
Эпигенетическая теория развития личности: Эпигенетическая теория развития личности Эриксона
1.2 Теория развития личности Эрика Эриксона среди других подходов. Теория развития личности Э. Эриксона
Теория развития личности Э. Эриксона
курсовая работа
Эпигенетическая теория развития личности Э. Эриксона — одна из наиболее авторитетных, зарекомендовавших себя теорий развития личности. Важно при этом понять, какой подход она реализует, на какие вопросы она отвечает, а на какие — нет.
Развитие личности интересно не только для психологов. Развитие личности важно и для педагогов, развивающих личность детей разного возраста, развитие личности важно для бизнесменов, заинтересованных в развитии личности своих сотрудников, развитие личности важно и просто для людей, которые хотят развивать свою личность.
Важно отметить, что концепция Э. Эриксона почти полностью находится в психологическом поле, в малой степени ориентируясь на педагогические запросы. Как писал А.В. Петровский, «Следует различать собственно психологический подход к развитию личности и строящуюся на его основе периодизацию возрастных этапов и собственно педагогический подход к последовательному вычленению социально обусловленных задач формирования личности на этапах онтогенеза.
Первый из них ориентирован на то, что реально обнаруживает психологическое исследование на ступенях возрастного развития в соответствующих конкретно-исторических условиях, что есть («здесь и теперь») и что может быть в развивающейся личности в условиях целенаправленных воспитательных воздействий. Второй — на то, что и как должно быть сформировано в личности, чтобы она отвечала всем требованиям, которые на данной возрастной стадии предъявляет к ней общество». Петровский А.В., Ярошевский М.Г. Основы теоретической психологии. М.: ИНФРА-М, 1998.
Тем не менее, скрытым образом модель Эрика Эриксона предлагает некоторые педагогические решения. Перечисляя общие для всех людей, естественные этапы развития личности, Эриксон по факту подсказывает своим читателям, в каком возрасте следует принимать какие решения, чтобы следующий этап его жизни прошел благополучно, а не криво. Поскольку нет никаких данных, что подобные выборы могут делаться лишь бессознательно и не могут быть сделаны сознательно (действительно, а почему бы и да?), то определенный, замаскированный педагогический смысл в модели Э. Эриксона содержится.
Эриксона содержится.
С другой стороны, видимо, нет смысла изучать эту теорию тому, кто хочет стать Личностью с большой буквы, кто изучает пути саморазвития и самосовершенствования личности. Развитие личности может быть результатом как естественного, пассивного, — так и активного личностного роста и развития личности, происходящего вследствие авторских инициатив самого человека.
В пассивном личностном росте интеллект и психологическая культура растут естественно, так же как растет тело человека. Вместе с телом потихоньку развивается интеллект, постепенно в процессе жизни естественно формируется общая и психологическая культура. Психологическую картину этого процесса описывают разные психологи по своему, но общая линия одна: человек проходит свой естественный путь от детства к отрочеству, от юности к зрелости, а далее к старости.
Концепция Э. Эриксона не является пособием для тех, кто хочет развивать себя, как личность — концепция развития личности Э. Эриксона описывает естественный, пассивный рост естественной психологически здоровой личности, фиксируя лишь успешное прохождение ею важных этапов и отмечая, когда возможны сбои в психологическом здоровье и человеку требуется психотерапия. Хьелл Л., Зиглер Д. Теории личности. — 3-е изд. — СПб.: Питер, 2006. — 607.
Хьелл Л., Зиглер Д. Теории личности. — 3-е изд. — СПб.: Питер, 2006. — 607.
Делись добром 😉
«Идентичность» Эрика Эриксона
1. «Идентичность» Эрика Эриксона
В данной работе, посвященной проблеме идентичности, изучением которой занимался Эрик Эриксон, выдающийся психоаналитик, рассуждение будет строиться, в первую очередь, на основании работы вышеупомянутого автора — «Детство и общество»…
«Идентичность» Эрика Эриксона
2. Резюме по части «Идентичность» Эрика Эриксона
Во-первых, идентичность строится всю жизнь, независимо от того, на каком этапе развития находится индивид: в любой момент могут возникнуть изменения, как социальные, так и глубоко индивидуальные (которые понимаются как «свободный выбор»)…
Влияние фрейдизма: теории Э. Эриксона, К.Г. Юнга,Г. Мюррея
1. Эпигенетическая теория развития личности Э.Эриксона
Предметом этой психологии является развитие личности через кризисы. Задачи…
Кризис личности
1.
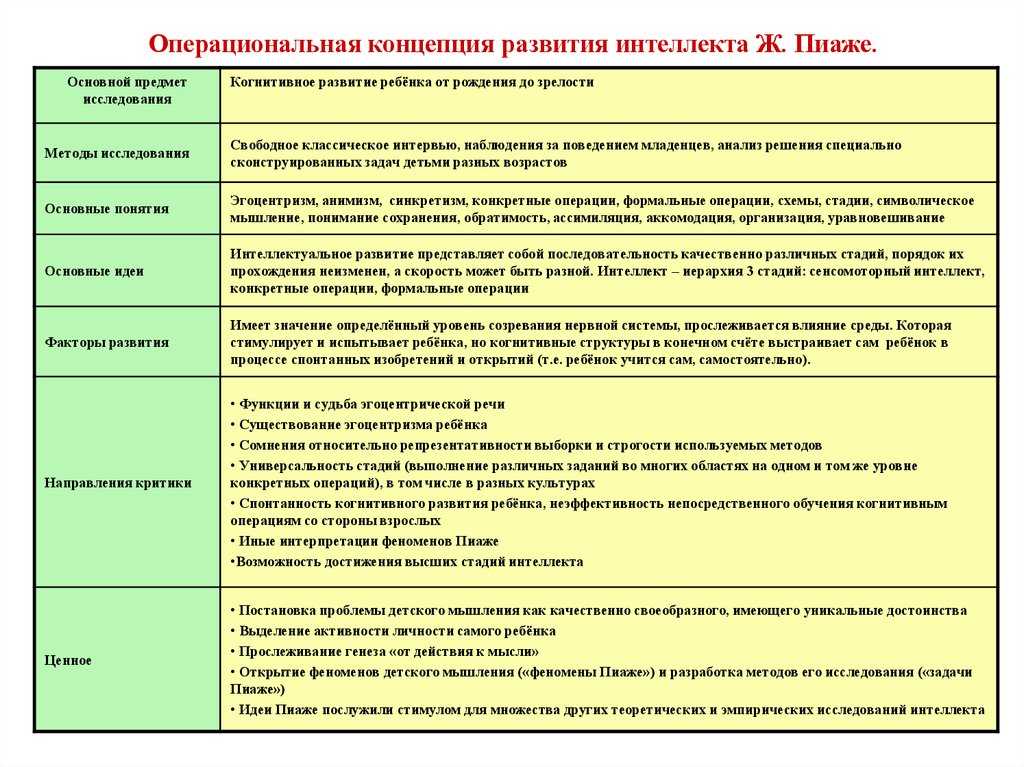 2.4 Теория Эрика Эриксона
2.4 Теория Эрика ЭриксонаТеория Эрика Эриксона о делении нашего жизненного цикла на восемь стадий является одним из крупнейших вкладов в психоанализ и в психологию в целом. В отличие от Фрейда, Эриксон полагает, что характер человека хотя и формируется в детстве…
Место и роль педагогической психологии в системе современных научных знаний
2.1 Педагогическая психология среди других человековедческих наук
В современной науке все нагляднее просматривается взаимодействие двух основных тенденций ее развития: интеграции и дифференциации научных отраслей, дисциплин, проблемных областей (наряду с такими тенденциями, как систематизация…
Особенности развития этнопсихологии
Место этнопсихологии среди других наук
Этнопсихология — междисциплинарная отрасль знания, изучающая этнокультурные особенности психики людей, психологические характеристики этносов, а также психологические аспекты межэтнических отношений…
Особенности структуры самосознания наркозависимых личностей
1.
 4 Теория поэтапного развития Я-концепции Э. Эриксона
4 Теория поэтапного развития Я-концепции Э. ЭриксонаКак формируется чувство собственного «Я»? Одна из наиболее авторитетных теорий, призванных ответить на этот вопрос, была предложена Эриком Эриксоном, автором теории поэтапного развития Я-концепции. Он утверждал, что, несмотря на то…
Принятие роли в юношеской группе в зависимости от уровня идентичности
3.1 Теория Э.Эриксона
В юношеском возрасте центральным моментом в развитии самосознания становится формирование эго-идентичности. В детстве мы судим о себе в соответствии с совокупностью различных ролей…
Проблема акмеологии кризиса на этапе взрослости. Особенности развития личности
1. Психосоциальная концепция развития личности Эриксона
Психосоциальная концепция развития личности, разработанная известным психологом Эриксоном, показывает тесную связь психики человека и характера общества, в котором он живет. Эриксон ввел понятие «групповая идентичность»…
Психологические закономерности делового общения
2.
 2 МЕСТО ЕСТЕСТВЕННОГО ЯЗЫКА СРЕДИ ДРУГИХ ЗНАКОВЫХ СИСТЕМ
2 МЕСТО ЕСТЕСТВЕННОГО ЯЗЫКА СРЕДИ ДРУГИХ ЗНАКОВЫХ СИСТЕМ коммуникация общение речь невербальный В семиотике выделяют несколько признаков естественного человеческого языка, отличающих его от других знаковых систем и, прежде всего от систем коммуникации у животных…
Психология как наука
2. Место психологии среди других наук
От того, какое место отводится психологии в системе наук, во многом зависит понимание возможностей использования психологических данных в других науках и, наоборот, понимание того, в какой мере правомочна психология использовать их результаты…
Современные теории личности
1. Современные теории личности: теория черт личности, фрейдизм, гуманистические теории личности, теория социального научения
…
Теории личности З. Фрейда и Э. Эриксона
Глава 2. Эпигенетическая периодизация развития личности Э. Эриксона
…
Теория развития личности Э. Эриксона
1. Психосоциальная теория развития личности э.
 эриксона
эриксона…
Теория развития личности Э. Эриксона
1.1 Теория психосоциального развития Э. Эриксона
Эрик Эриксон (1902-1994) считается неофрейдистом, потому что его исследование развития личности отталкивается от теории Фрейда, но в дальнейшем развилось в самостоятельное научное направление. Эрик-сон…
|
Главная | Контакты | FAQ |
||
|
Эпигенетическая теория развития личности Э.Эриксона – одна из наиболее авторитетных, зарекомендовавших себя теорий развития личности. Важно при этом понять, какой подход она реализует, на какие вопросы она отвечает, а на какие – нет. Развитие личности интересно не только для психологов. Развитие личности важно и для педагогов, развивающих личность детей разного возраста, развитие личности важно для бизнесменов, заинтересованных в развитии личности своих сотрудников, развитие личности важно и просто для людей, которые хотят развивать свою личность. Важно отметить, что концепция Э. Эриксона почти полностью находится в психологическом поле, в малой степени ориентируясь на педагогические запросы. Как писал А.В.Петровский, «Следует различать собственно психологический подход к развитию личности и строящуюся на его основе периодизацию возрастных этапов и собственно педагогический подход к последовательному вычленению социально обусловленных задач формирования личности на этапах онтогенеза. Первый из них ориентирован на то, что реально обнаруживает психологическое исследование на ступенях возрастного развития в соответствующих конкретно-исторических условиях, что есть («здесь и теперь») и что может быть в развивающейся личности в условиях целенаправленных воспитательных воздействий. Второй — на то, что и как должно быть сформировано в личности, чтобы она отвечала всем требованиям, которые на данной возрастной стадии предъявляет к ней общество». Тем не менее, скрытым образом модель Эрика Эриксона предлагает некоторые педагогические решения. С другой стороны, видимо, нет смысла изучать эту теорию тому, кто хочет стать Личностью с большой буквы, кто изучает пути саморазвития и самосовершенствования личности. Развитие личности может быть результатом как естественного, пассивного, — так и активного личностного роста и развития личности, происходящего вследствие авторских инициатив самого человека. В пассивном личностном росте интеллект и психологическая культура растут естественно, так же как растет тело человека. Вместе с телом потихоньку развивается интеллект, постепенно в процессе жизни естественно формируется общая и психологическая культура. Концепция Э.Эриксона не является пособием для тех, кто хочет развивать себя, как личность – концепция развития личности Э.Эриксона описывает естественный, пассивный рост естественной психологически здоровой личности, фиксируя лишь успешное прохождение ею важных этапов и отмечая, когда возможны сбои в психологическом здоровье и человеку требуется психотерапия. Клиенты Эриксона не читали А.Н.Леонтьева «Личностью не рождаются, личностью становятся!» и не ставили задачу «создать себя». К Эрику Эриксону приходили клиенты с переживанием «потери себя», с желанием найти себя, почувствовать собственную идентичность. Эриксону было что им предложить: созданная им концепция давала понятные для его клиентов, успокаивающие их объяснения («у вас закономерный для вашего возраста кризис, как у всех»), аккуратно подсказывая пути выхода по позитивному сценарию («Благоприятное разрешение вашего конфликта – любовь»). Концепция Эрика Эриксона напрямую не ставит перед собой педагогических, обучающих или развивающих задач, она констатирует существующее положение как норму и отмечает неудачные, неадаптивные, нежелательные варианты развития. Концепция Эрика Эриксона – пособие для скорее для психотерапевтической деятельности, а не специалистов по развитию личности. При этом эта концепция дала разумную, красивую, педагогически привлекательную картину развития личности и помогла очень многим людям. 23. Теория личности в гуманистической психологии (А.Г.Маслоу).
Гуманистическая психология подходит к человеку как существу, который от природы хорош и способен к самосовершенствованию. С точки зрения гуманистической психологии сама сущность человека постоянно движет его в направлении личного роста, творчества и самодостаточности, если только чрезвычайно сильные обстоятельства окружения не мешают этому. Гуманистическая теория личности А. Маслоу (1908 А. Маслоу полагал, что люди мотивированы для поиска личных целей, и это делает их жизнь значительной и осмысленной. А. Маслоу предположил, что все потребности человека врожденные и что они организованы в иерархическую систему приоритета или доминирования. В основе этой схемы лежит допущение, что доминирующие потребности, расположенные внизу, должны быть более или менее удовлетворены до того, как человек может осознать наличие и быть мотивированным потребностями, расположенными вверху. Следовательно, потребности одного типа должны быть удовлетворены полностью, прежде чем другая, расположенная выше, проявится и станет действовать. Потребности никогда не бывают удовлетворены по принципу – «все или ничего». Человек может быть мотивирован на двух и более уровнях. Средний человек удовлетворяет свои потребности примерно в следующей степени: • 85 % – физиологические; • 70 % – безопасности и защиты; • 50 % – любви, принадлежности; • 40 % – уважения; • 10 % – самоактуализации. Если потребности более низкого уровня перестанут удовлетворяться, человек вернется на данный уровень и останется там, пока эти потребности не будут в достаточной мере удовлетворены. Таблица 20 Система охраняемых территорий в США Изучение особо охраняемых природных территорий(ООПТ) США представляет особый интерес по многим причинам… Что способствует осуществлению желаний? Стопроцентная, непоколебимая уверенность в своем… Конфликты в семейной жизни. Как это изменить? Редкий брак и взаимоотношения существуют без конфликтов и напряженности. Через это проходят все… Что будет с Землей, если ось ее сместится на 6666 км? Что будет с Землей? — задался я вопросом… Не нашли то, что искали? Воспользуйтесь поиском гугл на сайте: |

 Перечисляя общие для всех людей, естественные этапы развития личности, Эриксон по факту подсказывает своим читателям, в каком возрасте следует принимать какие решения, чтобы следующий этап его жизни прошел благополучно, а не криво. Поскольку нет никаких данных, что подобные выборы могут делаться лишь бессознательно и не могут быть сделаны сознательно (действительно, а почему бы и да?), то определенный, замаскированный педагогический смысл в модели Э. Эриксона содержится.
Перечисляя общие для всех людей, естественные этапы развития личности, Эриксон по факту подсказывает своим читателям, в каком возрасте следует принимать какие решения, чтобы следующий этап его жизни прошел благополучно, а не криво. Поскольку нет никаких данных, что подобные выборы могут делаться лишь бессознательно и не могут быть сделаны сознательно (действительно, а почему бы и да?), то определенный, замаскированный педагогический смысл в модели Э. Эриксона содержится.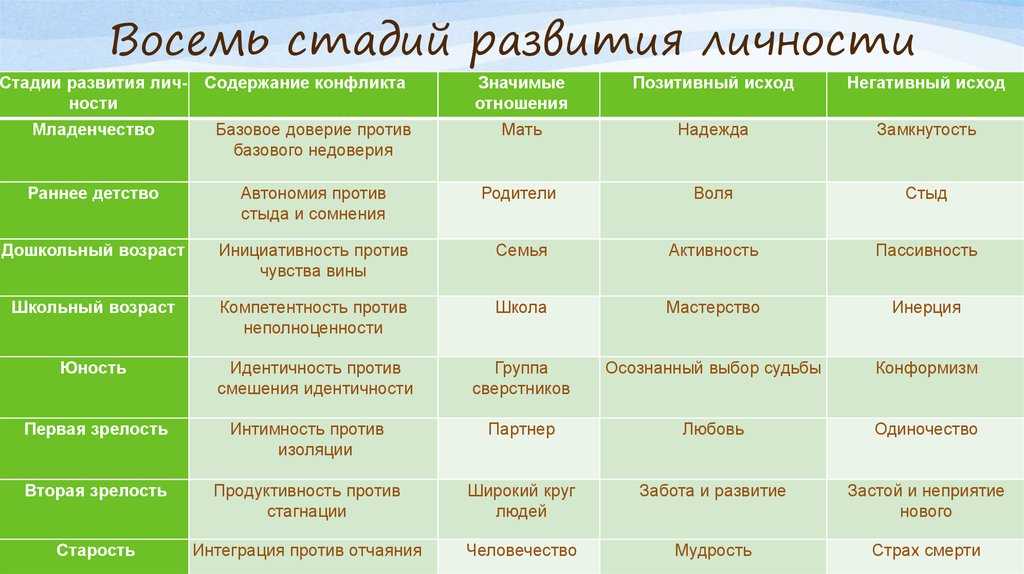

 С точки зрения гуманистической психологии, люди – в высшей степени сознательные и разумные создания без доминирующих бессознательных потребностей и конфликтов. Гуманистическая психология рассматривает людей как активных творцов собственной жизни, обладающих свободой выбирать и развивать стиль жизни, которая ограничена только физическими или социальными воздействиями.
С точки зрения гуманистической психологии, люди – в высшей степени сознательные и разумные создания без доминирующих бессознательных потребностей и конфликтов. Гуманистическая психология рассматривает людей как активных творцов собственной жизни, обладающих свободой выбирать и развивать стиль жизни, которая ограничена только физическими или социальными воздействиями. Удовлетворение потребностей, расположенных внизу иерархии, делает возможным осознание потребностей, расположенных выше в иерархии, и их участие в мотивации. Последовательное расположение основных нужд в иерархии является главным принципом, лежащим в основе организации мотивации человека. А. Маслоу исходил из того, что иерархия потребностей распространяется на всех людей, и чем выше человек может подняться в этой иерархии, тем большую индивидуальность, человеческие качества и психическое здоровье он продемонстрирует.
Удовлетворение потребностей, расположенных внизу иерархии, делает возможным осознание потребностей, расположенных выше в иерархии, и их участие в мотивации. Последовательное расположение основных нужд в иерархии является главным принципом, лежащим в основе организации мотивации человека. А. Маслоу исходил из того, что иерархия потребностей распространяется на всех людей, и чем выше человек может подняться в этой иерархии, тем большую индивидуальность, человеческие качества и психическое здоровье он продемонстрирует.
теории Э. Эриксона, К.Г. Юнга,Г. Мюррея
2
СОДЕРЖАНИЕ
1. Эпигенетическая теория развития личности Э.Эриксона 3
2.Персонология Г.Мюррея 6
3.Аналитическая теория К.Г. Юнга 7
СПИСОК ИСПОЛЬЗОВАННОЙ ЛИТЕРАТУРЫ 12
1. Эпигенетическая теория развития личности Э.Эриксона
Эпигенетическая теория развития личности Э.Эриксона
Предметом этой психологии является развитие личности через кризисы. Задачи, решаемые этим психологическим направлением:
1) исследование кризисных стадий в развитии личности;
2) исследование влияния социокультурного контекста на развитие личности. Развитие личности происходит через кризисы, результаты переживаний которых интегрируются в опыт личности.
С позиций Эриксона, Эго составляет основу поведения и функционирования человека и является автономной личностной структурой, основным направлением развития которой можно назвать социальную адаптацию. Эго взаимодействует с реальностью при помощи восприятия, мышления, внимания и памяти, способствуя возрастанию компетентности человека. Развитие Эго неизбежно связано с социокультурным контекстом и охватывает все жизненное пространство от рождения до смерти.
Человек в процессе жизни проходит восемь универсальных для всего человечества стадий, восемь возрастов.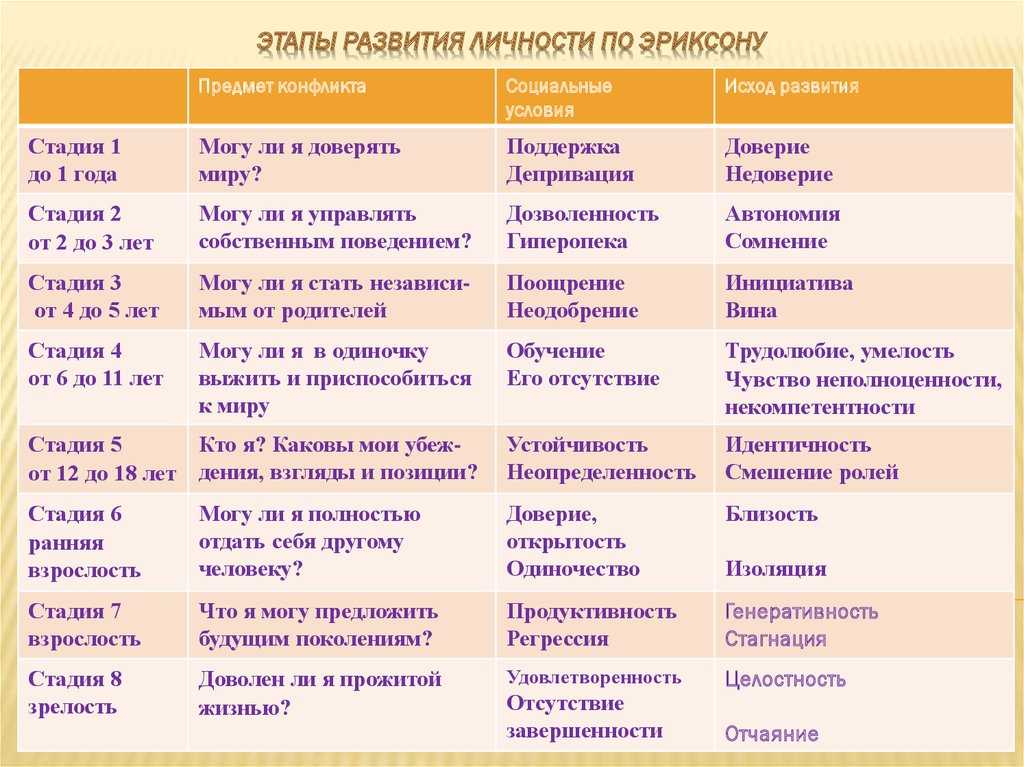 Эпигенетическая концепция развития базируется на представлении о том, что каждая стадия жизненного цикла наступает в определенное для нее время, а также о том, что полноценно функционирующая личность формируется только путем прохождения в своем развитии последовательно всех стадий.
Эпигенетическая концепция развития базируется на представлении о том, что каждая стадия жизненного цикла наступает в определенное для нее время, а также о том, что полноценно функционирующая личность формируется только путем прохождения в своем развитии последовательно всех стадий.
Каждая психосоциальная стадия сопровождается кризисом – поворотным моментом в жизни человека, возникающим как следствие достижения определенного уровня психологической зрелости. Любой кризис представляет собой своего рода вызов, приводящий человека к личностному росту и преодолению жизненных препятствий. На очередной фазе жизненного цикла личность решает специфичную для данного этапа развития эволюционную задачу. Кризис содержит позитивный и негативный компоненты. Если на предыдущей стадии Эго обогатилось новыми положительными качествами и конфликт решен удовлетворительно, то теперь Эго вбирает в себя новый позитивный компонент, например, базальное доверие и автономию, что гарантирует дальнейший личностный рост.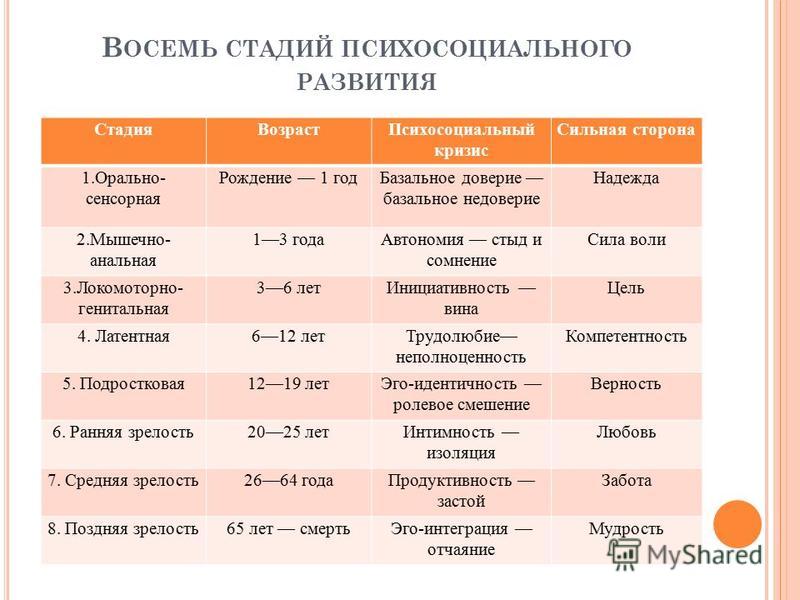 Напротив, если конфликт остался неразрешенным, в Эго встраивается негативный компонент, такой как базальное недоверие, стыд, сомнения.
Напротив, если конфликт остался неразрешенным, в Эго встраивается негативный компонент, такой как базальное недоверие, стыд, сомнения.
Для наступления каждого кризиса существует приоритетное время, обусловленное генетической последовательностью развития. Если первый кризис вовремя не разрешается (как и любой последующий), дилемма доверие-недоверие будет возникать снова и снова на каждой последующей стадии развития.
Обществу и человеку придается равное значение в формировании личности на всем протяжении жизни. Результаты первых четырех стадий развития практически полностью детерминированы влиянием социума, а разрешение конфликтов на более поздних возрастных этапах становится все более зависимым от внутренних факторов.
Э.Эриксон в своих взглядах на развитие придерживался так называемого эпигенетического принципа — генетической предопределенности стадий, которые в своем личностном развитии обязательно проходит человек от рождения до конца своих дней. Наиболее существенный вклад Э. Эриксона в теорию личностного развития состоит в выделении и описании восьми жизненных психологических кризисов, неизбежно наступающих у каждого человека:
Эриксона в теорию личностного развития состоит в выделении и описании восьми жизненных психологических кризисов, неизбежно наступающих у каждого человека:
1. Кризис доверия — недоверия (в течение первого года жизни).
2. Автономия в противоположность сомнениям и стыду (в возрасте около 2- 3 лет).
3. Появление инициативности в противовес чувству вины (примерно от 3 до 6 лет).
4. Трудолюбие в противоположность комплексу неполноценности (возраст от 7 до 12 лет).
5. Личностное самоопределение в противоположность индивидуальной серости и конформизму (от 12 до 18 лет).
6. Интимность и общительность в противовес личностной психологической изолированности (около 20 лет).
7. Забота о воспитании нового поколения в противоположность «погружению в себя» (между 30 и 60 годами).
8. Удовлетворенность прожитой жизнью в противоположность отчаянию (старше 60 лет).
Становление личности в концепции Эриксона понимается как смена этапов, на каждом из которых происходит качественное преобразование внутреннего мира человека и радикальное изменение его отношений с окружаюшими людьми. В результате этого он как личность приобретает нечто новое, характерное именно для данного этапа развития и сохраняющееся у него в течение всей жизни.
В результате этого он как личность приобретает нечто новое, характерное именно для данного этапа развития и сохраняющееся у него в течение всей жизни.
Сами личностные новообразования, по Э.Эриксону, возникают не на пустом месте — их появление на определенной стадии подготовлено всем процессом предшествующего развития личности. Новое в ней может возникнуть и утвердиться, лишь когда в прошлом уже были созданы соответствующие психологические и поведенческие условия.
Формируясь и развиваясь как личность, человек приобретает не только положительные качества, но и недостатки. Детально представить в единой теории всевозможные варианты индивидуального личностного развития по всевозможным сочетаниям положительных и отрицательных новообразований практически невозможно. Имея в виду эту трудность, Э.Эриксон изобразил в своей концепции только две крайние линии личностного развития: нормальную и аномальную. В чистом виде они в жизни почти не встречаются, но зато вмещают в себя всевозможные промежуточные, варианты личностного развития человека.
Предметом этой психологии является взаимодействие личности с окружающей средой через ее потребности. Основные задачи, решаемые этим направлением:
1) изучение структуры личности;
2)изучение динамики личности;
2)диагностика личности через ее потребности.
В персонологии Г.Мюррея – личность — это система, взаимодействующая с окружающей средой через ее потребности. Развитие личности происходит через развитие потребностей и побуждающие свойства внешней среды
Эта теория личности во многом заимствовала основные положения психоанализа 3. Фрейда.
Главный вклад в теорию Мюррея связан с разработкой понятия «потребность», которая определяется как сущность, организующая восприятие, апперцепцию, мышление, способность к волевому решению и действию таким способом, чтобы трансформировать в определенном направлении существующую неудовлетворительную ситуацию. На потребности влияют «прессы», т. е. реальные или воспринимаемые силы окружающей среды, имеющие побуждающие свойства. Автор предложил список сорока потребностей (в достижении, власти, аффилиации, агрессии, автономии, защите, уважении, опеке и др.), а также перечень прессов (отсутствие семейной поддержки, опасность беды, недостаток или потеря чего-либо, различные сдерживания, отказы и др.). Потребности и соответствующие им прессы объединяются в единице поведения, называемой «тема», которая вместе с набором устойчивых значений и образцов действия образует «потребностный интеграт». На основании этих базовых понятий Мюррей строит векторно-ценностную схему поведения личности.
Автор предложил список сорока потребностей (в достижении, власти, аффилиации, агрессии, автономии, защите, уважении, опеке и др.), а также перечень прессов (отсутствие семейной поддержки, опасность беды, недостаток или потеря чего-либо, различные сдерживания, отказы и др.). Потребности и соответствующие им прессы объединяются в единице поведения, называемой «тема», которая вместе с набором устойчивых значений и образцов действия образует «потребностный интеграт». На основании этих базовых понятий Мюррей строит векторно-ценностную схему поведения личности.
В структуре личности, так же как и в теории Фрейда, Мюррей выделяет Ид, Эго и Супер-эго. Однако его концепция имеет отличия от ортодоксального психоанализа: не все инстинкты Ид эгоистичны и социально неприемлемы, следовательно, функционирование личности не всегда определяется попыткой избежать конфликта между индивидом и обществом; функция Эго является не только защитной, но и организующей, контролирующей и планирующей; ценности и табу, которые содержит Супер-эго, не фиксируются в детстве, поэтому ровесники, значимые взрослые и даже литературные герои могут впоследствии влиять на Супер-эго; на основании этого влияния формируется Эго-идеал, который дает широкие возможности для личностных изменений даже на относительно поздних возрастных этапах.
Развитие личности, по теории Мюррея, состоит из семи этапов, полученных путем добавления к концепции развития личности Фрейда еще двух стадий: клаустральная (внутриутробный период жизни), в которой какая-либо определенная эрогенная зона не играет роли, но имеется пассивная зависимость от матери; уретральная (между оральной и анальной стадиями), в которой эрогенной зоной является мочевая система, и основные действия, связанные с сексуальным удовлетворением, включают мочевыделение.
Соответственно двум дополнительным стадиям Мюррей описывает два комплекса и соответствующие им типы характера:
1) клаустральный, в котором главным защитным механизмом является отрицание, а формирующиеся черты выражают пассивность и уход;
2) уретральный, в нем главные защиты напоминают анальный тип характера, и акцентируются черты, характеризующие дух соперничества или уступчивости. Мюррей разработал много диагностических методик для исследования личности, среди которых самой известной считается «Тематической апперцепции тест» (ТАТ).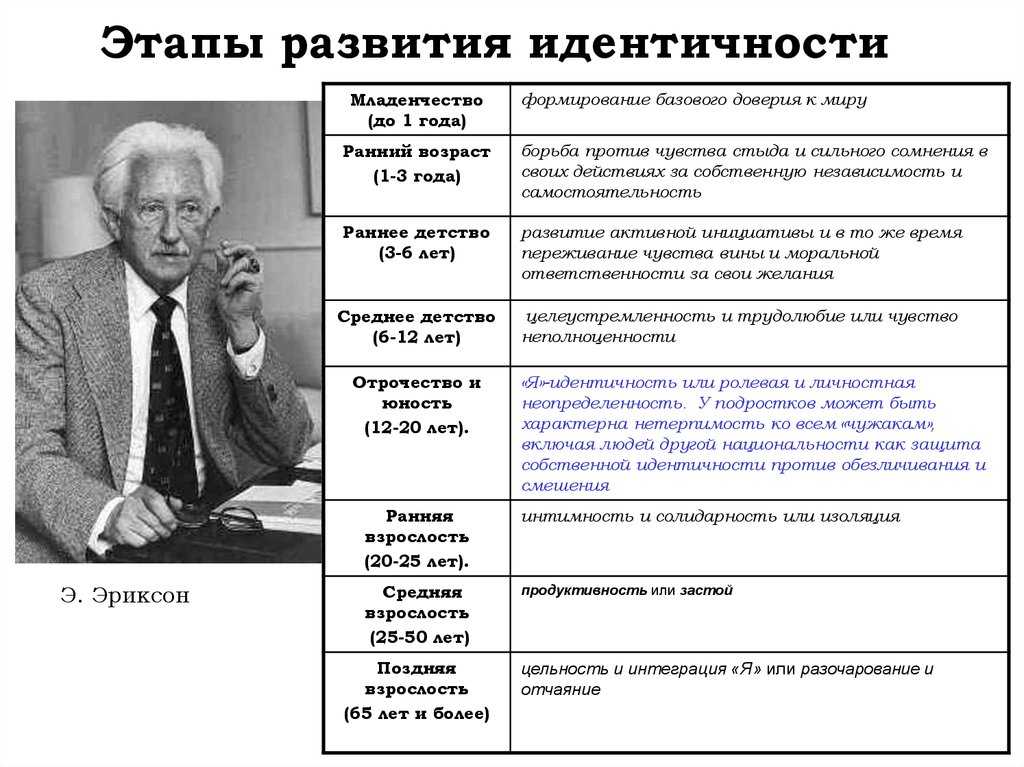
3.Аналитическая теория К.Г. Юнга
Предметом анаитической психологии являются бессознательные аспекты структуры и динамики личности. Основные задачи данного направления: 1)исследование роли коллективного бессознательного; 2)аналитическая терапия. Личность в целом, или психика, как называет ее Юнг, состоит из нескольких дифференцированных, но взаимосвязанных систем. Наиболее важные: Я, личное бессознательное и его комплексы, коллективное бессознательное и его архетипы, маска, анима и анимус, тень. Кроме этих взаимосвязанных систем, существуют установки – интроверсия и экстраверсия, и функции – мышление, чувство, ощущение и интуиция. Наконец, существует самость – центр всей личности.
Аналитическая психология создана швейцарским психиатром и психологом Юнгом и разрабатывается его многочисленными последователями. В психоанализе и аналитической психологии есть общие категории и объяснительные принципы. Динамический аспект функционирования психики в обоих направлениях представлен понятием либидо. Юнг доказал существование коллективного бессознательного, возникшего в результате эволюции биологического вида «человек» и представляющего собой накапливающийся опыт человечества в форме архетипов. Для объяснения движения энергии Юнг ввел принцип полярности: каждому желанию, свойству соответствует его противоположность. Если человек сознательно стремится к определенной цели, то в бессознательном присутствует противоположное намерение, чем объясняется бессознательный срыв сознательных намерений. Этот принцип распространяется и на взаимоотношения архетипических фигур (например, Тень – противоположность Эго и Персоны), и на психические функции, и на свойства (если мужчина сознательно культивирует маскулинность, то эмоциональность, импульсивность вытесняется им в бессознательное). Невротическое развитие рассматривается как односторонность, когда доминирует одна тенденция. Накопленная в индивидуальном и коллективном бессознательном энергия может переместиться в сознание и разрушить рациональный процесс, потому что ее действие примитивно.
Юнг доказал существование коллективного бессознательного, возникшего в результате эволюции биологического вида «человек» и представляющего собой накапливающийся опыт человечества в форме архетипов. Для объяснения движения энергии Юнг ввел принцип полярности: каждому желанию, свойству соответствует его противоположность. Если человек сознательно стремится к определенной цели, то в бессознательном присутствует противоположное намерение, чем объясняется бессознательный срыв сознательных намерений. Этот принцип распространяется и на взаимоотношения архетипических фигур (например, Тень – противоположность Эго и Персоны), и на психические функции, и на свойства (если мужчина сознательно культивирует маскулинность, то эмоциональность, импульсивность вытесняется им в бессознательное). Невротическое развитие рассматривается как односторонность, когда доминирует одна тенденция. Накопленная в индивидуальном и коллективном бессознательном энергия может переместиться в сознание и разрушить рациональный процесс, потому что ее действие примитивно. Например, человек стремится быть уступчивым, мягким. Тогда активность, твердость может проявиться в несоответствующей ситуации агрессивной реакцией. В то же время противоположности, компенсируя друг друга, создают напряжение, способствующее росту. Если человек сознательно игнорирует какую-либо сферу, например, сексуальность, то бессознательное через сновидения напоминает о ней. Единство личности обеспечивается трансцендентной функцией. Она преодолевает амбивалентные тенденции, внутренний конфликт, связывая полярности психики через символическое выражение психического содержания. Юнг подчеркивал ее естественный характер, не основанный на сознательном намерении. Проявляясь для преодоления односторонности в развитии, трансцендентная функция дает новую перспективу, которая не предполагалась, не была целью личности и не может быть исчерпывающе понята при помощи сознания. Наиболее реально ее действие обнаруживается в описании опыта просветления в восточных текстах. Основные понятия аналитической психологии:
Например, человек стремится быть уступчивым, мягким. Тогда активность, твердость может проявиться в несоответствующей ситуации агрессивной реакцией. В то же время противоположности, компенсируя друг друга, создают напряжение, способствующее росту. Если человек сознательно игнорирует какую-либо сферу, например, сексуальность, то бессознательное через сновидения напоминает о ней. Единство личности обеспечивается трансцендентной функцией. Она преодолевает амбивалентные тенденции, внутренний конфликт, связывая полярности психики через символическое выражение психического содержания. Юнг подчеркивал ее естественный характер, не основанный на сознательном намерении. Проявляясь для преодоления односторонности в развитии, трансцендентная функция дает новую перспективу, которая не предполагалась, не была целью личности и не может быть исчерпывающе понята при помощи сознания. Наиболее реально ее действие обнаруживается в описании опыта просветления в восточных текстах. Основные понятия аналитической психологии:
Либидо – У Юнга либидо понимается не как сексуальная энергия, а как гипотетическая категория, описывающая различные виды психической активности. Либидо существует в потенциальной форме (желание) и форме активных действий.
Либидо существует в потенциальной форме (желание) и форме активных действий.
Эго – архетип выбора, принятия решения.
Я – это сознающий ум. Оно строится из сознательных перцепций, воспоминаний, мыслей и чувств. Я отвечает за чувство самотождественности и непрерывности и с точки зрения индивидуального человека рассматривается как центр сознания.
Личное бессознательное – бессознательное отдельной личности, индивидуальное, неуниверсальное. Личное бессознательное – это регион, примыкающий к Я. Оно состоит из переживаний, некогда бывших сознательными, но вытесненных, подавленных, забытых или игнорируемых, и из переживаний, которые при появлении своем были слишком слабы, чтобы произвести впечатление на уровне сознания.
Коллективное бессознательное – Юнг доказал существование коллективного бессознательного, возникшего в результате эволюции биологического вида «человек» и представляющего собой накапливающийся опыт человечества в форме архетипов.
Комплекс – это организованная группа или чувств, мыслей, перцепций, воспоминаний, существующая в личном бессознательном. У него есть ядро, действующее как своего рода магнит, притягивающий или различные переживания.
У него есть ядро, действующее как своего рода магнит, притягивающий или различные переживания.
Архетипы – Архетипы являются формами восприятия и осмысления действительности. Они повторяют, разрабатывают сходный опыт переживаний в разных поколениях. Человек, не реализующий архетипы, не дающий им выхода через сознание, создает напряжение, которое может разрушить Эго.
Психические функции – интуиция, ощущения, мышление и чувство.
Индивидуация – Развитие личности, проявляющееся в сближении сознания и бессознательного, Юнг назвал индивидуацией. Индивидуация протекает как осознание ранее неосознанных потребностей, стремлений, чувств, дальнейшее их расщепление на сознательную и бессознательную части. Такая дифференциация частей психики позволяет человеку преодолеть полярности, более тонко воспринимать мир и выражать себя. Индивидуация основывается на ассимиляции индивидуального и коллективного опыта, в результате которого формируются баланс и целостность личности.
Самость – Уникальный потенциал, заложенный в центре личности, открывающийся при индивидуации.
Тень – один из важнейших архетипов, обусловливающих принятие или непринятие себя человеком. Юнг относил к проявлениям Тени разнообразные асоциальные проявления и поступки человека. Также он считал Тень энергетическим источником человека.
В аналитической психологии предполагается, что развитие личности имеет определенное направление, основывающееся на действии трансцендентной функции. На пути индивидуации личность переживает столкновение с архетипическими фигурами. В них сконцентрированы наиболее важные мотивы, коллизии человеческой жизни: отношение к другим, их влияние на человека (Персона), выбор, принятие решений (Эго) отношение к себе, отвержение или принятие себя (Тень), половая идентичность как деление на мужское и женское начала (Анима/Анимус), отношение к сверхъестественному, находящемуся за пределами понимания человека (Самость). Индивидуация рассматривается как проявление уникального потенциала, заложенного в центре личности – Самости.
Динамика личности может происходить по противоположному пути. При идентификации Эго с Самостью происходит регрессия к коллективному бессознательному. Такой способ развития Юнг определил как инфляцию. Захваченный архетипами человек может переживать власть над собой чего-то мощного и надличного, потерю себя либо собственную надличную силу. При инфляции Эго может идентифицироваться с Самостью и человек чувствует себя всесильным, избранным, наделенным особыми способностями и властью. Идеи и образы коллективной психики кажутся порождением собственного сознания, неоспоримыми истинами.
При идентификации Эго с Самостью происходит регрессия к коллективному бессознательному. Такой способ развития Юнг определил как инфляцию. Захваченный архетипами человек может переживать власть над собой чего-то мощного и надличного, потерю себя либо собственную надличную силу. При инфляции Эго может идентифицироваться с Самостью и человек чувствует себя всесильным, избранным, наделенным особыми способностями и властью. Идеи и образы коллективной психики кажутся порождением собственного сознания, неоспоримыми истинами.
Эпигенетическая теория развития личности Э.Эриксона
Детство и ранние годы
Эрик Эриксон родился в г. Франкфурт в Германии, в семье Карлы Абрахамсен и биржевого брокера, еврея Вальдемара Исидора Саломонсена. На тот момент, когда мальчик появился на свет, его родители уже несколько месяцев не видели друг друга. Записали его как Эрика Саломонсена, однако подлинной информации о его биологическом отце нет. Вскоре после рождения сына, его мать перебирается в Карлсруэ, где устраивается работать медсестрой и выходит замуж второй раз, за педиатра Теодора Хомбургера.
В 1911 г. Хомбургер официально усыновляет мальчика, и тот становится Эриком Хомбургером. Историю его рождения от него тщательно скрывают, и мальчик растёт, не зная, кто его настоящий отец.
Основные стадии развития личности по Эрику Эриксону
стадии развития личности
Каждая фаза формирования стадии развития личности (по Эриксону) подразумевает, что человек будет испытывать кризис, который повлечет за собой положительный или отрицательный результат. Эти кризисы имеют психосоциальную природу, потому что они связаны с психологическими потребностями индивида, противоречащими общественным потребностям.
Согласно теории, благополучное завершение каждой стадии приводит к формированию здоровой личности и приобретению основных добродетелей. Основные добродетели – это характерные сильные стороны, которые эго может использовать для разрешения последующих кризисов.
Доверие и недоверие
В течение первого года жизни основной упор делается на воспитательные способности матери и отца, особенно в плане визуального контакта и осязания. Ребенок будет развивать оптимизм, доверие, уверенность и безопасность, если о нем будут заботиться и общаться с ним. Если ребенок не испытывает доверия, у него могут развиться такие качества, как незащищенность, бесполезность и общее недоверие к миру.
Ребенок будет развивать оптимизм, доверие, уверенность и безопасность, если о нем будут заботиться и общаться с ним. Если ребенок не испытывает доверия, у него могут развиться такие качества, как незащищенность, бесполезность и общее недоверие к миру.
Самостоятельность (автономия) и нерешительность (стыд и сомнения)
Второй этап происходит от 18 месяцев до 3 лет. На этом этапе ребенок развивается физически, становится более мобильным и обнаруживает, что у него много навыков и способностей, таких как надевание одежды и обуви, игра с игрушками и т. д. Такие навыки иллюстрируют растущее осознание независимости и самостоятельности ребенка.
Эриксон утверждает, что необходимо, чтобы родители позволили детям изучить свои собственные границы в благоприятной среде, терпимой к неудачам. Т. о., родители должны поощрять малютку становиться более независимым и в то же время защищать ребенка, чтобы он не испытывал постоянных неудач.
Требуется тонкий баланс от родителей. Они должны стараться не делать все за ребенка, но если дитя не справляется с какой-либо задачей, они не должны критиковать его за неудачи (особенно при приучении к туалету). Целью должен быть самоконтроль без потери самооценки. Успех на этом этапе приведет к приобретению такой добродетели, как воля.
Целью должен быть самоконтроль без потери самооценки. Успех на этом этапе приведет к приобретению такой добродетели, как воля.
Если дети подвергаются критике, чрезмерному контролю или не имеют возможности заявить о себе, они становятся чрезмерно зависимыми от других, испытывают чувство стыда или сомнения в своих силах, им сложно найти себя в мире.
Предприимчивость и чувство вины (в другом переводе – неадекватность)
Эриксон считает, что третий психосоциальный кризис происходит во время «игрового возраста», или в дошкольные годы (примерно от 3,5 до 6 лет). Во время этой фазы формирования счастливый ребенок учится:
- представлять, расширять свои навыки посредством всевозможных активных игр, в том числе фантазий;
- сотрудничать с другими;
- руководить другими.
Научная деятельность
Эриксон преподаёт в частной школе в Вене, где знакомится с Анной Фрейд, дочерью Зигмунда Фрейда. Именно она разжигает в нём интерес к психоанализу, и Эриксон отправляется постигать эту науку в Венский институт психоанализа.
В 1933 г., во время его учёбы в институте, в Германии к власти приходит нацистская партия, и Эриксону приходится бежать из страны. Вначале он отправляется в Данию, а после переезжает в США, где становится первым детским психоаналитиком в г. Бостоне.
Проработав там некоторое время, Эриксон меняет должности в различных институтах, в том числе и Общеклинической больнице штата Массачусетс, Центре семейного воспитания судьи Бейкера, Гарвардской медицинской школе и Психологической клинике и т.д.
В 1936 г. Эриксон преподаёт в Гарвардской медицинской школе, а также трудится в Институте межличностных отношений при этом университете. Он также находит время для обучения группы детей в резервации сиу в Южной Дакоте.
В 1937 гг. Эриксон оставляет Гарвард и поступает в штат Калифорнийского университета. Он тесно сотрудничает с Институтом социальной защиты детей и занимается частной практикой. Часть своего времени Эриксон посвящает обучению детей племени юрок.
В 1950 г. его личный опыт общения с представителями разных рас, проживающих в различных социальных условиях, приводит к написанию самой известной книги за всю его научную деятельность, «Детство и общество». В этой книге автор и представляет миру собственную теорию «личностного кризиса».
В этой книге автор и представляет миру собственную теорию «личностного кризиса».
Уволившись из Калифорнийского университета, Эриксон начинает работать и преподавать в Центре Остен Риггс – главном психиатрическом лечебном заведении г. Стокбридж, штат Массачусетс. Там, по роду своей деятельности, он сталкивается с психически неуравновешенными подростками.
В 1960 г. Эриксон возвращается в Гарвардский университет, где проработает до пенсии, после чего, совместно со своей женой, займётся написанием работ на различные темы в психологии.
Основные тезисы теории личности Э.Эриксона
Как и большинство своих современников, Эриксон был психоаналитиком, но его взгляды расходились с классической фрейдовской теорией. Как и Фрейд, Эриксон рассматривал структуру личности исходя из её трёх основных составляющих: Ид, Эго и Супер-Эго. Но в отличие от основоположника психоанализа, Эриксон отдавал ведущую роль в формировании и становлении личности Эго, а не бессознательным инстинктам.
Кроме этого, учёный не был согласен с фрейдовской градацией основных жизненных кризисов, которая насчитывала 5 этапов, а само развитие личности по Фрейду завершалось в пубертальном возрасте. Эриксон был убеждён, что человек развивается всю жизнь и в процессе своего становления переживает 8 кризисов, каждый из которых соответствует определённому возрасту и отвечает за формирование отдельных черт характера.
К тому же учёный полагал, что важную роль в становлении личности играет общество, в котором человек живёт. Эриксон доказывал существование тесной связи между психическим развитием индивида и влиянием социумом. Поэтому хоть главная роль в его теории и принадлежит Эго, но само Я формируется под воздействием специфических культурных и социальных ценностей окружения личности. К такому выводу учёный пришёл, проанализировав особенности воспитания детей в двух различных культурах: коренной индейской и «белой» американской. Эриксон отметил, что каждый стиль воспитания принимается в своём социуме, как единственно правильный и именно он определяет ожидания общества, которые ребёнок должен оправдать. С этим механизмом тесно связано понятие «групповой идентичности» — чувства единения с окружающими людьми. Позже в процессе своего развития, у человека начинает формироваться «эгоидентичность» — чувство целостности и непрерывности своего «Я».
С этим механизмом тесно связано понятие «групповой идентичности» — чувства единения с окружающими людьми. Позже в процессе своего развития, у человека начинает формироваться «эгоидентичность» — чувство целостности и непрерывности своего «Я».
ЭРИКСОН
ЭРИКСОН (Erikson) Эрик Гомбургер (15 июня 1902, Франкфурт-на-Майне – 12 мая 1994, Гарвич, Массачусетс) – немецко-американский психолог и психотерапевт, теоретик неофрейдизма [НЕОФРЕЙДИЗМ]; создатель жанра психоистории. В юности был странствующим художником, в 1927–33 преподавал рисование в школе А.Фрейд [ФРЕЙД А.], одновременно посещал семинары Венского института психоанализа. С 1933 – член Венского психоаналитического общества и Международной психоаналитической ассоциации. После прихода к власти нацистов эмигрировал в США; в 30–40-е гг. занимался психоаналитической практикой в Бостоне, работал в качестве консультанта-психотерапевта в Гарвардской психологической клинике и Йельском институте человеческих отношений. В этот же период участвовал в полевых исследованиях группы культурантропологов в резервациях Южной Дакоты; впоследствии эти наблюдения составили основу его первой книги «Детство и общество» (Childhood and society, 1950, рус. пер. 1996). В 1939–50 преподавал в Калифорнийском (Беркли) и Питтсбургском университетах. На рубеже 50–60-х гг. разработал основные положения своей философско-психологической концепции, в 1958 опубликовал первый опыт психоисторического исследования – книгу «Молодой Лютер» (Young man Luther, рус. пер. 1996). В 1960 возобновил академическую деятельность в Гарварде, где читал курс лекций по теории жизненного цикла. Лауреат премии Пулитцера и Национальной премии США (за книгу «Правда Ганди» – Gandhi’s truth, 1969).
В этот же период участвовал в полевых исследованиях группы культурантропологов в резервациях Южной Дакоты; впоследствии эти наблюдения составили основу его первой книги «Детство и общество» (Childhood and society, 1950, рус. пер. 1996). В 1939–50 преподавал в Калифорнийском (Беркли) и Питтсбургском университетах. На рубеже 50–60-х гг. разработал основные положения своей философско-психологической концепции, в 1958 опубликовал первый опыт психоисторического исследования – книгу «Молодой Лютер» (Young man Luther, рус. пер. 1996). В 1960 возобновил академическую деятельность в Гарварде, где читал курс лекций по теории жизненного цикла. Лауреат премии Пулитцера и Национальной премии США (за книгу «Правда Ганди» – Gandhi’s truth, 1969).
В духе общей для неофрейдизма тенденции к социокультурному переосмыслению классического психоанализа [ПСИХОАНАЛИЗ], под влиянием идей эгопсихологии(А.Фрейд и др.) и сравнительной культурантропологии (М.Мид) Эриксон приходит к выводу о необходимости включить в теорию Фрейда понятия «социальное окружение» и «исторический момент». В центре его философско-психологической концепции – понятие психосоциальной идентичности, обнимающее как субъективное чувство, так и объективно наблюдаемое качество самотождественности и целостности индивидуального Я, сопряженного с верой этого Я в тождественность и целостность того или иного разделяемого с другими образа мира и человека. Идентичность включает такие аспекты, как непрерывность самосознания на фоне меняющихся объектов опыта («эгоидентичность»), норма индивидуального психического развития в онтогенезе («личностная идентичность»), вовлеченность индивидуального бытия в конкретный сегмент истории («психоисторическая идентичность»), наконец, психологическая устойчивость человека перед лицом небытия («экзистенциальная идентичность»).
В центре его философско-психологической концепции – понятие психосоциальной идентичности, обнимающее как субъективное чувство, так и объективно наблюдаемое качество самотождественности и целостности индивидуального Я, сопряженного с верой этого Я в тождественность и целостность того или иного разделяемого с другими образа мира и человека. Идентичность включает такие аспекты, как непрерывность самосознания на фоне меняющихся объектов опыта («эгоидентичность»), норма индивидуального психического развития в онтогенезе («личностная идентичность»), вовлеченность индивидуального бытия в конкретный сегмент истории («психоисторическая идентичность»), наконец, психологическая устойчивость человека перед лицом небытия («экзистенциальная идентичность»).
Эриксон рассматривает жизненный цикл личности как совокупность восьми сменяющих друг друга стадий развития, причем определяющим фактором этого развития личности выступает ее общение со значимыми другими. Кульминацией становления идентичности является юность.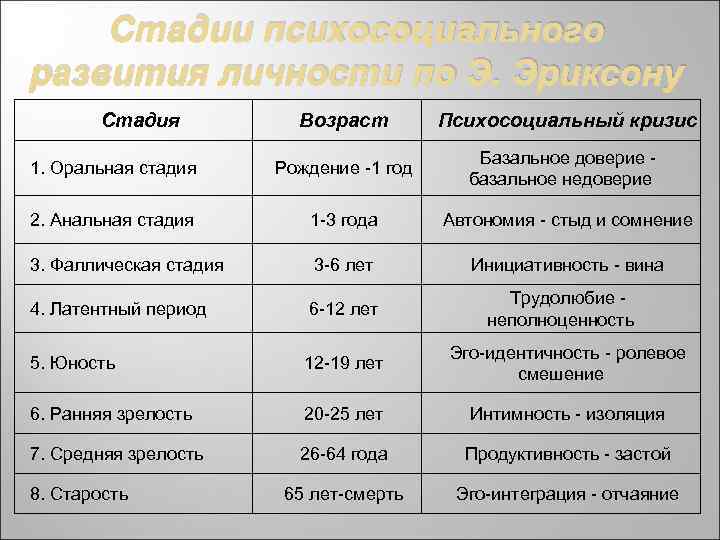 В ходе нормативного юношеского кризиса идентичности возникает качественно иная личностная целостность, открытая последующим трансформациям в ответ на требования «исторического момента». Только в юности бессознательный процесс формирования идентичности может стать фактором социально-исторического самосознания человека и эпохи. Как в психологическом, так и в историческом отношении юность опосредует связь между прошлым и будущим. С этой точки зрения история может быть интерпретирована как континуум индивидуальных жизненных циклов, а историческая эволюция осмыслена в терминах метаболизма поколений. Это образует пограничную область психоистории.
В ходе нормативного юношеского кризиса идентичности возникает качественно иная личностная целостность, открытая последующим трансформациям в ответ на требования «исторического момента». Только в юности бессознательный процесс формирования идентичности может стать фактором социально-исторического самосознания человека и эпохи. Как в психологическом, так и в историческом отношении юность опосредует связь между прошлым и будущим. С этой точки зрения история может быть интерпретирована как континуум индивидуальных жизненных циклов, а историческая эволюция осмыслена в терминах метаболизма поколений. Это образует пограничную область психоистории.
Психоистория выступает вспомогательным инструментом традиционного исторического исследования. Отказавшись от сведения фактов мировой истории культуры к иррациональным импульсам психики гениев, Эриксон рассматривает внутренние конфликты творческих личностей – М.Лютера, Τ.Джефферсона, М.Ганди, Б.Шоу, М.Горького – как выражение социально-психологических и нравственных проблем их времени и культуры. Историческую значимость личности и успех ее социально-культурных и мировоззренческих инноваций Эриксон связывает с адекватной интериоризацией ею духовных противоречий эпохи. Предмет психоистории составляют умонастроения, верования, эмоциональные и нравственные стереотипы, включая их иррациональные составляющие, и преломление этого содержания в исканиях творческой личности.
Историческую значимость личности и успех ее социально-культурных и мировоззренческих инноваций Эриксон связывает с адекватной интериоризацией ею духовных противоречий эпохи. Предмет психоистории составляют умонастроения, верования, эмоциональные и нравственные стереотипы, включая их иррациональные составляющие, и преломление этого содержания в исканиях творческой личности.
Принципы психоисторического анализа получили широкое распространение в американском обществоведении 1970–80-х гг.
Сочинения:
1. Identity and the Life Cycle. N.Y., 1959. Life History and the Historical Moment. N. Y., 1975;
2. в рус. пер.: Идентичность: юность и кризис. М., 1996.
Литература:
1. Анцыферова Л.И. Эпигенетическая концепция развития личности Эрика Г.Эриксона. – В сб.: Принцип развития в психологии. М., 1978, с. 212–242;
2. Якимова Е.В. Критика неофрейдистской концепции человека Э.Г.Эриксона. – «Вестник МГУ», серия философии. М., 1980, № 5, с. 67–75.
67–75.
Е.В.Якимова
Источник: Новая философская энциклопедия на Gufo.me
Значения в других словарях
- Эриксон — (Erixon) Сигурд (26. 3. 1888, Сёдерчёпинг, — 18. 2. 1968, Стокгольм), шведский этнограф. Основал этнографические журналы «Народная жизнь» («Folk-Liv»), «Европейская этнология» («Ethnologia Europaea»), «Лаос» («Laos»), создал школу… Большая советская энциклопедия
- ЭРИКСОН — ЭРИКСОН (Erikson) Эрик Хомбергер (1902-94) — американский психолог и психоаналитик, представитель эгопсихологии. Развил теорию стадиального формирования психосоциальной «групповой идентичности» и параллельно «эгоидентичности» (8 стадий развития личности). Труды по детской психологии. Большой энциклопедический словарь
- ЭРИКСОН — (Erixon), Сигурд (26.III.1888 — 18.II.1968) — швед. этнограф. Автор ряда трудов по материальной культуре, семейному и обществ. быту народов Скандинавии. Сотрудник «Северного музея» (1914-55) и этно-парка «Скансен» (с 1924) в Стокгольме. Советская историческая энциклопедия
- Эриксон — Эрик Хомбергер Эриксон (Erikson) (1902-1994) ☼ амер.
 психолог и психотерапевт, автор одной из первых психол. теорий жизненного цикла, создатель психоистор. модели социального познания. В 30-е гг. Словарь по культурологии
психолог и психотерапевт, автор одной из первых психол. теорий жизненного цикла, создатель психоистор. модели социального познания. В 30-е гг. Словарь по культурологии - ЭРИКСОН — ЭРИКСОН (Erikson) Эрик (1902 — 1994) — американский психолог, социолог и психоаналитик. Основатель психоистории. Профессор Гарвардского университета (I960). Детство и юность провел в Германии. Увлекался живописью. Новейший философский словарь
- Блог
- Ежи Лец
- Контакты
- Пользовательское соглашение
© 2005—2020 Gufo.me
Эпигенетика в расстройствах личности: современные взгляды
Введение
Эпигенетический взгляд на гены, предположительно связанные с психическими расстройствами, вызывает все больший академический интерес и позволяет получить дополнительные сведения о патогенезе конкретного заболевания. Тяжелые психические расстройства оси-I, такие как большое депрессивное расстройство (БДР), в настоящее время широко исследуются на эпигенетическом уровне, и результаты объясняют существенное патогенетическое влияние эпигенетических модификаций генов.
Эпигенетические механизмы в целом функционируют для гомеостатического контроля доступности генов и функциональности транскрипции. Таким образом, функция гена может быть организована как строго запрограммированной и сохраняющейся на протяжении всей жизни, так и реагирующей на окружающую среду (1). Транскриптом как непосредственное представление геномной активности регулируется (i) контролем доступа генов к механизму транскрипции посредством конденсации хроматина и модификации гистонов, такой как (де-)ацетилирование, (де-)фосфорилирование, сумоилирование, (ii) не- кодирование и микроРНК, которые влияют на образование хроматина, а также на трансляцию и деградацию РНК, и (iii) ковалентные изменения ДНК за счет метилирования цитозинового нуклеотида, что затрудняет доступ транскриптазы к метилированной области и может активировать ферменты, которые взаимодействуют, подавляя специфические ген [рассмотрено в (2)].
В то время как эпигенетические паттерны стабильно определяют устойчивую к митозу дифференцировку клеток как предварительное условие эмбрионального развития без изменения последовательности ДНК, эпигеном также представляет собой динамическую адаптацию к условиям окружающей среды. В последние два десятилетия появляется все больше свидетельств того, что ранний жизненный опыт может влиять на долгосрочные траектории соматического и психического здоровья у животных и людей, воздействуя на эпигенетический паттерн и, таким образом, воздействуя на структуру и доступность генома. Исследования на грызунах продемонстрировали решающее влияние материнской заботы и ранних социальных невзгод на развитие потомства и фенотипы взрослых особей. Ранние неблагоприятные условия жизни (ELA) у крыс приводили к увеличению экспрессии гиппокампального глюкокортикоидного рецептора (GR), нарушению функции оси гипоталамус-гипофиз-надпочечники (HPA) и изменению метилирования ДНК гена GR (9).0009 NR3C1) в гиппокампе (3), промотора гена мозгового нейротропного фактора ( BDNF ) в префронтальной коре (4) и гена аргинин-вазопрессина ( AVP ) (5). У людей ELA был связан со снижением экспрессии мРНК GR и гиперметилированием NR3C1 в посмертном человеческом мозге (6). Имеющиеся к настоящему времени данные показывают, что эпигенетические паттерны, обнаруживаемые в связи с ELA у животных и людей, не ограничиваются функциональными генами, вызывающими подозрение на заболевание, но распространяются на весь геном (7, 8) и не являются строго тканеспецифичными (9).
В последние два десятилетия появляется все больше свидетельств того, что ранний жизненный опыт может влиять на долгосрочные траектории соматического и психического здоровья у животных и людей, воздействуя на эпигенетический паттерн и, таким образом, воздействуя на структуру и доступность генома. Исследования на грызунах продемонстрировали решающее влияние материнской заботы и ранних социальных невзгод на развитие потомства и фенотипы взрослых особей. Ранние неблагоприятные условия жизни (ELA) у крыс приводили к увеличению экспрессии гиппокампального глюкокортикоидного рецептора (GR), нарушению функции оси гипоталамус-гипофиз-надпочечники (HPA) и изменению метилирования ДНК гена GR (9).0009 NR3C1) в гиппокампе (3), промотора гена мозгового нейротропного фактора ( BDNF ) в префронтальной коре (4) и гена аргинин-вазопрессина ( AVP ) (5). У людей ELA был связан со снижением экспрессии мРНК GR и гиперметилированием NR3C1 в посмертном человеческом мозге (6). Имеющиеся к настоящему времени данные показывают, что эпигенетические паттерны, обнаруживаемые в связи с ELA у животных и людей, не ограничиваются функциональными генами, вызывающими подозрение на заболевание, но распространяются на весь геном (7, 8) и не являются строго тканеспецифичными (9). ), поскольку было показано, что клетки периферической крови (ПБЦ), особенно Т-клеточные лимфоциты, отражают эпигенетические паттерны, сходные с нервными клетками в культуре и ткани головного мозга (9–11).
), поскольку было показано, что клетки периферической крови (ПБЦ), особенно Т-клеточные лимфоциты, отражают эпигенетические паттерны, сходные с нервными клетками в культуре и ткани головного мозга (9–11).
Преобразование сигналов окружающей среды в эпигенетическую информацию может запускаться активностью нейронов, которая инициирует внутриклеточные пути, такие как ацетилирование гистонов, опосредованное передачей сигналов цАМФ, или которая влияет и взаимодействует с другими эпигенетическими процессами (12–14). Кроме того, активность AVP 9Промотор 0010 регулируется метил-CpG-связывающим белком 2 (MeCp2), который фосфорилируется и активируется за счет деполяризации нейронов гипоталамуса и, в свою очередь, сдерживает деметилирование промотора BDNF (5, 15, 16). В целом, хотя многие детали молекулярных механизмов остаются неизвестными, настоящие открытия подтверждают и уточняют идею о том, как сигналы окружающей среды могут транслироваться во внутриклеточную информацию и молекулярную память.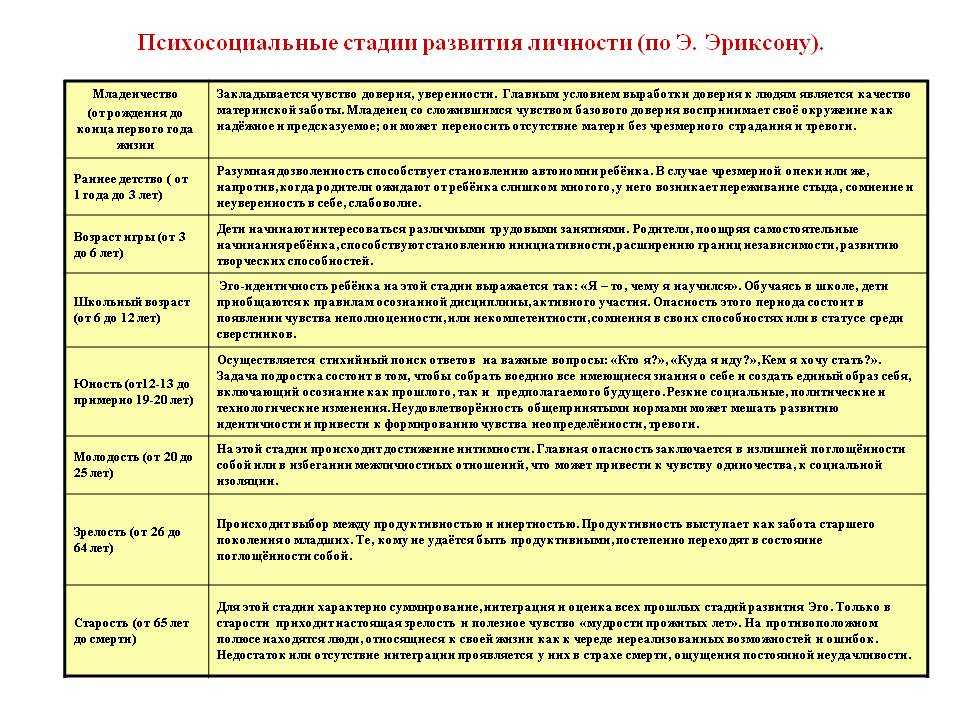
В настоящее время значительное число исследований посвящено изучению эпигенетических изменений, связанных с поведением или влияющих на трудности, такие как агрессия или страх, у людей и животных, особенно в связи с нарушениями серотонинергической системы, которая, как хорошо известно, играет решающую роль в раннем развитии головного мозга. разработка. Целью данной работы был обзор современных оригинальных публикаций об эпигенетических модификациях, связанных с расстройствами личности (PD) у человека.
Методы
Поиск литературы проводился как систематический обзор в соответствии с рекомендациями по предпочтительным элементам записи для систематических обзоров и метаанализов (PRISMA-P) (17). Мы основывали наш поиск на базе данных PubMed Central Национальной медицинской библиотеки Национального института здравоохранения США (NIH/NLM) с использованием терминов, ориентированных на медицинские предметные рубрики (MeSH) библиотеки NCBI.
Для поиска ключевые слова были вставлены в двойную или тройную комбинацию, чтобы получить исчерпывающие совпадения.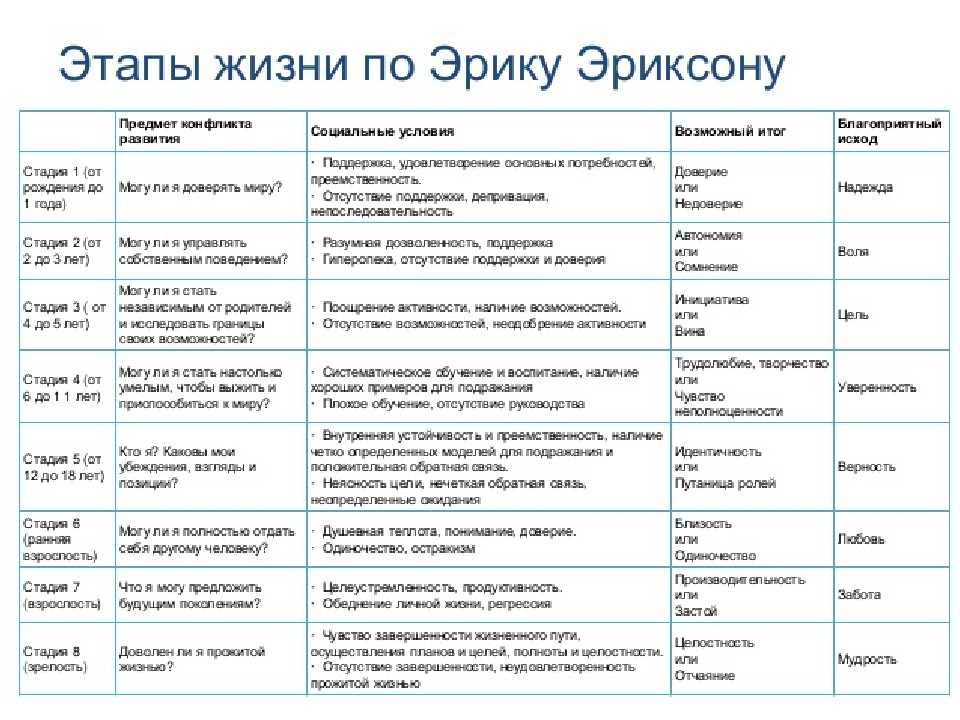 Были использованы следующие ключевые слова: «личность», «расстройство личности», «черта личности», каждое из которых сочеталось с «эпигенетический», «метилирование», «ацетилирование», «фосфорилирование», «убиквитация», «сумоилирование», « микроРНК», «хроматин» и «ремоделирование хроматина» соответственно, а также с одним из ключевых слов «агрессивность», «ананкастность», «асоциальный», «тревожный», «избегающий», «пограничный», «зависимый», «эксцентричный», «эмоционально неустойчивый», «истеричный», «пассивно-агрессивный», «импульсивный», «нарциссический», «нарциссический», «параноидальный», «шизоидный» и «шизотипический» соответственно. В поиск включены публикации до 15 мая 2018 г.
Были использованы следующие ключевые слова: «личность», «расстройство личности», «черта личности», каждое из которых сочеталось с «эпигенетический», «метилирование», «ацетилирование», «фосфорилирование», «убиквитация», «сумоилирование», « микроРНК», «хроматин» и «ремоделирование хроматина» соответственно, а также с одним из ключевых слов «агрессивность», «ананкастность», «асоциальный», «тревожный», «избегающий», «пограничный», «зависимый», «эксцентричный», «эмоционально неустойчивый», «истеричный», «пассивно-агрессивный», «импульсивный», «нарциссический», «нарциссический», «параноидальный», «шизоидный» и «шизотипический» соответственно. В поиск включены публикации до 15 мая 2018 г.
Всего поиск дал 345 различных статей. Во-вторых, мы просмотрели полученные статьи, просмотрев их заголовки, рефераты и полные тексты, чтобы определить, какие статьи соответствуют нашему буквальному исследовательскому вопросу. Таким образом, исследования отбирались последовательно, если они соответствовали критериям (1) психиатрическая тема [ n = 257], (2) расстройство личности или определенная черта личности [ n = 72 из (1)], (3) исследование на людях испытуемых [ n = 61 из (2)], (4) эпигенетических анализов [ n = 43 из (3)] и (5) оригинальное исследование [ n = 23 из (4)]. Следуя этим критериям отбора, осталось 23 статьи в соответствии с намеченной целью этого обзора (рис. 1).
Следуя этим критериям отбора, осталось 23 статьи в соответствии с намеченной целью этого обзора (рис. 1).
Рисунок 1 . Блок-схема поиска исследований и последовательного отбора.
Одновременно мы использовали функцию поиска PubMed f1000prime (Faculty of 1000 Limited, Лондон, Великобритания). При этом мы нашли еще одно подходящее исследование, отвечающее всем описанным критериям включения (18). Наконец, в этот обзор были включены 24 оригинальных исследования.
Результаты
Среди включенных исследований 13 изучали эпигенетическое влияние на ПТ (два на импульсивность, шесть на антисоциальные черты, семь на агрессию) и 11 изучали эпигенетические различия в расстройствах личности (два в антисоциальном расстройстве личности (АСРЛ), 9 при пограничном расстройстве личности (ПРЛ)).
Размер, состав изучаемых групп и количество исследованных генов различались между исследованиями и варьировались от анализа отдельных генов до анализа метилирования всего генома (GWA) с последствиями для статистической мощности.
Расстройства личности
Антисоциальное расстройство личности
Что касается антисоциальной болезни Паркинсона, исследовано только метилирование гена моноаминоксидазы А ( MAOA ). Два исследования решительно различались по дизайну исследования и результатам метилирования.
Филибер и др. рассмотрели известную область вариабельного нуклеотидного повтора (VTNR) MAOA и ввели новую область VTNR выше сайта начала транскрипции (TSS) гена ( MAOA VTNR P2). Они обнаружили генотип-зависимый уровень метилирования и активность генов, но только у самок (19). В общей сложности у 571 хорошо охарактеризованного субъекта (312 женщин) в исследовании усыновления в Айове (IAS) они измерили количество симптомов ASPD в течение жизни в линейном режиме в соответствии с критериями DSM-IV. Паттерны метилирования анализировали на двух ассоциированных с промотором CpG-островках MAOA в ДНК, экстрагированной из трансформированных EBV клеточных линий лимфобластов из периферической крови.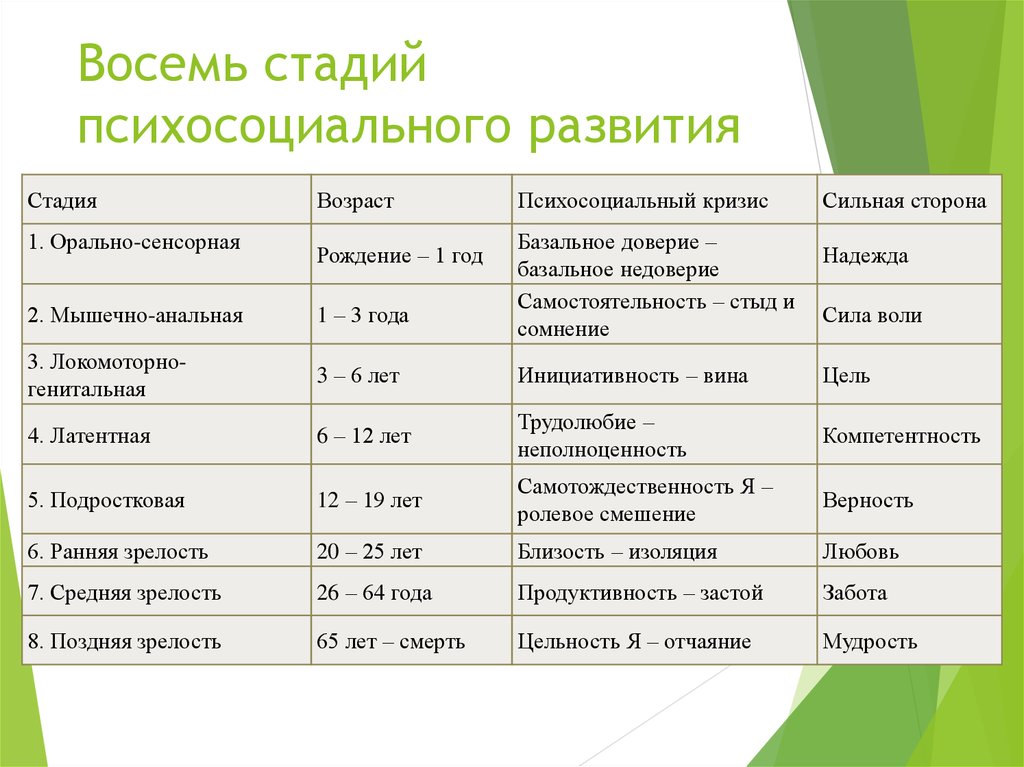 Анализ последовательности VTNR P2 выявил пять генотипов, каждый из которых содержит от семи до одиннадцати одиннадцати повторов (7R, 8R, 9R, 10R, 11R), из которых генотип 9R показал наименьшее метилирование у гомозиготных самок и наибольшую активность гена при функциональном анализе с помощью люциферазы. Наличие аллеля 10R с низкой активностью было связано с более высокой уязвимостью к вредным последствиям сексуального и физического насилия в детстве и в значительной степени объясняло различия в тяжести симптомов ASPD у женщин. У мужчин не было обнаружено значительного влияния генотипа P2 на статус метилирования MAOA .
Анализ последовательности VTNR P2 выявил пять генотипов, каждый из которых содержит от семи до одиннадцати одиннадцати повторов (7R, 8R, 9R, 10R, 11R), из которых генотип 9R показал наименьшее метилирование у гомозиготных самок и наибольшую активность гена при функциональном анализе с помощью люциферазы. Наличие аллеля 10R с низкой активностью было связано с более высокой уязвимостью к вредным последствиям сексуального и физического насилия в детстве и в значительной степени объясняло различия в тяжести симптомов ASPD у женщин. У мужчин не было обнаружено значительного влияния генотипа P2 на статус метилирования MAOA .
Напротив, в популяции мужчин-заключенных ( n = 86), отвечающих критериям DSM-IV для ASPD, Checknita et al. обнаружили значительное общее гиперметилирование области промотора MAOA в группе ASPD по сравнению со здоровым контролем ( n = 93) со значительными различиями в уровнях метилирования на 34 из 71 различных сайтов CpG промотора MAOA . В своем анализе они не учитывали тяжесть симптомов или невзгоды детства. Метилирование 9Промотор 0009 MAOA ассоциировался со сниженной активностью гена (анализ люциферазы) и положительно коррелировал с уровнями 5-гидрокситриптофана в крови, что предполагает функциональную значимость (20).
В своем анализе они не учитывали тяжесть симптомов или невзгоды детства. Метилирование 9Промотор 0009 MAOA ассоциировался со сниженной активностью гена (анализ люциферазы) и положительно коррелировал с уровнями 5-гидрокситриптофана в крови, что предполагает функциональную значимость (20).
Пограничное расстройство личности
Что касается эпигенетических модификаций при ПРЛ, предполагается, что различные гены участвуют в индивидуальном развитии, а также в фенотипических характеристиках расстройства (таблица 1). Помимо полногеномного анализа (GWA), основное внимание в теоретических эпигенетических исследованиях уделялось участкам генов, кодирующим BDNF, глюкокортикоидный рецептор (9).0009 NR3C1 ), дофаминовые и серотониновые рецепторы, МАОА и катехол-О-метилтрансфераза ( COMT ). Большинство исследований, посвященных генам-мишеням, рассматривали раннюю детскую травму в анамнезе как фактор, вмешивающийся в их анализ. Разнообразие в определении контрольной группы в отдельных исследованиях отражает борьбу за подходящий дизайн исследования, который позволяет изолировать специфические для расстройства характеристики. Аберрации метилирования в основном оценивались с диагностической, но также и с прогностической точки зрения (21).
Аберрации метилирования в основном оценивались с диагностической, но также и с прогностической точки зрения (21).
Таблица 1 . Исследования эпигенетики при расстройствах личности.
Perroud и др. изучали статус метилирования гена BDNF в лейкоцитах периферической крови и его модуляцию с помощью целенаправленного терапевтического вмешательства с интенсивной диалектической поведенческой терапией (I-DBT) продолжительностью 4 недели у амбулаторных пациентов с БЛД ( n = 115). В отличие от здоровых субъектов, богатые CpG области в экзоне 1 и экзоне 4 были значительно более высоко метилированы у субъектов с ПРЛ до терапевтического вмешательства. Количество различных видов детской травмы (ДТ) по Опроснику детской травмы (CTQ) положительно коррелировало со средним процентом метилирования обеих CpG-областей. После интенсивной ДПТ статус метилирования рассматриваемых CpG-сайтов значительно повышался у пациентов с БЛД. Этот эффект можно было проследить до тех, кто не ответил, тогда как у ответивших наблюдалось снижение метилирования. Однако сравнение 9Статус метилирования 0009 BDNF с уровнями белка BDNF в периферической сыворотке не выявил значимой связи (21).
Однако сравнение 9Статус метилирования 0009 BDNF с уровнями белка BDNF в периферической сыворотке не выявил значимой связи (21).
Что касается исследований метилирования NR3C1 и PD, два исследования были сосредоточены на промоторе экзона 1F, который является важным с функциональной точки зрения. В когорте амбулаторных пациентов с БЛД ( n = 281) Martin-Blanco и соавт. (22) обнаружили значительную положительную корреляцию между общим уровнем метилирования NR3C1 экзона 1F лейкоцитов периферической крови и клинической тяжестью. Метилирование экзона 1F также было в значительной степени связано с физическим насилием в детстве. Отдельные сайты CpG были связаны с определенными подшкалами CTQ.
Perroud et al. (23) считали, что текущее тяжелое психическое заболевание может иметь эпигенетические последствия per se и может запутать анализы, направленные на выделение характеристик метилирования, специфичных для ПРЛ. Таким образом, при сравнении субъектов с БЛД ( n = 101) и пациентов с БДР ( n = 99) их результаты показали более высокие общие уровни метилирования экзона 1F NR3C1 при БЛД, чем у субъектов с БДР в лейкоцитах периферической крови. Кроме того, метилирование было связано с CT, масштабируемым с помощью CTQ, и коррелировало с сексуальным, физическим и эмоциональным насилием в детстве и физическим и эмоциональным пренебрежением соответственно, а также с количеством этих типов CT. 9Гиперметилирование экзона 1F 0009 NR3C1 у субъектов с пограничным расстройством личности было все еще значительным после поправки на жестокое обращение в детстве.
Кроме того, метилирование было связано с CT, масштабируемым с помощью CTQ, и коррелировало с сексуальным, физическим и эмоциональным насилием в детстве и физическим и эмоциональным пренебрежением соответственно, а также с количеством этих типов CT. 9Гиперметилирование экзона 1F 0009 NR3C1 у субъектов с пограничным расстройством личности было все еще значительным после поправки на жестокое обращение в детстве.
Что касается генов моноаминовых рецепторов, в крупномасштабном исследовании с участием пациентов с расстройствами спектра булимии (BSD) с коморбидным ПРЛ и без него Groleau et al. (24) обнаружили значительное, но незначительно повышенное метилирование промоторной области экзона 1 гена рецептора дофамина D2 ( DRD2 ) в ДНК цельных клеток периферической крови (PBC) у субъектов с BSD и BPD по сравнению с таковой у субъектов только с BSD.
Метилирование гена рецептора серотонина 3A ( 5HTR3A ) было обнаружено Perroud et al. (25) коррелирует с клинической тяжестью ПРЛ и других психических расстройств. Авторы сравнили уровни метилирования ДНК ПБХ восьми сайтов CpG в гене 5HT3A у пациентов с ПРЛ ( n = 116), синдромом дефицита внимания и гиперактивности (СДВГ) ( n = 111) и биполярным расстройством (BD) ( n = 122). Они также рассматривали однонуклеотидные варианты (SNP) гена и историю CT в качестве дополнительных факторов и исследовали ассоциации с уровнями метилирования конкретных сайтов CpG.
Авторы сравнили уровни метилирования ДНК ПБХ восьми сайтов CpG в гене 5HT3A у пациентов с ПРЛ ( n = 116), синдромом дефицита внимания и гиперактивности (СДВГ) ( n = 111) и биполярным расстройством (BD) ( n = 122). Они также рассматривали однонуклеотидные варианты (SNP) гена и историю CT в качестве дополнительных факторов и исследовали ассоциации с уровнями метилирования конкретных сайтов CpG.
Уровни метилирования между группами пациентов значительно различались в большинстве CpG-сайтов и демонстрировали отчетливую картину гипер- и гипометилирования при некоторых нарушениях в выбранных CpG-сайтах. Для ПРЛ субъекты показали самые высокие баллы в CTQ и самый высокий уровень метилирования между группами пациентов. CT был связан со средним статусом метилирования, а общий балл CTQ и физическое насилие были связаны с разными выбранными сайтами CpG. Кроме того, КТ была связана с более высокой тяжестью заболевания. Носительство СС-аллеля достоверно ассоциировалось с метилированием одного специфического сайта CpG независимо от CT (при всех заболеваниях).
Дамманн и др. (26) проанализировали пять психоневрологических генов, предположительно имеющих значение для психопатологического фенотипа, в частности гены, кодирующие растворимую катехол-О-метилтрансферазу ( s-COMT ), рецептор серотонина 2A ( HTR2A ), NR3C1 и X. -хромосомы MAOA и MAOB . Уровни метилирования каждого промотора гена были количественно определены в ДНК PBC индивидуумов с ПРЛ ( n = 26, 24 женщины) и в здоровой контрольной группе ( n = 11, все женщины). По сравнению со здоровым контролем количественный анализ метилирования ДНК показал значительное повышение общего уровня метилирования у субъектов с ПРЛ в пределах HTR2A, NR3C1 и s-COMT . Метилирование генов MAOA и MAOB можно было проанализировать только у женщин, а метилирование MAOA было значительно выше при БЛД ( MAOB только по тенденции). Принимая во внимание все 27 отдельных CpG-сайтов по пяти исследованным генам, средний уровень метилирования во всех количественно определяемых областях был значительно выше у пациентов с ПРЛ по сравнению с контрольной группой. Влияние сопутствующих данных на историю травм субъектов не было представлено в рамках исследования, но было отмечено, что аберрантное метилирование не было связано с травматическим опытом в соответствующих статистических тестах.
Влияние сопутствующих данных на историю травм субъектов не было представлено в рамках исследования, но было отмечено, что аберрантное метилирование не было связано с травматическим опытом в соответствующих статистических тестах.
В то же время эпигенетические анализы включают полногеномные ассоциативные исследования (GWA), которые указывают на конкретные сайты CpG с аберрантными уровнями метилирования во всей геномной ДНК и облегчают комплексные аспекты эпигенетических оценок.
В дополнение к вышеупомянутому исследованию Teschler et al. (27) выполнили GWA в ДНК PBC между женщинами с BPD ( n = 24) и HC ( n = 11). Результаты показали в общей сложности 256 значительно гиперметилированных сайтов CpG при ПРЛ, но значимость каждого из них не сохранялась после коррекции Бонферрони. Исследовательская группа выбрала семь гиперметилированных генов для проверочного анализа и могла подтвердить повышенное метилирование в пяти из генов с определенными сайтами CpG, связанными с членом семейства А, связывающим белок-предшественник бета-амилоида (A4) 2 (9). 0009 APBA2 ) и члена 3 ( APBA3 ), ген калиевого потенциалзависимого канала KQT-подобного подсемейства члена 1 ( KCNQ1 ), ген трансформирующей последовательности, полученный из клеточной линии MCF2 ( MCF2 ), и ген ниндзюрина 2 ( НИНДЖ2 ). Гены GATA-связывающего белка 4 ( GATA4 ) и холокарбоксилазы синтетазы ( HLCS ) показали повышенное метилирование при БЛД в GWA, но не в проверочном анализе.
0009 APBA2 ) и члена 3 ( APBA3 ), ген калиевого потенциалзависимого канала KQT-подобного подсемейства члена 1 ( KCNQ1 ), ген трансформирующей последовательности, полученный из клеточной линии MCF2 ( MCF2 ), и ген ниндзюрина 2 ( НИНДЖ2 ). Гены GATA-связывающего белка 4 ( GATA4 ) и холокарбоксилазы синтетазы ( HLCS ) показали повышенное метилирование при БЛД в GWA, но не в проверочном анализе.
Исследования метилирования гена рибосомной РНК ( промотор рДНК ), 5′-транскрибируемый внешний спейсерный ген ( 5 ‘ ETS ) и пролин богатого пролином мембранного якоря 1 промотора ( PRIMA1 ) в ДНК PBC женщин с ПРЛ ( n = 24 ) и HC ( n = 11) выявили значительно на меньше метилирования промоторной области рДНК при ПРЛ по сравнению с субъектами HC и гипометилирование 5 ′ ETS при ПРЛ по тренду. PRIMA1 показал более высокое метилирование у субъектов с ПРЛ (28).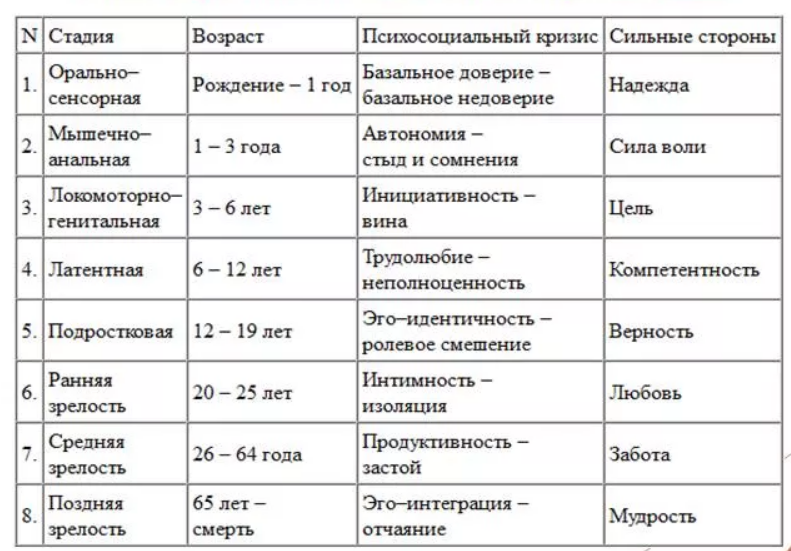
Другой GWA с ДНК PBC был выполнен Prados et al. (29) у субъектов с пограничным расстройством личности, страдающих высоким уровнем неблагополучия в детстве ( n = 96), и у субъектов с БДР и низким уровнем неблагополучия в детстве в анамнезе ( n = 93). Однофакторный и многофакторный анализы выявили значительные различия в метилировании значительного числа конкретных CpG-сайтов, связанных с ПРЛ, по сравнению с субъектами с БДР или связанными с жестоким обращением в детстве, соответственно. В отличие от пациентов с БЛД и пациентов с БДР, наиболее значимые результаты многомерного анализа показали, что метилированные CpG-сайты значительно различаются, например, в пределах гена, кодирующего лиганд тирозинкиназ рецептора, родственного Eph (9).0009 EFBN1 ), близко к гену, кодирующему члена семейства супрессора передачи сигналов цитокинов (SOCS) ( SPSB2 ), и рядом с геном, кодирующим белок, подобный мышиному цистатину 9 ( CST9L ). Однофакторный анализ выявил гипометилированные сайты CpG при БЛД, т. е. рядом с кодирующей областью микроРНК 124 ( miR124-3 ), которая нацелена на несколько генов, которые, как было описано, коррелируют с БЛД (включая NR3C1 ), рядом с геном, кодирующим повторный домен WD 60 ( WD60 ), и сайт CpG в гене семейства со сходством последовательностей 163 члена A ( FAM163A ). Многие из значительно гиперметилированных участков были обнаружены на хромосоме X. При нацеливании на КТ результаты многомерного анализа с помощью CTQ выявили сильные ассоциации с локусами внутри или рядом с генами человеческого гомолога мыши p (разведение для розовых глаз) ( OCA2 ), микрофибриллярно-ассоциированный белок 2 ( MFAP2 ), CST9L , E1A-связывающий белок p400 ( EP400 ), KCNQ2 , альфа-2-макроглобулин-подобный 1 ( A2ML1 ), 5′-нуклеотидный домен, содержащий 2 ( NT5DC2 ) и рабфилин 3A-подобный ( RPh4AL ). Однофакторный анализ выявил наиболее значительные изменения, локализованные в гене, кодирующем рецептор интерлейкина 17 A ( IL17RA ), в межгенной области на хромосоме 6p22.
е. рядом с кодирующей областью микроРНК 124 ( miR124-3 ), которая нацелена на несколько генов, которые, как было описано, коррелируют с БЛД (включая NR3C1 ), рядом с геном, кодирующим повторный домен WD 60 ( WD60 ), и сайт CpG в гене семейства со сходством последовательностей 163 члена A ( FAM163A ). Многие из значительно гиперметилированных участков были обнаружены на хромосоме X. При нацеливании на КТ результаты многомерного анализа с помощью CTQ выявили сильные ассоциации с локусами внутри или рядом с генами человеческого гомолога мыши p (разведение для розовых глаз) ( OCA2 ), микрофибриллярно-ассоциированный белок 2 ( MFAP2 ), CST9L , E1A-связывающий белок p400 ( EP400 ), KCNQ2 , альфа-2-макроглобулин-подобный 1 ( A2ML1 ), 5′-нуклеотидный домен, содержащий 2 ( NT5DC2 ) и рабфилин 3A-подобный ( RPh4AL ). Однофакторный анализ выявил наиболее значительные изменения, локализованные в гене, кодирующем рецептор интерлейкина 17 A ( IL17RA ), в межгенной области на хромосоме 6p22. 1 и закрытой miR124-3 и miR137 . Метилирование miR124-3 было связано как с тяжестью невзгод в детстве (более высокое метилирование), так и с ПРЛ (более низкое метилирование).
1 и закрытой miR124-3 и miR137 . Метилирование miR124-3 было связано как с тяжестью невзгод в детстве (более высокое метилирование), так и с ПРЛ (более низкое метилирование).
Таким образом, большая часть исследований БП была сосредоточена на отдельных генах, включая MAOA при ASPD и BDNF, NR3C1, DRD2 и HTR3A при ПРЛ, или на наборе предполагаемых генов, основанных на теории. (26). Они выявили значительные различия в метилировании ДНК клеток крови у субъектов с соответствующей болезнью Паркинсона. Во всех независимых исследованиях, нацеленных на гены, гиперметилирование NR3C1 наиболее часто и неизменно связывалось с ПРЛ; поэтому текущие результаты наиболее сильно указывают на NR3C1 , связанный с ПРЛ. NR3C1 , кроме того, был единственным геном, который также был затронут в одном из GWA (29), однако это было опосредовано различиями в метилировании miR124-3 , которые нацелены, среди других генов, на NR3C1 . Напротив, исследования GWA, проведенные Teschler и Prados, обнаружили большое количество генов, которые были по-разному метилированы при ПРЛ, что указывает на общесистемное участие в БП, включая гены, связанные с иммунным ответом, передачей клеточных сигналов или контролем транскрипции (27–27). 29). Транскрипционная значимость соответствующих различий метилирования была подтверждена только двумя авторами. Так, Checknita et al. обнаружили косвенную отрицательную связь гиперметилирования промотора MAOA с уровнем 5-HT в сыворотке in vitro (20). В противном случае Perroud et al. (21) не обнаружили связи между метилированием промотора BDNF и уровнем BDNF в сыворотке периферических белков. Все исследования были основаны на ДНК, выделенной из ПБЦ, в большинстве случаев из клеток цельной крови, частично только из лейкоцитов (21–23) или селективно из лимфоцитов (19).). Тем не менее, исследования особенно различаются по дизайну испытуемых и контрольной группы и, по крайней мере, по учету факторов окружающей среды, влияющих на эпигенетическую модуляцию, таких как невзгоды в раннем детстве, что затрудняет четкую интерпретацию в большинстве исследований (таблица 1).
Напротив, исследования GWA, проведенные Teschler и Prados, обнаружили большое количество генов, которые были по-разному метилированы при ПРЛ, что указывает на общесистемное участие в БП, включая гены, связанные с иммунным ответом, передачей клеточных сигналов или контролем транскрипции (27–27). 29). Транскрипционная значимость соответствующих различий метилирования была подтверждена только двумя авторами. Так, Checknita et al. обнаружили косвенную отрицательную связь гиперметилирования промотора MAOA с уровнем 5-HT в сыворотке in vitro (20). В противном случае Perroud et al. (21) не обнаружили связи между метилированием промотора BDNF и уровнем BDNF в сыворотке периферических белков. Все исследования были основаны на ДНК, выделенной из ПБЦ, в большинстве случаев из клеток цельной крови, частично только из лейкоцитов (21–23) или селективно из лимфоцитов (19).). Тем не менее, исследования особенно различаются по дизайну испытуемых и контрольной группы и, по крайней мере, по учету факторов окружающей среды, влияющих на эпигенетическую модуляцию, таких как невзгоды в раннем детстве, что затрудняет четкую интерпретацию в большинстве исследований (таблица 1). , столбец «Ограничения»).
, столбец «Ограничения»).
Черты личности
В большинстве оригинальных публикаций о метилировании и чертах личности были обнаружены антисоциальные или агрессивные черты в связи с эпигенетическими изменениями, связанными с функциональностью серотонинергической системы или функциональностью гипоталамо-гипофизарно-надпочечниковой (ГГН-) оси, т.е. основная нейроэндокринная система регуляции реакции на стресс и адаптации. Соответственно, в теоретических исследованиях изучались гены, связанные с этими системами, такие как ген переносчика серотонина (9).0009 SLC6A4 ), гены рецепторов допамина и серотонина ( DRD1, HTR1B, HTR1D, HTR3A ), MAOA и NR3C1 промоторный экзон 1F. Что касается его роли в социально-аффективном восприятии и обработке, существуют некоторые исследования, касающиеся антисоциального поведения и генов окситоцина и рецепторов окситоцина ( OXT, OXTR ). Далее рассматривалась роль цитокинов и других факторов.
Антисоциальные черты
В соответствии с большим объемом литературы, в которой серотонинергическая система участвует в регуляции тревоги, агрессии и реакции на стресс, эпигенетические изменения в гене переносчика серотонина ( SLC6A4 ) были связаны с историей сексуального насилия над детьми, а также с антиобщественным поведением во взрослом возрасте Beach et al. (30). Авторы провели анализ метилирования ДНК лимфоцитов периферической крови участников, которые были трансформированы ВЭБ в клеточные линии лимфобластов. Участники ( n = 155, все женщины) постепенно диагностировались с помощью оценки симптомов ASPD. Сексуальное насилие в детстве было тесно связано со средним уровнем метилирования, а метилирование было в значительной степени связано с симптомами ASPD, тем самым играя модулирующую роль в развитии антисоциальных черт после сексуального насилия в детстве. Интересно, что влияние 9Метилирование 0009 SLC6A4 на тяжесть ASPD зависело от генотипа, поскольку связь метилирования с генотипом ss и sl SLC6A4 была значимой, но не с генотипом ll. Эти результаты предполагают отягчающий эффект метилирования в SLC6A4 аллелях риска (s) для ASPD, кроме того, метилирование в большей степени ассоциировалось с ASPD у носителей с большим количеством аллелей s.
(30). Авторы провели анализ метилирования ДНК лимфоцитов периферической крови участников, которые были трансформированы ВЭБ в клеточные линии лимфобластов. Участники ( n = 155, все женщины) постепенно диагностировались с помощью оценки симптомов ASPD. Сексуальное насилие в детстве было тесно связано со средним уровнем метилирования, а метилирование было в значительной степени связано с симптомами ASPD, тем самым играя модулирующую роль в развитии антисоциальных черт после сексуального насилия в детстве. Интересно, что влияние 9Метилирование 0009 SLC6A4 на тяжесть ASPD зависело от генотипа, поскольку связь метилирования с генотипом ss и sl SLC6A4 была значимой, но не с генотипом ll. Эти результаты предполагают отягчающий эффект метилирования в SLC6A4 аллелях риска (s) для ASPD, кроме того, метилирование в большей степени ассоциировалось с ASPD у носителей с большим количеством аллелей s.
Моул и др. обнаружили антисоциальные черты, связанные с генетической и эпигенетической модуляцией SNP rs11568817 в Промоторная область HTR1B (31). В исследовании с мальчиками ( n = 117), проявляющими бессердечно-неэмоциональные (CU) черты и проблемы с антисоциальным поведением, и сгруппированными по сильным и менее сильным CU чертам, они обнаружили более низкие уровни метилирования HTTR1B в ДНК клеток слюны в группа признаков с высоким CU. В свою очередь, метилирование решительно сдерживалось SNP генотипом rs11568817 и осуществлялось двумя эксклюзивными сайтами CpG (CpG12 и CpG14), индивидуальные уровни метилирования которых были отрицательно связаны с общими уровнями метилирования в этой области гена. Авторы предполагают два пути риска для признаков с высоким CU: во-первых, носительство рискового (минорного) аллеля (аллелей) с низким уровнем метилирования в CpG-сайтах 12 и 14 (что было связано с высоким общим метилированием промотора) и, во-вторых, носительство без аллеля риска, но с высоким метилированием в CpG 12 и CpG14 (что было связано с низким общим метилированием) и высокой экспрессией HTR1B (31).
В исследовании с мальчиками ( n = 117), проявляющими бессердечно-неэмоциональные (CU) черты и проблемы с антисоциальным поведением, и сгруппированными по сильным и менее сильным CU чертам, они обнаружили более низкие уровни метилирования HTTR1B в ДНК клеток слюны в группа признаков с высоким CU. В свою очередь, метилирование решительно сдерживалось SNP генотипом rs11568817 и осуществлялось двумя эксклюзивными сайтами CpG (CpG12 и CpG14), индивидуальные уровни метилирования которых были отрицательно связаны с общими уровнями метилирования в этой области гена. Авторы предполагают два пути риска для признаков с высоким CU: во-первых, носительство рискового (минорного) аллеля (аллелей) с низким уровнем метилирования в CpG-сайтах 12 и 14 (что было связано с высоким общим метилированием промотора) и, во-вторых, носительство без аллеля риска, но с высоким метилированием в CpG 12 и CpG14 (что было связано с низким общим метилированием) и высокой экспрессией HTR1B (31).
В той же выборке участников, стратифицированных по уровню баллов CU и возрасту, вышеупомянутая исследовательская группа проанализировала характеристики метилирования гена рецептора оцитоцина ( OXTR ) в ДНК PBC. Они выявили высокие признаки CU, связанные с повышенным метилированием в двух из шести проанализированных локусов CpG в промоторе OXTR и коррелирующие со снижением уровня окситоцина в крови. Тем не менее, при разделении по возрастным группам эти данные достигли значимости только в старшей группе (возраст 9 лет).–16 лет), а не у детей препубертатного возраста (возраст 4–8 лет) (здесь только тенденция). Точно так же уровни окситоцина в сыворотке значимо отрицательно коррелировали с уровнем метилирования генов у мальчиков старшего возраста (32).
Далее, в проспективном лонгитюдном исследовании Cecil et al. (18) изучили 84 молодых человека с ранними и стойкими проблемами поведения в отношении рисков в раннем возрасте и изменений метилирования OXTR в ДНК ПБХ. Клинические опросы проводились при рождении, в возрасте 7, 9 и 13 лет, и собирали оценку риска окружающей среды (вид и время фактора риска), диагностику проблем поведения, черт CU и проблем интернализации. В равной степени анализы метилирования проводились продольно с отбором проб при рождении (пуповинная кровь) и в возрасте 7 и 9 лет.лет (периферическая кровь). Наконец, для оценки испытуемые были разделены по серьезности проблем интернализации, собранных по отчетам матерей в каждый период исследования.
Клинические опросы проводились при рождении, в возрасте 7, 9 и 13 лет, и собирали оценку риска окружающей среды (вид и время фактора риска), диагностику проблем поведения, черт CU и проблем интернализации. В равной степени анализы метилирования проводились продольно с отбором проб при рождении (пуповинная кровь) и в возрасте 7 и 9 лет.лет (периферическая кровь). Наконец, для оценки испытуемые были разделены по серьезности проблем интернализации, собранных по отчетам матерей в каждый период исследования.
Результаты для молодежи с низкими проблемами интернализации показали, что гиперметилирование OXTR при рождении было значимо связано с более высокими чертами CU в возрасте 13 лет. виктимизация. Единственный экологический фактор риска, связанный с Метилирование OXTR при рождении было пренатальным родительским риском, т. е. материнской психопатологией, преступным поведением и употреблением психоактивных веществ. У молодежи с высокими проблемами интернализации признаки CU не показали значительной связи с метилированием гена OXTR , но положительно коррелировали с пренатальными, особенно межличностными рисками, такими как насилие со стороны интимного партнера или семейные конфликты. Интересно, что черты CU отрицательно коррелировали с постнатальными рисками, в частности с жизненными событиями, такими как смерть родственника, несчастный случай или болезнь. Эти разные результаты корреляции наводят на мысль о различающих путях развития признаков CU, один из которых эпигенетически детерминирован, а другой зависит от различных факторов окружающей среды. Если связано с OXTR гиперметилирование при рождении, признаки CU могут защищать от проблем интернализации и от переживаемой виктимизации, что авторы рассматривают как возможную вызывающую воспоминания корреляцию (18).
Интересно, что черты CU отрицательно коррелировали с постнатальными рисками, в частности с жизненными событиями, такими как смерть родственника, несчастный случай или болезнь. Эти разные результаты корреляции наводят на мысль о различающих путях развития признаков CU, один из которых эпигенетически детерминирован, а другой зависит от различных факторов окружающей среды. Если связано с OXTR гиперметилирование при рождении, признаки CU могут защищать от проблем интернализации и от переживаемой виктимизации, что авторы рассматривают как возможную вызывающую воспоминания корреляцию (18).
Принимая во внимание симптомы ASPD, Philibert et al. (33) исследовали генотип полиморфизма MAOA VTNR и статус метилирования в трансформированных EBV клеточных линиях лимфобластов в отношении тяжести симптомов ASPD и расстройства, связанного с употреблением психоактивных веществ, определяемого как алкогольная или никотиновая зависимость, в коллективе из 19 человек.1 испытуемый (96 женщин). Уровень метилирования был неизменно выше у женщин, чем у мужчин, в каждом анализируемом CpG-сайте. Наблюдалась только статистическая тенденция для женщин, гомозиготных по аллелю 3,3, с более высоким средним метилированием, чем по аллелю 4,4. Исследование не выявило связи между количеством симптомов ASPD и уровнями метилирования ни у мужчин, ни у женщин (33).
Уровень метилирования был неизменно выше у женщин, чем у мужчин, в каждом анализируемом CpG-сайте. Наблюдалась только статистическая тенденция для женщин, гомозиготных по аллелю 3,3, с более высоким средним метилированием, чем по аллелю 4,4. Исследование не выявило связи между количеством симптомов ASPD и уровнями метилирования ни у мужчин, ни у женщин (33).
Обращая внимание на многогранные особенности у субъектов с ранними проблемами поведения, такие как повышенная раздражительность, тревога, импульсивность, возможно связанные с нарушением функции гипоталамо-гипофизарно-надпочечниковой системы (ГГН), Dadds et al. (34) исследовали паттерны метилирования Промотор экзона 1F NR3C1 в ДНК клеток цельной крови и слюны и утренние уровни кортизола в плазме в исследовательском коллективе ( n = 241, 51 женщина) с полными критериями или признаками расстройства поведения (CD) или оппозиционно-вызывающего расстройства (ODD) ) в отличие от австралийской нормативной выборки в качестве контроля (35).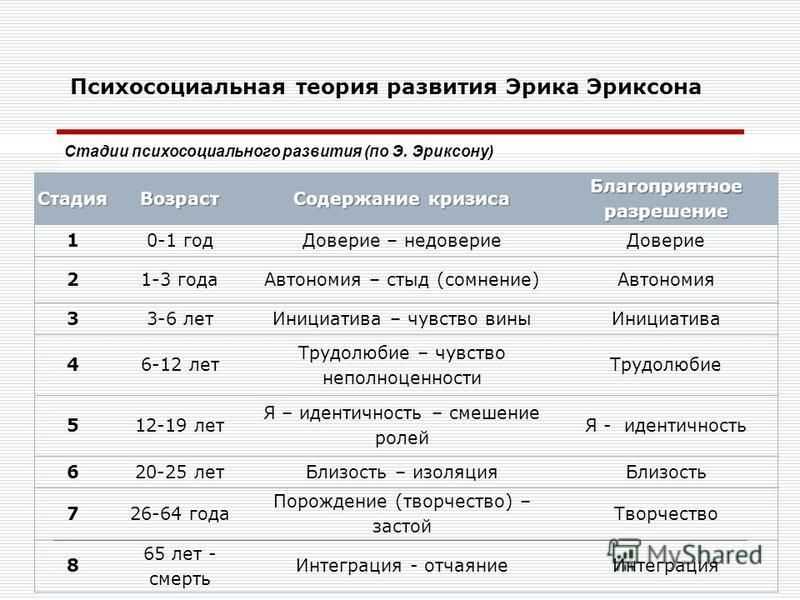 Психопатологические симптомы были отнесены к более интернализирующим расстройствам, в отличие от признаков экстернализирующих расстройств, таких как черты CU или проблемы с поведением. Общий уровень метилирования 9Экзон 1 0009 NR3C1 в пробах ДНК крови и слюны был связан с более высоким уровнем интернализирующих расстройств. Отчетливо CpG10 показал связь повышенного метилирования с более высокими показателями симптомов интернализации. Напротив, уровни метилирования CpG3 и CpG4 в пробах ДНК слюны предсказывали тяжесть экстернализации (34). Уровни периферического утреннего кортизола в плазме были связаны с гиперметилированием CpG9 только в ДНК PBC.
Психопатологические симптомы были отнесены к более интернализирующим расстройствам, в отличие от признаков экстернализирующих расстройств, таких как черты CU или проблемы с поведением. Общий уровень метилирования 9Экзон 1 0009 NR3C1 в пробах ДНК крови и слюны был связан с более высоким уровнем интернализирующих расстройств. Отчетливо CpG10 показал связь повышенного метилирования с более высокими показателями симптомов интернализации. Напротив, уровни метилирования CpG3 и CpG4 в пробах ДНК слюны предсказывали тяжесть экстернализации (34). Уровни периферического утреннего кортизола в плазме были связаны с гиперметилированием CpG9 только в ДНК PBC.
Кроме того, Radtke et al. (36) исследовали взаимное влияние истории жестокого обращения в детстве, метилирования 9Промотор 0009 NR3C1 в лимфоцитах периферической крови и психопатологическая нагрузка у субъектов условной выборки ( n = 46, 28 женщин). Их результаты не указывают на значительную корреляцию между средним уровнем метилирования промоторной области NR3C1 с тяжестью жестокого обращения в детстве или с баллами собранных психометрических данных. Тем не менее, с помощью отдельных анализов они изобразили два из 41 выбранного сайта CpG, которые взаимодействовали с жестоким обращением в детстве и уязвимостью к психопатологии. Один из двух упомянутых CpG-сайтов располагался в промоторе экзона 1F, и его метилирование высоко достоверно коррелировало с количеством пережитых детских невзгод по данным CTQ, а также с симптомами ПРЛ. С помощью линейной модели авторы выявили независимо сильное влияние жестокого обращения в детстве и статуса метилирования этого CpG-сайта на развитие симптомов, связанных с ПРЛ, и обнаружили аддитивный эффект обоих этих факторов (36).
Тем не менее, с помощью отдельных анализов они изобразили два из 41 выбранного сайта CpG, которые взаимодействовали с жестоким обращением в детстве и уязвимостью к психопатологии. Один из двух упомянутых CpG-сайтов располагался в промоторе экзона 1F, и его метилирование высоко достоверно коррелировало с количеством пережитых детских невзгод по данным CTQ, а также с симптомами ПРЛ. С помощью линейной модели авторы выявили независимо сильное влияние жестокого обращения в детстве и статуса метилирования этого CpG-сайта на развитие симптомов, связанных с ПРЛ, и обнаружили аддитивный эффект обоих этих факторов (36).
Таким образом, все исследования эпигенетических ассоциаций с антисоциальными чертами были сосредоточены на отдельных генах. Каждое исследование выявило значительные модификации метилирования в промоторе SLC6A4 (30) и в HTR1B (31), оба во взаимодействии с вариантами генотипа рассматриваемого гена. Два исследования были посвящены OXTR с разными результатами. В первом исследовании высокие признаки CU были связаны с гиперметилированием OXTR , что коррелировало с более низким уровнем OXT в сыворотке (32). В другом лонгитюдном исследовании 9Гиперметилирование 0009 OXTR при рождении было связано с более высокими чертами CU в препубертатном возрасте у молодежи с низкими проблемами интернализации, но метилирование OXTR не было связано с чертами CU у молодых людей с высокими проблемами интернализации, в которых они положительно коррелировали с пренатальными межличностными рисками. и отрицательно с негативными жизненными событиями (18). Это предполагает другой (эпи-)генетический патогенез признаков ХК. Напротив, среднее метилирование MAOA не было связано с оценкой симптомов ASPD (33) и не было значимо связано с MAOA Генотипы VTNR. Однако в двух независимых исследованиях с NR3C1 получены разные результаты. Следовательно, гиперметилирование отдельных CpG-сайтов в экзоне 1F промотора NR3C1 , но не общее метилирование, было связано с симптомами экстернализации (34).
В первом исследовании высокие признаки CU были связаны с гиперметилированием OXTR , что коррелировало с более низким уровнем OXT в сыворотке (32). В другом лонгитюдном исследовании 9Гиперметилирование 0009 OXTR при рождении было связано с более высокими чертами CU в препубертатном возрасте у молодежи с низкими проблемами интернализации, но метилирование OXTR не было связано с чертами CU у молодых людей с высокими проблемами интернализации, в которых они положительно коррелировали с пренатальными межличностными рисками. и отрицательно с негативными жизненными событиями (18). Это предполагает другой (эпи-)генетический патогенез признаков ХК. Напротив, среднее метилирование MAOA не было связано с оценкой симптомов ASPD (33) и не было значимо связано с MAOA Генотипы VTNR. Однако в двух независимых исследованиях с NR3C1 получены разные результаты. Следовательно, гиперметилирование отдельных CpG-сайтов в экзоне 1F промотора NR3C1 , но не общее метилирование, было связано с симптомами экстернализации (34).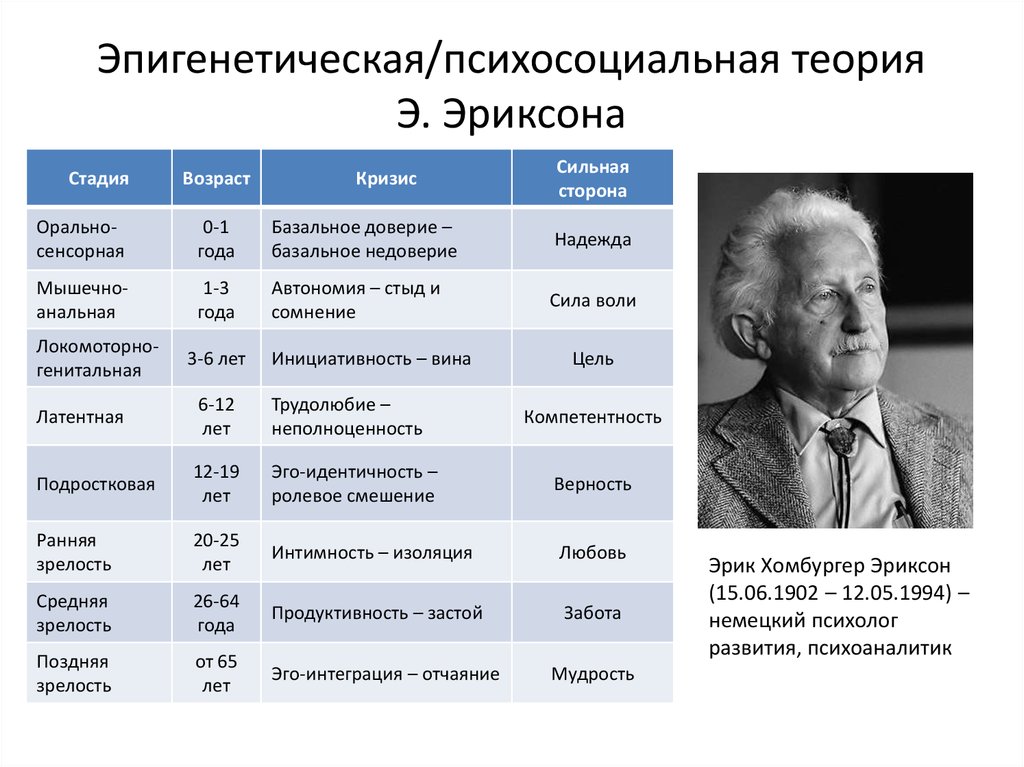 Это согласуется с результатами другого исследования, в котором метилирование экзона 1F промотора NR3C1 не показало корреляции с психометрическими показателями, но с учетом жестокого обращения в детстве оба фактора взаимодействовали при прогнозировании уязвимости к психопатологии (36). Интересно, NR3C1 был наиболее изучен при ПРЛ (-> пограничное расстройство личности). Только два исследования рассматривали ELA как независимый фактор (30, 36). Во всех исследованиях не удалось получить здоровую контрольную группу; большинство исследований ограничено только мужчинами или женщинами. Только в одном исследовании анализировалась транскрипционная значимость обнаруженных изменений метилирования с помощью уровня белка (32). Таким образом, представленные исследования антисоциальных и ассоциированных с ПРЛ черт личности пока не могут служить основой для общих выводов.
Это согласуется с результатами другого исследования, в котором метилирование экзона 1F промотора NR3C1 не показало корреляции с психометрическими показателями, но с учетом жестокого обращения в детстве оба фактора взаимодействовали при прогнозировании уязвимости к психопатологии (36). Интересно, NR3C1 был наиболее изучен при ПРЛ (-> пограничное расстройство личности). Только два исследования рассматривали ELA как независимый фактор (30, 36). Во всех исследованиях не удалось получить здоровую контрольную группу; большинство исследований ограничено только мужчинами или женщинами. Только в одном исследовании анализировалась транскрипционная значимость обнаруженных изменений метилирования с помощью уровня белка (32). Таким образом, представленные исследования антисоциальных и ассоциированных с ПРЛ черт личности пока не могут служить основой для общих выводов.
Агрессия
В рамках 21-летнего лонгитюдного исследования ранней агрессии исследовательская группа Wang et al. классифицировали детей по выраженности и стойкости агрессивного поведения в возрасте 6–15 лет и установили четыре состояния траекторий детской агрессии: первое состояло из испытуемых без физической агрессии в любые моменты времени (отсутствие физической агрессии, НПА), второе — у субъекты с агрессией от низкой до умеренной, которая снизилась в возрасте 10–15 лет (низкая физическая агрессия, LPA), третье место среди субъектов с высоким уровнем агрессии, которое впоследствии снизилось (высокая физическая агрессия, ограниченная детством, CLHPA), и, наконец, субъекты со стабильно высоким уровнем агрессии. физическая агрессия до 15 лет (хроническая высокая физическая агрессия, CPA).
классифицировали детей по выраженности и стойкости агрессивного поведения в возрасте 6–15 лет и установили четыре состояния траекторий детской агрессии: первое состояло из испытуемых без физической агрессии в любые моменты времени (отсутствие физической агрессии, НПА), второе — у субъекты с агрессией от низкой до умеренной, которая снизилась в возрасте 10–15 лет (низкая физическая агрессия, LPA), третье место среди субъектов с высоким уровнем агрессии, которое впоследствии снизилось (высокая физическая агрессия, ограниченная детством, CLHPA), и, наконец, субъекты со стабильно высоким уровнем агрессии. физическая агрессия до 15 лет (хроническая высокая физическая агрессия, CPA).
В описанной когорте исследования Wang et al. (37) изучали метилирование SLC6A4 в сравнении здоровых молодых взрослых субъектов мужского пола с ограниченной в детстве высокой физической агрессией (CLHPA, n = 7) и лиц с отсутствием и низкой физической агрессией в детстве, объединив последних в качестве контрольной группы. ( n = 18). Они проанализировали ДНК, выделенную из изолированных периферических моноцитов и Т-клеток. Не в Т-клетках, а в моноцитах авторы обнаружили значительно более высокий средний уровень метилирования по всем сайтам CpG, и авторы предполагают, что моноциты более надежны в изучении их ассоциации. Однако функциональный анализ люциферазы показал значительное снижение Активность транскрипции гена SLC6A4 как для Т-клеток, так и для моноцитов в группе CLHPA. Что касается конкретных сайтов CpG по отдельности, метилирование в CpG11 и CpG12 имело высокую положительную корреляцию и значительно выше в группе CLHPA. Индивидуальное метилирование в CpG5 и CpG6 было сильно коррелировано и выше у субъектов CLHPA. Не было обнаружено связи между средними уровнями метилирования SLC6A4 с уровнем мРНК и с существующим генотипом полиморфной области, связанной с переносчиком серотонина (5-HTTLPR). В дополнение, при ПЭТ-сканировании черепа авторы наблюдали значительную отрицательную связь между уровнями метилирования в CpG11 и CpG12 в Т-клетках или в CpG5 и CpG6 в моноцитах, соответственно, и in vivo Синтез 5-HT в латеральной левой и правой орбитофронтальной коре (OBFC), что указывает на долгосрочные функциональные эффекты in vivo .
( n = 18). Они проанализировали ДНК, выделенную из изолированных периферических моноцитов и Т-клеток. Не в Т-клетках, а в моноцитах авторы обнаружили значительно более высокий средний уровень метилирования по всем сайтам CpG, и авторы предполагают, что моноциты более надежны в изучении их ассоциации. Однако функциональный анализ люциферазы показал значительное снижение Активность транскрипции гена SLC6A4 как для Т-клеток, так и для моноцитов в группе CLHPA. Что касается конкретных сайтов CpG по отдельности, метилирование в CpG11 и CpG12 имело высокую положительную корреляцию и значительно выше в группе CLHPA. Индивидуальное метилирование в CpG5 и CpG6 было сильно коррелировано и выше у субъектов CLHPA. Не было обнаружено связи между средними уровнями метилирования SLC6A4 с уровнем мРНК и с существующим генотипом полиморфной области, связанной с переносчиком серотонина (5-HTTLPR). В дополнение, при ПЭТ-сканировании черепа авторы наблюдали значительную отрицательную связь между уровнями метилирования в CpG11 и CpG12 в Т-клетках или в CpG5 и CpG6 в моноцитах, соответственно, и in vivo Синтез 5-HT в латеральной левой и правой орбитофронтальной коре (OBFC), что указывает на долгосрочные функциональные эффекты in vivo . Авторы обсуждают свои результаты с осторожностью из-за небольшого размера выборки (37). В связи с вышеупомянутым исследованием Provençal et al. (38) сравнили взрослых субъектов мужского пола с хронической высокой физической агрессией (CPA, n = 8) с субъектами объединенной когорты трех других определенных выше групп агрессии (37), у которых во взрослом возрасте развились нормальные траектории агрессии (9).0009 н = 12). Они изучили статус метилирования ДНК, выделенной из периферических моноцитов и Т-клеток, с помощью всеобъемлющего микрочипа, охватывающего всю область генома, включая цитокины IL1A, IL6, IL8, IL4 и IL10, а также субъединицу 1 ядерного фактора каппа-В факторов транскрипции ( NFkB1 ), ядерный фактор активированных Т-клеток 5 ( NFAT5 ), преобразователь сигнала и активатор транскрипции 6 ( STAT6 ) в обеих группах исследования. Результаты показали значительные различия в метилировании каждого из проанализированных локусов генов цитокинов. У субъектов CPA общий уровень метилирования был значительно снижен в IL1A, IL4, IL6 и IL8 , и были значительно повышены в IL10 .
Авторы обсуждают свои результаты с осторожностью из-за небольшого размера выборки (37). В связи с вышеупомянутым исследованием Provençal et al. (38) сравнили взрослых субъектов мужского пола с хронической высокой физической агрессией (CPA, n = 8) с субъектами объединенной когорты трех других определенных выше групп агрессии (37), у которых во взрослом возрасте развились нормальные траектории агрессии (9).0009 н = 12). Они изучили статус метилирования ДНК, выделенной из периферических моноцитов и Т-клеток, с помощью всеобъемлющего микрочипа, охватывающего всю область генома, включая цитокины IL1A, IL6, IL8, IL4 и IL10, а также субъединицу 1 ядерного фактора каппа-В факторов транскрипции ( NFkB1 ), ядерный фактор активированных Т-клеток 5 ( NFAT5 ), преобразователь сигнала и активатор транскрипции 6 ( STAT6 ) в обеих группах исследования. Результаты показали значительные различия в метилировании каждого из проанализированных локусов генов цитокинов. У субъектов CPA общий уровень метилирования был значительно снижен в IL1A, IL4, IL6 и IL8 , и были значительно повышены в IL10 . Метилирование положительно коррелировало с уровнями сывороточного белка IL1A (значительный), IL4 и IL-6 и отрицательно коррелировало с уровнями IL-8 и IL-10 соответственно. Что касается факторов транскрипции, то два региона, близко расположенные к TSS каждой изоформы STAT6 , были значительно более метилированы в группе CPA, что было достоверно связано с более низкой транскрипцией IL4. Далее, Провансаль и др. провел GWA (39) в связи с CPA путем сравнения молодых взрослых мужчин с ранним развитием CPA ( n = 8) с субъектами без какой-либо истории высокой физической агрессии в детстве (NPA, n = 16) в соответствии с предыдущим определением исследования ( 37, 38). Они проанализировали ДНК периферических Т-клеток и обнаружили, что раннее начало CPA связано с кластерными и общегеномными различиями в метилировании в 900 сайтах в пределах 448 различных промоторов генов. Выявленные гены включали несколько генов, которые ранее были связаны с агрессией: рецептор аргинин-вазопрессина 1А ( AVPR1A ), серотонинового рецептора 1D ( HTR1D ) и глутаматного метаботропного рецептора 5 ( GRM5 ) с меньшим метилированием в группе CPA, и ген дофаминового рецептора D1 ( DRD1 ) и SLC069 с более высоким метилированием в группе СРА.
Метилирование положительно коррелировало с уровнями сывороточного белка IL1A (значительный), IL4 и IL-6 и отрицательно коррелировало с уровнями IL-8 и IL-10 соответственно. Что касается факторов транскрипции, то два региона, близко расположенные к TSS каждой изоформы STAT6 , были значительно более метилированы в группе CPA, что было достоверно связано с более низкой транскрипцией IL4. Далее, Провансаль и др. провел GWA (39) в связи с CPA путем сравнения молодых взрослых мужчин с ранним развитием CPA ( n = 8) с субъектами без какой-либо истории высокой физической агрессии в детстве (NPA, n = 16) в соответствии с предыдущим определением исследования ( 37, 38). Они проанализировали ДНК периферических Т-клеток и обнаружили, что раннее начало CPA связано с кластерными и общегеномными различиями в метилировании в 900 сайтах в пределах 448 различных промоторов генов. Выявленные гены включали несколько генов, которые ранее были связаны с агрессией: рецептор аргинин-вазопрессина 1А ( AVPR1A ), серотонинового рецептора 1D ( HTR1D ) и глутаматного метаботропного рецептора 5 ( GRM5 ) с меньшим метилированием в группе CPA, и ген дофаминового рецептора D1 ( DRD1 ) и SLC069 с более высоким метилированием в группе СРА.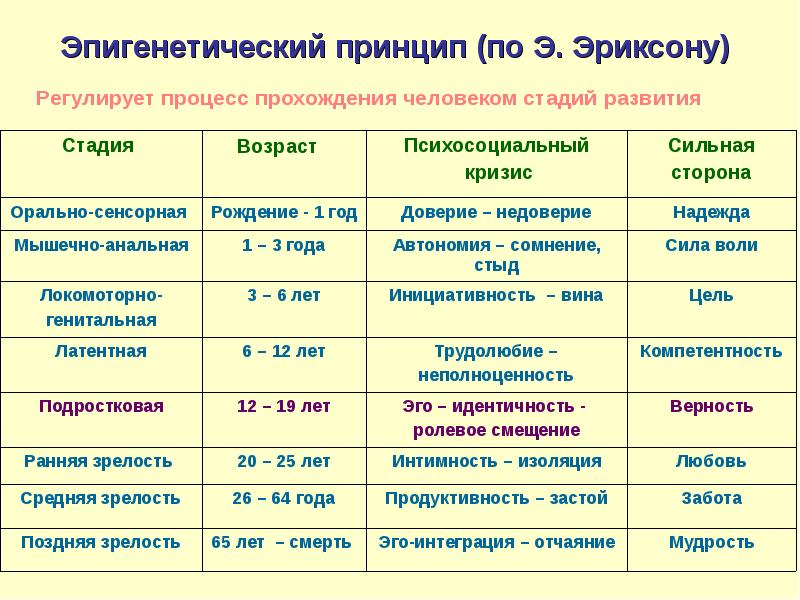 В целом, большинство функциональных категорий генов с разным метилированием при ЦПА включали поведение (гиперактивность), метаболические и неврологические заболевания, воспалительный ответ (хемотаксис и фагоциты), клеточный рост и пролиферацию, а также экспрессию генов (транскрипционные факторы, преобразователь сигнала как 9).0009 СТАТ6 ). Рассматриваемые специфические канонические пути включали передачу сигналов цитокинов и передачу сигналов рецепторов, связанных с G-белком.
В целом, большинство функциональных категорий генов с разным метилированием при ЦПА включали поведение (гиперактивность), метаболические и неврологические заболевания, воспалительный ответ (хемотаксис и фагоциты), клеточный рост и пролиферацию, а также экспрессию генов (транскрипционные факторы, преобразователь сигнала как 9).0009 СТАТ6 ). Рассматриваемые специфические канонические пути включали передачу сигналов цитокинов и передачу сигналов рецепторов, связанных с G-белком.
В другом GWA Guillemin et al. изучали аберрации метилирования в ДНК изолированных периферических Т-клеток у женщин с траекториями CPA с детства ( n = 5) по сравнению с женщинами без какой-либо агрессии в анамнезе ( n = 16) (40). В общей сложности 917 зондов, соответствующих 430 различным промоторам генов, были дифференциально метилированы, многие из которых вовлечены в иммунные и воспалительные реакции, такие как FOS, GATA1, GATA3 , фактор роста гепатоцитов ( HGF ), TNF, IFNG, IL1A, IL1B, IL10, IL13, IL17R и IL18 .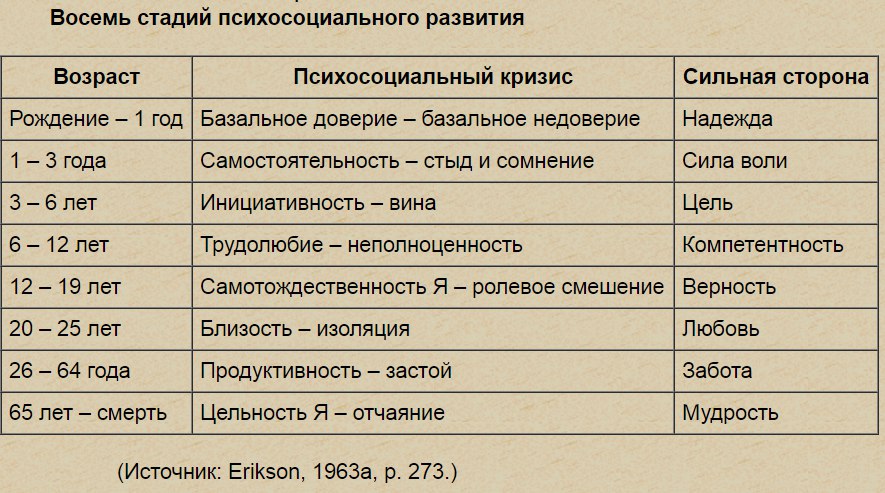 Ранее было показано, что некоторые из них связаны с агрессивным поведением, включая триптофангидроксилазу 2 ( TPh3 ), NR3C1 и белок, связывающий гормон, высвобождающий кортикотропин ( CRHBP ). субъектам ЦПУ.
Ранее было показано, что некоторые из них связаны с агрессивным поведением, включая триптофангидроксилазу 2 ( TPh3 ), NR3C1 и белок, связывающий гормон, высвобождающий кортикотропин ( CRHBP ). субъектам ЦПУ.
На втором этапе исследователи сравнили свои результаты с их предыдущими данными о GWA у мужчин из того же основного лонгитюдного исследования, проведенного Provençal et al. (39). Уровни метилирования промоторов 31 гена, как оказалось, связаны с физической агрессией как у женщин, так и у мужчин, и, таким образом, они составляют значительное перекрытие, включая TPh3, CRHBP и NR3C1 . В области промотора гена белка цинковых пальцев 366 ( ZNF366 ) связанное с агрессией метилирование было по-разному направлено как у женщин, так и у мужчин (40).
Ван Донген и др. (41) изучали различия в метилировании в отношении агрессивного поведения у 40 монозиготных близнецов (20 пар), резко дискордантных по агрессивным признакам. Они выполнили GWA с ДНК PBC и получили около 24 сайтов метилирования с высокой разницей в метилировании, но различия, как правило, были специфичны для пары близнецов; таким образом, в этом образце не было выявлено статистически значимых различий в метилировании по всему геному.
С учетом основных результатов исследований агрессивных признаков четыре из пяти исследований были получены в одной и той же исследовательской группе. Два исследования были основаны на теории и нацелены на гены. Их результаты указывают на повышенное метилирование SLC6A4 у здоровых мужчин с историей CLHPA с последствиями для транскрипционной активности in vitro и показали ассоциации с 5-НТ головного мозга в левом латеральном OBFC (37). Более полный анализ области генома, включая IL1A, IL4, IL6, IL8, IL10 и факторы транскрипции NFkB1, NFAT5 и STAT6 выявили значительные различия в метилировании у мужчин с CPA в релевантных для транскрипции сайтах вблизи или внутри каждого из исследованных генов. Однако корреляция с уровнями интерлейкина в сыворотке плазмы была значимой только для IL1A . Многие из идентифицированных аберраций метилирования касались участков, расположенных за пределами внутренних последовательностей генов, что подчеркивает актуальность анализов метилирования, которые не ограничиваются целевыми последовательностями генов (38). Дополнительное исследование у женщин с CPA показало аналогичные результаты для иммунных и воспалительных генов (40). Результаты GWA подтвердили участие всего генома в различиях метилирования в отношении агрессивных черт у мужчин (39).). Упомянутые исследования проводились на небольшой выборке из 8 (мужчины) и 5 (женщин) испытуемых соответственно в группе агрессии. Ни в одном из исследований не рассматривался опыт детской травмы.
Дополнительное исследование у женщин с CPA показало аналогичные результаты для иммунных и воспалительных генов (40). Результаты GWA подтвердили участие всего генома в различиях метилирования в отношении агрессивных черт у мужчин (39).). Упомянутые исследования проводились на небольшой выборке из 8 (мужчины) и 5 (женщин) испытуемых соответственно в группе агрессии. Ни в одном из исследований не рассматривался опыт детской травмы.
В целом, представленные исследования подтверждают необходимость полногеномных анализов метилирования или, по крайней мере, сосредоточения внимания на комплексном наборе функционально ассоциированных генов. Будущие исследования должны учитывать детскую травму и определять подходящую контрольную группу.
Импульсивность
Выполняя GWA Ruggeri et al. (42) изучали аберрации метилирования в ДНК ПБЦ монозиготных пар близнецов ( n = 18) дискордантно для расстройства, связанного с употреблением алкоголя. Авторы получили 77 дифференциально метилированных участков, ассоциированных с 62 генами. Повторение этих результатов на микрочипе выявило значительные изменения метилирования только в одном сайте CpG в 3’UTR гена 3′-протеин-фосфатазы-1G ( PPM1G ). Гиперметилирование в PPM1G положительно коррелировало с более низкими уровнями мРНК PPM1G и в значительной степени ассоциировалось с ранней эскалацией употребления алкоголя, а также с повышенной импульсивностью. Дальнейшие анализы показали, что Гиперметилирование PPM1G было независимо связано как с импульсивностью признаков, так и с активацией правого субталамического ядра, предположительно из-за повышенных усилий по осуществлению контрольного торможения. Поскольку PPM1G состоит из пяти SNP, авторы провели регрессионный анализ, который мог исключить значительное влияние вариаций генотипа на метилирование PPM1G . Кроме того, не было обнаружено корреляции SNP с чертой импульсивности (42).
Повторение этих результатов на микрочипе выявило значительные изменения метилирования только в одном сайте CpG в 3’UTR гена 3′-протеин-фосфатазы-1G ( PPM1G ). Гиперметилирование в PPM1G положительно коррелировало с более низкими уровнями мРНК PPM1G и в значительной степени ассоциировалось с ранней эскалацией употребления алкоголя, а также с повышенной импульсивностью. Дальнейшие анализы показали, что Гиперметилирование PPM1G было независимо связано как с импульсивностью признаков, так и с активацией правого субталамического ядра, предположительно из-за повышенных усилий по осуществлению контрольного торможения. Поскольку PPM1G состоит из пяти SNP, авторы провели регрессионный анализ, который мог исключить значительное влияние вариаций генотипа на метилирование PPM1G . Кроме того, не было обнаружено корреляции SNP с чертой импульсивности (42).
Исследования личностных качеств обобщены в таблице 2.
Таблица 2 . Исследования по эпигенетике в чертах личности.
Исследования по эпигенетике в чертах личности.
Обсуждение
Цель этого обзора состояла в том, чтобы обобщить первое полное количество современных оригинальных публикаций о расстройствах личности человека и личностных чертах в связи с эпигенетическими модификациями, а также обсудить результаты в отношении патогенетического воздействия и выводов, относящихся к лечению. Примечательно, что поиск литературы не дал ни одного исследования эпигенетических оценок нарциссических, истерических, ананкастических, избегающих, зависимых, эксцентричных, параноидальных, шизоидных и шизотипических аспектов личности. Не было ни результатов с каким-либо ключевым словом по модификации гистонов, ни результатов исследований микроРНК, а только аберраций метилирования. Преимущественно изучались агрессивность и антисоциальные черты, возможно, предпочтительные, поскольку эти поведенческие характеристики имеют высокую генетическую нагрузку [например, (43)].
Большинство теоретических исследований были сосредоточены на локусах генов, участвующих в серотонинергической, дофаминергической или норадренергической системе нейромедиаторов, например, рецепторе и транспортере серотонина, рецепторе дофамина и МАОА. Дальнейший критический интерес представляла эпигенетическая модуляция функциональности нейротрофических факторов, контура оси HPA и окситоцинергической системы. Это кажется последовательным, поскольку эти генные продукты конститутивны для функционирования мозга. Таким образом, MAOA, фермент, кодируемый X-хромосомой, отвечает за окислительный распад моноаминовых нейротрансмиттеров, то есть в мозге, в частности, серотонина, адреналина, норадреналина и дофамина. Мутации в гене MAOA с нарушением активности гена приводят к избытку серотонина и норадреналина, что связано с нарушением контроля над импульсивностью и агрессией пораженного субъекта (NIH, Genetics Home Reference). BDNF участвует в росте, созревании и поддержании нервных клеток и играет решающую роль в построении синапсов и синаптической пластичности. Полиморфизмы в гене BDNF связаны с повышенным риском психических расстройств, таких как биполярное расстройство, тревога и расстройства пищевого поведения. Однако результаты полногеномных ассоциативных исследований относятся к эпигенетическим изменениям генов, которые, помимо нейрофункциональных генов, включают геномные области, влияющие на гены, участвующие в воспалении, передаче клеточных сигналов, метаболизме и генах, кодирующих сами белки транскрипционного аппарата (27, 29).
Дальнейший критический интерес представляла эпигенетическая модуляция функциональности нейротрофических факторов, контура оси HPA и окситоцинергической системы. Это кажется последовательным, поскольку эти генные продукты конститутивны для функционирования мозга. Таким образом, MAOA, фермент, кодируемый X-хромосомой, отвечает за окислительный распад моноаминовых нейротрансмиттеров, то есть в мозге, в частности, серотонина, адреналина, норадреналина и дофамина. Мутации в гене MAOA с нарушением активности гена приводят к избытку серотонина и норадреналина, что связано с нарушением контроля над импульсивностью и агрессией пораженного субъекта (NIH, Genetics Home Reference). BDNF участвует в росте, созревании и поддержании нервных клеток и играет решающую роль в построении синапсов и синаптической пластичности. Полиморфизмы в гене BDNF связаны с повышенным риском психических расстройств, таких как биполярное расстройство, тревога и расстройства пищевого поведения. Однако результаты полногеномных ассоциативных исследований относятся к эпигенетическим изменениям генов, которые, помимо нейрофункциональных генов, включают геномные области, влияющие на гены, участвующие в воспалении, передаче клеточных сигналов, метаболизме и генах, кодирующих сами белки транскрипционного аппарата (27, 29). , 39–42). Таким образом, различия в метилировании представляют собой сложный паттерн, который исключает возможные тесты функциональной проверки соответствующих генных локусов, поскольку это усложняет окончательную интерпретацию. Все рассмотренные здесь исследования GWA, за исключением Prados et al., не учитывают дополнительные факторы, возможно влияющие на эпигенетическую конфигурацию, особенно опыт раннего детства, который также представляет основной интерес для понимания развития БП. Например, невзгоды в раннем возрасте были связаны с метилированием SLC6A4 , что, в свою очередь, показало связь со структурой и функцией мозга (44). Напротив, большинство исследований БП и анализов отдельных генов рассматривали детскую травму как фактор, вмешивающийся в их эпигенетический анализ. Согласно предыдущим данным, эти целевые исследования генов подтвердили значимость серотонинергической системы для аффективной регуляции и выявили нарушения метилирования генов переносчика серотонина ( SLC6A4 ) и рецепторов ( 5HTR1B, 5HTR2A и 9).
, 39–42). Таким образом, различия в метилировании представляют собой сложный паттерн, который исключает возможные тесты функциональной проверки соответствующих генных локусов, поскольку это усложняет окончательную интерпретацию. Все рассмотренные здесь исследования GWA, за исключением Prados et al., не учитывают дополнительные факторы, возможно влияющие на эпигенетическую конфигурацию, особенно опыт раннего детства, который также представляет основной интерес для понимания развития БП. Например, невзгоды в раннем возрасте были связаны с метилированием SLC6A4 , что, в свою очередь, показало связь со структурой и функцией мозга (44). Напротив, большинство исследований БП и анализов отдельных генов рассматривали детскую травму как фактор, вмешивающийся в их эпигенетический анализ. Согласно предыдущим данным, эти целевые исследования генов подтвердили значимость серотонинергической системы для аффективной регуляции и выявили нарушения метилирования генов переносчика серотонина ( SLC6A4 ) и рецепторов ( 5HTR1B, 5HTR2A и 9).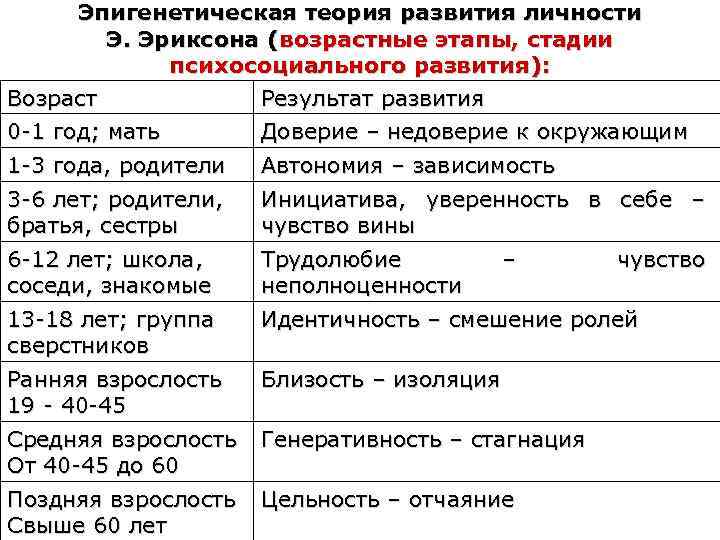 0009 5HTR3A ), чтобы быть связанным с антисоциальными чертами (30, 31) и ПРЛ (25, 26). Также было показано, что для DRD2 статус метилирования связан с ПРЛ (24). Основываясь на его влиянии на пути каждого серотонина, дофамина и норадреналина, было обнаружено, что эпигенетические изменения MAOA конгруэнтно связаны с антисоциальными чертами личности, ASPD и BPD (19, 20, 26, 33). Исследования также указывают на значительную роль статуса метилирования экзона 1 промотора NR3C1 в антисоциальных чертах, а также, в сочетании с невзгодами в раннем возрасте, в ПРЛ (22, 26). В самом деле, с пониманием участия эпигенетически модифицированных локусов генов в масштабах всего генома достоверность исследований конкретных генов сегодня, по-видимому, ограничена.
0009 5HTR3A ), чтобы быть связанным с антисоциальными чертами (30, 31) и ПРЛ (25, 26). Также было показано, что для DRD2 статус метилирования связан с ПРЛ (24). Основываясь на его влиянии на пути каждого серотонина, дофамина и норадреналина, было обнаружено, что эпигенетические изменения MAOA конгруэнтно связаны с антисоциальными чертами личности, ASPD и BPD (19, 20, 26, 33). Исследования также указывают на значительную роль статуса метилирования экзона 1 промотора NR3C1 в антисоциальных чертах, а также, в сочетании с невзгодами в раннем возрасте, в ПРЛ (22, 26). В самом деле, с пониманием участия эпигенетически модифицированных локусов генов в масштабах всего генома достоверность исследований конкретных генов сегодня, по-видимому, ограничена.
Помимо общей связи эпигенетических изменений с расстройствами личности и чертами, результаты исследований сильно различаются в отношении конкретных участков генов эпигенетических различий. Одной из основных причин может быть отсутствие или несопоставимость соответствующей контрольной группы в каждом исследовании. Другим методическим недостатком является непостоянный учет факторов внешней среды, в частности неблагоприятных условий раннего детства. Эпигенетически уязвимые факторы также включают воздействие насилия со стороны интимного партнера во время беременности, а также настроение, привычки курения или диеты матери во время беременности, а также стресс у субъекта во время зародыша или в раннем периоде жизни (45–49).). Примечательно, что во многих исследованиях размер выборки очень мал и может повлиять на результаты.
Другим методическим недостатком является непостоянный учет факторов внешней среды, в частности неблагоприятных условий раннего детства. Эпигенетически уязвимые факторы также включают воздействие насилия со стороны интимного партнера во время беременности, а также настроение, привычки курения или диеты матери во время беременности, а также стресс у субъекта во время зародыша или в раннем периоде жизни (45–49).). Примечательно, что во многих исследованиях размер выборки очень мал и может повлиять на результаты.
Во всех исследованиях для анализа метилирования использовалась ДНК, выделенная из клеток периферической крови, хотя и различных клеточных фракций, предпочтительно клеток цельной крови, лейкоцитов, лимфоцитов, Т-клеток. Имеются данные о том, что периферические мононуклеарные клетки, особенно лимфоциты, демонстрируют сходный эпигенетический паттерн с тканями головного мозга [т.е. (9)], и использование клеток периферической крови получило признание для эпигенетических исследований нервно-психических расстройств. Но результаты также указывают на различия в эпигенетических паттернах между линиями лейкоцитов крови (37, 38), что также может объяснить расхождения в исследованиях. Будущие исследования должны учитывать тип клеток и препарат из цельной крови, поскольку клеточный стресс может вызвать эпигенетические изменения в клетках (50).
Но результаты также указывают на различия в эпигенетических паттернах между линиями лейкоцитов крови (37, 38), что также может объяснить расхождения в исследованиях. Будущие исследования должны учитывать тип клеток и препарат из цельной крови, поскольку клеточный стресс может вызвать эпигенетические изменения в клетках (50).
Высокие требования к техническим методам и целесообразному дизайну исследования делают распутывание личностно-специфических паттернов метилирования сложной задачей, тем более что может существовать более одного способа (эпи-)генетически ускорить развитие БП. Это также может быть причиной разнообразия конечных клинических эпифенотипов этих расстройств. И наоборот, разнообразие патогенеза и индивидуально развивающихся клинических форм затрудняет отнесение возможно выявленных генетических или эпигенетических аберраций к определенной черте личности или расстройству.
Если функциональная атрибуция эпигенетических аберраций с расстройствами личности так сильно затруднена, какую пользу мы можем получить при этом?
Возможность эпигенетического подтипа помогает оценить индивидуальное патофизиологическое состояние, а также улучшить дедуктивные терапевтические подходы. Выявление чьего-либо профиля эпигенетического риска может инициировать усилия по установлению стандартов интенсивного клинического наблюдения за больными детьми и методов раннего обучения родителей или преждевременного вмешательства с целью предотвращения дальнейшего развития полномасштабной клинической картины БП. Как убедительно показали Cecil et al., различные эпигенетические состояния у психопатологически сходных субъектов могут вызывать различные последствия для индивидуальных потребностей в лечении (18).
Выявление чьего-либо профиля эпигенетического риска может инициировать усилия по установлению стандартов интенсивного клинического наблюдения за больными детьми и методов раннего обучения родителей или преждевременного вмешательства с целью предотвращения дальнейшего развития полномасштабной клинической картины БП. Как убедительно показали Cecil et al., различные эпигенетические состояния у психопатологически сходных субъектов могут вызывать различные последствия для индивидуальных потребностей в лечении (18).
Как и при других психических расстройствах, эпигенетические паттерны могут помочь предсказать реакцию на лечение. Таким образом, при депрессии отдельные аберрации метилирования могут предсказать различную реакцию на терапию эсциталопрамом и между эсциталопрамом и нортриптилином (51, 52). При пограничном расстройстве личности уровни метилирования APBA3 и MCF2 были предикторами исхода психотерапии (53).
Современная современная терапия рака включает эпигенетические препараты, нацеленные, т. е. на подавленные гены, подавляющие опухоль. Из-за множества и индивидуального разнообразия генов, вовлеченных в психические расстройства, использование деметилирующих препаратов не может быть нацелено на отдельные гены. Побочные эффекты при общем гипометилировании могут противоречить ожидаемой пользе. Но есть данные о том, что такие метилдонорские компоненты, как вальпроевая или фолиевая кислота, сами по себе, а также усиливая действие флуоксетина, обладают положительным терапевтическим эффектом при психических расстройствах (54–57).
е. на подавленные гены, подавляющие опухоль. Из-за множества и индивидуального разнообразия генов, вовлеченных в психические расстройства, использование деметилирующих препаратов не может быть нацелено на отдельные гены. Побочные эффекты при общем гипометилировании могут противоречить ожидаемой пользе. Но есть данные о том, что такие метилдонорские компоненты, как вальпроевая или фолиевая кислота, сами по себе, а также усиливая действие флуоксетина, обладают положительным терапевтическим эффектом при психических расстройствах (54–57).
Помимо основного нейробиологического эффекта, психотропные препараты могут дополнительно оказывать прямое эпигенетическое действие. Исследования антидепрессантов и антипсихотических препаратов, т. е. показали их способность модулировать эпигеном путем ацетилирования связанных с генами гистонов, изменений метилирования в дофаминовых путях (58), увеличения экспрессии ДНК-метилтрансфераз и индукции ремоделирования хроматина (58, 59). ). Аберрации метилирования также можно ремодулировать с помощью психофармакологического лечения или психотерапии (60, 61).
Эти чисто показательные данные свидетельствуют о высокой динамике эпигенетических процессов при психических расстройствах и их течении. Взаимодействие психотропных препаратов и других терапевтических вмешательств с эпигенетическим ремоделированием кажется еще недостаточно изученным. Наконец, факт этого взаимодействия можно было бы использовать в качестве интраиндивидуального контроля длительной реакции на терапию.
Последствия и варианты на будущее
Патофизиологический принцип взаимодействия генов со средой не только объясняет очевидные различия в выраженности или сочетании личностных черт человека, но и предполагает существование генетических и эпигенетических факторов риска и защитных факторов при формировании конечная структура личности. До сих пор эпигенетические исследования подчеркивали влияние невзгод в раннем возрасте на многофакторный путь к БП. Они выявили несколько соответствующих генных локусов, которые эпигенетически затрагиваются при БП, но различаются между исследованиями. Частично это можно объяснить многофакторным и многоэтапным генезом каждой болезни Паркинсона, что приводит к различным патогенетическим подтипам.
Частично это можно объяснить многофакторным и многоэтапным генезом каждой болезни Паркинсона, что приводит к различным патогенетическим подтипам.
Эпигенетический анализ в связи с болезнью Паркинсона представляет собой сложную, но подходящую поправку к выяснению развития личности и представляет собой ценный диагностический шаг в спецификации преморбидных рисков человека и для разработки индивидуально адаптированных терапевтических стратегий. Они могут сыграть ценную роль в будущей разработке и наблюдении за ранним и персонализированным терапевтическим вмешательством и, таким образом, могут помочь предотвратить развитие тяжелых дисфункциональных расстройств поведения или расстройств личности у субъектов риска.
Вклад автора
DG провела полный поиск литературы, прочитала все найденные статьи, написала рукопись. К.К. критически переработал фокус и рукопись. TH и HF предоставили экспертные консультации по эпигенетике. JK и TF рассмотрели рукопись и совместно интерпретировали результаты и участвовали в обсуждении.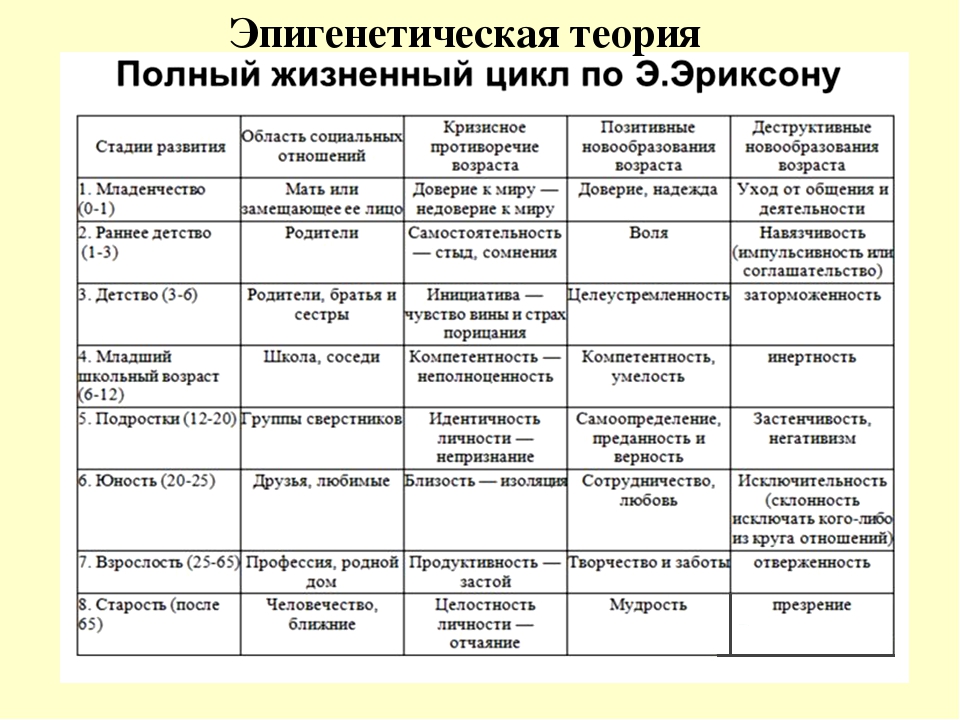
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Ссылки
1. Шиф М., Макгоуэн П., Мини М. Социальная среда и эпигеном. Environ Mol Мутаген. (2008) 49:46–60. doi: 10.1002/em.20357
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
2. Шиф М., Бик Дж. Метилирование ДНК: механизм встраивания опыта ранней жизни в геном. Детская разработка. (2013) 84:49–57. doi: 10.1111/j.1467-8624.2012.01793.x
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
3. Weaver IC, Cervoni N, Champagne FA, D’Alessio AC, Sharma S, Seckl JR, et al. Эпигенетическое программирование материнским поведением. Нат Неврологи. (2004) 7:847–54. doi: 10.1038/nn1276
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
4. Рот Т.Л., Любин Ф.Д., Функ А.Дж., Суитт Д.Д. Длительное эпигенетическое влияние невзгод в раннем возрасте на ген BDNF. Биол Психиатрия (2009) 65:760–9. doi: 10.1016/j.biopsych.2008.11.028
Рот Т.Л., Любин Ф.Д., Функ А.Дж., Суитт Д.Д. Длительное эпигенетическое влияние невзгод в раннем возрасте на ген BDNF. Биол Психиатрия (2009) 65:760–9. doi: 10.1016/j.biopsych.2008.11.028
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
5. Murgatroyd C, Patchev AV, Wu Y, Micale V, Bockmuhl Y, Fischer D, et al. Динамическое метилирование ДНК программирует стойкие неблагоприятные последствия стресса в раннем возрасте. Нат Неврологи. (2009) 12:1559–66. doi: 10.1038/nn.2436
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
6. McGowan PO, Sasaki A, D’Alessio AC, Dymov S, Labonte B, Szyf M, et al. Эпигенетическая регуляция глюкокортикоидного рецептора в мозге человека связана с жестоким обращением в детстве. Нат Неврологи. (2009) 12:342–8. doi: 10.1038/nn.2270
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
7. McGowan PO, Suderman M, Sasaki A, Huang TC, Hallett M, Meaney MJ, et al. Широкая эпигенетическая подпись материнской заботы в мозгу взрослых крыс. PLoS ONE (2011) 6:e14739. doi: 10.1371/journal.pone.0014739
Широкая эпигенетическая подпись материнской заботы в мозгу взрослых крыс. PLoS ONE (2011) 6:e14739. doi: 10.1371/journal.pone.0014739
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
8. Suderman M, McGowan PO, Sasaki A, Huang TC, Hallett M, Meaney MJ, et al. Сохраненная эпигенетическая чувствительность к раннему жизненному опыту в гиппокампе крысы и человека. Proc Natl Acad Sci USA . (2012) 109 (Приложение 2): 17266–72. doi: 10.1073/pnas.1121260109
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
9. Provençal N, Suderman MJ, Guillemin C, Massart R, Ruggiero A, Wang D, et al. Признак материнского воспитания в метиломе префронтальной коры макаки-резуса и Т-клетках. Дж. Неврологи. (2012) 32:15626–42. doi: 10.1523/JNEUROSCI.1470-12.2012
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
10. Klengel T, Mehta D, Anacker C, Rex-Haffner M, Pruessner JC, Pariante CM, et al. Аллель-специфическое деметилирование ДНК FKBP5 опосредует взаимодействие генов и детской травмы. Нат Неврологи. (2013) 16:33–41. doi: 10.1038/nn.3275
Аллель-специфическое деметилирование ДНК FKBP5 опосредует взаимодействие генов и детской травмы. Нат Неврологи. (2013) 16:33–41. doi: 10.1038/nn.3275
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
11. Auta J1, Smith RC, Dong E, Tueting P, Sershen H, Boules S, et al. Дисрегуляция сети генов метилирования ДНК в лимфоцитах периферической крови больных шизофренией. Шизофр Рез. (2013) 150:312–8. doi: 10.1016/j.schres.2013.07.030
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
12. Юань Л.В., Гамби Дж.Э. Ацетилирование гистонов p300 участвует в CREB-опосредованной транскрипции на хроматине. Биохим Биофиз Акта (2001) 1541:161–9. doi: 10.1016/S0167-4889(01)00141-0
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
13. Weaver IC, Hellstrom IC, Brown SE, Andrews SD, Dymov S, Diorio J, et al. Белок, связывающий метилированную ДНК MBD2, усиливает опосредованную NGFI-A (egr-1) транскрипционную активацию глюкокортикоидного рецептора. Philos Trans R Soc Lond B Biol Sci. (2014) 369:20130513. doi: 10.1098/rstb.2013.0513
Philos Trans R Soc Lond B Biol Sci. (2014) 369:20130513. doi: 10.1098/rstb.2013.0513
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
14. Weaver IC, D’Alessio AC, Brown SE, Hellstrom IC, Dymov S, Sharma S, et al. Фактор транскрипции, индуцируемый фактором роста нервов белок а, опосредует эпигенетическое программирование: изменение эпигенетических меток с помощью непосредственно-ранних генов. Дж. Неврологи. (2007) 27:1756–68. doi: 10.1523/JNEUROSCI.4164-06.2007
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
15. Zhou Z, Hong EJ, Cohen S, Zhao WN, Ho HY, Schmidt L, et al. Специфическое для мозга фосфорилирование MeCP2 регулирует зависимую от активности транскрипцию Bdnf, рост дендритов и созревание шипов. Нейрон (2006) 52:255–69. doi: 10.1016/j.neuron.2006.09.037
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
16. Chen WG, Chang Q, Lin Y, Meissner A, West AE, Griffith EC, et al. Дерепрессия транскрипции BDNF включает кальций-зависимое фосфорилирование MeCP2. Наука (2003) 302:885–9. doi: 10.1126/science.1086446
Дерепрессия транскрипции BDNF включает кальций-зависимое фосфорилирование MeCP2. Наука (2003) 302:885–9. doi: 10.1126/science.1086446
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
17. Шамсир Л., Мохер Д., Кларк М., Герси Д., Либерати А., Петтикрю М. и др. Предпочтительные элементы отчетности для протоколов систематического обзора и метаанализа (PRISMA-P) 2015: разработка и объяснение. BMJ (2015) 349:g7647. doi: 10.1136/bmj.g7647
Полный текст CrossRef | Google Scholar
18. Сесил К.А., Лысенко Л.Дж., Джаффи С.Р., Пинго Дж.Б., Смит Р.Г., Релтон С.Л. и другие. Экологический риск, метилирование гена рецептора окситоцина (OXTR) и бессердечно-неэмоциональные черты молодежи: 13-летнее продольное исследование. Mol Psychiatry (2014) 19:1071–7. doi: 10.1038/mp.2014.95
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
19. Филиберт Р.А., Вернетт П., Плюм Дж., Пакер Х., Броди Г.Х., Бич С. Р. Взаимодействия генной среды с новым вариабельным усилителем транскрипции моноаминоксидазы А связаны с антисоциальным расстройством личности. Биол Психол. (2011) 87:366–71. doi: 10.1016/j.biopsycho.2011.04.007
Р. Взаимодействия генной среды с новым вариабельным усилителем транскрипции моноаминоксидазы А связаны с антисоциальным расстройством личности. Биол Психол. (2011) 87:366–71. doi: 10.1016/j.biopsycho.2011.04.007
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
20. Checknita D, Maussion G, Labonté B, Comai S, Tremblay RE, Vitaro F, et al. Метилирование промотора гена моноаминоксидазы А и подавление транскрипции в популяции правонарушителей с антисоциальным расстройством личности. Br J Психиатрия (2015) 206: 216–22. doi: 10.1192/bjp.bp.114.144964
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
21. Perroud N, Salzmann A, Prada P, Nicastro R, Hoeppli ME, Furrer S, et al. Ответ на психотерапию при пограничном расстройстве личности и статус метилирования гена BDNF. Transl Psychiatry (2013) 3:e207. doi: 10.1038/tp.2012.140
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
22. Мартин-Бланко А., Феррер М., Солер Дж., Салазар Дж., Вега Д., Андион О. и другие. Связь между метилированием гена глюкокортикоидного рецептора, жестоким обращением в детстве и клинической тяжестью пограничного расстройства личности. J Psychiatr Res. (2014) 57:34–40. doi: 10.1016/j.jpsychires.2014.06.011
Мартин-Бланко А., Феррер М., Солер Дж., Салазар Дж., Вега Д., Андион О. и другие. Связь между метилированием гена глюкокортикоидного рецептора, жестоким обращением в детстве и клинической тяжестью пограничного расстройства личности. J Psychiatr Res. (2014) 57:34–40. doi: 10.1016/j.jpsychires.2014.06.011
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
23. Perroud N, Paoloni-Giacobino A, Prada P, Olié E, Salzmann A, Nicastro R, et al. Повышенное метилирование гена глюкокортикоидного рецептора (NR3C1) у взрослых с жестоким обращением в детстве: связь с тяжестью и типом травмы. Перевод психиатрии (2011) 1:e59. doi: 10.1038/tp.2011.60
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
24. Groleau P, Joober R, Israel M, Zeramdini N, DeGuzman R, Steiger H. Метилирование промотора гена дофаминового рецептора D2 (DRD2) у женщин с расстройством булимического спектра: связь с пограничным расстройством личности и подверженность жестокому обращению в детстве.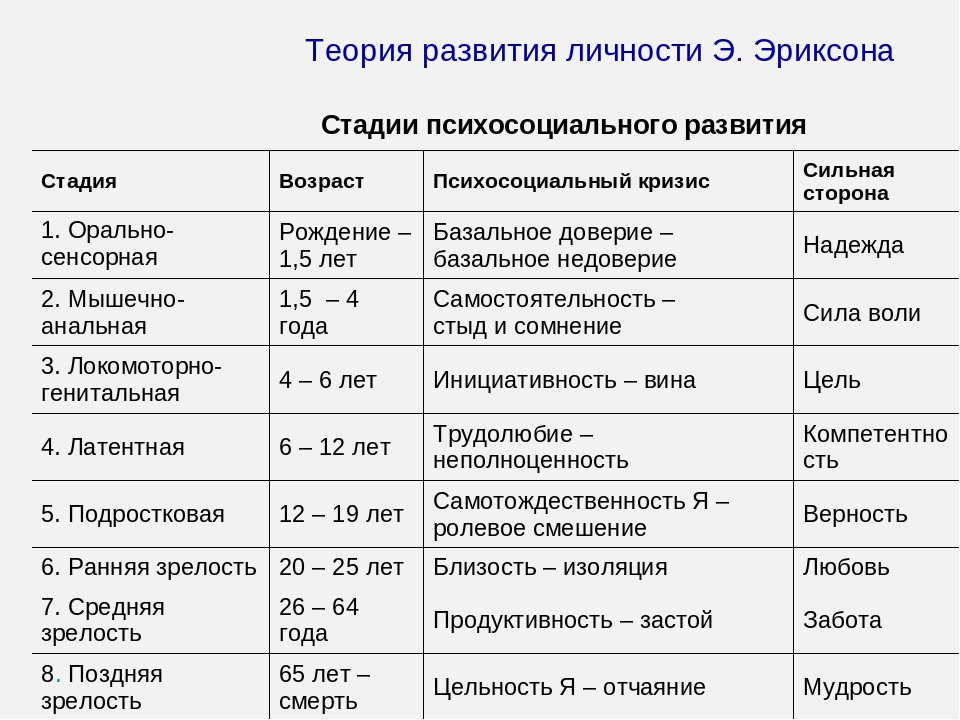 J Psychiatr Res. (2014) 48:121–7. doi: 10.1016/j.jpsychires.2013.10.003
J Psychiatr Res. (2014) 48:121–7. doi: 10.1016/j.jpsychires.2013.10.003
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
25. Perroud N, Zewdie S, Stenz L, Adouan W, Bavamian S, Prada P, et al. Метилирование серотонинового рецептора 3А при СДВГ, пограничной личности и биполярных расстройствах: связь с тяжестью расстройств и жестоким обращением в детстве. Депрессивное беспокойство (2016) 33:45–55. doi: 10.1002/da.22406
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
26. Дамманн Г., Тешлер С., Хааг Т., Альтмюллер Ф., Тучек Ф., Дамманн Р.Х. Повышенное метилирование ДНК нейропсихиатрических генов происходит при пограничном расстройстве личности. Эпигенетика (2011) 6:1454–62. doi: 10.4161/epi.6.12.18363
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
27. Teschler S, Bartkuhn M, Künzel N, Schmidt C, Kiehl S, Dammann G, et al. Аберрантное метилирование CpG-сайтов, ассоциированных с генами, происходит при пограничном расстройстве личности. PLoS ONE (2013) 8:e84180. doi: 10.1371/journal.pone.0084180
PLoS ONE (2013) 8:e84180. doi: 10.1371/journal.pone.0084180
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
28. Тешлер С., Готтхардт Дж., Дамманн Г., Дамманн Р.Х. Аберрантное ДНК-метилирование рДНК и PRIMA1 при пограничном расстройстве личности. Int J Mol Sci. (2016) 17:E67. doi: 10.3390/ijms17010067
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
29. Prados J, Stenz L, Courtet P, Prada P, Nicastro R, Adouan W, et al. Пограничное расстройство личности и жестокое обращение в детстве: анализ метилирования всего генома. Гены Поведение мозга. (2015) 14:177–88. doi: 10.1111/gbb.12197
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
30. Бич С.Р., Броуди Г.Х., Тодоров А.А., Гюнтер Т.Д., Филиберт Р.А. Метилирование в 5HTT опосредует влияние сексуального насилия над детьми на антиобщественное поведение женщин: исследование образца усыновленного ребенка из Айовы. Психосом Мед. (2011) 73:83–7. doi: 10.1097/PSY.0b013e3181fdd074
Психосом Мед. (2011) 73:83–7. doi: 10.1097/PSY.0b013e3181fdd074
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
31. Моул С., Добсон-Стоун С., Бреннан Дж., Хоуз Д.Дж., Даддс М.Р. Метилирование гена рецептора серотонина 1B (HTR1B) как фактор риска бездушно-неэмоциональных черт у антисоциальных мальчиков. PLoS ONE (2015) 10:e0126903. doi: 10.1371/journal.pone.0126903
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
32. Dadds MR, Moul C, Cauchi A, Dobson-Stone C, Hawes DJ, Brennan J, et al. Метилирование гена рецептора окситоцина и уровень окситоцина в крови при развитии психопатии. Дев Психопат. (2014) 26:33–40. doi: 10.1017/S09545700497
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
33. Philibert RA, Gunter TD, Beach SR, Brody GH, Madan A. Метилирование MAOA связано с никотиновой и алкогольной зависимостью у женщин. Нейропсихиатр Жен. (2008) 147B: 565–70. doi: 10.1002/ajmg.b.30778
doi: 10.1002/ajmg.b.30778
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
34. Dadds MR, Moul C, Hawes DJ, Mendoza Diaz A, Brennan J. Индивидуальные различия детских поведенческих расстройств, связанных с эпигенетической модуляцией гена рецептора кортизола. Детская разработка. (2015) 86:1311–20. doi: 10.1111/cdev.12391
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
35. Меллор Д. Нормативные данные для опросника сильных сторон и трудностей в Австралии. Aust Psychol. (2005) 40:215–22. doi: 10.1080/00050060500243475
CrossRef Full Text
36. Radtke KM, Schauer M, Gunter HM, Ruf-Leuschner M, Sill J, Meyer A, et al. Эпигенетические модификации гена глюкокортикоидного рецептора связаны с уязвимостью к психопатологии при жестоком обращении в детстве. Transl Psychiatry (2015) 5:e571. doi: 10.1038/tp.2015.63
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
37. Wang D, Szyf M, Benkelfat C, Provençal N, Turecki G, Caramaschi D, et al. Метилирование периферической ДНК SLC6A4 связано с in vivo показателями синтеза серотонина в мозге человека и физической агрессией в детстве. PLoS ONE (2012) 7:e39501. doi: 10.1371/journal.pone.0039501
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
38. Provençal N, Suderman MJ, Caramaschi D, Wang D, Hallett M, Vitaro F, et al. Дифференциальные области метилирования ДНК в геномных локусах цитокинов и факторов транскрипции связаны с физической агрессией в детстве. PLoS ONE (2013) 8:e71691. doi: 10.1371/journal.pone.0071691
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
39. Provençal N, Suderman MJ, Guillemin C, Vitaro F, Côté SM, Hallett M, et al. Ассоциация хронической физической агрессии в детстве с характерным метилированием ДНК в Т-клетках взрослого человека. PLoS ONE (2014) 9:e89839. doi: 10.1371/journal. pone.0089839
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
40. Guillemin C, Provençal N, Suderman M, Côté SM, Vitaro F, Hallett M, et al. Признак метилирования ДНК хронической физической агрессии в детстве в Т-клетках мужчин и женщин. PLoS ONE (2014) 9:e86822. doi: 10.1371/journal.pone.0086822
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
41. Van Dongen J, Nivard MG, Baselmans BM, Zilhão NR, Ligthart L, BIOS Consortium, et al. Эпигеномное исследование ассоциации агрессивного поведения. Twin Res Hum Genet. (2015) 18:686–98. doi: 10.1017/thg.2015.74
CrossRef Полный текст | Google Scholar
42. Ruggeri B, Nymberg C, Vuoksimaa E, Lourdusamy A, Wong CP, Carvalho FM, et al. Ассоциация протеинфосфатазы PPM1G с расстройством, связанным с употреблением алкоголя, и активностью мозга во время поведенческого контроля в анализе метилирования всего генома. Am J Psychiatry (2015) 172: 543–52. doi: 10.1176/appi.ajp.2014.14030382
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
43. Viding E, Blair RJR, Moffitt TE, Plomin R. Доказательства значительного генетического риска психопатии у 7-летних детей. J Детская психологическая психиатрия (2005) 46:592–7. doi: 10.1111/j.1469-7610.2004.00393.x
Полный текст CrossRef | Google Scholar
44. Frodl T, Szyf M, Carballedo A, Ly V, Dymov S, Vaisheva F, et al. Метилирование ДНК гена переносчика серотонина (SLC6A4) связано с функцией мозга, участвующей в обработке эмоциональных стимулов. J Психиатрия Neurosci. (2015) 40: 296–305. doi: 10.1503/jpn.140180
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
45. Рейк В. Стабильность и гибкость эпигенетической регуляции генов в развитии млекопитающих. Природа (2007) 447:425–32. doi: 10.1038/nature05918
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
46. Фейл Р., Фрага М.Ф. Эпигенетика и окружающая среда: новые модели и последствия. Nat Rev Genet (2012) 13: 97–109. doi: 10.1038/nrg3142
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
47. Radtke KM, Ruf M, Gunter HM, Dohrmann K, Schauer M, Meyer A, et al. Трансгенерационное влияние насилия со стороны интимного партнера на метилирование промотора глюкокортикоидного рецептора. Перевод психиатрии (2011) 1:e21. doi: 10.1038/tp.2011.21
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
48. Zeilinger S, Kühnel B, Klopp N, Baurecht H, Kleinschmidt A, Gieger C, et al. Табакокурение приводит к обширным полногеномным изменениям метилирования ДНК. PLoS ONE (2013) 8:e63812. doi: 10.1371/journal.pone.0063812
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
49. McGowan PO, Meaney MJ, Szyf M. Диета и эпигенетическое (пере) программирование фенотипических различий в поведении. Мозг Res. (2008) 1237:12–24. doi: 10.1016/j.brainres.2008.07.074
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
50. Philibert R, Caspers K, Beach SRH, Bakermans-Kranenburg MJ, van Ijzendoorn MH. Ответ на: Вирус Эпштейна-Барр трансформировал ДНК как источник ложноположительных результатов в исследованиях метилирования психических состояний. Biol Psychiatry (2011) 70:e27–8. doi: 10.1016/j.biopsych.2011.02.029
CrossRef Полный текст | Google Scholar
51. Домшке К., Тидоу Н., Шварте К., Декерт Дж., Леш К.П., Арольт В. и др. Гипометилирование гена переносчика серотонина предсказывает нарушение ответа на лечение антидепрессантами. Int J Neuropsychopharmacol . (2014) 17:1167–76. doi: 10.1017/S146114571400039X
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
52. Уэр Р., Уэзо-Диас П., Перроуд Н., Смит Р., Ритшель М., Морс О. и др. Генетические предикторы реакции на антидепрессанты в проекте GENDEP. Pharmacogenomics J. (2009) 9:225–33. doi: 10.1038/tpj.2009.12
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
53. Knoblich N, Gundel F, Brückmann C, Becker-Sadzio J, Frischholz C, Nieratschker V. Метилирование ДНК APBA3 и MCF2 при пограничном расстройстве личности: потенциальные биомаркеры реакции на психотерапию. Евро Нейропсихофармакол. (2018) 28: 252–63. doi: 10.1016/j.euroneuro.2017.12.010
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
54. Моррис Д.В., Триведи М.Х., Раш А.Дж. Фолаты и униполярная депрессия. J Altern Complement Med. (2008) 14: 277–85. doi: 10.1016/j.jad.2011.04.032
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
55. Фарах А. Роль L-метилфолата при депрессивных расстройствах. Спектр ЦНС. (2009) 14:2–7. doi: 10.1017/S10
3473
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
56. Коппен А., Бейли Дж. Усиление антидепрессивного действия флуоксетина фолиевой кислотой: рандомизированное плацебо-контролируемое исследование. J Аффективное расстройство. (2000) 60:121–30. doi: 10.1016/S0165-0327(00)00153-1
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
57. Godfrey PS, Toone BK, Carney MW, Flynn TG, Bottiglieri T, Laundy M, et al. Улучшение выздоровления от психических заболеваний с помощью метилфолата. Ланцет (1990) 336:392–5. doi: 10.1016/0140-6736(90)
-4
CrossRef Full Text | Google Scholar
58. Swathy B, Banerjee M. Понимание эпигенетики шизофрении на фоне ее антипсихотической лекарственной терапии. Эпигеномика (2017) 9:721–36. doi: 10.2217/epi-2016-0106
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
59. Zimmermann N, Zschocke J, Perisic T, Yu S, Holsboer F, Rein T. Антидепрессанты ингибируют ДНК-метилтрансферазу 1 за счет снижения уровня G9a. Biochem J. (2012) 448:93–102. doi: 10.1042/BJ20120674
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
60. Vialou V, Feng J, Robison AJ, Nestler EJ. Эпигенетические механизмы депрессии и действия антидепрессантов. Энн Рев Фармакол Токсикол . (2013) 53:59–87. doi: 10.1146/annurev-pharmtox-010611-134540
CrossRef Полный текст | Google Scholar
61. Yehuda R, Daskalakis NP, Desarnaud F, Makotkine I, Lehrner AL, Koch E, et al. Эпигенетические биомаркеры как предикторы и корреляты улучшения симптомов после психотерапии у ветеранов боевых действий с посттравматическим стрессовым расстройством. Передняя психиатрия (2013) 4:118. doi: 10.3389/fpsyt.2013.00118
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
какое влияние это оказывает на нашу психологию?
В битве между природой и воспитанием у воспитания появился новый рекрут: эпигенетика, взятая из молекулярной биологии, чтобы придать научную значимость аргументу о том, что гены не являются судьбой. Неопровержимые данные о генетическом влиянии на наши психологические черты вызывают у многих людей фаталистическое видение, согласно которому мы — рабы своей биологии, не контролирующие собственную психику и собственное поведение. Эпигенетика, механизм регуляции экспрессии генов, по-видимому, предлагает бегство от генетического детерминизма, средство преодолеть наши врожденные предрасположенности и изменить себя.
Эту точку зрения хорошо представляют Дипак Чопра, доктор медицины, и Рудольф Танзи, доктор медицины, профессор неврологии Гарвардской медицинской школы, которые пишут:
Каждый день приносит новые доказательства того, что связь между разумом и телом напрямую связана с активностью наших генов. То, как эта деятельность меняется в ответ на наш жизненный опыт, называется «эпигенетикой». Независимо от природы генов, которые мы унаследовали от родителей, динамические изменения на этом уровне позволяют нам практически безгранично влиять на свою судьбу.
Эта надежда связана с исследованием, которое предполагает, что определенные виды опыта у животных действительно могут привести к прикреплению эпигенетической метки к определенным генам с долгосрочными последствиями для поведения. Таким образом, эпигенетика придает некоторые механистические подтверждения идее о том, что мы можем переопределить или перезаписать гены, которые в противном случае диктовали бы наши врожденные черты и предрасположенности.
Однако в этой идее есть неотъемлемое противоречие, заключающееся в том, что механизм, придающий восприимчивость к опыту, должен в то же время фиксировать результирующие изменения. Есть даже исследования, предполагающие, что такие эпигенетические метки могут передаваться от родителей их детям и даже внукам, заставляя их вести себя определенным образом в ответ на опыт, который был у их предков. Это иронически детерминистская идея — что поведение человека будет так сильно зависеть от опыта его предков — особенно для механизма, который должен обеспечивать неограниченную поведенческую гибкость.
Чтобы оценить заявления о том, что эпигенетика может освободить нас от предопределенных психологических черт, нам нужно подробно рассмотреть, как наши гены влияют на эти черты, и что на самом деле влечет за собой эпигенетика.
Все мы закодировали в своем геноме программу создания человека с человеческим мозгом, который придает нам общую человеческую природу. Но эта программа различается у разных людей из-за многих миллионов генетических различий, которые мы все несем. Так что программа для изготовления моего мозга отличается от программы для изготовления вашего. И то, как именно работает программа, варьируется от запуска к запуску, поэтому результат различается даже между генетически идентичными близнецами. Так что наша индивидуальная природа — это уникальная вариация на общую тему.
Мы устроены по-разному: врожденные предрасположенности влияют на наш интеллект, личность, сексуальность и даже на то, как мы воспринимаем мир. Эти врожденные психологические черты не обязательно определяют наше поведение от момента к моменту, но они влияют на него как в любой данный момент, так и направляя развитие наших привычек и появление других аспектов нашего характера на протяжении всей жизни. . Но может ли эпигенетика действительно переписать эти генетические эффекты на нашу психологию?
В молекулярной биологии эпигенетика относится к клеточному механизму контроля экспрессии генов. Это особенно важно для образования различных типов клеток во время эмбрионального развития. Все наши клетки содержат один и тот же геном, содержащий около 20 000 генов, каждый из которых кодирует определенный белок, такой как коллаген, ферменты печени или рецепторы нейротрансмиттеров. Разным типам клеток для выполнения своей работы требуется разное подмножество этих белков. Итак, в каждом типе клеток некоторые гены «включаются», то есть ген транскрибируется ферментом в информационную РНК, которая затем транслируется в соответствующий белок. Другие «выключены», так что этот фрагмент ДНК просто сидит там, а белок на самом деле не производится.
Во время развития эмбриона определенные клетки получают сигнал стать мышечными, нервными или кожными клетками. Этот сигнал индуцирует экспрессию одних генов и репрессию других. Но эти сигналы часто преходящи и не сохраняются после развития, в то время как клетки все еще должны оставаться мышечными, кожными или нервными клетками. Эпигенетические механизмы включают упаковку ДНК в активное или неактивное состояние, так что исходные профили экспрессии генов сохраняются в течение всей жизни клеток. Таким образом, он действует как своего рода клеточная память. Эпигенетическое состояние клетки может даже передаваться через клеточные деления.
Неверно истолковано
К сожалению, некоторые термины в этом описании могут быть неверно истолкованы. Во-первых, это сам термин «ген». Первоначальное значение этого слова пришло из науки о наследственности и относилось к какой-то физической вещи, которая передавалась от родителей потомству и контролировала какую-то наблюдаемую черту. Теперь мы знаем, что гены в смысле наследственности на самом деле являются вариациями в последовательности ДНК, кодирующей некоторый белок. Например, «ген» серповидноклеточной анемии на самом деле является мутацией в гене, кодирующем белок гемоглобин. У всех нас одинаковый набор генов, просто разные их версии.
Во-вторых, когда мы говорим, что ген «экспрессируется», мы имеем в виду это с точки зрения молекулярной биологии. Может показаться, что речь идет о наследственности, как будто речь идет о влиянии генетической вариации на какой-то признак, который либо очевиден, либо нет. Но это совсем не одно и то же. На самом деле связь между уровнями экспрессии любого данного гена и нашими чертами обычно очень сложна и косвенна.
В-третьих, термин «клеточная память» неизбежно предполагает, что эпигенетика может лежать в основе психологической памяти и, таким образом, формировать основу нашей реакции на опыт. Хотя для формирования воспоминаний необходимы динамические изменения в экспрессии генов, нет никаких доказательств того, что сами воспоминания хранятся в паттернах экспрессии генов. Вместо этого они воплощаются в изменениях силы связей между нервными клетками, опосредованных очень локальными, субклеточными изменениями в нейроанатомии.
Наконец, идея о том, что эпигенетические модификации ДНК могут «передаваться», предназначена для клеточного деления, но звучит так, будто эпигенетические реакции на опыт могут передаваться от организма к его потомству. Хотя такой механизм действительно существует у растений и нематод, нет убедительных доказательств того, что это имеет место у млекопитающих, особенно у людей.
Довольно причудливо
Рассмотрим простой пример. Если я проведу некоторое время на солнце, у меня появится загар. По сути, это эпигенетический процесс, включающий изменения в экспрессии генов, которые увеличивают выработку меланина в моей коже, что приводит к потемнению тона кожи. Здесь существует довольно простая, прямая и непосредственная связь между экспрессией соответствующих генов и признаком цвета кожи. Этот клеточный ответ на опыт длится от недель до месяцев, но не дольше. И это не будет передано моим детям или внукам.
Загар: одна вещь, на которую действительно влияет эпигенетика. ProStockStudio/Shutterstock Существует несколько нейронных функций, в которых могут быть важны эпигенетические воздействия на небольшое количество генов, например, регуляция реакции на стресс и зависимость от наркотиков. Но психологические черты, такие как интеллект и личность, не определяются постоянным действием нескольких генов.
Во-первых, эти черты вообще не детерминированы генетически — большая часть вариаций не имеет генетического происхождения. Кроме того, генетические эффекты возникают из-за вариаций тысяч генов, и эта вариация в основном влияет на процессы развития мозга. Эти эффекты возникают не потому, что наши гены экспрессируются определенным образом прямо сейчас, а потому, что они определенным образом экспрессировались во время развития.
Это привело к тому, что наш мозг был устроен определенным образом, так что наши различные нейронные цепи имеют тенденцию работать определенным образом, что приводит к различиям в когнитивных функциях и принятии решений в различных сценариях, проявляющихся в характерных моделях поведения. Это ужасно долгий и сложный путь от генов к психологическим чертам. Таким образом, идея о том, что мы можем изменить эти черты, изменив экспрессию некоторых генов у взрослых — например, получить загар — довольно фантастична.
Использование клеточного механизма эпигенетики не делает его менее причудливым. Также нет никаких реальных доказательств того, что такие переживания, как травма, вызывают эпигенетические изменения, которые влияют на детей или внуков пострадавшего, поведенческие или каким-либо другим образом.
Однако ничто из этого не означает, что мы являемся генетически запрограммированными автоматами, поведение которых запрограммировано с рождения. У нас, безусловно, есть врожденные предрасположенности, но они обеспечивают лишь основу для нашего поведения. На самом деле мы запрограммированы учиться на собственном опыте — именно так мы приспосабливаемся к нашим конкретным обстоятельствам и так возникают наши модели поведения. Но это происходит за счет изменений в нашей нейроанатомии, а не в паттернах экспрессии генов.
И эти конструкции не зафиксированы. Изменения остаются возможными. Мы по-прежнему можем контролировать свое поведение. Мы можем работать над тем, чтобы отменить и изменить наши привычки. Мы можем до некоторой степени превзойти наши собственные подсознательные склонности. Это требует самосознания, дисциплины и усилий. Единственное, чего он не требует, так это эпигенетики.
Эпигенетические механизмы в теории депрессии, связанной с развитием нервной системы
На этой странице
РезюмеВведениеКонфликты интересовСсылкиАвторское правоСтатьи по теме
Геном (гены), эпигеном и окружающая среда работают вместе с самых ранних стадий человеческой жизни, создавая фенотип здоровья или болезни человека. Эпигенетические модификации, включая среди прочего: метилирование ДНК, модификации гистонов и структуры хроматина, а также функции некодирующих РНК, отвечают за специфические паттерны экспрессии генов. Это относится и к психическим расстройствам, в том числе к депрессивным расстройствам. Переживания раннего детства, сопровождаемые сильными стрессорами (рассматриваемыми как фактор риска депрессии во взрослой жизни), связаны с изменениями в экспрессии генов. К ним относятся гены, участвующие в реакции на стресс (гипоталамо-гипофизарно-надпочечниковая ось, HPA), связанные с гиперактивностью вегетативной нервной системы и с корковыми и подкорковыми процессами нейропластичности и нейродегенерации. К ним, среди прочего, относятся: ген, кодирующий глюкокортикоидный рецептор, ген, кодирующий белок 5, связывающий FK506 (FKBP5), ген, кодирующий альфа-рецептор аргинина, вазопрессина и эстрогена, ген переносчика 5-гидрокситриптамина (SLC6A4) и ген, кодирующий нейротрофический фактор головного мозга. Как насчет личности? Могут ли опыт, уникальный для каждого человека, история его развития и взаимодействие генов и окружающей среды посредством эпигенетических механизмов формировать черты нашей личности? Можем ли мы передать эти функции будущим поколениям? Следовательно, заложен ли риск депрессии в нашей биологической природе? Можем ли мы изменить свою судьбу?
1. Введение
Геном (гены), эпигеном (химические модификации ДНК и хроматина) и окружающая среда работают вместе с самых ранних стадий жизни человека, создавая определенный фенотип здоровья или болезни человека (рис. 1). Термин «эпигенетика» впервые был использован Уоддингтоном в конце 1930-х годов для описания явления, связанного с тем, что фенотипические изменения не всегда идут рука об руку с изменениями генотипа [1]. Уоддингтон предположил, что истоки развития исходят от взаимодействия исходных материалов в оплодотворенной яйцеклетке. Эти взаимодействия позволяют создавать что-то новое. Более того, он предположил, что это повторяющийся процесс, который приводит к образованию нового организма [1]. Однако исследования, сообщающие о важности эпигенетики в этиологии психических расстройств, появились лишь недавно [2–4].
Геном человека состоит примерно из 25 000 генов, кодирующих белок [5]. В каждой клетке экспрессируется только часть из них. Вышеупомянутые эпигенетические модификации, включая метилирование ДНК, модификации структур гистонов и хроматина, а также функции некодирующих РНК, частично ответственны за специфические паттерны экспрессии генов [6]. Каждый из трех указанных выше процессов, в отличие от генетических изменений, не связан с изменением последовательности ДНК [7]. На эти изменения в наибольшей степени влияют факторы внешней среды. Влияя на транскрипцию генов, они модифицируют наш фенотип [8].
Уникальный для каждого человека опыт, история роста, а также взаимодействия между генами и окружающей средой — посредством эпигенетических механизмов — считаются ключевым механизмом, запускающим симптомы многих соматических и психических заболеваний, включая депрессию [9] . Эти взаимодействия генов и окружающей среды вызывают эпигенетические изменения в паттернах экспрессии генов, при которых гены переключаются в состояние «включено» или «выключено», изменяя способ функционирования клеток и, таким образом, влияя на нашу предрасположенность к заболеваниям [10, 11].
Депрессия является многофакторным заболеванием. В теории нейроразвития депрессии [12] авторы подчеркивали важность ранних стадий развития для появления симптомов болезни во взрослой жизни. В данной статье основное внимание будет уделено вопросам, связанным с взаимодействием матери и ребенка, с попыткой продемонстрировать влияние их эпигенетических механизмов (главным образом, метилирования ДНК), приводящих в детстве и во взрослом возрасте к возникновению депрессивных симптомов [13].
2. Эпигенетика депрессии
Переживания в раннем детстве, связанные с сильными стрессорами (которые считаются фактором риска депрессии во взрослой жизни), связаны с модификациями экспрессии генов [14, 15]. Изменения в объеме экспрессии генов затрагивают гены, участвующие в реакции на стресс (гипоталамо-гипофизарно-надпочечниковая ось, HPA), связанные с гиперактивностью вегетативной нервной системы и корковыми и подкорковыми процессами нейропластичности и нейродегенерации [3], в том числе среди других генов, кодирующих глюкокортикоидный рецептор, FK506-связывающий белок 5 (FKBP5) [16], аргинин-вазопрессин и альфа-рецептор эстрогена, ген транспортера 5-гидрокситриптамина (SLC6A4) [17] и нейротрофические факторы головного мозга [18-20].
Story-Jovanova et al. [21] перечисляют 3 сайта метилирования, связанные с возникновением депрессии во взрослом возрасте и тяжестью ее симптомов (в исследовании приняли участие 7948 жителей Европы): cg04987734 (; ; ген CDC42BPB), cg12325605 (; ; ген ARHGEF3) и межгенный сайт CpG cg14023999 (; ; ). Все три этих сайта метилирования связаны с аксональной проводимостью. В течении депрессии у пожилых людей (>65 лет) может иметь значение сниженный уровень метилирования гена интерлейкина 6 (IL-6), который значительно повышается в результате лечения антидепрессантами [22].
Кроме того, исследовательская группа Park et al. [23] и Misra et al. [24] подтвердили, что связанные со стрессом эпигенетические изменения в следующих генах: NRC31, SLCA4, BDNF, FKBP5, SKA2, OXTR, LINGO3, POU3F1, ID3, TPPP, GRIN1 и ITGB1 коррелируют с возникновением депрессии (соответственно, Misra и др., [24]). Кроме того, было отмечено, что негативные переживания детства через эпигенетические изменения могут оказывать существенное влияние на эффективность фармакотерапии депрессии [25].
2.1. Метилирование ДНК
Метилирование включает ферментативную модификацию ДНК после репликации (связывание метильной группы с углеродом в положении 5 цитозинового кольца). Метилирование цитозина до 5-метилцитозина представляет собой пострепликационную модификацию ДНК, которая играет важную роль в подавлении транскрипции. Этот процесс включает ковалентную связь метильной группы с цитозином в составе динуклеотидов CpG [26]. Реакция метилирования ДНК катализируется ДНК-метилтрансферазами, которые переносят метильную группу с S-аденозил-L-метионина на оставшийся в ДНК цитозин. Паттерн метилирования ДНК устанавливается на ранних стадиях эмбрионального роста и поддерживается в течение индивидуальной жизни с помощью ДНК-метилтрансфераз (DNMT) [4]. В нормальных условиях метилирование ДНК используется в клетке для сайленсинга многочисленных повторяющихся последовательностей, родительского импринтинга и выключения второй Х-хромосомы в женских клетках. Метилирование остатков цитозина в ДНК играет важную роль в контроле процессов, определяющих уровень экспрессии генов в клетках в эмбриогенезе млекопитающих и в дальнейшем в процессе клеточной дифференцировки [27]. Функционирование и развитие мозга на всех этапах жизни требуют метилирования ДНК в головном мозге [28]. Следует подчеркнуть, что функция метилирования ДНК варьируется в зависимости от геномного контекста. Метилирование ДНК регуляторных областей, включающих промоторы, обычно связано с подавлением экспрессии нижестоящих генов, хотя этот эффект не является абсолютным. Напротив, метилирование ДНК тел генов связано с активной транскрипцией генов [29].].
2.2. Экспрессия miRNA (MicroRNA)
miRNAs представляют собой небольшие (состоящие примерно из 22 нуклеотидов), одноцепочечные некодирующие РНК, которые действуют как эпигенетические регуляторы благодаря своему влиянию на посттранскрипционную экспрессию генов [30]. Они присутствуют в клетках растений, животных и человека [31]. К настоящему времени идентифицировано более 2000 зрелых микроРНК, транскрибируемых в геноме человека [32]. Одна микроРНК может модулировать тысячи генов, распознавая комплементарные последовательности в конце 3UTR мРНК-мишени. Эндогенные микроРНК влияют на многочисленные процессы, происходящие в клетках, например, на пролиферацию, репарацию ДНК, клеточную дифференцировку, метаболизм и апоптоз, а также регулируют воспалительные процессы [33]. В нашей предыдущей статье мы указывали на возможное значение микроРНК-370, микроРНК-411, микроРНК-433, микроРНК-487b и микроРНК-539.для этиологии и течения депрессии [34]. Между тем, Нарахари и соавт. [33] идентифицируют miRNA-16 как посттранскрипционный репрессор транспортера серотонина (SERT).
Не так много исследований взаимосвязи между миРНК и пренатальными стрессорами; однако в некоторых работах указывается на значительное значение этого феномена для возникновения депрессивных симптомов во взрослой жизни [35].
2.3. Modification of Histone Proteins
Посттрансляционная модификация гистонов является еще одним эпигенетическим механизмом, участвующим в нормальном развитии и поддержании паттернов экспрессии генов, которые могут быть важны в этиологии депрессии [36]. Модификации гистонов влияют на организацию структуры хроматина локально и глобально, тогда как эффекты модификаций гистонов могут влиять друг на друга антагонистически или синергически, регулируя доступ хроматин-связывающих белков, что определяет переход между транскрипционно активным и неактивным хроматином [37]. Они могут приводить к активации или подавлению экспрессии генов в зависимости от типа присоединенной функциональной группы и типа модифицируемого аминокислотного остатка [38, 39].].
3. Депрессия: можно ли обмануть судьбу?
Согласно теории привязанности Боулби, эмоциональная связь между матерью и ребенком формируется на первом году жизни [40]. Однако, согласно положениям неврологической теории депрессии, этот процесс начинается значительно раньше, поскольку ключевое значение для возникновения заболевания имеют эмоциональные переживания трех периодов жизни (пренатальный период, раннее детство и подростковый возраст) [12]. Эмоциональная связь между матерью и ребенком не только становится предтечей более поздних социальных отношений, но оказывается, что ее характер влияет на формирование постоянных биологических путей и нейрогормональных реакций, которые во взрослой жизни становятся «топливом» для развития депрессивных и депрессивных состояний. тревожные расстройства или, наоборот, представляют собой специфический защитный механизм в борьбе с их возникновением [12] (рис. 2). Следовательно, если модели наших эмоциональных реакций (в том числе подверженность депрессивным расстройствам) посредством биологических механизмов формируются уже на стадии внутриутробного развития, обречены ли мы на депрессию? Можем ли мы передать склонность к депрессивным реакциям будущим поколениям? Результаты следующих исследований показывают, что это так.
Депрессия во время беременности и послеродовая депрессия матери оказывают разнонаправленное влияние на возникновение депрессивных расстройств у ребенка. Не только биологические факторы и связанные с ними нарушения отношений мать-ребенок и связи между ними (депрессивные матери менее чувствительны к потребностям развития своих детей), но и нарушения в функционировании семейной системы и, наконец, нарушения в супружеская диада важна [41]. В табл. 1 показаны другие возможные взаимосвязи между депрессией матери и возникновением депрессии у ребенка [41].
Исследования, проведенные на животных моделях (исследования на грызунах), выявили, что отделение от матери на раннем этапе жизни потомства приводит к стойким нейроэндокринным изменениям, проявляющимся во взрослой жизни в виде когнитивного, эмоционального и социального дефицита [11, 42]. Эти дефициты образуют набор симптомов, соответствующих тревожным и депрессивным расстройствам. Такое поведение связано с изменениями функции ГГАС как у матери, так и у новорожденного [42]. В исследованиях на людях, проведенных Koutra et al. [43] показано, что выраженность симптомов послеродовой депрессии у матери и степень переживаемой ею тревожности как постоянной черты ее личности связаны с качеством нервно-психического развития детей. Более того, эмоциональная близость родителей и детей в раннем детстве была фактором, существенно влияющим на объем коры в области лобных извилин у потомков и коррелирующим с личностными чертами, способствующими развитию депрессии у детей [44].
В метаанализе, проведенном Elwood et al. [45] выявлена положительная корреляция между наличием у матери симптомов послеродовой депрессии и полиморфизмов генов HMNC1, COMT, MAOT, PRKCB, ESR1, SLC6A4 и наличием жизненных событий, считающихся стрессовыми. В тех случаях, когда постнатальный период приходился на осенние и зимние месяцы, аналогичная зависимость касалась полиморфизма гена BDNF, а пережитое матерью насилие в детстве коррелировало с полиморфизмом генов OXT и OXTR. По мнению авторов [45], женщины, подверженные эпизодам послеродовой депрессии, «эпигенетически» более чувствительны к физиологическим факторам, связанным с родами. С другой стороны, Lambert и Gressier [46] подчеркивают, что повышенный уровень СРБ непосредственно перед и сразу после окончания активных родов является фактором риска послеродовой депрессии. По их мнению, эпигенетические механизмы могут приводить к патологической активности ГГАС и провоспалительному состоянию.
3.1. Эпигенетический возраст
Пренатальная материнская депрессия может вызывать эпигенетические модификации в ДНК новорожденного [47]. Суарес и др. [48] и Вольф и соавт. [49] используют новый для психиатрии термин «эпигенетический гестационный возраст» (ГВ). Под ним следует понимать эпигенетический возраст при рождении, оцениваемый по уровню метилирования в пуповинной крови плода. В настоящее время этот возраст рассчитывается на основе двух методов: предиктора Hannum et al. [50] (оценка 71 сайта CpG в цельной крови людей в возрасте 19 лет). -101 год) и так называемые часы Хорвата [51] (оценка 353 сайтов CpG в тканях органов у людей в возрасте 0-100 лет). Оба эти биомаркера молекулярного старения сильно коррелируют с хронологическим возрастом человека. Чен и др. [8] говорят, что включение оценок эпигенетического возраста улучшило способность прогнозировать смертность [8]. В исследовании, проведенном Suarez et al. [48], как симптомы депрессии во время беременности, так и послеродовая депрессия матерей коррелировали с более низким эпигенетическим возрастом потомства при рождении, что, в свою очередь, ассоциировалось с более высокой вероятностью психических расстройств в детстве у мальчиков. Между тем, Вольф и др. [49] оценили группу из 179 ветеранов войны из Ирака и Афганистана. Авторы подчеркнули, что часы Хорвата положительно коррелируют с тяжестью симптомов посттравматического стрессового расстройства, таких как избегание и эмоциональное онемение. Хан и др. [27] подчеркивают, что в группе лиц, страдающих депрессией, эпигенетический возраст значительно выше, чем в сравнительной группе лиц без психиатрического лечения в анамнезе. Более того, более высокий эпигенетический возраст был статистически связан с переживанием травм раннего детства.
FKBP5 является одним из белков, известных как иммунофилины, то есть белки, влияющие на процесс иммунного ответа клетки. У людей с депрессией, перенесших насилие в детстве, обнаружено более низкое метилирование гена, кодирующего белок FKBP5, в сочетании с редукцией серого вещества в области лобных извилин (с обеих сторон) [16].
В исследовании, проведенном Hein et al. [52] (19 женщин в возрасте 17–29,5 лет с симптомами депрессии и их матери в возрасте 36–51 год) уровень метилирования ДНК в периферических Т-лимфоцитах у обследованных матерей был связан с тяжестью депрессии у их дочерей. Гиперметилирование ДНК в группе матерей также коррелировало с «негативным воспитанием». По мнению авторов, такое негативное воспитание может быть модулятором между эпигеномом матери и депрессией потомства. «Негативное воспитание» включало материнское поведение, такое как враждебность и агрессия по отношению к ребенку, равнодушие, пренебрежение и активное неприятие ребенка (9). 0009 Анкета родительского принятия-отвержения , PARQ). Аналогичное влияние на эпигенетические изменения у потомства могут вызывать и депрессивные симптомы у отца, присутствующие после рождения ребенка, часто связанные с негативным родительским влиянием отца на потомство [53, 54] (согласно Нараянану и Нэрде [53, 54]). 55], депрессивная симптоматика у отца чаще является причиной агрессивного поведения у детей). Однако, согласно Tissot et al. [56], симптомы депрессии у матери имеют ключевое значение для заботливого поведения обоих родителей.
Кроме того, поведение матерей в раннем постнатальном периоде с высоким уровнем вовлеченности в уход за ребенком передается следующим поколениям [57]. Аналогичные отношения существуют и в случае отказа матери от своего потомства. Тот факт, что такое поведение встречается у усыновленных (и не только биологических) потомков, свидетельствует о значительной доле средовых факторов в приобретении этого поведения [58]. В исследованиях, основанных на моделях животных, отмечается, что указанные выше модели поведения связаны с изменениями в метилировании генов, кодирующих глюкокортикоидный рецептор, наблюдаемыми в области гиппокампа исследованных животных [20].
В исследовании, проведенном Stonawski et al. [19] (167 детей в возрасте 6-9 лет) диагноз депрессии во время беременности был связан со сниженным метилированием в гене, кодирующем глюкокортикоидный рецептор (NR3C1), в гене, кодирующем минералокортикоидный рецептор (NR3C2), и в гене рецептор серотонина (SLC6A4). В этом исследовании были отобраны гены, связанные с работой оси HPA.
Лайонс и др. [59] оценили взаимосвязь между качеством ранней помощи и маркерами активации симпатической нервной системы и хронического воспаления (уровень С-реактивного белка, СРБ) на выборке из 52 матерей и их детей дошкольного возраста. Матери, которые проявляли больше внимания, тепла и поддержки автономии своих детей, характеризовались более низким потенциалом покоя симпатической нервной системы и достоверно более низкими показателями хронического воспалительного процесса по уровню СРБ. Значения СРБ также были связаны с более низким потенциалом покоя симпатической нервной системы у их детей как во время релаксации, так и при контакте с матерью или женщиной, незнакомой ребенку. Таким образом, можно сделать вывод, что поддерживающее поведение матери способствовало положительному восприятию ребенком новых социальных взаимодействий и сопровождалось снижением возбудимости симпатической нервной системы.
Муг и др. [60] оценили взаимосвязь между пережитым матерями насилием в детстве и общим объемом серого вещества в мозгу их детей. Установлено, что объем серого вещества у детей матерей, ставших жертвами насилия, был достоверно ниже, чем в контрольной группе (; ). Этот эффект не был обусловлен наличием других потенциально значимых переменных, таких как социально-экономический статус матери, перинатальные осложнения, ожирение матери, насилие во время беременности, тяжелый перинатальный стресс, пол ребенка. новорожденного ребенка, возраст матери или возраст ребенка в день визуализирующего обследования. По мнению авторов, этот эффект может быть результатом эпигенетических факторов.
Интересные результаты исследований представлены Serpeloni et al. [61], с участием женщин, подвергшихся насилию со стороны своих интимных партнеров во время беременности и их детей. Уровень метилирования гена NR3C1, кодирующего глюкокортикоидный рецептор, и гена FKPB5, отвечающего за способность организма реагировать на стресс, оценивали по регуляции экспрессии гормона стресса. Показано, что жертва насилия во время беременности не только увеличивает риск развития симптомов депрессии и тревоги у женщин после родов, но и связана с изменением уровня метилирования NR3C1 и FKBP5. Аналогичные зависимости наблюдались у детей, матери которых подвергались насилию со стороны партнеров только после рождения, а не во время беременности. Эта связь свидетельствует о действии эпигенетических механизмов, обеспечивающих адаптацию новорожденных детей к неблагоприятным условиям внешней среды. Следует также отметить, что то, что называется «неблагополучием в раннем возрасте» (ELA), включая различные формы жестокого обращения с детьми, такие как физическое насилие, сексуальное насилие, психическое и эмоциональное насилие и пренебрежение, также рассматривается как фактор риска. при депрессии [2].
Также оказывается, что пренатальное воздействие СИОЗС (селективных ингибиторов обратного захвата серотонина) через эпигенетические механизмы может изменять активность гипоталамо-гипофизарно-надпочечниковой оси (ГГН) [62, 63], которая особенно активна в ответ на стрессовые стимулы (данный эффект бывает прямым и косвенным, через модуляцию секреции глюкокортикостероидов) [64]. Серотонин выполняет несколько важных функций во время эмбрионального и фетального развития мозга, включая созревание нейронов, миграцию, синаптогенез и дифференцировку клеток нервного гребня [65], в то время как переносчик серотонина (5-НТТ) был выделен в плаценте человека [66]. Таким образом, СИОЗС, проникающие через гематоэнцефалический барьер плода, посредством модификации передачи сигналов серотонина потенциально изменяют поведение в детстве, подростковом и взрослом возрасте [64]. Более того, по данным Ornoy и Koren [62], применение СИОЗС матерями в последние месяцы беременности может вызывать так называемый синдром плохой адаптации новорожденных, который включает раздражительность, повышенную плаксивость, слабое мышечное напряжение, нарушения дыхания. Однако сообщения по этому вопросу противоречивы [67]. Интересно, что Bleker et al. [68] подчеркивают положительное влияние когнитивно-поведенческой психотерапии у женщин во время беременности на уровень метилирования отдельных генов у их детей.
3.2. Окситоцин: социальный нейропептид?
Окситоцин — нейрогормон — 9-аминокислотный нейропептид, вырабатываемый гипоталамусом, активность которого тесно связана с ранее упомянутым аргинином вазопрессином [69, 70]. Наличие окситоцина и его рецепторов (OTR) наблюдается в структурах головного мозга, важных для установления и поддержания социальных отношений и развития депрессивных расстройств, таких как миндалевидное тело и гиппокамп, прилежащее ядро и дорсальное ядро блуждающего нерва. [71, 72]. Целевой зоной его действия также являются дорсальная часть поясной извилины и орбитофронтальная кора [73].
Окситоцин определяет формирование привязанности между матерью и ребенком [74]. Его уровень у матери в постнатальном периоде влияет на выраженность сепарационной тревоги, симптомы депрессии, образ себя как матери, а также формирует тревожный стиль привязанности в отношениях мать—ребенок [74, 75].
Краузе и др. [76] подчеркивают, что ранняя детская травма в виде насилия оказывает негативное влияние на нейрогормональную систему окситоцина, что проявляется в снижении экспрессии гена рецептора окситоцина (OXTR) во взрослом возрасте. У детей в очаге войны и боевых действий снижение уровня окситоцина было связано с повышением фактора иммунодефицита (оценивалась концентрация иммуноглобулина А в слюне (s-IgA)). Эти переменные были связаны с более высокой интенсивностью тревожности и более частым возникновением посттравматических стрессовых расстройств в этой группе детей [77].
Также стоит отметить, что недавние исследования на животных моделях указывают на положительные эффекты антидепрессантов, которые устраняют негативные эффекты эпигенетических механизмов [78, 79]. Однако, как мы упоминали ранее, результаты исследований, проведенных до сих пор, не ясны.
3.3. Дальнейшие направления исследований
Следует отметить, что депрессия является основной причиной инвалидности во всем мире, и более половины пациентов не достигают ремиссии симптомов после лечения антидепрессантами [80]. Все больше данных свидетельствует о том, что эпигенетические факторы (включая метилирование ДНК и модификацию гистонов) играют ключевую роль в прогнозировании ответа на антидепрессанты [80]. Важна также роль малых некодирующих РНК (микроРНК, микроРНК) и длинных некодирующих РНК (днРНК) [81] в этиологии депрессивного ответа. miRNAs могут модулировать посттранскрипционную экспрессию генов, вмешиваясь в трансляцию, контролируемую мРНК [82]. lncRNAs в высокой степени экспрессируются в головном мозге и участвуют в различных нормальных функциях мозга, а также в нейропсихиатрических расстройствах [83]. Подчеркнута роль специфических классов длинных некодирующих РНК в устойчивости или предрасположенности к развитию депрессии с реципрокным ответом на лечение антидепрессантами [83]. Эпигенетические изменения в сигнальном пути глюкокортикостероидов (например, NR3C1, FKBP5), в отношении серотонинергической нейротрансмиссии (например, SLC6A4) и в области генов, кодирующих нейротрофические факторы (например, BDNF), представляются наиболее многообещающими терапевтическими мишенями для будущие исследования [23, 84].
Конфликт интересов
Автор(ы) заявляют(ют), что у них нет конфликта интересов.
Ссылки
-
C. H. Waddington, «The Epigenotype», International Journal of Epidemiology , vol. 41, нет. 1, стр. 10–13, 2012 г.
Посмотреть по адресу:
Сайт издателя | Google Scholar
-
A. Brown, L. M. Fiori и G. Turecki, «Соединение фундаментальных и клинических исследований в области неблагоприятных условий раннего детства, метилирования ДНК и большого депрессивного расстройства», стр. 9.0009 Frontiers in Genetics , vol. 10, с. 229, 2019.
Посмотреть по адресу:
Сайт издателя | Google Scholar
-
J. Chiarella, RE Tremblay, M. Szyf, N. Provençal и L. Booij, «Влияние раннего окружения на психическое здоровье детей: уроки исследований метилирования ДНК с монозиготными близнецами», Twin Research and Генетика человека , том.
 18, нет. 6, стр. 623–634, 2015.
18, нет. 6, стр. 623–634, 2015. Посмотреть по адресу:
Сайт издателя | Академия Google
-
Ф. Кадер, М. Гай и Л. Махарадж, «Влияние метилирования ДНК на психологию человека», Behavioral Brain Research , vol. 346, стр. 47–65, 2018.
Посмотреть по адресу:
Сайт издателя | Google Scholar
-
С. Л. Зальцберг, «Открытые вопросы: сколько у нас генов?» BMC Biology , vol. 16, нет. 1, с. 94, 2018.
Посмотреть по адресу:
Сайт издателя | Google Scholar
-
Б. Грубер, «Эпигенетика и этиология нейродегенеративных заболеваний», стр. Postępy Gigieny i Medycyny Doświadczalnej , vol. 65, стр. 542–551, 2011.
Посмотреть по адресу:
Сайт издателя | Google Scholar
-
M.
Olszewska и M. Kurpisz, «Метилирование ДНК — это эпигенетическая модификация, имеющая решающее значение для гаметического импринтинга», Postepy higieny i medycyny doswiadczalnej , vol. 64, pp. 642–649, 2010. Просмотр по адресу:
Google Scholar
-
Б. Х. Чен, Р. Э. Мариони, Э. Колицино и др., «Измерения биологического возраста на основе метилирования ДНК: прогнозирование метаанализа время до смерти» Старение , об. 8, нет. 9, стр. 1844–1865, 2016.
Посмотреть по адресу:
Сайт издателя | Google Scholar
-
Э. Страчан, Дж. Чжао, П. П. Рой-Бирн, Э. Фаулер и Т. Бакус, «Дизайн исследования и обоснование исследования настроения и метилирования: платформа для мультиомного исследования депрессии у близнецы», Twin Research and Human Genetics , vol. 21, нет. 6, стр. 507–513, 2018.
Посмотреть по адресу:
Сайт издателя | Академия Google
-
K.
Olden, N. Freudenberg, J. Dowd и A. E. Shields, «Открытие того, как воздействие окружающей среды изменяет гены, может привести к новым методам лечения хронических заболеваний», Health Affairs , vol. 30, нет. 5, стр. 833–841, 2011. Посмотреть по адресу:
Сайт издателя | Google Scholar
-
А. М. Вайзерман и А. К. Колиада, «Невзгоды в раннем возрасте и долгосрочные нейроповеденческие последствия: эпигеном как мост?» Геномика человека , том. 11, нет. 1, с. 34, 2017.
Посмотреть по адресу:
Сайт издателя | Google Scholar
-
П. Галецки и М. Таларовска, «Нейроразвитие теории депрессии», Progress in Neuro-Psychopharmacology & Biological Psychiatry , vol. 80, стр. 267–272, 2018.
Посмотреть по адресу:
Сайт издателя | Google Scholar
-
Дж.
Хилл, А. Пиклз, Н. Райт, Э. Брейтуэйт и Х. Шарп, «Предсказания детской эмоциональности на основе эволюционных и эпигенетических гипотез», Научные отчеты , том. 9, нет. 1, с. 2519, 2019. Посмотреть по адресу:
Сайт издателя | Google Scholar
-
И. Ярич, Д. Рокс, Х. Чам, А. Херчек и М. Кундакович, «Влияние полового и эстрального циклов на фенотипы, связанные с тревогой и депрессией, в модели стресса развития с двумя ударами, Frontiers in Molecular Neuroscience , vol. 12, с. 74, 2019.
Посмотреть по адресу:
Сайт издателя | Google Scholar
-
A.C. Viuff, G.C. Sharp, D. Rai et al., «Материнская депрессия во время беременности и метилирование ДНК пуповинной крови: результаты исследования Avon Longitudinal Study of Parents and Children», стр. Трансляционная психиатрия , том. 8, нет.
1, с. 244, 2018. Посмотреть по адресу:
Сайт издателя | Google Scholar
-
L. Tozzi, C. Farrell, L. Booij et al., «Эпигенетические изменения FKBP5 как связующее звено, связывающее генетические и экологические факторы риска со структурными и функциональными изменениями мозга при большой депрессии», Neuropsychopharmacology , об. 43, нет. 5, стр. 1138–1145, 2018.
Посмотреть по адресу:
Сайт издателя | Академия Google
-
L. Provenzi, R. Giorda, S. Beri и R. Montirosso, «Метилирование SLC6A4 как эпигенетический маркер воздействия жизненных невзгод на людей: систематический обзор литературы», Neuroscience and Biobehavioral Reviews , vol. 71, стр. 7–20, 2016 г.
Посмотреть по адресу:
Сайт издателя | Google Scholar
-
M.
S. Mendonça, P. M. Mangiavacchi, P. F. de Sousa et al., «Эпигенетическая изменчивость промотора гена SLC6A4 в парах мать-ребенок с большим депрессивным расстройством», Журнал аффективных расстройств , том. 245, стр. 716–723, 2019. Посмотреть по адресу:
Сайт издателя | Google Scholar
-
В. Стонавски, С. Фрей, Ю. Голуб и др., «Ассоциации пренатальных депрессивных симптомов с метилированием ДНК генов, связанных с осью HPA, и суточными профилями кортизола у детей младшего школьного возраста», Развитие и Психопатология , том. 31, нет. 2, стр. 419–431, 2019.
Посмотреть по адресу:
Сайт издателя | Академия Google
-
Р. Вентура-Хунка и Л. М. Эррера, «Эпигенетические изменения, связанные со стрессовыми событиями в раннем возрасте», Acta Neuropsychiatrica , vol. 24, нет. 5, стр.
255–265, 2012 г. Посмотреть по адресу:
Сайт издателя | Google Scholar
-
О. С. Йованова, И. Неделькович, Д. Шпилер и др., «Сигнатуры метилирования ДНК симптомов депрессии у людей среднего и пожилого возраста: метаанализ мультиэтнических эпигеномных исследований», JAMA Psychiatry , том. 75, нет. 9, стр. 949–959, 2018.
Посмотреть по адресу:
Сайт издателя | Google Scholar
-
J. Ryan, L. Pilkington, K. Neuhaus, K. Ritchie, M.L. Ancelin и R. Saffery, «Изучение эпигенетического профиля воспалительного гена IL-6 при депрессии в пожилом возрасте», BMC Психиатрия , том. 17, нет. 1, 2017.
Посмотреть по адресу:
Сайт издателя | Google Scholar
-
C. Park, JD Rosenblat, E. Brietzke et al., «Стресс, эпигенетика и депрессия: систематический обзор», стр.
9.0009 Neuroscience and Biobehavioral Reviews , vol. 102, стр. 139–152, 2019. Посмотреть по адресу:
Сайт издателя | Google Scholar
-
P. Misra, S. Liu и X. Meng, «Какие модификации метилирования ДНК и/или генетические вариации взаимодействуют с жестоким обращением в детстве при развитии депрессии: систематический обзор», Journal of Affective Disorders , том. 252, стр. 325–333, 2019.
Посмотреть по адресу:
Сайт издателя | Академия Google
-
P. Jaworska-Andryszewska и JK Rybakowski, «Детская травма при расстройствах настроения: нейробиологические механизмы и значение для лечения», Pharmacological Reports , vol. 71, нет. 1, стр. 112–120, 2019.
Посмотреть по адресу:
Сайт издателя | Google Scholar
-
Т.
С. Доэрти и Т. Л. Рот, «Эпигенетические ландшафты мозга, подверженного неблагоприятным воздействиям», Progress in Molecular Biology and Translational Science , vol. 157, стр. 1–19., 2018. Посмотреть по адресу:
Сайт издателя | Google Scholar
-
Л. К. М. Хан, М. Агаджани, С. Л. Кларк и др., «Эпигенетическое старение при большом депрессивном расстройстве», Американский журнал психиатрии , том. 175, нет. 8, стр. 774–782, 2018 г.
Посмотреть по адресу:
Сайт издателя | Google Scholar
-
Кюнер Дж. Н., Брюггеман Э. К., Вен З. и Яо Б., «Эпигенетические регуляции нейропсихиатрических расстройств», Frontiers in Genetics , том. 10, 2019.
Посмотреть по адресу:
Сайт издателя | Google Scholar
-
Т.
Е. Келлер, П. Хан и С. В. Йи, «Эволюционный переход метилирования ДНК промотора и тела гена через границу между беспозвоночными и позвоночными», Molecular Biology and Evolution , vol. 33, нет. 4, стр. 1019–1028, 2016. Посмотреть по адресу:
Сайт издателя | Google Scholar
-
J. Tavakolizadeh, K. Roshanaei, A. Salmaninejad et al., «МикроРНК и экзосомы при депрессии: потенциальные диагностические биомаркеры», Журнал клеточной биохимии , том. 119, нет. 5, стр. 3783–3797, 2018.
Посмотреть по адресу:
Сайт издателя | Google Scholar
-
К. Сааведра, А. Молина-Маркес, Н. Сааведра, Т. Замбрано и Л. Салазар, «Эпигенетические модификации большого депрессивного расстройства», International Journal of Molecular Sciences , vol. 17, нет. 8, с. 1279, 2016.
Посмотреть по адресу:
Сайт издателя | Google Scholar
-
«miRBase 22.
0: база данных микроРНК», декабрь 2018 г. http://www.mirbase.org/index.shtml. Просмотр по адресу:
Google Scholar
-
А. Нарахари, М. Хуссейн и В. Шрирам, «МикроРНК как биомаркеры психических состояний: обзор текущих исследований», Innovations in Clinical Neuroscience , vol. 14, нет. 1–2, pp. 53–55, 2017.
Посмотреть по адресу:
Google Scholar
-
значение микроРНК в ходе rDD», Фармакологические отчеты , том. 69, нет. 2, стр. 206–212, 2017 г.
Посмотреть по адресу:
Сайт издателя | Google Scholar
-
Л. Стенц, Д. С. Шехтер, С. Р. Серпа и А. Паолони-Джакобино, «Передача между поколениями признаков метилирования ДНК, связанных со стрессом в раннем возрасте», Current Genomics , vol. 19, нет.
8, стр. 665–675, 2018. Посмотреть по адресу:
Сайт издателя | Google Scholar
-
JM Deussing and M. Jakovcevski, «Модификации гистонов в большом депрессивном расстройстве и родственных моделях грызунов», Достижения экспериментальной медицины и биологии , vol. 978, стр. 169–183, 2017.
Посмотреть по адресу:
Сайт издателя | Google Scholar
-
Э. Пишва, Б. П. Ф. Руттен и Д. ван ден Хоув, «Метилирование ДНК при большом депрессивном расстройстве», Успехи экспериментальной медицины и биологии , том. 978, стр. 185–196, 2017.
Посмотреть по адресу:
Сайт издателя | Google Scholar
-
Чистяков Д. А., Бобрышев Ю. В., Чехонин В. П. Эпигенетические изменения в ДНК и модификации гистонов, вызванные депрессией и антидепрессантами: уроки на моделях грызунов, Текущий фармацевтический дизайн , том.
23, нет. 44, стр. 6828–6840, 2017. Посмотреть по адресу:
Сайт издателя | Google Scholar
-
P. Misztak, P. Pańczyszyn-Trzewik и M. Sowa-Kućma, «Деацетилазы гистонов (HDAC) как терапевтическая мишень при депрессивных расстройствах», Pharmacological Reports , vol. 70, нет. 2, стр. 398–408, 2018 г.
Посмотреть по адресу:
Сайт издателя | Google Scholar
-
Дж. Чемберс, «Нейробиология привязанности: от младенчества до клинических результатов», Психодинамическая психиатрия , том. 45, нет. 4, стр. 542–563, 2017.
Посмотреть по адресу:
Сайт издателя | Google Scholar
-
Т. Л. Тайра, «Депрессивные родители и их дети», Przegląd Psychologiczny , vol. 43, pp. 323–337, 2000.
Посмотреть по адресу:
Google Scholar
-
М.
Ниши, Н. Хории-Хаяши и Т. Сасагава, «Влияние неблагоприятных событий в раннем возрасте на мозг: последствия из моделей материнского отделения у грызунов» Frontiers in Neuroscience , vol. 8, 2014. Посмотреть по адресу:
Сайт издателя | Google Scholar
-
К. Коутра, Т. Румелиотаки, А. Кириклаки и др., «Материнская депрессия и личностные черты в связи с нейропсихологическим и поведенческим развитием ребенка в дошкольном возрасте: когорта матери и ребенка (исследование Rhea) на Крите, Греция», Журнал аффективных расстройств , том. 217, стр. 89–98, 2017.
Посмотреть по адресу:
Сайт издателя | Академия Google
-
Дж. Ян, П. Инь, Д. Вей, К. Ван, Ю. Ли и Дж. Цю, «Влияние родительской эмоциональной теплоты на взаимосвязь между региональным объемом серого вещества и личностными чертами, связанными с депрессией, Социальная нейронаука , том.
12, нет. 3, стр. 337–348, 2017. Посмотреть по адресу:
Сайт издателя | Google Scholar
-
Дж. Элвуд, Э. Мюррей, А. Белл, М. Синклер, У. Г. Кернохан и Дж. Стокдейл, «Систематический обзор, посвященный изучению того, связаны ли генетические или эпигенетические маркеры с послеродовой депрессией», стр. 9.0009 Журнал аффективных расстройств , том. 253, стр. 51–62, 2019.
Посмотреть по адресу:
Сайт издателя | Google Scholar
-
М. Ламберт и Ф. Грессье, «Biomarqueurs de L’inflammation et Dépression du Post-Partum. Une Revue Systématique De la Littérature», Canadian Journal of Psychiatry , vol. 26, 2019.
Посмотреть по адресу:
Сайт издателя | Google Scholar
-
З. Немода и М. Шиф, «Эпигенетические изменения и пренатальная материнская депрессия», Исследование врожденных дефектов , vol.
109, нет. 12, стр. 888–897, 2017. Посмотреть по адресу:
Сайт издателя | Google Scholar
-
А. Суарес, Дж. Лахти, Д. Чамара и др., «Эпигенетические часы при рождении: связь с материнской антенатальной депрессией и детскими психическими проблемами», Журнал Американской академии детской и подростковой психиатрии , том. 57, нет. 5, стр. 321–328.e2, 2018.
Посмотреть по адресу:
Сайт издателя | Академия Google
-
Э. Дж. Вольф, М. В. Лог, Ф. Г. Моррисон и др., «Посттравматическая психопатология и ритм эпигенетических часов: продольное исследование», Psychological Medicine , vol. 49, нет. 5, стр. 791–800, 2019.
Посмотреть по адресу:
Сайт издателя | Google Scholar
-
G. Hannum, J. Guinney, L. Zhao et al.
, «Полногеномные профили метилирования раскрывают количественные представления о темпах старения человека», Molecular Cell , vol. 49, нет. 2, стр. 359–367, 2013. Посмотреть по адресу:
Сайт издателя | Google Scholar
-
С. Хорват, «Возраст метилирования ДНК тканей и типов клеток человека», Genome Biology , vol. 14, нет. 10, с. R115, 2013.
Посмотреть по адресу:
Сайт издателя | Google Scholar
-
С. Хайн, Т. Томас, О. Ю. Наумова, С. С. Лутар и Е. Л. Григоренко, «Негативное воспитание модулирует связь между профилями метилирования ДНК матери и депрессией у взрослых потомков», Психобиология развития , том. 61, нет. 2, стр. 304–310, 2018.
Посмотреть по адресу:
Сайт издателя | Google Scholar
-
С.
Нат, Г. Рассел, В. Кайкен, Л. Психогиу и Т. Форд, «Опосредует ли конфликт между отцом и ребенком связь между послеродовыми депрессивными симптомами отцов и проблемами адаптации детей в возрасте 7 лет?» ?» Психологическая медицина , том. 46, нет. 8, стр. 1719–1733, 2016. Посмотреть по адресу:
Сайт издателя | Академия Google
-
S. Yeshurun, J. Rogers, A.K. Short, T. Renoir, T.Y. Pang и A.J. Hannan, «Повышенное воздействие глюкокортикоидов на отца изменяет сохранение памяти у потомства женского пола», Psychoneuroendocrinology , vol. 83, стр. 9–18, 2017 г.
Посмотреть по адресу:
Сайт издателя | Google Scholar
-
М. К. Нараянан и А. Нэрде, «Связи между материнскими и отцовскими депрессивными симптомами и проблемами поведения детей в раннем возрасте: тестирование взаимно скорректированной проспективной продольной модели», Журнал аффективных расстройств , том.
196, стр. 181–189, 2016. Посмотреть по адресу:
Сайт издателя | Google Scholar
-
H. Tissot, N. Favez, P. Ghisletta, F. Frascarolo и JN Despland, «Продольное исследование симптомов депрессии у родителей и совместного воспитания в первые 18 месяцев», Family Process , vol. 56, нет. 2, стр. 445–458, 2017.
Посмотреть по адресу:
Сайт издателя | Google Scholar
-
И. К. Г. Уивер, «Эпигенетические эффекты глюкокортикоидов», Семинары по фетальной и неонатальной медицине , vol. 14, нет. 3, стр. 143–150, 2009 г.
Посмотреть по адресу:
Сайт издателя | Google Scholar
-
Р. К. Багот, Ф. Н. ван Хасселт, Д. Л. Шампань, М. Дж. Мини, Х. Дж. Крюгерс и М. Джоэлс, «Материнская забота определяет быстрое воздействие медиаторов стресса на синаптическую пластичность зубчатой извилины гиппокампа взрослых крыс», Нейробиология Обучение и память , том.
92, нет. 3, стр. 292–300, 2009. Просмотр:
Сайт издателя | Google Scholar
-
E. R. Lyons, J. Norman Wells, C. M. Scholtes, B. Mintz, R. J. Giuliano и E. A. Skowron, «Воспоминания о положительном раннем уходе связаны с активацией симпатической нервной системы и хроническим воспалением в последующих поколениях», Developmental Психобиология , том. 61, нет. 2, стр. 261–274, 2019.
Посмотреть по адресу:
Сайт издателя | Google Scholar
-
Н. К. Муг, С. Энтринджер, Дж. М. Расмуссен и др., «Влияние между поколениями материнского жестокого обращения в детстве на анатомию мозга новорожденного», Биологическая психиатрия , том. 83, нет. 2, стр. 120–127, 2018 г.
Посмотреть по адресу:
Сайт издателя | Google Scholar
-
Ф.
Серпелони, К. М. Радтке, Т. Хекер и др., «Формирует ли пренатальный стресс постнатальную устойчивость? — эпигеномное исследование насилия и психического здоровья у людей», Frontiers in Genetics , vol. 10, 2019. Посмотреть по адресу:
Сайт издателя | Google Scholar
-
А. Орной и Г. Корен, «СИОЗС и СИОЗСН (СИОЗС) при беременности: влияние на течение беременности и потомство: насколько мы далеки от ответов на все вопросы?» Международный журнал молекулярных наук , том. 20, нет. 10, с. 2370, 2019.
Посмотреть по адресу:
Сайт издателя | Google Scholar
-
А. Орной и Г. Корен, «Селективные ингибиторы обратного захвата серотонина при беременности человека: на пути к разрешению разногласий», семинаров по фетальной и неонатальной медицине , том. 19, нет. 3, стр. 188–194, 2014.
Посмотреть по адресу:
Сайт издателя | Google Scholar
-
С.
Браммельте, Э. М. Гланаги, А. Боннин и Т. Ф. Оберландер, «Развитие изменений в передаче сигналов серотонина: последствия для ранней функции мозга, поведения и адаптации», Неврология , том. 342, стр. 212–231, 2017. Посмотреть по адресу:
Сайт издателя | Google Scholar
-
Т. Ф. Оберландер, «Передача сигналов серотонина плода: установление путей развития и поведения в раннем детстве», The Journal of Adolescent Health , vol. 51, стр. 9–16, 2012.
Просмотр по адресу:
Google Scholar
-
С. Рамамурти, Ф. Х. Лейбах, В. Б. Махеш и В. Ганапати, «Частичная очистка и характеристика плацентарного переносчика серотонина человека, Плацента , об. 14, нет. 4, стр. 449–461, 1993.
Посмотреть по адресу:
Сайт издателя | Google Scholar
-
A.
Cardenas, S. Faleschini, A.C. Hidalgo et al., «Пренатальные материнские антидепрессанты, тревога и депрессия и метилирование ДНК потомства: эпигеномные ассоциации при рождении и сохранение в раннем детстве», Clinical Epigenetics , том. 11, нет. 1, с. 56, 2019. Посмотреть по адресу:
Сайт издателя | Академия Google
-
Л. С. Блекер, Дж. Милгром, А. Секстон-Оутс и др., «Изучение влияния дородового лечения депрессии на эпигенетические профили детей: результаты экспериментального рандомизированного контролируемого исследования», Clinical Epigenetics , vol. 11, нет. 1, с. 18, 2019.
Посмотреть по адресу:
Сайт издателя | Google Scholar
-
М. М. Перес-Родригес, К. Махон, М. Руссо, А. К. Унгар и К. Э. Бердик, «Окситоцин и социальное познание при аффективных и психотических расстройствах», стр.
9.0009 Европейская нейропсихофармакология , том. 25, нет. 2, стр. 265–282, 2015 г. Посмотреть по адресу:
Сайт издателя | Google Scholar
-
Л. М. Сиппель, К. Э. Аллингтон, Р. Х. Петрзак, И. Харпаз-Ротем, Л. К. Мэйс и М. Олф, «Окситоцин и связанные со стрессом расстройства: нейробиологические механизмы и возможности лечения», Chronic Stress , vol. . 1, 2017.
Посмотреть по адресу:
Сайт издателя | Google Scholar
-
М. Киммел, М. Клайв, Ф. Гиспен и др., «Метилирование ДНК рецептора окситоцина при послеродовой депрессии», стр. 9.0009 Психонейроэндокринология , том. 69, стр. 150–160, 2016.
Посмотреть по адресу:
Сайт издателя | Google Scholar
-
A. Meyer-Lindenberg, G. Domes, P. Kirsch и M. Heinrichs, «Окситоцин и вазопрессин в человеческом мозгу: социальные нейропептиды для трансляционной медицины», Nature Reviews Neuroscience , vol.
12, нет. 9, стр. 524–538, 2011. Посмотреть по адресу:
Сайт издателя | Google Scholar
-
Т. Т. Поль, Л. Дж. Янг и О. Дж. Бош, «Потерянные связи: окситоцин и нервные, физиологические и поведенческие последствия нарушенных отношений», стр. 9.0009 Международный журнал психофизиологии , том. 136, стр. 54–63, 2019.
Посмотреть по адресу:
Сайт издателя | Google Scholar
-
Б. Л. Мах, М. Дж. Бейкерманс-Краненбург, М. Х. ван Эйзендорн и Р. Смит, «Окситоцин способствует защитному поведению у депрессивных матерей: экспериментальное исследование с восторженной парадигмой незнакомца», , Депрессия и тревога, , том. 32, нет. 2, стр. 76–81, 2015 г.
Посмотреть по адресу:
Сайт издателя | Академия Google
-
Дж.
Колхофф, В. Ипен, М. Даддс, Ф. Хан, Д. Силов и Б. Барнетт, «Окситоцин в постнатальный период: ассоциации с привязанностью и материнской заботой», Комплексная психиатрия , том . 76, стр. 56–68, 2017. Посмотреть по адресу:
Сайт издателя | Google Scholar
-
S. Krause, C. Boeck, AM Gumpp et al., «Жестокое обращение с детьми связано со снижением рецептора окситоцина в мононуклеарных клетках периферической крови», Границы психологии , том. 9, 2018.
Посмотреть по адресу:
Сайт издателя | Google Scholar
-
А. Ульмер-Янив, А. Джаловски, К. Йирмия, Г. Халеви, О. Загури-Шарон и Р. Фельдман, «Материнские иммунные и аффилиативные биомаркеры и чувствительное воспитание опосредуют эффекты хронического раннего травма детской тревожности», Psychological Medicine , vol. 48, нет. 6, стр. 1020–1033, 2018.
Посмотреть по адресу:
Сайт издателя | Академия Google
-
С. В. Парк, М. К. Сео, Дж. Г. Ли, Л. Т. Хиен и Ю. Х. Ким, «Влияние разлуки с матерью и приема антидепрессантов на эпигенетическую регуляцию промотора экзона I мозгового нейротрофического фактора в гиппокампе взрослой крысы», Психиатрия. и клиническая нейронаука , том. 72, нет. 4, стр. 255–265, 2018 г.
Посмотреть по адресу:
Сайт издателя | Google Scholar
-
A. J. Sales and S. R. L. Joca, «Введение антидепрессантов модулирует индуцированное стрессом метилирование ДНК и экспрессию ДНК-метилтрансфераз в префронтальной коре и гиппокампе крыс», Поведенческие исследования мозга , том. 343, стр. 8–15, 2018 г.
Посмотреть по адресу:
Сайт издателя | Google Scholar
-
Л.
М. Хак, Г. Р. Фрайс, Х. А. Эйр и др., «Перемещение инструментов фармакоэпигенетики депрессии к клиническому использованию», Journal of Affective Disorders , vol. 249, стр. 336–346, 2019. Посмотреть по адресу:
Сайт издателя | Google Scholar
-
Q. Wang, B. Roy и Y. Dwivedi, «Моделирование сети коэкспрессии идентифицирует ключевые длинные некодирующие модули РНК и мРНК в изменении молекулярного фенотипа для развития вызванной стрессом депрессии у крыс», Трансляционная психиатрия , том. 9, нет. 1, 2019.
Посмотреть по адресу:
Сайт издателя | Google Scholar
-
Г. Р. Фрайс, В. Чжан, Д. Беневенуто и Дж. Кеведо, «МикроРНК при большом депрессивном расстройстве», Достижения в области экспериментальной медицины и биологии , том. 1118, стр. 175–190, 2019.
Посмотреть по адресу:
Сайт издателя | Google Scholar
-
B.
Roy, Q. Wang и Y. Dwivedi, «Длинные некодирующие РНК-ассоциированные транскриптомные изменения устойчивости или восприимчивости к депрессии и реакции на лечение антидепрессантами», Международный журнал нейропсихофармакологии , том. 21, нет. 5, стр. 461–472, 2018. Посмотреть по адресу:
Сайт издателя | Google Scholar
-
A.W.M. Spithoven, S. Cacioppo, L. Goossens, and JT Cacioppo, «Генетические вклады в одиночество и их отношение к эволюционной теории одиночества», Perspectives on Psychological Science , vol. 14, нет. 3, стр. 376–396, 2019.
Посмотреть по адресу:
Сайт издателя | Академия Google
Copyright
Copyright © 2020 Моника Таларовска. Это статья с открытым доступом, распространяемая в соответствии с лицензией Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии надлежащего цитирования оригинальной работы.
Ранний жизненный опыт, эпигенетика и развивающийся мозг
Аннотация
Развитие — это динамический процесс, который включает взаимодействие между генами и окружающей средой. У млекопитающих качество постнатальной среды формируется взаимодействиями родителей и потомства, которые способствуют росту и выживанию и могут приводить к расходящимся траекториям развития с последствиями для нейробиологических и поведенческих характеристик в более позднем возрасте. Появляющиеся данные свидетельствуют о том, что эпигенетические факторы (например, метилирование ДНК, посттрансляционные модификации гистонов и малые некодирующие РНК) могут играть решающую роль в этих эффектах родительской заботы. Хотя эти данные получены в основном из исследований на грызунах, все больше подтверждаются данные об этих эффектах у людей. Благодаря этим молекулярным механизмам могут возникать различия в риске психопатологии, особенно в результате пренебрежения и жестокого обращения в раннем возрасте. Здесь мы выделим доказательства динамических эпигенетических изменений в развивающемся мозге в ответ на изменение качества постнатальных взаимодействий родитель-потомок. Задействование эпигенетических путей для биологического закрепления раннего жизненного опыта может также иметь трансгенерационные последствия, и мы опишем и сопоставим два пути, по которым может происходить эта передача: зависимый от опыта 9Наследование зародышевой линии 0009 против . Наконец, мы порассуждаем о будущих направлениях эпигенетических исследований и о том, как они могут помочь нам лучше понять генетическое происхождение психической дисфункции.
ВВЕДЕНИЕ
Развитие обычно происходит в социальном контексте, включающем родителей, братьев и сестер и других опекунов или членов группы. Этот контекст обеспечивает потребности развития потомства, такие как кормление, сенсорная стимуляция и эмоциональное тепло, но также может служить важным сигналом к типу среды, в которой будет происходить непосредственное и более позднее развитие. Например, хотя основные потребности могут быть удовлетворены усилиями одного человека, осуществляющего уход, отсутствие других лиц (например, друзей и родственников) может указывать на отсутствие социальной поддержки или финансовые ограничения, которые могут быть типичными условиями, в которых приходится сталкиваться на протяжении всей жизни. срок жизни. Учитывая потенциальную ценность этих сигналов окружающей среды для прогнозирования как текущего, так и будущего социально-экономического контекста, возможно, неудивительно, что качество окружающей среды в раннем возрасте может оказывать устойчивое влияние на нашу биологию, приводя к расходящимся траекториям развития. В частности, качество взаимодействия родителей и потомков может формировать развивающийся мозг с долгосрочными последствиями для его функций и поведения. Важнейший вопрос, поднятый десятилетиями исследований влияния воспитания на развитие, касается механизма: как опыт разнообразия родительской заботы приводит к пожизненным изменениям в нейробиологии и поведении?
В последнее десятилетие в свете современных достижений в области молекулярной биологии и визуализации мозга, которые указывают на пути, которые могут быть общими для многих переживаний раннего периода жизни, поиск механизмов, лежащих в основе влияния родителей на потомство, быстро продвигался вперед. т. е. воздействие токсинов, изменение питания и стресс). В частности, очевидно, что качество взаимодействия родитель-потомок вызывает эпигенетические изменения в развивающемся мозге, которые объясняют вариации в ответ на стресс, познание, социальность и репродуктивное поведение. Эпигенетика — это развивающаяся область исследований, которая фокусируется не на последовательности ДНК в наших геномах, а скорее на регуляции «активности» генов. Эти эпигенетические механизмы могут помочь нам лучше понять биологическое воздействие широкого спектра воздействий окружающей среды, включая качество родительской заботы. Более того, ясно, что влияние раннего жизненного опыта может повлиять на последующие поколения потомства. В этом обзоре мы выделим доказательства нейробиологического и поведенческого влияния взаимодействий родителей и потомства, происходящих в постнатальный период, эпигенетические механизмы, с помощью которых эти эффекты могут быть достигнуты, и исследуем трансгенерационные последствия родительской заботы и невзгод в раннем возрасте.
ПРОДОЛЖИТЕЛЬНОЕ ВОЗДЕЙСТВИЕ РОДИТЕЛЬСКОЙ ЗАБОТЫ
Контекст развития может значительно различаться у разных людей. Хотя источники этой изменчивости могут включать в себя широкие экологические факторы, такие как климат, плотность населения и социально-экономический статус, с точки зрения развивающегося человека эти контекстуальные факторы ощущаются косвенно через влияние окружающей среды на взаимодействие родителей и потомков. Зависимость потомства от этих взаимодействий может выходить за рамки выживания. Социально-эмоциональные системы, управляющие саморегуляцией и реакцией на стресс, могут развиваться в процессе ожидания опыта, который зависит от видоспецифичных форм социальных взаимодействий в раннем возрасте. Этот процесс развития продемонстрирован в исследованиях воздействия изменчивости или нарушения качества взаимодействия родитель-потомок у людей, приматов и грызунов.
Родительская забота, пренебрежение и жестокое обращение с людьми
Исследования влияния поведения родителей на развитие человека были сосредоточены на качестве привязанности, чувствительности к сигналам младенца, родительской привязанности и нарушениях отношений между родителями и детьми из-за отсутствия родителей. (т. е. институциональное воспитание), пренебрежение и жестокий уход. Изменения в отношениях привязанности (обычно наблюдаемые между матерью и младенцем) связаны либо с устойчивостью к психологическому дистрессу (например, надежная привязанность), либо с повышенной частотой психопатологий (например, дезорганизованная привязанность) (Sroufe, 2005). Дезорганизованная привязанность является предиктором повышенного уровня расстройств личности, диссоциации, членовредительства и повышения уровня кортизола в слюне во время воздействия стрессоров, что предполагает повышенную реактивность гипоталамо-гипофизарно-надпочечниковой (ГГН) реакции на стресс (Bernard and Dozier, 2010; Карлсон, 19 лет98; Карлсон и др., 2009 г.; ван Айзендорн и др., 1999). Среди младенцев, чьи матери характеризуются низкой ( против ) высокой частотой чувствительного/ненавязчивого ухода, отмечаются повышенные реакции страха и сниженная лобная электроэнцефалографическая асимметрия (предиктор повышенной реактивности ГГН; см. рис. 1а) в возрасте 9 месяцев. (Hane and Fox, 2006), чем сохраняются в возрасте 2–3 лет (см. рисунок 1b; Hane et al, 2010). Родительская привязанность, которая включает в себя показатели степени заботы и чрезмерной защиты, является важным предиктором депрессии в более позднем возрасте (связанной с низкой заботой и высокой степенью чрезмерной защиты; Паркер, 19 лет).90) и может вызывать изменения объема серого вещества в префронтальной коре и гиппокампе, а также изменение дофаминергической и кортизоловой реакции на стресс (Engert et al, 2010; Narita et al, 2010, 2012; Pruessner et al, 2004). Таким образом, даже отклонения в нормальном диапазоне родительского поведения могут иметь значительное влияние на нейроповеденческие результаты.
Влияние низкого по сравнению с высокого качества материнской помощи у людей на (а) исходы в младенчестве и (б) в детстве. Адаптировано из Hane and Fox (2006) и Hane et al (2010).
Слайд PowerPoint
Изображение в полный размер
Было обнаружено, что неблагоприятный детский опыт, который включает пренебрежение и жестокое обращение в детстве, предсказывает снижение когнитивных способностей, нарушение социального развития и рост расстройств личности (Johnson et al, 1999; Trickett and McBride-Chang, 1995) . Эти результаты могут быть частично обусловлены нарушением регуляции HPA, которое характерно для людей, испытавших пренебрежение и/или жестокое обращение (Carpenter et al, 2011; Heim et al, 2010). В исследованиях визуализации головного мозга эмоциональное жестокое обращение в детстве связано с меньшим размером медиальной префронтальной коры (van Harmelen et al, 2010), а изменения объема и активации в ответ на сигналы угрозы были обнаружены в нескольких областях мозга, включая миндалевидное тело и гиппокамп (De Bellis, 2005; Maheu и др., 2010). Серьезное пренебрежение, с которым сталкиваются младенцы, находящиеся в специализированных учреждениях (например, румынские сироты), приводит к задержке развития, нарушению исполнительных функций, усилению аутистического поведения и повышенному уровню кортизола в слюне (Gunnar et al, 2001; Hostinar et al, 2012; O’Connor et al. и др., 2000; Раттер и О’Коннор, 2004). Подобно младенцам, не находящимся в интернатных учреждениях, которые в прошлом подвергались пренебрежению/насилию, у детей, воспитанных в детских домах, повышена активность миндалевидного тела (Tottenham et al, 2011). Повышенная мозговая активность, особенно в миндалевидном теле и гиппокампе, в ответ на угрожающие сигналы выступает как общий нейробиологический маркер нарушения отношений между родителями и ребенком, что приводит к нарушению социальных взаимодействий (Maheu et al, 2010).
Хотя нейробиологические последствия низкой родительской заботы, жестокого обращения и пренебрежения часто концептуализируются как отдельные формы раннего жизненного опыта, существует значительная взаимосвязь между этими формами невзгод. Дезорганизованная привязанность наблюдается у 48–80% младенцев, подвергшихся жестокому обращению/насилию (Johnson et al, 1999). Институциональное воспитание связано с дезорганизованными отношениями привязанности, и вмешательства, направленные на создание большей стабильности во взаимоотношениях опекуна и младенца, могут усилить привязанность и улучшить долгосрочные результаты (Smyke et al, 2002). Более того, качество отношений привязанности может быть медиатором долгосрочных эффектов социальной депривации в раннем возрасте на симптомы психопатологии (McGoron et al, 2012). Надежность привязанности определяется качеством родительской заботы, включая чувствительность и синхронность взаимодействий (De Wolff and van Ijzendoorn, 19).97). Таким образом, этот ранний жизненный опыт может быть коллективно частью каскада развития, который способствует уязвимости к нейробиологическим и поведенческим нарушениям.
Влияние родительского поведения на приматов
Исследования на нечеловеческих приматах показывают, что степень контакта между матерью и младенцем и показатели отвержения/жестокого обращения могут иметь длительные поведенческие и нейробиологические последствия. Материнская депривация, достигаемая за счет питомникового выращивания, оказывает глубокое влияние на развитие макак-резусов, что приводит к нарушениям социального поведения, гиперактивности и чувствительности к стрессорам (Harlow et al, 19).65; Суоми и др., 1971). Среди выращенных матерью макак против макак, выращенных сверстниками (Suomi, 1991), у которых младенцы имеют социальный контакт со сверстниками, но не с матерью, у выращенных сверстниками младенцев обнаруживаются долгосрочные нарушения поведения, нейроэндокринной реакции на стресс и развития мозга. (Деттмер и др., 2012; Спинелли и др., 2009, 2010). Качество и количество взаимодействий матери и детеныша у приматов также можно изменить, изменяя продолжительность времени, необходимого для кормления каждый день (Coplan et al, 2006). Переменная потребность в пище приводит к снижению чувствительности матерей к сигналам младенца, что приводит к повышенному уровню тревожного поведения, снижению социального поведения и нарушениям развития и функционирования гиппокампа у потомства (Coplan et al, 2005; Gorman et al, 2002; Jackowski). и др., 2011). В колониях макак жестокое обращение со стороны матери (например, таскание младенцев и наступление на них) происходит с частотой 2–10% и связано с задержкой начала социальных игр и повышенной агрессией в новых условиях (McCormack et al, 2006). ). Жестокие матери также часто подвергаются материнскому отторжению, что является предиктором нарушения ГГН, морфологии мозга и серотонинергической функции у потомства (Howell et al, 2013; Koch et al, 2014; Maestripieri et al, 2006; Sanchez et al, 2010). ). Также было замечено, что у нечеловеческих приматов частота контактов мать-младенец различается (т. е. чрезмерно опекающий стиль воспитания), а высокий уровень контактов предсказывает снижение исследования новой среды (Fairbanks and McGuire, 19).88). У мартышек выращивание происходит в семейном контексте, и более высокие показатели отказа от потомства со стороны матерей, отцов или братьев и сестер в течение первых 2 месяцев жизни приводят к повышенному стресс-индуцированному ответу кортизола (Birnie et al, 2013). Таким образом, социальный контекст воспитания может оказать длительное влияние на стресс-реактивность.
Влияние материнского поведения на развитие лабораторных грызунов
Изучение родительской заботы у лабораторных грызунов включает различные экспериментальные подходы, в том числе разлучение или лишение матери в раннем возрасте, индуцированное нарушение или усиление взаимодействия мать-младенец и естественные различия в материнской заботе. У грызунов длительное послеродовое разлучение с матерью вызывает повышение уровня глюкокортикоидного гормона (Stanton et al, 19). 88), повышенная реакция на стресс и нарушение когнитивных функций (Andersen et al, 1999; Lehmann et al, 1999). Полная депривация матери, сравнимая с опытом детей, находящихся в специализированных учреждениях, или питомников/обезьян, выращенных сверстниками, может быть реализована у крыс путем выращивания детенышей в отсутствие материнского контакта (искусственное выращивание). Щенки, выращенные в таких условиях, демонстрируют нарушение социального обучения (Levy et al, 2003), снижение внимания (Lovic and Fleming, 2004) и повышенную импульсивность (Lovic et al, 2011). В головном мозге детенышей, лишенных матери, снижены нейротрофические факторы, что может привести к нарушению развития нервной системы и снижению пластичности (Chatterjee et al, 2007). Снижение нейротропных факторов в префронтальной коре также наблюдается у детенышей, подвергшихся жестокому обращению в младенчестве (Roth et al, 2009).). Как и в случае привязанности у людей, фрагментация взаимодействия мать-младенец может быть обычным путем во многих из этих моделей неблагоприятных обстоятельств и играть опосредующую роль в прогнозировании долгосрочных результатов у потомства (Baram et al, 2012). Эта непредсказуемость ухода может быть достигнута за счет удаления подстилки из домашней клетки (Rice et al, 2008) и ведет к уменьшению заботливости и усилению оскорбительного взаимодействия с матерью (например, наступание на щенков, агрессивный уход и транспортировка щенков со стороны матери). конечность) с потомством, а также повышенный базальный уровень кортикостерона и нарушение обучения/памяти, вероятно, опосредованное передачей сигналов рецептора 1 рилизинг-фактора кортикотропина переднего мозга (Rice et al, 2008; Roth et al, 2009).; Ван и др., 2011).
Изучение естественных вариаций материнской заботы у грызунов предоставило привлекательную модель для изучения воздействия взаимодействия мать-младенец в пределах нормы. Наблюдения за крысами Long-Evans в послеродовой период показывают, что облизывание/груминг (LG) детенышей, одна из основных форм тактильной стимуляции во время постнатального развития, значительно различается у лактирующих самок (Champagne et al, 2003a). Во взрослом возрасте потомство с низким уровнем ЛГ по сравнению с высоким имеет повышенный стресс-индуцированный уровень глюкокортикоидов (Liu et al, 19). 97), снижение пластичности гиппокампа (Bagot et al, 2009) и нарушение обучения/памяти (Liu et al, 2000). Эти функциональные результаты связаны со сниженным содержанием белка глюкокортикоидного рецептора (GR) и мРНК в гиппокампе потомства, выращенного самками с низким уровнем LG (Francis et al, 1999; Liu et al, 1997). GR гиппокампа играет критическую роль в качестве отрицательной обратной связи в ответе HPA на стресс (Sapolsky et al, 1985), и сниженные уровни GR могут объяснить повышенную реакцию на стресс, наблюдаемую у потомков с низким уровнем LG (см. Рисунок 2). Подобно другим моделям грызунов невзгод в раннем возрасте, нейротрофические факторы уменьшаются в мозге потомства, выращенного матерью с низким LG (Liu et al, 2000). Эти материнские фенотипы также передаются потомству женского пола, так что опыт низкого LGG предсказывает низкий LGG у потомства (Champagne et al, 2003a). Напротив, потомство женского пола с низким LG демонстрирует повышенное сексуальное поведение, что позволяет предположить, что количество материнской заботы, испытываемой в младенчестве, приводит к сдвигам в репродуктивной стратегии потомства (Cameron et al, 2005, 2008). Эти материнские/репродуктивные фенотипы связаны со специфическими для областей мозга эффектами на уровни окситоцина и рецепторов эстрогена (ER) (Cameron et al, 2008; Champagne et al, 2001, 2003b). Важно отметить, что исследования перекрестного воспитания подтверждают, что эти нейробиологические эффекты являются следствием различий в послеродовом уходе за матерью (Champagne et al, 2006b; Francis et al, 19).99)
Схематическая иллюстрация влияния низкого уровня по сравнению с высоким уровнем у матери на уровни GR в гиппокампе. Низкий уровень материнского ЛГ связан со снижением уровня ГР, что приводит к нарушению отрицательной обратной связи на реакцию ГГН на стресс. CA1, cornu ammonis 1; CA3, cornu ammonis 3; ДГ, зубчатая извилина.
Слайд PowerPoint
Изображение полного размера
Качество материнской помощи в послеродовой период можно регулировать с помощью различных манипуляций с окружающей средой. У крыс материнский LG может быть увеличен путем содержания самок после отлучения от груди в условиях повышенного социального и физического обогащения (например, групповое содержание с игрушками и сложным дизайном) до спаривания (Champagne and Meaney, 2007). И наоборот, LG может быть снижена, если женщины испытывают длительные периоды социальной изоляции или стресса во время беременности (Champagne and Meaney, 2006a; Champagne 9).0009 и др. , 2007). Было продемонстрировано, что короткие периоды разлуки с матерью (т. е. продолжительностью 5–20 минут) увеличивают материнскую LG (Lee and Williams, 1974; Liu et al, 1997) и ослабляют стрессовую реакцию потомства (Ader, 1970). Этот эффект был приписан влиянию повышенной материнской стимуляции потомства на уровни GR в гиппокампе (Meaney et al, 1985). Кратковременное разлучение с матерью во время постнатального развития также продемонстрировало усиление памяти, зависящей от гиппокампа (Tang, 2001), хотя опыт новой среды во время разлуки может быть критическим фактором в прогнозировании этих эффектов (Tang et al, 2006). ). У мышей можно стимулировать поведение взрослых самок, направленное на детеныш, при помещении их в общее гнездо, состоящее из нескольких воспитателей и выводков (Curley et al, 2009).). Потомство, выращенное в общем гнезде, лучше ориентируется в новой среде, демонстрирует более высокий уровень социального поведения и повышенный уровень нейротрофинов в гиппокампе (Branchi et al, 2006a, 2006b, 2013; Curley et al, 2009). Во взрослом возрасте самки, воспитываемые в общине, проявляют более высокий уровень материнской заботы о собственном потомстве и имеют повышенный уровень гипоталамических рецепторов окситоцина (Curley et al, 2009). Таким образом, существуют поразительные параллели между опытом совместного воспитания и воспитанием матери с высоким уровнем LG.
Исследования влияния отца на развитие потомства у грызунов
Хотя в лабораторных исследованиях влияния родителей преобладают исследования материнской заботы, важно отметить, что отцы могут аналогичным образом влиять на развитие потомства. У социально моногамных и двуродительских видов лишение родительской заботы может привести к снижению социального признания (Cao et al, 2014), изменению развития игрового поведения (Wang et al, 2012), нарушению образования пар (Yu et al, 2012), и повышенное тревожное поведение (Jia et al, 2009).). Эти поведенческие нарушения связаны с нейробиологическими изменениями, вызванными отцовской депривацией, включая специфические для пола эффекты на экспрессию генов дофаминовых рецепторов в прилежащем ядре (Yu et al, 2012), снижение уровней рецептора окситоцина и ER альфа (ERα) (Cao et al. , 2014), изменение развития нейроэндокринных путей, участвующих в реакции на стресс (Seidel et al, 2011), и снижение сложности нейронов (Braun et al, 2013; Pinkernelle et al, 2009). Было показано, что у двухродительских калифорнийских мышей отцовское поведение изменяет развитие нейронных систем, участвующих в агрессии, что влияет на изменение отцовского поведения у потомства мужского пола (Frazier et al, 2006; Gleason and Marler, 2013). Манипулирование количеством отцовской заботы у калифорнийских мышей за счет использования высокой потребности в пище приводит к ухудшению обучения и памяти потомства, увеличению показателей тревожно-подобного поведения и изменению синаптического развития (Bredy et al, 2007). Таким образом, у видов, у которых отцы вносят свой вклад в заботу о потомстве, лишение отца может привести к нейробиологическим и поведенческим нарушениям у потомства.
ЭПИГЕНЕТИЧЕСКИЕ ПУТИ В РАЗВИВАЮЩЕМСЯ МОЗГЕ
Критический вопрос касается механизмов, с помощью которых вариации во взаимодействии родителей и потомков приводят к долговременным сдвигам в функциях и поведении мозга. Поиски механизма все больше указывали на молекулярные пути, регулирующие активность генов, включая метилирование ДНК, посттрансляционные модификации гистонов и некодирующие РНК, которые играют критическую роль в развитии мозга.
Метилирование ДНК
Добавление метильной группы к цитозинам в ДНК, особенно в динуклеотиды CpG, обычно считается стабильной и устойчивой эпигенетической модификацией (Разин, 1998), хотя следует отметить, что имеются данные о динамических изменениях этой эпигенетической метки. в ответ на острое воздействие окружающей среды (Barres et al, 2012) и формирование памяти (Day and Sweatt, 2010) во взрослом мозге. Когда цитозины метилируются, доступ к ДНК, как правило, снижается, и, следовательно, считается, что метилирование ДНК связано с молчанием генов (Razin, 19).98). Метилированная ДНК может рекрутировать метил CpG-связывающие белки (например, MeCP2), которые группируются вокруг ДНК и привлекают гистон-модифицирующие ферменты, которые индуцируют репрессивную структуру хроматина в окружающей генной области (Fan and Hutnick, 2005). Во время клеточной дифференцировки надежная передача паттернов метилирования ДНК от материнских клеток к дочерним способствует возникновению клеточного разнообразия и поддержанию клеточных фенотипов среди клеток, содержащих идентичные последовательности ДНК. Таким образом, эпигенетическая подпись клетки определяет клеточные фенотипы, которые могут наследоваться посредством митоза (Taylor and Jones, 19).85). Быстрая пролиферация и дифференцировка клеток, которые происходят во время пренатального и постнатального развития, требуют как de novo , так и поддерживающего метилирования ДНК. Нарушение этих процессов путем направленной делеции генов, кодирующих ДНК-метилтрансферазы (Dnmt1 или Dnmt3a/3b), приводит к гипометилированию всего генома, нарушению роста и эмбриональной или постнатальной летальности (Jackson-Grusby et al, 2001; Okano et al, 1999). ). У человека были идентифицированы полиморфизмы в генах DNMT (El-Maarri et al, 2009).), и эти генотипы могут быть важны для понимания индивидуальных различий в эпигенетической дисрегуляции и исходах развития нервной системы.
Посттрансляционные модификации гистонов
Экспрессия генов требует, чтобы ДНК стала доступной для факторов транскрипции и для общего механизма транскрипции, который включает РНК-полимеразу. Одной из стратегий достижения этого результата является модификация гистоновых белков в хроматине, чтобы ослабить физическое притяжение/взаимодействие между ДНК и гистоновыми аминокислотными «хвостами». Добавление или удаление химических групп из гистоновых хвостов может приводить к динамическим изменениям транскрипционной активности в зависимости от типа группы и места добавления/удаления (Peterson and Laniel, 2004). Например, триметилирование лизина 4 в хвосте гистона h4 (h4K4) связано с повышенной экспрессией генов, тогда как триметилирование лизина 9в гистоне h4 (h4K9) соответствует пониженной экспрессии гена (Barski et al, 2007; Koch et al, 2007). Есть много химических групп, которые могут быть добавлены или удалены из гистоновых хвостов, и в совокупности эти посттрансляционные модификации создают сложную стратегию динамической регуляции экспрессии генов (Jenuwein and Allis, 2001). Нарушение ферментативного механизма, который способствует этим эпигенетическим процессам, оказывает глубокое влияние на развитие. Например, мутация в JARID1C, который кодирует гистондеметилазу, связана с нарушением развития нервной системы, включая аутизм (Adegbola et al, 2008) и умственную отсталость (Tahiliani et al, 2007).
Некодирующие РНК
Хотя большая часть генома не кодирует белковый продукт, это не означает, что большая часть генома не выполняет никакой функции. Это понятие становится все более очевидным в случае малых некодирующих молекул РНК (например, микроРНК (миРНК) и пиРНК). miRNAs могут вызывать посттранскрипционное замалчивание генов путем спаривания оснований с РНК-мишенью (mRNAs), предотвращая трансляцию, расщепляя мРНК или способствуя деградации мРНК (Sato et al, 2011). Эти РНК, благодаря их способности нацеливаться на несколько генов, могут служить критическими эпигенетическими регуляторами. Более того, изменения в экспрессии специфических микроРНК связаны с переходом от материнского к зиготическому контролю транскрипции, детерминации клеточных судеб и формированию паттернов развития в развивающемся организме (Pauli et al, 2011), а также мутациям, ингибирующим продукцию Dicer, фермент, который облегчает продукцию зрелой микроРНК, может нарушать развитие нервной системы (Bernstein et al, 2003; Davis et al, 2008).
ЭПИГЕНЕТИЧЕСКОЕ ПРОГРАММИРОВАНИЕ ЧЕРЕЗ ИЗМЕНЕНИЯ В МАТЕРИНСКОЙ ЗАБОТЕ
Объединение данных о влиянии взаимодействия родителей и потомства на развитие нервной системы с достижениями в нашем понимании молекулярных основ регуляции генов (краткое описание см. проникновения в биологическое воплощение раннего жизненного опыта. В одном из новаторских исследований, связывающих качество постнатальной среды с эпигенетическими вариациями, изучается связь между опытом жизни в условиях низкой против high-LG у крыс и метилирование ДНК в промоторной области гена GR ( Nr3c1 ) (Weaver et al, 2004). Повышенное метилирование ДНК Nr3c1 наблюдается в ткани гиппокампа взрослых потомков мужского пола с низким уровнем LG по сравнению с матерями с высоким уровнем LG. В соответствии с результатами исследований по перекрестному приему, эти эффекты, связанные с материнской заботой, не проявляются ни в пренатальном периоде, ни при рождении. Скорее, повышенное метилирование ДНК Nr3c1 возникает в течение первой постнатальной недели и стабильно сохраняется во взрослом возрасте. Ацетилирование гистонов и связывание факторов транскрипции с Промоторная область Nr3c1 также редуцирована среди потомков самок с низким LG (см. рис. 3), что согласуется с гипотезой о том, что повышенное метилирование ДНК в этой геномной области приводит к сайленсингу транскрипции (Weaver et al, 2004). Хотя GR представляет собой ключевой регулятор в путях реакции на стресс, важно отметить, что вариации материнского LG связаны с изменениями в транскрипции сотен генов (Weaver et al, 2006) и с широкими эпигенетическими изменениями в гиппокампе, которые включают как метилирование ДНК, так и ацетилирование гистонов (h4K9) (Макгоуэн и др., 2011). Другие гены-мишени с эпигенетическими вариациями, связанными с опытом низкого против высокого LG, включают глутаматдекарбоксилазу ( Gad1 ) и Grm1 , которая кодирует метаботропный рецептор глутамата I типа (mGluR1). Низкие уровни постнатального материнского LG связаны с повышенным метилированием ДНК регуляторных областей этих генов в гиппокампе, а также со сниженными уровнями ацетилирования h4K9 и триметилирования h4K4 (Bagot et al, 2012; Zhang et al, 2010). Экспрессия DNMT1 увеличивается в гиппокампе потомства с низким уровнем LG, указывая на потенциальный механизм этих широких эпигенетических эффектов (Zhang et al, 2010).
Сводка эпигенетических эффектов взаимодействий между родителем и потомством Полноразмерная таблица
Рисунок 3Схематическая иллюстрация влияния материнского LG с низким и высоким на метилирование ДНК в промоторной области Nr3c1 . Повышенные уровни метилирования ДНК Nr3c1 связаны с эпигенетическим молчанием экспрессии генов посредством ингибирования связывания факторов транскрипции (TF) и РНК-полимеразы (RNApol).
Слайд PowerPoint
Изображение в полный размер
Эпигенетические вариации, связанные с опытом материнской заботы, могут также объяснить сниженные уровни ER гипоталамуса, наблюдаемые у потомства женского пола с низким уровнем LG по сравнению с матерями с высоким уровнем LG. Низкие уровни LG в раннем постнатальном периоде связаны со снижением транскрипции Esr1 , гена, кодирующего ERα, в медиальной преоптической области гипоталамуса потомства взрослых самок (Champagne et al, 2003b). Эпигенетический анализ Промотор Esr1 указывает на то, что низкий уровень LG связан с повышенным метилированием ДНК, пониженным триметилированием h4K4 и повышенным триметилированием h4K9 в этой геномной области (Champagne et al, 2006b; Pena et al, 2013). Подобно эпигенетическим изменениям в пределах Nr3c1 , вариации метилирования ДНК и модификаций гистонов в пределах Esr1 появляются в постнатальный период и отсутствуют при рождении (Pena et al, 2013). Половое эпигенетическое программирование Esr1 может объяснять половой диморфизм в нейронных цепях, и может случиться так, что LG, испытываемые во время постнатального развития, способствуют этим устойчивым эффектам (Kurian et al, 2010). Среди совместно выращенных мышей, у которых наблюдается усиление LG, наблюдается повышенное ацетилирование h4 в гиппокампе в некоторых промоторах генов нейротрофического фактора головного мозга ( Bdnf ), что может привести к временному увеличению уровней белка BDNF (Branchi et al, 2011).
ЭПИГЕНЕТИЧЕСКОЕ ВОЗДЕЙСТВИЕ МАТЕРИНСКОЙ БЕЗРАБОТКИ И НАСИЛИЯ
Глубокие последствия раннего жизненного опыта наблюдаются в исследованиях пренебрежения и жестокого обращения, и растет поддержка участия эпигенетических путей в долгосрочных последствиях этих форм невзгод (обобщение см. в Таблице 1).
Материнское пренебрежение у грызунов и приматов
У мышей разлучение с матерью вызывает повышенную реакцию на стресс у потомства, что может быть связано с экспрессией генов и эпигенетическими изменениями в связанных с HPA генах, таких как вазопрессин ( Avp ), фактор высвобождения кортикотропина ( Crf ) и Nr3c1 . Например, повышенная чувствительность к стрессу у потомства может быть связана с повышенной экспрессией гипоталамического AVP, поскольку разделение матери связано со снижением метилирования ДНК и снижением уровней связывания MeCP2 (метил-Cpg-связывающий белок 2) в пределах регуляторной области Avp . в этой области мозга (Murgatroyd et al, 2009). У взрослых мышей, переживших постнатальное отделение от матери, наблюдается активация CRF в гиппокампе, и эта повышенная генная активность связана с повышенным ацетилированием h4 и снижением метилирования ДНК Crf промоутер. Интересно, что эпигенетические изменения в Crf , связанные с разлучением с матерью, являются обратимыми в ответ на обогащение окружающей среды после отъема (т. е. доступ к повышенной сенсорной стимуляции и упражнениям), что приводит к улучшению нейробиологических и поведенческих нарушений, вызванных разлукой с матерью (Wang et al. др., 2014). Повышенное метилирование ДНК Nr3c1 гиппокампа также наблюдалось после разлучения с матерью у мышей, что позволяет предположить, что это может быть общим эпигенетическим путем для многих форм невзгод в раннем возрасте (Kember et al, 2012; Kundakovic et al, 2013).
Помимо связанных со стрессом генов в гиппокампе и гипоталамусе, нарушение взаимодействия мать-младенец в раннем возрасте может изменить эпигенетическое состояние различных генов-мишеней, связанных с функцией развития нервной системы во многих областях мозга. Постнатальное отделение от матери изменяет триметилирование h4K27 в гене, кодирующем N -метил-D-аспартатный рецептор ( Grin2b ) в гиппокампе с последствиями для обучения и памяти (Rodenas-Ruano et al, 2012). Было обнаружено, что в миндалевидном теле отделение от матери увеличивает метилирование ДНК рецептора нейротензина (9).0009 Ntsr1 ), связанный с уменьшением мРНК Ntsr1 и повышенным поведением замирания во время кондиционирования страха (Toda et al, 2014). В гипофизе у мышей, разлученных с матерью, наблюдается долгосрочное снижение цитозинового метилирования гена проопиомеланокортина ( Pomc ), ключевого фактора нейроэндокринных стрессовых реакций (Wu et al, 2014). В рамках этих исследований может быть важно рассмотреть временную динамику эпигенетических изменений, вызванных окружающей средой. Исследования, изучающие влияние разлучения с матерью у мышей на эпигенетическую изменчивость в пределах Ген Bdnf в ткани гиппокампа свидетельствует о двухфазном ответе, таком как снижение диметилирования h4K9 (репрессивная хроматиновая метка) при отъеме и у молодых взрослых особей и повышение диметилирования h4K9 у стареющих (15-месячных) мышей (Suri et al. , 2009). др., 2013).
У макак-резусов глобальные изменения метилирования ДНК, которые сохраняются во взрослом возрасте, наблюдались в префронтальной коре в ответ на воспитание сверстников. Интересно, что этот эпигенетический признак материнской депривации в раннем возрасте также наблюдается в образцах крови детей, воспитанных сверстниками 9.0009 против макак, выкормленных матерями, что свидетельствует о потенциальной ценности периферических тканей при оценке длительного воздействия невзгод в раннем возрасте (Provencal et al, 2012). Индуцированные разделением эффекты на эпигенетическое состояние SLC6A4 , гена, кодирующего переносчик серотонина (5-HTT), также наблюдались (Kinnally et al, 2010). Подобно людям, макаки-резусы обладают полиморфизмом SLC6A4 , который связан с изменением риска или устойчивости к стрессовым факторам, таким как жестокое обращение в детстве (Caspi et al, 2003). Макаки-резусы, выращенные в условиях отделения от матери и обладающие 9Аллель 0009 SLC6A4 повышает риск метилирования ДНК SLC6A4 в клетках крови (Kinnally et al, 2010). Это взаимодействие между генетическими и эпигенетическими вариациями, возникающими в ответ на невзгоды в раннем возрасте, может способствовать нашему пониманию механизмов, связанных с взаимодействием генов с окружающей средой, и станет важным фактором в исследованиях человека невзгод в раннем возрасте.
Эпигенетические эффекты жестокого обращения с животными у грызунов
Послеродовое жестокое обращение, вызванное удалением постельного материала, связано со снижением экспрессии BDNF в префронтальной коре во взрослом возрасте (Roth et al, 2009).). Среди потомков, подвергшихся жестокому обращению, наблюдается повышенное метилирование ДНК промоторных областей Bdnf гена, что может привести к снижению пластичности нейронов (см. Рисунок 4). Анализ эпигенетических изменений, связанных с оскорбительным уходом за ребенком во время развития, позволяет предположить сложные взаимодействия между полом и возрастом в проявлении реакции на неблагоприятный опыт (Blaze and Roth, 2013).
Схематическая иллюстрация влияния жестокого обращения и заботы о матери на метилирование ДНК в Промоторная область Bdnf . Повышенные уровни метилирования ДНК Bdnf, вызванные злоупотреблением, связаны с эпигенетическим подавлением экспрессии генов посредством ингибирования связывания РНК-полимеразы (RNApol) с сайтом начала транскрипции.
Слайд PowerPoint
Изображение в натуральную величину
Эпигенетические последствия невзгод детства у людей
Проблема перевода этих лабораторных данных в исследования на людях связана, прежде всего, с невозможностью изучения эпигенетических вариаций в мозге живых людей. Однако в случае жестокого обращения в детстве в нескольких исследованиях изучались эпигенетические вариации, присутствующие в посмертной ткани мозга. Анализ ткани головного мозга жертв самоубийств с жестоким обращением в детстве или без него, а также контрольных субъектов, не совершавших самоубийств, показывает, что экспрессия ГР гиппокампа снижается у лиц, подвергшихся насилию, и это снижение транскрипционной активности связано с повышенным метилированием ДНК и снижением фактора транскрипции. обязательный в течение промотор гена NR3C1 (McGowan et al., 2009). Таким образом, представляется, что исследования, проведенные на лабораторных грызунах, имеют трансляционную значимость. Полногеномные анализы метилирования ДНК показывают, что эти эффекты не специфичны для промотора гена NR3C1 и включают как гипометилирование, так и гиперметилирование сотен промоторов генов в гиппокампе (Labonte et al, 2012). Однако опора на посмертные ткани, особенно от жертв самоубийства, является значительным препятствием для расширения изучения эпигенетической изменчивости у людей, и это привело к увеличению зависимости от образцов периферической ДНК (например, из крови или слюны). Хотя эпигенетические профили различают типы клеток, может случиться так, что эти периферические источники ДНК можно использовать в качестве биомаркера неблагополучия в раннем возрасте (Uher and Weaver, 2014). Анализы 9Промотор гена 0009 NR3C1 с использованием образцов крови указывает на то, что жестокое обращение в детстве (например, физическое, сексуальное или эмоциональное насилие) предсказывает повышенное метилирование ДНК NR3C1 (Perroud et al, 2014). Более того, увеличение метилирования ДНК NR3C1 , связанное с невзгодами детства, предсказывает стрессовую реактивность, которая может иметь последствия для риска психопатологии (Tyrka et al, 2012). Подобно результатам, полученным на моделях грызунов, эпигенетическая изменчивость человека в ответ на невзгоды детства выходит за пределы NR3C1 . Среди детей, воспитывающихся в детских домах, в клетках крови выявляется полногеномное гиперметилирование (Наумова и др., 2012). Широкие эпигенетические вариации, включая как гипометилирование ДНК, так и гиперметилирование ДНК, очевидны в образцах крови взрослых, связанных с жестоким обращением в детстве (Suderman et al, 2014). Интересно, что гены-мишени, идентифицированные в этих анализах, включали несколько miRNAs, тем самым создавая потенциал для экстенсивной эпигенетической дисрегуляции после изменений в метилировании ДНК. Таким образом, в периферических тканях существуют эпигенетические признаки социальных невзгод в раннем возрасте. Существуют также периферические эпигенетические маркеры психического расстройства (D’Addario et al, 2012; Fuchikami et al, 2011). Недавнее исследование показало, что некоторые межиндивидуальные вариации метилирования ДНК коррелируют между мозгом и кровью человека (Davies et al, 2012), что также указывает на то, что эпигенетическое профилирование периферических тканей может быть полезным для изучения нарушений головного мозга. Важным вопросом, который необходимо решить, является прогностическая ценность эпигенетических вариаций в крови или слюне, вызванных несчастными случаями, для этих исходов развития нервной системы и психики.
ОТ ОДНОГО ПОКОЛЕНИЯ К СЛЕДУЮЩЕМУ
Появляется все больше данных о влиянии невзгод в раннем возрасте на потомство и внуков (Champagne, 2008; Curley et al, 2011), однако остается вопрос, как это достигается . Здесь мы кратко рассмотрим два пути, с помощью которых эти эффекты могут быть достигнуты, и то, как эпигенетическая изменчивость, вызванная качеством постнатальной среды, может способствовать этой передаче.
Эпигенетическая передача, зависящая от опыта
Нервные системы, которые регулируют родительское поведение, чувствительны к качеству ухода за ребенком в младенчестве (Champagne, 2008). У крыс опыт низкого уровня материнской заботы смещает развитие гипоталамической окситоциновой и ER систем, делая потомство менее чувствительным к гормональному праймингу, тем самым снижая материнский LG (см. Рисунок 5). Эти нейроэндокринные последствия материнской заботы приводят к передаче LG от одного поколения к другому (Champagne, 2008). С передачей материнской ЛГ также имеет место передача от нескольких поколений исходов, связанных с переживанием низких уровней ЛГ, таких как снижение пластичности нейронов и повышенная реакция ГГА на стресс (Francis et al, 19).99). Жестокая забота может также изменить развитие материнского мозга потомства, что приведет к усилению жестокой заботы среди детей, подвергшихся жестокому обращению (Roth et al, 2009). У биродительских видов отцы могут аналогичным образом изменять нейроэндокринные цепи, регулирующие отцовское поведение у потомства мужского пола, что приводит к передаче вариаций отцовского поведения из поколения в поколение (Bester-Meredith and Marler, 2003). Эти эффекты развития взаимодействия родителей и потомства могут объяснить межпоколенческую передачу широкого спектра поведенческих фенотипов. Сохранение этих эффектов развития во взрослом возрасте посредством эпигенетических изменений формирует контекст развития следующего поколения потомства. В отличие от генетического пути наследования, эта передача поведения очень пластична в ответ на окружающую среду, с которой сталкиваются в более позднем возрасте (т. е. стресс, социальная изоляция и социальные взаимодействия), так что, когда среда меняется, то же самое может происходить и с родительскими качествами. взаимодействие потомков (Champagne and Meaney, 2006a; Champagne и др. , 2007).
Цикл эпигенетических, нейроэндокринных и поведенческих изменений, вызванных низким уровнем ЛГ у матери. Эпигенетическая регуляция Esr1 материнской заботой может привести к каскаду изменений, которые изменяют развитие F1 (потомство) и F2 (внучатое потомство) с потенциалом для непрерывности вариаций материнского поведения в потомстве поколения F3.
Слайд PowerPoint
Изображение в натуральную величину
Эпигенетическое наследование зародышевой линии
Появляется все больше предположений о том, что эпигенетические модификации, наследуемые во время митоза, также могут наследоваться последующими поколениями (Daxinger and Whitelaw, 2012). Несмотря на то, что на этапах развития после оплодотворения происходит полногеномное эпигенетическое перепрограммирование, наблюдение трансгенерационного воздействия родительского опыта, которое может сохраняться даже у потомков поколения F3, подняло вопрос о том, не полностью ли стираются индуцированные окружающей средой эпигенетические изменения. и передаются через зародышевую линию. Доказательства, подтверждающие эти трансгенерационные эффекты, обычно сосредоточены на передаче отцовской эпигенетической информации (Anway and Skinner, 2006; Morgan and Bale, 2011; Saavedra-Rodriguez and Feig, 2013). У самцов мышей опыт разлучения с матерью во время постнатального развития связан с нарушением социального поведения и измененными тревожными и депрессивными реакциями, которые сохраняются из поколения в поколение и совпадают с измененной серотонинергической функцией (Franklin et al, 2010, 2011). В сперме этих самцов постнатальное отделение матери вызывает повышенное метилирование ДНК в Mecp2 и гена каннабиноидного рецептора типа 1 ( Cb1 ), а также снижение метилирования ДНК в гене рецептора 2 рилизинг-фактора кортикотропина ( Crfr2 ) (Franklin et al, 2010). Эти эпигенетические изменения также сохраняются в коре и сперматозоидах потомства этих самцов. miRNAs также могут играть критическую роль в передаче эффектов отделения от матери (Gapp et al, 2014). Эти результаты дополняют растущее количество данных о том, что широкий спектр воздействий (например, токсины, питание и стресс) может иметь эпигенетические последствия для зародышевой линии (Curley et al, 2011).
Ключевой вопрос, относящийся как к эпигенетической передаче, зависящей от опыта, так и к эпигенетическому наследованию зародышевой линии, касается того, как приобретаются эпигенетические метки. В случае изменений в метилировании ДНК Nr3c1 гиппокампа, in vitro и in vivo исследования показывают, что активация нейронов, включающая передачу сигналов серотонинергических гормонов и гормонов щитовидной железы, может быть предшественником клеточных и молекулярных изменений, которые приводят к Nr3c1 гиперметилирование как следствие опыта низкого уровня материнской заботы (Champagne, 2013). Однако было замечено, что ослабление материнской заботы, лишение матери и жестокое обращение вызывают эпигенетические последствия для всего генома (Labonte et al, 2012; Naumova et al, 2012; Provencal et al, 2012; Weaver et al, 2006), присутствующие как в головного мозга и на периферии. Кроме того, хотя многие исследования были сосредоточены на эпигенетических вариациях в гиппокампе, ясно, что многие области мозга чувствительны к эпигенетическим нарушениям, вызванным ранним жизненным опытом (например, гипоталамус, префронтальная кора и миндалевидное тело; см. Таблицу 1). Таким образом, для понимания процесса биологического закрепления переживаний ранней жизни потребуется более всесторонний анализ каскада молекулярных изменений во многих тканях-мишенях, происходящих после этого переживания. В свете двухфазных эпигенетических изменений, наблюдаемых в мозге в ответ на отделение матери (Suri et al, 2013), повторные измерения с течением времени будут необходимы для этого понимания. Наконец, понимание механизмов, с помощью которых индуцируется наследуемая эпигенетическая изменчивость, потребует тщательного рассмотрения взаимосвязей между мозгом, эндокринной системой и иммунными реакциями, которые могут координировать клеточные и молекулярные реакции в тканях (т. е. нейрональные и гаметы).
ПОСЛЕДСТВИЯ И НАПРАВЛЕНИЯ НА БУДУЩЕЕ
За последнее десятилетие мы значительно продвинулись в понимании роли эпигенетических путей в связи раннего опыта с исходами развития нервной системы, и становится все более очевидным, что эти эпигенетические изменения могут лежать в основе риска в более позднем возрасте. психопатологии. Однако этот подход к происхождению психической дисфункции все еще находится в зачаточном состоянии, и многие вопросы остаются без ответа. Хотя феномен, посредством которого вариации в детском опыте связаны с эпигенетическими вариациями, продолжает устанавливаться и уточняться, пока что у нас есть ограниченное понимание процесса, посредством которого достигаются эти эпигенетические эффекты. Понимание механизмов, вызывающих эти эффекты, потребует интеграции высокоточных и чувствительных измерений нескольких слоев эпигенетической информации (т.е. метилирования ДНК, модификаций гистонов и микроРНК). Особое значение при рассмотрении применения эпигенетики для изучения развития нервной системы человека будет иметь изучение взаимодействия между вариациями последовательности ДНК и эпигенетическими вариациями, индуцированными окружающей средой. Доверие к периферическим измерениям эпигенетической изменчивости также потребует дальнейшей проверки для решения ключевых вопросов в этой области: каково значение этих биомаркеров для психической дисфункции? Насколько предсказуемы эти периферические эпигенетические модификации сходных эпигенетических профилей в головном мозге? Получение ответов на эти вопросы и установление причинно-следственной роли эпигенетических вариаций в развитии риска заболеваний в более позднем возрасте создаст серьезные проблемы в этой области, но потенциально может определить новые терапевтические цели и стратегии.
В соответствии с этими эмпирическими вопросами будущие исследования влияния детского опыта на развитие потребуют теоретической основы, которая объясняет более широкий эволюционный вопрос о том, почему задействованы эти эпигенетические пути. Пластичность развивающейся особи в ответ на окружающую среду представляет собой адаптивную систему, которая лучше подготавливает особь к жизни и размножению в этой среде. Лишение заботливой родительской заботы можно рассматривать как сигнал о качестве окружающей среды, а способность реагировать на этот сигнал можно считать важным эволюционным преимуществом. Наблюдается, что в случае опыта с низким уровнем LG потомство принимает когнитивные и репродуктивные стратегии, которые могут лучше подходить для среды с высокой угрозой (Cameron et al, 2005; Champagne et al, 2008). Интересно, что многие животные модели, используемые в изучении влияния родительской заботы на потомство, манипулируют доступностью ресурсов (т. е. за счет уменьшения количества подстилки или изменяющейся потребности в корме), создавая сильные параллели с обедненной социально-экономической средой, в которой нарушается взаимодействие между родителем и потомством человека. Молекулярная и нейробиологическая адаптация к этим средам может выглядеть как дисфункция вне контекста, и появляется все больше предположений о том, что именно несоответствие между качеством окружающей среды в раннем возрасте и в более позднем возрасте приводит к возникновению нейробиологических расстройств (Schmidt, 2011). Таким образом, стабильность качества окружающей среды, такая, что окружающая среда в раннем возрасте подобна той, которая будет испытана в более поздней жизни, может быть важным фактором в этих исследованиях развития. Эту стабильность и предсказуемость качества окружающей среды необходимо учитывать, когда последствия родительской заботы передаются из поколения в поколение. Хотя трансгенерационные эффекты раннего жизненного опыта могут адаптировать потомство и его потомство, польза от этой адаптации будет зависеть от того, насколько хорошо среда предков предсказывает среду потомков. У видов с короткой продолжительностью жизни и ранним началом размножения (например, у лабораторных грызунов) эта эпигенетическая передача может иметь значение. Однако споры относительно применимости этих эффектов к людям будут оставаться до тех пор, пока мы не достигнем более глубокого понимания механизмов индуцированной окружающей средой эпигенетической изменчивости, степени пластичности/обратимости этих молекулярных изменений и компромиссов, которые происходят для поддержания этой динамики. путь наследования.
ФИНАНСИРОВАНИЕ И РАСКРЫТИЕ ИНФОРМАЦИИ
Авторы заявляют об отсутствии конфликта интересов.
Ссылки
-
Адегбола А., Гао Х., Соммер С., Браунинг М. (2008). Новая мутация в JARID1C/SMCX у пациента с расстройством аутистического спектра (РАС). Am J Med Genet A 146A : 505–511.
КАС пабмед Статья Google ученый
-
Адер Р. (1970). Влияние раннего опыта на адренокортикальный ответ на различную величину стимуляции. Physiol Behav 5 : 837–839.
КАС пабмед Статья Google ученый
-
Андерсен С.Л., Лисс П.Дж., Дюмон Н.Л., Тейхер М.Х. (1999). Стойкие нейрохимические эффекты раннего отделения матери от лимбических структур. Ann NY Acad Sci 877 : 756–759.
КАС пабмед Статья Google ученый
-
Anway MD, Skinner MK (2006).
Эпигенетические трансгенерационные действия эндокринных разрушителей. Эндокринология 147 : S43–S49. КАС пабмед Статья Google ученый
-
Bagot RC, van Hasselt FN, Champagne DL, Meaney MJ, Krugers HJ, Joels M (2009). Материнская забота определяет быстрое воздействие медиаторов стресса на синаптическую пластичность зубчатой извилины гиппокампа взрослых крыс. Neurobiol Learn Mem 92 : 292–300.
ПабМед Статья Google ученый
-
Bagot RC, Zhang TY, Wen X, Nguyen TT, Nguyen HB, Diorio J et al (2012). Изменения в послеродовом материнском уходе и эпигенетическая регуляция экспрессии метаботропного глутаматного рецептора 1 и функции гиппокампа у крыс. Proc Natl Acad Sci USA 109 (Приложение 2): 17200–17207.
КАС пабмед Статья ПабМед Центральный Google ученый
-
Барам Т.
З., Дэвис Э.П., Обенаус А., Сэндман К.А., Смолл С.Л., Солодкин А. и др. (2012). Фрагментарность и непредсказуемость раннего жизненного опыта при психических расстройствах. Am J Психиатрия 169 : 907–915. ПабМед ПабМед Центральный Статья Google ученый
-
Баррес Р., Ян Дж., Иган Б., Трибак Дж. Т., Расмуссен М., Фриц Т. и др. (2012). Острые физические нагрузки реконструируют метилирование промотора в скелетных мышцах человека. Cell Metab 15 : 405–411.
КАС пабмед Статья Google ученый
-
Barski A, Cuddapah S, Cui K, Roh TY, Schones DE, Wang Z et al (2007). Профилирование метилирования гистонов в геноме человека с высоким разрешением. Сотовый 129 : 823–837.
КАС Статья пабмед Google ученый
-
Бернар К.
, Дозье М. (2010). Изучение реакции кортизола младенцев на лабораторные задачи среди детей с различной дезорганизацией привязанности: стрессовая реакция или возвращение к исходному уровню? Dev Psychol 46 : 1771–1778. ПабМед ПабМед Центральный Статья Google ученый
-
Bernstein E, Kim SY, Carmell MA, Murchison EP, Alcorn H, Li MZ et al (2003). Дайсер необходим для развития мышей. Nat Genet 35 : 215–217.
КАС пабмед Статья Google ученый
-
Бестер-Мередит Дж. К., Марлер К.А. (2003). Вазопрессин и передача отцовского поведения из поколения в поколение у спаренных мышей Peromyscus, воспитанных перекрестно. Behav Neurosci 117 : 455–463.
КАС пабмед Статья Google ученый
-
Бирни А.К., Тейлор Дж.Х., Кавано Дж., Френч Дж.
А. (2013). Качество материнской и отцовской заботы предсказывает более позднюю реакцию на стресс у совместно размножающихся мартышек (Callithrix geoffroyi). Психонейроэндокринология 38 : 3003–3014. ПабМед Статья Google ученый
-
Блейз Дж., Рот Т.Л. (2013). Воздействие жестокого обращения со стороны опекунов изменяет уровни экспрессии эпигенетических регуляторов в медиальной префронтальной коре. Int J Dev Neurosci 31 : 804–810.
КАС пабмед Статья Google ученый
-
Бранчи И., Керли Дж. П., Д’Андреа И., Чирулли Ф., Шампань Ф.А., Аллева Э. (2013). Раннее взаимодействие с матерью и сверстниками независимо формирует социальные навыки взрослых и формирует уровни BDNF и рецепторов окситоцина в мозгу. Психонейроэндокринология 38 : 522–532.
КАС пабмед Статья Google ученый
-
Бранчи И.
, Д’Андреа И., Фиоре М., Ди Фаусто В., Алоэ Л., Аллева Э. (2006a). Раннее социальное обогащение формирует социальное поведение и уровни фактора роста нервов и мозгового нейротрофического фактора в мозге взрослых мышей. Биол Психиатрия 60 : 690–696. КАС пабмед Статья Google ученый
-
Бранчи И., Д’Андреа И., Сиетзема Дж., Фиоре М., Ди Фаусто В., Алоэ Л. и др. (2006b). Раннее социальное обогащение увеличивает уровни BDNF в гиппокампе взрослых и выживаемость BrdU-позитивных клеток, одновременно усиливая тревожное и «депрессивное» поведение. J Neurosci Res 83 : 965–973.
КАС Статья пабмед Google ученый
-
Бранчи И., Карпова Н.Н., Д’Андреа И., Кастрен Э., Аллева Э. (2011). Эпигенетические модификации, вызванные ранним обогащением, связаны с изменениями времени индукции экспрессии BDNF. Neurosci Lett 495 : 168–172.
КАС пабмед Статья Google ученый
-
Браун К., Зайдель К., Холечка Р., Гроегер Н., Поегель Г. (2013). Отцовская депривация изменяет развитие катехоламинергической иннервации в префронтальной коре и связанных с ней лимбических областях мозга. Функция структуры мозга 218 : 859–872.
КАС пабмед Статья Google ученый
-
Бреди Т.В., Браун Р.Е., Мини М.Дж. (2007). Влияние доступности ресурсов на заботу о двух родителях, а также на нервное и поведенческое развитие потомства калифорнийской мыши (Peromyscus californicus). Eur J Neurosci 25 : 567–575.
ПабМед Статья Google ученый
-
Кэмерон Н., Дель Корпо А., Диорио Дж., Макаллистер К., Шарма С., Мини М.Дж. (2008). Материнское программирование полового поведения и гипоталамо-гипофизарно-гонадная функция у самок крыс.
PLoS One 3 : e2210. ПабМед ПабМед Центральный Статья КАС Google ученый
-
Кэмерон Н.М., Шампань Ф.А., Родитель К., Фиш Э.В., Одзаки-Курода К., Мини М.Дж. (2005). Программирование индивидуальных различий в защитных реакциях и репродуктивных стратегиях у крыс посредством различий в материнской заботе. Neurosci Biobehav Rev 29 : 843–865.
ПабМед Статья Google ученый
-
Cao Y, Wu R, Tai F, Zhang X, Yu P, An X et al (2014). Неонатальная депривация отца ухудшает социальное признание и изменяет уровни экспрессии окситоцина и альфа-мРНК рецептора эстрогена в MeA и NAcc, а также окситоцина в сыворотке у мандариновых полевок. Horm Behav 65 : 57–65.
КАС пабмед Статья Google ученый
-
Карлсон Э.А. (1998). Проспективное лонгитюдное исследование дезорганизации/дезориентации привязанности.
Ребенок Дев 69 : 1107–1128. КАС пабмед Статья Google ученый
-
Карлсон Э.А., Эгеланд Б., Сроуф Л.А. (2009). Проспективное исследование развития пограничных симптомов личности. Дев-психопат 21 : 1311–1334.
ПабМед Статья Google ученый
-
Карпентер Л.Л., Шаттак Т.Т., Тырка А.Р., Герациоти Т.Д., Прайс Л.Х. (2011). Влияние физического насилия в детстве на реакцию кортизола на стресс. Психофармакология (Берл) 214 : 367–375.
КАС Статья Google ученый
-
Caspi A, Sugden K, Moffitt TE, Taylor A, Craig IW, Harrington H et al (2003). Влияние жизненного стресса на депрессию: модерация полиморфизмом в гене 5-НТТ. Наука 301 : 386–389.
КАС Статья пабмед Google ученый
-
Champagne DL, Bagot RC, van Hasselt F, Ramakers G, Meaney MJ, de Kloet ER и др.
(2008). Материнская забота и пластичность гиппокампа: свидетельство зависимой от опыта структурной пластичности, измененного синаптического функционирования и дифференциальной реакции на глюкокортикоиды и стресс. J Neurosci 28 : 6037–6045. КАС пабмед Статья ПабМед Центральный Google ученый
-
Шампанское Ф., Диорио Дж., Шарма С., Мини М.Дж. (2001). Естественные вариации материнского поведения у крыс связаны с различиями в индуцируемых эстрогеном центральных окситоциновых рецепторах. Proc Natl Acad Sci USA 98 : 12736–12741.
КАС пабмед ПабМед Центральный Статья Google ученый
-
Шампанское FA (2008 г.). Эпигенетические механизмы и трансгенерационные эффекты материнской заботы. Передний нейроэндокринол 29 : 386–397.
КАС пабмед ПабМед Центральный Статья Google ученый
-
Шампанское FA (2013).
Ранняя среда, глюкокортикоидные рецепторы и поведенческая эпигенетика. Behav Neurosci 127 : 628–636. ПабМед Статья Google ученый
-
Шампанское Ф.А., Фрэнсис Д.Д., Мар А., Мини М.Дж. (2003a). Вариации материнской заботы у крысы как опосредующее влияние среды на развитие. Physiol Behav 79 : 359–371.
КАС пабмед Статья Google ученый
-
Шампанское FA, Meaney MJ (2006a). Стресс во время беременности изменяет послеродовой уход матери и развитие потомства в модели грызунов. Биол Психиатрия 59 : 1227–1235.
КАС Статья пабмед Google ученый
-
Шампанское FA, Мини М.Дж. (2007). Трансгенерационное влияние социальной среды на различия в материнской заботе и поведенческой реакции на новизну. Behav Neurosci 121 : 1353–1363.
ПабМед Статья Google ученый
-
Шампанское Ф.А., Уивер И.С., Диорио Дж., Дымов С., Шиф М., Мини М.Дж. (2006b). Материнская забота связана с метилированием промотора рецептора эстрогена-альфа1b и экспрессией рецептора эстрогена-альфа в медиальной преоптической области потомства женского пола. Эндокринология 147 : 2909–2915.
КАС пабмед Статья Google ученый
-
Шампанское Ф.А., Уивер И.С., Диорио Дж., Шарма С., Мини М.Дж. (2003b). Естественные вариации материнской заботы связаны с экспрессией альфа-рецептора эстрогена и чувствительностью к эстрогену в медиальной преоптической области. Эндокринология 144 : 4720–4724.
КАС пабмед Статья Google ученый
-
Чаттерджи Д., Чаттерджи-Чакраборти М., Рис С., Коши Дж., Медейрос де Медейрос С.Б., Флеминг А.
С. (2007). Материнская изоляция изменяет экспрессию нейронных белков во время развития: стимуляция «поглаживанием» обращает эти эффекты вспять. Brain Res 1158 : 11–27. КАС пабмед Статья Google ученый
-
Coplan JD, Altemus M, Mathew SJ, Smith EL, Sharf B, Coplan PM и др. (2005). Синхронизированное повышение концентрации CRF CRF приматов у матери и ребенка в ответ на переменный спрос на корм. ЦНС Спектр 10 : 530–536.
ПабМед Статья Google ученый
-
Coplan JD, Smith EL, Altemus M, Mathew SJ, Perera T, Kral JG et al (2006). Реакция матери и ребенка на переменную потребность в пище у нечеловеческих приматов: влияние времени стрессора на фактор, высвобождающий кортикотропин, в спинномозговой жидкости и концентрации циркулирующих глюкокортикоидов. Ann NY Acad Sci 1071 : 525–533.
КАС пабмед Статья Google ученый
-
Керли Дж. П., Дэвидсон С., Бейтсон П., Шампань Ф.А. (2009). Социальное обогащение во время постнатального развития вызывает трансгенерационные эффекты на эмоциональное и репродуктивное поведение мышей. Front Behav Neurosci 3 : 25.
PubMed ПабМед Центральный Статья КАС Google ученый
-
Curley JP, Mashoodh R, Champagne FA (2011). Эпигенетика и происхождение отцовских эффектов. Horm Behav 59 : 306–314.
ПабМед Статья Google ученый
-
D’Addario C, Dell’Osso B, Palazzo MC, Benatti B, Lietti L, Cattaneo E и др. (2012). Селективное метилирование ДНК промотора BDNF при биполярном расстройстве: различия между пациентами с BDI и BDII. Нейропсихофармакология 37 : 1647–1655.
ПабМед ПабМед Центральный Статья КАС Google ученый
-
Дэвис М.
Н., Вольта М., Пидсли Р., Луннон К., Диксит А., Лавстон С. и др. (2012). Функциональная аннотация метилома головного мозга человека идентифицирует тканеспецифические эпигенетические вариации в мозге и крови. Геном Биол 13 : R43. КАС пабмед ПабМед Центральный Статья Google ученый
-
Davis TH, Cuellar TL, Koch SM, Barker AJ, Harfe BD, McManus MT et al (2008). Условная потеря Dicer нарушает морфогенез клеток и тканей в коре и гиппокампе. J Neurosci 28 : 4322–4330.
КАС пабмед ПабМед Центральный Статья Google ученый
-
Дасингер Л., Уайтлоу Э. (2012). Понимание трансгенерационного эпигенетического наследования через гаметы у млекопитающих. Nat Rev Genet 13 : 153–162.
КАС пабмед Статья Google ученый
-
Дэй Дж.
Дж., Суитт Дж. Д. (2010). Метилирование ДНК и формирование памяти. Nat Neurosci 13 : 1319–1323. КАС пабмед ПабМед Центральный Статья Google ученый
-
Де Беллис, доктор медицины (2005 г.). Психобиология пренебрежения. Жестокое обращение с детьми 10 : 150–172.
ПабМед Статья Google ученый
-
De Wolff MS, van Ijzendoorn MH (1997). Чувствительность и привязанность: метаанализ родительских предшественников детской привязанности. Ребенок Дев 68 : 571–591.
КАС пабмед Статья Google ученый
-
Деттмер А.М., Новак М.А., Суоми С.Дж., Мейер Дж.С. (2012). Физиологическая и поведенческая адаптация к стрессу при переезде у дифференцированно выращенных макак-резусов: кортизол в волосах как биомаркер реакций, связанных с тревогой.
Психонейроэндокринология 37 : 191–199. КАС пабмед Статья Google ученый
-
Эль-Маарри О, Карета М.С., Микеска Т., Беккер Т., Диас-Лакава А., Джунен Дж. и др. (2009). Систематический поиск полиморфизмов ДНК-метилтрансферазы выявляет редкий вариант DNMT3L, связанный с субтеломерным гипометилированием. Hum Mol Genet 18 : 1755–1768.
КАС пабмед Статья Google ученый
-
Энгерт В., Ефанов С.И., Дедович К., Дюшен А., Дагер А., Прюсснер Дж. К. (2010). Воспринимаемая материнская забота в раннем возрасте и реакция кортизола на повторяющийся психосоциальный стресс. J Psychiatry Neurosci 35 : 370–377.
ПабМед ПабМед Центральный Статья Google ученый
-
Фэрбенкс, Л.А., Макгуайр, М.Т. (1988). Долгосрочные эффекты раннего материнского поведения на реакцию мартышек на окружающую среду.
Dev Psychobiol 21 : 711–724. КАС пабмед Статья Google ученый
-
Фан Г., Хатник Л. (2005 г.). Белки, связывающие метил-CpG, в нервной системе. Cell Res 15 : 255–261.
КАС пабмед Статья Google ученый
-
Фрэнсис Д., Диорио Дж., Лю Д., Мини М.Дж. (1999). Негеномная передача через поколения материнского поведения и реакции на стресс у крыс. Наука 286 : 1155–1158.
КАС Статья пабмед Google ученый
-
Франклин Т.Б., Линдер Н., Рассиг Х., Тони Б., Мансуй И.М. (2011). Влияние раннего стресса на социальные способности и серотонинергические функции у мышей в разных поколениях. PLoS One 6 : e21842.
КАС пабмед ПабМед Центральный Статья Google ученый
-
Franklin TB, Russig H, Weiss IC, Graff J, Linder N, Michalon A et al (2010).
Эпигенетическая передача влияния раннего стресса из поколения в поколение. Биол Психиатрия 68 : 408–415. ПабМед Статья Google ученый
-
Frazier CR, Trainor BC, Cravens CJ, Whitney TK, Marler CA (2006). Отцовское поведение влияет на развитие агрессии и экспрессию вазопрессина у потомства самцов калифорнийских мышей. Horm Behav 50 : 699–707.
КАС пабмед Статья Google ученый
-
Fuchikami M, Morinobu S, Segawa M, Okamoto Y, Yamawaki S, Ozaki N et al (2011). Профили метилирования ДНК гена нейротрофического фактора головного мозга (BDNF) как мощного диагностического биомаркера при большой депрессии. PLoS One 6 : e23881.
КАС пабмед ПабМед Центральный Статья Google ученый
-
Гапп К., Джаваид А., Саркис П., Бохачек Дж.
, Пелчар П., Прадос Дж. и др. (2014). Роль РНК сперматозоидов в трансгенерационном наследовании последствий ранней травмы у мышей. Nat Neurosci 17 : 667–669. КАС пабмед ПабМед Центральный Статья Google ученый
-
Глисон Э.Д., Марлер К.А. (2013). Негеномная передача отцовского поведения между отцами и сыновьями у моногамных и двуродительских калифорнийских мышей. Proc Biol Sci 280 : 20130824.
PubMed ПабМед Центральный Статья Google ученый
-
Горман Дж. М., Мэтью С., Коплан Дж. (2002). Нейробиология стресса в раннем возрасте: модели приматов. Semin Clin Нейропсихиатрия 7 : 96–103.
ПабМед Статья Google ученый
-
Gunnar MR, Morison SJ, Chisholm K, Schuder M (2001). Уровни кортизола в слюне у детей, усыновленных из румынских детских домов.
Дев-психопат 13 : 611–628. КАС пабмед Статья Google ученый
-
Хане А.А., Фокс Н.А. (2006). Обычные различия в материнском уходе влияют на стрессовую реакцию младенцев. Psychol Sci 17 : 550–556.
ПабМед Статья Google ученый
-
Хейн А.А., Хендерсон Х.А., Риб-Сазерленд Б.К., Фокс Н.А. (2010). Обычные вариации материнской заботы человека в младенчестве и биоповеденческого развития в раннем детстве: последующее исследование. Dev Psychobiol 52 : 558–567.
ПабМед ПабМед Центральный Статья Google ученый
-
Harlow HF, Dodsworth RO, Harlow MK (1965). Полная социальная изоляция у обезьян. Proc Natl Acad Sci USA 54 : 90–97.
КАС пабмед Статья ПабМед Центральный Google ученый
-
Хейм С.
, Шугарт М., Крейгхед В.Е., Немерофф С.Б. (2010). Нейробиологические и психиатрические последствия жестокого обращения с детьми и безнадзорности. Dev Psychobiol 52 : 671–690. ПабМед Статья Google ученый
-
Hostinar CE, Stellern SA, Schaefer C, Carlson SM, Gunnar MR (2012). Ассоциации между невзгодами в раннем возрасте и исполнительной функцией у детей, усыновленных на международном уровне из детских домов. Proc Natl Acad Sci USA 109 (Приложение 2): 17208–17212.
КАС пабмед Статья ПабМед Центральный Google ученый
-
Howell BR, McCormack KM, Grand AP, Sawyer NT, Zhang X, Maestripieri D et al (2013). Изменения микроструктуры белого вещества головного мозга у макак-резусов-подростков, подвергшихся стрессу в раннем возрасте: связь с высоким уровнем кортизола в младенчестве. Biol Mood Anxiety Disord 3 : 21.
PubMed ПабМед Центральный Статья КАС Google ученый
-
Яковски А., Перера Т.Д., Абдаллах К.Г., Гарридо Г., Тан С.И., Мартинес Дж. и др. (2011). Стресс в раннем возрасте, развитие мозолистого тела, волюметрия гиппокампа и тревожное поведение у самцов нечеловеческих приматов. Psychiatry Res 192 : 37–44.
ПабМед ПабМед Центральный Статья Google ученый
-
Jackson-Grusby L, Beard C, Possemato R, Tudor M, Fambrough D, Csankovszki G et al (2001). Потеря геномного метилирования вызывает р53-зависимый апоптоз и эпигенетическую дерегуляцию. Nat Genet 27 : 31–39.
КАС пабмед Статья Google ученый
-
Jenuwein T, Allis CD (2001). Трансляция гистонового кода. Наука 293 : 1074–1080.
КАС пабмед Статья Google ученый
-
Цзя Р.
, Тай Ф., Ан С., Чжан С., Бродерс Х. (2009). Влияние неонатальной депривации отца или ранней депривации на тревожность и социальное поведение взрослых особей мандариновых полевок. Процессы поведения 82 : 271–278. ПабМед Статья Google ученый
-
Джонсон Дж.Г., Коэн П., Браун Дж., Смейлс Э.М., Бернштейн Д.П. (1999). Жестокое обращение в детстве увеличивает риск расстройств личности в раннем взрослом возрасте. Arch General Psychiatry 56 : 600–606.
КАС пабмед Статья Google ученый
-
Кембер Р.Л., Демпстер Э.Л., Ли Т.Х., Шалквик Л.С., Милл Дж., Фернандес С. (2012). Материнское отделение связано со специфическими для штамма ответами на стресс и эпигенетическими изменениями Nr3c1, Avp и Nr4a1 у мышей. Поведение мозга 2 : 455–467.
КАС пабмед ПабМед Центральный Статья Google ученый
-
Kinnally EL, Capitanio JP, Leibel R, Deng L, LeDuc C, Haghighi F и др.
(2010). Эпигенетическая регуляция экспрессии и поведения переносчика серотонина у детенышей макак-резусов. Гены, мозг и поведение 9 : 575–582. КАС Статья Google ученый
-
Koch CM, Andrews RM, Flicek P, Dillon SC, Karaoz U, Clelland GK et al (2007). Ландшафт модификаций гистонов в 1% генома человека в пяти клеточных линиях человека. Геном Res 17 : 691–707.
КАС пабмед ПабМед Центральный Статья Google ученый
-
Кох Х., Маккормак К., Санчес М.М., Маэстрипьери Д. (2014). Развитие гипоталамо-гипофизарно-надпочечниковой оси у макак-резусов: влияние возраста, пола и раннего опыта. Dev Psychobiol 56 : 86–95.
КАС пабмед Статья Google ученый
-
Кундакович М., Лим С., Гудснюк К., Шампань Ф.А. (2013). Влияние невзгод в раннем возрасте на поведенческие и эпигенетические результаты, зависящее от пола и напряжения.
Front Psychiatry 4 : 78. PubMed ПабМед Центральный Статья Google ученый
-
Куриан Дж. Р., Олесен К. М., Оже А. П. (2010). Половые различия в эпигенетической регуляции промотора альфа-рецептора эстрогена в развивающейся преоптической области. Эндокринология 151 : 2297–2305.
КАС пабмед ПабМед Центральный Статья Google ученый
-
Лабонте Б., Судерман М., Мауссион Г., Наваро Л., Йерко В., Махар I и др. (2012). Полногеномная эпигенетическая регуляция ранними травмами. Arch General Psychiatry 69 : 722–731.
КАС пабмед ПабМед Центральный Статья Google ученый
-
Lee MHS, Williams DI (1974). Изменения в облизывающем поведении крысиной матери после обращения с детенышами. Anim Behav 22 : 679–681.
Артикул Google ученый
-
Леманн Дж., Прайс К.Р., Беттшен Д., Фелдон Дж. (1999). Парадигма материнского разделения и взрослая эмоциональность и познание у самцов и самок крыс Wistar. Pharmacol Biochem Behav 64 : 705–715.
КАС пабмед Статья Google ученый
-
Леви Ф., Мело А.И., Галеф Б.Г. мл., Мэдден М., Флеминг А.С. (2003). Полная материнская депривация влияет на социальное, но не пространственное обучение взрослых крыс. Dev Psychobiol 43 : 177–191.
КАС пабмед Статья Google ученый
-
Лю Д., Диорио Дж., Дэй Дж.С., Фрэнсис Д.Д., Мини М.Дж. (2000). Материнская забота, синаптогенез гиппокампа и когнитивное развитие у крыс. Nat Neurosci 3 : 799–806.
КАС пабмед Статья Google ученый
-
Liu D, Diorio J, Tannenbaum B, Caldji C, Francis D, Freedman A et al (1997).
Материнская забота, глюкокортикоидные рецепторы гиппокампа и реакции гипоталамо-гипофизарно-надпочечниковой системы на стресс. Наука 277 : 1659–1662. КАС пабмед Статья Google ученый
-
Лович В., Флеминг А.С. (2004). Искусственно выращенные самки крыс демонстрируют пониженное торможение предимпульса и дефицит в задаче переключения внимания — изменение эффектов при материнской стимуляции лизания. Behav Brain Res 148 : 209–219.
ПабМед Статья Google ученый
-
Лович В., Кин Д., Флетчер П.Дж., Флеминг А.С. (2011). Разлука с матерью в раннем возрасте и социальная изоляция вызывают рост импульсивных действий, но не импульсивного выбора. Behav Neurosci 125 : 481–491.
ПабМед Статья Google ученый
-
Маэстрипьери Д., Хигли Дж.Д.
, Линделл С.Г., Ньюман Т.К., МакКормак К.М., Санчес М.М. (2006). Раннее неприятие матери влияет на развитие моноаминергических систем и жестокое воспитание во взрослом возрасте у макак-резусов (Macaca mulatta). Behav Neurosci 120 : 1017–1024. ПабМед Статья Google ученый
-
Maheu FS, Dozier M, Guyer AE, Mandell D, Peloso E, Poeth K et al (2010). Предварительное исследование функции медиальной височной доли у молодых людей с историей лишения опекуна и эмоционального пренебрежения. Cogn Affect Behav Neurosci 10 : 34–49.
ПабМед ПабМед Центральный Статья Google ученый
-
Маккормак К., Санчес М.М., Барди М., Маэстрипьери Д. (2006). Образцы ухода за матерью и поведенческое развитие младенцев макак-резусов, подвергшихся жестокому обращению, в первые 6 месяцев жизни. Dev Psychobiol 48 : 537–550.
КАС пабмед Статья Google ученый
-
McGoron L, Gleason MM, Smyke AT, Drury SS, Nelson CA 3rd, Gregas MC et al (2012). Восстановление после ранней депривации: привязанность опосредует влияние заботы на психопатологию. J Am Acad Детская подростковая психиатрия 51 : 683–693.
ПабМед ПабМед Центральный Статья Google ученый
-
McGowan PO, Sasaki A, D’Alessio AC, Dymov S, Labonte B, Szyf M et al (2009). Эпигенетическая регуляция глюкокортикоидного рецептора в мозге человека связана с жестоким обращением в детстве. Nat Neurosci 12 : 342–348.
КАС пабмед ПабМед Центральный Статья Google ученый
-
McGowan PO, Suderman M, Sasaki A, Huang TC, Hallett M, Meaney MJ et al (2011). Широкая эпигенетическая подпись материнской заботы в мозгу взрослых крыс.
PLoS One 6 : e14739. КАС пабмед ПабМед Центральный Статья Google ученый
-
Мини М.Дж., Эйткен Д.Х., Боднофф С.Р., Ини Л.Дж., Татаревич Дж.Е., Сапольский Р.М. (1985). Раннее постнатальное обращение изменяет концентрацию глюкокортикоидных рецепторов в отдельных областях мозга. Behav Neurosci 99 : 765–770.
КАС Статья пабмед Google ученый
-
Morgan CP, Bale TL (2011). Ранний пренатальный стресс эпигенетически программирует дисмаскулинизацию у потомства во втором поколении по отцовской линии. J Neurosci 31 : 11748–11755.
КАС пабмед ПабМед Центральный Статья Google ученый
-
Murgatroyd C, Patchev AV, Wu Y, Micale V, Bockmuhl Y, Fischer D et al (2009). Динамическое метилирование ДНК программирует стойкие неблагоприятные последствия стресса в раннем возрасте.
Nat Neurosci 12 : 1559–1566. КАС Статья пабмед Google ученый
-
Нарита К., Фуджихара К., Такей Ю., Суда М., Аояма Ю., Уэхара Т. и др. (2012). Связь между родительским опытом в детстве и подростковом возрасте, гипоталамо-гипофизарно-надпочечниковой гипоактивностью и уменьшением объема серого вещества гиппокампа у молодых людей. Hum Brain Map 33 : 2211–2223.
ПабМед Статья Google ученый
-
Нарита К., Такей Ю., Суда М., Аояма Ю., Уэхара Т., Косака Х. и др. (2010). Связь стилей родительской привязанности с объемом серого вещества дорсолатеральной префронтальной коры у молодых людей. Прог Нейропсихофармакол Биол Психиатрия .
-
Наумова О.Ю., Ли М., Копосов Р., Шиф М., Дозье М., Григоренко Е.Л. (2012). Дифференциальные паттерны метилирования полногеномной ДНК у детей, находящихся в специализированных учреждениях, и детей, воспитанных их биологическими родителями.
Дев-психопат 24 : 143–155. ПабМед Статья Google ученый
-
О’Коннор Т.Г., Раттер М., Беккет С., Кивени Л., Креппнер Дж.М. (2000). Влияние глобальных тяжелых лишений на когнитивную компетентность: расширение и лонгитюдное наблюдение. Группа по изучению усыновленных детей на английском и румынском языках. Ребенок Дев 71 : 376–390.
ПабМед Статья Google ученый
-
Окано М., Белл Д.В., Хабер Д.А., Ли Э. (1999). ДНК-метилтрансферазы Dnmt3a и Dnmt3b необходимы для метилирования de novo и развития млекопитающих. Сотовый 99 : 247–257.
КАС пабмед Статья Google ученый
-
Паркер Г. (1990). Инструмент родительской привязанности: десятилетие исследований. Soc Psychiatry Psychiatr Epidemiol 25 : 281–282.
КАС пабмед Статья Google ученый
-
Паули А.
, Ринн Дж.Л., Шир А.Ф. (2011). Некодирующие РНК как регуляторы эмбриогенеза. Nat Rev Genet 12 : 136–149. КАС пабмед ПабМед Центральный Статья Google ученый
-
Pena CJ, Neugut YD, Champagne FA (2013). Сроки развития влияния материнской заботы на экспрессию генов и эпигенетическую регуляцию уровней гормональных рецепторов у самок крыс. Эндокринология 154 : 4340–4351.
КАС пабмед ПабМед Центральный Статья Google ученый
-
Perroud N, Dayer A, Piguet C, Nallet A, Favre S, Malafosse A et al (2014). Жестокое обращение в детстве и метилирование гена глюкокортикоидного рецептора NR3C1 при биполярном расстройстве. Br J Психиатрия 204 : 30–35.
ПабМед Статья Google ученый
-
Peterson CL, Laniel MA (2004). Гистоны и модификации гистонов.
Карр Биол 14 : R546–R551. КАС пабмед Статья Google ученый
-
Пинкернелл Дж., Абрахам А., Зайдель К., Браун К. (2009). Отцовская депривация вызывает дендритные и синаптические изменения и полушарную асимметрию пирамидных нейронов в соматосенсорной коре. Дев Нейробиол 69 : 663–673.
ПабМед Статья Google ученый
-
Provencal N, Suderman MJ, Guillemin C, Massart R, Ruggiero A, Wang D et al (2012). Признак материнского воспитания в метиломе префронтальной коры макаки-резуса и Т-клетках. J Neurosci 32 : 15626–15642.
КАС пабмед ПабМед Центральный Статья Google ученый
-
Pruessner JC, Champagne F, Meaney MJ, Dagher A (2004). Высвобождение дофамина в ответ на психологический стресс у людей и его связь с уходом за матерью в раннем возрасте: исследование позитронно-эмиссионной томографии с использованием [11C]раклоприда.
J Neurosci 24 : 2825–2831. КАС пабмед ПабМед Центральный Статья Google ученый
-
Разин А (1998). Метилирование CpG, структура хроматина и сайленсинг генов — трехсторонняя связь. EMBO J 17 : 4905–4908.
КАС пабмед ПабМед Центральный Статья Google ученый
-
Rice CJ, Sandman CA, Lenjavi MR, Baram TZ (2008). Новая мышиная модель острых и длительных последствий стресса в раннем возрасте. Эндокринология 149 : 4892–4900.
КАС пабмед ПабМед Центральный Статья Google ученый
-
Роденас-Руано А., Чавес А.Е., Коссио М.Дж., Кастильо П.Е., Зукин Р.С. (2012). REST-зависимое эпигенетическое ремоделирование способствует онтогенетическим переключениям синаптических рецепторов NMDA. Nat Neurosci 15 : 1382–1390.
КАС пабмед ПабМед Центральный Статья Google ученый
-
Рот Т.Л., Любин Ф.Д., Функ А.Дж., Суитт Д.Д. (2009). Длительное эпигенетическое влияние невзгод в раннем возрасте на ген BDNF. Биол Психиатрия 65 : 760–769.
КАС пабмед ПабМед Центральный Статья Google ученый
-
Раттер М., О’Коннор Т.Г. (2004). Существуют ли эффекты биологического программирования для психологического развития? Результаты исследования румынских приемных детей. Dev Psychol 40 : 81–94.
ПабМед Статья Google ученый
-
Сааведра-Родригес Л., Фейг Л.А. (2013). Хроническая социальная нестабильность вызывает беспокойство и неполноценные социальные взаимодействия между поколениями. Биол Психиатрия 73 : 44–53.
ПабМед Статья Google ученый
-
Санчес М.
М., Маккормак К., Гранд А.П., Фулкс Р., Графф А., Маэстрипьери Д. (2010). Влияние секса и жестокого обращения с матерью в раннем возрасте на реакцию адренокортикотропного гормона и кортизола на введение кортикотропин-высвобождающего гормона в течение первых 3 лет жизни у макак-резусов, живущих в группах. Дев-психопат 22 : 45–53. ПабМед ПабМед Центральный Статья Google ученый
-
Сапольски Р.М., Мини М.Дж., Макьюэн Б.С. (1985). Развитие системы глюкокортикоидных рецепторов в лимбическом мозге крыс. III. Регулирование с отрицательной обратной связью. Мозг Res 350 : 169–173.
КАС пабмед Статья Google ученый
-
Сато Ф., Цутия С., Мельцер С.Дж., Симидзу К. (2011). МикроРНК и эпигенетика. FEBS J 278 : 1598–1609.
КАС Статья пабмед Google ученый
-
Шмидт М.
В. (2011). Животные модели депрессии и гипотеза несоответствия болезни. Психонейроэндокринология 36 : 330–338. ПабМед Статья Google ученый
-
Зайдель К., Поегель Г., Холечка Р., Хельмеке К., Браун К. (2011). Отцовская депривация влияет на развитие экспрессирующих кортикотропин-рилизинг-фактор нейронов в префронтальной коре, миндалевидном теле и гиппокампе двухродительского Octodon degus. J Нейроэндокринол 23 : 1166–1176.
КАС пабмед Статья Google ученый
-
Смайк А.Т., Думитреску А., Зеана К.Х. (2002). Нарушения привязанности у детей раннего возраста. I: континуум ухода за пострадавшим. J Am Acad Детская подростковая психиатрия 41 : 972–982.
ПабМед Статья Google ученый
-
Spinelli S, Chefer S, Carson RE, Jagoda E, Lang L, Heilig M et al (2010).
Влияние стресса в раннем возрасте на рецепторы серотонина (1А) у молодых макак-резус, измеренное с помощью позитронно-эмиссионной томографии. Биол Психиатрия 67 : 1146–1153. КАС пабмед ПабМед Центральный Статья Google ученый
-
Спинелли С., Чефер С., Суоми С.Дж., Хигли Дж.Д., Барр С.С., Штейн Э. (2009). Стресс в раннем возрасте вызывает долгосрочные морфологические изменения в мозге приматов. Arch General Psychiatry 66 : 658–665.
ПабМед Статья Google ученый
-
Сроуф Л.А. (2005). Привязанность и развитие: проспективное лонгитюдное исследование от рождения до взрослой жизни. Прикрепить Hum Dev 7 : 349–367.
ПабМед Статья Google ученый
-
Стэнтон М.Е., Гутьеррес Ю.Р., Левин С. (1988). Материнская депривация потенцирует стрессовые реакции гипофиза и надпочечников у крысят.
Behav Neurosci 102 : 692–700. КАС пабмед Статья Google ученый
-
Suderman M, Borghol N, Pappas JJ, Pinto Pereira SM, Pembrey M, Hertzman C и др. (2014). Жестокое обращение в детстве связано с метилированием множества локусов в ДНК взрослых. BMC Med Genomics 7 : 13.
PubMed ПабМед Центральный Статья КАС Google ученый
-
Suomi SJ (1991). Ранний стресс и взрослая эмоциональная реактивность у макак-резусов. Ciba Found Symp 156 : 171–183.
КАС пабмед Google ученый
-
Suomi SJ, Harlow HF, Kimball SD (1971). Поведенческие эффекты длительной частичной социальной изоляции у макак-резусов. Psychol Rep 29 : 1171–1177.
КАС пабмед Статья Google ученый
-
Сури Д.
, Винит В., Саркар А., Тиагараджан Д., Кумар А., Нестлер Э.Дж. и др. (2013). Ранний стресс вызывает возрастные двухфазные изменения нейрогенеза гиппокампа, экспрессии BDNF и когнитивных функций. Биол Психиатрия 73 : 658–666. КАС пабмед Статья Google ученый
-
Тахилиани М., Мэй П., Фанг Р., Леонор Т., Рутенберг М., Симидзу Ф. и др. (2007). Деметилаза SMCX гистона h4K4 связывает гены-мишени REST с Х-сцепленной умственной отсталостью. Природа 447 : 601–605.
КАС Статья пабмед Google ученый
-
Тан AC (2001). Неонатальное воздействие новой окружающей среды усиливает функцию памяти, зависящую от гиппокампа, в младенчестве и во взрослом возрасте. Узнать память 8 : 257–264.
КАС пабмед ПабМед Центральный Статья Google ученый
-
Танг А.
С., Акерс К.Г., Риб Б.К., Ромео Р.Д., Макьюэн Б.С. (2006). Программирование социального, когнитивного и нейроэндокринного развития путем раннего знакомства с новизной. Proc Natl Acad Sci USA 103 : 15716–15721. КАС пабмед Статья ПабМед Центральный Google ученый
-
Тейлор С.М., Джонс П.А. (1985). Клеточная дифференцировка. Int J Obes 9 (дополнение 1): 15–21.
КАС пабмед Google ученый
-
Тода Х., Боку С., Накагава С., Иноуэ Т., Като А., Такамура Н. и др. (2014). Разлучение с матерью усиливает условный страх и снижает уровень мРНК гена рецептора нейротензина 1 с гиперметилированием этого гена в миндалевидном теле крысы. PLoS One 9 : e97421.
ПабМед ПабМед Центральный Статья КАС Google ученый
-
Тоттенхэм Н.
, Хэйр Т.А., Миллнер А., Гилхули Т., Зевин Д.Д., Кейси Б.Дж. (2011). Повышенная реакция миндалевидного тела на лица после ранней депривации. Dev Sci 14 : 190–204. КАС пабмед ПабМед Центральный Статья Google ученый
-
Трикетт П., Макбрайд-Чанг С. (1995). Влияние на развитие различных форм жестокого обращения с детьми и безнадзорности. Devel Rev 15 : 11–37.
Google ученый
-
Tyrka AR, Price LH, Marsit C, Walters OC, Carpenter LL (2012). Невзгоды детства и эпигенетическая модуляция глюкокортикоидного рецептора лейкоцитов: предварительные данные у здоровых взрослых. PLoS One 7 : e30148.
КАС пабмед ПабМед Центральный Статья Google ученый
-
Уэр Р., Уивер И.С. (2014). Эпигенетические следы жестокого обращения в детстве в периферической крови: новая стратегия изучения взаимодействия генов и окружающей среды.
Br J Психиатрия 204 : 3–5. ПабМед Статья Google ученый
-
ван Хармелен А.Л., ван Тол М.Дж., ван дер Ви Н.Дж., Вельтман Д.Дж., Алеман А., Спинховен П. и др. (2010). Уменьшение объема медиальной префронтальной коры у взрослых, сообщивших о жестоком эмоциональном обращении в детстве. Биол Психиатрия 68 : 832–838.
ПабМед Статья Google ученый
-
ван Айзендорн М.Х., Шуенгель С., Бейкерманс-Краненбург М.Ю. (1999). Дезорганизованная привязанность в раннем детстве: метаанализ предшественников, сопутствующих факторов и последствий. Дев-психопат 11 : 225–249.
КАС пабмед Статья Google ученый
-
Ван А, Не В, Ли Х, Хоу Ю, Ю З, Фан Кью и др. (2014). Эпигенетическая активация кортикотропин-рилизинг-гормона опосредует постнатальный дефицит памяти, вызванный разлукой с матерью.
PLoS One 9 : e. ПабМед ПабМед Центральный Статья КАС Google ученый
-
Ван Дж., Тай Ф., Ян С., Ю. П. (2012). Отцовская депривация изменяет игровые бои, уровень кортикостерона в сыворотке и экспрессию гипоталамического вазопрессина и окситоцина у молодых самцов мандариновых полевок. J Comp Physiol A Neuroethol Sens Neural Behav Physiol 198 : 787–796.
КАС пабмед Статья Google ученый
-
Wang XD, Rammes G, Kraev I, Wolf M, Liebl C, Scharf SH и др. (2011). CRF(1) переднего мозга модулирует запрограммированный стрессом когнитивный дефицит в раннем возрасте. J Neurosci 31 : 13625–13634.
КАС пабмед ПабМед Центральный Статья Google ученый
-
Weaver IC, Cervoni N, Champagne FA, D’Alessio AC, Sharma S, Seckl JR et al (2004).
Эпигенетическое программирование материнским поведением. Nat Neurosci 7 : 847–854. КАС Статья пабмед Google ученый
-
Weaver IC, Meaney MJ, Szyf M (2006). Влияние материнской заботы на транскриптом гиппокампа и поведение, опосредованное тревогой, у потомства обратимо во взрослом возрасте. Proc Natl Acad Sci USA 103 : 3480–3485.
КАС пабмед ПабМед Центральный Статья Google ученый
-
Ву Ю, Патчев А.В., Даниэль Г., Алмейда О.Ф., Шпенглер Д. (2014). Стресс в раннем возрасте снижает метилирование ДНК гена pomc у самцов мышей. Эндокринология 155 : 1751–1762.
ПабМед Статья КАС Google ученый
-
Yu P, An S, Tai F, Zhang X, He F, Wang J et al (2012). Влияние неонатальной депривации отца на образование пар, экспрессию мРНК дофаминового рецептора NAcc и уровень кортикостерона в сыворотке у мандариновых полевок.
Horm Behav 61 : 669–677. КАС пабмед Статья Google ученый
-
Чжан TY, Hellstrom IC, Bagot RC, Wen X, Diorio J, Meaney MJ (2010). Материнский уход и метилирование ДНК промотора декарбоксилазы 1 глутаминовой кислоты в гиппокампе крысы. J Neurosci 30 : 13130–13137.
КАС пабмед ПабМед Центральный Статья Google ученый
Ссылки на скачивание
Благодарности
Это исследование поддерживается финансированием NIMH 1P50MH0
-01A1.
Информация о авторе
Авторы и принадлежности
-
Департамент психологии, Колумбийский университет, Нью -Йорк, Нью -Йорк, США
Marija Kundakovic & Frances A Champagne
9105
9995aknaknaknaknaknaknaknaknaknaknaknaknaknejavics 9105. Вы также можете искать этого автора в
PubMed Google Академия
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
Автор, ответственный за корреспонденцию
Фрэнсис Шампанское.
Слайды PowerPoint
Слайд PowerPoint для рис. 1
Слайд PowerPoint для рис. 2
Слайд PowerPoint для рис.0057
Слайд PowerPoint для рис. 5
Права и разрешения
Перепечатка и разрешения
Об этой статье
Ненадежность материнской привязанности, жестокое обращение в анамнезе и депрессивные симптомы связаны с широкими признаками метилирования ДНК у младенцев.
- Талия К. Робакис
- Марисса С. Рот
- Ян Х. Готлиб
Молекулярная психиатрия (2022)
Типоспецифические эпигенетические эффекты стресса в раннем возрасте на мозг
- Мули Ф. Рахман
- Патрик О. Макгоуэн
Трансляционная психиатрия (2022)
Семейный контекст и симптомы СДВГ в среднем детстве: объяснительная модель
- Флоренсия Баррето-Сарса
- Мануэль Санчес де Мигель
- Хесус Ибарлузеа
Журнал детских и семейных исследований (2022)
Frühe Hilfen: Был ли müssen Kinderärzte wissen?
- Сабина Хаас
- Ульрике Мецгер
- Мехтильд Пауль
Monatsschrift Kinderheilkunde (2022)
Жесткое воспитание предсказывает метилирование нового гена рецептора HPA, а метилирование NR3C1 предсказывает ежедневный наклон уровня кортизола в среднем детстве
- Кэндес Р. Льюис
- Рейган С. Брайтенштейн
- Кэтрин Лемери-Чалфант
Клеточная и молекулярная нейробиология (2021)
Эпигенетика | Психология Вики | Фэндом
Оценка |
Биопсихология |
Сравнительный |
Познавательный |
Развивающие |
Язык |
Индивидуальные различия |
Личность |
Философия |
Социальные |
Методы |
Статистика |
Клинический |
Образовательные |
промышленный |
Профессиональные товары |
Мировая психология |
Биологический: Поведенческая генетика · Эволюционная психология · Нейроанатомия · Нейрохимия · Нейроэндокринология · Неврология · Психонейроиммунология · Физиологическая психология · Психофармакология (Индекс, Структура)
. Пожалуйста, помогите улучшить эту страницу самостоятельно, если можете.
В биологии, и особенно в генетике, эпигенетика — это изучение наследственных изменений в экспрессии генов или клеточного фенотипа, вызванных механизмами, отличными от изменений в основной последовательности ДНК — отсюда и название эпи- (греч. επί — над, над, наружным) — генетика . Это относится к функционально значимым модификациям генома, не связанным с изменением последовательности нуклеотидов. Примерами таких изменений являются метилирование ДНК и модификация гистонов, которые служат для регуляции экспрессии генов без изменения основной последовательности ДНК. Убедительные доказательства, подтверждающие эпигенетику, показывают, что эти механизмы могут способствовать передаче эффектов родительского опыта последующим поколениям.
Эти изменения могут сохраняться при клеточных делениях до конца жизни клетки, а также могут сохраняться в течение нескольких поколений. Однако в основной последовательности ДНК организма нет изменений; [1] вместо этого негенетические факторы заставляют гены организма вести себя (или «выражать себя») по-разному. [2]
Одним из примеров эпигенетических изменений в биологии эукариот является процесс клеточной дифференцировки. Во время морфогенеза тотипотентные стволовые клетки становятся различными плюрипотентными клеточными линиями эмбриона, которые, в свою очередь, становятся полностью дифференцированными клетками. Другими словами, одна оплодотворенная яйцеклетка — зигота — превращается во множество типов клеток, включая нейроны, мышечные клетки, эпителий, эндотелий кровеносных сосудов и т. д., поскольку она продолжает делиться. Он делает это, активируя одни гены и подавляя другие. [3]
В 2011 году было показано, что метилирование мРНК играет критическую роль в энергетическом гомеостазе человека. Показано, что связанный с ожирением ген FTO способен деметилировать N6-метиладенозин в РНК. Это открыло смежную область эпигенетики РНК. [4] [5]
Содержание
- 1 Этимология и определения
- 2 Молекулярные основы эпигенетики
- 3 Механизма
- 3.1 Метилирование ДНК и ремоделирование хроматина
- 3.2 Транскрипты РНК и кодируемые ими белки
- 3.3 Прионы
- 3.4 Системы структурного наследования
- 4 Функции и последствия
- 4.1 Развитие
- 4.2 Медицина
- 4.3 Эволюция
- 5 Эпигенетические эффекты у человека
- 5.1 Геномный импринтинг и родственные расстройства
- 5.2 Трансгенерационные эпигенетические наблюдения
- 5.3 Исследования близнецов
- 6 См. также
- 7 Внешние ссылки
- 8 Каталожные номера
Этимология и определения
Файл:Эпигенетические механизмы.jpgЭпигенетические механизмы
Эпигенетика (как в «эпигенетическом ландшафте») была придумана Ч. Х. Уоддингтоном в 1942 году как сочетание слов эпигенезис и 90.010. [6] Эпигенез — это старое [7] слово, которое использовалось совсем недавно (см. preformationism для исторической справки) для описания дифференциации клеток от их начального тотипотентного состояния в эмбриональном развитии. Когда Уоддингтон ввел этот термин, физическая природа генов и их роль в наследственности еще не были известны; он использовал ее как концептуальную модель того, как гены могут взаимодействовать со своим окружением, создавая фенотип.
Робин Холлидей определил эпигенетику как «изучение механизмов временного и пространственного контроля активности генов во время развития сложных организмов». [8] Таким образом, эпигенетический может использоваться для описания всего, кроме последовательности ДНК, которая влияет на развитие организма.
Современное использование этого слова в научном дискурсе является более узким, относящимся к наследственным признакам (в ходе клеточного деления, а иногда и из поколения в поколение), которые не связаны с изменениями в основной последовательности ДНК. [9] Греческий префикс эпи- в эпигенетика подразумевает признаки, которые находятся «поверх» или «в дополнение» к генетике; таким образом эпигенетических признаков существуют поверх или в дополнение к традиционной молекулярной основе наследования.
Сходство слова «генетика» породило множество параллельных употреблений. «Эпигеном» аналогичен слову «геном» и относится к общему эпигенетическому состоянию клетки. Фраза «генетический код» также была адаптирована — «эпигенетический код» использовался для описания набора эпигенетических признаков, которые создают разные фенотипы в разных клетках. Доведенный до крайности, «эпигенетический код» мог бы представлять общее состояние клетки, с положением каждой молекулы в эпигеномная карта , схематическое изображение экспрессии генов, метилирования ДНК и статуса модификации гистонов конкретной области генома. Чаще всего этот термин используется в отношении систематических усилий по измерению конкретных, релевантных форм эпигенетической информации, таких как гистоновый код или паттерны метилирования ДНК.
Психолог Эрик Эриксон использовал термин эпигенетический в своей книге Идентичность: Молодость и кризис (1968). Эриксон пишет, что эпигенетический принцип заключается в том, что «все, что растет, имеет основной план, и что из этого основного плана возникают части, каждая часть имеет свое время особого господства, пока все части не возникнут, чтобы сформировать функционирующее целое». [10] Однако такое употребление представляет прежде всего исторический интерес. [11]
Молекулярные основы эпигенетики
Эпигенетические изменения могут модифицировать активацию определенных генов, но не последовательность ДНК. Кроме того, белки хроматина, связанные с ДНК, могут быть активированы или заглушены. Вот почему дифференцированные клетки многоклеточного организма экспрессируют только те гены, которые необходимы для их собственной активности. Эпигенетические изменения сохраняются при делении клеток. Большинство эпигенетических изменений происходят только в течение жизни одного отдельного организма, но, если мутация в ДНК была вызвана в сперме или яйцеклетке, что приводит к оплодотворению, то некоторые эпигенетические изменения наследуются от одного поколения к другому. [12] Это поднимает вопрос о том, могут ли эпигенетические изменения в организме изменить основную структуру его ДНК (см. Эволюция ниже), форма ламаркизма.
Специфические эпигенетические процессы включают парамутацию, закладку, импринтинг, замалчивание генов, инактивацию Х-хромосомы, эффект положения, перепрограммирование, трансвекцию, материнские эффекты, развитие канцерогенеза, многие эффекты тератогенов, регуляцию модификаций гистонов и гетерохроматина, а также технические ограничения, влияющие на партеногенез и клонирование.
В эпигенетических исследованиях используется широкий спектр молекулярно-биологических методов для дальнейшего понимания эпигенетических явлений, включая иммунопреципитацию хроматина (вместе с ее крупномасштабными вариантами ChIP-on-chip и ChIP-Seq), флуоресцентную гибридизацию in situ, чувствительную к метилированию ферменты рестрикции, идентификация ДНК-аденинметилтрансферазы (DamID) и бисульфитное секвенирование. Кроме того, все большую роль играет использование биоинформационных методов (вычислительная эпигенетика).
Механизмы
Несколько типов эпигенетических систем наследования могут играть роль в так называемой клеточной памяти: [13]
Метилирование ДНК и ремоделирование хроматина
ДНК связывается с гистоновыми белками с образованием хроматина.
Поскольку фенотип клетки или индивидуума зависит от того, какие из ее генов транскрибируются, наследуемые состояния транскрипции могут вызывать эпигенетические эффекты. Существует несколько уровней регуляции экспрессии генов. Одним из способов регуляции генов является ремоделирование хроматина. Хроматин представляет собой комплекс ДНК и гистоновых белков, с которыми он связан. Белки-гистоны — это маленькие сферы, вокруг которых обернута ДНК. Если способ, которым ДНК оборачивается вокруг гистонов, изменится, экспрессия генов также может измениться. Ремоделирование хроматина осуществляется посредством двух основных механизмов:
- Первый способ — это посттрансляционная модификация аминокислот, входящих в состав гистоновых белков. Гистоновые белки состоят из длинных цепочек аминокислот. Если аминокислоты в цепи изменены, форма гистоновой сферы может измениться. ДНК полностью не раскручивается во время репликации. Таким образом, возможно, что модифицированные гистоны могут быть перенесены в каждую новую копию ДНК. Оказавшись там, эти гистоны могут действовать как шаблоны, инициируя формирование окружающих новых гистонов по-новому. Изменяя форму гистонов вокруг себя, эти модифицированные гистоны гарантируют, что дифференцированная клетка останется дифференцированной и не превратится обратно в стволовую клетку.
- Второй способ заключается в добавлении метильных групп к ДНК, в основном в сайтах CpG, для превращения цитозина в 5-метилцитозин. 5-Метилцитозин работает так же, как обычный цитозин, в паре с гуанином. Однако некоторые области генома метилированы более сильно, чем другие, а сильно метилированные области имеют тенденцию быть менее транскрипционно активными из-за механизма, который до конца не изучен. Метилирование цитозинов также может сохраняться из зародышевой линии одного из родителей в зиготу, отмечая хромосому как унаследованную от этого родителя (генетический импринтинг).
То, как клетки остаются дифференцированными в случае метилирования ДНК, нам более ясно, чем в случае гистоновой формы. В основном, некоторые ферменты (например, DNMT1) имеют более высокое сродство к метилированному цитозину. Если этот фермент достигает «гемиметилированной» части ДНК (где метилцитозин находится только в одной из двух цепей ДНК), фермент метилирует другую половину.
Хотя модификации гистонов происходят по всей последовательности, неструктурированные N-концы гистонов (называемые гистоновыми хвостами) особенно сильно модифицируются. Эти модификации включают ацетилирование, метилирование, убиквитилирование, фосфорилирование и сумоилирование. Ацетилирование является наиболее изученной из этих модификаций. Например, ацетилирование K14 и K9лизин хвоста гистона h4 ферментами гистон-ацетилтрансфераз (HATs) обычно коррелирует с транскрипционной компетентностью.
Один из способов мышления состоит в том, что эта тенденция ацетилирования, связанная с «активной» транскрипцией, имеет биофизическую природу. Поскольку в норме лизин имеет положительно заряженный азот на конце, он может связывать отрицательно заряженные фосфаты остова ДНК. Событие ацетилирования превращает положительно заряженную аминогруппу на боковой цепи в нейтральную амидную связь. Это удаляет положительный заряд, тем самым отделяя ДНК от гистона. Когда это происходит, такие комплексы, как SWI/SNF и другие транскрипционные факторы, могут связываться с ДНК и обеспечивать возможность транскрипции. Это «цис» модель эпигенетической функции. Другими словами, изменения гистоновых хвостов напрямую влияют на саму ДНК.
Другой моделью эпигенетической функции является «транс» модель. В этой модели изменения гистоновых хвостов косвенно воздействуют на ДНК. Например, ацетилирование лизина может создать сайт связывания ферментов, модифицирующих хроматин (а также базальный аппарат транскрипции). Затем этот моделлер хроматина может вызывать изменения в состоянии хроматина. Действительно, бромодомен — белковый сегмент (домен), специфически связывающий ацетиллизин, — обнаружен во многих ферментах, помогающих активировать транскрипцию, в том числе в комплексе SWI/SNF (на белке полибромо). Возможно, что ацетилирование действует таким и предыдущим способом, помогая активации транскрипции.
Идея о том, что модификации действуют как стыковочные модули для родственных факторов, также подтверждается метилированием гистонов. Метилирование лизина 9 гистона h4 долгое время ассоциировалось с конститутивно транскрипционно молчащим хроматином (конститутивным гетерохроматином). Было установлено, что хромодомен (домен, который специфически связывает метиллизин) в транскрипционно-репрессивном белке HP1 рекрутирует HP1 в метилированные области K9. Одним из примеров, который, по-видимому, опровергает эту биофизическую модель ацетилирования, является то, что триметилирование гистона h4 по лизину 4 сильно связано (и необходимо для полной) активации транскрипции. Триметилирование в этом случае внесло бы фиксированный положительный заряд на хвост.
Было показано, что гистон-лизин-метилтрансфераза (КМТ) отвечает за эту активность метилирования в структуре гистонов h4 и h5. Этот фермент использует каталитически активный сайт, называемый доменом SET (подавитель пестроты, усилитель пикантности, Trithorax). Домен SET представляет собой последовательность из 130 аминокислот, участвующую в модуляции активности генов. Было показано, что этот домен связывается с гистоновым хвостом и вызывает метилирование гистона. [14]
Различные модификации гистонов, вероятно, функционируют по-разному; ацетилирование в одном положении, вероятно, действует иначе, чем ацетилирование в другом положении. Кроме того, одновременно могут происходить множественные модификации, и эти модификации могут работать вместе, чтобы изменить поведение нуклеосомы. Идея о том, что множественные динамические модификации регулируют транскрипцию генов систематическим и воспроизводимым образом, называется гистоновым кодом.
Метилирование ДНК часто происходит в повторяющихся последовательностях и помогает подавить экспрессию и подвижность «мобильных элементов»: [15] Поскольку 5-метилцитозин может спонтанно дезаминироваться (замещая азот кислородом) до тимидина, сайты CpG часто мутируют и становятся редкими в геноме, за исключением островков CpG, где они остаются неметилированными. Таким образом, эпигенетические изменения этого типа могут направлять увеличение частоты перманентных генетических мутаций. Известно, что паттерны метилирования ДНК устанавливаются и модифицируются в ответ на факторы окружающей среды в результате сложного взаимодействия по крайней мере трех независимых ДНК-метилтрансфераз, DNMT1, DNMT3A и DNMT3B, потеря любой из которых приводит к летальному исходу у мышей. [16] DNMT1 является наиболее распространенной метилтрансферазой в соматических клетках, [17] локализуется в очагах репликации, [18] имеет 10-40-кратное предпочтение гемиметилированной ДНК и взаимодействует с ядерным антигеном пролиферирующих клеток ( ПКНА). [19]
Преимущественно модифицируя гемиметилированную ДНК, DNMT1 переносит образцы метилирования на вновь синтезированную цепь после репликации ДНК, и поэтому ее часто называют «поддерживающей» метилтрансферазой. [20] DNMT1 необходим для правильного эмбрионального развития, импринтинга и Х-инактивации. [16] [21]
Гистоны h4 и h5 также можно манипулировать путем деметилирования с использованием гистон-лизин-деметилазы (KDM). Этот недавно идентифицированный фермент имеет каталитически активный сайт, называемый доменом Jumonji (JmjC). Деметилирование происходит, когда JmjC использует несколько кофакторов для гидроксилирования метильной группы, тем самым удаляя ее. JmjC способен деметилировать моно-, ди- и триметилированные субстраты. [22]
Хромосомные области могут принимать стабильные и наследуемые альтернативные состояния, что приводит к бистабильной экспрессии генов без изменений в последовательности ДНК. Эпигенетический контроль часто связан с альтернативными ковалентными модификациями гистонов. [23] Часто считается, что стабильность и наследуемость состояний более крупных хромосомных регионов связаны с положительной обратной связью, когда модифицированные нуклеосомы привлекают ферменты, которые аналогичным образом модифицируют близлежащие нуклеосомы. Упрощенная стохастическая модель для этого типа эпигенетики находится здесь. [24] [25]
Поскольку метилирование ДНК и ремоделирование хроматина играют центральную роль во многих типах эпигенного наследования, слово «эпигенетика» иногда используется как синоним этих процессов. Однако это может ввести в заблуждение. Ремоделирование хроматина не всегда передается по наследству, и не всякое эпигенетическое наследование включает ремоделирование хроматина. [26]
Было высказано предположение, что гистоновый код может быть опосредован действием малых РНК. Недавнее открытие и характеристика обширного массива малых (от 21 до 26 нуклеотидов) некодирующих РНК позволяет предположить, что существует компонент РНК, возможно участвующий в регуляции эпигенетического гена. Малые интерферирующие РНК могут модулировать экспрессию генов транскрипции посредством эпигенетической модуляции промоторов-мишеней. [27]
Транскрипты РНК и кодируемые ими белки
Иногда ген после включения транскрибирует продукт, который (прямо или косвенно) поддерживает активность этого гена. Напр., Hnf4 и MyoD усиливают транскрипцию многих генов, специфичных для печени и мышц, соответственно, включая их собственные, за счет активности фактора транскрипции белков, которые они кодируют. Передача сигналов РНК включает дифференциальное рекрутирование иерархии родовых комплексов, модифицирующих хроматин, и ДНК-метилтрансфераз в специфические локусы с помощью РНК во время дифференцировки и развития. [28] Другие эпигенетические изменения опосредуются продукцией различных форм сплайсинга РНК или образованием двухцепочечной РНК (РНКи). Потомки клетки, в которой ген был включен, унаследуют эту активность, даже если первоначальный стимул для активации гена больше не присутствует. Эти гены чаще всего включаются или выключаются путем передачи сигнала, хотя в некоторых системах, где важны синцитии или щелевые соединения, РНК может распространяться непосредственно в другие клетки или ядра путем диффузии. Большое количество РНК и белка вносится в зиготу матерью во время оогенеза или через питающие клетки, что приводит к фенотипам с материнским эффектом. Меньшее количество РНК сперматозоидов передается от отца, но недавно появились доказательства того, что эта эпигенетическая информация может привести к видимым изменениям в нескольких поколениях потомства. [29]
Прионы
- Подробнее по этой теме см. Прионы.
Прионы представляют собой инфекционные формы белков. Как правило, белки складываются в отдельные единицы, которые выполняют определенные клеточные функции, но некоторые белки также способны формировать инфекционное конформационное состояние, известное как прион. Хотя прионы часто рассматриваются в контексте инфекционных заболеваний, они более свободно определяются по их способности каталитически преобразовывать другие версии того же белка в нативном состоянии в инфекционное конформационное состояние. Именно в этом последнем смысле их можно рассматривать как эпигенетические агенты, способные вызывать фенотипические изменения без модификации генома. [30]
Грибковые прионы считаются эпигенетическими, поскольку инфекционный фенотип, вызванный прионом, может передаваться по наследству без модификации генома. PSI+ и URE3, обнаруженные у дрожжей в 1965 и 1971 годах, являются двумя наиболее изученными представителями этого типа прионов. [31] [32] Прионы могут иметь фенотипический эффект за счет связывания белка в агрегатах, тем самым снижая активность этого белка. В клетках PSI+ потеря белка Sup35 (который участвует в терминации трансляции) приводит к тому, что рибосомы имеют более высокую скорость считывания стоп-кодонов, эффект, который приводит к подавлению нонсенс-мутаций в других генах. [33] Способность Sup35 образовывать прионы может быть консервативным признаком. Это может дать адаптивное преимущество, давая клеткам возможность переключаться в состояние PSI+ и выражать неактивные генетические особенности, которые обычно заканчиваются преждевременными мутациями стоп-кодона. [34] [35]
Системы структурного наследования
- Дополнительные сведения по этой теме см. в разделе Структурное наследование.
Инфузории, такие как Tetrahymena и Paramecium , генетически идентичные клетки обнаруживают наследуемые различия в структуре ресничных рядов на своей клеточной поверхности. Экспериментально измененные паттерны могут быть переданы дочерним клеткам. Кажется, что существующие структуры действуют как шаблоны для новых структур. Механизмы такого наследования неясны, но существуют основания предполагать, что многоклеточные организмы также используют существующие клеточные структуры для сборки новых. [36] [37] [38]
Функции и последствия
Развитие
Соматическая эпигенетическая наследственность посредством эпигенетических модификаций, особенно посредством метилирования ДНК и ремоделирования хроматина, очень важна для развития многоклеточных эукариотических организмов. Последовательность генома статична (за некоторыми заметными исключениями), но клетки дифференцируются во множество различных типов, которые выполняют разные функции и по-разному реагируют на окружающую среду и межклеточную передачу сигналов. Таким образом, по мере развития индивидуумов морфогены активируют или заглушают гены эпигенетически наследуемым образом, давая клеткам «память». У млекопитающих большинство клеток окончательно дифференцируются, и только стволовые клетки сохраняют способность дифференцироваться в несколько типов клеток («тотипотентность» и «мультипотентность»). У млекопитающих некоторые стволовые клетки продолжают продуцировать новые дифференцированные клетки на протяжении всей жизни, но млекопитающие не способны реагировать на потерю некоторых тканей, например, неспособностью к регенерации конечностей, на что способны некоторые другие животные. В отличие от животных растительные клетки окончательно не дифференцируются, оставаясь тотипотентными со способностью давать начало новому индивидуальному растению. Хотя растения действительно используют многие из тех же эпигенетических механизмов, что и животные, такие как ремоделирование хроматина, было высказано предположение, что растительные клетки не имеют «памяти», сбрасывая свои паттерны экспрессии генов при каждом делении клетки, используя информацию о положении из окружающей среды и окружающих клеток. определить их судьбу. [39]
Медицина
Эпигенетика имеет много различных потенциальных медицинских применений, поскольку она имеет многомерный характер. [40] Врожденные генетические заболевания хорошо изучены, и также ясно, что эпигенетика может играть роль, например, в случае синдрома Ангельмана и синдрома Прадера-Вилли. Это нормальные генетические заболевания, вызванные генными делециями или инактивацией генов, но они необычайно распространены, потому что люди по существу гемизиготны из-за геномного импринтинга, и поэтому достаточно нокаута одного гена, чтобы вызвать заболевание, тогда как в большинстве случаев потребуются обе копии. быть выбитым. [41]
Эволюция
Эпигенетические механизмы были необходимой частью эволюционного происхождения дифференцировки клеток. [42] Хотя обычно считается, что эпигенетика многоклеточных организмов является механизмом, участвующим в дифференцировке, с «перезапуском» эпигенетических паттернов при размножении организмов, были некоторые наблюдения трансгенерационного эпигенетического наследования (например, явление парамутации, наблюдаемое у кукурузы ). Хотя большинство этих многопоколенческих эпигенетических признаков постепенно утрачиваются в течение нескольких поколений, остается возможность того, что эпигенетика нескольких поколений может быть еще одним аспектом эволюции и адаптации. Секвестрированная зародышевая линия или барьер Вейсмана специфичны для животных, и ожидается, что эпигенетическая наследственность будет гораздо более распространена у растений и микробов. Эти эффекты могут потребовать усовершенствования стандартной концептуальной основы современного эволюционного синтеза. [43] [44]
Эпигенетические особенности могут играть роль в краткосрочной адаптации видов, допуская обратимую изменчивость фенотипа. Модификация эпигенетических особенностей, связанных с участком ДНК, позволяет организмам во временной шкале нескольких поколений переключаться между фенотипами, экспрессирующими и репрессирующими этот конкретный ген. [45] Когда последовательность ДНК региона не мутирована, это изменение является обратимым. Также предполагалось, что организмы могут использовать дифференциальную скорость мутаций, связанную с эпигенетическими особенностями, для контроля скорости мутаций определенных генов. [45] Интересно, что недавние анализы показали, что члены семейства цитозиндеаминаз APOBEC/AID способны одновременно обеспечивать генетическое и эпигенетическое наследование с использованием сходных молекулярных механизмов. [46]
File:Epigenesis.jpg Эволюционную эпигенетику можно разделить на предопределенный и вероятностный эпигенез. Предопределенный эпигенез представляет собой однонаправленное движение от структурного развития в ДНК к функциональному созреванию белка. «Предопределенный» здесь означает, что разработка запрограммирована и предсказуема. Вероятностный эпигенез, с другой стороны, представляет собой двунаправленное структурно-функциональное развитие с опытом и развитием внешнего формирования. [47]
Эпигенетические изменения также наблюдались в ответ на воздействие окружающей среды — например, у мышей, получавших некоторые пищевые добавки, наблюдались эпигенетические изменения, влияющие на экспрессию гена агути, который влияет на цвет их меха, вес и склонность к развиться рак. [48] [49]
Сообщалось о более чем 100 случаях феномена трансгенерационного эпигенетического наследования у широкого круга организмов, включая прокариоты, растения и животных. [50]
Эпигенетические эффекты у человека
Геномный импринтинг и родственные расстройства
Некоторые нарушения у человека связаны с геномным импринтингом, явлением у млекопитающих, когда отец и мать вносят разные эпигенетические паттерны для определенных геномных локусов в их зародышевых клетках . [51] Наиболее известным случаем импринтинга при заболеваниях человека являются синдром Ангельмана и синдром Прадера-Вилли — оба могут быть вызваны одной и той же генетической мутацией, частичной делецией хромосомы 15q, и конкретный синдром, который разовьется, зависит от унаследована ли мутация от матери ребенка или от отца. [52] Это связано с наличием геномного импринтинга в регионе. Синдром Беквита-Видеманна также связан с геномным импринтингом, часто вызванным аномалиями материнского геномного импринтинга области на хромосоме 11.
Трансгенерационные эпигенетические наблюдения
См. основную статью Трансгенерационная эпигенетика
В исследовании Överkalix Маркус Пембри и его коллеги заметили, что внуки по отцовской (но не по материнской) линии [53] шведских мужчины, подвергшихся голоду в подростковом возрасте в 19 веке, имели меньше шансов умереть от сердечно-сосудистых заболеваний. Если пищи было много, то смертность от диабета у внуков увеличивалась, что позволяет предположить, что это было трансгенерационное эпигенетическое наследование. [54] У самок наблюдался противоположный эффект — внучки по отцовской (но не по материнской) линии женщин, переживших голод в утробе матери (и, следовательно, во время формирования яйцеклеток), жили в среднем меньше. [55]
Исследования близнецов
Недавние исследования с участием как дизиготных, так и монозиготных близнецов дали некоторые доказательства эпигенетического влияния на людей. [56] [57] [58]
См. также
- Болдуинская эволюция
- Поведенческая эпигенетика
- Центромера
- Вычислительная эпигенетика
- Деметилирование ДНК
- Возникновение
- Эпигенетика в обучении и памяти
- Эпигенетика в психологии
- Эпигенетика аутизма
- Эпигенетика кокаиновой зависимости
- Эпигенетическая теория
- Эпигеномика
- Эволюционная емкость
- Эволюционная психология развития
- Внеядерное наследование
- Код гистона
- Геном человека
- Впечатывание
- Ламаркизм
- Материнский эффект
- Молекулярная биология
- Нутриэпигеномика
- Парамутация
- Прион
- Мягкое наследство
- Соматический эпитип
- Барьер Вейсмана
- Barbara McClintock
Внешние ссылки
- The Epigenome Network of Excellence (NoE)
- Научный веб-сайт для эпигенома NoE
- Консорциум эпигенома человека
- BBC Horizon 2005, документальный фильм «Призрак в твоих генах» об эпигенетике.
- Код 2, статья (образ жизни и активация генов)
- Эпигенетика и ее приложения
- Новости эпигенетики
- Эпигенетический центр протокола и информационных ресурсов
- Epigentek, крупнейший производитель продуктов для эпигенетических исследований
Ссылки
- ↑ Bird A (май 2007 г.). Представления об эпигенетике. Природа 447 (7143): 396–8.
- ↑ Специальный репортаж: «Что помнят гены» Филипа Хантера | Журнал «Проспект», выпуск 146, май 2008 г. Web.archive.org. URL-адрес получен 26 июля 2012 г.
- ↑ Рейк В. (май 2007 г.). Стабильность и гибкость эпигенетической регуляции генов в развитии млекопитающих. Природа 447 (7143): 425–32.
- ↑ Цзя, Гуйфан, Фу, Е, Чжао, Сюй, Дай, Цин, Чжэн, Гуаньцюнь, Ян, Ин, И, Чэнци, Линдал, Томас, Пан, Тао, Ян, Юнь-Гуй, Хэ, Чуань (16 октябрь 2011 г.). N6-метиладенозин в ядерной РНК является основным субстратом FTO, ассоциированного с ожирением. Природа Химия Биология 7 (12): 885–887.
- ↑ Новое исследование связывает распространенную модификацию РНК с ожирением. Physorg.com. URL-адрес получен 26 июля 2012 г.
- ↑ Waddington CH (1942). Эпигенотип. Индевор 1 : 18–20.
- ↑ Согласно Оксфордскому словарю английского языка:
Также стоит процитировать это краткое изложение определения, данного там (а именно, «образование органического зародыша как нового продукта»):Слово используется У. Харви, Exercitationes 1651, p. 148, и в английских анатомических упражнениях 1653, с. 272. Объясняется, что это означает «partium super-exorientium additamentum», «сложение частей, отпочковавшихся одна от другой».
теория эпигенеза : теория о том, что зародыш создан последовательными срастаниями), а не просто развитыми в процессе воспроизводства. […] Противоположная теория ранее была известна как «теория эволюции»; чтобы избежать двусмысленности этого названия, теперь о ней говорят главным образом как о «теории преформации», иногда как о «оболочке» или «внедрении».
- ↑ Холлидей Р. (ноябрь 1990 г.). Механизмы контроля активности генов в процессе развития. Biol Rev Camb Philos Soc 65 (4): 431–71.
- ↑ Риггс А.Д., Руссо ВЭА, Мартиенссен Р.А. (1996). Эпигенетические механизмы регуляции генов , Плейнвью, Нью-Йорк: Издательство лаборатории Колд-Спринг-Харбор.
- ↑ Эриксон, Эрик (1968). Личность: Молодежь и кризис , Глава 3: В.В. Нортон и Компания, Инк..
- ↑ Эпигенетика. Био-Медицина.org. URL-адрес получен 21 мая 2011 г.
- ↑ Чендлер В.Л. (февраль 2007 г.). Парамутация: от кукурузы к мышам. Сотовый 128 (4): 641–5.
- ↑ Яблонка Э., Лэмб М.Дж., Лахманн М. (сентябрь 1992 г.). Доказательства, механизмы и модели наследования приобретенных признаков. Ж. Теорет. биол. 158 (2): 245–268.
- ↑ Jenuwein T, Laible G, Dorn R, Reuter G (19 января98). Белки домена SET модулируют домены хроматина в эу- и гетерохроматине. Сотовый. Мол. Жизнь наук. 54 (1): 80–93.
- ↑ Слоткин Р.К., Мартиенссен Р. (апрель 2007 г.). Мобильные элементы и эпигенетическая регуляция генома. Нац. Преподобный Жене. 8 (4): 272–85.
- ↑ 16,0 16,1 Ли Э., Бестор Т.Х., Йениш Р. (июнь 1992 г.). Целенаправленная мутация гена ДНК-метилтрансферазы приводит к эмбриональной летальности. Сотовый 69 (6): 915–26.
- ↑ Робертсон К.Д., Узволгий Э., Лян Г., Талмадж С., Сумеги Дж., Гонсалес Ф.А., Джонс П.А. (июнь 1999 г.). ДНК-метилтрансферазы человека (DNMTs) 1, 3a и 3b: координируют экспрессию мРНК в нормальных тканях и сверхэкспрессию в опухолях. Рез. нуклеиновых кислот. 27 (11): 2291–8.
- ↑ Leonhardt H, Page AW, Weier HU, Bestor TH (ноябрь 1992 г.). Целевая последовательность направляет ДНК-метилтрансферазу к местам репликации ДНК в ядрах млекопитающих. Сотовый 71 (5): 865–73.
- ↑ Чуанг Л.С., Ян Х.И., Кох Т.В., Нг Х.Х., Сюй Г., Ли Б.Ф. (сентябрь 1997 г.). Комплекс ДНК-(цитозин-5) метилтрансфераза-PCNA человека как мишень для p21WAF1. Наука 277 (5334): 1996–2000.
- ↑ Робертсон К.Д., Вольф А.П. (октябрь 2000 г.). Метилирование ДНК в норме и при патологии. Нац. Преподобный Жене. 1 (1): 11–9.
- ↑ Li E, Beard C, Jaenisch R (19 ноября93). Роль метилирования ДНК в геномном импринтинге. Природа 366 (6453): 362–5.
- ↑ Nottke A, Colaiácovo MP, Shi Y (март 2009 г.). Роль гистон-лизин-деметилазы в развитии. Развитие 136 (6): 879–89.
- ↑ Розенфельд Дж.А., Ван З., Шонес Д.Е., Чжао К., ДеСалле Р., Чжан М.К. (2009). Определение обогащенных модификаций гистонов в негенных участках генома человека. BMC Геномика 10 .
- ↑ Эпигенетическая клеточная память. Смоль.нби.дк. URL-адрес получен 26 июля 2012 г.
- ↑ Додд И.Б., Мичелсен М.А., Снеппен К., Тон Г. (май 2007 г.). Теоретический анализ эпигенетической клеточной памяти путем модификации нуклеосом. Сотовый 129 (4): 813–22.
- ↑ Пташне М. (апрель 2007 г.). Об употреблении слова «эпигенетический». Курс. биол. 17 (7): R233–6.
- ↑ Моррис К.Л. (2008). «Эпигенетическая регуляция экспрессии генов» РНК и регуляция экспрессии генов: скрытый уровень сложности , Норфолк, Англия: Caister Academic Press.
- ↑ Mattick JS, Amaral PP, Dinger ME, Mercer TR, Mehler MF (январь 2009 г.). РНК-регуляция эпигенетических процессов. BioEssays 31 (1): 51–9.
- ↑ Чой CQ. Ученый: РНК может быть наследственной молекулой. Ученый. URL-адрес, полученный в 2006 г.
- ↑ Юл А., Эдмундс В.Дж. (1998). Эпигенетическая наследственность и прионы. Журнал эволюционной биологии 11 (2): 241–242.
- ↑ Кокс Б.С. (1965). [PSI], цитоплазматический супрессор суперсупрессии у дрожжей. Наследственность 20 (4): 505–521.
- ↑ Лакрут Ф. (май 1971 г.). Неменделевская мутация, обеспечивающая поглощение уреидоянтарной кислоты дрожжами. J. Бактериол. 106 (2): 519–22.
- ↑ Либман С.В., Шерман Ф. (сентябрь 1979 г.). Внехромосомная детерминанта psi+ подавляет нонсенс-мутации у дрожжей. J. Бактериол. 139 (3): 1068–71.
- ↑ True HL, Lindquist SL (сентябрь 2000 г.). Дрожжевой прион обеспечивает механизм генетической изменчивости и фенотипического разнообразия. Природа 407 (6803): 477–83.
- ↑ Шортер Дж., Линдквист С. (июнь 2005 г.). Прионы как адаптивные проводники памяти и наследственности. Нац. Преподобный Жене. 6 (6): 435–50.
- ↑ Сапп Дж. (1991). Понятия организации. Рычаги инфузорий простейших. Дев. биол. (Нью-Йорк) 7 : 229–58.
- ↑ Сапп Дж. (2003). Бытие: эволюция биологии , Оксфорд [Оксфордшир]: Издательство Оксфордского университета.
- ↑ Грей Р.Д., Ояма С., Гриффитс ЧП (2003). Циклы непредвиденных обстоятельств: системы развития и эволюция (жизнь и разум: философские вопросы биологии и психологии) , Кембридж, Массачусетс: MIT Press.
- ↑ Коста С., Шоу П. (март 2007 г.). «Открытые» клетки: как клетки могут изменить судьбу. Trends Cell Biol. 17 (3): 101–6.
- ↑ Чахван Р., Вонтакал С.Н., Роа С. (март 2011 г.). Многомерный характер эпигенетической информации и ее роль в заболевании. Discov Med 11 (58): 233–43.
- ↑ ОМИМ 105830
- ↑ Hoekstra RF (2000). Эволюция: введение , Оксфорд [Оксфордшир]: Издательство Оксфордского университета.
- ↑ Лэмб М.Дж., Яблонка Э. (2005). Эволюция в четырех измерениях: генетические, эпигенетические, поведенческие и символические вариации в истории жизни , Кембридж, Массачусетс: MIT Press.
- ↑ См. также Denis Noble Музыка жизни см. особенно стр. 93–8 и стр. 48, где он цитирует обзор «Яблонка и ягненок» и Массимо Пильуччи о Яблоньке и ягненке в журнале Nature 9.1019 435 , 565–566 (2 июня 2005 г.)
- ↑ 45,0 45,1 Рандо О.Дж., Верстрепен К.Дж. (февраль 2007 г.). Временные шкалы генетического и эпигенетического наследования. Сотовый 128 (4): 655–68.
- ↑ Чахван Р., Вонтакал С.Н., Роа С. (октябрь 2010 г.). Перекрестные помехи между генетической и эпигенетической информацией посредством дезаминирования цитозина. Тенденции Жене. 26 (10): 443–8.
- ↑ Griesemer J, Haber MH, Yamashita G, Gannett L (март 2005 г. ). Критическое замечание: циклы непредвиденных обстоятельств — системы развития и эволюция. Биология и философия 20 (2–3): 517–544.
- ↑ Куни К.А., Дэйв А.А., Вольф Г.Л. (август 2002 г.). Материнские метиловые добавки у мышей влияют на эпигенетическую изменчивость и метилирование ДНК потомства. Дж. Нутр. 132 (8 Дополнение): 2393S–2400S.
- ↑ Waterland RA, Jirtle RL (август 2003 г.). Мобильные элементы: мишени для раннего воздействия питания на регуляцию эпигенетического гена. Мол. Клетка. биол. 23 (15): 5293–300.
- ↑ Яблонка Э., Раз Г. (июнь 2009 г.). Трансгенерационное эпигенетическое наследование: распространенность, механизмы и значение для изучения наследственности и эволюции. Q Rev Biol 84 (2): 131–76.
- ↑ Вуд А.Дж., Оки Р.Дж. (ноябрь 2006 г.). Геномный импринтинг у млекопитающих: новые темы и устоявшиеся теории. Генетика PLoS. 2 (11): e147.
- ↑ Нолл Дж. Х., Николлс Р. Д., Магенис Р. Е., Грэм Дж. М., Лаланд М., Латт С. А. (19 февраля89). Синдромы Ангельмана и Прадера-Вилли имеют общую делецию хромосомы 15, но различаются родительским происхождением делеции. утра. Дж. Мед. Жене. 32 (2): 285–90.
- ↑ Внук лица по отцовской линии является сыном сына этого лица; внук по материнской линии — сын дочери.
- ↑ Пембри М.Е., Бигрен Л.О., Каати Г., Эдвинссон С., Нортстоун К., Шёстрём М., Голдинг Дж. (февраль 2006 г.). Половые трансгенерационные реакции по мужской линии у людей. Евро. Дж. Хам. Жене. 14 (2): 159–66. Роберт Уинстон ссылается на это исследование в своей лекции; см. также обсуждение в Университете Лидса, здесь [1]
- ↑ НОВА | Стенограммы | Призрак в твоих генах. ПБС. URL-адрес получен 26 июля 2012 г.
- ↑ includeonly>О’Коннор, Анахад. «Утверждение: идентичные близнецы имеют одинаковую ДНК», New York Times , 11 марта 2008 г. Проверено 2 мая 2010 г.
- ↑ Каминский З.А., Танг Т., Ван С.К., Птак С., Ох Г.Х., Вонг А.Х., Фельдкамп Л.А., Виртанен С., Хальварсон Дж., Тиск С., МакРэй А.Ф., Висшер П.М., Монтгомери Г.В., Готтесман И.И., Мартин Н.Г., Петронис А. (февраль 2009 г.). Профили метилирования ДНК у монозиготных и дизиготных близнецов. Нац. Жене. 41 (2): 240–5.
- ↑ Фрага М.Ф., Баллестар Э., Пас М.Ф., Роперо С., Сетьен Ф., Баллестар М.Л., Хайне-Суньер Д., Сигудоса Х.К., Уриосте М., Бенитес Х., Буа-Шорне М., Санчес-Агилера А., Линг К., Карлссон Э. , Поульсен П., Вааг А., Стефан З., Спектор Т.Д., Ву Ю.З., Пласс С., Эстеллер М. (июль 2005 г.). Эпигенетические различия возникают в течение жизни монозиготных близнецов. Проц. Натл. акад. науч. США 102 (30): 10604–9.
- Оскар Хертвиг, 1849–1922 гг. Биологическая проблема современности: преформация или эпигенез? Основа теории органического развития . В. Хайнеманн: Лондон, 1896 г. .
- R. Jaenisch и A. Bird (2003) Эпигенетическая регуляция экспрессии генов: как геном интегрирует внутренние сигналы и сигналы окружающей среды. Нац. Жене. 33 (дополнение) 245-254.
- Джошуа Ледерберг, «Значение эпигенетики», Ученый 15 (18):6, 17 сентября 2001 г.
- Р. Дж. Симс III, К. Нишиока и Д. Рейнберг (2003) Метилирование лизина гистонов: признак функции хроматина. Тенденции Жене. 19 , 629-637.
- Б. Д. Страль и К. Д. Аллис (2000) Язык ковалентных модификаций гистонов. Природа 403 , 41-45.
- Ч.Х. Уоддингтон (1942), «Эпигенотип». Индевор 1 , 18–20.
- Р.А. Уотерленд, Р.Л. Джиртл, «Мобильные элементы: мишени для раннего воздействия питания на регуляцию эпигенетического гена», Molecular and Cellular Biology 2003, 1 августа; 23 (15):5293-5300.
- Б. МакКлинток (1978) Механизмы быстрой реорганизации генома. Stadler Symposium vol 10:25-48
- Г.В. Граймс; К.Дж. Ауфдерхайде; Клеточные аспекты формирования узора: проблема сборки. Монографии по биологии развития, Vol. 22. Каргер, Базель (1991)
- Li E, Bestor TH и Jaenisch R. Целевая мутация гена ДНК-метилтрансферазы приводит к гибели эмбрионов. Сотовый 69, 915-926 (1992)
- Li E, Beard C и Jaenisch R. Роль метилирования ДНК в геномном импринтинге. Природа 366, 362-365 (1993)
- Робертсон К.Д., Узёлджи Э., Лян Г., Талмадж С., Сумеги Дж., Гонсалес Ф.А. и Джонс П.А. ДНК-метилтрансферазы человека (DNMT) 1, 3a, 3b: координируют экспрессию мРНК в нормальных тканях и сверхэкспрессию в опухолях. Рез. нуклеиновых кислот. 27, 2291-2298 (1999)
- Леонхардт Х., Пейдж А., Вейер Х. и Бестор Т.Х. Целевая последовательность направляет ДНК-метилтрансферазу к местам репликации ДНК в ядрах млекопитающих. Сотовый 71, 865-873 (1992)
- Chuang, L. Комплекс ДНК человека-(цитозин-5)метилтрансфераза-PCNA является мишенью для p21 Waf1. Наука 277 , 1996-2000 (1997)
- Робертсон, К.Д. и Wolffe, AP Метилирование ДНК в норме и болезни. Nature Rev. Genet. 1 , 11-19(2000)
File:Epigenome NoE tag1.gif
| edit Развитие фенотипа |
|---|
| Ключевые понятия: Генотип-фенотип различия | Нормы реакции | Взаимодействие генов с окружающей средой | Наследуемость | Количественная генетика |
| Генетическая архитектура: Отношения доминирования | Эпистаз | Полигенное наследование | плейотропия | Пластичность | Канализация | Фитнес-ландшафт |
| Негенетические влияния: Эпигенетическое наследование | Эпигенетика | Материнский эффект | теория двойного наследования |
| Архитектура разработки: Сегментация | Модульность |
| Эволюция генетических систем: Эволюционность | Мутационная устойчивость | Эволюция секса |
| Влиятельные фигуры: CH Waddington | Ричард Левонтин |
| Дебаты: Природа против воспитания |
| Список тем эволюционной биологии |
де:Эпигенетика es: Эпигенетика fr: Эпигенетическая он: אפיגנטיקה hu: Эпигенетика nl: Эпигенетика sl:Эпигенетика fi:Epigeneettinen perytyminen sv: Эпигенетика zh:表觀遺傳學
| На этой странице используется лицензированный Creative Commons контент из Википедии (просмотреть авторов).
|
Эпигенетические эффекты материнского поведения… Очень похоже на теорию привязанности
Сэмми Ахмед
В сфере человеческого развития ведутся большие споры о том, что больше влияет на различные аспекты человеческого развития: «природа» или «воспитание». Область психологии развития признает как природу, так и воспитание как детерминированные силы, однако уделяет большое внимание влияниям, связанным с «воспитанием», предоставляя доказательства того, что постнатальная семейная среда является основной силой в развитии индивидуальных различий личности (Чжан и Мини, 2010). С другой стороны, биологи развития склонны рассматривать человеческое развитие через призму «природы». Например, биологические или эволюционные подходы понимают мозг и его развитие как подчиненные эволюционным силам вне непосредственных факторов окружающей среды. Кроме того, генетики предоставляют доказательства того, что вариации в поведении — это просто вариации в последовательности ДНК (Zhang & Meaney, 2010). Хотя эти две противоположные точки зрения делают зонтичную область человеческого развития очень динамичной и всеобъемлющей, зарождающаяся область эпигенетики может объединить эти две дисциплины и обеспечить единое понимание человеческого развития.
Эпигенетика — это область эволюционной биологии, которая фокусируется на ненаследственных модификациях генетического материала и различных факторах, которые могут изменить экспрессию генов (Lillycrop et al., 2007), таких как метилирование ДНК. Когда участки генов метилированы, факторы, необходимые для экспрессии, ингибируются. Наоборот, деметилирование участков генов усиливает экспрессию генов (Henckel, Toth, & Arnaud, 2007). Другими словами, метилирование ДНК вызывает «включение или выключение» определенных генов без изменения фактической последовательности ДНК (Moalem & Prince, 2007). Чтобы лучше понять, как работает эпигенетика, рассмотрите знаменитое «исследование тощих коричневых мышей» в Университете Дьюка. В ходе этого исследования группа ученых использовала жирных желтых мышей для проверки своей эпигенетической гипотезы (Moalem & Prince, 2007). Эти толстые желтые мыши обычно рождались толстыми желтыми мышами из-за гена, называемого агути, который придавал им пухлые/желтые характеристики (Moalem & Prince, 2007). Однако, когда экспериментальная группа перешла на другую диету, они родили тощих коричневых мышей. Интересно, что при анализе генетического кода тощих коричневых мышей исследователи обнаружили, что ген агути все еще присутствует, но характеристики, которые обеспечивает ген, больше не проявляются (Moalem & Prince, 2007). Исследователи пришли к выводу, что изменение рациона мыши-матери привело к отключению гена. Другими словами, новая диета, в которую входили «доноры метила», запускала метилирование генов агути и подавляла ген, что приводило к тощим коричневым мышам (Moalem & Prince, 2007).
Вскоре после «эксперимента с тощими коричневыми мышами» область эпигенетики взорвалась. Было очевидно, что генетическая последовательность не высечена на камне, и факторы окружающей среды могут изменить экспрессию определенных генов. Просто краткий обзор генетического кодирования и экспрессии. Генетический состав/код называется генотипом, он определяет приобретаемые гены, но не обязательно экспрессируемые гены; экспрессия генов называется фенотипом (Mustard, 2010). Например, однояйцевые близнецы имеют одинаковую ДНК (генотип), но часто имеют разный опыт, что приводит к различиям в экспрессии генов (фенотип). Исследования показывают, что у однояйцевых близнецов разница в поведении во взрослом возрасте может достигать 30%. Согласно эпигенетической теории, это различие, вероятно, связано с эпигенетическими аффектами в раннем развитии (Mustard, 2010). Было проведено множество исследований, изучающих эпигенетические эффекты на различные психологические последствия. Например, исследования показали, что эпигенетика может быть фактором шизофрении, биполярных расстройств и синдрома дефицита внимания с гиперактивностью (Mustard, 2010).
В то время как «эксперимент с тощими коричневыми мышами» показал, как факторы до рождения могут влиять на характеристики потомства, исследования также показали, что эпигенетические модификации могут происходить после рождения. Фактически, исследование в Университете Макгилла было направлено на изучение того, могут ли быть эпигенетические изменения после рождения. Согласно Meaney & Szyf (2005), взаимодействие между матерями и их потомством может спровоцировать эпигенетические изменения в фенотипах их потомства. Результаты их эксперимента показывают, что, когда крысята получали разное внимание от своих матерей, они росли с относительно разными темпераментами. Крысы, которые получали больше внимания от своих матерей вскоре после рождения, становились расслабленными, общительными и справлялись со стрессом лучше, чем их собратья, лишенные внимания, которые вырастали нервными и более восприимчивыми к стрессу (Meaney & Szyf, 2005; Moalem & Prince). , 2007). Чтобы исключить генетические эффекты, исследователи поменяли крысят; они давали крысят от матерей, которые были менее внимательными, к более внимательным матерям. Независимо от биологической матери, дети, получавшие больше материнской заботы, росли хорошо приспособленными и обладали «лучшими социальными навыками» (Moalem & Prince, 2007). Кроме того, последующий генный анализ показал, что у крыс, которые получали больше материнского внимания, было снижено количество метильных продуцентов генов, связанных с развитием мозга, поэтому исследователи пришли к выводу, что материнская забота удалила метильные маркеры, которые в противном случае вызвали бы их нервозность. темперамент (Moalem & Prince, 2007).
Интересно, что результаты эксперимента Мини очень похожи на этологическую теорию привязанности Джона Боулби. Экологическая теория привязанности была создана в 1969 году и стала важной теорией в психологии развития, поскольку она пытается понять, как связь младенца и опекуна связана с чувством безопасности ребенка, его эмоциональным развитием и будущими отношениями. Понятие установления надежной привязанности к основному опекуну имеет первостепенное значение для этой теории и связано с различными социальными и эмоциональными результатами. На этапе до привязанности (от рождения до 6 недель) наличие внимательного опекуна имеет решающее значение для установления этой привязанности, поскольку это помогает новорожденным чувствовать связь и комфорт (Berk, 2009). ). Согласно Sroufe (2005), дети, у которых была надежная привязанность к своим опекунам, демонстрировали более высокий уровень самооценки, социальной компетентности и эмпатии, чем дети с ненадежной привязанностью в дошкольном возрасте. При повторном тестировании в возрасте 11 лет дети, у которых была надежная привязанность к своему основному опекуну, продемонстрировали лучшие социальные навыки и более тесные отношения со сверстниками; эти дети продолжали извлекать пользу из своих надежных привязанностей в детстве в подростковом и взрослом возрасте (Sroufe, 2005).
Хотя эпигенетические принципы выявили некоторые важные открытия, связанные с психологическим развитием, проблема заключается в интеграции генетических и биологических принципов в дисциплину, основанную на окружающей среде, такую как психология развития. Полученные данные не обязательно согласуются с основами влияния окружающей среды на психологические процессы и развитие; однако показать, как генетика и окружающая среда взаимодействуют друг с другом и вместе играют большую роль в развитии человека. Принятие целостного подхода означает лучшее понимание динамики человеческого развития и имеет решающее значение для продвижения вперед как в биологических, так и в экологических дисциплинах.
Ссылки
Berk, LE (2009). Развитие ребенка (8-е изд.).
Бостон, Массачусетс: Pearson Publishers.
Хенкель, А., Тот, С., и Арно, П. (2007). Раннее развитие эмбриона мыши: может ли эпигенетика влиять на определение клеточной судьбы? Bioessays, 29 (6), 520-524.
Лилликроп, К.А., Слейтер-Джеффрис, Дж.Л., Хэнсон, Массачусетс, Годфри, К.М., Джексон, А.А., и Бердж, Г.К. (2007). Индукция измененной эпигенетической регуляции печеночного глюкокортикоидного рецептора у потомства крыс, получавших во время беременности диету с ограниченным содержанием белка, позволяет предположить, что снижение экспрессии ДНК-метилтрансферазы-1 связано с нарушением метилирования ДНК и изменениями модификаций гистонов. Br Journal of Nutrition, 97 (6), 1064-1073.
Мини, М. Дж., и Шиф, М. (2005). Материнская забота как модель пластичности хроматина, зависящей от опыта? Trends in Neuroscience, 28 (9), 456-463.
Моалем, С., и Принс, Дж. (2007). Выживание самых больных: удивительная связь между болезнями и долголетием . Нью-Йорк: Издательство HarperCollins.
Горчица, JF (2010). Раннее развитие мозга и развитие человека. Энциклопедия раннего развития детей. (опубликовано в Интернете 17 февраля 2010 г.)
Sroufe, LA (2005). Привязанность и развитие: проспективное лонгитюдное исследование от рождения до взрослой жизни. Привязанность и развитие человека, 7 , 349-367
Чжан, Т., и Мини, М. Дж. (2010). Эпигенетика и экологическая регуляция генома и его функции. Ежегодный обзор психологии, 61, 439–66. doi: 10.1146/annurev.psych.60.110707.163625
Биография автора
Сэмми Ахмед является выпускником программы прикладной психологии с отличием.