
Эпигенетический принцип это: Эпигенетический принцип это
Эпигенетический принцип
Многие психологи,психотерапевтыипсихоаналитикипослеФрейда, такие какЭрих Фромм и Карен Хорни, пытались пересмотреть психоанализ, чтобы показать значение связанных сЯ процессов и проследить психологиюих развития. Наиболее выдающимся из так называемыхэго-психологов был Эрик Эриксон. Как и для других постфрейдистов, для Эриксона наибольшее значение имело Я и его адаптивные способности (в том числе, в преодолении невроза,депрессии,фобии, последствийстресса) в связи с проблемойразвития индивидуума и психологической помощиему (психологическое консультирование,психотерапия). Однако из этого не следует, что в своей теории он пренебрег биологическими или социальными факторами. По сути дела, Эриксон настаивал на том, что любой психологический феномен может быть понят в контексте согласованного взаимодействия биологических, поведенческих, эмпирических и социальных факторов. К другим особенностям теоретической ориентации Эриксона относятся следующие:
1) акцент на изменениях, происходящих в процессе развития на протяжении всей жизни человека;
2) упор на «нормальном», или «здоровом», а не на патологическом;
3) особое значение, придаваемое им достижению ощущения самотождественности, своеобразия;
4)
попытки сочетать клинические наблюдения
с изучением культурных и исторических
факторов в объяснении структуры личности
и путей помощи
психолога.
Описанные Эриксоном «восемь возрастных периодов человека» представляют его наиболее оригинальный и важный вклад в теорию личности, оказавший большое влияние на практически каждого профессионального психолога-консультанта. Его попытка показать влияние культуры на развитие личности явилась стимулом для всех, кто изучает поведение человека, разрабатывает новые подходы в изучении главных психологических проблем, с которыми сталкивается сегодня человечество.
Суммируя 15 лет практической и теоретической работы, он выдвинул три новых положения, ставшие тремя важными вкладами в изучение человеческого «Я».Во-первых, Эриксон предположил, что наряду с описанными Фрейдом фазами психосексуального развития, в ходе которого меняется направленность влечения, существуют и психологические стадии развития «Я», в ходе которого индивид устанавливает основные ориентиры по отношению к себе и своей социальной среде.
Во-вторых,
Эриксон утверждал, что становление
личности не заканчивается в подростковом
возрасте, но растягивается на весь
жизненный цикл.
И, наконец, Эриксон говорил, что каждой стадии присущи свои собственные параметры развития, способные принимать положительные и отрицательные значения.
Центральным для созданной Эриксоном теории развития Я является положение о том, что человек в течение жизни проходит через несколько универсальных для всего человечества стадий. Процесс развертывания этих стадий регулируется в соответствии с
«1) в принципе, личность развивается ступенчато, переход от одной ступени к другой предрешен готовностью личности двигаться в направлении дальнейшего роста, расширения сознаваемого социального кругозора и радиуса социального взаимодействия;
2)
общество, в принципе, устроено так, что
развитие социальных возможностей
человека принимается одобрительно,
общество пытается способствовать
сохранению этой тенденции, а также
поддерживать как надлежащий темп, так
и правильную последовательность
развития» (Erikson, 1963a, р. 270).
270).
 Характерные для индивидуума модели
поведения обусловлены тем, каким образом
в конце концов разрешается каждая из
этих задач или как преодолевается
кризис. Конфликты играют жизненно важную
роль в теории Эриксона, потому что рост
и расширение сферы межличностных
отношений связаны с растущей уязвимостью
функций Я на каждой стадии. В то же время
он отмечает, что кризис означает «не
угрозу катастрофы, а поворотный пункт,
и тем самым онтогенетический источник
как силы, так и недостаточной адаптации».
Характерные для индивидуума модели
поведения обусловлены тем, каким образом
в конце концов разрешается каждая из
этих задач или как преодолевается
кризис. Конфликты играют жизненно важную
роль в теории Эриксона, потому что рост
и расширение сферы межличностных
отношений связаны с растущей уязвимостью
функций Я на каждой стадии. В то же время
он отмечает, что кризис означает «не
угрозу катастрофы, а поворотный пункт,
и тем самым онтогенетический источник
как силы, так и недостаточной адаптации».|
Стадия |
Возраст |
Психосоциальный кризис |
Сильная сторона |
|
1. Орально-сенсорная |
Рождение – 1 год |
Базальное доверие – базальное недоверие |
Надежда |
|
2. |
1 – 3 года |
Самостоятельность – стыд и сомнение |
Сила воли |
|
3. Локомоторно-генитальная |
3 – 6 лет |
Инициативность – вина |
Цель |
|
4. Латентная |
6 – 12 лет |
Трудолюбие – неполноценность |
Компетентность |
|
5. Подростковая |
12 – 19 лет |
Самотождественность Я – ролевое смешение |
Верность |
|
6. Ранняя зрелость |
20 – 25 лет |
Близость – изоляция |
Любовь |
|
7. Средняя зрелость |
26 – 64 года |
Продуктивность – застой |
Забота |
|
8. |
65 лет – смерть |
Цельность Я – отчаяние |
Мудрость |
 Мышечно-анальная
Мышечно-анальная Поздняя зрелость
Поздняя зрелостьДоверие и недоверие. Первая стадия развития человека соответствует оральной фазе классического психоанализа и обычно охватывает первый год жизни. В этот период, считает Эриксон, развивается параметр социального взаимодействия, положительным полюсом которого служит доверие, а отрицательным — недоверие.
Степень
доверия, которым ребенок проникается
к окружающему миру, к другим людям и к
самому себе, в значительной степени
зависит от проявляемой к нему заботы.
Младенец, который получает все, что
хочет, потребности которого быстро
удовлетворяются, который никогда долго
не испытывает недомогания, которого
баюкают и ласкают, с которым играют
и разговаривают, чувствует, что мир, в
общем, место уютное, а люди—существа
отзывчивые и услужливые. Если же ребенок
не получает должного ухода, не встречает
любовной заботы, то в нем вырабатывается
недоверие — боязливость и подозрительность
по отношению к миру вообще, к людям в
частности, и недоверие это он несет
с собой в другие стадии его развития.
Необходимо подчеркнуть, однако, что вопрос о том, какое начало одержит верх, не решается раз и навсегда в первый год жизни, но возникает заново на каждой последующей стадии развития. Это и несет надежду и таит угрозу. Ребенок, который приходит в школу с чувством настороженности, может постепенно проникнуться доверием к какой-нибудь учительнице, не допускающей несправедливости по отношению к детям. При этом он может преодолеть первоначальную недоверчивость. Но зато и ребенок, выработавший в младенчестве доверчивый подход к жизни, может проникнуться к ней недоверием на последующих стадиях развития, если, скажем, в случае развода родителей в семье создается обстановка, переполненная взаимными обвинениями и скандалами.
Эпигенетический принцип Эриксона
Эпигенетический принцип
Центральным для созданной Эриксоном теории развития эго является положение о том, что человек в течение жизни проходит через несколько универсальных для всего человечества стадий. Процесс развертывания этих стадий регулируется в соответствии с эпигенетическим принципом созревания. Под этим Эриксон понимает следующее:
Процесс развертывания этих стадий регулируется в соответствии с эпигенетическим принципом созревания. Под этим Эриксон понимает следующее:
1) в принципе, личность развивается ступенчато, переход от одной ступени к другой предрешен готовностью личности двигаться в направлении дальнейшего роста, расширения осознаваемого социального кругозора и радиуса социального взаимодействия;
2) общество, в принципе, устроено так, что развитие социальных возможностей человека принимается одобрительно, общество пытается способствовать сохранению этой тенденции, а также поддерживать как надлежащий темп, так и правильную последовательность развития»
В книге «Детство и общество» (1963а) Эриксон разделил жизнь человека на восемь отдельных стадий психосоциального развития эго (как говорят, на «восемь возрастов человека»). Согласно его утверждению, эти стадии являются результатом эпигенетически развертывающегося «плана личности», который наследуется генетически. Эпигенетическая концепция развития (по-гречески «επι» означает «после», a «γενεσιζ» — «рождение, происхождение») базируется на представлении о том, что каждая стадия жизненного цикла наступает в определенное для нее время («критический период»), а также о том, что полноценно функционирующая личность формируется только путем прохождения в своем развитии последовательно всех стадий.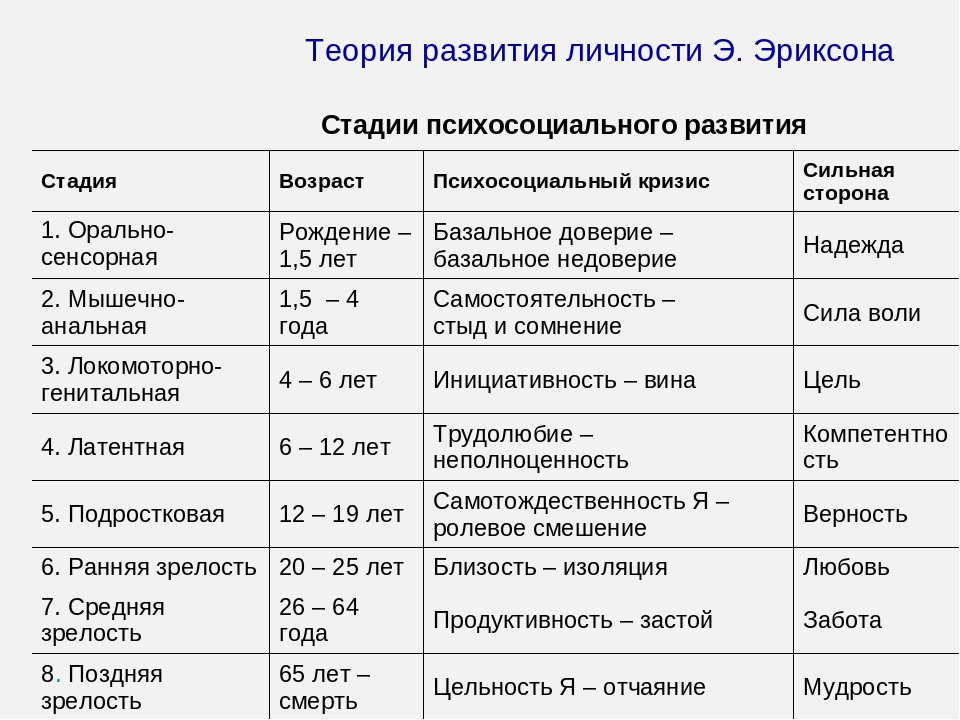 Кроме того, согласно Эриксону, каждая психосоциальная стадия сопровождается кризисом — поворотным моментом в жизни индивидуума, который возникает как следствие достижения определенного уровня психологической зрелости и социальных требований, предъявляемых к индивидууму на этой стадии. Иначе говоря, каждая из восьми фаз жизненного цикла человека характеризуется специфичной именно для данной фазы («фазо-специфической») эволюционной задачей — проблемой в социальном развитии, которая в свое время предъявляется индивидууму, но не обязательно находит свое разрешение. Характерные для индивидуума модели поведения обусловлены тем, каким образом в конце концов разрешается каждая из этих задач или как преодолевается кризис. Конфликты играют жизненно важную роль в теории Эриксона, потому что рост и расширение сферы межличностных отношений связаны с растущей уязвимостью функций эго на каждой стадии. В то же время он отмечает, что кризис означает «не угрозу катастрофы, а поворотный пункт, и тем самым онтогенетический источник как силы, так и недостаточной адаптации» [Erikson, 1968, р.
Кроме того, согласно Эриксону, каждая психосоциальная стадия сопровождается кризисом — поворотным моментом в жизни индивидуума, который возникает как следствие достижения определенного уровня психологической зрелости и социальных требований, предъявляемых к индивидууму на этой стадии. Иначе говоря, каждая из восьми фаз жизненного цикла человека характеризуется специфичной именно для данной фазы («фазо-специфической») эволюционной задачей — проблемой в социальном развитии, которая в свое время предъявляется индивидууму, но не обязательно находит свое разрешение. Характерные для индивидуума модели поведения обусловлены тем, каким образом в конце концов разрешается каждая из этих задач или как преодолевается кризис. Конфликты играют жизненно важную роль в теории Эриксона, потому что рост и расширение сферы межличностных отношений связаны с растущей уязвимостью функций эго на каждой стадии. В то же время он отмечает, что кризис означает «не угрозу катастрофы, а поворотный пункт, и тем самым онтогенетический источник как силы, так и недостаточной адаптации» [Erikson, 1968, р. 286].
286].
Каждый психосоциальный кризис, если рассматривать его с точки зрения оценки, содержит и позитивный, и негативный компоненты. Если конфликт разрешен удовлетворительно (то есть на предыдущей стадии эго обогатилось новыми положительными качествами), то теперь эго вбирает в себя новый позитивный компонент (например, базальное доверие и автономию), и это гарантирует здоровое развитие личности в дальнейшем. Напротив, если конфликт остается неразрешенным или получает неудовлетворительное разрешение, развивающемуся эго тем самым наносится вред, и в него встраивается негативный компонент (например, базальное недоверие, стыд и сомнения). Хотя на пути развития личности возникают теоретически предсказуемые и вполне определенные конфликты, из этого не следует, что на предшествующих стадиях успехи и неудачи обязательно одни и те же. Качества, которые эго приобретает на каждой стадии, не снижают его восприимчивости к новым внутренним конфликтам или меняющимся условиям [Erikson, 1964a]. Задача состоит в том, чтобы человек адекватно разрешал каждый кризис, и тогда у него будет возможность подойти к следующей стадии развития более адаптивной и зрелой личностью.
Задача состоит в том, чтобы человек адекватно разрешал каждый кризис, и тогда у него будет возможность подойти к следующей стадии развития более адаптивной и зрелой личностью.
Развитие личности: психосоциальные стадии
1. Младенчество: базальное доверие—базальное недоверие
Первая психосоциальная стадия соответствует оральной стадии по Фрейду и охватывает первый год жизни. По Эриксону, в этот период краеугольным камнем формирования здоровой личности является общее чувство доверия; другие ученые называют ту же самую характеристику «уверенностью». Младенец, имеющий базальное чувство «внутренней определенности», воспринимает социальный мир как безопасное, стабильное место, а людей как заботливых и надежных. Это чувство определенности лишь частично осознается в период младенчества.
Согласно Эриксону, степень развития у ребенка чувства доверия к другим людям и миру зависит от качества получаемой им материнской заботы.
«Я полагаю, что матери формируют чувство доверия у своих детей благодаря такому обращению, которое по своей сути состоит из чуткой заботы об индивидуальных потребностях ребенка и отчетливого ощущения того, что она сама — тот человек, которому можно доверять, в том понимании слова «доверие», которое существует в данной культуре применительно к данному стилю жизни. Благодаря этому у ребенка закладывается основа для чувства «все хорошо»; для появления чувства тождества; для становления тем, кем он станет, согласно надеждам других» [Erikson, 1963a, р. 249].
Благодаря этому у ребенка закладывается основа для чувства «все хорошо»; для появления чувства тождества; для становления тем, кем он станет, согласно надеждам других» [Erikson, 1963a, р. 249].
Таким образом, чувство доверия не зависит от количества пищи или от проявлений родительской нежности; скорее оно связано со способностью матери передать своему ребенку чувство узнаваемости, постоянства и тождества переживаний. Эриксон также подчеркивает: младенцы должны доверять не только внешнему миру, но также и миру внутреннему, они должны научиться доверять себе и в особенности должны приобрести способность к тому, чтобы их органы эффективно справлялись с биологическими побуждениями. Подобное поведение мы наблюдаем тогда, когда младенец может переносить отсутствие матери без чрезмерного страдания и тревоги по поводу «отделения» от нее.
Вопрос о том, что является причиной первого важного психологического кризиса, глубоко проанализирован Эриксоном. Он связывает этот кризис с качеством материнского ухода за ребенком — причиной кризиса является ненадежность, несостоятельность матери и отвергание ею ребенка. Это способствует появлению у него психосоциальной установки страха, подозрительности и опасений за свое благополучие. Данная установка направлена как на мир в целом, так и на отдельных людей; она будет проявляться во всей своей полноте на более поздних стадиях личностного развития. Эриксон также считает, что чувство недоверия может усилиться тогда, когда ребенок перестает быть для матери главным центром внимания; когда она возвращается к тем занятиям, которые оставила на время беременности (скажем, возобновляет прерванную карьеру), или рожает следующего ребенка. Наконец, родители, придерживающиеся противоположных принципов и методов воспитания, или чувствующие себя неуверенно в роли родителей, или те, чья система ценностей находится в противоречии с общепринятым в данной культуре стилем жизни, могут создавать для ребенка атмосферу неопределенности, двусмысленности, в результате чего у него появляется чувство недоверия. Согласно Эриксону, поведенческими последствиями подобного неблагополучного развития являются острая депрессия у младенцев и паранойя у взрослых.
Это способствует появлению у него психосоциальной установки страха, подозрительности и опасений за свое благополучие. Данная установка направлена как на мир в целом, так и на отдельных людей; она будет проявляться во всей своей полноте на более поздних стадиях личностного развития. Эриксон также считает, что чувство недоверия может усилиться тогда, когда ребенок перестает быть для матери главным центром внимания; когда она возвращается к тем занятиям, которые оставила на время беременности (скажем, возобновляет прерванную карьеру), или рожает следующего ребенка. Наконец, родители, придерживающиеся противоположных принципов и методов воспитания, или чувствующие себя неуверенно в роли родителей, или те, чья система ценностей находится в противоречии с общепринятым в данной культуре стилем жизни, могут создавать для ребенка атмосферу неопределенности, двусмысленности, в результате чего у него появляется чувство недоверия. Согласно Эриксону, поведенческими последствиями подобного неблагополучного развития являются острая депрессия у младенцев и паранойя у взрослых.
Основная предпосылка психосоциальной теории состоит в том, что кризис «доверие — недоверие» (или любой другой последующий кризис) не всегда находит разрешение в течение первого или второго года жизни. В соответствии с эпигенетическим принципом, дилемма доверие—недоверие будет проявляться снова и снова на каждой последующей стадии развития, хотя она и является центральной для периода младенчества. Адекватное разрешение кризиса доверия имеет важные последствия для развития личности ребенка в дальнейшем. Укрепление доверия к себе и к матери дает возможность ребенку переносить состояния фрустрации, которые он неизбежно будет переживать на протяжении следующих стадий своего развития.
Как отмечает Эриксон, здоровое развитие младенца не является результатом исключительно чувства доверия, но скорее обусловлено благоприятным соотношением доверия и недоверия. Понять, чему не следует доверять, так же важно, как и понять, чему доверять необходимо. Эта способность предвидеть опасность и дискомфорт также важна для совладания с окружающей реальностью и для эффективного принятия решений; поэтому базальное доверие не стоит трактовать в контексте шкалы достижений. Эриксон заявлял, что животные обладают почти инстинктивной готовностью к приобретению психосоциальных навыков, а у людей психосоциальные способности приобретаются в процессе научения. Кроме того, он утверждал, что в различных культурах и социальных классах матери учат доверию и недоверию по-разному. Но путь приобретения базального доверия по самой сути своей универсален; человек доверяет социуму подобно тому, как он доверяет собственной матери, словно она вот-вот вернется и накормит его в подходящее время подходящей пищей.
Эриксон заявлял, что животные обладают почти инстинктивной готовностью к приобретению психосоциальных навыков, а у людей психосоциальные способности приобретаются в процессе научения. Кроме того, он утверждал, что в различных культурах и социальных классах матери учат доверию и недоверию по-разному. Но путь приобретения базального доверия по самой сути своей универсален; человек доверяет социуму подобно тому, как он доверяет собственной матери, словно она вот-вот вернется и накормит его в подходящее время подходящей пищей.
Положительное психосоциальное качество, приобретаемое в результате успешного разрешения конфликта «доверие—недоверие», Эриксон обозначает термином надежда. Иначе говоря, доверие переходит в способность младенца надеяться, что, в свою очередь, у взрослого может составлять основу веры в соответствии с какой-либо официальной формой религии. Надежда, это первое положительное качество эго, поддерживает убежденность человека в значимости и надежности общего культурного пространства.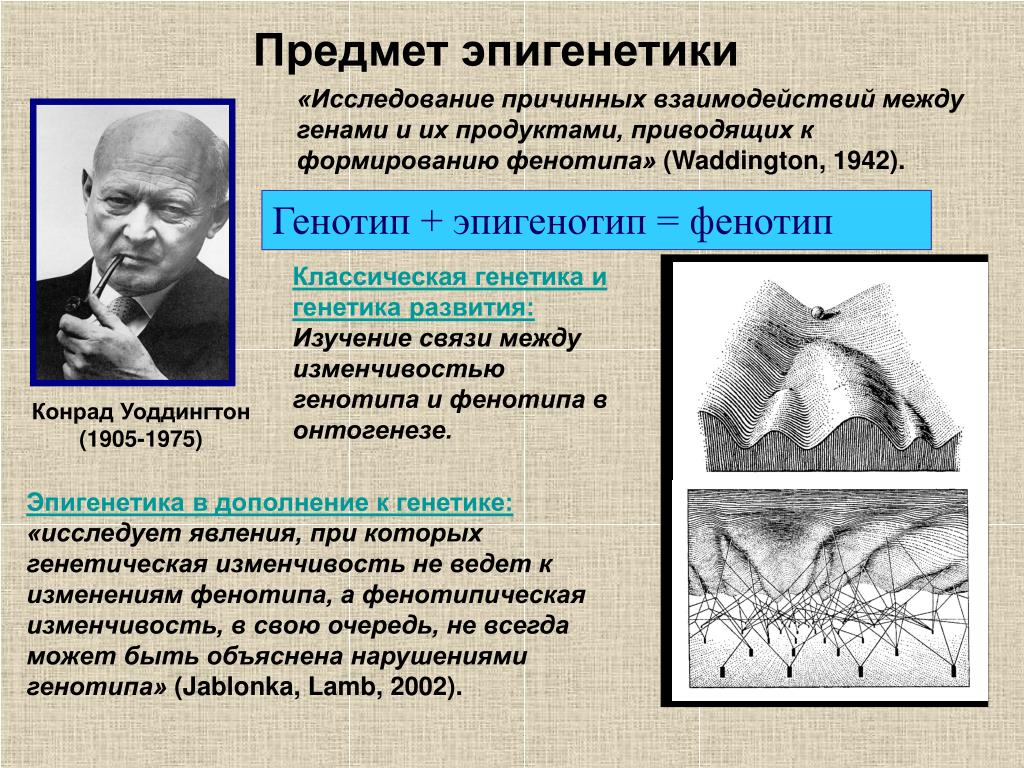 Эриксон подчеркивает: когда институт религии утрачивает для индивидуума свое ощутимое значение, он становится неуместным, устаревает и, возможно, на смену ему даже приходят другие, более значимые источники веры и уверенности в будущем (например, достижения науки, искусства и общественной жизни).
Эриксон подчеркивает: когда институт религии утрачивает для индивидуума свое ощутимое значение, он становится неуместным, устаревает и, возможно, на смену ему даже приходят другие, более значимые источники веры и уверенности в будущем (например, достижения науки, искусства и общественной жизни).
2. Раннее детство: автономия—стыд и сомнение
Приобретение чувства базального доверия подготавливает почву для достижения определенной автономии и самоконтроля, избегания чувств стыда, сомнения и унижения. Этот период соответствует анальной стадии, по Фрейду, и продолжается в течение второго и третьего годов жизни. Согласно Эриксону, ребенок, взаимодействуя с родителями в процессе обучения туалетному поведению, обнаруживает, что родительский контроль бывает разным: с одной стороны, он может проявляться как форма заботы, с другой — как деструктивная форма обуздания и мера пресечения. Ребенок также научается различать предоставление свободы типа «пусть попробует» и, напротив, попустительство как деструктивную форму избавления от хлопот. Эта стадия становится решающей для установления соотношения между добровольностью и упрямством. Чувство самоконтроля без потери самооценки является онтогенетическим источником уверенности в свободном выборе; чувство чрезмерного постороннего контроля и одновременная потеря самоконтроля может послужить толчком для постоянной склонности к сомнениям и стыду [Erikson, 1968b].
Эта стадия становится решающей для установления соотношения между добровольностью и упрямством. Чувство самоконтроля без потери самооценки является онтогенетическим источником уверенности в свободном выборе; чувство чрезмерного постороннего контроля и одновременная потеря самоконтроля может послужить толчком для постоянной склонности к сомнениям и стыду [Erikson, 1968b].
До наступления этой стадии дети почти полностью зависят от заботящихся о них людей. Однако, поскольку у них быстро развивается нервно-мышечная система, речь и социальная избирательность, они начинают исследовать свое окружение и взаимодействовать с ним более независимо. В особенности они гордятся своими только что обнаруженными локомоторными навыками и все хотят делать сами (например, умываться, одеваться и есть). Мы наблюдаем у них огромное желание исследовать предметы и манипулировать ими, а также установку по отношению к родителям: «Я сам» и «Я — то, что я могу».
С точки зрения Эриксона, удовлетворительное разрешение психосоциального кризиса на этой стадии зависит прежде всего от готовности родителей постепенно предоставлять детям свободу самим осуществлять контроль над своими действиями. В то же время он подчеркивает, что родители должны ненавязчиво, но четко ограничивать ребенка в тех сферах жизни, которые потенциально или актуально представляются опасными как для самих детей, так и для окружающих. Автономия не означает, что ребенок получает неограниченную свободу. Скорее она означает, что родители должны удерживать возрастающую способность ребенка делать выбор в пределах определенных «степеней свободы».
В то же время он подчеркивает, что родители должны ненавязчиво, но четко ограничивать ребенка в тех сферах жизни, которые потенциально или актуально представляются опасными как для самих детей, так и для окружающих. Автономия не означает, что ребенок получает неограниченную свободу. Скорее она означает, что родители должны удерживать возрастающую способность ребенка делать выбор в пределах определенных «степеней свободы».
Эриксон рассматривает переживание стыда у ребенка как нечто родственное гневу, направленному на самого себя, когда ребенку не разрешается развивать свою автономию и самоконтроль. Стыд может появиться, если родители нетерпеливо, раздраженно и настойчиво делают за детей что-то, что те могут сделать сами; или, наоборот, когда родители ожидают, что дети сделают то, чего они еще сами сделать не в состоянии. Разумеется, каждый родитель хотя бы однажды подталкивал своего ребенка к действиям, которые на самом деле лежат за пределами разумных ожиданий. Но только в тех случаях, когда родители постоянно чрезмерно опекают ребенка или остаются глухими к его потребностям, у него появляется либо преобладающее чувство стыда перед другими, либо сомнения в своей способности контролировать окружающий мир и владеть собой.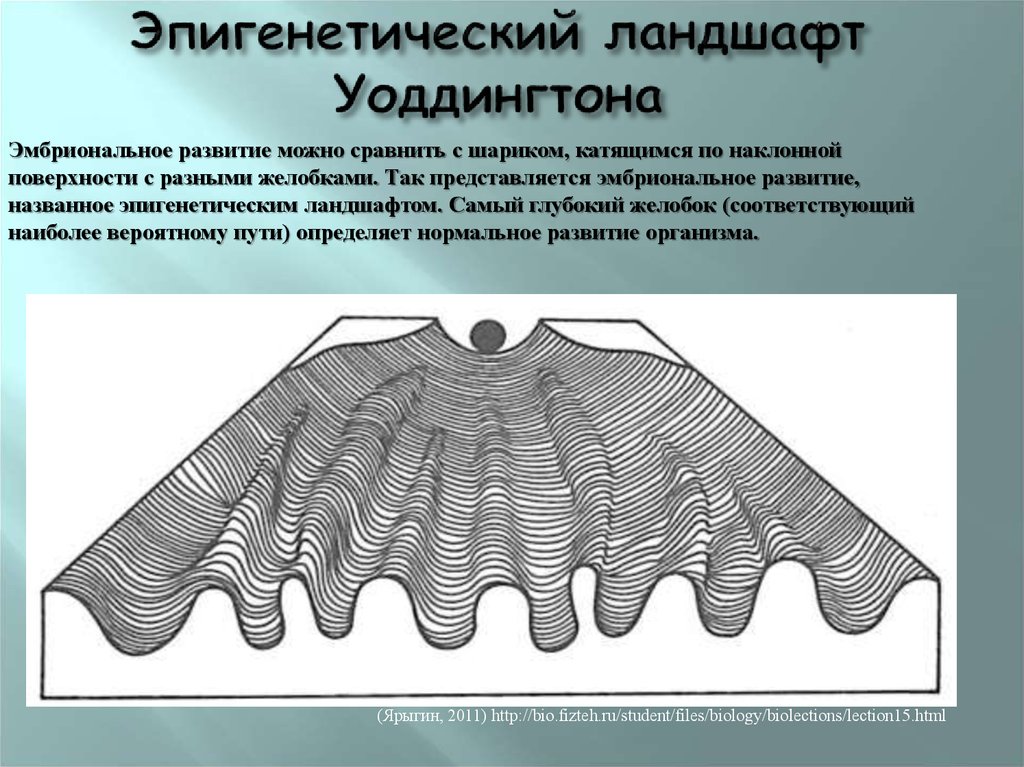 Вместо того, чтобы быть уверенными в себе и ладить с окружением, такие дети думают, что другие пристально их рассматривают, относятся с подозрением и неодобрением; или же они считают себя совершенно несчастными. У них слабая «сила воли» — они пасуют перед теми, кто над ними главенствует или их эксплуатирует. В результате формируются такие черты, как неуверенность в себе, приниженность и слабоволие.
Вместо того, чтобы быть уверенными в себе и ладить с окружением, такие дети думают, что другие пристально их рассматривают, относятся с подозрением и неодобрением; или же они считают себя совершенно несчастными. У них слабая «сила воли» — они пасуют перед теми, кто над ними главенствует или их эксплуатирует. В результате формируются такие черты, как неуверенность в себе, приниженность и слабоволие.
По убеждению Эриксона, приобретение ребенком постоянного чувства автономии в значительной степени укрепляет у него чувство доверия. Эта взаимозависимость доверия и автономии может иногда замедлять будущее психическое развитие. Например, дети с неустойчивым чувством доверия могут на стадии автономии стать нерешительными, робкими, могут бояться отстаивать свои права, так что будут искать помощи и поддержки у окружающих. В зрелости у таких людей наиболее вероятно проявится обсессивно-компульсивная симптоматика (что обеспечивает им необходимый контроль) или паранойяльный страх преследования.
Социальное дополнение автономии — система правопорядка. Эриксон использует термины «право» и «порядок», невзирая на возможные эмоциональные коннотации. Согласно его теории, родители должны быть всегда справедливыми и уважать права и привилегии других, если они хотят, чтобы их дети были готовы» в зрелом возрасте принять ограниченную автономию.
3. Возраст игры: инициативность—вина
Конфликт между инициативой и виной — последний психосоциальный конфликт в дошкольном периоде, который Эриксон называл «возрастом игры». Он соответствует фаллической стадии в теории Фрейда и длится от четырех лет до поступления ребенка в школу. В это время социальный мир ребенка требует от него активности, решения новых задач и приобретения новых навыков; похвала является наградой за успехи. Кроме того, у детей появляется дополнительная ответственность за себя и за то, что составляет их мир (игрушки, домашние животные и, возможно, братья и сестры). Они начинают интересоваться трудом других, пробовать новое и допускать, что и на других людях в их окружении лежит определенная ответственность. Успехи в освоении речи и развитие моторики дают возможность контактировать со сверстниками и более старшими детьми за пределами дома, что позволяет им участвовать в разнообразных общественных играх. Это возраст, когда дети начинают чувствовать, что их воспринимают как людей и считаются с ними и что жизнь для них имеет цель. «Я — то, что я буду» — становится у ребенка главным чувством идентичности во время периода игры. Процитируем Эриксона:
Успехи в освоении речи и развитие моторики дают возможность контактировать со сверстниками и более старшими детьми за пределами дома, что позволяет им участвовать в разнообразных общественных играх. Это возраст, когда дети начинают чувствовать, что их воспринимают как людей и считаются с ними и что жизнь для них имеет цель. «Я — то, что я буду» — становится у ребенка главным чувством идентичности во время периода игры. Процитируем Эриксона:
Эпигенетика: принципы и практика — PMC
- Список журналов
- Копать Дис
- PMC3134032
В качестве библиотеки NLM предоставляет доступ к научной литературе. Включение в базу данных NLM не означает одобрения или согласия с
содержание NLM или Национальных институтов здравоохранения.
Узнайте больше о нашем отказе от ответственности.
Копать Дис. 2011 июль; 29(2): 130–135.
doi: 10.1159/000323874
Информация об авторе Информация об авторских правах и лицензиях Отказ от ответственности
Эпигенетика определяется как наследуемые изменения в экспрессии генов, которые, в отличие от мутаций, не связаны с изменениями в последовательности ДНК. Преобладающими эпигенетическими механизмами являются метилирование ДНК, модификации хроматина, потеря импринтинга и некодирующая РНК. Эпигенетическая регуляция экспрессии генов оказывает долгосрочное и широкомасштабное влияние на здоровье. Диета и воздействие окружающей среды потенциально могут изменить уровень и объем эпигенетической регуляции, поэтому интересные разработки в области изучения эпигенетики могут объяснить корреляции, обнаруженные исследователями между образом жизни и риском заболевания. Аберрантные эпигенетические паттерны связаны с рядом заболеваний органов пищеварения, включая пищевод Барретта, цирроз печени, воспалительные заболевания кишечника и многочисленные злокачественные новообразования желудочно-кишечного тракта.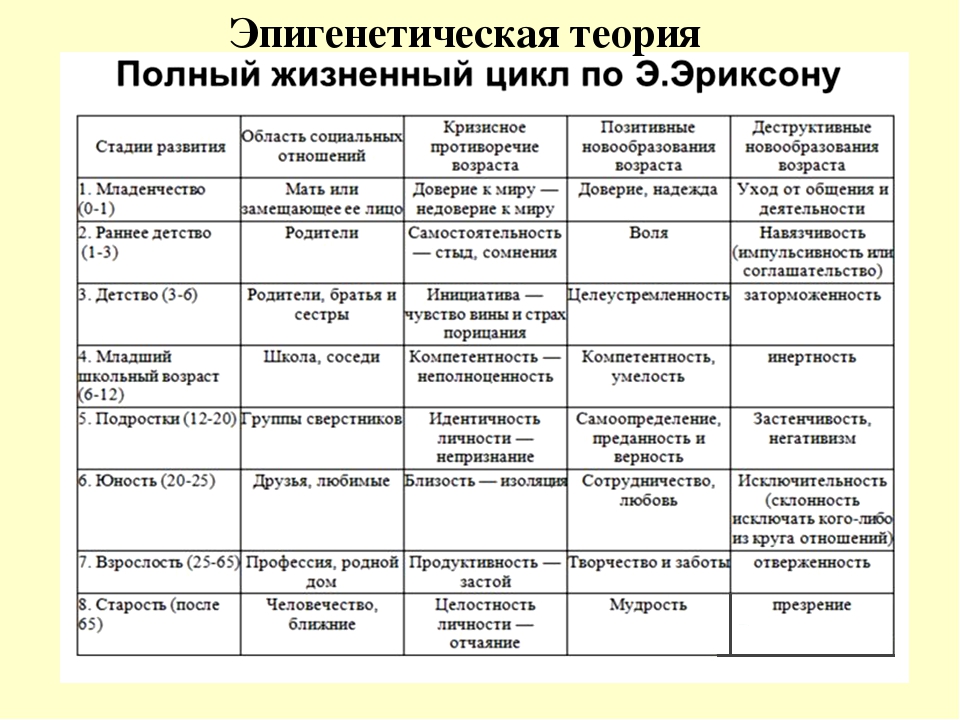
Ключевые слова: Эпигенетика, гиперметилирование, гипометилирование, потеря импринтинга, модификации гистонов, микроРНК
Эпигенетика определяется как наследуемые изменения в экспрессии генов, которые, в отличие от мутаций, не связаны с изменениями в последовательности ДНК. Преобладающими эпигенетическими механизмами являются метилирование ДНК, модификации структуры хроматина, потеря импринтинга и некодирующая РНК [1].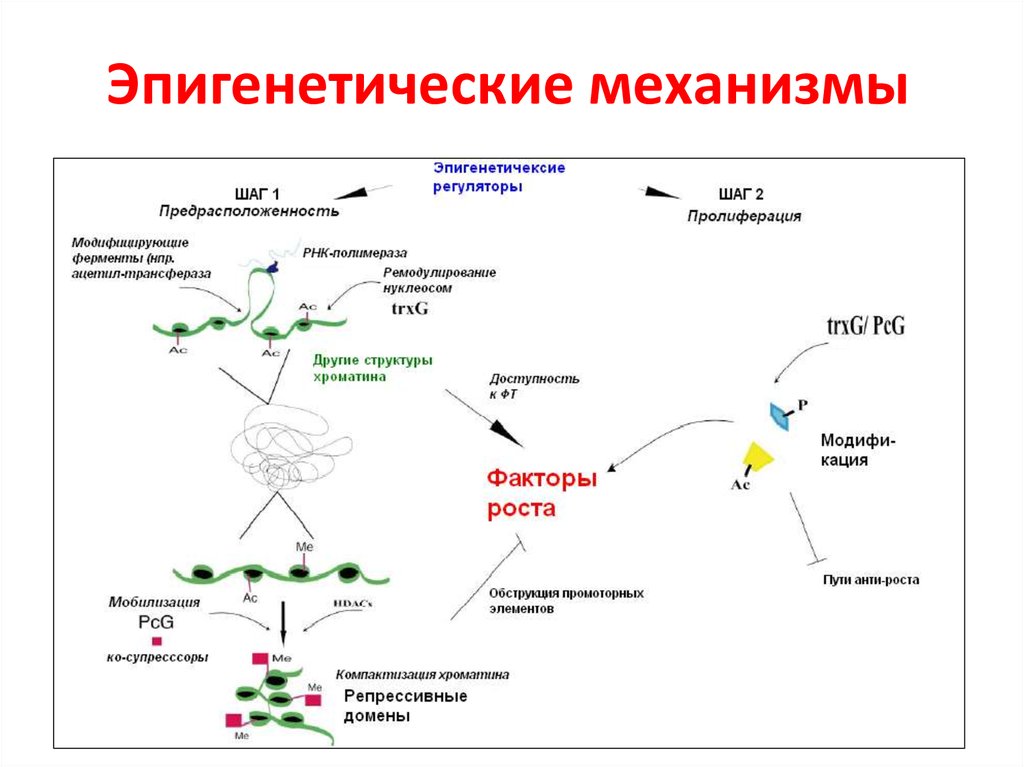 Важной особенностью эпигенетических модификаций является то, что они могут наследоваться между материнскими и дочерними клетками (митотическое наследование) и между поколениями (мейотическое наследование). Эпигенетика является одним из объяснений того, как клетки и организмы с идентичной ДНК могут иметь такие разительные фенотипические различия. Диета и воздействие окружающей среды потенциально могут изменить уровень и объем эпигенетической регуляции, поэтому интересные разработки в области изучения эпигенетики могут объяснить корреляции, обнаруженные исследователями между образом жизни и риском заболевания [2]. Кроме того, эпигенетическая регуляция экспрессии генов стала фундаментальным путем в патогенезе многих заболеваний, в частности злокачественных новообразований [3,4]. Заболевания пищеварительной системы не являются исключением, ведь многие захватывающие открытия в области эпигенетики в целом были сделаны при изучении рака желудочно-кишечного тракта, печени и гепатобилиарного дерева.
Важной особенностью эпигенетических модификаций является то, что они могут наследоваться между материнскими и дочерними клетками (митотическое наследование) и между поколениями (мейотическое наследование). Эпигенетика является одним из объяснений того, как клетки и организмы с идентичной ДНК могут иметь такие разительные фенотипические различия. Диета и воздействие окружающей среды потенциально могут изменить уровень и объем эпигенетической регуляции, поэтому интересные разработки в области изучения эпигенетики могут объяснить корреляции, обнаруженные исследователями между образом жизни и риском заболевания [2]. Кроме того, эпигенетическая регуляция экспрессии генов стала фундаментальным путем в патогенезе многих заболеваний, в частности злокачественных новообразований [3,4]. Заболевания пищеварительной системы не являются исключением, ведь многие захватывающие открытия в области эпигенетики в целом были сделаны при изучении рака желудочно-кишечного тракта, печени и гепатобилиарного дерева.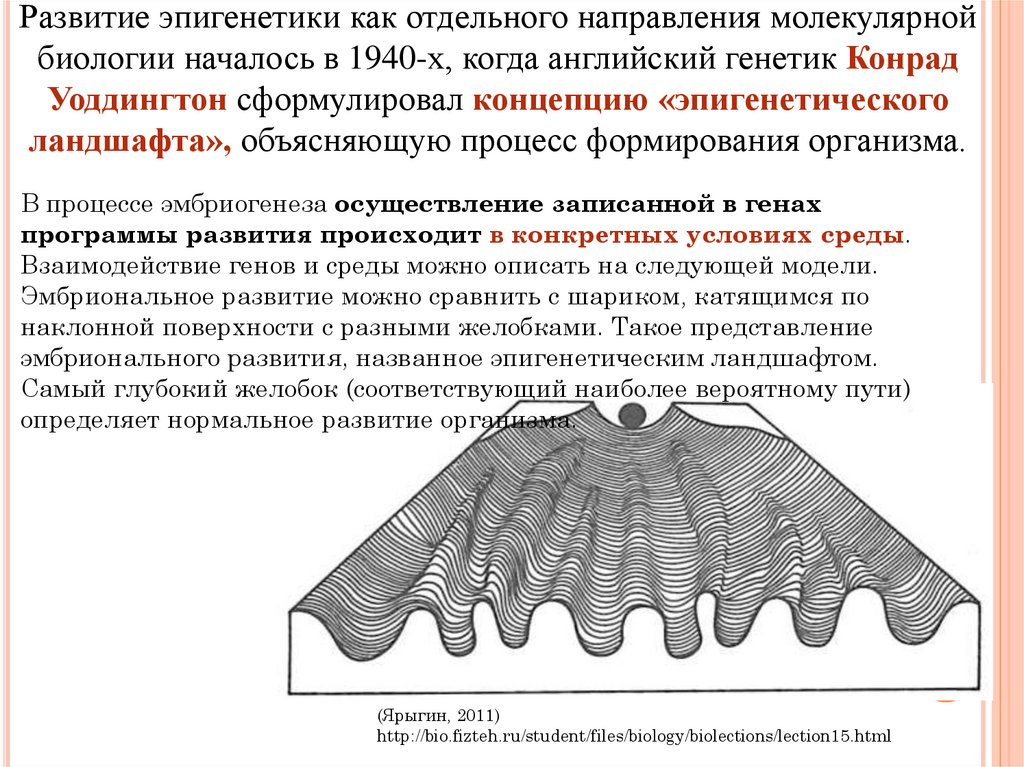 Эпигенетические модификации ДНК дают надежду и обещают новые биомаркеры для раннего обнаружения, прогнозирования и ответа на лечение рака. Кроме того, обращение эпигенетических изменений представляет собой потенциальную цель новых терапевтических стратегий и дизайна лекарств [5]. В последнее десятилетие было опубликовано множество данных и исследований, касающихся эпигенетических механизмов и их участия в развитии фенотипов и заболеваний. Настоящий отчет призван представить введение в область эпигенетики с примерами некоторых наиболее ярких и провокационных исследований, связанных с этой темой.
Эпигенетические модификации ДНК дают надежду и обещают новые биомаркеры для раннего обнаружения, прогнозирования и ответа на лечение рака. Кроме того, обращение эпигенетических изменений представляет собой потенциальную цель новых терапевтических стратегий и дизайна лекарств [5]. В последнее десятилетие было опубликовано множество данных и исследований, касающихся эпигенетических механизмов и их участия в развитии фенотипов и заболеваний. Настоящий отчет призван представить введение в область эпигенетики с примерами некоторых наиболее ярких и провокационных исследований, связанных с этой темой.
Метилирование ДНК представляет собой ковалентное присоединение или вычитание метильной группы к цитозиновому нуклеотиду в последовательности ДНК. Метилирование контролируется семейством специфических ферментов, известных как ДНК-метилтрансферазы (DNMT). У позвоночных добавление метильной группы происходит только к цитозину, предшествующему гуанину (динуклеотид CpG) [1]. Области генома, богатые последовательностями цитозина, предшествующего гуанину, известны как островки CpG.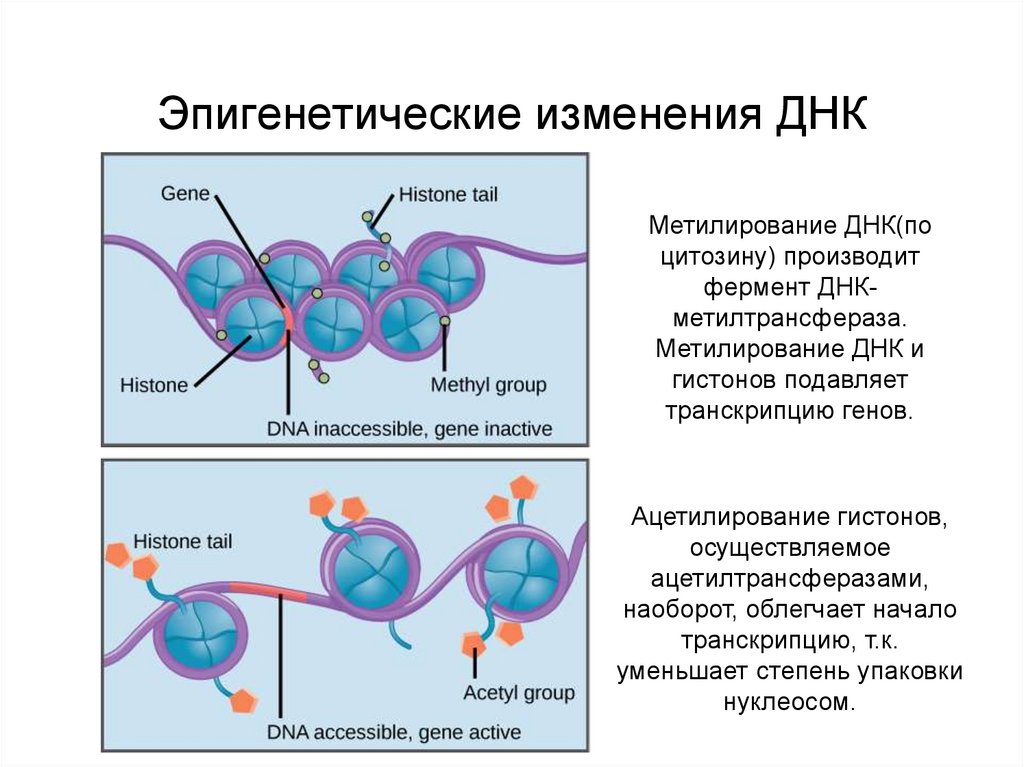 Фактически островки CpG существуют в промоторных областях примерно половины всех генов.
Фактически островки CpG существуют в промоторных областях примерно половины всех генов.
Гипометилирование
Приблизительно 80% динуклеотидов CpG за пределами промоторных областей метилированы при нормальных физиологических условиях. Полногеномное снижение метилирования или гипометилирования наиболее функционально значимо, когда оно происходит в кодирующих областях генов, приводя к альтернативным версиям или уровням матричной РНК. Глобальное гипометилирование ДНК из опухолей больных раком толстой кишки было одной из первых описанных эпигенетических аномалий [6]. Предполагается, что гипометилирование способствует канцерогенезу, способствуя митотической рекомбинации, что приводит к делециям, транслокациям и хромосомным перестройкам, что в совокупности известно как нестабильность генома. Гипометилирование ДНК (также известное как деметилирование) также связано с активацией протоонкогенов, таких как c-Jun, c-Myc и c-Ha-Ras [7]. В целом, большая часть ДНК в опухолях гипометилирована, с редким геноспецифическим гиперметилированием (рис.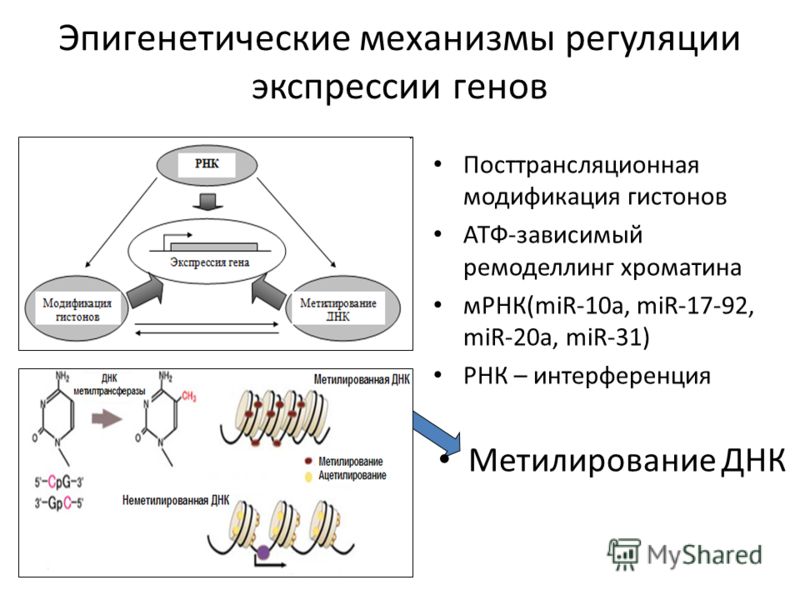 ) [8]. В целом гипометилирование увеличивается по мере прогрессирования опухоли. Соответственно, гипометилирование повторяющихся элементов ДНК, таких как SAT2, LINE1 и ALU, происходит в многоступенчатом процессе гепатоканцерогенеза и коррелирует с неблагоприятным прогнозом [9].].
) [8]. В целом гипометилирование увеличивается по мере прогрессирования опухоли. Соответственно, гипометилирование повторяющихся элементов ДНК, таких как SAT2, LINE1 и ALU, происходит в многоступенчатом процессе гепатоканцерогенеза и коррелирует с неблагоприятным прогнозом [9].].
Открыть в отдельном окне
Гипо- и гиперметилирование ДНК. Черная линия представляет ген, а серый прямоугольник — промотор. В нормальных условиях CpG вне промоторной области метилированы. При раке может происходить метилирование CpG в промоторной области и гипометилирование CpG вне промоторной области. Гиперметилирование CpG в промоторной области может приводить к репрессии транскрипции. Адаптировано из Herman and Baylin [10].
Гиперметилирование
Добавление метильных групп или гиперметилирование может быть очень специфичным для конкретного гена. Гиперметилирование островков CpG в промоторной области гена может привести к подавлению транскрипции гена и последующей потере экспрессии белка. Таким образом, гиперметилирование генов-супрессоров опухолей в настоящее время признано средством подавления генов, альтернативным мутации или потере аллелей [10]. Метильные группы выступают в большую бороздку ДНК, тем самым изменяя биофизические свойства ДНК. Эти изменения могут положительно или отрицательно влиять на связывающие способности ДНК и некоторых белков. Например, если РНК-полимераза 1 не может правильно связываться с ДНК, то нарушается процесс транскрипции ДНК в РНК. Во время митоза паттерны метилирования копируются с материнской цепи ДНК на дочернюю цепь ДНК с помощью DNMT1.
Таким образом, гиперметилирование генов-супрессоров опухолей в настоящее время признано средством подавления генов, альтернативным мутации или потере аллелей [10]. Метильные группы выступают в большую бороздку ДНК, тем самым изменяя биофизические свойства ДНК. Эти изменения могут положительно или отрицательно влиять на связывающие способности ДНК и некоторых белков. Например, если РНК-полимераза 1 не может правильно связываться с ДНК, то нарушается процесс транскрипции ДНК в РНК. Во время митоза паттерны метилирования копируются с материнской цепи ДНК на дочернюю цепь ДНК с помощью DNMT1.
Гиперметилирование генов, участвующих в клеточном цикле, репарации ДНК, ангиогенезе, метаболизме канцерогенов, апоптозе и межклеточном взаимодействии вовлечено в канцерогенез. Следует отметить, что гиперметилирование также происходит как нормальный физиологический процесс, например, при инактивации второй Х-хромосомы (тельце Барра) у женщин. Кроме того, гиперметилирование является физиологическим процессом, связанным со старением, и индуцированная метилированием репрессия транскрипции повторяющихся элементов ДНК помогает предотвратить хромосомную нестабильность.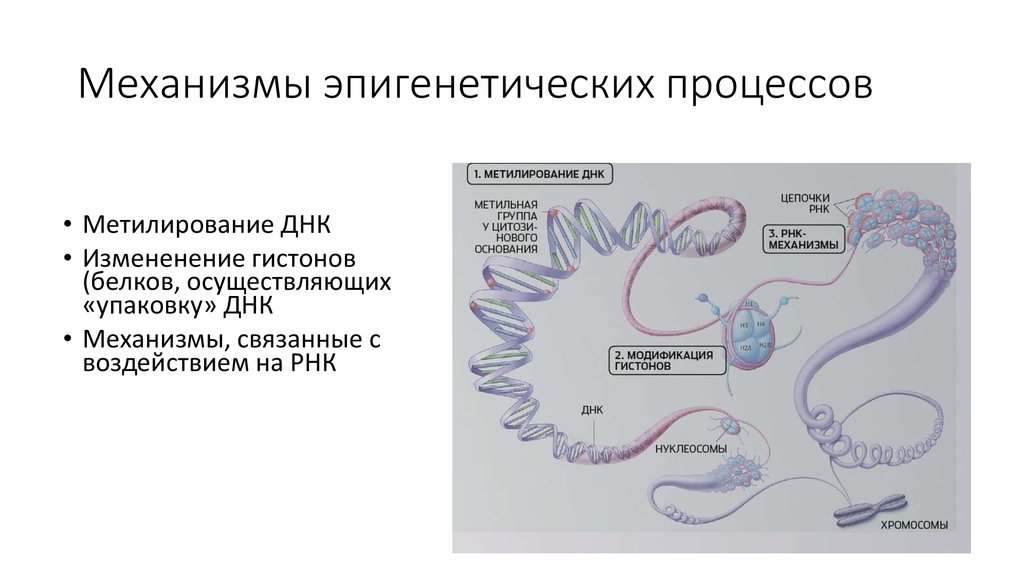
Методы, используемые для обнаружения метилирования
В лаборатории метилирование ДНК можно измерить различными методами в тканях, а иногда и в периферической крови или других выделениях организма, таких как желчь. Одним из основных преимуществ попыток обнаружения метилирования является присущая ДНК стабильность. До появления методов секвенирования метилирования ДНК для обнаружения метилирования ДНК использовались изошизомеры с различной чувствительностью к метилированию. Основным недостатком этого метода является то, что в любой заданной последовательности ДНК можно оценить менее 5% метилированных цитозинов. С начала 1990-х годов, ключевой метод, с помощью которого определяются уровни метилирования, требует начальной бисульфитной конверсии ДНК. Эта обработка бисульфитом превращает неметилированные, но не метилированные цитозины в урацилы. Последующее геноспецифическое метилирование можно определить с помощью качественной или количественной ПЦР, специфичной к метилированию (MSP), с использованием праймеров и зондов, специфичных к соответствующей метилированной последовательности ДНК [11]. Преимущества MSP заключаются в том, что он дает положительное представление о метилированных цитозинах и обеспечивает полный профиль метилирования для определенной последовательности ДНК, а не оценку только нескольких цитозинов в последовательности. Автор предпочитает количественный MSP в реальном времени, поскольку он определяет фактический процент метилированных аллелей в данном образце.
Преимущества MSP заключаются в том, что он дает положительное представление о метилированных цитозинах и обеспечивает полный профиль метилирования для определенной последовательности ДНК, а не оценку только нескольких цитозинов в последовательности. Автор предпочитает количественный MSP в реальном времени, поскольку он определяет фактический процент метилированных аллелей в данном образце.
Секвенирование ДНК также может быть выполнено на бисульфит-конвертированной ДНК для определения конкретных областей гипер- или гипометилирования. Это особенно полезный метод для определения областей дифференциального метилирования и помощи в разработке праймеров и зондов для более специфического MSP. Пиросеквенирование — это метод секвенирования ДНК в реальном времени, основанный на активности ДНК-полимеразы и основанный на люминометрическом обнаружении высвобождения пирофосфата после включения нуклеотида [12]. Преимущество пиросеквенирования заключается в том, что оно сочетает в себе высокую производительность технологий на основе ПЦР с возможностью анализа всех отдельных CpG данного региона [12].
Ограничение технологий на основе ПЦР заключается в том, что они специально нацелены на представляющие интерес гены-кандидаты. Совсем недавно разработка высокопроизводительных полногеномных платформ микрочипов была разработана в попытке определить глобальный паттерн метилирования опухолей. Иммунопреципитация метилированной ДНК (MeDIP) представляет собой иммунологический подход, который обогащает метилированную ДНК и основан на том принципе, что геномная ДНК случайным образом расщепляется ультразвуком и может подвергаться иммунопреципитации антителом, которое специально нацелено на 5-метилцитидин. Этот метод можно использовать для создания полных профилей метилирования геномной ДНК и для выявления аномально (гипер- или гипо-) метилированных генов [13].
Метод амплификации метилированных CpG-островков (MCA) основан на расщеплении геномной ДНК чувствительным к метилированию рестрикционным ферментом Sma I, который разрезает только неметилированные сайты, оставляя тупые концы между C и G. Затем ДНК расщепляется нечувствительным к метилированию изошизомером Sma I, Xma I, который оставляет выступ из четырех оснований. За этими двумя последовательными перевариваниями следует лигирование адаптеров с выступающими частями и, наконец, выполнение специфичной для адаптера ПЦР-амплификации [14]. Таким образом, этот метод приводит к обогащению и амплификации только метилированных фрагментов ДНК. Эти метилированные фрагменты ДНК затем используются для опросов с помощью платформ микрочипов.
Затем ДНК расщепляется нечувствительным к метилированию изошизомером Sma I, Xma I, который оставляет выступ из четырех оснований. За этими двумя последовательными перевариваниями следует лигирование адаптеров с выступающими частями и, наконец, выполнение специфичной для адаптера ПЦР-амплификации [14]. Таким образом, этот метод приводит к обогащению и амплификации только метилированных фрагментов ДНК. Эти метилированные фрагменты ДНК затем используются для опросов с помощью платформ микрочипов.
Дополнительный метод, с помощью которого можно обнаружить новые мишени индуцированного метилированием транскрипционного молчания, заключается в обработке раковых клеточных линий агентами, которые обращают эпигенетические события, а затем в проведении микрочипов экспрессии генов, чтобы определить, какие гены активируются.
Хроматин состоит из гистонов и ДНК. Гистоны — это белковые компоненты хроматина, структуры, вокруг которой намотана ДНК. Гистоны представляют собой октомеры с вариабельными хвостами, которые выходят за пределы комплекса ДНК/гистон (нуклеосома). Существует несколько типов посттрансляционных модификаций, которые могут влиять на хвосты гистонов, включая метилирование, ацетилирование, фосфорилирование и убиквитинирование. Эти модификации могут влиять на взаимодействие между ДНК и гистонами, приводя к изменениям транскрипции генов, репарации ДНК, репликации ДНК и даже организации хромосом [4].
Существует несколько типов посттрансляционных модификаций, которые могут влиять на хвосты гистонов, включая метилирование, ацетилирование, фосфорилирование и убиквитинирование. Эти модификации могут влиять на взаимодействие между ДНК и гистонами, приводя к изменениям транскрипции генов, репарации ДНК, репликации ДНК и даже организации хромосом [4].
Одной из наиболее изученных модификаций гистонов является ацетилирование остатка лизина. В общем, ацетилирование гистонов связано с активацией транскрипции, а деацетилирование связано с репрессией транскрипции (рис. 1). Ацетилирование нейтрализует положительно заряженный остаток лизина в гистоновом хвосте, уменьшая силу связи между гистоновым хвостом и ДНК. Это явление открывает комплекс ДНК/гистон, делая его доступным для факторов транскрипции [1]. Метилирование гистонов может положительно или отрицательно влиять на транскрипцию. Также может иметь место сотрудничество между метилированием ДНК и модификациями гистонов. При всех различных модификациях и комбинациях изменений сложность модификаций хроматина поразительна и до сих пор является областью интенсивных исследований [15].
Открыть в отдельном окне
Ацетилирование гистонов. На верхнем кадре ДНК (черные линии) плотно намотана на гистоны (кружки), образуя нуклеосомный комплекс. Хвосты гистонов (кривые линии) не ацетилированы. В нижней рамке ацетилирование (Ac) гистоновых хвостов «открывает» ДНК для доступа к факторам транскрипции.
Методы обнаружения модификаций хроматина
В лаборатории модификации гистонов легко и точно обнаруживаются с помощью масс-спектрометрии. Однако этот метод трудоемок и требует высокоспециализированного обучения и оборудования [16]. Чтобы определить истинное биологическое значение модификации гистонов, также требуется информация о последовательности ДНК. Оптимальный метод, иммунопреципитация хроматина (ChIP), сочетает технологию секвенирования с ДНК, которая была иммунопреципитирована с антителами против специфических модификаций гистонов. Полногеномные исследования модификаций гистонов теперь возможны благодаря использованию так называемых исследований «ЧИП на чипе», которые сочетают иммунопреципитацию с платформами для секвенирования микрочипов. Текущие ограничения этого метода связаны с качеством поликлональных антител, созданных против модификаций гистонов [16].
Текущие ограничения этого метода связаны с качеством поликлональных антител, созданных против модификаций гистонов [16].
Импринтированные гены имеют моноаллельную экспрессию – имеется только 1 копия (от одного из родителей) вместо 2. Импринтируется примерно 1% аутосомных генов. Метилирование ДНК и ацетилирование гистонов маркируют импринтированный ген и препятствуют транскрипции гена. Поскольку экспрессия зависит только от одного родителя, экспрессия в текущем поколении зависит от среды, в которой проживало предыдущее поколение, и любых эпигенетических меток, которые могли возникнуть [2]. Потеря импринтинга означает, что либо имеется биаллельная экспрессия гена, либо обе копии не экспрессируются. Функциональное гаплоидное состояние этих генов делает их исключительно восприимчивыми к дальнейшим эпигенетическим изменениям или мутациям [3]. Некоторые нарушения развития связаны с импринтированными генами, такие как синдром Ангельмана, синдром Прадера-Вилли и синдром Беквита-Видеманна. Основываясь на родительском типе наследования, предполагается, что биполярное расстройство, аутизм, шизофрения и синдром Туретта также могут быть связаны с потерей импринтинга [2].
Основываясь на родительском типе наследования, предполагается, что биполярное расстройство, аутизм, шизофрения и синдром Туретта также могут быть связаны с потерей импринтинга [2].
Наиболее изученными некодирующими РНК являются микроРНК (миРНК). Это последовательности длиной примерно 22 нуклеотида, которые кодируются длинными некодирующими РНК или интронами генов. miRNA транскрибируются в ядре и претерпевают несколько модификаций до своего созревания. miRNA может ингибировать трансляцию мРНК в белок двумя способами. Если миРНК представляет собой прямую последовательность, комплементарную мРНК, то миРНК связывается с мРНК и разрушает ее посредством действия комплекса RISC. Если микроРНК не полностью соответствует мРНК, то микроРНК частично связывается с 3′-концом мРНК и запрещает действие транспортной РНК [1]. Было обнаружено, что миРНК аномально экспрессируется при множестве желудочно-кишечных заболеваний, включая воспалительные заболевания кишечника [17,18], холангиокарциному [19), аденокарциному пищевода [20] и гепатоцеллюлярную карциному [5,15].
Появляется все больше данных, свидетельствующих о том, что воздействие окружающей среды на раннем этапе развития играет определенную роль в заболевании в более позднем возрасте. Именно эпигенетика обеспечивает правдоподобную связь между воздействием окружающей среды и риском заболевания [2]. Эпигенетика также может объяснить, как риск определенного заболевания передается из поколения в поколение. Классическим примером того, как воздействия окружающей среды могут влиять на эпигенетические изменения, является история мыши агути. Аллель 9 мышиного агути0051 Avy имеет мобильный элемент (названный IAP) перед своим промотором и может регулироваться метилированием ДНК. Аллель дикого типа a кодирует коричневый цвет шерсти, а аллель Avy кодирует желтый цвет шерсти, ожирение и диабет. В исходных условиях мышь агути демонстрирует химерный фенотип со смешанным коричневым и желтым окрасом шерсти. Естественное метилирование элемента IAP приводит к увеличению коричневого цвета шерсти и мышечной массы тела. В одном эксперименте a 9Матери 0052 / a были скрещены с самцами a / Avy и дополнены донорами метила бетаином, холином, витамином B 12 и фолиевой кислотой. Потомство матерей, получавших добавки, было более худым и имело больше коричневого цвета в шерсти. Кроме того, было обнаружено, что элементы IAP у животных, получавших добавки, имели повышенное метилирование, чем у потомства матерей, не получавших добавки [2]. Этот эксперимент показал, что воздействие внутриутробно может повлиять на эпигенетический статус потомства, а также повлиять на фенотип. Эта же исследовательская группа смогла вызвать метилирование элемента IAP с помощью растительного эстрогена генистен в тех же дозах, что и у людей, которые потребляют пищу на основе сои. Примечательно, что прием матерями генистена защищал потомство от диабета [2].
В одном эксперименте a 9Матери 0052 / a были скрещены с самцами a / Avy и дополнены донорами метила бетаином, холином, витамином B 12 и фолиевой кислотой. Потомство матерей, получавших добавки, было более худым и имело больше коричневого цвета в шерсти. Кроме того, было обнаружено, что элементы IAP у животных, получавших добавки, имели повышенное метилирование, чем у потомства матерей, не получавших добавки [2]. Этот эксперимент показал, что воздействие внутриутробно может повлиять на эпигенетический статус потомства, а также повлиять на фенотип. Эта же исследовательская группа смогла вызвать метилирование элемента IAP с помощью растительного эстрогена генистен в тех же дозах, что и у людей, которые потребляют пищу на основе сои. Примечательно, что прием матерями генистена защищал потомство от диабета [2].
Другим примером воздействия окружающей среды и его влияния на эпигенетику является голландское исследование зимнего голода 1944–1945 годов. В это время в оккупированной Голландии голландцы находились на строгом пайке. Ежедневная еда состояла из двух картофелин, двух ломтиков хлеба и кусочка свеклы. Шестьдесят лет спустя, по сравнению с их однополыми братьями и сестрами, потомки, происходящие от беременных матерей голода, продолжали иметь биаллельную экспрессию импринтированного гена IGFR2 [2]. Предполагается, что к этому явлению привел дефицит фолиевой кислоты, вызванный голодом. Считается, что диеты, богатые обработанными продуктами, такие как те, которыми пользуются в западном мире, содержат недостаточно фолиевой кислоты, бетаина и холина, что потенциально предрасполагает людей к эпигенетическим аномалиям.
В это время в оккупированной Голландии голландцы находились на строгом пайке. Ежедневная еда состояла из двух картофелин, двух ломтиков хлеба и кусочка свеклы. Шестьдесят лет спустя, по сравнению с их однополыми братьями и сестрами, потомки, происходящие от беременных матерей голода, продолжали иметь биаллельную экспрессию импринтированного гена IGFR2 [2]. Предполагается, что к этому явлению привел дефицит фолиевой кислоты, вызванный голодом. Считается, что диеты, богатые обработанными продуктами, такие как те, которыми пользуются в западном мире, содержат недостаточно фолиевой кислоты, бетаина и холина, что потенциально предрасполагает людей к эпигенетическим аномалиям.
Многие другие воздействия окружающей среды связаны с изменением эпигенетических паттернов в течение жизни и последующим риском заболевания. Конкретные примеры включают табачный дым, алкоголь, вирусный гепатит, промышленные загрязнители и выбросы углерода. В одном исследовании изучалось метилирование и модификации гистонов у монозиготных близнецов на разных этапах жизни. В начале жизни эпигенетические признаки почти идентичны. Однако в возрасте 50 лет были обнаружены существенные различия, что позволяет предположить, что воздействие окружающей среды изменило эпигеном [3].
В начале жизни эпигенетические признаки почти идентичны. Однако в возрасте 50 лет были обнаружены существенные различия, что позволяет предположить, что воздействие окружающей среды изменило эпигеном [3].
Эпигенетические события, которые регулируют экспрессию генов, ясно проявились как фундаментальный механизм в биологии развития и в патогенезе болезней человека. Например, многие гены, влияющие на многочисленные клеточные пути, подавляются гиперметилированием при раке, и изучение этих генов расширило наше понимание того, как развивается и прогрессирует рак. Однако большинство исследований сосредоточено на метилировании одного гена или группы генов без подробного изучения функциональной значимости сайленсинга генов. С появлением полногеномных микрочиповых платформ «метилом» получит дальнейшее определение. Молекулярная информация, полученная в результате эпигенетических исследований, в сочетании с другой генетической информацией может быть использована для разработки новой системы классификации опухолей и заболеваний. Эта теоретическая система классификации может быть разработана для отражения так называемой «биологии опухоли», которая может предсказывать клинические исходы, такие как общий прогноз, риск рецидива после операции или ответ на химиотерапию.
Эта теоретическая система классификации может быть разработана для отражения так называемой «биологии опухоли», которая может предсказывать клинические исходы, такие как общий прогноз, риск рецидива после операции или ответ на химиотерапию.
Естественно, с расширением знаний о патогенезе заболеваний есть надежда на разработку новой эры новых терапевтических средств, которые могут эффективно лечить пациентов. Прелесть эпигенных модификаций ДНК в том, что их можно предотвратить или даже обратить. Например, блокирование метилирования ДНК путем ингибирования DNMT приводит к деметилированию CpG-островков в дочерних клетках с последующим восстановлением экспрессии генов-супрессоров опухолей и прекращением роста опухоли. Есть несколько соединений, деметилирующих ДНК, которые активно исследуются, включая 5-азацитидин (AzaC), 5-аза-2′-дезоксицитидин, зебуларин, прокаинамид и прокаин [8]. К сожалению, основным ограничивающим фактором этих препаратов является токсичность, и в настоящее время они показаны только пациентам с выраженными миелодиспластическими синдромами [21]. Вместо системного введения терапевтических средств активно разрабатываются и изучаются специализированные системы доставки, которые специально нацелены на аберрантное метилирование в опухолях. Также было показано, что аномальное метилирование промотора коррелирует с устойчивостью к химиотерапии и радиации [22]. В будущем возможно использование деметилирующих агентов для повышения эффективности традиционной химиотерапии [21].
Вместо системного введения терапевтических средств активно разрабатываются и изучаются специализированные системы доставки, которые специально нацелены на аберрантное метилирование в опухолях. Также было показано, что аномальное метилирование промотора коррелирует с устойчивостью к химиотерапии и радиации [22]. В будущем возможно использование деметилирующих агентов для повышения эффективности традиционной химиотерапии [21].
Существует также множество ингибиторов гистондеацетилазы (HDAC), которые изучаются для лечения рака. Примеры этих ингибиторов HDAC включают субероиланилид гидроксамовой кислоты, трихостатин А, вальпроевую кислоту и бутират натрия [8]. Эти агенты приводят к увеличению ацетилирования гистонов за счет блокирования действия нескольких HDAC и обычно используются в лабораторных экспериментах для устранения эпигенетически индуцированного сайленсинга генов.
Многие из описанных эпигенетических событий были обнаружены в человеческих образцах предраковых тканей. В частности, некоторые гиперметилированные гены могут быть обнаружены даже в сыворотке пациентов до клинического выявления злокачественного новообразования. Другим примером является прогноз развития прогрессирования дисплазии при ассоциированной с болезнью Барретта аденокарциноме пищевода. В рандомизированном двойном слепом многоцентровом исследовании панель из восьми гиперметилированных генов предсказывала прогрессирование дисплазии лучше, чем традиционные клинические факторы риска [23]. По этой причине метилирование ДНК, гипометилирование или модификация гистонов являются потенциальными кандидатами в качестве биомаркеров для раннего выявления заболевания.
В частности, некоторые гиперметилированные гены могут быть обнаружены даже в сыворотке пациентов до клинического выявления злокачественного новообразования. Другим примером является прогноз развития прогрессирования дисплазии при ассоциированной с болезнью Барретта аденокарциноме пищевода. В рандомизированном двойном слепом многоцентровом исследовании панель из восьми гиперметилированных генов предсказывала прогрессирование дисплазии лучше, чем традиционные клинические факторы риска [23]. По этой причине метилирование ДНК, гипометилирование или модификация гистонов являются потенциальными кандидатами в качестве биомаркеров для раннего выявления заболевания.
Таким образом, эпигенетика стала важным связующим звеном между природой и воспитанием. С невероятной степенью сложности эпигенетические явления оказывают глубокое влияние на регуляцию того, как генетическая информация транскрибируется и транслируется в белки и фенотипы. Благодаря передовым технологиям и знаниям возможны будущие открытия в отношении патогенеза многофакторных и ранее идиопатических заболеваний.
Автор заявляет об отсутствии финансового или иного конфликта интересов в отношении содержания статьи.
Эта работа поддерживается K08DK083774, спонсируемым Национальным институтом диабета и Отделением расстройств пищеварения и почек Национального института здравоохранения (США).
1. Гибни Э.Р., Нолан К.М. Эпигенетика и экспрессия генов. Наследственность. 2010; 105:4–13. [PubMed] [Google Scholar]
2. Jirtle RL, Skinner MK. Экологическая эпигеномика и восприимчивость к болезням. Нат Рев Жене. 2007; 8: 253–262. [Бесплатная статья PMC] [PubMed] [Google Scholar]
3. Feinberg AP. Эпигенетика в эпицентре современной медицины. ДЖАМА. 2008;299:1345–1350. [PubMed] [Google Scholar]
4. Эстеллер М. Эпигенетика рака. N Engl J Med. 2008; 358:1148–1159. [PubMed] [Google Scholar]
5. Селару Ф.М., Дэвид С., Мельцер С.Дж., Гамильтон Дж.П. Эпигенетические события при раке желудочно-кишечного тракта. Am J Гастроэнтерол. 2009; 104:1910–1912. [PMC free article] [PubMed] [Google Scholar]
6. Feinberg AP, Vogelstein B. Гипометилирование отличает гены некоторых видов рака человека от их нормальных аналогов. Природа. 1983; 301: 89–92. [PubMed] [Академия Google]
Feinberg AP, Vogelstein B. Гипометилирование отличает гены некоторых видов рака человека от их нормальных аналогов. Природа. 1983; 301: 89–92. [PubMed] [Академия Google]
7. Calvisi DF, Ladu S, Gorden A, Farina M, Lee JS, Conner EA, et al. Механистическое и прогностическое значение аберрантного метилирования в молекулярном патогенезе гепатоцеллюлярной карциномы человека. Джей Клин Инвест. 2007; 117: 2713–2722. [Бесплатная статья PMC] [PubMed] [Google Scholar]
8. Roberts LR, Gores GJ. Гепатоцеллюлярная карцинома: молекулярные пути и новые терапевтические мишени. Семин Печень Дис. 2005; 25: 212–225. [PubMed] [Google Scholar]
9. Lee HS, Kim BH, Cho NY, Yoo EJ, Choi M, Shin SH, et al. Прогностические последствия и взаимосвязь между гиперметилированием CpG-островков и повторяющимся гипометилированием ДНК при гепатоцеллюлярной карциноме. Клин Рак Рез. 2009 г.;15:812–820. [PubMed] [Google Scholar]
10. Герман Дж.Г., Бейлин С.Б. Сайленсинг генов при раке в связи с гиперметилированием промотора. N Engl J Med. 2003; 349:2042–2054. [PubMed] [Google Scholar]
N Engl J Med. 2003; 349:2042–2054. [PubMed] [Google Scholar]
11. Герман Дж.Г., Графф Дж.Р., Миоханен С., Нелкин Б.Д., Байлин С.Б. Метилирование-специфическая ПЦР: новый ПЦР-анализ статуса метилирования CpG-островков. Proc Natl Acad Sci USA. 1996; 93:9821–9826. [Бесплатная статья PMC] [PubMed] [Google Scholar]
12. Dupont JM, Tost J, Jammes H, Gut IG. Количественное бисульфитное секвенирование de novo с использованием технологии пиросеквенирования. Анальная биохимия. 2004;333:119–127. [PubMed] [Google Scholar]
13. Weber M, Davies JJ, Wittig D, Oakeley EJ, Haase M, Lam WL, et al. Полнохромосомный и специфичный для промотора анализ выявляют сайты дифференциального метилирования ДНК в нормальных и трансформированных клетках человека. Нат Жене. 2005; 37: 853–862. [PubMed] [Google Scholar]
14. Toyota M, Ho C, Ahuja N, Jair KW, Li Q, Ohe-Toyota M, et al. Идентификация дифференциально метилированных последовательностей при колоректальном раке путем амплификации метилированных CpG-островков. Рак рез. 1999;59:2307–2312. [PubMed] [Google Scholar]
Рак рез. 1999;59:2307–2312. [PubMed] [Google Scholar]
15. Hamilton JP. Эпигенетические механизмы, участвующие в патогенезе гепатобилиарных злокачественных новообразований. Эпигеномика. 2010;2:233–243. [Бесплатная статья PMC] [PubMed] [Google Scholar]
16. Эстеллер М. Эпигеномика рака: метиломы ДНК и карты модификации гистонов. Нат Рев Жене. 2007; 8: 286–298. [PubMed] [Google Scholar]
17. Wu F, Zhang S, Dassopoulos T, Harris ML, Bayless TM, Meltzer SJ, et al. Идентификация микроРНК, связанных с болезнью Крона подвздошной и толстой кишки. Воспаление кишечника Dis. 2010;16:1729–1738. [Бесплатная статья PMC] [PubMed] [Google Scholar]
18. Олару А.В., Селару Ф.М., Мори Ю., Васкес С., Дэвид С., Паун Б. и соавт. Динамические изменения экспрессии микроРНК-31 при неопластической трансформации, связанной с воспалительным заболеванием кишечника. Воспаление кишечника Dis. 2011;17:221–231. [Бесплатная статья PMC] [PubMed] [Google Scholar]
19. Selaru FM, Olaru AV, Kan T, David S, Cheng Y, Mori Y, et al. МикроРНК-21 сверхэкспрессируется при холангиокарциноме человека и регулирует запрограммированную гибель клеток 4 и тканевой ингибитор металлопротеиназы 3. Гепатология. 2009 г.;49:1595–1601. [Бесплатная статья PMC] [PubMed] [Google Scholar]
МикроРНК-21 сверхэкспрессируется при холангиокарциноме человека и регулирует запрограммированную гибель клеток 4 и тканевой ингибитор металлопротеиназы 3. Гепатология. 2009 г.;49:1595–1601. [Бесплатная статья PMC] [PubMed] [Google Scholar]
20. Kan T, Sato F, Ito T, Matsuura N, David S, Cheng Y, et al. Полицистрон miR-106b-25, активируемый геномной амплификации, функционирует как онкоген, подавляя p21 и Bim. Гастроэнтерология. 2009; 136:1689–1700. [Статья бесплатно PMC] [PubMed] [Google Scholar]
21. Boland CR, Shin SK, Goel A. Метилирование промотора в генезе рака желудочно-кишечного тракта. Yonsei Med J. 2009; 50: 309–321. [Бесплатная статья PMC] [PubMed] [Google Scholar]
22. Hamilton JP, Sato F, Greenwald BD, Suntharalingam M, Krasna MJ, Edelman MJ, et al. Метилирование промотора и ответ на химиотерапию и облучение при раке пищевода. Клин Гастроэнтерол Гепатол. 2006; 4: 701–708. [PubMed] [Google Scholar]
23. Jin Z, Cheng Y, Gu W, Zheng Y, Sato F, Mori Y, et al. Многоцентровое двойное слепое проверочное исследование биомаркеров метилирования для прогнозирования прогрессирования пищевода Барретта. Рак рез. 2009;69:4112–4115. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Многоцентровое двойное слепое проверочное исследование биомаркеров метилирования для прогнозирования прогрессирования пищевода Барретта. Рак рез. 2009;69:4112–4115. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Статьи по заболеваниям пищеварения (Базель, Швейцария) предоставлены здесь Karger Publishers
Эпигенетический принцип
Эпигенетический принцип относится к биологическому плану роста, при котором каждая функция возникает в систематической последовательности до тех пор, пока не разовьется полностью функционирующий организм. В теории Эриксона это идея о том, что у каждой психосоциальной силы есть свой особый период особой важности, идея о том, что развитие направляется основным планом, в котором определенные вопросы имеют свое особое время важности. (См. Психосоциальную теорию Эриксона)
- Пред.
- Следующий
|
Статьи по теме | |
Инициатива против чувства вины (3-5 лет) на сайте психологии-глоссария. | |
| — Инициатива против Вина (3-5 лет): — Инициатива vs. — Вина (3-5 лет) относится к 3-й стадии Эрика. . . Подробнее | |
| Доверие и недоверие на сайте психологии-глоссария.com | ■■■■■ |
| Доверие против недоверия: ; Доверие против недоверия (0–1 год) относится к первому этапу теории Эрика Эриксона. . . Подробнее | |
| Автономия против стыда и сомнений (1–3 года) на сайте психологии-глоссария. лет): — Автономия против стыда и сомнения (1-3 года) относится ко 2-му. . . Подробнее | |
| Трудолюбие против неполноценности (5-12 лет) на сайте психологии-глоссария.com | ■■■■■ |
| — Трудолюбие против неполноценности (5-12 лет): ; Трудолюбие против неполноценности (5–12 лет) относится к 4-му этапу. . . Подробнее | |
Идентичность и спутанность идентичности (подростковый возраст) на сайте психологии-глоссария.
| |