [Google Scholar]
93. Sheehan GJ, Harding GK, Haase DA, et al. Двойное слепое рандомизированное сравнение 24-недельного курса норфлоксацина и 12-недельного курса норфлоксацина с последующим 12-недельным курсом плацебо при лечении осложненной инфекции мочевыводящих путей. Противомикробные агенты Chemother 1988; 32:1292-3. [Бесплатная статья PMC] [PubMed] [Google Scholar]
94. Boerema JB, van Saene HK. Лечение норфлоксацином осложненной инфекции мочевыводящих путей. Scand J Infect Dis 1986; 48:20-6. [PubMed] [Google Scholar]
95. Leigh DA, Emmanuel FX, Petch VJ. Терапия ципрофлоксацином при осложненных инфекциях мочевыводящих путей, вызванных Pseudomonas aeruginosa и другими резистентными бактериями. J Antimicrob Chemother 1986;18(Приложение D):117-21. [PubMed] [Google Scholar]
96. Leigh DA, Emmanuel FX. Лечение Pseudomonas aeruginosa инфекции мочевыводящих путей с норфлоксацином. J Antimicrob Chemother 1984;13(Приложение B):85-8. [PubMed] [Google Scholar]
97. Cox CE. Терапия азтреонамом при осложненных инфекциях мочевыводящих путей, вызванных бактериями с множественной лекарственной устойчивостью. Rev Infect Dis 1985; 7 (Приложение 4): S767-71. [PubMed] [Google Scholar]
98. Уорд Т.Т., Амон М.Б., Краузе Л.К. Комбинированная терапия амдиноциллином и цефокситином мультирезистентных Serratia marcescens инфекций мочевыводящих путей. Am J Med. 1983;75(Приложение 2А):85-9. [PubMed] [Google Scholar]
99. Kauffman CA, Vazquez JA, Sobel JD, et al. Проспективное многоцентровое эпиднадзорное исследование грибка у госпитализированных пациентов. Группа по изучению микозов Национального института аллергии и инфекционных заболеваний (NIAID). Clin Infect Dis 2000;30:14-8. [PubMed] [Google Scholar]
100. Schwab U, Chernomas F, Larcom L, Weems J. Молекулярное типирование и чувствительность к флуконазолу мочевых изолятов Candida glabrata от госпитализированных пациентов. Диагностика Microbiol Infect Dis 1997;29:11-17. [PubMed] [Google Scholar]
101. Sobel JD, Kauffman CA, McKinsey D, et al. Кандидурия: рандомизированное двойное слепое исследование лечения флуконазолом и плацебо. Группа по изучению микозов Национального института аллергии и инфекционных заболеваний (NIAID). Clin Infect Dis 2000; 30:19-24. [PubMed] [Google Scholar]
102. Leu HS, Huang CT. Избавление от грибка с помощью краткосрочных противогрибковых схем: проспективное рандомизированное контролируемое исследование. Clin Infect Dis 1995; 20:1152-7. [PubMed] [Академия Google]
103. Jacobs LG, Skidmore EA, Freeman K, Lipschultz D, Fox N. Пероральный флуконазол по сравнению с промыванием мочевого пузыря амфотерицином B для лечения грибковых инфекций мочевыводящих путей у пожилых пациентов. Clin Infect Dis 1996; 22:30-5. [PubMed] [Google Scholar]
104. Fan-Havard P, O’Donovan C, Smith SM, Oh J, Bamberger M, Eng RH. Пероральный флуконазол в сравнении с промыванием мочевого пузыря амфотерицином В для лечения кандидозной фунгурии. Clin Infect Dis 1995; 21:960-5. [PubMed] [Академия Google]
105. Bennett WM, Craven R. Инфекции мочевыводящих путей у пациентов с тяжелой почечной недостаточностью. Лечение ампициллином и триметоприм-сульфаметоксазолом. ЯМА 1976; 236:946-8. [PubMed] [Google Scholar]
106. Кио Б., Раддок Г., Ирвин Дж., Уотсон А., Кин КТ. Лечение инфекции мочевыводящих путей цефуроксимом у пациентов с почечной недостаточностью. Ir Med J 1981; 74: 205-7. [PubMed] [Google Scholar]
107. Bailey RR, Peddie B, Blake E. Концентрация цефоперазона в сыворотке и моче при тяжелой хронической почечной недостаточности. Наркотики 1981;22(Приложение 1):46-51. [PubMed] [Google Scholar]
108. Westenfelder SR, Welling G, Madsen PO. Эффективность и фармакокинетика тобрамицина у пациентов с хроническими инфекциями мочевыводящих путей и различной степенью почечной недостаточности. Инфекция 1974; 2:76-9. [PubMed] [Google Scholar]
109. Spring PJ, Sharpe DM, Hayes MW. Нитрофурантоин и периферическая невропатия: забытая проблема? Med J Aust 2001; 174:153-4. [PubMed] [Google Scholar]
110. Маунтокалакис Т., Скунакис М., Целентис Дж. Краткосрочная и длительная системная антибиотикопрофилактика у пациентов, получающих постоянные катетеры. Дж Урол 1985;134:506-8. [PubMed] [Google Scholar]
111. Найрен П., Рунеберг Л., Костиала А.И., Ренконен О.В., Ройне Р. Профилактическое применение метенамина гиппурата или нитрофурантоина у пациентов с постоянным мочевым катетером. Энн Клин Рез 1981; 13:16-21. [PubMed] [Google Scholar]
112. Батлер Х.К., Кунин С.М. Оценка специфической системной антимикробной терапии у пациентов, находящихся на закрытом катетерном дренировании. Дж. Урол 1968; 100:567-72. [PubMed] [Google Scholar]
113. Gribble MJ, Puterman ML. Профилактика инфекции мочевыводящих путей у лиц с недавней травмой спинного мозга: проспективное рандомизированное двойное слепое плацебо-контролируемое исследование триметоприм-сульфаметоксазола. Am J Med 1993;95:141-52. [PubMed] [Google Scholar]
114. Райли Д.К., Классен Д.К., Стивенс Л.Е., Берк Дж.П. Большое рандомизированное клиническое исследование мочевого катетера, пропитанного серебром: отсутствие эффективности и стафилококковая суперинфекция. Am J Med 1995; 98: 349-56. [PubMed] [Google Scholar]
115. Linsenmeyer TA, Harrison B, Oakley A, Kirshblum S, Stock JA, Millis SR. Оценка добавки клюквы для снижения инфекций мочевыводящих путей у лиц с нейрогенным мочевым пузырем, вторичным по отношению к травме спинного мозга. Проспективное двойное слепое плацебо-контролируемое перекрестное исследование. J Медицина спинного мозга 2004; 27:29-34. [PubMed] [Google Scholar]
116. Cardenas DD, Hoffman JM, Kelly E, Mayo ME. Влияние образовательной программы по инфекциям мочевыводящих путей у лиц с травмой спинного мозга. J Spinal Cord Med 2004; 27:47-54. [PubMed] [Google Scholar]
117. Morton SC, Shekelle PG, Adams JL, et al. Антимикробная профилактика инфекции мочевыводящих путей у лиц с дисфункцией спинного мозга. Arch Phys Med Rehabil 2002; 83: 129-38. [PubMed] [Google Scholar]
118. Thompson RL, Haley CE, Searcy MA, et al. Катетерассоциированная бактериурия. Неспособность снизить частоту приступов с помощью периодических инстилляций дезинфицирующего средства в мочевыводящие системы. ДЖАМА 1984;251:747-51. [PubMed] [Google Scholar]
119. Gillespie WA, Simpson RA, Jones JE, Nashef L, Teasdale C, Speller DC. Предотвращает ли добавление дезинфицирующего средства в мочеприемники инфекции у катетеризированных пациентов? Ланцет 1983; 1:1037-9. [PubMed] [Google Scholar]
120. Леоне М., Гарнье Ф., Дубук М., Бимар М.С., Мартин С. Профилактика внутрибольничной инфекции мочевыводящих путей у пациентов в отделении интенсивной терапии: сравнение эффективности двух мочевых дренажных систем. Грудь 2001;120:220-4. [PubMed] [Академия Google]
121. Huth TS, Burke JP, Larsen RA, Classen DC, Stevens LE. Рандомизированное исследование ухода за полостью рта кремом с сульфадиазином серебра для профилактики катетер-ассоциированной бактериурии. J Infect Dis 1992;165:14-8. [PubMed] [Google Scholar]
122. Burke JP, Jacobson JA, Garibaldi RA, Conti MT, Alling DW. Оценка ежедневного ухода за полостью рта полиантибиотическими мазями в профилактике бактериурии, связанной с мочевым катетером. Дж. Урол 1983; 129:331-4. [PubMed] [Google Scholar]
123. Burke JP, Garibaldi RA, Britt MR, Jacobson JA, Conti M, Alling DW. Профилактика катетер-ассоциированных инфекций мочевыводящих путей. Эффективность ежедневных режимов ухода за мясом. Am J Med 1981;70:655-8. [PubMed] [Google Scholar]
124. Schierholz JM, Yucel N, Rump AF, Beuth J, Pulverer G. Противоинфекционные и антикоррозийные материалы — мифы и факты. Int J Antimicrob Agents 2002;19:511-6. [PubMed] [Google Scholar]
Развитие толерантности к опиоидам: фармакокинетическая/фармакодинамическая перспектива
1. Inturrisi C. E. Клиническая фармакология опиоидов при боли. клин. Дж. Боль. 2002;18(4 Приложение):S3–S13. [PubMed] [Google Scholar]
2. Всемирная организация здравоохранения. «Облегчение боли при раке», второе издание , Всемирная организация здравоохранения: Женева, Швейцария, 1996 г., стр. 1–69.
3. Р. Т. Джонс. Euphoria против концентрации кокаина в плазме после назального введения (20% раствор). В CN Chiang, RL Hawks (eds.), NIDA Research Monograph 99 (Результаты исследований курения веществ, вызывающих злоупотребление) , 1990, стр. 30–41.
4. Siegel S. Толерантность к морфиновым анальгетикам: ее ситуационная специфичность поддерживает павловскую модель кондиционирования. Наука. 1976;193(4250):323–325. [PubMed] [Google Scholar]
5. Ван Ри Дж. М., Герритс М. А., Вандершурен Л. Дж. Опиоиды, вознаграждение и зависимость: встреча биологии, психологии и медицины. Фармакол. Ред. 1999; 51 (2): 341–396. [PubMed] [Google Scholar]
6. Э. М. Росс и Т. П. Кенакин. Фармакодинамика: механизмы действия лекарств и взаимосвязь между концентрацией лекарства и эффектом. В JG Hardman и L.E. Limbird (eds.), Goodman and Gilman’s The Pharmacological Basis of Therapeutics , 10-е изд., McGraw Hill, 2001, стр. 31–43.
7. Гибальди М., Леви Г. Дозозависимое снижение фармакологических эффектов препаратов с линейными фармакокинетическими характеристиками. Дж. Фарм. науч. 1972; 61 (4): 567–569. [PubMed] [Google Scholar]
8. Бернард С., Невилл К. А., Нгуен А. Т., Флокхарт Д. А. Межэтнические различия в генетических полиморфизмах CYP2D6 в популяции США: клинические последствия. Онколог. 2006;11(2):126–135. [PubMed] [Google Scholar]
9. Desmeules J., Gascon M.P., Dayer P., Magistris M. Влияние факторов окружающей среды и генетики на обезболивание кодеином. Евро. Дж. Клин. Фармакол. 1991;41(1):23–26. [PubMed] [Google Scholar]
10. Берковиц Б. А. Связь фармакокинетики с фармакологической активностью: морфин, метадон и налоксон. клин. Фармакокинетика. 1976; 1 (3): 219–230. [PubMed] [Google Scholar]
11. Саве Дж. Высокие дозы морфина и метадона у больных раком. Клинические фармакокинетические аспекты перорального лечения. клин. Фармакокинетика. 1986;11(2):87–106. doi: 10.2165/00003088-198611020-00001. [PubMed] [CrossRef] [Академия Google]
12. Фредхейм О. М., Борхгревинк П. К., Клепстад П., Кааса С., Дейл О. Метадон длительного действия при хронической боли: пилотное исследование фармакокинетических аспектов. Евро. Дж. Боль. 2007;11(6):599–604. [PubMed] [Google Scholar]
13. Sawe J., Svensson J. O., Rane A. Метаболизм морфина у онкологических больных при увеличении пероральных доз — нет доказательств аутоиндукции или дозозависимости. бр. Дж. Клин. Фармакол. 1983;16(1):85–93. [Бесплатная статья PMC] [PubMed] [Google Scholar]
14. Dagenais C., Graff C.L., Pollack G.M. Вариабельная модуляция поглощения опиоидов мозгом P-гликопротеином у мышей. Биохим. Фармакол. 2004;67(2):269–276. [PubMed] [Google Scholar]
15. Matheny C.J., Lamb M.W., Brouwer; К. Р., Поллак Г. М. Фармакокинетические и фармакодинамические последствия модуляции P-гликопротеина. Фармакотерапия. 2001;21(7):778–796. [PubMed] [Google Scholar]
16. Кинг М., Су В., Чанг А., Цукерман А., Пастернак Г.В. Транспорт опиоидов из головного мозга на периферию с помощью Р-гликопротеина: периферические действия центральных препаратов. Нац. Неврологи. 2001;4(3):268–274. [PubMed] [Google Scholar]
17. Дж. К. Калвасс, Э. О. Олсон и Г. М. Поллак. Влияние Р-гликопротеина гематоэнцефалического барьера на проникновение в головной мозг и антиноцицептивные эффекты модельных опиоидов. ААПС J . 7 (S2) (2005).
18. Bauer B., Yang X., Hartz A.M., Olson E.R., Zhao R., Kalvass J.C., Pollack G.M., Miller D.S. -регуляция гликопротеина. Мол. Фармакол. 2006;70(4):1212–1219. [PubMed] [Google Scholar]
19. Hassan H. E., Myers A. L., Lee I. J., Coop A., Eddington N. D. Оксикодон индуцирует сверхэкспрессию P-гликопротеина (ABCB1) и влияет на распределение паклитаксела в тканях у крыс Sprague Dawley. Дж. Фарм. науч. 2007;96(9):2494–506. [Бесплатная статья PMC] [PubMed] [Google Scholar]
20. Aquilante C.L., Letrent S.P., Pollack G.M., Brouwer K.L. Повышение P-гликопротеина мозга у устойчивых к морфину крыс. Жизнь. науч. 2000;66(4):PL47–PL51. [PubMed] [Google Scholar]
21. Виттвер Э., Керн С. Э. Роль метаболитов морфина в обезболивании: концепции и противоречия. ААПС. Дж. 2006;8(2):E348–E352. [Бесплатная статья PMC] [PubMed] [Google Scholar]
22. Смит Г. Д., Смит М. Т. Морфин-3-глюкуронид: доказательства, подтверждающие его предполагаемую роль в развитии толерантности к антиноцицептивным эффектам морфина у крыс. Боль. 1995;62(1):51–60. [PubMed] [Google Scholar]
23. Gardmark M., Karlsson M.O., Jonsson F., Hammarlund-Udenaes M. Морфин-3-глюкуронид оказывает незначительное влияние на антиноцицепцию морфина. Фармакодинамическое моделирование. Дж. Фарм. науч. 1998;87(7):813–820. [PubMed] [Google Scholar]
24. Уэлле Д. М., Поллак Г. М. Влияние предшествующего воздействия морфина-3-глюкуронида на расположение морфина и антиноцицепцию. Биохим. Фармакол. 1997;53(10):1451–1457. [PubMed] [Google Scholar]
25. Смит М. Т. Нейровозбуждающие эффекты морфина и гидроморфона: доказательства причастности метаболитов 3-глюкуронида. клин. Эксп. Фармакол. Физиол. 2000;27(7):524–528. [PubMed] [Академия Google]
26. Bartlett S.E., Cramond T., Smith M.T. Возбуждающие эффекты морфина-3-глюкуронида ослабляются LY274614, конкурентным антагонистом рецептора NMDA, и мидазоламом, агонистом бензодиазепинового сайта в рецепторном комплексе ГАМК. Жизнь. науч. 1994;54(10):687–694. [PubMed] [Google Scholar]
27. Evans CJ, Keith D.E., Jr., Morrison H., Magendzo K., Edwards R.H. Клонирование дельта-опиоидного рецептора путем функциональной экспрессии. Наука. 1992; 258 (5090): 1952–1955. [PubMed] [Академия Google]
28. Chen Y., Mestek A., Liu J., Hurley J.A., Yu L. Молекулярное клонирование и функциональная экспрессия мю-опиоидного рецептора из мозга крысы. Мол. Фармакол. 1993;44(1):8–12. [PubMed] [Google Scholar]
29. Li S., Zhu J., Chen C., Chen YW, Deriel J.K., Ashby B., Liu-Chen L.Y. Молекулярное клонирование и экспрессия крысиного каппа-опиоидного рецептора. Биохим. Дж. 1993; 295 (часть 3): 629–633. [Бесплатная статья PMC] [PubMed] [Google Scholar]
30. Киффер Б. Л. Опиоиды: первые уроки нокаутированных мышей. Тенденции. Фармакол. науч. 1999;20(1):19–26. [PubMed] [Google Scholar]
31. Matthes H.W., Maldonado R., Simonin F., Valverde O., Slowe S., Kitchen I., Befort K., Dierich A., Le Meur M., Dolle P. , Tzavara E., Hanoune J., Roques B.P., Kieffer B.L. Потеря обезболивания, вызванного морфином, эффект вознаграждения и симптомы отмены у мышей, у которых отсутствует ген мю-опиоидного рецептора. Природа. 1996;383(6603):819–823. [PubMed] [Google Scholar]
32. Sora I., Takahashi N., Funada M., Ujike H., Revay R.S., Donovan D.M., Miner L.L., Uhl G.R. Мыши с нокаутом опиатных рецепторов определяют роли мю-рецепторов в эндогенных ноцицептивных реакциях. и морфин-индуцированная анальгезия. проц. Натл. акад. науч. США 1997; 94 (4): 1544–1549. [Бесплатная статья PMC] [PubMed] [Google Scholar]
33. Уильямс Дж. Т., Кристи М. Дж., Мандзони О. Клеточные и синаптические адаптации, опосредующие опиоидную зависимость. Физиол. 2001;81(1):299–343. [PubMed] [Google Scholar]
34. Цо П. Х., Вонг Ю. Х. Молекулярные основы опиоидной зависимости: роль регуляции сигналов G-белками. клин. Эксп. Фармакол. Физиол. 2003;30(5–6):307–316. [PubMed] [Google Scholar]
35. Smart D., Hirst R.A., Hirota K., Grandy D.K., Lambert D.G. Влияние активации рекомбинантного крысиного мю-опиоидного рецептора в клетках CHO на фосфолипазу C, [Ca2+]i и аденилил циклаза. бр. Дж. Фармакол. 1997;120(6):1165–1171. [PMC free article] [PubMed] [Google Scholar]
36. Нестлер Э. Дж., Агаджанян Г. К. Молекулярно-клеточные основы зависимости. Наука. 1997;278(5335):58–63. [PubMed] [Google Scholar]
37. Тейлор Д. А., Флеминг В. В. Объединение взглядов на механизмы, лежащие в основе развития толерантности и физической зависимости от опиоидов. Дж. Фармакол. Эксп. тер. 2001;297(1):11–18. [PubMed] [Google Scholar]
38. Zhang L., Yu Y., Mackin S., Weight F. F., Uhl G. R., Wang J. B. Дифференциальное фосфорилирование и десенсибилизация мю-опиатных рецепторов, вызванное агонистами и эфирами форбола. Дж. Биол. хим. 1996;271(19):11449–11454. [PubMed] [Google Scholar]
39. Yu Y., Zhang L., Yin X., Sun H., Uhl G.R., Wang JB. Фосфорилирование опиоидных рецепторов Mu, десенсибилизация и эффективность лиганда. Дж. Биол. хим. 1997;272(46):28869–28874. [PubMed] [Google Scholar]
40. Zhang J., Ferguson S.S., Barak L.S., Bodduluri S.R., Laporte S.A., Law P.Y., Caron M.G. Роль киназы рецептора, связанного с G-белком, в агонист-специфической регуляции мю-опиоидного рецептора ответная реакция. проц. Натл. акад. науч. США 1998;95(12):7157–7162. [PMC free article] [PubMed] [Google Scholar]
41. Бон Л. М., Гайнетдинов Р. Р., Лин Ф. Т., Лефковиц Р. Дж., Карон М. Г. Десенсибилизация мю-опиоидных рецепторов бета-аррестином-2 определяет толерантность к морфину, но не зависимость. Природа. 2000;408(6813):720–723. [PubMed] [Google Scholar]
42. Бон Л. М., Лефковиц Р. Дж., Гайнетдинов Р. Р., Пеппель К., Карон М. Г., Лин Ф. Т. Усиленная морфиновая анальгезия у мышей, лишенных бета-аррестина 2. Наука. 1999;286(5449):2495–2498. [PubMed] [Google Scholar]
43. Томпсон Д., Пуш М., Уистлер Дж. Л. Изменения в сродстве к рецепторам, связанным с G-белком, регулируют постэндоцитарное нацеливание на рецепторы, связанные с G-белком. Дж. Биол. хим. 2007;282(40):29178–29185. [PubMed] [Google Scholar]
44. Narita M., Suzuki M., Narita M., Niikura K., Nakamura A., Miyatake M., Yajima Y., Suzuki T. мю-опиоидные рецепторы, зависящие от интернализации и -независимые механизмы развития толерантности к агонистам мю-опиоидных рецепторов: сравнение эторфина и морфина. Неврология. 2006;138(2):609–619. [PubMed] [Google Scholar]
45. Данг В. К., Уильямс Дж. Т. Морфин-индуцированная десенсибилизация мю-опиоидных рецепторов. Мол. Фармакол. 2005;68(4):1127–1132. [PubMed] [Google Scholar]
46. Хе Л., Фонг Дж., фон Застров М., Уистлер Дж. Л. Регулирование транспорта опиоидных рецепторов и толерантности к морфину путем олигомеризации рецепторов. Клетка. 2002;108(2):271–282. [PubMed] [Google Scholar]
47. Джордан Б. А., Деви Л. А. Гетеродимеризация рецептора, связанного с G-белком, модулирует функцию рецептора. Природа. 1999;399(6737):697–700. [Бесплатная статья PMC] [PubMed] [Google Scholar]
48. Bailey C.P., Couch D., Johnson E., Griffiths K., Kelly E., Henderson G. Десенсибилизация мю-опиоидных рецепторов в нейронах зрелых крыс: отсутствие взаимодействие между ДАМГО и морфином. Дж. Нейроски. 2003;23(33):10515–10520. [Бесплатная статья PMC] [PubMed] [Google Scholar]
49. Stafford K., Gomes A.B., Shen J., Yoburn B.C. Понижающая регуляция мю-опиоидных рецепторов способствует толерантности к опиоидам in vivo . Фармакол. Биохим. Поведение 2001;69(1–2): 233–237. [PubMed] [Google Scholar]
50. Финн А. К., Уистлер Дж. Л. Эндоцитоз мю-опиоидного рецептора снижает толерантность и является клеточным признаком отмены опиатов. Нейрон. 2001;32(5):829–839. [PubMed] [Google Scholar]
51. Sim-Selley L.J., Scoggins K.L., Cassidy M.P., Smith L.A., Dewey W.L., Smith F.L., Selley D.E. Регионально-зависимое ослабление активации G-белка, опосредованной мю-опиоидным рецептором, в ЦНС мыши. в зависимости от толерантности к морфину. бр. Дж. Фармакол. 2007;151(8):1324–1333. [Бесплатная статья PMC] [PubMed] [Google Scholar]
52. Лафорж К. С., Юферов В., Крик М. Дж. Полиморфизмы опиоидных рецепторов и пептидных генов: потенциальные последствия для зависимостей. Евро. Дж. Фармакол. 2000;410(2–3):249–268. [PubMed] [Google Scholar]
53. Крик М.Дж., Барт Г., Лилли К., Лафорж К.С., Нильсен Д.А. Фармакогенетика и молекулярная генетика человека опиатной и кокаиновой зависимости и их лечение. Фармакол. 2005; 57(1):1–26. [PubMed] [Google Scholar]
54. Bond C., LaForge K.S., Tian M., Melia D., Zhang S., Borg L., Gong J., Schluger J., Strong J.A., Leal S.M., Tischfield J.A. , Крик М.Дж., Ю Л. Однонуклеотидный полиморфизм в гене мю-опиоидного рецептора человека изменяет связывание и активность бета-эндорфина: возможные последствия для опиатной зависимости. проц. Натл. акад. науч. США 1998;95(16):9608–9613. [Бесплатная статья PMC] [PubMed] [Google Scholar]
55. Surratt C.K., Johnson P.S., Moriwaki A., Seidleck B.K., Blaschak CJ, Wang JB, Uhl G.R. −mu опиатный рецептор. Аминокислоты с заряженными трансмембранными доменами имеют решающее значение для распознавания агонистов и внутренней активности. Дж. Биол. хим. 1994;269(32):20548–20553. [PubMed] [Google Scholar]
56. Romberg R.R., Olofsen E., Bijl H., Taschner P.E., Teppema L.J., Sarton E.Y., van Kleef J.W., Dahan A. Полиморфизм гена мю-опиоидного рецептора (OPRM1:c. 118A>G) не защищает от угнетения дыхания, вызванного опиоидами, несмотря на сниженный анальгетический ответ. Анестезиология. 2005;102(3):522–530. [PubMed] [Академия Google]
57. Oertel B.G., Schmidt R., Schneider A., Geisslinger G., Lotsch J. Полиморфизм гена мю-опиоидного рецептора 118A>G истощает анальгезию, индуцированную альфентанилом, и защищает от угнетения дыхания у гомозиготных носителей. Фармакогенет. Геномика. 2006;16(9):625–36. [PubMed] [Google Scholar]
58. Befort K., Filliol D., Decaillot F.M., Gaveriaux-Ruff C., Hoehe M.R., Kieffer B.L. Однонуклеотидная полиморфная мутация в мю-опиоидном рецепторе человека серьезно нарушает передачу сигналов рецептора. Дж. Биол. хим. 2001;276(5):3130–3137. [PubMed] [Академия Google]
59. Beyer A., Koch T., Schroder H., Schulz S., Hollt V. Влияние полиморфизма A118G на аффинность связывания, активность и опосредованный агонистом эндоцитоз, десенсибилизацию и ресенсибилизацию мю-опиоидного рецептора человека . Дж. Нейрохим. 2004;89(3):553–560. [PubMed] [Google Scholar]
60. Ван Д., Куиллан Дж. М., Винанс К., Лукас Дж. Л., Сэди В. Однонуклеотидные полиморфизмы в гене мю-опиоидного рецептора человека изменяют базальное связывание G-белка и связывание кальмодулина. Дж. Биол. хим. 2001;276(37):34624–34630. [PubMed] [Академия Google]
61. Millan M.J. Опосредованная каппа-опиоидными рецепторами антиноцицепция у крыс. I. Сравнительное действие мю- и каппа-опиоидов на вредные термические, сдавливающие и электрические раздражители. Дж. Фармакол. Эксп. тер. 1989;251(1):334–341. [PubMed] [Google Scholar]
62. Миллан М.Дж., Члонковски А., Липковски А., Герц А. Опосредованная каппа-опиоидными рецепторами антиноцицепция у крыс. II. Супраспинальные в дополнение к спинальным сайтам действия. Дж. Фармакол. Эксп. тер. 1989;251(1):342–350. [PubMed] [Академия Google]
63. Heyman J.S., Mulvaney S.A., Mosberg H.I., Porreca F. Участие опиоидных дельта-рецепторов в супраспинальной и спинальной антиноцицепции у мышей. Мозг. Рез. 1987; 420(1):100–108. [PubMed] [Google Scholar]
64. Виттерт Г., Хоуп П. , Пайл Д. Тканевое распределение экспрессии генов опиоидных рецепторов у крыс. Биохим. Биофиз. Рез. коммун. 1996;218(3):877–881. [PubMed] [Google Scholar]
65. Leighton G.E., Rodriguez R.E., Hill R.G., Hughes J. Агонисты каппа-опиоидов вызывают антиноцицепцию после внутривенного введения. и i.c.v. но не интратекальное введение крысе. бр. Дж. Фармакол. 1988;93(3):553–560. [Бесплатная статья PMC] [PubMed] [Google Scholar]
66. Нарита М., Хотиб Дж., Судзуки М., Одзаки С., Ядзима Ю., Судзуки Т. Адаптация гетерологичного мю-опиоидного рецептора путем повторной стимуляции каппа -опиоидный рецептор: активация G-белка и антиноцицепция. Дж. Нейрохим. 2003;85(5):1171–1179. [PubMed] [Google Scholar]
67. McLaughlin J. P., Myers L. C., Zarek P. E., Caron M. G., Lefkowitz R. J., Czyzyk T. A., Pintar J. E., Chavkin C. толерантность. Дж. Биол. хим. 2004;279(3): 1810–1818 гг. [PMC free article] [PubMed] [Google Scholar]
Агонист-специфическое подавление дельта-опиоидных рецепторов человека. Евро. Дж. Фармакол. 2003;459(1):9–16. [PubMed] [Google Scholar]
69. Lecoq I., Marie N., Jauzac P., Allouche S. Различная регуляция дельта-опиоидных рецепторов человека с помощью SNC-80 [(+)-4-[(alphaR)- альфа-((2S,5R)-4-аллил-2,5-диметил-1-пиперазинил)-3-метоксибензил]-N,N-диэтилбензамид] и эндогенные энкефалины. Дж. Фармакол. Эксп. тер. 2004;310(2):666–677. [PubMed] [Академия Google]
70. де Штутц Н. Д., Бруэра Э., Суарес-Алмазор М. Ротация опиоидов для снижения токсичности у больных раком в терминальной стадии. Дж. Боль. Симптом. Управлять. 1995;10(5):378–384. [PubMed] [Google Scholar]
71. Райли Дж., Росс Дж. Р., Раттер Д., Уэллс А. У., Голлер К., Дюбуа Р., Уэлш К. Морфин не облегчает боль? Индивидуальная вариабельность чувствительности к морфину и необходимость перехода на альтернативный опиоид у онкологических больных. Поддерживать. Уход. Рак. 2006;14(1):56–64. [PubMed] [Академия Google]
72. Athanasos P., Smith C.S., White J.M., Somogyi A.A., Bochner F. , Ling W. Пациенты, поддерживающие лечение метадоном, обладают перекрестной толерантностью к антиноцицептивным эффектам очень высоких концентраций морфина в плазме. Боль. 2006;120(3):267–275. [PubMed] [Google Scholar]
73. Duttaroy A., Yoburn B.C. Влияние внутренней эффективности на толерантность к опиоидам. Анестезиология. 1995;82(5):1226–1236. [PubMed] [Google Scholar]
74. Уокер Э. А., Ричардсон Т. М., Янг А. М. Толерантность и перекрестная толерантность к морфиноподобным стимулирующим эффектам мю-опиоидов у крыс. Психофармакология. (Берл) 1997;133(1):17–28. [PubMed] [Google Scholar]
75. Пастернак Г. В. Множественные опиатные рецепторы: снова дежавю. Нейрофармакология. 2004; 47 (Приложение 1): 312–323. [PubMed] [Google Scholar]
76. Гупта А., Декайо Ф. М., Деви Л. А. Ориентация на гетеродимеры опиоидных рецепторов: стратегии скрининга и разработки лекарств. ААПС. Дж. 2006;8(1):E153–E159. [Бесплатная статья PMC] [PubMed] [Google Scholar]
77. Зарриндаст М. Р., Алаей-Ниа К., Шафизаде М. О механизме толерантности к морфин-индуцированной реакции хвоста Штрауба у мышей. Фармакол. Биохим. Поведение 2001;69(3–4): 419–424. [PubMed] [Google Scholar]
78. Luccarini P., Perrier L., Degoulange C., Gaydier A.M., Dallel R. Синергический антиноцицептивный эффект амитриптилина и морфина в орофациальном формалиновом тесте крыс. Анестезиология. 2004; 100(3):690–696. [PubMed] [Google Scholar]
79. Sawynok J., Esser MJ, Reid A.R. Антидепрессанты как анальгетики: обзор центральных и периферических механизмов действия. Дж. Психиатрия. Неврологи. 2001;26(1):21–29. [Бесплатная статья PMC] [PubMed] [Google Scholar]
80. Грейси К. Н., Свингос А. Л., Пикель В. М. Двойная ультраструктурная локализация мю-опиоидных рецепторов и глутаматных рецепторов NMDA-типа в оболочке прилежащего ядра крысы. Дж. Нейроски. 1997;17(12):4839–4848. [Бесплатная статья PMC] [PubMed] [Google Scholar]
81. Commons KG, van Bockstaele EJ, Pfaff DW. Частая совместная локализация мю-опиоидных и глутаматных рецепторов NMDA-типа в постсинаптических участках периакведуктальных серых нейронов. Дж. Комп. Нейрол. 1999;408(4):549–559. [PubMed] [Академия Google]
82. Чжао М., Джу Д. Т. Субпопуляция нейронов задних рогов демонстрирует усиленную функцию рецептора N-метил-D-аспартата после хронического воздействия морфина. Анестезиология. 2006;104(4):815–825. [PubMed] [Google Scholar]
83. Снайдер С. Х. Опиатные рецепторы и не только: 30 лет исследований нейронной сигнализации. Нейрофармакология. 2004; 47 (Приложение 1): 274–285. [PubMed] [Google Scholar]
84. Колесников Ю. А., Пик С. Г., Цишевска Г., Пастернак Г. В. Блокада толерантности к морфину, но не к каппа-опиоидам, ингибитором синтазы оксида азота. проц. Натл. акад. науч. США 1993;90(11):5162–5166. [PMC free article] [PubMed] [Google Scholar]
85. Бабей А. М., Колесников Ю., Ченг Дж., Интурриси С. Э., Трифиллетти Р. Р., Пастернак Г. В. Оксид азота и толерантность к опиоидам. Нейрофармакология. 1994;33(11):1463–1470. [PubMed] [Google Scholar]
86. Кемп Дж. А., МакКернан Р. М. Пути рецепторов NMDA как мишени для лекарств. Нац. Неврологи. 2002; 5 (прил.): 1039–1042. [PubMed] [Google Scholar]
87. Герман Б. Х., Воччи Ф., Бридж П. Влияние антагонистов рецепторов NMDA и ингибиторов синтазы оксида азота на толерантность к опиоидам и синдром отмены. Проблемы разработки лекарственных препаратов для лечения опиоидной зависимости. Нейропсихофармакология. 1995;13(4):269–293. [PubMed] [Google Scholar]
88. Лауретти Г. Р., Лима И. К., Рейс М. П., Прадо В. А., Перейра Н. Л. Пероральный кетамин и трансдермальный нитроглицерин в качестве анальгетических адъювантов пероральной терапии морфином для лечения боли при раке. Анестезиология. 1999; 90(6):1528–1533. [PubMed] [Google Scholar]
89. Аллен Р. М., Дайкстра Л. А. Ослабление толерантности к мю-опиоидам и перекрестной толерантности с помощью конкурентного антагониста рецептора N-метил-D-аспартата LY235959 связано с величиной толерантности и перекрестной толерантности. Дж. Фармакол. Эксп. тер. 2000;295(3):1012–1021. [PubMed] [Google Scholar]
90. Фишер Б. Д., Карриган К. А., Дайкстра Л. А. Влияние антагонистов рецепторов N-метил-D-аспартата на острую антиноцицептивную реакцию, индуцированную морфином и метадоном, у мышей. Дж. Боль. 2005;6(7):425–433. [PubMed] [Google Scholar]
91. Медведев И. О., Малышкин А. А., Белозерцева И. В., Сухотина И. А., Севостьянова Н. Ю., Алиев К., Звартау Э. Э., Парсонс К. Г., Даныш В., Беспалов А. Ю. Эффекты канала низкоаффинного рецептора NMDA блокаторов в двух крысиных моделях хронической боли. Нейрофармакология. 2004;47(2):175–183. [PubMed] [Академия Google]
92. Suzuki M., Kinoshita T., Kikutani T., Yokoyama K., Inagi T., Sugimoto K., Haraguchi S., Hisayoshi T., Shimada Y. Определение концентрации кетамина в плазме, которая усиливает эпидуральное введение бупивакаина. и морфин-индуцированная анальгезия. Анест. аналг. 2005;101(3):777–784. [PubMed] [Google Scholar]
93. Sethna N. F., Liu M., Gracely R., Bennett G. J., Max M. B. Анальгетические и когнитивные эффекты внутривенных комбинаций кетамина и альфентанила по сравнению с любым препаратом в отдельности после внутрикожного введения капсаицина у нормальных субъектов. Анест. аналг. 1998;86(6):1250–1256. [PubMed] [Google Scholar]
94. Редвин К. Э., Трухильо К. А. Влияние антагонистов рецепторов NMDA на острую мю-опиоидную анальгезию у крыс. Фармакол. Биохим. Поведение 2003;76(2):361–372. [PubMed] [Google Scholar]
95. Willetts J., Balster R.L., Leander J.D. Поведенческая фармакология антагонистов рецепторов NMDA. Тренды Фармакол. науч. 1990;11(10):423–428. [PubMed] [Google Scholar]
96. Чен Х. С., Липтон С. А. Химическая биология клинически переносимых антагонистов рецепторов NMDA. Дж. Нейрохим. 2006;97 (6): 1611–1626. [PubMed] [Google Scholar]
97. Косман К. М., Бойл Л. Л., Порстейнссон А. П. Мемантин в лечении болезни Альцгеймера легкой и средней степени тяжести. Эксперт. мнение Фармацевт. 2007;8(2):203–214. [PubMed] [Google Scholar]
98. Coggeshall R.E., Zhou S., Carlton S.M. Опиоидные рецепторы на периферических сенсорных аксонах. Мозг. Рез. 1997; 764 (1–2): 126–132. [PubMed] [Google Scholar]
99. Carlton S. M., Zhou S., Coggeshall R.E. Доказательства взаимодействия рецепторов глутамата и NK1 на периферии. Мозг. Рез. 1998;790(1–2):160–169. [PubMed] [Google Scholar]
100. Carlton S.M., Hargett G.L., Coggeshall R.E. Локализация и активация глутаматных рецепторов в немиелинизированных аксонах гладкой кожи крысы. Неврологи. лат. 1995;197(1):25–28. [PubMed] [Google Scholar]
101. Carlton S.M., Coggeshall R.E. Вызванные воспалением изменения в популяциях периферических глутаматных рецепторов. Мозг. Рез. 1999;820(1–2):63–70. [PubMed] [Google Scholar]
102. Pol O., Puig M.M. Экспрессия опиоидных рецепторов при периферическом воспалении. Курс. Верхний. Мед. хим. 2004;4(1):51–61. [PubMed] [Академия Google]
103. Колесников Ю.А., Джайн С., Уилсон Р., Пастернак Г.В. Периферическая морфиновая анальгезия: синергия с центральными участками и цель толерантности к морфину. Дж. Фармакол. Эксп. тер. 1996;279(2):502–506. [PubMed] [Google Scholar]
104. Райхерт Дж. А., Дочери Р. С., Ривард Р., Симона Д. А. Периферическая и упреждающая опиоидная антиноцицепция на модели висцеральной боли у мышей. Боль. 2001;89(2–3):221–227. [PubMed] [Google Scholar]
105. Shannon H.E., Lutz E.A. Сравнение периферических и центральных эффектов опиоидных агонистов лоперамида и морфина в формалиновом тесте у крыс. Нейрофармакология. 2002;42(2):253–261. [PubMed] [Академия Google]
106. Колесников Ю., Кристеа М., Оксман Г., Торосян А., Уилсон Р. Оценка формалинового теста на хвосте у мышей как новой модели для оценки местного обезболивающего действия. Мозг. Рез. 2004;1029(2):217–223. [PubMed] [Google Scholar]
107. Гардмарк М., Бринн Л., Хаммарлунд-Уденаес М., Карлссон М. О. Взаимозаменяемость и прогностическая эффективность эмпирических моделей толерантности. клин. Фармакокинетика. 1999;36(2):145–167. [PubMed] [Google Scholar]
108. Porchet H.C., Benowitz N.L., Sheiner L.B. Фармакодинамическая модель толерантности: приложение к никотину. Дж. Фармакол. Эксп. тер. 1988;244(1):231–236. [PubMed] [Google Scholar]
109. Ouellet D.M., Pollack G.M. Фармакодинамика и развитие толерантности при многократном внутривенном болюсном введении морфина крысам. Дж. Фармакол. Эксп. тер. 1997;281(2):713–720. [PubMed] [Google Scholar]
110. Lutfy K., Hurlbut D.E., Weber E. Блокада индуцированной морфином анальгезии и толерантности у мышей с помощью MK-801. Мозг. Рез. 1993;616(1–2):83–88. [PubMed] [Google Scholar]
111. Бхаргава Х. Н., Шарма С. С., Биан Дж. Т. Доказательства роли рецепторов N-метил-D-аспартата в индуцированном L-аргинином ослаблении антиноцицептивного действия морфина. Мозг. Рез. 1998;782(1–2):314–317. [PubMed] [Google Scholar]
112. Бхаргава Х. Н., Биан Дж. Т., Кумар С. Механизм ослабления антиноцицептивного действия морфина при хроническом лечении L-аргинином. Дж. Фармакол. Эксп. тер. 1997;281(2):707–712. [PubMed] [Google Scholar]
113. Хайнцен Э. Л., Поллак Г. М. Развитие антиноцицептивной толерантности к морфину у мышей с дефицитом синтазы оксида азота. Биохим. Фармакол. 2004;67(4):735–741. [PubMed] [Google Scholar]
114. Хайнцен Э. Л., Поллак Г. М. Использование электрохимического датчика оксида азота для обнаружения выработки оксида азота нейронами у находящихся в сознании крыс без ограничений. Дж. Фармакол. Токсикол. Методы. 2002;48(3):139–146. [PubMed] [Google Scholar]
115. Хайнцен Э. Л., Поллак Г. М. Фармакокинетика и фармакодинамика L-аргинина у крыс: модель стимулированного нейронального синтеза оксида азота. Мозг. Рез. 2003;989(1):67–75. [PubMed] [Google Scholar]
116. Хайнцен Э. Л., Поллак Г. М. Фармакодинамика индуцированной морфином продукции нейронов оксида азота и развития антиноцицептивной толерантности. Мозг. Рез. 2004;1023(2):175–184. [PubMed] [Google Scholar]
117. Хайнцен Э. Л., Бут Р. Г., Поллак Г. М. Нейрональный оксид азота модулирует антиноцицептивную толерантность к морфину за счет усиления конститутивной активности мю-опиоидного рецептора. Биохим. Фармакол. 2005;69(4): 679–688. [PubMed] [Google Scholar]
118. Ouellet D.M., Pollack G.M. Фармакокинетико-фармакодинамическая модель толерантности к анальгезии морфина во время инфузии у крыс. Дж. Фармакокинетика. Биофарм. 1995;23(6):531–549. [PubMed] [Google Scholar]
Причины, типы, симптомы и диагностика
Человек с пищевой непереносимостью испытывает трудности с перевариванием определенных продуктов. Важно отметить, что пищевая непереносимость отличается от пищевой аллергии.
Часто встречается пищевая непереносимость. По некоторым оценкам, они могут поражать 15–20% населения.
Пищевая непереносимость чаще встречается у людей с расстройствами пищеварительной системы, такими как синдром раздраженного кишечника (СРК). По данным сети IBS, большинство людей с IBS имеют пищевую непереносимость.
В то время как пищевая аллергия возникает в результате реакции иммунной системы на определенный продукт, пищевая непереносимость обычно затрагивает пищеварительную систему, а не иммунную систему.
В этой статье рассматриваются причины, типы, симптомы и диагностика пищевой непереносимости, а также объясняется, как с ней справиться.
Человек с пищевой непереносимостью часто испытывает дискомфорт вскоре после употребления определенных продуктов. Симптомы разнообразны и обычно связаны с пищеварительной системой.
Common symptoms of food intolerance include:
- bloating
- excess gas
- stomach pain
- diarrhea
- migraine
- headaches
- a runny nose
- malaise, which is a general feeling of being under the weather
У людей с пищевой непереносимостью количество съеденной пищи определяет тяжесть симптомов.
Симптомы пищевой непереносимости могут проявиться через некоторое время. Начало может произойти через несколько часов после приема пищи, а симптомы могут сохраняться в течение нескольких часов или дней.
Может быть трудно определить, есть ли у кого-то пищевая непереносимость или аллергия, поскольку признаки и симптомы этих состояний совпадают.
Пищевая непереносимость возникает, если организм не может переваривать определенную пищу. Это нарушение может быть связано с недостатком пищеварительных ферментов или чувствительностью к определенным химическим веществам.
Пищевые продукты, обычно вызывающие пищевую непереносимость, включают:
- молоко
- глютен
- пищевые красители и консерванты
- сульфиты
- другие соединения, такие как кофеин и фруктоза. пищевая непереносимость.
Непереносимость лактозы
Организм использует пищеварительные ферменты для расщепления пищи. Если человеку не хватает определенных ферментов, он может быть менее способен переваривать определенные продукты.
Лактоза — это сахар, содержащийся в молоке. Людям с непереносимостью лактозы не хватает лактазы — фермента, расщепляющего лактозу на более мелкие молекулы, которые организм может легко усваивать через кишечник.
Если лактоза остается в пищеварительном тракте, это может вызвать спазмы, боль в животе, вздутие живота, диарею и газы.
Непереносимость фруктозы
Фруктоза — это сахар, содержащийся во фруктах, некоторых овощах и меде.
Непереносимость фруктозы также может быть связана с отсутствием фермента, хотя это бывает редко. В таких случаях говорят о наследственной непереносимости фруктозы. Мальабсорбция фруктозы, при которой организму не хватает белка, позволяющего ему поглощать сахар из кишечника, встречается гораздо чаще.
У этих людей фруктоза в пищевых продуктах ферментируется в кишечнике, что приводит к газообразованию, полноте, вздутию живота, судорогам и диарее.
Непереносимость глютена
Глютен — это белок, содержащийся в некоторых злаках, включая пшеницу, ячмень и рожь. Человек с непереносимостью глютена испытывает дискомфорт, такой как боль, вздутие живота или тошнота, после употребления продуктов, содержащих глютен.
Gluten intolerance is also associated with nondigestive symptoms, such as:
- brain fog
- headaches
- joint pain
- fatigue
- depression
- anxiety
- a general lack of well-being
Gluten intolerance is отличается от глютеновой болезни, которая является реакцией аутоиммунной системы на глютен, и аллергии на пшеницу, которая является аллергической реакцией на пшеницу.
Однако симптомы этих состояний могут быть схожими. Непереносимость глютена также известна как непереносимость глютена без целиакии.
Симптомы глютеновой болезни и непереносимости глютена обычно улучшаются, когда человек исключает глютен из рациона, но возвращаются, когда он снова вводит его.
Непереносимость салицилатов
Салицилаты представляют собой соединения, которые встречаются во многих растительных продуктах, включая фрукты, овощи, травы и специи. Они также распространены в искусственных ароматизаторах и консервантах, в том числе в зубной пасте, жевательной резинке и конфетах.
Большинство людей могут переносить умеренное количество салицилатов в своем рационе, но у некоторых людей толерантность снижена.
Symptoms can include:
- hives
- rashes
- stomach pain
- diarrhea
- fatigue
- a runny nose
- wheezing
Food poisoning
Some foods have naturally occurring chemicals that have a toxic effect on человека, вызывая диарею, тошноту, сыпь и рвоту.
Например, недоваренные бобы содержат афлатоксины, которые могут вызвать очень неприятные проблемы с пищеварением. Полностью приготовленные бобы не содержат токсина. Поскольку время приготовления может повлиять на количество токсинов, люди могут обнаружить, что они реагируют на бобы после одного приема пищи, но не после другого.
Точно так же употребление в пищу некоторых видов испорченной рыбы может привести к отравлению скумброидной рыбой. Эта токсическая реакция возникает из-за употребления в пищу рыбы с высоким содержанием гистамина в результате неправильного хранения или обработки. Он может имитировать сильную аллергическую реакцию.
Многие люди обеспокоены тем, что у них может быть непереносимость пищевых добавок. Однако, по данным Американского фонда астмы и аллергии, большинство исследований показывают, что некоторые добавки вызывают проблемы и что эти проблемы затрагивают относительно небольшое количество людей.
Производители продуктов питания часто используют добавки для улучшения вкуса, придания продуктам более привлекательного вида и увеличения срока их хранения.
Примеры пищевых добавок включают: - антиоксиданты
- искусственные красители
- искусственные ароматизаторы
- эмульгаторы
- усилители вкуса
- консерванты
- подсластители
Эксперты считают, что из тысяч добавок, используемых в пищевой промышленности, лишь относительно небольшое количество вызывает проблемы. Следующие пищевые добавки могут вызывать побочные реакции у некоторых людей:
- Нитраты: Эти консерванты распространены в переработанном мясе, а симптомы непереносимости могут включать головные боли и крапивницу.
- Глутамат натрия (MSG): Этот усилитель вкуса может вызывать головную боль, стеснение в груди, тошноту и диарею у людей с непереносимостью.
- Сульфиты: Обычные источники этих консервантов включают вино, сухофрукты, свежие креветки, а также некоторые джемы и желе. Люди с непереносимостью могут испытывать стеснение в груди, крапивницу, диарею, а иногда и анафилаксию.
Пищевая непереносимость и пищевая аллергия могут иметь схожие симптомы, поэтому диагностировать эти состояния может быть сложно. Диагностика также усложняется, когда у человека несколько пищевых непереносимостей.
Симптомы пищевой непереносимости могут также имитировать симптомы хронических заболеваний пищеварительного тракта, таких как СРК. Однако определенные закономерности в симптомах могут помочь врачу различить их.
Люди могут вести дневник питания, чтобы записывать, какие продукты они едят, какие симптомы проявляются и когда. Эти данные могут помочь людям и их лечащим врачам выяснить, какие продукты вызывают побочные реакции.
Помимо непереносимости лактозы и глютеновой болезни, не существует точных, надежных и проверенных тестов для выявления пищевой непереносимости. Лучшим диагностическим инструментом является исключающая диета, также известная как элиминационная или диагностическая диета.
Врач может порекомендовать сделать кожную пробу или анализ крови, чтобы исключить пищевую аллергию.
Для проведения прик-теста медицинский работник кладет небольшое количество пищи на спину или предплечье пациента и прокалывает кожу иглой. Кожная реакция указывает на наличие аллергии.
Анализ крови позволяет определить уровни антител иммуноглобулина Е (IgE). Высокий уровень может указывать на аллергию.
Некоторые люди обнаруживают, что если они какое-то время воздерживаются от определенной пищи, то не реагируют на нее снова. Это известно как толерантность. Поддержание толерантности часто зависит от того, как долго воздерживаться от употребления пищи и сколько съедать при ее повторном введении. Медицинский работник может помочь.
Эти состояния имеют некоторые общие симптомы. Однако симптомы пищевой непереносимости, как правило, проявляются дольше, чем симптомы пищевой аллергии.
В то время как пищевая аллергия возникает в результате реакции иммунной системы на определенный продукт, пищевая непереносимость затрагивает пищеварительную систему, а не иммунную систему.
При пищевой непереносимости количество съедаемой пищи связано с тяжестью симптомов. С другой стороны, при пищевой аллергии даже небольшое количество пищи может вызвать сильную аллергическую реакцию.
Узнайте больше о пищевой непереносимости и пищевой аллергии здесь.
Пищевая непереносимость возникает, когда организм не может правильно переваривать определенные продукты. Непереносимость лактозы и непереносимость глютена являются распространенными типами.
В настоящее время нет лекарства от пищевой непереносимости. Лучший способ избежать симптомов пищевой непереносимости — избегать определенных продуктов или есть их реже и в меньших количествах. Люди также могут принимать добавки для улучшения пищеварения.
Эффективное нацеливание белкового антигена на рецептор дендритных клеток DEC-205 в стабильном состоянии приводит к презентации антигена на продуктах класса I главного комплекса гистосовместимости и толерантности периферических CD8+ Т-клеток | Журнал экспериментальной медицины
Пропустить пункт назначения навигацииСтатья| 16 декабря 2002 г.
Лаура Бонифаз,
Дэвид Бонняй,
Карстен Манке,
Мигель Ривера,
Мишель К. Нуссенцвейг,
Ральф М. Штейнман
Информация об авторе и статье
Адресная переписка с Ральфом М. Стейнманом, Лаборатория клеточной физиологии и иммунологии, Университет Рокфеллера, 1230 York Ave., Box 176, New York, NY 10021. Телефон: 212-327-8106; Факс: 212-327-8875; Электронная почта: [email protected]
Л. Бонифаз, Д. Бонняй и К. Манке внесли равный вклад в эту работу.
*
Сокращения, используемые в данном документе: CFSE, сукцинимидиловый эфир карбоксифлуоресцеина диацетата; ДК, дендритная клетка.
Полученный: 10 сентября 2002 г.
Полученная редакция: 20 октября 2002 г.
Принято: 12 ноября 2002 г.
Интернет номер: 1540-9538
Издательство печати: 0022-1007
The Rockefeller University Press
2002
J Exp Med (2002) 196 (12): 1627–1638.
https://doi.org/10.1084/jem.20021598
История статьи
Получено:
10 сентября 2002 г.
Пересмотрено:
20 октября 2002 г.
Принято:
12 ноября 2002 г. Стандартный вид 906:40
-
Просмотры
- Содержание артикула
- Рисунки и таблицы
- Видео
- Аудио
- Дополнительные данные
- Экспертная оценка
-
Делиться
- MailTo
- Твиттер
-
Инструменты
-
Получить разрешения
-
Иконка Цитировать Цитировать
-
- Поиск по сайту
Citation
Лаура Бонифаз, Дэвид Бонняй, Карстен Манке, Мигель Ривера, Мишель К. Нуссенцвейг, Ральф М. Штейнман; Эффективное нацеливание белкового антигена на рецептор дендритных клеток DEC-205 в стабильном состоянии приводит к презентации антигена на продуктах главного комплекса гистосовместимости класса I и периферических CD8 + Толерантность Т-клеток . J Exp Med 16 декабря 2002 г.; 196 (12): 1627–1638. doi: https://doi.org/10.1084/jem.20021598
Скачать файл цитаты:
- Рис (Зотеро)
- Менеджер ссылок
- EasyBib
- Подставки для книг
- Менделей
- Бумаги
- Конечная примечание
- RefWorks
- Бибтекс
Расширенный поиск
Чтобы идентифицировать эндоцитарные рецепторы, которые позволяют дендритным клеткам (ДК) захватывать и представлять антигены на продуктах главного комплекса гистосовместимости (МНС) класса I in vivo, мы оценили DEC-205, который в изобилии присутствует на ДК в лимфоидных тканях. Белок овальбумин (OVA) при химическом соединении с моноклональным антителом αDEC-205 был представлен CD11c + ДК лимфатических узлов, но не CD11c — клеток, к OVA-специфическим, CD4 + и CD8 + Т-лимфоцитам. Представление, опосредованное рецептором, было по крайней мере в 400 раз более эффективным, чем неконъюгированный OVA, и для MHC класса I DCs должны были экспрессировать переносчики транспортеров антигенных пептидов (TAP). Когда αDEC-205:OVA вводили подкожно, белок OVA идентифицировали в течение 4–48 ч в ДК, преимущественно в лимфатических узлах, дренирующих место инъекции. In vivo белок OVA был селективно представлен DCs трансгенному CD8 9 TCR.0939 + клеток, опять же по меньшей мере в 400 раз более эффективно, чем растворимый OVA, и ТАР-зависимым образом. Нацеливание αDEC-205:OVA на ДК в стационарном состоянии первоначально индуцировало 4–7 циклов деления Т-клеток, но затем Т-клетки удалялись, и мыши становились специфически невосприимчивыми к повторному заражению OVA в полном адъюванте Фрейнда. Напротив, одновременная доставка стимула созревания DC через CD40 вместе с αDEC-205:OVA индуцировала сильный иммунитет. CD8 + Т-клетки, реагирующие в присутствии агонистического антитела αCD40, продуцировали большое количество интерлейкина 2 и интерферона γ, приобретали цитолитическую функцию in vivo, эмигрировали в больших количествах в легкие и энергично реагировали на повторное введение OVA. Следовательно, DEC-205 обеспечивает эффективный механизм на основе рецептора для DCs для процессинга белков для презентации MHC класса I in vivo, что приводит к толерантности в устойчивом состоянии и иммунитету после созревания DC.
Иммунная система сопротивляется патогенам, оставаясь терпимой к собственным антигенам и безвредным белкам в окружающей среде. Иммунная толерантность может поддерживаться как центральными, так и периферическими механизмами. Центральная толерантность к Т-клеткам устанавливается в тимусе, где аутореактивные Т-клетки могут делетироваться (1–3), например, после встречи с антигеном на поверхности медуллярных дендритных клеток (ДК) тимуса * (4–3). 6). Механизмы периферической толерантности необходимы в ситуациях, когда центральная толерантность является неполной, а также к антигенам, которые не попадают в тимус, например, к белкам окружающей среды, обычно обнаруживаемым на поверхностях тела, таких как дыхательные пути и кишечник (обзоры см. в ссылках 7–10). ДК на периферии непрерывно захватывают белки из дыхательных путей и кишечника (11, 12) и анатомически расположены для представления этих антигенов Т-клеткам в лимфоидных органах (13, 14). Хорошо изученная модель включает презентацию антигенов инсулин-продуцирующих β-клеток островков поджелудочной железы на продуктах MHC класса I и II дендритных клеток в лимфатических узлах поджелудочной железы (15–18). Однако экспериментальная индукция периферической толерантности к белкам, представленным MHC класса I, обычно требует введения больших количеств препроцессированных пептидов, а не небольших количеств интактных белков (19).). Эффективные молекулярные пути для развития толерантности к белкам окружающей среды и собственным белкам, присутствующим в продуктах MHC класса I, еще предстоит определить.
ДК имеют ряд рецепторов для адсорбционного захвата антигенов. Некоторые из них являются общими с другими клетками, например, рецепторы Fcγ (20–23) и маннозный рецептор макрофагов (24). Другие рецепторы более ограничены ДК, например, лангерин или CD207 (25), DC-SIGN или CD209 (26, 27), асиалогликопротеиновый рецептор (28) и BDCA-2 (29), но в настоящее время эти рецепторы имеют не было показано, что он опосредует избирательную презентацию антигенов на продуктах MHC класса I и II DCs in vivo. Напротив, DEC-205, эндоцитарный рецептор с 10 наружными от мембраны смежными лектиновыми доменами С-типа (30, 31), опосредует эффективный процессинг и презентацию антигенов на продуктах МНС класса II in vivo (32). DEC-205 экспрессируется на высоких уровнях на ДК в областях Т-клеток лимфоидных органов (13, 33), и доступны антитела αDEC-205, которые избирательно воздействуют на эти ДК у мышей (32). Здесь мы химически связываем белок с αDEC-205 и демонстрируем способность этого рецептора доставлять эндоцитированный белок в отсутствие инфекции к TAP-зависимому пути загрузки пептида MHC класса I. Небольшие количества инъецируемого антигена, нацеленного на ДК посредством адсорбционного пути DEC-205, способны индуцировать солидный периферический CD8 9.0939 + Толерантность Т-клеток. Эти результаты идентифицируют эффективный механизм на основе рецептора для DC, чтобы постоянно захватывать антигены для представления на продуктах MHC класса I в устойчивом состоянии и блокировать развитие реактивности CD8 + T-клеток.
Антитела.
Антитела к CD45.1 (A20) и другим маркерам клеточной поверхности (CD8α/53–6.7, CD62L/MEL-14, CD80/16–10A1, CD86/GL1, MHC class II I-A b /AF6–120.1) включая специфичность TCR (Vβ 5,1/5,2 /MR9–4; Vα 2 /B20.1) были приобретены у BD Biosciences. Гибридомы, продуцирующие антитела, были получены из Американской коллекции типов культур, включая DEC-205/CD205 (NLDC-145 HB 290), GL-117 (нереактивный крысиный изотип IgG2a, соответствующий DEC-205), CD107a/LAMP-1, MHC класса II. (M5/114, TIB120), F4/80 (HB 198), NK1.1, B220 (RA3–6B2), CD4 (GK1.5, TIB 207), CD8 (53.6.7 TIB 105) и CD11c (N418 , НВ224). Агонистичное mAb αCD40 FGK45.5 было предоставлено доктором Т. Ролинком (Базельский институт иммунологии, Базель, Швейцария). Магнитные микрогранулы были получены от Miltenyi Biotec.
Мыши. Мыши
C57BL/6 (B6) были получены от Taconic, а мыши TAP -/- — от The Jackson Laboratory. Мыши OT-I были предоставлены доктором Ф. Карбоне (Университет Мельбурна, Парквилл, Виктория, Австралия), а мыши OT-II были получены из Института Трюдо. Мыши CD45.1 + OT-I были получены путем скрещивания мышей OT-I с мышами B6.SJL-Ptprc (The Jackson Laboratory).
Конъюгация OVA с моноклональными антителами.
Очищенные IgG были конъюгированы с OVA, активированным малеимидом (Pierce Chemical Co.) или без LPS OVA (Seikagaku Corporation), активированным Sulfo-SMCC (сульфосукцинимидил 4-(N-малеимидометил)циклогексан-1-карбоксилат; Pierce Chemical Co. ) с использованием протокола производителя. Конъюгаты антитело:OVA, полученные с OVA из обоих источников, давали сходные результаты в отношении нацеливания на ДК и ответов Т-клеток in vivo. Хотя трудно исключить некоторую контаминацию ЛПС в наших препаратах, ДК у мышей, которым вводили конъюгаты антител, не демонстрировали признаков созревания, как это происходит, когда мышам вводят ЛПС. Для связывания антитела восстанавливали с использованием 2-меркаптоэтаноламина:HCl и отделяли от восстанавливающего агента на обессоливающей колонке. Затем активированный малеимидом OVA смешивали с восстановленными антителами в течение ночи при 4°С. Конъюгаты антитело:OVA пропускали через колонку с белком G для удаления неконъюгированного OVA, концентрировали на спин-колонках (Millipore) и оценивали с помощью спектрофотометрии и SDS-PAGE.
Выделение ДК и антиген-специфических Т-клеток.
Подколенные, паховые, подмышечные и плечевые ЛУ удаляли у взрослых самок мышей B6 или TAP -/- . Суспензии отдельных клеток готовили с использованием 400 ед/мл коллагеназы в течение 25 мин (Roche). Клетки инкубировали с анти-мышиными CD11c или CD19 микробусинами MACS ® в течение 30 мин на льду. CD11c + (обогащенные ДК), CD19 + (обогащенные В-клетками) и двойные негативные клетки разделяли с помощью магнитного поля и распределяли в 48-луночные планшеты при 5 × 10 5 клеток/лунку. Клетки культивировали с антигеном в течение ночи (16–20 ч) при 37°C и дважды промывали в PBS перед использованием в качестве антигенпрезентирующих клеток для наивных трансгенных TCR, OVA-специфических Т-клеток. ДК костного мозга готовили с GM-CSF, как описано ранее (34). Через 6 дней после забора костного мозга DC удаляли, промывали и подвергали импульсной обработке конъюгатом антитело:OVA (10 мкг/мл) в течение 6 часов перед кокультивированием с Т-клетками. Т-клетки CD8 + или CD4 + получали из мышей OT-I или OT-II соответственно. Т-клетки OT-I и OT-II очищали из суспензий одиночных клеток лимфатических узлов или клеток селезенки путем негативной селекции с использованием супернатантов гибридом (выше), направленных против MHC-II, F4/80, B220, NK 1. 1 и CD4 или CD8 и козы против крыс Dynabeads ® (Dynal) в соотношении 4 гранулы на 1 клетку-мишень. Культуральная среда представляла собой RPMI, 7% FBS, 100 ед/мл пенициллин-стрептомициновой смеси, 0,25 мг/мл фунгизона, 10 мМ HEPES и 55 мкМ β-меркаптоэтанола. Для идентификации клеток OT-I в легких готовили суспензии отдельных клеток путем промывания сосудистой системы легкого 10 мл PBS и 10 ЕД/мл гепарина (Elkins-Sinn) через правый желудочек и диссоциации ткани в 70 мкм Cell Strainer ® (Becton Dickinson).
Захват антител in vivo: конъюгаты OVA.
B6 или TAP -/- мышам вводили градуированные дозы конъюгатов IgG:OVA или растворимого OVA в лапы подкожно. Лимфатические узлы (подколенные, паховые, подмышечные и плечевые) собирали через 1–4 дня и выделяли ДК (как указано выше) для тестирования презентации антигена клеткам OT-I in vitro. 10 5 Т-клеток добавляли к градуированным дозам CD11c + DC, CD11c — неDC или CD11c — CD5 — В-клеток в круглодонных 96-луночных планшетах. [ 3 Н]тимидин (1 мкКи; Amersham Biosciences) добавляли через 48–72 ч для обнаружения включения в ДНК. Показанные данные являются средними значениями для трех повторностей, где стандартное отклонение составляло <10% от среднего числа импульсов в минуту. Для обнаружения OVA клетки CD11c + или CD11c — промывали, лизировали в буфере для образцов SDS для SDS-PAGE, а затем аликвоты, соответствующие 10 6 клеток, переносили на мембраны PVDF (Hybond-P; Amersham Biosciences). Для иммуноблоттинга мы использовали конъюгированные с HRP поликлональные кроличьи анти-OVA (Research Diagnostics, Inc.), разведенные в Roti-Block 9.0939® (Carl Roth), визуализированный с помощью ECL ® (Amersham Biosciences).
Проточная цитометрия.
Многоцветная проточная цитометрия использовалась для наблюдения за тремя функциональными ответами, как описано в следующих разделах: пролиферация Т-клеток, меченных карбоксифлуоресцеиндиацетат-сукцинимидиловым эфиром (CFSE), оцениваемая по постепенному уменьшению вдвое количества флуоресценции на клетку; созревание ДК, оцениваемое по активизации поверхностных антигенов, таких как CD80, CD86 и MHC II; и индукция эффекторной функции Т-клеток, т. е. накопление внутриклеточных цитокинов и проявление активности ЦТЛ. Мы использовали FACSCalibur™ (BD Biosciences) с последующим анализом данных в CELLQuest™ (BD Biosciences) или FlowJo 9.0939 ® (Звезда Дерева).
Мечение Т-клеток с помощью CFSE для определения пролиферативных ответов Т-клеток in vivo.
Т-клетки OT-I при 10 7 клеток/мл инкубировали с CFSE (Molecular Probes; 5 мкМ) в течение 10 мин при 37°C. Добавляли равный объем FCS и клетки дважды промывали PBS/0,1% BSA и один раз PBS. 2 × 10 6 меченых OT-I Т-клеток вводили внутривенно реципиентам B6. Через 24 ч конъюгаты антитело:OVA инъецировали во все 4 лапы с агонистическим антителом αCD40 или без него, 100 мкг подкожно. Через 3 дня суспензии лимфатических узлов окрашивали на Vβ 9.0999 5,1/5,2 , Vα 2 и CD8 и оценивали с помощью многоцветной проточной цитометрии.
Созревание DC in vivo.
100 мкг αCD40 вводили подкожно мышам B6, которые получали или не получали конъюгаты антитело:OVA и Т-клетки OT-I. Через 1–3 дня клетки лимфоузла CD11c + выделяли путем окрашивания PE-конъюгированным αCD11c и разделения с помощью анти-PE-микрогранул и MACS ® . Клетки CD11c + окрашивали на CD80, CD86 или I-A b (MHC II) и оценивали с помощью проточной цитометрии. В некоторых экспериментах клетки CD11c + были дважды помечены биотином-αDEC-205, чтобы следовать субпопуляциям DC с высокой и низкой экспрессией DEC-205.
Эффекторные ответы Т-клеток in vivo.
10 6 CD45.1 + Т-клетки OT-I очищали (выше) и вводили внутривенно мышам B6. Через 24 часа вводили инъекцией конъюгаты антитело:OVA с αCD40 или без него. Через 3 и 12 дней суспензии лимфатических узлов окрашивали на поверхностные CD45.1, Vβ 5.1/5.2 , CD62L, внутриклеточный IL-2 (JES6–5h5) или IFN-γ (XMG1.2). 5 × 10 6 клеток лимфатических узлов подвергали пульсации в 24-луночных чашках с родственным пептидом OT-I (SIINFEKL) в течение 5 часов в присутствии брефельдина А (Sigma-Aldrich). Затем клетки собирали, дважды промывали PBS/2% FBS и окрашивали на внеклеточный CD45.1 и Vβ 5,1/5,2 . Затем эти клетки фиксировали и окрашивали на цитокины с помощью стартового набора для окрашивания внутриклеточных цитокинов BD Biosciences в соответствии с протоколом производителя.
Анализы ЦТЛ in vivo проводили, как описано ранее (35), путем инъекции смесей 1:1 сингенных спленоцитов с пептидным импульсом и без него (3 × 10 6 каждый), меченных 5 мкМ (CFSE hi ) и 0,5 мкМ (CFSE lo ) CFSE соответственно, как описано выше. Через 12 ч суспензии одиночных клеток из лимфатических узлов оценивали с помощью проточной цитометрии. Специфическое умерщвление оценивали по сокращению популяции CFSE hi без какого-либо сокращения CFSE 9.популяции 0939- по сравнению с контрольными мышами, у которых отсутствовали Т-клетки OT-I.
Тесты на иммунную толерантность.
Мышам вводили Т-клетки OT-I и конъюгаты IgG:OVA с αCD40 или без него (см. выше). Через 12 дней после инъекции антигена мышам повторно вводили белок OVA (50 мкг подкожно; Calbiochem), суспендированный в полном адъюванте Фрейнда (Difco). Через 3 дня мышей умерщвляли и собирали лимфатические узлы либо для повторной стимуляции in vitro и внутриклеточного окрашивания IL-2 и IFN-γ, либо для анализа ЦТЛ in vivo (см. выше).
Нацеливание белка на DC In Situ через DEC-205.
Чтобы изучить потенциальную роль эндоцитарного рецептора DEC-205 в процессинге белков посредством пути МНС класса I, мы сначала химически соединили цельный белок OVA с антителом αDEC-205. Известно, что OVA презентируется на молекуле H-2K b MHC класса I CD8 + T-клеткам, включая TCR-трансгенные OT-I T-клетки (36). После связывания OVA с крысиным антимышиным антителом DEC-205 свободный OVA удаляли путем очистки конъюгата на колонках с белком G, получая смеси конъюгатов IgG:OVA ~1:1 (молекулярная масса ~200 кДа) и неконъюгированного антитела (рис. 1). А). Основываясь на наблюдаемой эффективности конъюгации и массовом отношении IgG к OVA 4:1, мы оценили, что <10% конъюгата состоит из белка OVA.
Чтобы убедиться, что αDEC-205:OVA нацелен на ДК у мышей, мы вводили 1 мкг конъюгатов подкожно и исследовали белок OVA в изолированных CD11c + DC-обогащенных (14, 37) и CD11c — DC- истощенные клетки лимфатических узлов. Предыдущая работа продемонстрировала захват крысиного антитела αDEC-205 большинством ДК (32), но здесь мы проследили за захватом белка OVA. Поглощение ДК стабилизировалось в дренирующем узле в течение 12–24 ч после инъекции (рис. 1, Б–Г). Иммунореактивный OVA размером 45 кД в ДК был подобен нативному OVA, что позволяет предположить, что большая часть белка высвобождается из его конъюгации с инъецированным антителом (рис. 1В). Инъекция 10 мкг растворимого OVA подкожно не давала обнаруживаемого антигена в лимфатических узлах, дренирующих место инъекции (фиг. 1C, левая дорожка). В случае конъюгата αDEC-205:OVA OVA был обнаружен только в CD11c + DC-обогащенная фракция, а не CD11c — DC-истощенные клетки (рис. 1C). Меньшие количества OVA были также обнаружены в ДК в дистальных участках, таких как лимфатические узлы и селезенка (рис. 1D). Количество захваченного OVA соответствовало ~10 нг белка на 10 6 ДК или 10 5 молекул/ДК (рис. 1 Б). Мы пришли к выводу, что белок, связанный с антителом αDEC-205, эффективно доставляется в ДК in situ.
ДК изолированных лимфатических узлов представляют конъюгаты αDEC-205:OVA на MHC класса I через TAP.
Чтобы определить, может ли антиген, доставляемый αDEC-205 в ДК, обрабатываться для представления MHC класса I, мы сначала выделили ДК из лимфатических узлов и обработали их αDEC-205:OVA in vitro. Конъюгаты антитело:OVA инкубировали с клетками лимфатических узлов CD11c + и CD11c — в течение ночи, избыток конъюгата удаляли промыванием, а затем клетки совместно культивировали в течение 2 дней с OVA-специфическими трансгенными TCR T-клетками (CD8 + MHC класса I – ограниченный OT-I или CD4 + MHC класса II – рестриктированные Т-клетки OT-II). Мы обнаружили, что OVA был представлен CD11c + DC на продуктах MHC класса I и класса II (рис. 2А). ДК, подвергшиеся воздействию 0,1 мкг/мл αDEC-205:OVA (с <10 нг/мл белка OVA), были более активны в представлении Т-клеток CD8 + OT-I, ограниченных MHC I класса, чем ДК, которые были подвергнуты воздействию 30 мкг/мл растворимого OVA (рис. 2А). Поглощение через DEC-205 увеличивало эффективность презентации по сравнению с неконъюгированным OVA по крайней мере в 1000 раз для MHC I и в 300 раз для MHC II (рис. 2А). Напротив, ни CD19В-клетки + и АПК, лишенные клеток CD11c + и CD19 + , были способны стимулировать пролиферацию Т-клеток выше фонового уровня, даже при клеточных дозах, в 30 раз превышающих дозы, необходимые для наблюдения представления ДК (рис. 2). Б). Конъюгаты изотипического контроля IgG:OVA были гораздо менее активны, чем конъюгаты αDEC-205:OVA (рис. 2В).
Было выполнено несколько дополнительных контролей (данные не показаны) для проверки функции рецептора DEC-205 в представлении белка, конъюгированного с соответствующим моноклональным антителом. Поскольку ДК могут использовать рецепторы Fcγ для представления иммунных комплексов на продуктах МНС класса I (20, 21), мы показали, что блокирование этих рецепторов (по крайней мере, CD16 и CD32) моноклональным антителом 2.4G2 αFcγR (38) не снижает представление αDEC- 205:ОВА. Чтобы оценить другой конъюгат антитело:OVA, который нацелен на эндоцитарную систему, мы использовали LAMP-1, ассоциированный с лизосомой мембранный белок, экспрессируемый в DC и других клетках. Конъюгат αLAMP-1:OVA был сравним только с изотипическим контрольным антителом:OVA в обеспечении презентации пептидов OVA в контексте либо MHC-I, либо -II с помощью CD11c + ДК. Конъюгация была необходима для эффективного нацеливания на OVA, поскольку смеси неконъюгированного αDEC-205 и растворимого OVA были представлены только с эффективностью, сравнимой с эквивалентными дозами растворимого OVA, что исключало возможность того, что αDEC-205 просто усиливал представление неконъюгированного OVA. Таким образом, рецептор DEC-205 опосредует эффективную презентацию белковых антигенов как на продуктах MHC класса I, так и на продуктах II ДК лимфатических узлов.
Чтобы установить, что презентация MHC класса I с помощью αDEC-205:OVA зависит от TAP, мы провели аналогичные эксперименты с DC, полученными от мышей с дефицитом TAP (TAP -/- ; Рис. 2 С). В отсутствие TAP процессированные протеосомами пептиды в DCs не могут перемещаться в эндоплазматический ретикулум для ассоциации с молекулами MHC I. Мы обнаружили, что TAP -/- DC были неспособны представлять OVA на MHC класса I после поглощения αDEC-205:OVA, но те же клетки представляли OVA на MHC класса II (рис. 2C). Кроме того, TAP -/- DC могут презентировать предварительно обработанный пептид OVA рестриктированным MHC класса I Т-клеткам OT-I (данные не показаны). Таким образом, антигены, эндоцитированные через DEC-205, направляются к аппарату процессинга MHC класса I по пути, который требует транспорта пептидов из цитоплазмы в эндоплазматический ретикулум.
В отличие от ДК, полученных из лимфатических узлов, экспрессирующие DEC-205 ДК, полученные из клеток-предшественников костного мозга с помощью GM-CSF, были способны презентировать конъюгаты αDEC-205:OVA только с OVA-специфическим CD4 + OT-II T клетки, но не CD8 + OT-I Т-клетки (рис. 2D). Следовательно, тип DC влияет на способность DEC-205 доставлять антигены для TAP-зависимой презентации MHC класса I.
DEC-205-опосредованная презентация MHC класса I In Vivo.
Чтобы определить, представляют ли ДК, на которые нацелен αDEC-205:OVA, также пептиды OVA на MHC класса I in vivo, мы затем выделили CD11c + и CD11c — клеток (или В-клеток, обогащенных популяцией CD11c — путем истощения CD5 + Т-клеток) от мышей, которым подкожно вводили конъюгаты или растворимый OVA, а затем мы анализировали презентацию антигена OT -I клеток in vitro без дальнейшего добавления антигена. После инъекции αDEC-205:OVA мы обнаружили сильную презентацию через 4 и 24 ч после инъекции, но только ДК (рис. 3 A , слева), а не CD11c — или обогащенные В-клетки (рис. 3). А, правильно). В-клетки также были неактивны, когда животным давали антитело αCD40 (вместе с антителом:OVA) для усиления их костимулирующих свойств (данные не представлены). Для обнаружения презентации требовались 100-кратно более высокие дозы растворимого OVA по сравнению с αDEC-205:OVA (который составляет <10% OVA), но DC снова избирательно презентировали антиген. Это представление было сильнее, когда клетки выделяли через 24 часа, а не через 4 часа после инъекции (рис. 3А). После инъекции αDEC-205:OVA презентация легко определялась в течение 4 дней (рис. 3В). Презентация DCs in vivo зависела от TAP (фиг. 3C). Напротив, конъюгаты антитело:OVA, направленные на другие маркеры DC, например, MHC-II и LAMP-1, не усиливали представление OVA (рис. 3D). Во всех экспериментах захват изотип-контроль: конъюгат OVA был незначительным или отсутствовал (рис. 3, B-D).
Для проверки того, что ДК, на которые нацелен αDEC-205:OVA, также могут презентировать антиген Т-клеткам in situ , , мы перенесли 2 × 10 6 CFSE-меченых, антигенспецифических Т-клеток (CD8 + OT-I клетки) за 1 день до инъекции αDEC-205:OVA, изотипа:OVA или растворимого OVA. Через 3 дня лимфатические узлы (рис. 3E), селезенка и кровь (данные не показаны) оценивали на предмет пролиферации OT-I, что определяли с помощью разведения CFSE. Практически все клетки OT-I в лимфатическом узле вступили в клеточный цикл и подверглись 3–7 клеточным делениям после дозы всего 1,0 мкг конъюгата αDEC-205:OVA (<100 нг OVA) на мышь (рис. 3E). Для растворимого OVA требовалось по крайней мере в 400 раз больше белка, чтобы вызвать сравнимые пролиферативные ответы, и, опять же, изотип-контроль: конъюгат OVA вызывал небольшую пролиферацию или не вызывал ее вообще (рис. 3E). Чтобы доказать, что DEC-205, а не рецепторы Fcγ опосредовали презентацию, мы подтвердили, что презентация отменялась с помощью DC из DEC-205 9.0939 -/- мышей (данные не представлены). Таким образом, αDEC-205 эффективно нацеливается на антигены для презентации экзогенным путем в MHC класса I in vivo.
αDEC-205:OVA не созревает ДК in vivo.
Чтобы выяснить, приводит ли обработка αDEC-205:OVA к созреванию ДК в присутствии или в отсутствие OVA-специфических Т-клеток OT-I, мы провели исследования FACS ® ДК от мышей, которым вводили конъюгаты 1 или 3 дня назад под разнообразные условия. Как показано на фиг. 4А, поверхностная экспрессия CD80, CD86, а также продуктов MHC класса II не изменилась у мышей, которым инъецировали αDEC-205:OVA, независимо от того, получали ли они Т-клетки OT-1 или нет. Количество ДК также не изменилось у мышей, получавших αDEC-205:OVA. Однако совместное введение агонистического антитела к αCD40 (FGK-45.5) в качестве адъюванта активировало DCs in situ в течение 3 дней и увеличивало их количество примерно в два раза. Степень созревания с αCD40 была одинаковой в отсутствие или в присутствии антигена (αDEC-205:OVA) или Т-клеток OT-I (фиг. 4А). Созревание было обнаружено в CD11c + ДК, которые имели низкий и высокий уровни DEC-205, но уровни CD86 были выше в высокой фракции DEC-205 (рис. 4Б). Таким образом, хотя ДК лимфатических узлов в стационарном состоянии экспрессируют молекулы, используемые для активации Т-клеток, такие как CD86, эти ДК, по-видимому, не дифференцируются дальше при воздействии αDEC-205:OVA, но дифференцируются в ответ на агонистическое антитело αCD40.
Различный ответ Т-клеток in vivo на антиген, представленный в стационарном состоянии, и созревание DC на основе CD40.
Чтобы проследить судьбу Т-клеток OT-I, которые пролиферировали в ответ на антиген, нацеленный на ДК in vivo , , мы отследили перенесенные Т-клетки с помощью проточной цитометрии с использованием комбинации CD45.1 и Vβ 5,1/5,2 маркеры, и мы сравнили ответы в стационарном состоянии с ответами после созревания DC, индуцированного αCD40. Через 3 дня после инъекции αDEC-205:OVA мы обнаружили сильные пролиферативные ответы в присутствии или отсутствии созревания ДК (рис. 5А). Однако у мышей, получавших αCD40, также наблюдалось значительно повышенное производство Т-клетками IL-2 и IFN-γ (фиг. 5В, нижний ряд).
Когда мы проследили количество инъецированных Т-клеток OT-I через 3 дня и через 12–14 дней в нескольких лимфоидных тканях (рис. 6, A и B), мы обнаружили, что αDEC-205:OVA в стационарном состоянии сначала увеличились клетки OT-I, но к 12–14 дню Т-клетки практически полностью отсутствовали в лимфатических узлах, селезенке или крови (рис. 6А, сравните левую и правую панели и рис. 6В). Однако, если конъюгат αDEC-205:OVA давали с αCD40, клетки OT-I размножались по сравнению с контролем PBS (фиг. 6, A и B) или конъюгатом изотип-контроль:OVA (данные не показаны). Точно так же, когда продукцию IFN-γ контролировали с помощью FACS ® (фиг. 6C) или с помощью анализов ELISPOT (данные не показаны), комбинация αDEC-205:OVA и αCD40 индуцировала сильную экспансию эффекторов, продуцирующих цитокины, в то время как клетки OT-I, продуцирующие цитокины, практически удалялись из лимфатический узел, когда мыши получали αDEC-205:OVA (фиг. 6C). Поскольку было возможно, что Т-клетки перераспределялись по тканям в стационарном состоянии, а не подвергались истинной делеции, мы выделили клетки из одного крупного периферического органа, легкого. У мышей, подвергшихся воздействию αDEC-205:OVA, мы не смогли обнаружить никаких Т-клеток OT-I в легких на 10-й день, но многие клетки OT-I были обнаружены в легких мышей, получавших αDEC-205:OVA плюс αCD40 (рис. 6 Д). Увеличение числа клеток в лимфатических узлах и легких в ответ на αDEC-205:OVA плюс αCD40 включало большую фракцию клеток, лишенных селектина CD62L для самонаведения в лимфатические узлы, что является типичной особенностью активированных Т-клеток (рис. 6D). Таким образом, доставка белковых антигенов в ДК приводит к делеции Т-клеток, рестриктированных по МНС класса I, но доставка антигена вместе со стимулом созревания приводит к экспансии Т-клеток, продукции IFN-γ и экспорту Т-клеток в периферические клетки. ткани.
Нацеливание антигена на ДК индуцирует толерантность периферических CD8
+ Т-клеток. Важнейшим критерием индукции периферической толерантности является неспособность реагировать на повторное введение антигена вместе с сильным адъювантом. Чтобы определить, становятся ли мыши, обработанные αDEC-205:OVA, толерантными к OVA, мы иммунизировали 50 мкг белка OVA, суспендированного в CFA, через 12 дней после введения конъюгата. Через 3 дня после OVA при иммунизации CFA мышей умерщвляли и иммунный ответ оценивали с использованием либо анализов ELISPOT (данные не показаны), либо FACS 9Анализы 0939® на внутриклеточную продукцию цитокинов (фиг. 7А). Контрольные мыши, предварительно обработанные PBS или изотип-контроль: OVA, могли быть примированы к OVA в CFA (фиг. 7A). Напротив, мыши, предварительно обработанные αDEC-205:OVA, были неспособны реагировать на заражение OVA (фиг. 7A, верхняя правая панель в каждой группе). Этих же мышей можно было иммунизировать белками, отличными от OVA, например, антигеном PPD в CFA (данные не представлены). Далее мы оценили образование эффекторных ЦТЛ, используя недавно описанный анализ функции ЦТЛ in vivo (35). Мыши, получившие толерантность αDEC-205:OVA за 12 дней до этого, не могли образовывать ЦТЛ, но мыши, которым вводили PBS или комбинацию αDEC-205:OVA с αCD40, вызывали сильные ответы ЦТЛ (фиг. 7В). Мы пришли к выводу, что нацеливание антигенов на ДК с помощью DEC-205 в стационарном состоянии приводит к индукции периферической толерантности без образования эффекторных Т-клеток и памяти, тогда как нацеливание в присутствии стимула созревания ДК приводит к иммунизации.
Эти находки дополняют доказательства того, что ДК в периферических лимфоидных органах конститутивно процессируют антигены в стационарном состоянии, и следствием этого проявления является периферическая толерантность. С идентификацией эффективного пути, опосредованного рецептором, DEC-205, становится ясно, что небольшие количества растворимого белка могут привести к представлению на MHC класса I и толерантности к CD8 + Т-клеткам. Предыдущие исследования с растворимыми белками подчеркивали необходимость высоких доз антигена для толерантности иммунной системы, и в первую очередь для продуктов MHC класса I.
Эндоцитарный рецептор DEC-205 играет несколько важных ролей в поглощении и процессинге антигена DCs. Во-первых, рецептор нацелен на MHC class II + , поздние эндосомальные/лизосомальные компартменты в культивируемых DCs, происходящих из костного мозга (31). Напротив, большинство рециклирующих рецепторов эндоцитоза перемещаются через периферические или ранние эндосомы. Позднее эндосомальное/лизосомальное нацеливание DEC-205 связано с гораздо более эффективным процессингом и презентацией антигена посредством пути МНС класса II. Эти свойства опосредованы цитозольным доменом DEC-205, особенно кислой последовательностью EDE (31). Во-вторых, антитела αDEC-205 в небольших количествах (<1 мкг/мышь) можно использовать для нацеливания на антигены для представления DCs in vivo. Когда антигенный пептид из лизоцима куриного яйца встраивается в тяжелую цепь антитела αDEC-205, антитело и пептид избирательно доставляются к ДК (32). Здесь αDEC-205 был химически конъюгирован с полноразмерным OVA и также селективно доставляет белок в DC in vivo. В-третьих, теперь мы обнаружили, что DEC-205 опосредует презентацию белковых антигенов посредством экзогенного, но TAP-зависимого пути MHC class I. Поглощение через DEC-205 подтверждает, что ДК лимфатических узлов в стационарном состоянии способны обрабатывать антигены на МНС класса I (35, 39).–42), а также продукты класса MHC II (18, 32). Напротив, мы не наблюдаем презентации αDEC-205:OVA на МНС класса I с использованием ДК, полученных из костного мозга, хотя эта популяция ДК представляет экзогенные белки на продуктах МНС класса II (43). Напротив, DCs из лимфатических узлов кажутся компетентными в представлении MHC класса I и II в стационарном состоянии in situ, без добавления стимулов созревания.
В стационарном состоянии DEC-205 представляет собой специфический рецептор для ДК, индуцирующий периферическую толерантность к растворимым антигенам как для CD4, так и для + (32) и CD8 + Т-клетки (эта статья). ДК также индуцируют толерантность к ассоциированным с клеткой антигенам in vivo (35, 39–41), но ответственные рецепторы до сих пор не идентифицированы. Хотя DEC-205 значительно усиливает презентацию белковых антигенов, возможно, что рецептор функционирует просто для усиления захвата антигена, а не для последующих событий, необходимых для презентации на МНС класса I. Кроме того, когда подходящие реагенты станут доступными, можно будет оценить, другие рецепторы на DCs (см. Введение) функционируют для усиления представления антигена и периферической толерантности in vivo. В настоящее время моноклональные антитела в первую очередь доступны для рецептора DEC-205, и, как упоминалось выше, ранее было показано, что они эффективно и избирательно нацеливаются на DC in vivo и усиливают представление MHC класса II. Ключевыми новыми моментами в этой статье являются, во-первых, то, что ДК обладают эффективными механизмами на основе рецепторов для улучшения презентации продуктов MHC класса I in vivo, во-вторых, они работают в устойчивом состоянии, и, в-третьих, следствием презентации является периферическая толерантность в CD8 9.0939 + отсек делеционным механизмом.
Наличие механизмов захвата, опосредованных рецепторами, должно позволить ДК играть ценную роль в подавлении реакции на безвредные аутоантигены и белки окружающей среды. Однократная доза <0,1 мкг OVA, конъюгированного с антителом αDEC-205, может удалить и сделать толерантными значительное количество (>10 6 ) инъецированных OVA-специфических CD8 + Т-клеток, что соответствует >1% от ~10 8 Т-клеток у мыши. Это число велико, так как <1/10 000 наивных Т-клеток обычно способны реагировать на специфический антиген. В результате способность DC представлять антигены экзогенным путем значительно превышает репертуар Т-клеток, которые должны быть активированы, по крайней мере, в отношении антигена, который эффективно распознается, т. Будет важно протестировать другие белковые антигены и более широкий репертуар Т-клеток, поскольку наши исследования на данный момент сосредоточены на Т-клетках OT-I, чтобы получить прямые данные о количестве и функции антиген-специфических Т-клеток. Тем не менее, наши результаты показывают, что CD8 + Толерантность Т-клеток (а также иммунитет) может быть достигнута in situ с помощью низких доз интактных белковых антигенов.
ДК в лимфатических узлах и селезенке проявляют многие признаки, указывающие на то, что они являются «незрелыми», то есть способны захватывать антигены, но не способны стимулировать иммунитет (44, 45). ДК активны в эндоцитозе (40, 46), а также реагируют in vivo на микробные, воспалительные и Т-клеточные стимулы, продуцируя цитокины (47) и активируя несколько костимулирующих молекул (48-50). Исследования с αDEC-205:OVA подтверждают, что ДК in vivo в стационарном состоянии не стимулируют иммунный ответ, даже если они эффективно представляют антиген и индуцируют обширную пролиферацию Т-клеток. Однако Т-клетки не могут дифференцироваться и удаляются, если также не предоставляется стимул к созреванию. На сегодняшний день мы сосредоточились на созревании ДК на основе CD40. Тем не менее полученные данные согласуются с мнением о том, что иммуногенная функция ДК, т. е. индукция эффекторных Т-клеток и развитие памяти, требует, чтобы имели место по крайней мере два набора событий. Один включает захват антигена и успешный процессинг с образованием комплексов MHC-пептид; это происходит в ДК лимфатических узлов в стационарном состоянии. Другое требование влечет за собой сложный процесс созревания, который изменяет DC разными способами, например, увеличивая костимулирующие молекулы Т-клеток, индуцируя цитокины, такие как IL-12 и IL-2, и изменяя экспрессию рецепторов хемокинов.
Определены два пути презентации антигенов на продуктах МНС класса I. Классический или «эндогенный» путь происходит из вновь синтезированных белков, особенно белков, происходящих из дефектных продуктов инициации рибосом (51). Этот путь обеспечивает элегантный и установленный механизм защитного иммунитета, направляя цитотоксические Т-лимфоциты, рестриктированные MHC класса I, к пептидам, продуцируемым в инфицированных и злокачественных клетках. Распознавая комплексы MHC-пептид, представленные на клеточной поверхности, CD8 9Цитолитическая реакция 0939 + сосредоточена на клетках, несущих патогены, а не на невинных клетках-свидетелях (52). Также существует второй «экзогенный» путь, позволяющий презентировать эндоцитированные нереплицирующиеся белки на MHC класса I. Экзогенный путь априори может позволить неинфицированным клеткам поглощать белок и становиться мишенями для MHC класса I, специфичного для CD8 + цитолитических T лимфоциты. Эта потенциально серьезная проблема была бы предотвращена, если бы экзогенный путь экспрессировался в первую очередь в DCs. Во время инфекции процессинг умирающих инфицированных клеток экзогенным путем дает ДК возможность инициировать CD8 9.0939 + Т-клеточный иммунитет к патогенам, которые не продуктивно инфицируют их. Сходным образом, в стационарном состоянии DCs в лимфоидных органах могут использовать экзогенный путь для представления собственных пептидов на MHC класса I (16, 40, 53; и эта статья) и, таким образом, индуцировать толерантность. Следовательно, для индукции толерантности не требуется, чтобы все клетки были способны обрабатывать антигены экзогенным путем; скорее, наличие эффективных рецепторов, таких как DEC-205, для экзогенной презентации in vivo обеспечивает специфический механизм для DCs, чтобы постоянно удалять репертуар реактивности периферических Т-клеток к низким уровням собственных белков и белков окружающей среды.
При поддержке грантов AI13013 и AI051573 Национальных институтов здравоохранения Р.М. Steinman, благодаря стипендии Instituto Mexicano del Seguro Social Л. Бонифазу и гранту CA09673 Национального института здравоохранения на обучение Д. Бонняю.
Л. Бонифаз, Д. Бонняй и К. Манке внесли равный вклад в эту работу.
*
Сокращения, используемые в данной статье: CFSE, сукцинимидиловый эфир карбоксифлуоресцеина диацетата; ДК, дендритная клетка.
1
Капплер, Дж. В., Н. Рем и П. Маррак.
1987
. Толерантность Т-клеток путем клональной элиминации в тимусе.
Сотовый.
49
:
273
–280.
2
Киселов, П., Х. Блатманн, У.Д. Штаерц, М. Штайнмец и Х. фон Бемер.
1988
. Толерантность у мышей, трансгенных по Т-клеточному рецептору, включает делецию незрелых CD4+8+ тимоцитов.
Природа.
333
:
742
–746.
3
Буно, К., П. Курильский и П. Буссо.
2000
. Влияние отрицательного отбора на репертуар Т-клеток, реагирующих на собственный пептид: большая часть клонов Т-клеток избегает клонального обнаружения.
Иммунитет.
13
:
829
–840.
4
Матцингер П. и С. Гердер.
1989
. Требуется ли для толерантности Т-клеток специальная антигенпрезентирующая клетка?
Природа.
338
:
74
–76.
5
Зал Т., А. Фолькманн и Б. Стокингер.
1994
. Механизмы индукции толерантности в рестриктированных Т-клетках главного комплекса гистосовместимости II класса, специфичных к аутоантигену, переносимому кровью.
J. Расшир. Мед.
180
:
2089
–2099.
6
Фолькманн, А., Т. Зал и Б. Стокингер.
1997
. Антиген-презентирующие клетки в тимусе, которые могут негативно отбирать МНС-рестриктированные Т-клетки класса II, распознающие циркулирующий аутоантиген.
Дж. Иммунол.
158
:
693
–706.
7
Стокингер, Б.
1999
. Толерантность Т-лимфоцитов: от делеции тимуса до механизмов периферического контроля.
Доп. Иммунол.
71
:
229
–265.
8
Сакагучи С.
2000
. Регуляторные Т-клетки: ключевые регуляторы иммунологической самопереносимости.
Сотовый.
101
:
455
–458.
9
Камрадт Т. и Н. А. Митчисон.
2001
. Толерантность и аутоиммунитет.
Н. англ. Дж. Мед.
344
:
655
–664.
10
Уокер, Л.С.К. и А.К. Аббас.
2002
. Враг внутри: сдерживание самореактивных Т-клеток на периферии.
Нац. Преподобный Иммунол.
2
:
11
–19.
11
Vermaelen, K.Y., I. Carro-Muino, B.N. Ламбрехт и Р.А. Пауэлс.
2001
. Специфические мигрирующие дендритные клетки быстро транспортируют антиген из дыхательных путей в грудные лимфатические узлы.
J. Расшир. Мед.
193
:
51
–60.
12
Хуанг, Ф.-П., Н. Платт, М. Вайкс, Дж. Р. Мейджор, Т.Дж. Пауэлл, К.Д. Дженкинс и Г.Г. Макферсон.
2000
. Дискретная субпопуляция дендритных клеток транспортирует апоптотические эпителиальные клетки кишечника в области Т-клеток брыжеечных лимфатических узлов.
J. Расшир. Мед.
191
:
435
–442.
13
Крааль, Г., М. Брил, М. Янсе и Г. Брюин.
1986
. Клетки Лангерганса, завуалированные клетки и межпальцевые клетки у мыши распознаются моноклональным антителом.
J. Расшир. Мед.
163
:
981
–997.
14
Метлай, Дж.П., М.Д. Витмер-Пак, Р. Аггер, М.Т. Кроули, Д. Лоулесс и Р.М. Штейнман.
1990
. Отдельные лейкоцитарные интегрины дендритных клеток селезенки мыши, идентифицированные с новыми моноклональными антителами хомяка.
J. Расшир. Мед.
171
:
1753
–1771.
15
Хит, В.Р. и Ф.Р. Карбон.
2001
. Кросс-презентация вирусного иммунитета и аутотолерантности.
Нац. Преподобный Иммунол.
1
:
126
–134.
16
Куртс К., М. Каннариле, И. Клебба и Т. Брокер.
2001
. Дендритных клеток достаточно для перекрестного представления аутоантигенов Т-клеткам CD8 in vivo.
Дж. Иммунол.
166
:
1439
–1442.
17
Морган, Д.Дж., Х.Т. Кройвель и Л. А. Шерман.
1999
. Концентрация антигена и частота предшественников определяют степень толерантности CD8+ Т-клеток к периферически экспрессируемым антигенам.
Дж. Иммунол.
163
:
723
–727.
18
Hugues, S., E. Mougneau, W. Ferlin, D. Jeske, P. Hofman, D. Homann, L. Beaudoin, C. Schrike, M. Von Herrath, A. Lehuen и N. Глайхенхаус.
2002
. Толерантность к островковым антигенам и профилактика диабета, вызванного ограниченным апоптозом β-клеток поджелудочной железы.
Иммунитет.
16
:
169
–181.
19
Айхеле П., К. Брдуша-Рим, Р.М. Цинкернагель, Х. Хенгартнер и Х. Пирхер.
1995
. Примирование Т-клеток по сравнению с толерантностью Т-клеток, индуцированной синтетическими пептидами.
J. Расшир. Мед.
182
:
261
–266.
20
Рено, А., Д. Ланкар, В. Лакабанн, А. Родригес, К. Тери, М. Решиньо, Т. Сайто, С. Вербек, К. Боннеро, П. Риччарди-Кастаньоли и С. , Амигорена.
1999
. Fcγ-рецептор-опосредованная индукция созревания дендритных клеток и главный комплекс гистосовместимости класса I – ограниченная презентация антигена после интернализации иммунного комплекса.
J. Расшир. Мед.
189
:
371
–380.
21
Родригес А., А. Реньо, М. Клеймер, П. Рикарди-Кастаньоли и С. Амигорена.
1999
. Селективный транспорт интернализованных антигенов в цитозоль для презентации МНС класса I в дендритных клетках.
Нац. Клеточная биол.
1
:
362
–368.
22
Дходапкар К.М., Дж. Красовский, Б. Уильямсон и М.В. Дходапкар.
2002
. Противоопухолевые моноклональные антитела усиливают перекрестную презентацию клеточных антигенов и образование специфичных для миеломы Т-клеток-киллеров дендритными клетками.
Дж. Эксп. Мед.
195
:
125
–133.
23
Калергис, А. М. и Дж. В. Равеч.
2002
. Индукция противоопухолевого иммунитета за счет избирательного взаимодействия активирующих рецепторов Fcγ на дендритных клетках.
J. Расшир. Мед.
195
:
1653
–1659.
24
Саллусто Ф., М. Целла, К. Даниэли и А. Ланцавеккья.
1995
. Дендритные клетки используют макропиноцитоз и маннозный рецептор для концентрации антигена в главном компартменте класса II гистосовместимости. Понижающая регуляция цитокинами и бактериальными продуктами.
J. Расшир. Мед.
182
:
389
–400.
25
Валладо, Дж., О. Равель, К. Дезуттер-Дамбуянт, К. Мур, М. Кляймер, Ю. Лю, В. Дювер-Фрэнсис, К. Винсент, Д. Шмитт, Дж. Даву, и другие.
2000
. Лангерин, новый лектин С-типа, специфичный для клеток Лангерганса, представляет собой эндоцитозный рецептор, который индуцирует образование гранул Бирбека.
Иммунитет.
12
:
71
–81.
26
Квон Д.С., Г. Грегарио, Н. Биттон, В.А. Хендриксон и Д.Р. Литтман.
2002
. Опосредованная DC-SIGN интернализация ВИЧ необходима для усиления Т-клеточной инфекции.
Иммунитет.
16
:
135
–144.
27
Энгеринг А., Т.Б. Гейтенбек, С.Дж. ван Влит, М. Вайерс, Э. ван Лимпт, Н. Деморекс, А. Ланзавеккья, Дж. Франсен, К.Г. Фигдор, В. Пиге и Ю. ван Койк.
2002
. Рецептор адгезии DC-SIGN, специфичный для дендритных клеток, интернализует антиген для представления Т-клеткам.
Дж. Иммунол.
168
:
2118
–2126.
28
Валладо, Ж. , В. Дювер-Франсис, Ж.-Ж. Pin, MJ Kleijmeer, S. Ait-Yahia, O. Ravel, C. Vincent, F. Vega, Jr., A. Helms, D. Gorman, et al.
2001
. Незрелые дендритные клетки человека экспрессируют изоформы рецептора асиалогликопротеина для эффективного эндоцитоза, опосредованного рецептором.
Дж. Иммунол.
167
:
5767
–5774.
29
Дзионек А., Ю. Сохма, Дж. Нагафуне, М. Селла, М. Колонна, Ф. Факкетти, Г. Гюнтер, И. Джонстон, А. Ланзавеккья, Т. Нагасака и др.
2001
. BDCA-2, новый лектин С-типа типа II, специфичный для плазмоцитоидных дендритных клеток, опосредует захват антигена и является мощным ингибитором индукции интерферона-α/β.
J. Расшир. Мед.
194
:
1823
–1834.
30
Цзян В., В.Дж. Свиггард, К. Хойфлер, М. Пэн, А. Мирза, Р.М. Штейнман и М.К. Нуссенцвейг.
1995
. Рецептор DEC-205, экспрессируемый дендритными клетками и эпителиальными клетками тимуса, участвует в процессинге антигена.
Природа.
375
:
151
–155.
31
Манке, К., М. Гуо, С. Ли, Х. Сепульведа, С.Л. Суэйн, М. Нуссенцвейг и Р.М. Штейнман.
2000
. Рецептор дендритных клеток для эндоцитоза, DEC-205, может рециркулировать и усиливать представление антигена через главный комплекс гистосовместимости класса II-позитивные лизосомальные компартменты.
J. Cell Biol.
151
:
673
–683.
32
Хавигер Д., К. Инаба, Ю. Дорсетт, К. Гуо, К. Манке, М. Ривера, Дж. В. Раветч, Р.М. Штейнман и М.К. Нуссенцвейг.
2001
. Дендритные клетки индуцируют невосприимчивость периферических Т-клеток в стационарных условиях in vivo.
Дж. Эксп. Мед.
194
:
769
–780.
33
Witmer-Pack, MD, WJ Swiggard, A. Mirza, K. Inaba и R.M. Штейнман.
1995
. Распределение белка DEC-205 в тканях, выявляемое моноклональным антителом NLDC-145. II. Экспрессия in situ в лимфоидных и нелимфоидных тканях.
Сотовый. Иммунол.
163
:
157
–162.
34
Инаба, К. , М. Инаба, Н. Романи, Х. Ая, М. Дегучи, С. Икехара, С. Мурамацу и Р.М. Штейнман.
1992
. Получение большого количества дендритных клеток из культур костного мозга мышей с добавлением гранулоцитарно-макрофагального колониестимулирующего фактора.
J. Расшир. Мед.
176
:
1693
–1702.
35
Эрнандес, Дж., С. Аунг, В.Л. Редмонд и Л.А. Шерман.
2001
. Фенотипический и функциональный анализ CD8+ Т-клеток, подвергающихся периферической делеции в ответ на перекрестную презентацию собственного антигена.
Дж. Эксп. Мед.
194
:
707
–718.
36
Хогквист, К. А., С.С. Джеймсон, В.Р. Хит, Дж.Л. Ховард, М.Дж. Беван и Ф.Р. Карбон.
1994
. Пептиды-антагонисты Т-клеточных рецепторов индуцируют положительную селекцию.
Сотовый.
76
:
17
–27.
37
Кроули М.Т., К. Инаба, М.Д. Витмер-Пак, С. Гезелтер и Р.М. Штейнман.
1990
. Использование сортировщика клеток с активированной флуоресценцией для обогащения дендритных клеток из селезенки мыши.
Дж. Иммунол. Методы.
133
:
55
–66.
38
Ункелесс, Дж. К.
1979
. Характеристика моноклонального антитела, направленного против мышиных макрофагов и рецепторов Fc лимфоцитов.
J. Расшир. Мед.
150
:
580
–596.
39
Куртс К., Х. Косака Ф.Р. Карбоне, J.F.A.P. Миллер и У. Р. Хит.
1997
. Ограниченная классом I перекрестная презентация экзогенных аутоантигенов приводит к делеции аутореактивных CD8+ Т-клеток.
J. Расшир. Мед.
186
:
239
–245.
40
Ийода Т., С. Симояма, К. Лю, Ю. Омацу, Ю. Маэда, К. Такахара, Ю. Акияма, Р.М. Штейнман и К. Инаба.
2002
. Субпопуляция дендритных клеток CD8+ избирательно эндоцитирует умирающие клетки в культуре и in vivo.
Дж. Эксп. Мед.
195
:
1289
–1302.
41
Лю, К., Т. Ийода, М. Сатернус, Ю. Кимура, К. Инаба и Р.М. Штейнман.
2002
. Иммунная толерантность после доставки умирающих клеток в дендритные клетки in situ.
J. Расшир. Мед.
196
:
1091
–1097.
42
Scheinecker, C., R. McHugh, E.M. Shevach, and R.N. Жермен.
2002
. Конститутивное представление естественного тканевого аутоантигена исключительно дендритными клетками в дренирующем лимфатическом узле.
J. Расшир. Мед.
196
:
1079
–1090.
43
Инаба, К., С. Терли, Т. Иода, Ф. Ямаиде, С. Шимояма, К. Рейс и Соуза, Р.Н. Жермен, И. Меллман и Р.М. Штейнман.
2000
. Образование иммуногенных пептидных лигандов МНС класса II в лизосомальных компартментах дендритных клеток регулируется воспалительными стимулами.
Дж. Эксп. Мед.
191
:
927
–936.
44
Инаба К., Г. Шулер, М.Д. Витмер, Дж. Валинский, Б. Атасси и Р.М. Штейнман.
1986
. Иммунологические свойства очищенных клеток Лангерганса: различные требования к стимуляции непримированных и сенсибилизированных Т-лимфоцитов.
J. Расшир. Мед.
164
:
605
–613.
45
Романи, Н. , С. Койде, М. Кроули, М. Витмер-Пак, А.М. Ливингстон, К.Г. Фатман, К. Инаба и Р.М. Штейнман.
1989
. Презентация экзогенных белковых антигенов дендритными клетками клонам Т-клеток: интактный белок лучше всего представлен незрелыми эпидермальными клетками Лангерганса.
J. Расшир. Мед.
169
:
1169
–1178.
46
Камат, А.Т., Дж. Пули, М.А. О’Киф, Д. Времец, Ю. Жан, А. Лью, А. Д’Амико, Л. Ву, Д.Ф. Жесткий, и К.С. Коротышка.
2000
. Развитие, созревание и скорость оборота популяций дендритных клеток селезенки мыши.
Дж. Иммунол.
165
:
6762
–6770.
47
Reis e Sousa, C. , S. Hieny, T. Scharton-Kersten, D. Jankovic, H. Charest, R.N. Жермен и А. Шер.
1997
. Микробная стимуляция in vivo индуцирует быструю CD40L-независимую продукцию IL-12 дендритными клетками и их перераспределение в области Т-клеток.
J. Расшир. Мед.
186
:
1819
–1829.
48
Де Смедт, Т., Б. Паяк, Э. Мурай, Л. Леспаньяр, Э. Хайнен, П. Де Бетселье, Дж. Урбен, О. Лео и М. Мозер.
1996
. Регуляция количества и созревания дендритных клеток липополисахаридом in vivo.
J. Расшир. Мед.
184
:
1413
–1424.
49
Спарвассер, Т., Р.М. Вабулас, Б. Виллмоу, Г.Б. Липфорд и Х. Вагнер.
2000
. Бактериальная CpG-ДНК активирует дендритные клетки in vivo: независимые от Т-хелперов цитотоксические Т-клеточные ответы на растворимые белки.
Евро. Дж. Иммунол.
30
:
3591
–3597.
50
Акбари О., Н. Панджвани, С. Гарсия, Р. Таскон, Д. Лоури и Б. Стокингер.
1999
. ДНК-вакцинация: трансфекция и активация дендритных клеток как ключевые события иммунитета.
J. Расшир. Мед.
189
:
169
–178.
51
Schubert, U., L.C. Антон, Дж. Гиббс, К.С. Норбери, Дж.В. Юделл и Дж. Р. Беннинк.
2000
. Быстрая деградация большой доли вновь синтезированных белков протеасомами.
Природа.
404
:
770
–774.
52
Цинкернагель Р.М. и П.К. Доэрти.
1979
. MHC-рестриктированные цитотоксические Т-клетки: исследования биологической роли полиморфных основных трансплантационных антигенов, определяющих рестрикционную специфичность, функцию и реактивность Т-клеток.
Доп. Иммунол.
27
:
51
–177.
53
den Haan, J., S. Lehar и M. Bevan.
2000
. CD8+, но не CD8- дендритные клетки перекрестно праймируют цитотоксические Т-клетки in vivo.
J. Расшир. Мед.
192
:
1685
–1696.
данные и цифрыДанные и цифры
содержимоеСодержимое
дополняетДополнения
ссылокСсылки
Рисунок 1.
Конъюгация и доставка OVA с помощью моноклональных антител. (A) OVA-конъюгированные и неконъюгированные IgG, разрешенные с помощью SDS-PAGE и окрашенные кумасси синим. (B) Иммуноблоттинг с анти-OVA антителом для обнаружения OVA в конъюгатах Ig:OVA и в CD11c + ДК лимфатических узлов (10 6 /дорожка), последний через 12 и 24 ч после подкожной инъекции 1 мкг αDEC-205:OVA или изотипический контроль:OVA. (C) Подкожная инъекция 1 мкг αDEC-205:OVA на подушечку стопы, но не 10 мкг растворимого OVA, нацелена на CD11c 9.0939 + ДК лимфатических узлов. 10 6 CD11c + клеток из дренирующих узлов на дорожку подвергали иммуноблотингу через 12–72 ч после инъекции. (D) Поглощение αDEC-205:OVA происходит главным образом в CD11c + DC в дренирующих лимфатических узлах. Как и в С, но фракции клеток получали из дренирующих лимфатических узлов, дистальных (Dist.) узлов и селезенки (Spl.).
Рисунок 1.
Конъюгация и доставка OVA с помощью моноклональных антител. (A) OVA-конъюгированные и неконъюгированные IgG, разрешенные с помощью SDS-PAGE и окрашенные кумасси синим. (B) Иммуноблоттинг с антителом против OVA для обнаружения OVA в конъюгатах Ig:OVA и в CD11c + ДК лимфатических узлов (10 6 /полоса), последние через 12 и 24 ч после подкожной инъекции 1 мкг αDEC-205:OVA или изотипический контроль:OVA. (C) Подкожная инъекция 1 мкг αDEC-205:OVA на подушечку стопы, но не 10 мкг растворимого OVA, нацелена на CD11c + ДК лимфатических узлов. 10 6 CD11c + клеток из дренирующих узлов на дорожку подвергали иммуноблотингу через 12–72 ч после инъекции. (D) Поглощение αDEC-205:OVA происходит главным образом в CD11c + DC в дренирующих лимфатических узлах. Как и в С, но фракции клеток получали из дренирующих лимфатических узлов, дистальных (Dist.) узлов и селезенки (Spl.).
Закрыть модальное окно
Рисунок 2.
Нацеливание αDEC-205:OVA на CD11c + ДК лимфатических узлов in vitro. (A) αDEC-205:OVA вызывает более сильную презентацию, чем только OVA, дозозависимым образом. Клетки CD11c + из лимфатических узлов C57BL/6 культивировали в течение 18 ч в градуированных дозах только αDEC-205:OVA или OVA, промывали и совместно культивировали с Т-клетками OT-I или OT-II перед измерением поглощения [ 3 Н]тимидин через 48–72 ч для оценки пролиферации Т-клеток. (B) Презентация пептидов, полученных из αDEC-205:OVA, ограничена CD11c + клетками лимфатических узлов, а не фракциями CD19 + или CD11c — CD19 — (двойные отрицательные). Как в А, но с αDEC-205:OVA или конъюгатом изотип:OVA в концентрации 10 мкг/мл. (C) Презентация пептидов, полученных из αDEC-205:OVA, зависит от TAP. То же, что и в B, но клетки CD11c + получали из C57BL/6 или TAP 9.0939 -/- мышей. (D) DCs костного мозга неспособны представить αDEC-205:OVA на продуктах MHC класса I. Клетки из культур d6 собирали и культивировали с конъюгатами антитело:OVA в течение 6 ч при 10 мкг/мл, промывали и совместно культивировали с Т-клетками, как показано на панели А. Данные являются репрезентативными для трех экспериментов.
Рисунок 2.
Нацеливание αDEC-205:OVA на CD11c + ДК лимфатических узлов in vitro. (A) αDEC-205:OVA вызывает более сильную презентацию, чем только OVA, дозозависимым образом. CD11c 9Клетки 0939 + из лимфатических узлов C57BL/6 культивировали в течение 18 ч в градуированных дозах только αDEC-205:OVA или OVA, промывали и совместно культивировали с Т-клетками OT-I или OT-II перед измерением поглощения [ 3 H ] тимидина через 48–72 ч для оценки пролиферации Т-клеток. (B) Презентация пептидов, полученных из αDEC-205:OVA, ограничена CD11c + клетками лимфатических узлов, а не фракциями CD19 + или CD11c — CD19 — (двойные отрицательные). Как в А, но с αDEC-205:OVA или конъюгатом изотип:OVA в концентрации 10 мкг/мл. (C) Презентация пептидов, полученных из αDEC-205:OVA, зависит от TAP. Как в B, но CD11c 9Клетки 0939+ получали из мышей C57BL/6 или TAP -/-. (D) DCs костного мозга неспособны представить αDEC-205:OVA на продуктах MHC класса I. Клетки из культур d6 собирали и культивировали с конъюгатами антитело:OVA в течение 6 ч при 10 мкг/мл, промывали и совместно культивировали с Т-клетками, как показано на панели А. Данные являются репрезентативными для трех экспериментов.
Закрыть модальное окно
Рисунок 3.
Нацеливание αDEC-205:OVA на лимфатический узел CD11c + ДК in vivo. ( A ) Только CD11c + DC лимфатических узлов эффективно представляют экзогенный αDEC-205: OVA и, в меньшей степени, OVA Т-клеткам OT-I. Мышам C57BL/6 вводили 4,0 мкг (1,0 мкг на подушечку лапы) конъюгата αDEC-205:OVA или 400 мкг (100 мкг на подушечку лапы) растворимого OVA подкожно за 4 и 24 часа до умерщвления. Фракции CD11c + и CD11c — CD5 — (В-клетки) были отсортированы по MACS ® из лимфатических узлов и оценены на презентацию Т-клеткам OT-I, как показано на рис. 2. максимальное титрование АПК (ДК, слева; В-клетки, справа) для каждой группы. (B) Как на панели A, но CD11c + ДК исследовали через 1 и 4 дня после подкожной инъекции 4,0 мкг (1,0 мкг/подушечка стопы) конъюгатов антитело:OVA. (C) Презентация DC пептидов OVA из C57BL/6, но не TAP -/- мышей, получавших 4,0 мкг (1,0 мкг/подушечка лапы) конъюгатов IgG:OVA подкожно за 4 дня до этого. (D) αDEC-205:OVA вызывает лучшую презентацию пептидов, полученных из OVA, чем другие конъюгаты, нацеленные на ДК, при подкожном введении 4,0 мкг (1,0 мкг/подушечка стопы) конъюгатов IgG:OVA за 4 дня до этого. (E) αDEC-205:OVA индуцирует более сильную пролиферацию OT-I Т-клеток in vivo, чем один OVA. Мышам C57BL/6 внутривенно вводили 2 × 10 6 CFSE-меченые Т-клетки OT-I, а затем градуированные дозы конъюгатов IgG:OVA или OVA подкожно через 24 часа. Через 3 дня после инъекции конъюгата собирали лимфатические узлы и оценивали размножение клеток CD8 + Vα 2 Vβ 5,1/5,2 с помощью проточной цитометрии для разбавления CFSE. Каждая панель представляет два или более эксперимента.
Рисунок 3.
Нацеливание αDEC-205:OVA на лимфатические узлы CD11c + DCs in vivo. (А) Только CD11c + ДК лимфатических узлов эффективно представляют экзогенный αDEC-205:OVA и, в меньшей степени, OVA Т-клеткам OT-I. Мышам C57BL/6 вводили 4,0 мкг (1,0 мкг на подушечку лапы) конъюгата αDEC-205:OVA или 400 мкг (100 мкг на подушечку лапы) растворимого OVA подкожно за 4 и 24 часа до умерщвления. Фракции CD11c + и CD11c — CD5 — (В-клетки) были отсортированы по MACS ® из лимфатических узлов и оценены на презентацию Т-клеткам OT-I, как показано на рис. 2. максимальное титрование АПК (ДК, слева; В-клетки, справа) для каждой группы. (B) Как на панели A, но CD11c + ДК исследовали через 1 и 4 дня после подкожной инъекции 4,0 мкг (1,0 мкг/подушечка стопы) конъюгатов антитело:OVA. (C) Презентация DC пептидов OVA из C57BL/6, но не TAP -/- мышей, получавших 4,0 мкг (1,0 мкг/подушечка лапы) конъюгатов IgG:OVA подкожно за 4 дня до этого. (D) αDEC-205:OVA вызывает лучшую презентацию пептидов, полученных из OVA, чем другие конъюгаты, нацеленные на ДК, при подкожном введении 4,0 мкг (1,0 мкг/подушечка стопы) конъюгатов IgG:OVA за 4 дня до этого. (E) αDEC-205:OVA индуцирует более сильную пролиферацию OT-I Т-клеток in vivo, чем один OVA. Мышам C57BL/6 внутривенно вводили 2 × 10 6 CFSE-меченые Т-клетки OT-I, а затем градуированные дозы конъюгатов IgG:OVA или OVA подкожно через 24 часа. Через 3 дня после инъекции конъюгата собирали лимфатические узлы и оценивали размножение клеток CD8 + Vα 2 Vβ 5,1/5,2 с помощью проточной цитометрии для разбавления CFSE. Каждая панель представляет два или более эксперимента.
Закрыть модальное окно
Рисунок 4.
Созревание ДК in vivo под действием агониста αCD40, но не под действием αDEC-205:OVA. (A) Мышам C57BL/6 подкожно вводили PBS или 4,0 мкг (1,0 мкг на подушечку стопы) конъюгата αDEC-205:OVA с αCD40 или без него (100 мкг FGK45.5 подкожно), за 1 и 3 дня до умерщвления. CD11c 9Клетки 0939+ сортировали с помощью MACS ® из лимфатических узлов и оценивали с помощью проточной цитометрии на экспрессию CD80, CD86 и МНС класса II. Перед инъекцией конъюгата OVA и αCD40 мышам вводили клетки PBS (-) или OT-I (+). Жирные символы представляют собой средние показатели флуоресценции клеток CD11c + в присутствии стимула созревания, в то время как серым жирным шрифтом на 3-й день указано значительное увеличение, согласующееся с созреванием. (B) Иллюстративные данные FACS ®, показывающие созревание DEC-205 hi CD11c + клеток и DEC-205 lo CD11c + клеток у мышей, обработанных за 3 дня до этого PBS и αCD40, как на панели A.
Рисунок 4.
Созревание ДК in vivo под действием агониста αCD40, но не под действием αDEC-205:OVA. (A) Мышам C57BL/6 подкожно вводили PBS или 4,0 мкг (1,0 мкг на подушечку стопы) конъюгата αDEC-205:OVA с αCD40 или без него (100 мкг FGK45.5 подкожно), за 1 и 3 дня до умерщвления. CD11c 9Клетки 0939+ сортировали с помощью MACS ® из лимфатических узлов и оценивали с помощью проточной цитометрии на экспрессию CD80, CD86 и МНС класса II. Перед инъекцией конъюгата OVA и αCD40 мышам вводили клетки PBS (-) или OT-I (+). Жирные символы представляют собой средние показатели флуоресценции клеток CD11c + в присутствии стимула созревания, в то время как серым жирным шрифтом на 3-й день указано значительное увеличение, согласующееся с созреванием. (B) Иллюстративные данные FACS ®, показывающие созревание DEC-205 hi CD11c + клеток и DEC-205 lo CD11c + клеток у мышей, обработанных за 3 дня до этого PBS и αCD40, как на панели A.
Закрыть модальное окно
Рисунок 5.
Контрастные ответы Т-клеток OT-I на αDEC-205:OVA в присутствии или в отсутствие αCD40-индуцированного созревания ДК. (A) αCD40 мало влияет на индуцированную αDEC-205:OVA пролиферацию Т-клеток OT-I. Как на фиг. 3Е, но мышам давали или не давали αCD40 (100 мкг FGK45.5 подкожно). (B) Дифференциальная продукция IL-2 и IFN-γ Т-клетками OT-I в ответ на изотип:OVA или αDEC-205:OVA с αCD40 или без него. Как и на панели А, но суспензии лимфатических узлов повторно стимулировали in vitro родственным пептидом OT-I в течение 5 ч в присутствии брефельдина А (5 мкг/мл) перед окрашиванием на внутриклеточный цитокин. Данные являются репрезентативными для трех экспериментов.
Рисунок 5.
Контрастные ответы Т-клеток OT-I на αDEC-205:OVA в присутствии или в отсутствие αCD40-индуцированного созревания ДК. (A) αCD40 мало влияет на индуцированную αDEC-205:OVA пролиферацию Т-клеток OT-I. Как на фиг. 3Е, но мышам давали или не давали αCD40 (100 мкг FGK45.5 подкожно). (B) Дифференциальная продукция IL-2 и IFN-γ Т-клетками OT-I в ответ на изотип:OVA или αDEC-205:OVA с αCD40 или без него. Как и на панели А, но суспензии лимфатических узлов повторно стимулировали in vitro родственным пептидом OT-I в течение 5 ч в присутствии брефельдина А (5 мкг/мл) перед окрашиванием на внутриклеточный цитокин. Данные являются репрезентативными для трех экспериментов.
Закрыть модальное окно
Рисунок 6.
Делеция Т-клеток OT-I в ответ на αDEC-205:OVA в стабильном состоянии. (A) Мышам C57BL/6 вводили CD45.1 + OT-I T-клеток и антиген, как описано в 3E, с αCD40 или без него. Через 3 и 12 дней лимфатические узлы, селезенку и кровь собирали и оценивали на наличие Т-клеток OT-I (CD45.1 + Vβ 5,1/5,2 + ) с помощью проточной цитометрии. (B) Данные о количестве клеток OT-I, выраженные в виде среднего процента CD8 + T-клетки из трех экспериментов типа, показанного на панели A. (C) αCD40-спасенные OT-I T-клетки примированы и секретируют IFN-γ. Мышам C57BL/6 вводили Т-клетки OT-I и антиген, как описано на панели A. Через 12 дней после введения антигена собирали лимфатические узлы и Т-клетки OT-I оценивали на секрецию IFN-γ, как описано на фиг. 5B. ( D) Т-клетки OT-I отсутствуют в периферической нелимфоидной ткани после презентации αDEC-205:OVA DCs в стационарном состоянии в отсутствие стимуляции αCD40. Как и на панели А, но легкие собирали через 10 дней после введения антигена и клетки OT-I оценивали на экспрессию CD62L и CD45.1.
Рисунок 6.
Делеция Т-клеток OT-I в ответ на αDEC-205:OVA в стабильном состоянии. (A) Мышам C57BL/6 вводили CD45.1 + OT-I T-клеток и антиген, как описано в 3E, с αCD40 или без него. Через 3 и 12 дней лимфатические узлы, селезенку и кровь собирали и оценивали на наличие Т-клеток OT-I (CD45.1 + Vβ 5,1/5,2 + ) с помощью проточной цитометрии. (B) Данные о количестве клеток OT-I, выраженные в виде среднего процента CD8 + T-клетки из трех экспериментов типа, показанного на панели A. (C) αCD40-спасенные OT-I T-клетки примированы и секретируют IFN-γ. Мышам C57BL/6 вводили Т-клетки OT-I и антиген, как описано на панели A. Через 12 дней после введения антигена собирали лимфатические узлы и Т-клетки OT-I оценивали на секрецию IFN-γ, как описано на фиг. 5B. ( D) Т-клетки OT-I отсутствуют в периферической нелимфоидной ткани после презентации αDEC-205:OVA DCs в стационарном состоянии в отсутствие стимуляции αCD40. Как и на панели А, но легкие собирали через 10 дней после введения антигена и клетки OT-I оценивали на экспрессию CD62L и CD45.1.
Закрыть модальное окно
Рисунок 7.
αDEC-205:OVA индуцирует периферическую толерантность к OVA в стабильном состоянии. (A) Мышам C57BL/6 вводили CD45.1 + OT-I T-клеток и антиген, как описано в 3E, с αCD40 или без него. Через 12 дней после введения антигена мышам повторно вводили 50 мкг белка OVA в полной CFA. Через 3 дня собирали лимфатические узлы и Т-клетки OT-I оценивали на секрецию IL-2 (вверху) или IFN-γ (внизу), как на рис. 5 B. (B) Мышей C57BL/6 лечили, как на панели. А, но через 3 дня после введения OVA в CFA мышам внутривенно вводили смесь сингенных спленоцитов, меченных CFSE, импульсно (CFSE hi ) или без (CFSE 10 ) родственного пептида OT-I (3×10 6 каждого). Через 12 ч потерю клеток CFSE hi в лимфатических узлах оценивали как меру специфической активности ЦТЛ. Наивные мыши не обнаруживают потери клеток, меченных CFSE (не показано). Результаты являются репрезентативными для трех экспериментов.
Рисунок 7.
αDEC-205:OVA индуцирует периферическую толерантность к OVA в стабильном состоянии. (A) Мыши C57BL/6 получали CD45.1 + OT-I T-клетки и антиген, как описано в 3E, с αCD40 или без него. Через 12 дней после введения антигена мышам повторно вводили 50 мкг белка OVA в полной CFA. Через 3 дня собирали лимфатические узлы и Т-клетки OT-I оценивали на секрецию IL-2 (вверху) или IFN-γ (внизу), как на рис. 5 B. (B) Мышей C57BL/6 лечили, как на панели. А, но через 3 дня после введения OVA в CFA мышам внутривенно вводили смесь сингенных спленоцитов, меченных CFSE, импульсно (CFSE hi ) или без (CFSE lo ) родственного пептида OT-I (3×10 6 каждого). Через 12 ч потерю клеток CFSE hi в лимфатических узлах оценивали как меру специфической активности ЦТЛ. Наивные мыши не обнаруживают потери клеток, меченных CFSE (не показано). Результаты являются репрезентативными для трех экспериментов.
Закрыть модальное окно
- Предыдущая статья 906:40
- Следующая статья
1
Капплер, Дж. В., Н. Рем и П. Маррак.
1987
. Толерантность Т-клеток путем клональной элиминации в тимусе.
Сотовый.
49
:
273
–280.
2
Киселов, П., Х. Блатманн, У.Д. Штаерц, М. Штайнмец и Х. фон Бемер.
1988
. Толерантность у мышей, трансгенных по Т-клеточному рецептору, включает делецию незрелых CD4+8+ тимоцитов.
Природа.
333
:
742
–746.
3
Буно, К., П. Курильский и П. Буссо.
2000
. Влияние отрицательного отбора на репертуар Т-клеток, реагирующих на собственный пептид: большая часть клонов Т-клеток избегает клонального обнаружения.
Иммунитет.
13
:
829
–840.
4
Матцингер П. и С. Гердер.
1989
. Требуется ли для толерантности Т-клеток специальная антигенпрезентирующая клетка?
Природа.
338
:
74
–76.
5
Зал Т., А. Фолькманн и Б. Стокингер.
1994
. Механизмы индукции толерантности в рестриктированных Т-клетках главного комплекса гистосовместимости II класса, специфичных к аутоантигену, переносимому кровью.
J. Расшир. Мед.
180
:
2089
–2099.
6
Фолькманн, А. , Т. Зал и Б. Стокингер.
1997
. Антиген-презентирующие клетки в тимусе, которые могут негативно отбирать МНС-рестриктированные Т-клетки класса II, распознающие циркулирующий аутоантиген.
Дж. Иммунол.
158
:
693
–706.
7
Стокингер, Б.
1999
. Толерантность Т-лимфоцитов: от делеции тимуса до механизмов периферического контроля.
Доп. Иммунол.
71
:
229
–265.
8
Сакагучи С.
2000
. Регуляторные Т-клетки: ключевые регуляторы иммунологической самопереносимости.
Сотовый.
101
:
455
–458.
9
Камрадт Т. и Н. А. Митчисон.
2001
. Толерантность и аутоиммунитет.
Н. англ. Дж. Мед.
344
:
655
–664.
10
Уокер, Л.С.К. и А.К. Аббас.
2002
. Враг внутри: сдерживание самореактивных Т-клеток на периферии.
Нац. Преподобный Иммунол.
2
:
11
–19.
11
Vermaelen, K.Y., I. Carro-Muino, B.N. Ламбрехт и Р.А. Пауэлс.
2001
. Специфические мигрирующие дендритные клетки быстро транспортируют антиген из дыхательных путей в грудные лимфатические узлы.
Дж. Эксп. Мед.
193
:
51
–60.
12
Хуанг, Ф.-П., Н. Платт, М. Вайкс, Дж. Р. Мейджор, Т.Дж. Пауэлл, К.Д. Дженкинс и Г.Г. Макферсон.
2000
. Дискретная субпопуляция дендритных клеток транспортирует апоптотические эпителиальные клетки кишечника в области Т-клеток брыжеечных лимфатических узлов.
J. Расшир. Мед.
191
:
435
–442.
13
Крааль Г., М. Брил, М. Янсе и Г. Брюин.
1986
. Клетки Лангерганса, завуалированные клетки и межпальцевые клетки у мыши распознаются моноклональным антителом.
J. Расшир. Мед.
163
:
981
–997.
14
Метлай, Дж.П., М.Д. Витмер-Пак, Р. Аггер, М.Т. Кроули, Д. Лоулесс и Р.М. Штейнман.
1990
. Отдельные лейкоцитарные интегрины дендритных клеток селезенки мыши, идентифицированные с новыми моноклональными антителами хомяка.
Дж. Эксп. Мед.
171
:
1753
–1771.
15
Хит, В.Р. и Ф.Р. Карбон.
2001
. Кросс-презентация вирусного иммунитета и аутотолерантности.
Нац. Преподобный Иммунол.
1
:
126
–134.
16
Куртс К., М. Каннариле, И. Клебба и Т. Брокер.
2001
. Дендритных клеток достаточно для перекрестного представления аутоантигенов Т-клеткам CD8 in vivo.
Дж. Иммунол.
166
:
1439
–1442.
17
Морган, Д.Дж., Х.Т. Кройвель и Л. А. Шерман.
1999
. Концентрация антигена и частота предшественников определяют степень толерантности CD8+ Т-клеток к периферически экспрессируемым антигенам.
Дж. Иммунол.
163
:
723
–727.
18
Hugues, S., E. Mougneau, W. Ferlin, D. Jeske, P. Hofman, D. Homann, L. Beaudoin, C. Schrike, M. Von Herrath, A. Lehuen и N. Глайхенхаус.
2002
. Толерантность к островковым антигенам и профилактика диабета, вызванного ограниченным апоптозом β-клеток поджелудочной железы.
Иммунитет.
16
:
169
–181.
19
Айхеле П., К. Брдуша-Рим, Р.М. Цинкернагель, Х. Хенгартнер и Х. Пирхер.
1995
. Примирование Т-клеток по сравнению с толерантностью Т-клеток, индуцированной синтетическими пептидами.
J. Расшир. Мед.
182
:
261
–266.
20
Рено, А., Д. Ланкар, В. Лакабанн, А. Родригес, К. Тери, М. Решиньо, Т. Сайто, С. Вербек, К. Боннеро, П. Риччарди-Кастаньоли и С. , Амигорена.
1999
. Fcγ-рецептор-опосредованная индукция созревания дендритных клеток и главный комплекс гистосовместимости класса I – ограниченная презентация антигена после интернализации иммунного комплекса.
J. Расшир. Мед.
189
:
371
–380.
21
Родригес А., А. Реньо, М. Клеймер, П. Рикарди-Кастаньоли и С. Амигорена.
1999
. Селективный транспорт интернализованных антигенов в цитозоль для презентации МНС класса I в дендритных клетках.
Нац. Клеточная биол.
1
:
362
–368.
22
Дходапкар К.М., Дж. Красовский, Б. Уильямсон и М.В. Дходапкар.
2002
. Противоопухолевые моноклональные антитела усиливают перекрестную презентацию клеточных антигенов и образование специфичных для миеломы Т-клеток-киллеров дендритными клетками.
Дж. Эксп. Мед.
195
:
125
–133.
23
Калергис, А. М. и Дж. В. Равеч.
2002
. Индукция противоопухолевого иммунитета за счет избирательного взаимодействия активирующих рецепторов Fcγ на дендритных клетках.
J. Расшир. Мед.
195
:
1653
–1659.
24
Саллусто Ф., М. Целла, К. Даниэли и А. Ланцавеккья.
1995
. Дендритные клетки используют макропиноцитоз и маннозный рецептор для концентрации антигена в главном компартменте класса II гистосовместимости. Понижающая регуляция цитокинами и бактериальными продуктами.
J. Расшир. Мед.
182
:
389
–400.
25
Валладо, Дж., О. Равель, К. Дезуттер-Дамбуянт, К. Мур, М. Кляймер, Ю. Лю, В. Дювер-Фрэнсис, К. Винсент, Д. Шмитт, Дж. Даву, и другие.
2000
. Лангерин, новый лектин С-типа, специфичный для клеток Лангерганса, представляет собой эндоцитозный рецептор, который индуцирует образование гранул Бирбека.
Иммунитет.
12
:
71
–81.
26
Квон Д.С., Г. Грегарио, Н. Биттон, В.А. Хендриксон и Д.Р. Литтман.
2002
. Опосредованная DC-SIGN интернализация ВИЧ необходима для усиления Т-клеточной инфекции.
Иммунитет.
16
:
135
–144.
27
Энгеринг А. , Т.Б. Гейтенбек, С.Дж. ван Влит, М. Вайерс, Э. ван Лимпт, Н. Деморекс, А. Ланзавеккья, Дж. Франсен, К.Г. Фигдор, В. Пиге и Ю. ван Койк.
2002
. Рецептор адгезии DC-SIGN, специфичный для дендритных клеток, интернализует антиген для представления Т-клеткам.
Дж. Иммунол.
168
:
2118
–2126.
28
Валладо, Ж., В. Дювер-Франсис, Ж.-Ж. Pin, MJ Kleijmeer, S. Ait-Yahia, O. Ravel, C. Vincent, F. Vega, Jr., A. Helms, D. Gorman, et al.
2001
. Незрелые дендритные клетки человека экспрессируют изоформы рецептора асиалогликопротеина для эффективного эндоцитоза, опосредованного рецептором.
Дж. Иммунол.
167
:
5767
–5774.
29
Дзионек А. , Ю. Сохма, Дж. Нагафуне, М. Селла, М. Колонна, Ф. Факкетти, Г. Гюнтер, И. Джонстон, А. Ланзавеккья, Т. Нагасака и др.
2001
. BDCA-2, новый лектин С-типа типа II, специфичный для плазмоцитоидных дендритных клеток, опосредует захват антигена и является мощным ингибитором индукции интерферона-α/β.
J. Расшир. Мед.
194
:
1823
–1834.
30
Цзян В., В.Дж. Свиггард, К. Хойфлер, М. Пэн, А. Мирза, Р.М. Штейнман и М.К. Нуссенцвейг.
1995
. Рецептор DEC-205, экспрессируемый дендритными клетками и эпителиальными клетками тимуса, участвует в процессинге антигена.
Природа.
375
:
151
–155.
31
Манке, К. , М. Гуо, С. Ли, Х. Сепульведа, С.Л. Суэйн, М. Нуссенцвейг и Р.М. Штейнман.
2000
. Рецептор дендритных клеток для эндоцитоза, DEC-205, может рециркулировать и усиливать представление антигена через главный комплекс гистосовместимости класса II-позитивные лизосомальные компартменты.
J. Cell Biol.
151
:
673
–683.
32
Хавигер Д., К. Инаба, Ю. Дорсетт, К. Гуо, К. Манке, М. Ривера, Дж. В. Раветч, Р.М. Штейнман и М.К. Нуссенцвейг.
2001
. Дендритные клетки индуцируют невосприимчивость периферических Т-клеток в стационарных условиях in vivo.
Дж. Эксп. Мед.
194
:
769
–780.
33
Witmer-Pack, MD, WJ Swiggard, A. Mirza, K. Inaba и R.M. Штейнман.
1995
. Распределение белка DEC-205 в тканях, выявляемое моноклональным антителом NLDC-145. II. Экспрессия in situ в лимфоидных и нелимфоидных тканях.
Сотовый. Иммунол.
163
:
157
–162.
34
Инаба, К., М. Инаба, Н. Романи, Х. Ая, М. Дегучи, С. Икехара, С. Мурамацу и Р.М. Штейнман.
1992
. Получение большого количества дендритных клеток из культур костного мозга мышей с добавлением гранулоцитарно-макрофагального колониестимулирующего фактора.
J. Расшир. Мед.
176
:
1693
–1702.
35
Эрнандес, Дж., С. Аунг, В.Л. Редмонд и Л.А. Шерман.
2001
. Фенотипический и функциональный анализ CD8+ Т-клеток, подвергающихся периферической делеции в ответ на перекрестную презентацию собственного антигена.
Дж. Эксп. Мед.
194
:
707
–718.
36
Хогквист, К.А., С.С. Джеймсон, В.Р. Хит, Дж.Л. Ховард, М.Дж. Беван и Ф.Р. Карбон.
1994
. Пептиды-антагонисты Т-клеточных рецепторов индуцируют положительную селекцию.
Сотовый.
76
:
17
–27.
37
Кроули М.Т., К. Инаба, М.Д. Витмер-Пак, С. Гезелтер и Р.М. Штейнман.
1990
. Использование сортировщика клеток с активированной флуоресценцией для обогащения дендритных клеток из селезенки мыши.
Дж. Иммунол. Методы.
133
:
55
–66.
38
Ункелесс, Дж. К.
1979
. Характеристика моноклонального антитела, направленного против мышиных макрофагов и рецепторов Fc лимфоцитов.
J. Расшир. Мед.
150
:
580
–596.
39
Куртс К., Х. Косака Ф.Р. Карбоне, J.F.A.P. Миллер и У. Р. Хит.
1997
. Ограниченная классом I перекрестная презентация экзогенных аутоантигенов приводит к делеции аутореактивных CD8+ Т-клеток.
J. Расшир. Мед.
186
:
239
–245.
40
Ийода Т., С. Симояма, К. Лю, Ю. Омацу, Ю. Маэда, К. Такахара, Ю. Акияма, Р.М. Штейнман и К. Инаба.
2002
. Субпопуляция дендритных клеток CD8+ избирательно эндоцитирует умирающие клетки в культуре и in vivo.
Дж. Эксп. Мед.
195
:
1289
–1302.
41
Лю, К., Т. Ийода, М. Сатернус, Ю. Кимура, К. Инаба и Р.М. Штейнман.
2002
. Иммунная толерантность после доставки умирающих клеток в дендритные клетки in situ.
J. Расшир. Мед.
196
:
1091
–1097.
42
Scheinecker, C., R. McHugh, E.M. Shevach, and R.N. Жермен.
2002
. Конститутивное представление естественного тканевого аутоантигена исключительно дендритными клетками в дренирующем лимфатическом узле.
J. Расшир. Мед.
196
:
1079
–1090.
43
Инаба, К., С. Терли, Т. Иода, Ф. Ямаиде, С. Шимояма, К. Рейс и Соуза, Р.Н. Жермен, И. Меллман и Р.М. Штейнман.
2000
. Образование иммуногенных пептидных лигандов МНС класса II в лизосомальных компартментах дендритных клеток регулируется воспалительными стимулами.
Дж. Эксп. Мед.
191
:
927
–936.
44
Инаба К., Г. Шулер, М.Д. Витмер, Дж. Валинский, Б. Атасси и Р.М. Штейнман.
1986
. Иммунологические свойства очищенных клеток Лангерганса: различные требования к стимуляции непримированных и сенсибилизированных Т-лимфоцитов.
J. Расшир. Мед.
164
:
605
–613.
45
Романи, Н., С. Койде, М. Кроули, М. Витмер-Пак, А.М. Ливингстон, К.Г. Фатман, К. Инаба и Р.М. Штейнман.
1989
. Презентация экзогенных белковых антигенов дендритными клетками клонам Т-клеток: интактный белок лучше всего представлен незрелыми эпидермальными клетками Лангерганса.
J. Расшир. Мед.
169
:
1169
–1178.
46
Камат, А.Т., Дж. Пули, М.А. О’Киф, Д. Времец, Ю. Жан, А. Лью, А. Д’Амико, Л. Ву, Д.Ф. Жесткий, и К.С. Коротышка.
2000
. Развитие, созревание и скорость оборота популяций дендритных клеток селезенки мыши.
Дж. Иммунол.
165
:
6762
–6770.
47
Reis e Sousa, C., S. Hieny, T. Scharton-Kersten, D. Jankovic, H. Charest, R.N. Жермен и А. Шер.
1997
. Микробная стимуляция in vivo индуцирует быструю CD40L-независимую продукцию IL-12 дендритными клетками и их перераспределение в области Т-клеток.
J. Расшир. Мед.
186
:
1819
–1829.
48
Де Смедт, Т. , Б. Паяк, Э. Мурай, Л. Леспаньяр, Э. Хайнен, П. Де Бетселье, Дж. Урбен, О. Лео и М. Мозер.
1996
. Регуляция количества и созревания дендритных клеток липополисахаридом in vivo.
J. Расшир. Мед.
184
:
1413
–1424.
49
Спарвассер, Т., Р.М. Вабулас, Б. Виллмоу, Г.Б. Липфорд и Х. Вагнер.
2000
. Бактериальная CpG-ДНК активирует дендритные клетки in vivo: независимые от Т-хелперов цитотоксические Т-клеточные ответы на растворимые белки.
Евро. Дж. Иммунол.
30
:
3591
–3597.
50
Акбари О., Н. Панджвани, С. Гарсия, Р. Таскон, Д. Лоури и Б. Стокингер.
1999
. ДНК-вакцинация: трансфекция и активация дендритных клеток как ключевые события иммунитета.
J. Расшир. Мед.
189
:
169
–178.
51
Schubert, U., L.C. Антон, Дж. Гиббс, К.С. Норбери, Дж.В. Юделл и Дж. Р. Беннинк.
2000
. Быстрая деградация большой доли вновь синтезированных белков протеасомами.
Природа.
404
:
770
–774.
52
Цинкернагель Р.М. и П.К. Доэрти.
1979
. MHC-рестриктированные цитотоксические Т-клетки: исследования биологической роли полиморфных основных трансплантационных антигенов, определяющих рестрикционную специфичность, функцию и реактивность Т-клеток.
Доп. Иммунол.
27
:
51
–177.
53
den Haan, J., S. Lehar и M. Bevan.
2000
. CD8+, но не CD8- дендритные клетки перекрестно праймируют цитотоксические Т-клетки in vivo.
J. Расшир. Мед.
192
:
1685
–1696.
APD — Агентство по делам инвалидов
Щелкните здесь, чтобы посмотреть специальное сообщение директора Палмера по случаю 32-й годовщины принятия Закона об американцах-инвалидах.
Агентство по делам инвалидов, профессиональной реабилитации, службы помощи слепым и другие организации инвалидов отметят выдающихся работодателей Флориды в октябре в рамках ежегодного празднования Месяца информирования о трудоустройстве инвалидов. Празднование запланировано на 6 октября в мэрии Таллахасси. Если вам известен работодатель, который имеет опыт найма и поддержки людей с ограниченными возможностями, пожалуйста, заполните форму номинации здесь.
Агентство по делам инвалидов ищет высококлассных сотрудников. Подать заявку на вакансии в Агентстве по делам инвалидов можно здесь:
https://jobs.myflorida.com/go/Agency-for-Persons-with-Disabilities/2814600/
На наш веб-сайт была добавлена новая страница с информацией о посещении объектов, которая включает в себя условия посещения резидентами Sunland Center в Марианне и Tacachale Center в Гейнсвилле.
Если у вас есть вопросы о готовности к стихийным бедствиям, посетите Флоридский отдел управления чрезвычайными ситуациями. Контактный номер информационной линии государственной помощи (SAIL): 1-800-342-3557. Ознакомьтесь с Руководством по обеспечению готовности к чрезвычайным ситуациям во Флориде на сайте FloridaHealth.gov. Также ознакомьтесь с Реестром лиц с особыми потребностями Флориды, чтобы получить важную информацию от местных должностных лиц по чрезвычайным ситуациям.
APD Recovery Toolkit
Агентство поддерживает лиц с нарушениями развития в жизни,
обучении и работе в их сообществах.
|
Щелкните здесь для просмотра местных ресурсов |
Щелкните здесь , чтобы получить оценочную карту WSC. |
Щелкните здесь для государственных ресурсов |
Консультации
|
|
Новости и объявления
-
В 2022 году Законодательное собрание Флориды поручило Агентству по делам инвалидов реализовать стоматологическую программу в масштабах штата.
Законодательное собрание Флориды выделило агентству 8,5 миллионов долларов для заключения на конкурсной основе контракта с некоммерческой организацией на программу стоматологических услуг штата для лиц с нарушениями развития. APD ищет информацию и идеи относительно реализации Государственной стоматологической программы для обслуживания лиц с нарушениями интеллекта и развития. Для получения дополнительной информации, пожалуйста, прочитайте RFI здесь и прочитайте рекламу MFMP здесь. -
Щелкните здесь для получения информации о COVID-19 для семей, поставщиков услуг и WSC. Щелкните здесь, чтобы получить информацию и ресурсы о вакцинах. Для получения дополнительной информации о COVID-19 обязательно посетите веб-сайт Министерства здравоохранения Флориды и веб-сайт CDC. Обязательно ознакомьтесь с набором инструментов CDC для людей с ограниченными возможностями.
-
ABLE United предлагает людям с ограниченными возможностями более разумный и безопасный способ сберегать безналоговые сбережения, сохраняя при этом важные льготы, на которые они полагаются.
Как правило, средства на счете ABLE не считаются ресурсом или активом для Medicaid или Supplemental Security Income. Средства могут быть использованы для различных расходов, которые помогают людям улучшить или сохранить свое здоровье, независимость или качество жизни. Есть вопросы? Посмотрите нашу записанную презентацию на английском и/или испанском языках, запланируйте беседу один на один с ABLE United или запросите презентацию для своей группы или организации, чтобы узнать больше о том, почему вы или те, кого вы обслуживаете, должны рассмотреть возможность открытия учетной записи ABLE сегодня. -
Программа инвалидов и здоровья (DHP) финансируется за счет пятилетнего гранта Центров по контролю и профилактике заболеваний (CDC). Целью программы является вовлечение жителей Флориды всех возрастов с ограниченными возможностями во все программы и мероприятия Министерства здравоохранения, направленные на укрепление здоровья, профилактику заболеваний, оздоровление и обеспечение готовности к стихийным бедствиям.