
Филогенез человека: Филогенез человека — Эволюция человека — ЭВОЛЮЦИЯ, ИЛИ ИСТОРИЯ ЖИЗНИ НА ЗЕМЛЕ — БИОЛОГИЯ Том 3 — Biological Science — руководство по общей биологии — Тейлор Д.
Онтогенез повторяет филогенез • Джеймс Трефил, энциклопедия «Двести законов мироздания»
200 законов мироздания > Взгляд в прошлое
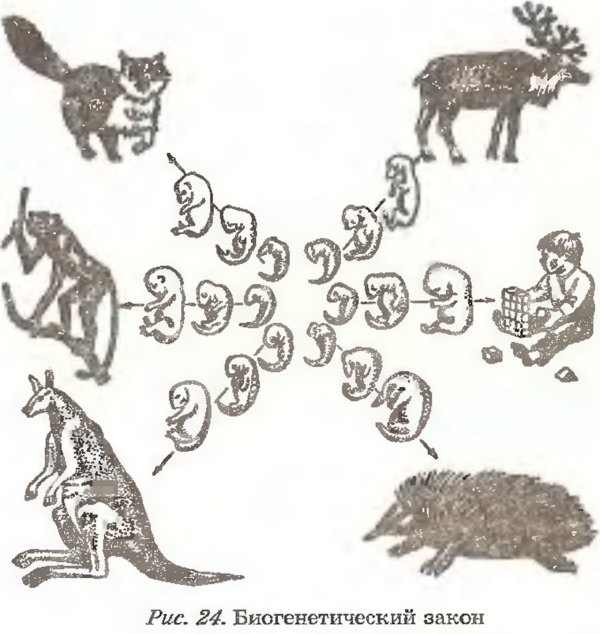
Зародыш в своем развитии проходит весь путь эволюции своего вида.
В XIX веке ученые, изучавшие внутриутробное развитие человеческого эмбриона, заметили, что в первые месяцы жизни он обладает поразительным сходством с другими позвоночными. Например, в месячном возрасте у человеческого эмбриона в области шеи заметны щели, во всех отношениях похожие на зачаточные жабры. Позднее зародыш имеет сходство с земноводными, затем с птицами и наконец — с другими млекопитающими. Это сходство привело к появлению приведенного выше изречения, сделанного немецким натуралистом Эрнстом Геккелем (Ernst Haeckel, 1834–1919) в его книге «Решето вселенной», опубликованной в 1899 году. Имеется в виду, что онтогенез живого существа (развитие индивида) повторяет путь филогенеза (развития типа, класса или вида — см. Система классификации Линнея). Так, человеческий эмбрион сначала похож на эмбрион рыбы, затем рептилии и так далее — до тех пор, пока не проявится его принадлежность к роду людей. Такова одна из этих идей — ясных, красивых, разумных — и в корне неверных.
Система классификации Линнея). Так, человеческий эмбрион сначала похож на эмбрион рыбы, затем рептилии и так далее — до тех пор, пока не проявится его принадлежность к роду людей. Такова одна из этих идей — ясных, красивых, разумных — и в корне неверных.
На самом деле у человеческого зародыша никогда не бывает жабр или каких-либо других придатков, которые ему следовало бы в соответствии с этой концепцией иметь на той или иной стадии развития. Появляющиеся жаброподобные щели называются вторая жаберная дуга. У рыб эти образования действительно развиваются в жабры, но у человека они служат предшественниками частей головы и шеи. Точно так же, как теория эволюции предполагает не то, что человек произошел от приматов, а то, что он имеет с ними общего предка, — так и эмбриология утверждает не то, что человеческий зародыш в своем развитии проходит все ступени эволюции, а просто то, что в нем развиваются другие органы из тех же зародышевых клеток. (Идея о том, что «онтогенез повторяет филогенез», чем-то напоминает столь же неверную теорию Триединого мозга.
Удивительно, но несмотря на то, что эта идея, которая удостоилась даже статуса закона биогенетики, была опровергнута почти сразу после того, как была выдвинута, она тем не менее смогла просуществовать до наших дней (ее даже можно найти еще в некоторых учебниках!). Между онтогенезом и филогенезом действительно есть связь, но нет эмбриологического повторения. К очевидным вещам иногда полезно относиться скептически!
15
Показать комментарии (15)
Свернуть комментарии (15)
-
tulpan 31.01.2007 19:27 Ответить
<Онтогенез повторяет филогенез>
«К очевидным вещам иногда полезно относиться скептически…»
Добрый вечер,всем
Когда вы будете знакомиться с Царь-Пушкой, которая в Кремле или изображена на открытке, НЕ ВЕРЬТЕ своим глазам (и знаниям, полученным из школьной геометрии), что ФИГУРА из шаров в её основании является ПИРАМИДОЙ.

На самом деле она является РЕФЕРЕНТОМ реальности со свойствами, отвечающими фундаментальной гипотезе нашего выдающегося учёного-энциклопедиста Владимира Ивановича Вернадского, которую он высказал, завершая свой жизненный и творческий пути, в одной из неопубликованных статей 1941-42 гг.Кстати, В.И.Вернадский пророчески заявил: «Царство моих идей впереди!»
Приглашаю всех принять участие в изучении и освоении этого референта и той РЕАЛЬНОСТИ, которую он представляет.
Было бы замечательно, если проведённую непростую работу опубликовать совместно ко дню его рождения в 2008 или 2013 году
31 январь 2007 Всего доброго
ИСТИНЫ
С искренним уважением ко всем искателям
Анатолий ИвановичОтветить
-
тюльпан tulpan 19.02.2007 17:06 Ответить
Удивительно: не вижу реакции на своё сообщение от 31.01.07
Тема не интересна? Или что-то иное?
Очень похоже, что на «Элементах» мало любознательных
человеков. Или вас здесь вообще нет?
Или вас здесь вообще нет?
Надеюсь, что ошибаюсь
Тюльпан 19 февраль 2007Ответить
-
Факир 05.05.2007 04:15 Ответить
Почтеннейшие, господа. Я всю жизнь прожил в уверенности, что «онтогенез повторяет филогенез». И теперь это возвышавшее меня в собственных глазах знание оказалось заблуждением? Но доказательного опровержения я не увидел. А прочитал нечто похожее на приказ по науке: «Считать такое-то утверждение ошибочным». А между тем опровергаемый закон так красив, что может оказаться истиной. Я раздосадован и остаюсь верен своему «заблуждению».
Ответить
-
eugeneol Факир 06.07.2007 23:47 Ответить
Уважаемый Факир,
полностью Вас поддерживаю и требую от автора статьи доказательств!!!
Ответить
-
sanchik Факир 24.
 01.2008 02:15
Ответить
01.2008 02:15
Ответить
поддерживаю факира!
и пусть в нас кинут камниОтветить
-
evolucionism2021
Факир 21.03.2021 14:42
Ответить
Этот закон близок к истине, но именно что близок. Неточный и не всеобщий.
Ответить
tr1 04.01.2010 21:05 Ответить
«Между онтогенезом и филогенезом действительно есть связь, но нет эмбриологического повторения.»
Точно это же самое написано в учебнике биологии для средней школы конца прошлого века. В начале перевирается и вульгаризируется классическое определение. Изалагается ошибочная точка зрения «типичного троечника».( который не удосужился ознакомиться с тем, что из каких жаберных дуг формируется)А потом классическая формулировка выдается за «научную новсть» брр.
Уся новость имеется в БСЭ.
Биогенетический закон
Ответить
-
Andrey2 tr1 18.08.2010 16:36 Ответить
«Между онтогенезом и филогенезом действительно есть связь, но нет эмбриологического повторения.»
Если Я не повторяю филогенез в перинатальном онтогенезе, то Вы полагаете, что я его повторяю в постнатальном?Ответить
-
Andrey2 tr1 18.08.2010 16:36 Ответить
«Между онтогенезом и филогенезом действительно есть связь, но нет эмбриологического повторения.»
Если Я не повторяю филогенез в перинатальном онтогенезе, то Вы полагаете, что я его повторяю в постнатальном?Ответить
agalanin 11.02.2011 05:26 Ответить
Кстати от всех заметок, несмотря на их разнообразие, на этом сайте веет одним и тем-же — стенфордским университетом, вернее каким то примитивным учебником американским для студентов естественников.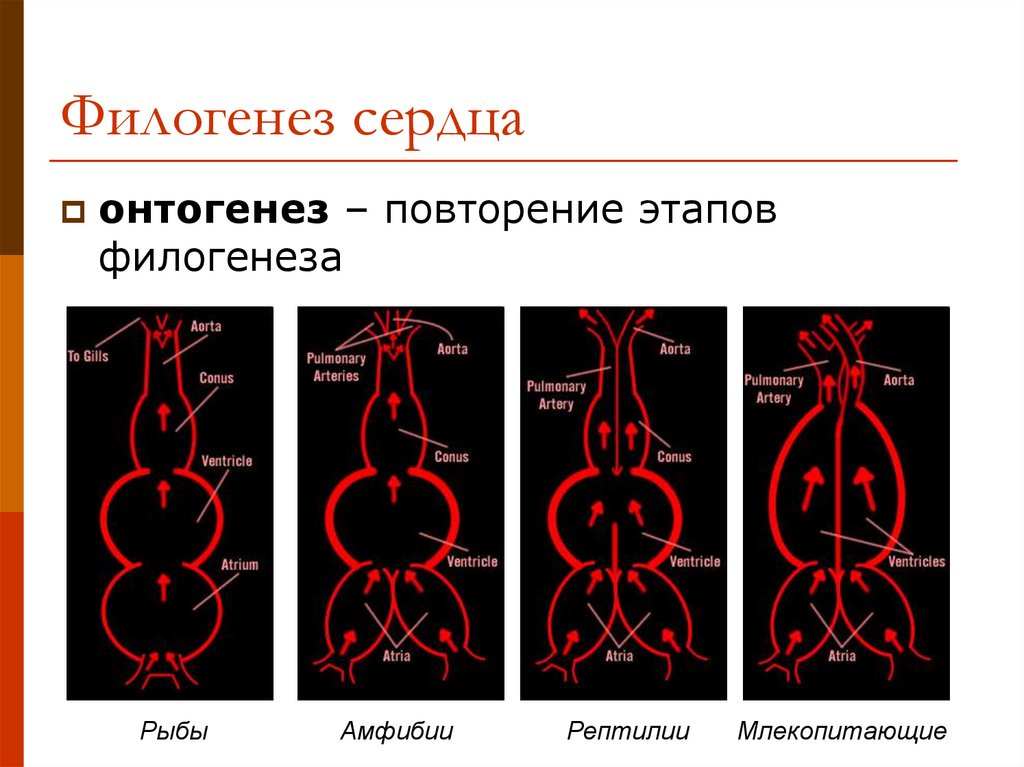 Одни верхи по сливкам. Автор этих заметок все это перепечатал и имеет весьма поверхностные несвязанные и неустоявшиеся знания. Несмотря на четкость статей, все они под одну гребенку с американским душком. Я много с такими общался. Мыслителей среди них почти что нет, да и те с восточной европы недавно приехали. Могу дать коментарий по всем статьям по их логике. Пустой звук и никакой для меня новой информации.
Одни верхи по сливкам. Автор этих заметок все это перепечатал и имеет весьма поверхностные несвязанные и неустоявшиеся знания. Несмотря на четкость статей, все они под одну гребенку с американским душком. Я много с такими общался. Мыслителей среди них почти что нет, да и те с восточной европы недавно приехали. Могу дать коментарий по всем статьям по их логике. Пустой звук и никакой для меня новой информации.
леша
Ответить
ортодокс 09.02.2014 17:46 Ответить
«…На самом деле у человеческого зародыша никогда не бывает жабр или каких-либо других придатков,которые ему следовало бы в соответствии с этой концепцией иметь на той или иной стадии развития.»
«…Этого не может быть потому что этого не может быть никогда!»
(Козьма Прутков)
Хорошо,что Автор статьи не требует в качестве неопровержимых доказательств оспариваемой им теории,снимки (сделанные,скажем,при помощи УЗИ) матки беременной женщины,на которых он мог бы увидеть в зависимости от срока беременности то маленькую рыбку,то лягушку,то птичку и т. д.,пока,наконец,не увидел бы воочию человеческого младенца…Впрочем,любые аргументы бессильны там,где силу «неоспоримых доказательств» должна заменить сила абстракции,присущая только человеческому разуму…
д.,пока,наконец,не увидел бы воочию человеческого младенца…Впрочем,любые аргументы бессильны там,где силу «неоспоримых доказательств» должна заменить сила абстракции,присущая только человеческому разуму…
Ответить
«Точно так же, как теория эволюции предполагает не то, что человек произошел от приматов, а то, что он имеет с ними общего предка,»
Не ожидал такого в этой замечательной книге. Утверждение, что «человек и примат имели общего предка» — это из разряда «крыса и грызун имели общего предка».
Ответить
evolucionism2021 21.03.2021 14:47 Ответить
Да и утверждения о неверности закона слишком твёрдые. От абсолютно принятых в науке законов Бэра он отличается не радикально.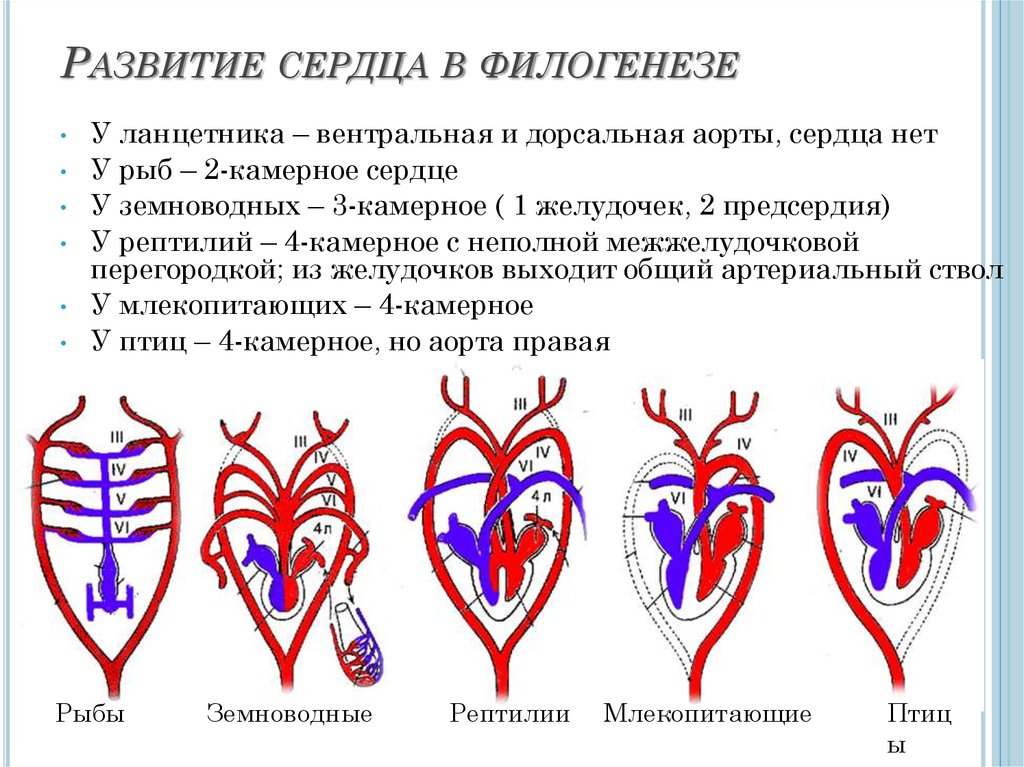
Ответить
makeyev 20.08.2022 21:47 Ответить
Несколько десятилетий в науке о развитии живых организмов торжествовало убеждение, что онтогенез повторяет филогенез.
Генетические участки ДНК, РНК частично или целиком могут перемещаться доль нити ДНК, РНК. Некоторые перестановки генов могут изменять течение онтогенеза живого существа. На эти миграции генов и участков генов могут накладываться выпадения некоторых генов или их участков, а также включения новых генов или участков новых генов. И ещё накладываются мутации нуклеотидов на каких-то участках в любых генах и как бы не несущих генетическую информацию больших участках ДНК, РНК, вследствие рождения новых нейтронов в веществе и включения этих нейтронов в состав ядер атомов разных изотопов разных элементов, в том числе входящих с ткани разных органов и систем живых организмов.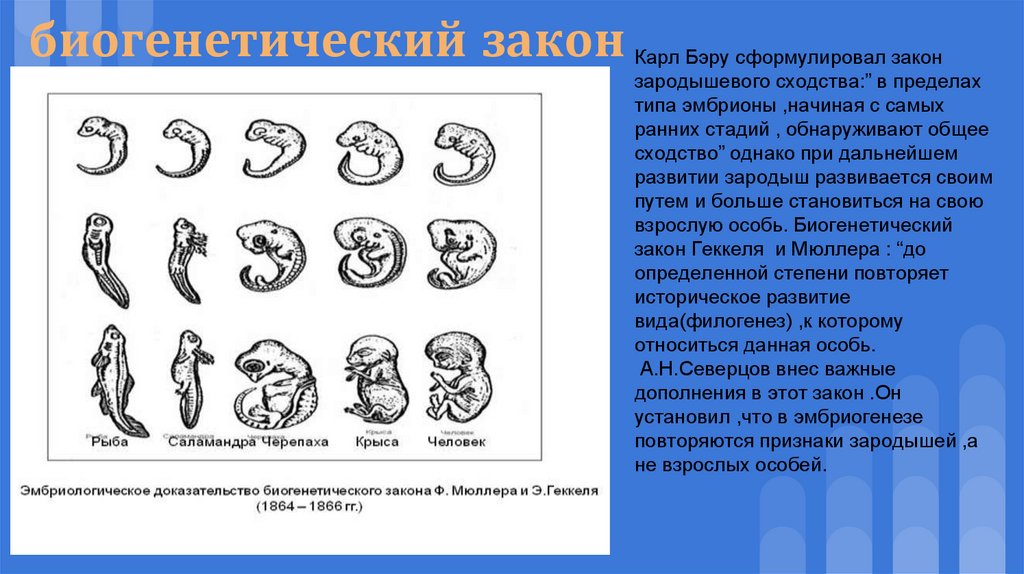 А именно в грамме любого вещества функция-работа процесса времени бытия рождает около 4,155 миллионов новых нейтронов в грамме любого вещества.
А именно в грамме любого вещества функция-работа процесса времени бытия рождает около 4,155 миллионов новых нейтронов в грамме любого вещества.
И какие-то такие пертурбации в генетической и негенетической информации могут изменять течение формирования тканей и органов зародышей живых организмов. Как у усложнение, так и в упрощение. Так какие-то органы могут формироваться из зачатков того, что у других типов, классов и видов живых организмов развиваются в другие ткани и органы.
Поэтому действительно, лишь частично соответствует реальности утверждение, что онтогенез (развитие зародыша) повторяет филогенез — историческое развитие живых организмов.
Ответить
Написать комментарий
1809 |
Ламаркизм |
ок. 1850 |
Социальный дарвинизм |
1859 |
Теория эволюции |
1899 |
Онтогенез повторяет филогенез |
сер. |
Триединый мозг |
 XX в.
XX в.1927
Опыт Дэвиссона—Джермера
1985
Озоновая дыра
Новостная рассылка
«Элементы» в соцсетях:
В рамках Научно-образовательного лектория обсудят современные проблемы эволюционной биологии
image/svg+xmlПриемная комиссия
+7 (831) 262-26-20
В рамках Научно-образовательного лектория обсудят современные проблемы эволюционной биологии
20 ноября 2018 г. в рамках Научно-образовательного лектория состоятся лекции доктора биологических наук, профессора, главного научного сотрудника Лаборатории микроэволюции млекопитающих ИПЭЭ РАН (г. Москва) Виктора Николаевича Орлова. Вниманию слушателей будет представлен микроцикл лекций о современных проблемах эволюционной биологии.
- центрсоцразвития
20 ноября 2018 г.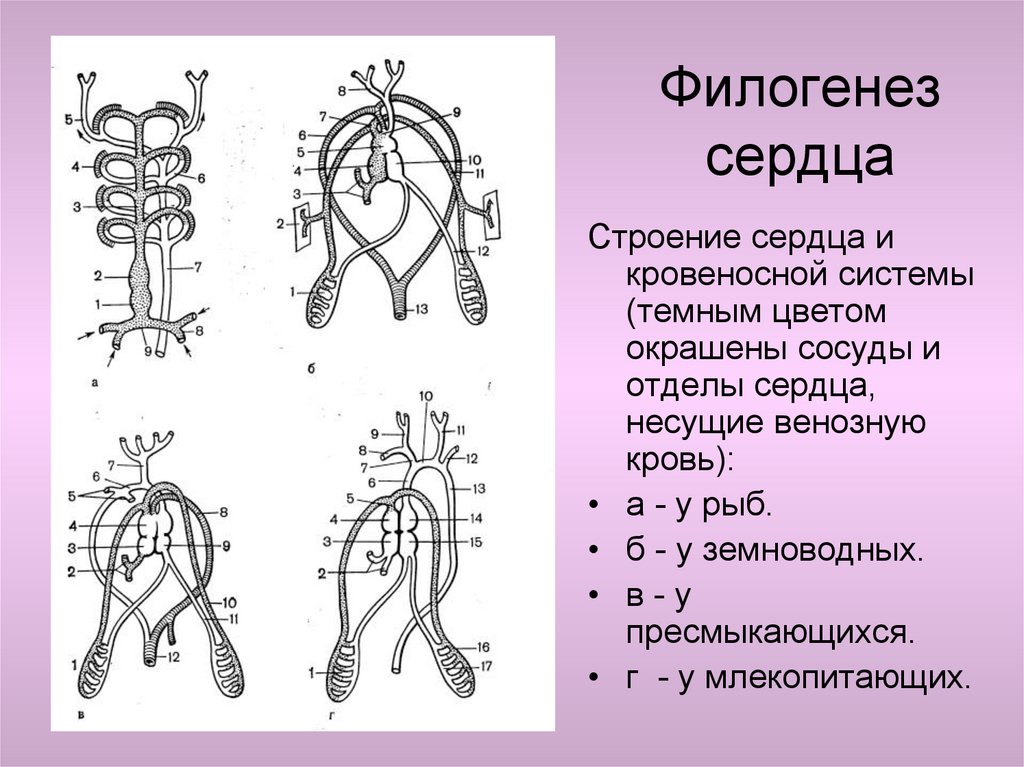 в рамках Научно-образовательного лектория состоятся лекции доктора биологических наук, профессора, главного научного сотрудника Лаборатории микроэволюции млекопитающих ИПЭЭ РАН (г. Москва) Виктора Николаевича Орлова. Вниманию слушателей будет представлен микроцикл лекций о современных проблемах эволюционной биологии.
в рамках Научно-образовательного лектория состоятся лекции доктора биологических наук, профессора, главного научного сотрудника Лаборатории микроэволюции млекопитающих ИПЭЭ РАН (г. Москва) Виктора Николаевича Орлова. Вниманию слушателей будет представлен микроцикл лекций о современных проблемах эволюционной биологии.
Аннотации лекций.
- Иерархическая классификация и филогенетика
Жизнь как контрапункт двух тем – бесконечного биологического разнообразия и удивительного сходства клеточного уровня строения живых организмов. Иерархическая классификация – способ описания биологического разнообразия. Таксоны и систематические категории. Филогенез – реконструкция исторического развития живых организмов. Метод тройного параллелизма (триада). Нестыковки классификации и филогенеза и возникающие из-за этого проблемы.
- Молекулярные методы в классификации и филогенетике
Молекулярные методы дополняют метод тройного параллелизма (от триады к квадриаде). Молекулярные методы — сравнение последовательностей нуклеотидов гомологичных генов и последовательности аминокислот в гомологичных белках. Гены и белки. Ядерные и митохондриальные гены. Молекулярные методы в классификации. Возникновение баркодинга – диагностики видов по митохондриальному гену COI. Пауль Геберт. Молекулярные методы в филогенетике. Возникновение филогеографии — географии генеалогических линий генов. Джон Эвайс. Алгоритм исследований в рамках баркодинга и филогеографии. Примеры.
Молекулярные методы — сравнение последовательностей нуклеотидов гомологичных генов и последовательности аминокислот в гомологичных белках. Гены и белки. Ядерные и митохондриальные гены. Молекулярные методы в классификации. Возникновение баркодинга – диагностики видов по митохондриальному гену COI. Пауль Геберт. Молекулярные методы в филогенетике. Возникновение филогеографии — географии генеалогических линий генов. Джон Эвайс. Алгоритм исследований в рамках баркодинга и филогеографии. Примеры.
- Микроэволюция и макроэволюция
Возникновении представлений о микро- и макроэволюции. Ю.А. Филипченко и Р. Гольдшмидт (20-40 годы). Микро- и макроэволюционные подходы, методы и возможности в изучении единого эволюционного процесса. Микроэволюция – эволюция популяций и видообразование. Примеры. Макроэволюция – дивергенция и направление эволюции видов и надвидовых таксонов. Примеры. Макроэволюционные изменения полностью определяются микроэволюционными. Критика «системной концепции» макроэволюции.
- Эволюция человека
Человек как модель в эволюционных исследованиях. Основные этапы филогенеза (исторического развития) рода людей, Homo. Два типа морфологических признаков людей. Первый – признаки, возникавшие на определенных этапах филогенеза рода (прямохождение, безволосость, морфологические предпосылки речи и др.). Второй – признаки, направленно эволюционировавшие на протяжении всего филогенеза рода людей, такие, как постепенное увеличение размеров и усложнения строения головного мозга. Молекулярные механизмы увеличения мозга и усложнения его структуры в филогенезе рода людей.
Место проведения: Ульянова, 1, ауд. 209.
Время проведения: 10:10-15:10.
Приглашаем всех желающих посетить лекции и поучаствовать в научной дискуссии.
Поделиться
- Просмотров: 1140
Другие новости по теме
В Мининском университете прошел Круглый стол «Перспективы взаимодействия Мининского университета с базовой школой №34 — Гагаринские высоты»
Мероприятие прошло 16 сентября.
Мининский университет улучшил позиции в международном рейтинге «зеленых» вузов — UI GreenMetric
По итогам ежегодного международного рейтинга «зеленых» вузов — UI GreenMetric Мининский университет улучшил свои позиции.
Компетенции управленческих команд образовательных организаций оценили в рамках флагманской программы «Лидерство в образовании»
На этой неделе в рамках очного модуля программы «Лидерство в образовании: командная траектория» обучение управленческих команд образовательных организаций было организовано на площадке Корпоративн
Учителей начали готовить к работе в Школе 800
С 01 декабря 2020 года стартовал цикл тренингов по подготовке будущих педагогических кадров
Ваше имя(*)
Вы не ввели Ваше Имя.
Телефон(*)
Вы не указали Ваш номер телефона.
Согласен на обработку персональных данных Подтвердите свое согласие на обработку персональных данных
Неверный ввод
Нажмите, чтобы прослушать выделенный текст Powered By GSpeechбиофак СПбГУ
биофак СПбГУ
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 gif»>
gif»> gif»>
gif»> gif»>
gif»> gif»>
gif»> gif»>
gif»> gif»>
gif»> gif»>
gif»> gif»>
gif»> gif»>
gif»>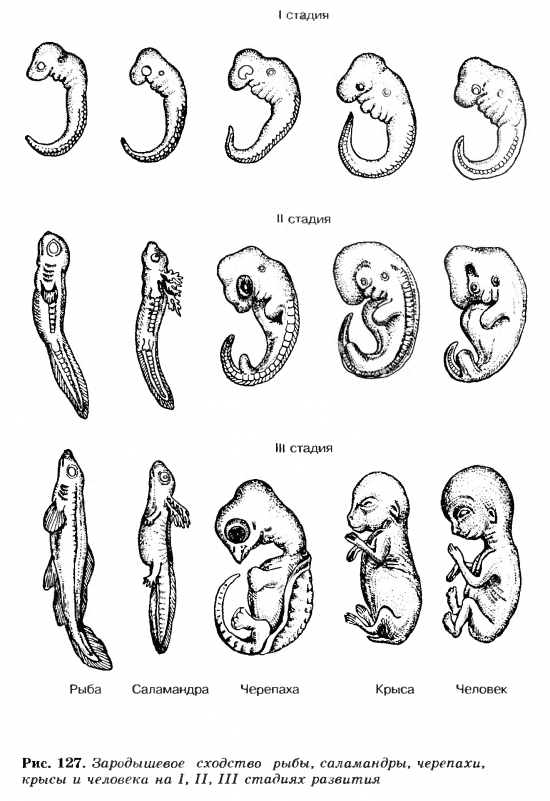 gif»>
gif»> gif»>
gif»> gif»>
gif»> gif»>
gif»>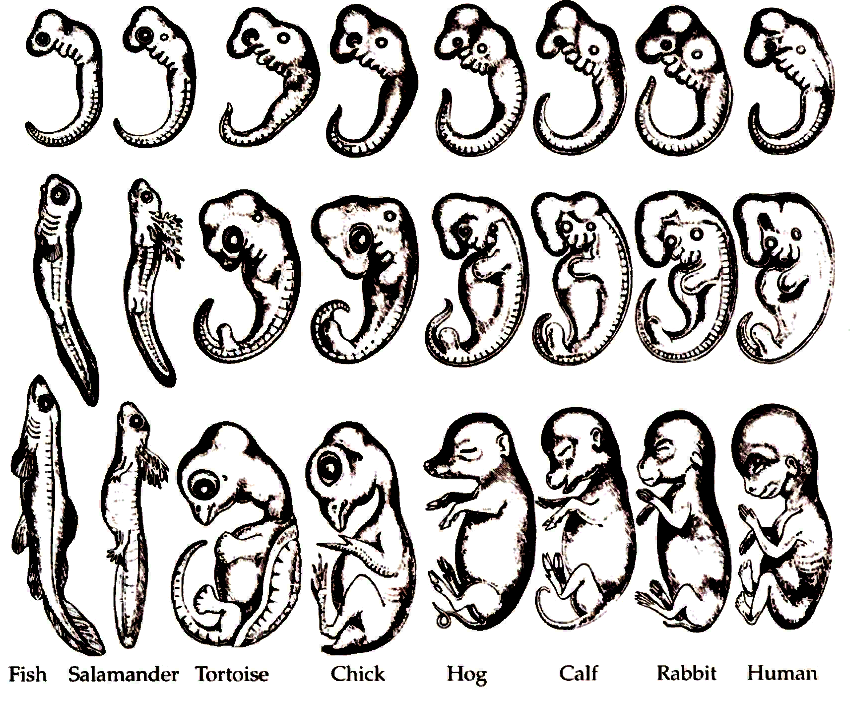 gif»>
gif»> gif»>
gif»>контакты карта сайта почтовый сервер управление поддержка
199034, Санкт-Петербург, Университетская наб., 7-9
© Санкт-Петербургский государственный университет, 2006-2017
биофак СПбГУ
биофак СПбГУ
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 gif»>
gif»> gif»>
gif»> gif»>
gif»> gif»>
gif»>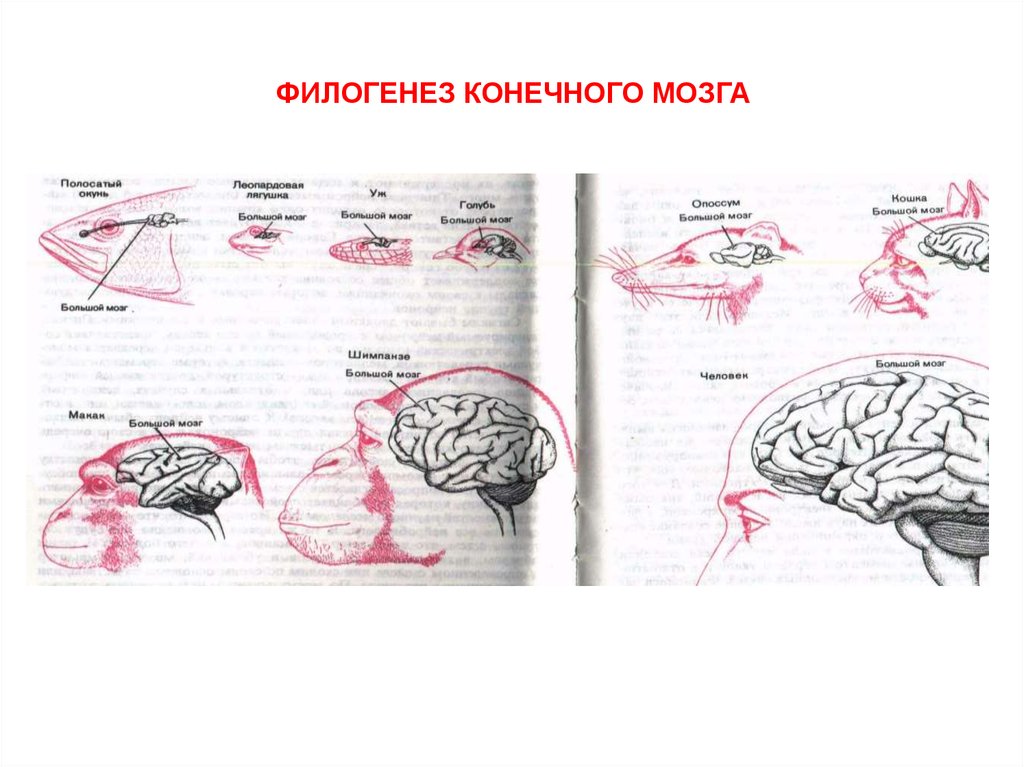 gif»>
gif»> gif»>
gif»> gif»>
gif»> gif»>
gif»> gif»>
gif»> gif»>
gif»> gif»>
gif»> gif»>
gif»> gif»>
gif»> gif»>
gif»>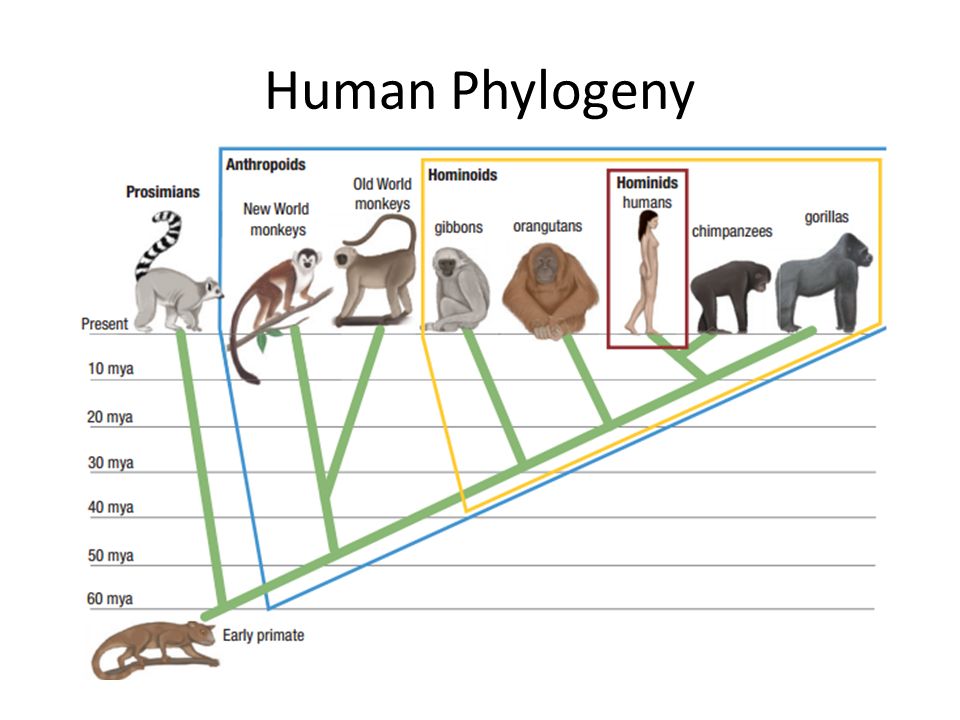 gif»>
gif»>контакты карта сайта почтовый сервер управление поддержка
199034, Санкт-Петербург, Университетская наб., 7-9
© Санкт-Петербургский государственный университет, 2006-2017
404 Cтраница не найдена
Размер:
AAA
Изображения Вкл. Выкл.
Обычная версия сайта
К сожалению запрашиваемая страница не найдена.
Но вы можете воспользоваться поиском или картой сайта ниже
|
|
 ru
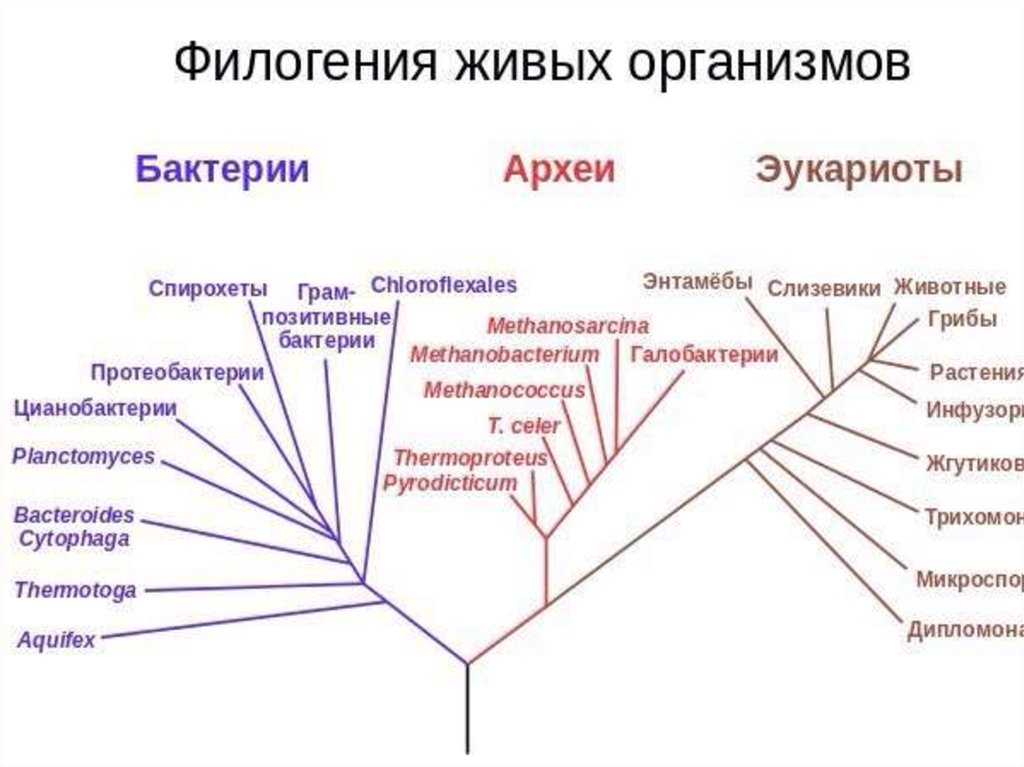
ruФилогенез.
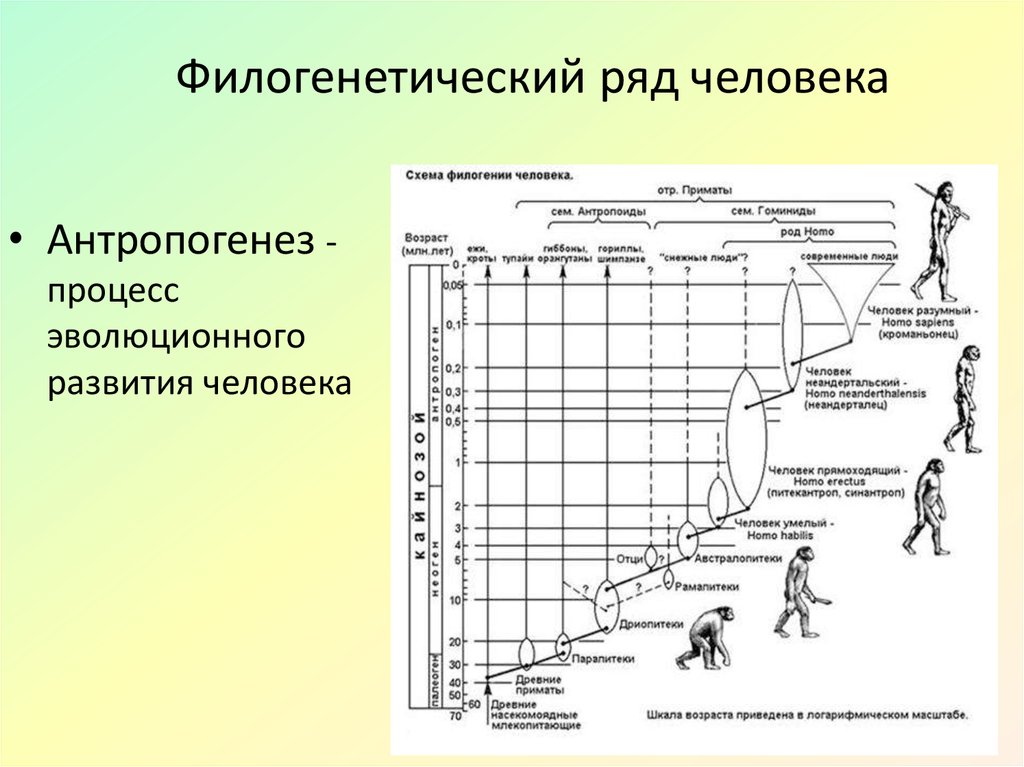 Эволюционные преобразования — презентация онлайн
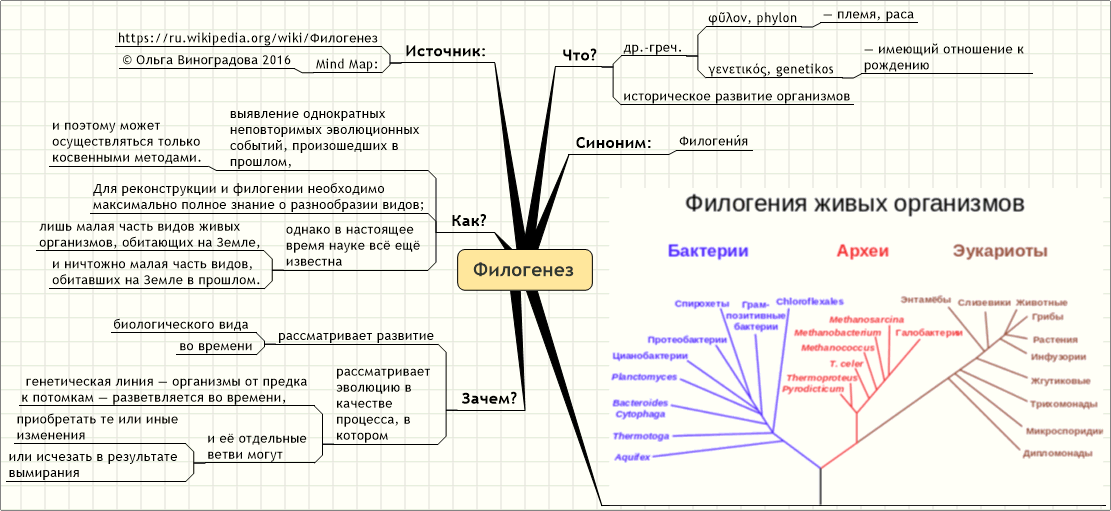
Эволюционные преобразования — презентация онлайн1. ФИЛОГЕНЕЗ
• Термин «Филогения» (филогенез) былвведен Э.Геккелем во 2 половине XIX века. В
широком понимании – филогения –это
эволюция в современном понимании, а в
узком смысле – это последовательность
ветвлений родословного древа.
• Эволюционные преобразования:
• — образование и вымирание видов
• — преобразование онтогенеза.
• Перестройки онтогенеза — предпосылка
филогенетических преобразований.
• «Геккелевская триада» — основа научного
подхода к реконструкции филогенеза,
базируется на комплексном использовании
методов:
• -эмбриологических,
• -палеонтологических
• -сравнительно-анатомических
• Биогенетический закон Геккеля, Мюллера:
«Онтогения есть краткое повторение
филогении».
• Биогенетический закон был широко
использован при создании современной
классификации организмов (таксономии),
построении схем филогенетического
родства организмов.

• При обосновании биогенетического закона в 1866
• Э. Геккель ввел термины «Палингенезы» и
«Ценогенезы».
• «Палингенезы» по Геккелю — повторения
(рекапитуляция) более или менее далёких этапов
филогенеза в процессе зародышевого развития
особи.
• «Ценогенезы» по Геккелю — признаки, которые,
нарушают проявления палингенезов, не позволяют
проследить в ходе онтогенеза современных форм
последовательность этапов филогенеза их предков, т.
е. нарушают биогенетический закон.
• А.Н. Северцов переосмыслил значение явлений, которые Геккель
обозначил терминами «Палингенезы» и «Ценогенезы».
• Термин «Ценогенезы» А. Н. Северцов использовал при описании
провизорных приспособлений — эмбрио-адаптаций. Т.е., это
приспособительные признаки, возникающие у зародышей и
личинок, адаптирующие их к особенностям среды обитания. У
взрослых организмов ценогенезы не сохраняются.
• Ценогенезы у амниот:
• -зародышевые оболочки
• -желточный мешок
• -аллантоис
• Эволюционное значение Ценогенезов — они проявляются только
на ранних стадиях онтогенеза, не изменяют типа организации
взрослого организма, но обеспечивают более высокую
вероятность выживания потомства.

• Э. Геккель рассматривал эволюцию взрослых организмов в отрыве
от эволюции зародышей. В сравнительной эмбриологии Геккель
делал главный акцент на рекапитуляции — «Палингенезах».
• А.Н. Северцов, вводя в 1922 году новый термин
«Филэмбриогенезы», подчеркивал, что «Филэмбриогенезы»,
напротив — отклонения от онтогенеза, характерного для предков,
проявляющиеся в эмбриогенезе, но имеющие адаптивное
значение у взрослых форм.
• А.Н. Северцов указал, что отношения между онтогенезом и
филогенезом гораздо сложнее, чем это описано в
Биогенетическом законе, отмечая:
• Во-первых, различия в сроках появления новых признаков в
онтогенезе, которые далеко не всегда «надставляют»
(рекапитулируют) ряд последовательных стадий онтогенеза.
• Во-вторых, вся последовательность стадий онтогенеза может
оказаться радикально преобразованной.
• Таким образом, согласно теории А.Н. Северцова
Филэмбриогенезы – это:
• — изменения, возникающие в результате перестройки
генотипа в ходе эмбрионального развития и имеющие
филогенетическое значение.
• — перестройки генотипа, которые могут происходить на
основе мутационного процесса и естественного отбора не
только в конце развития органа, как считал Геккель, но и
начальных, средних, и конечных этапах онтогенеза.
• — изменения, которые могут активно влиять на эволюцию
данного вида.
• Процесс превращения «мутаций в адаптации» протекает
под контролем естественного отбора и затрагивает все
стадии онтогенеза.
• 1.Анаболии – изменения, возникающие к концу эмбриогенеза, когда
формирование органов и систем почти завершено. К анаболиям
относят такие явления, как приобретение специфической формы тела
камбалой лишь после того, как из икринки вылупляется малек,
неотличимый от других рыб, а также появление изгибов позвоночника,
сращение швов в мозговом черепе, окончательное перераспределение
кровеносных сосудов в организме млекопитающих и человека.
• 2.Девиация – изменения, возникающие на средних стадиях
эмбриогенеза. Пример — развитие сердца в онтогенезе
млекопитающих, у которых оно рекапитулирует стадию трубки,
двухкамерное и трехкамерное строение, но стадия формирования
неполной перегородки, характерной для пресмыкающихся,
вытесняется развитием перегородки, построенной и расположенной
иначе и характерной только для млекопитающих.

3.Архаллаксис – изменения, возникающие на ранних этапах
эмбриогенеза. Например, классическим примером архаллаксиса
является развитие волос у млекопитающих, закладка которых наступает
на очень ранних стадиях развития.
• В основе Филэмбриогенезов — те же механизмы,
которые обусловливают врожденные пороки
развития. От пороков развития их отличает адаптивная
ценность, т.е. полезность и закрепленность
естественным отбором в филогенезе.
• За счет анаболии в онтогенезах потомков полностью
реализуется основной биогенетический закон, т.е.
происходят рекапитуляции всех предковых стадий
развития.
• При девиациях рекапитулируют только ранние
предковые стадии
• Архаллаксисы полностью не допускают
рекапитуляции
• Кроме ценогенезов и филэмбриогенезов в эволюции онтогенеза могут
обнаруживаться отклонения:
• -времени закладки органов — гетерохронии
• -места их развития — гетеротопии.
• Гетерохронии и гетеротопии приводят к изменению
взаимосоответствия развивающихся структур и проходят жёсткий
контроль естественного отбора.
• Гетерохронии — сдвиги во времени закладок наиболее важных органов
в группах, эволюционирующих по типу ароморфоза. Так, у
млекопитающих, и в особенности у человека, дифференцировка
переднего мозга существенно опережает развитие других его отделов.
• Гетеротопии — формирование новых пространственных и
функциональных связей между органами, обеспечивая в дальнейшем
их совместную эволюцию. Например, сердце, располагающееся у рыб
под глоткой, обеспечивает эффективное поступление крови в
жаберные артерии для газообмена. Перемещаясь в загрудинную
область у наземных позвоночных, оно развивается и функционирует
уже в едином комплексе с новыми органами дыхания — лёгкими,
выполняя и здесь, в частности, функцию доставки крови к дыхательной
системе для газообмена.
В свою очередь, указанные выше изменения хода
онтогенеза могут реализоваться уже на уровне
филогенеза в виде:
-ароморфоза
-идиоадаптации
-дегенерации
• Ароморфозы — направление эволюции, при котором
появляются качественно новые морфофизиологические
особенности, приводящие к резкому повышению уровня их
организации.

• Ароморфозы позволяют организмам заселять принципиально
новые адаптивные зоны.
• Это путь морфофизиологического прогресса, ведущий
к возникновению организмов все более сложных и менее
зависимых от условий внешней среды.
• Благодаря появлению и накоплению ароморфозов возникают
крупные таксоны, такие как класс, отдел, тип, царство.
• Идиоадаптации — приспособление организмов к конкретным
условиям среды, которое не ведет к существенным изменениям
уровня организации. Благодаря идиоадаптациям возникают
такие таксоны, как род, семейство.
• Идиоадаптации способствуют появлению у животных и растений
не общих, а частных изменений
• Идиоадаптации могут затрагивать: 1).Изменение строения тела.
2).Изменение цвета наружных покров. 3).Характер
передвижения.
Еще один путь к достижению биологического прогресса – Дегенерация. Это
морфофизиологический регресс, упрощение организации, утрата отдельных органов или систем
(редукция) в связи с переходом организмов к сидячему образу жизни или паразитизму.

Например:
1). Паразитические ленточные черви утратили нервную, мышечную и даже пищеварительную
системы. Питательные вещества поступают в их организм через покровы. У этих паразитов очень
сильно развита способность к размножению.
2).Сидячие, прикрепленные формы могут испытывать редукцию (недоразвитие или полное
исчезновение) нервной и опорно-двигательной систем. Например, двустворчатые моллюски,
перешедшие к пассивной фильтрации, утратили не только мозг, но и голову как таковую. В то
время, как их активно плавающие сородичи осьминоги за высокое развитие головного мозга и глаз
названы «приматами мира беспозвоночных».
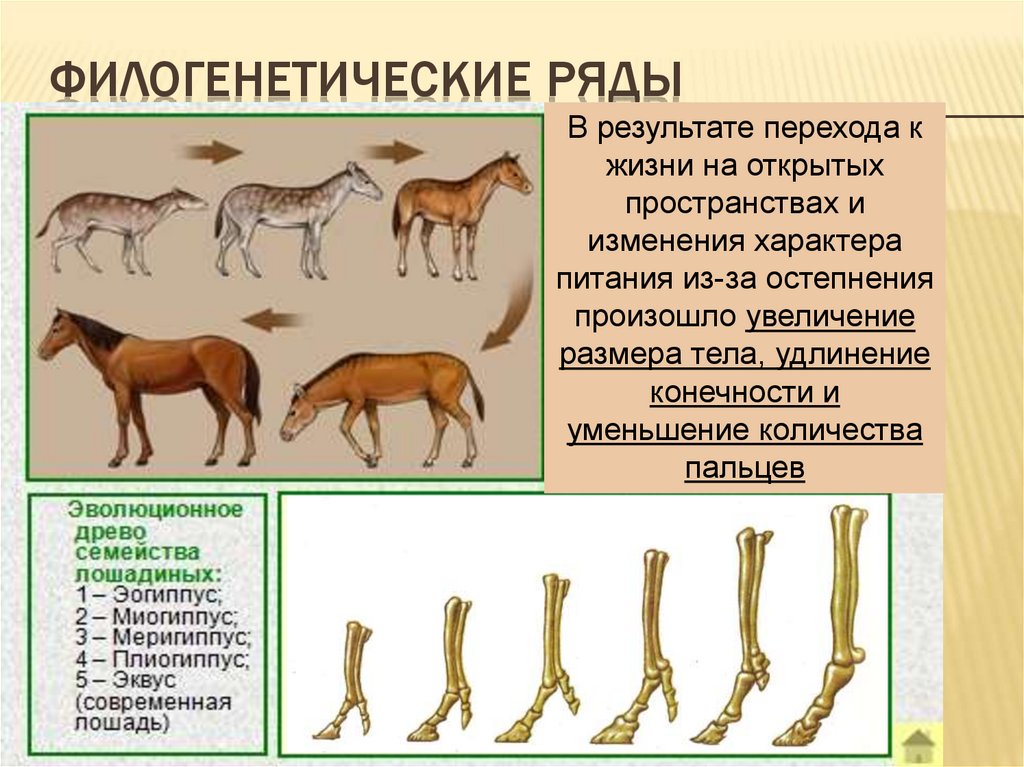
21. ОБЩАЯ СХЕМА НАПРАВЛЕНИЙ ФИЛОГЕНЕЗА СИСТЕМ ОРГАНОВ ПОЗВОНОЧНЫХ
ОБЩАЯ СХЕМА НАПРАВЛЕНИЙ ФИЛОГЕНЕЗА СИСТЕМ ОРГАНОВ ПОЗВОНОЧНЫХ22. Эволюция органов дыхания. Жабры Легкие
Плавательный пузырь рыб (А—костной;Б—кистеперой) и развитие легких у человека
(В—ранние стадии): 1—средняя кишка,
2—плавательный пузырь, 3—глотка,
4—развивающиеся легкие
23.
 ЛЕГКИЕ АМФИБИЙ И РЕПТИЛИЙСхема легких хвостатых амфибий и рептилий (а — Necturus;
ЛЕГКИЕ АМФИБИЙ И РЕПТИЛИЙСхема легких хвостатых амфибий и рептилий (а — Necturus;б — саламандра и рептилия; в — ящерица; г 2 черепаха): 1 — бронхи; 2 — внутрилегочний бронх; З — трахея
24. СТРОЕНИЕ АЛЬВЕОЛ
1 — венозная кровь; 2 — воздух; 3 – артериальная кровь;4 — легочные капиляры; 5 — воздух; 6 — венозная кровь легочных артерий;
7 — оксигенация крови;
8 — эритроциты; 9 — стенки альвеол.
25. ОСНОВНЫЕ НАПРВЛЕНИЯ ЭВОЛЮЦИИ КРОВЕНОСНОЙ СИСТЕМЫ ПОЗВОНОЧНЫХ
Строение кровеносной системы водных (А) и наземных (Б) позвоночных:1—жаберные артерии, 2—сонная артериям—передняя кардинальная вена,
4— задняя кардинальная вена, 5—спинная аорта, 6—кювьеров проток, 7—подкишечная вена,
8—печеночная вена, 9—брюшная аорта, 10—задняя (нижняя) полая вена, 11—воротная вена печени,
12—легочная вена, 13—легочная артерия
26. ИЗМЕНЕНИЯ КРОВЕНОСНОЙ СИСТЕМЫ ПОЗВОНОЧНЫХ ПРИ ВЫХОДЕ НА СУШУ
27. ИЗМЕНЕНИЯ КРОВЕНОСНОЙ СИСТЕМЫ ПОЗВОНОЧНЫХ ПРИ ПОЯВЛЕНИИ ТЕПЛОКРОВНЫХ ЖИВОТНЫХ
28.
 КОЖА АМФИБИИ
КОЖА АМФИБИИ29. КОЖА РЕПТИЛИИ
Поперечный разрез кожи рептилии:1 — эпидермис; 2 — кориум; 3 — роговой слой; 4 — пигментные клетки; 5 — кожные
окостенения.
30. КОЖА МЛЕКОПИТАЮЩИХ:
• I ороговевшийэпителий; II
эпидермис; III
кориум; IV подкожная
клетчатка;
• 1 — волос; 2 — сальные
железы; 3 — мышцы
волоса; 4 — волосяная
сумка;
• 5 — потовая железа; 6
— рецепторы кожи; 7 жировая ткань; 8 кровеносные сосуды.
31. ЭВОЛЮЦИЯ ГОЛОВНОГО МОЗГА ПОЗВОНОЧНЫХ
• I — костистаярыба; II электрический
скат; III — лягушка;
IV — рептилии;
• V — птицы: VI –
млекопитающие
• 1 — передний
мозг; 2 — эпифиз;
З — гипофиз; 4 средний мозг; 5 мозжечок; 6 продолговатый
мозг; 7 промежуточный
мозг; 8 – мантия
32. ЭВОЛЮЦИЯ СКЕЛЕТА ПОЗВОНОЧНЫХ
Сравнительная характеристика скелета позвоночных (а 2 скелет рыбы; б 2 скелет наземного животного):1 — черепная коробка;
2 — висцеральные дуги; З — висцеральная часть черепа; 4 — позвоночник;
5 — ребра; 6 — скелет непарных плавников; 7 — грудные плавники;
7а — передние конечности; 8 — плечевой пояс; 9 — брюшные плавники;
9а — задние конечности; 10 — тазовый пояс.

СПАСИБО ЗА ВНИМАНИЕ!
Инверсия филогении человека: Homo покинул Африку как прямоходящий, вернулся как sapiens sapiens
1. Cann RL, Stoneking M, Wilson AC. Митохондриальная ДНК и эволюция человека. Природа. 1987; 325:31–36. doi: 10.1038/325031a0. [PubMed] [CrossRef] [Google Scholar]
2. Vigilant L, Stoneking M, Harpending H, Hawkes K, Wilson AC. Африканские популяции и эволюция митохондриальной ДНК человека. Наука. 1991; 253:1503–1507. doi: 10.1126/science.1840702. [PubMed] [CrossRef] [Академия Google]
3. Мэддисон Д.Р. Пересмотрено африканское происхождение митохондриальной ДНК человека. Сист Зоол. 1991; 40: 355–363. дои: 10.2307/2992327. [CrossRef] [Google Scholar]
4. Темплтон А.Р. Происхождение человека и анализ последовательностей митохондриальной ДНК. Наука. 1992; 255:737. [PubMed]
5. Хеджес Б.С., Кумар С., Тамура К., Стоункинг М. Происхождение человека и анализ последовательностей митохондриальной ДНК. Наука. 1992; 255: 737–739. doi: 10.1126/science.1738849. [PubMed] [CrossRef] [Академия Google]
doi: 10.1126/science.1738849. [PubMed] [CrossRef] [Академия Google]
6. Арнасон Ю. Гипотеза выхода из Африки и происхождение недавних людей: cherchez la femme (et l’homme). Ген. 2016; 585:9–12. [PubMed]
7. Арнасон Ю. Филогенетический взгляд на гипотезы выхода из Азии/Евразии и Африки в свете недавних молекулярных и палеонтологических находок. Ген. 2017; 627: 473–476. doi: 10.1016/j.gene.2017.07.006. [PubMed] [CrossRef] [Google Scholar]
8. Mallick S, Li H, Lipson M, Mathieson I, Gymrek M, Racimo F, et al. Проект Саймонса по разнообразию генома: 300 геномов из 142 различных популяций. Природа. 2016; 538: 201–206. дои: 10.1038/nature18964. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
9. Pagani L, Lawson DJ, Jagoda E, Mörseburg A, Eriksson A, Mitt M, et al. Геномные анализы информируют о миграционных событиях при заселении Евразии. Природа. 2016; 538: 238–242. doi: 10.1038/nature19792. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
10. Meyer M, Kircher M, Gansauge MT, Li H, Racimo F, Mallick S, et al. Последовательность генома с высоким охватом архаичного денисовца. Наука. 2012; 338: 222–226. doi: 10.1126/science.1224344. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Meyer M, Kircher M, Gansauge MT, Li H, Racimo F, Mallick S, et al. Последовательность генома с высоким охватом архаичного денисовца. Наука. 2012; 338: 222–226. doi: 10.1126/science.1224344. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
11. Meyer M, Fu Q, Aximu-Petri A, Glocke I, Nickel B, Arsuaga J-L, et al. Последовательность митохондриального генома гоминина из Сима-де-лос-Уэсос. Природа. 2014; 505:403–405. doi: 10.1038/nature12788. [PubMed] [CrossRef] [Google Scholar]
12. Prüfer K, Racimo F, Patterson F, Jay F, Sankararaman S, Sawyer S, et al. Полная последовательность генома неандертальца из Горного Алтая. Природа. 2014; 505:43–49. doi: 10.1038/nature12886. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
13. Kuhlwilm M, Gronau I, Hubisz MJ, de Filippo C, Prado-Martinez J, Kircher M, et al. Поток древних генов от ранних современных людей к восточным неандертальцам. Природа. 2016; 530:429–433. doi: 10.1038/nature16544. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
14. Meyer M, Arsuaga J-L, de Filippo C, Nagel S, Aximu-Petri A, Nickel B, et al. Последовательности ядерной ДНК гомининов Sima de los Huesos среднего плейстоцена. Природа. 2016; 531: 504–507. doi: 10.1038/nature17405. [PubMed] [CrossRef] [Академия Google]
Meyer M, Arsuaga J-L, de Filippo C, Nagel S, Aximu-Petri A, Nickel B, et al. Последовательности ядерной ДНК гомининов Sima de los Huesos среднего плейстоцена. Природа. 2016; 531: 504–507. doi: 10.1038/nature17405. [PubMed] [CrossRef] [Академия Google]
15. Posth C, Wißing C, Kitagawa K, Pagani L, van Holstein L, Racimo F, et al. Глубоко расходящийся архаичный митохондриальный геном обеспечивает более низкую временную границу для потока африканских генов к неандертальцам. Связь с природой 2017. 10.1038/ncomms16046. [Бесплатная статья PMC] [PubMed]
16. Hubisz MJ, Williams AL, Siepel A. Картирование потока генов между древними гомининами посредством демографически-осведомленного вывода графа рекомбинации предков. bioRxiv. 2019. 10.1101/687368. [Бесплатная статья PMC] [PubMed]
17. Чен Л., Вольф А.Б., Фу В., Ли Л., Акей Дж.М. Выявление и интерпретация явного неандертальского происхождения у африканцев. Клетка. 2020; 180:1–11. doi: 10.1016/j.cell.2020.01.012. [PubMed] [CrossRef] [Google Scholar]
18. Bischoff JL, Williams RW, Rosenbauer RJ, Aranburu A, Arsuaga JL, García N, et al. Даты U-серии с высоким разрешением по гоминидам Сима-де-лос-Уэсос дают 600 +/- 66 тысяч лет: значение для эволюции ранней неандертальской линии. J Archaeol Sci. 2007; 34: 763–770. doi: 10.1016/j.jas.2006.08.003. [Перекрестная ссылка] [Академия Google]
Bischoff JL, Williams RW, Rosenbauer RJ, Aranburu A, Arsuaga JL, García N, et al. Даты U-серии с высоким разрешением по гоминидам Сима-де-лос-Уэсос дают 600 +/- 66 тысяч лет: значение для эволюции ранней неандертальской линии. J Archaeol Sci. 2007; 34: 763–770. doi: 10.1016/j.jas.2006.08.003. [Перекрестная ссылка] [Академия Google]
19. Арсуага Дж. Л., Мартинес И., Арнольд Л. Дж., Аранбуру А., Грасия-Теллес А., Шарп В. Д. и другие. Неандертальские корни: черепные и хронологические свидетельства из Сима-де-лос-Уэсос. Наука. 2014; 344:1358–1363. doi: 10.1126/science.1253958. [PubMed] [CrossRef] [Google Scholar]
20. Arsuaga JL, Carretero J-M, Lorenzo C, Gómez-Olivencia A, Pablos A, Rodríguez L, et al. Посткраниальная морфология людей среднего плейстоцена из Сима-де-лос-Уэсос, Испания. Proc Natl Acad Sci USA. 2015;112:11524–11529. doi: 10.1073/pnas.1514828112. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
21. Zanolli C, Martinón-Torres M, Bernardini F, Boschian G, Coppa A, Dreossi D, et al.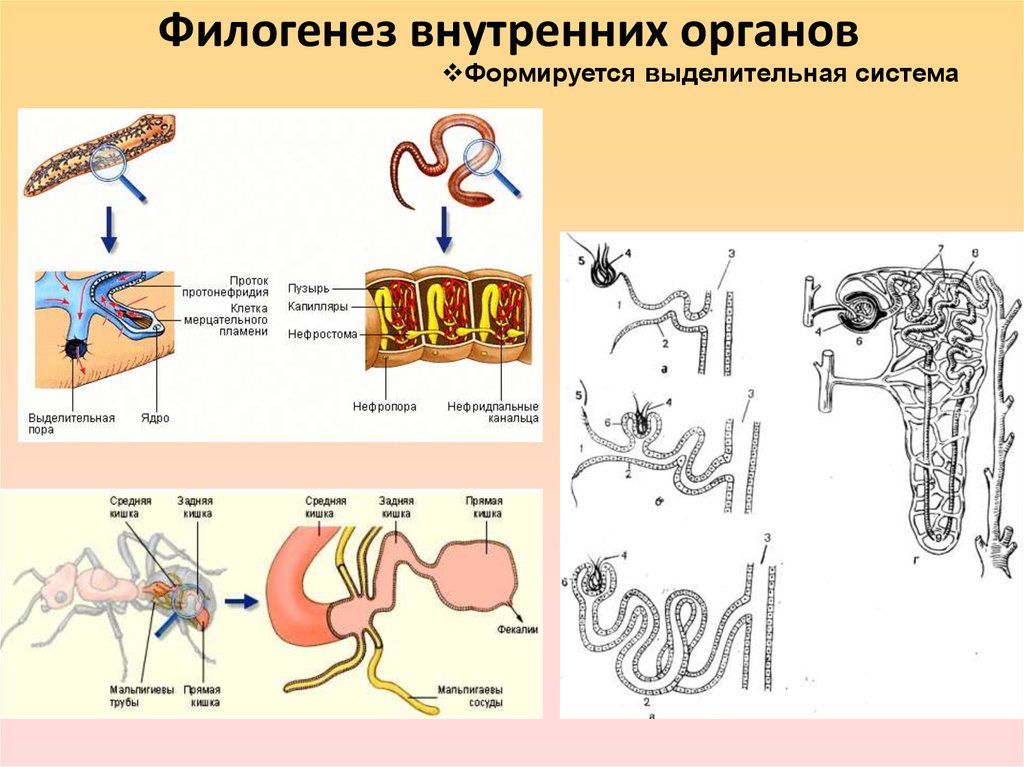 Зубные остатки человека среднего плейстоцена (MIS 12) из Фонтана-Рануччо (Лацио) и Визольяно (Фриули-Венеция-Джулия), Италия. Сравнительная эндоструктурная оценка высокого разрешения. ПЛОС ОДИН. 2018;13(10):e0189773. doi: 10.1371/journal.pone.0189773. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Зубные остатки человека среднего плейстоцена (MIS 12) из Фонтана-Рануччо (Лацио) и Визольяно (Фриули-Венеция-Джулия), Италия. Сравнительная эндоструктурная оценка высокого разрешения. ПЛОС ОДИН. 2018;13(10):e0189773. doi: 10.1371/journal.pone.0189773. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
22. Hershkovitz I, Weber GW, Quam R, Duval M, Grün R, Kinsley L, et al. Самые ранние современные люди за пределами Африки. Наука. 2018; 359: 456–459. doi: 10.1126/science.aap8369. [PubMed] [CrossRef] [Google Scholar]
23. Тринкаус Э. Углы шейки бедра у ранних современных людей Квафзех-Схул и уровни активности среди неполовозрелых гоминидов ближнего восточного среднего палеолита. Джей Хам Эвол. 1993; 25: 393–416. doi: 10.1006/jhev.1993.1058. [CrossRef] [Google Scholar]
24. Бермудес де Кастро Дж. М., Арсуага Дж. Л., Карбонелл Э., Росас А., Мартинес I, Москера М. Гоминид из нижнего плейстоцена Атапуэрки, Испания: возможный предок неандертальцев и современных людей. Наука. 1997;276:1392–1395. doi: 10.1126/science.276.5317.1392. [PubMed] [CrossRef] [Google Scholar]
Наука. 1997;276:1392–1395. doi: 10.1126/science.276.5317.1392. [PubMed] [CrossRef] [Google Scholar]
25. Стайнер М.С., Манро Н.Д., Суровелл Т.А., Чернов Э., Бар-Йосеф О. Палеолитические импульсы роста населения, свидетельствующие об эксплуатации мелких животных. Наука. 1999; 283:190–194. doi: 10.1126/science.283.5399.190. [PubMed] [CrossRef] [Google Scholar]
26. Niewoehner WA. Поведенческие выводы из ранней современной руки Схул / Кафзе сохранились. Proc Natl Acad Sci U S A. 2001; 98:2979–2984. doi: 10.1073/pnas.041588898. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
27. Shen GJ, Ku TL, Cheng H, Edwards RL, Yuan ZX, Wang Q. Высокоточное датирование U-серией местности 1 в Чжоукоудянь, Китай. Джей Хам Эвол. 2001; 41: 679–688. doi: 10.1006/jhev.2001.0516. [PubMed] [CrossRef] [Google Scholar]
28. Shen GJ, Wang W, Wang Q, Zhao JX, Collerson K. Датирование U-серией стоянки гоминидов Люцзян в Гуанси, Южный Китай. Джей Хам Эвол. 2002;43:817–829. doi: 10.1006/jhev.2002.0601. [PubMed] [CrossRef] [Академия Google]
doi: 10.1006/jhev.2002.0601. [PubMed] [CrossRef] [Академия Google]
29. Сяо Дж., Цзинь Ч.З., Чжу Ю.З. Возраст ископаемого человека Дали на севере Центрального Китая определен на основе хроностратиграфии лёссово-палеопочвенной последовательности. Quat Sci Rev. 2002; 21: 2191–2198. doi: 10.1016/S0277-3791(02)00011-2. [CrossRef] [Google Scholar]
30. Shen GJ, Cheng H, Edwards RL. Масс-спектрометрическое датирование U-серии новой пещеры в Чжоукоудяне, Китай. J Archaeol Sci. 2004; 31: 337–342. doi: 10.1016/j.jas.2003.09.004. [CrossRef] [Google Scholar]
31. Wu X. 2004. О происхождении современного человека в Китае. Кват Инт. 2004; 117: 131–140. doi: 10.1016/S1040-6182(03)00123-X. [Перекрестная ссылка] [Академия Google]
32. Carbonell E, Bermúdez de Castro JM, Arsuaga JL, Allue E, Bastir M, Benito A, et al. Раннеплейстоценовый гоминин из Атапуэрка-TD6, Испания. Proc Natl Acad Sci USA. 2005; 102: 5674–5678. doi: 10.1073/pnas.0501841102. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
33. Rosenberg R, Zuné L, Ruff CB. Размер тела, пропорции тела и энцефализация архаичного человека среднего плейстоцена из северного Китая. Proc Natl Acad Sci U S A. 2006;103:3552–3556. doi: 10.1073/pnas.0508681103. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Rosenberg R, Zuné L, Ruff CB. Размер тела, пропорции тела и энцефализация архаичного человека среднего плейстоцена из северного Китая. Proc Natl Acad Sci U S A. 2006;103:3552–3556. doi: 10.1073/pnas.0508681103. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
34. Карбонелл Э., Бермудес де Кастро Х.М., Парес Х.М., Перес-Гонсалес А., Куэнка-Бескос Г., Олле А. и др. Первый гоминин Европы. Природа. 2008; 452:465–469. doi: 10.1038/nature06815. [PubMed] [CrossRef] [Google Scholar]
35. de Lumley MA, Grimaud-Hervé D, Li T, Feng XB, Wang Z. Les cranes d’ Homo erectus du site de l’Homme de Yunxian, Quyanhe -kou, Qingqu, Yunxian I и Yunxian II. В: де Ламли Х., Ли Т., редакторы. Le site de l’Homme de Yunxian, Éditions Recherche sur les Civilisations. Выпуски CNRS. 2008. стр. 381–466. [Академия Google]
36. Zhu RX, Potts R, Pan YX, Yao HT, Lu LO, Zhao X. Ранние свидетельства существования рода Homo в Восточной Азии. Джей Хам Эвол. 2008;55:1075–1085. doi: 10.1016/j.jhevol.2008.08.005. [PubMed] [CrossRef] [Google Scholar]
doi: 10.1016/j.jhevol.2008.08.005. [PubMed] [CrossRef] [Google Scholar]
37. Shen GJ, Gao X, Granger DE. Возраст Чжоукоудяня Homo erectus определяется с помощью 26 Al/ 10 Датировка захоронения. Природа. 2009; 458:198–200. doi: 10.1038/nature07741. [PubMed] [CrossRef] [Google Scholar]
38. Stiner MC, Barkai R, Gopher A. Совместная охота и раздел мяса 400-200 тыс. лет назад в пещере Квесем, Израиль. Proc Natl Acad Sci USA. 2009 г.;106:13207–13212. doi: 10.1073/pnas.0
4106. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
39. Бермудес де Кастро Х.М., Мартинон-Торрес М., Гомес-Роблес А., Прадо-Симон Л., Карбонелл Э. Новые человеческие свидетельства раннего плейстоценового поселения Европы, с участка Сима-дель-Элефанте (Сьерра-де-Атапуэрка, Бургос, Испания) Quat Int. 2010; 223–224:431–433. doi: 10.1016/j.quaint.2009.07.024. [CrossRef] [Google Scholar]
40. Shen GJ, Fang YS, Bischoff JL, Feng YX, Zhao JX. Масс-спектрометрическое датирование U-серии стоянки гоминидов Чаосянь в Иньшане, восточный Китай. Кват Инт. 2010; 211:24–28. doi: 10.1016/j.quaint.2009.02.020. [CrossRef] [Google Scholar]
Кват Инт. 2010; 211:24–28. doi: 10.1016/j.quaint.2009.02.020. [CrossRef] [Google Scholar]
41. Vialet A, Guipert G, Jianing H, Xiaobo F, Zune L, Youping W. Homo erectus с китайских памятников Yunxian и Nankin: антропологические выводы с использованием методов трехмерного виртуального изображения. CR Палевол. 2010;9:331–339. doi: 10.1016/j.crpv.2010.07.017. [CrossRef] [Google Scholar]
42. Wagner GA, Krbetschek M, Degering D, Bahain JJ, Shao Q, Falguères C. Радиометрическое датирование типового участка для Homo heidelbergensis в Мауэре, Германия. Proc Natl Acad Sci USA. 2010;107:19726–19730. doi: 10.1073/pnas.1012722107. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
43. Бермудес де Кастро Х.М., Мартинон-Торрес М., Гомес-Роблес А., Прадо-Симон Л., Мартин-Франсес Л., Лапреса М. Человек раннего плейстоцена нижняя челюсть из пещеры Сима-дель-Элефанте (TE) в Сьерра-де-Атапуэрка (Испания): сравнительное морфологическое исследование. Джей Хам Эвол. 2011;61:12–25. doi: 10.1016/j.jhevol.2011.03.005. [PubMed] [CrossRef] [Google Scholar]
Джей Хам Эвол. 2011;61:12–25. doi: 10.1016/j.jhevol.2011.03.005. [PubMed] [CrossRef] [Google Scholar]
44. Манзи Г. По следам рода Homo между архаичной и производной морфологиями. J Anthropol Sci. 2012;90:99–116. [PubMed] [Google Scholar]
45. Розенберг К.Р., Ву С. Через нее протекает река: Происхождение современного человека в Восточной Азии. В: Smith FH, Ahern JCM, редакторы. Происхождение современного человека. 2. Хобокен: Уайли; 2013. С. 89–121. [Google Scholar]
46. Shen GJ, Wu XZ, Wang Q, Tu H, Feng YX, Zhao JX. Масс-спектрометрическое датирование U-серии пещеры Хуанлун в провинции Хубэй, Центральный Китай: свидетельство раннего присутствия современных людей в Восточной Азии. Джей Хам Эвол. 2013;65:162–167. doi: 10.1016/j.jhevol.2013.05.002. [PubMed] [CrossRef] [Академия Google]
47. Wu X, Athreya S. Описание геологического контекста, дискретных признаков и линейной морфометрии гоминина среднего плейстоцена из Дали, провинция Шэньси, Китай.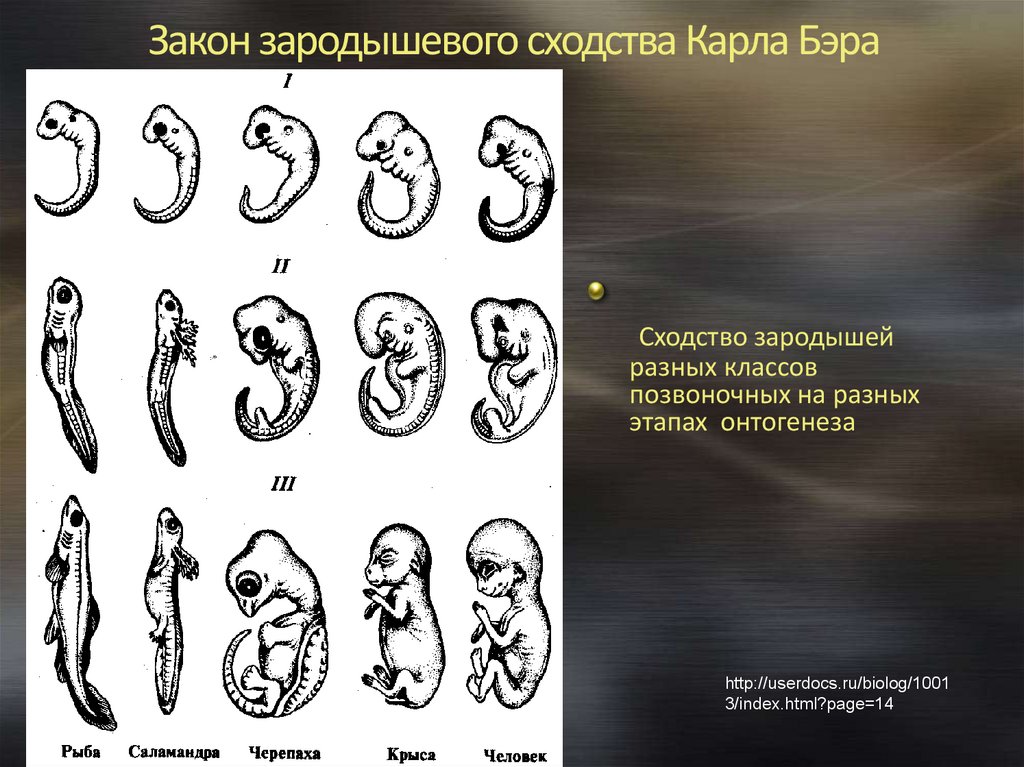 Am J Phys Антропол. 2013;150(1):141–157. doi: 10.1002/ajpa.22188. [PubMed] [CrossRef] [Google Scholar]
Am J Phys Антропол. 2013;150(1):141–157. doi: 10.1002/ajpa.22188. [PubMed] [CrossRef] [Google Scholar]
48. Антон С.К., Поттс Р., Айелло Л.С. Эволюция раннего Homo : интегрированная биологическая перспектива. Наука. 2014;345(6192):1236828. doi: 10.1126/science.1236828. [PubMed] [CrossRef] [Академия Google]
49. Шэнь Г., Ту Х., Сяо Д., Цю Л., Фэн Ю.С., Чжао Дж.С. Возраст стоянки гоминидов Маба на юге Китая: свидетельства датирования серии U пещеры Южного отделения. Кват Геохронол. 2014; 23:56–62. doi: 10.1016/j.quageo.2014.06.004. [CrossRef] [Google Scholar]
50. Wu XJ, Trinkaus E. Ветвь нижней челюсти Xujiayao 14 и плейстоценовая вариация нижней челюсти Homo . ЧР Палевол. 2014;13:333–341. doi: 10.1016/j.crpv.2013.10.002. [CrossRef] [Google Scholar]
51. Wu XJ, Crevecoeur I, Liu W, Xing S, Trinkaus E. Височные лабиринты восточно-евразийских плейстоценовых людей. Proc Natl Acad Sci U S A. 2014; 111:10509–10513. doi: 10.1073/pnas.1410735111. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
52. Xing S, Martinón-Torres M, de Castro JMB, Zhang Y, Fan X, Zheng L, et al. Зубы гоминида среднего плейстоцена из пещеры Лунтань, Хесянь, Китай. ПЛОС ОДИН. 2014;9(12):e114265. doi: 10.1371/journal.pone.0114265. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
53. Dennell RW. Субхрон Харамильо, горизонт геохронологического маркера в палеоантропологической летописи Китая. Квотерн Интернешнл. 2015;389: 241–254. doi: 10.1016/j.quaint.2014.11.006. [CrossRef] [Google Scholar]
54. Zhu ZY, Dennell R, Huang WW, Wu Y, Rao ZG, Qiu SF, et al. Новая датировка черепа Homo erectus из Ланьтяня (Гунванлин), Китай. Джей Хам Эвол. 2015; 78: 144–157. doi: 10.1016/j.jhevol.2014.10.001. [PubMed] [CrossRef] [Google Scholar]
55. Бермудес де Кастро Дж. М., Мартинон-Торрес М., Роселл Дж., Бласко Р., Арсуага Дж. Л., Карбонелл Э. Преемственность против Ранний плейстоцен и начало среднего плейстоцена. Нижнечелюстные доказательства. Quat Sci Rev. 2016; 153:51–62. doi: 10.1016/j.quascirev.2016.10.010. [Перекрестная ссылка] [Академия Google]
Нижнечелюстные доказательства. Quat Sci Rev. 2016; 153:51–62. doi: 10.1016/j.quascirev.2016.10.010. [Перекрестная ссылка] [Академия Google]
56. Ao H, Liu C-R, Roberts AP, Zhang P, Xu X. Обновленный возраст гоминина Xujiayao из бассейна Nihewan в Северном Китае: последствия для эволюции человека в среднем плейстоцене в Восточной Азии. Джей Хам Эвол. 2017;106:54–65. doi: 10.1016/j.jhevol.2017.01.014. [PubMed] [CrossRef] [Google Scholar]
57. Athreya S, Wu X. Многомерная оценка черепа гоминина Дали из Китая: морфологическое сходство и значение для эволюции плейстоцена в Восточной Азии. Am J Phys Антропол. 2017;164:679–701. doi: 10.1002/ajpa.23305. [PubMed] [CrossRef] [Google Scholar]
58. Бермудес де Кастро Х.М., Мартинон-Торрес М., Мартин-Франсес Л., Модесто-Мата М., Мартинес-де-Пинильос М., Гарсия С., Карбонелл Е. Homo antecessor : Состояние искусства восемнадцать лет спустя. Кват Инт. 2017; 433:22–31. doi: 10.1016/j.quaint.2015.03.049. [CrossRef] [Google Scholar]
59. Han F, Bahain JJ, Deng C, Boëda É, Hou Y, Wei G, et al. Самые ранние свидетельства поселения гоминидов в Китае: комбинированное датирование электронно-спиновым резонансом и урановой серией (ESR / U-серия) ископаемых зубов млекопитающих из пещеры Лунгупо. Кват Инт. 2017; 434:75–83. doi: 10.1016/j.quaint.2015.02.025. [Перекрестная ссылка] [Академия Google]
Han F, Bahain JJ, Deng C, Boëda É, Hou Y, Wei G, et al. Самые ранние свидетельства поселения гоминидов в Китае: комбинированное датирование электронно-спиновым резонансом и урановой серией (ESR / U-серия) ископаемых зубов млекопитающих из пещеры Лунгупо. Кват Инт. 2017; 434:75–83. doi: 10.1016/j.quaint.2015.02.025. [Перекрестная ссылка] [Академия Google]
60. Huguet R, Vallverdú J, Rodríguez-Alvarez XP, Terradillos-Bernal M, Bargalló A, Lombera-Hermida A, et al. Уровень TE9c Сима-дель-Элефанте (Сьерра-де-Атапуэрка, Испания): комплексный подход. Кват Инт. 2017; 433: 278–295. doi: 10.1016/j.quaint.2015.11.030. [CrossRef] [Google Scholar]
61. Li ZY, Wu XJ, Zhou LP, Liu W, Gao X, Nian XM, Trinkaus E. Архаичные человеческие черепа позднего плейстоцена из Сюйчана, Китай. Наука. 2017; 355:969–972. doi: 10.1126/science.aal2482. [PubMed] [CrossRef] [Академия Google]
62. Мартин-Альбаладехо, Мартинон-Торрес М., Гарсия-Гонсалес Р., Арсуага Х.Л., Бермудес де Кастро Х.М. Морфометрический анализ нижних первых моляров Атапуэрка-Сима-де-лос-Уэсос. Кват Инт. 2017; 433:156–162. doi: 10.1016/j.quaint.2015.11.126. [CrossRef] [Google Scholar]
Кват Инт. 2017; 433:156–162. doi: 10.1016/j.quaint.2015.11.126. [CrossRef] [Google Scholar]
63. Martinón-Torres M, Wu X, de Castro JMB, Xing S, Liu W. Homo sapiens в позднем плейстоцене Восточной Азии. Карр Антропол. 2017; 58 (Приложение 17): 434–448. дои: 10.1086/694449. [Перекрестная ссылка] [Академия Google]
64. Прат С. Первые поселения гоминидов за пределами Африки. Темп и режим разгона: обзор и перспективы. CR Палевол. 2017; 127:1–11. [Google Scholar]
65. Sun XF, Yi S, Lu H, Zhang W. Датировка TT-OSL и post-IR IRSL стоянки человека Дали в Центральном Китае. Кват Инт. 2017; 434:99–106. doi: 10.1016/j.quaint.2015.05.027. [CrossRef] [Google Scholar]
66. Duval M, Grün R, Parés JM, Martín-Francés L, Campana I, Rosell J, et al. Первая прямая ЭПР-датировка зуба гоминина из Атапуэрка-Гран-Долина TD-6 (Испания) подтверждает древность Homo antecessor . Кват Геохронол. 2018;47:120–137. doi: 10.1016/j.quageo.2018.05.001. [CrossRef] [Google Scholar]
67. Xing S, Martinón-Torres M, de Castro JMB. Ископаемые зубы пекинского человека. 2018. Научные отчеты, характер с41598–018-20432. [Бесплатная статья PMC] [PubMed]
Xing S, Martinón-Torres M, de Castro JMB. Ископаемые зубы пекинского человека. 2018. Научные отчеты, характер с41598–018-20432. [Бесплатная статья PMC] [PubMed]
68. Zhu Z, Dennell R, Huang W, Wu Y, Qiu S, Yang S, et al. Гоминиды заселили китайское лёссовое плато примерно 2,1 миллиона лет назад. Природа. 2018; 559: 608–612. doi: 10.1038/s41586-018-0299-4. [PubMed] [CrossRef] [Google Scholar]
69. Han F, Shao Q, Bahain JJ, Sun X, Yin G. Совмещенное ESR и U-серия датирования ископаемых зубов из стоянки гоминидов Июань, северный Китай. Кват Геохрон. 2019;49:291–296. doi: 10.1016/j.quageo.2018.02.004. [CrossRef] [Google Scholar]
70. Wu XJ, Pei SW, Cai YJ, Tong HW, Li Q, Dong Z и др. Архаичные человеческие останки из Хуалонгдонга, Китай, и преемственность и вариации человека среднего плейстоцена. Proc Natl Acad Sci U S A. 2019;116:9820–9824. doi: 10.1073/pnas.16116. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
71. de Lumley MA, Guipert G, de Lumley H, Protopapa N, Pitsios T. Apidima 1 и Apidima 2: Два антеандертальских черепа на Пелопоннесе , Греция. Антропология. 2020;124:102743. doi: 10.1016/j.anthro.2019.102743. [CrossRef] [Google Scholar]
Apidima 1 и Apidima 2: Два антеандертальских черепа на Пелопоннесе , Греция. Антропология. 2020;124:102743. doi: 10.1016/j.anthro.2019.102743. [CrossRef] [Google Scholar]
72. Yang S-X, Deng C-L, Zhu R-X, Petraglia MD. Палеолит в бассейне Нихэван, Китай: эволюционная история записи от раннего до позднего плейстоцена в Восточной Азии. Эвол Антропол. 2020;29: 125–142. doi: 10.1002/evan.21813. [PubMed] [CrossRef] [Google Scholar]
73. Welker F, Ramos-Madrigal J, Gutenbrunner P, Mackie M, Tiwary S, Jersie-Christensen RR, et al. Зубной протеом Homo antecessor . Природа. 2020; 580: 235–238. doi: 10.1038/s41586-020-2153-8. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
74. Arnason U, Xu X, Gullberg A. Сравнение полной последовательности митохондриальной ДНК Homo и обыкновенного шимпанзе на основе нехимерных последовательностей. Дж Мол Эвол. 1996;42:145–152. doi: 10.1007/BF02198840. [PubMed] [CrossRef] [Google Scholar]
75. Green RE, Malaspinas A-S, Krause J, Briggs AW, Johnson PLF, Uhler C, et al. Полная последовательность митохондриального генома неандертальца, определенная с помощью высокопроизводительного секвенирования. Клетка. 2008; 134:416–426. doi: 10.1016/j.cell.2008.06.021. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Полная последовательность митохондриального генома неандертальца, определенная с помощью высокопроизводительного секвенирования. Клетка. 2008; 134:416–426. doi: 10.1016/j.cell.2008.06.021. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
76. Krause J, Fu Q, Good JM, Viola B, Shunkov MV, Деревянко AP, et al. Полный митохондриальный ДНК-геном неизвестного гоминина из Южной Сибири. Природа. 2010;464:894–897. doi: 10.1038/nature08976. [PubMed] [CrossRef] [Google Scholar]
77. Маласпинас А.С., Westaway MC, Muller C, Sousa VC, Lao O, Alves I, et al. Геномная история аборигенов Австралии. Природа. 2016; 538: 207–214. doi: 10.1038/nature18299. [PubMed] [CrossRef] [Google Scholar]
78. Jacobs GS, Hudjashow G, Saag L, Kusuma P, Darusallam CC, Lawson DJ, et al. Множественные глубоко расходящиеся денисовские предки папуасов. Клетка. 2019;177:1010–1021. doi: 10.1016/j.cell.2019.02.035. [PubMed] [CrossRef] [Академия Google]
79. Райх Д., Паттерсон Н., Кирхер М. , Дельфин Ф., Нандинени М.Р., Пугач И., Ко А.М., Ко Ю.С., Джинам Т.А., Фиппс М.Э., и соавт. Примесь Денисовой и первые современные расселения людей в Юго-Восточную Азию и Океанию. Am J Hum Genet. 2011; 89: 516–528. doi: 10.1016/j.ajhg.2011.09.005. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
, Дельфин Ф., Нандинени М.Р., Пугач И., Ко А.М., Ко Ю.С., Джинам Т.А., Фиппс М.Э., и соавт. Примесь Денисовой и первые современные расселения людей в Юго-Восточную Азию и Океанию. Am J Hum Genet. 2011; 89: 516–528. doi: 10.1016/j.ajhg.2011.09.005. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
80. Browning SR, Browning BL, Zhou Y, Tucci S, Akey JM. Анализ данных секвенирования человека выявил два импульса архаичной денисовской примеси. Клетка. 2018;173:53–61. doi: 10.1016/j.cell.2018.02.031. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
81. Арнасон У., Адегоке Дж. А., Галлберг А., Харли Э. Х., Янке А., Куллберг М. Митогеномные отношения плацентарных млекопитающих и молекулярные оценки их расхождений. Ген. 2008; 585:9–12. doi: 10.1016/j.gene.2016.03.018. [PubMed] [CrossRef] [Google Scholar]
82. Anderson S, Bankier AT, Barrell BG, de Bruin MHL, Coulson AR, Drouin J, et al. Последовательность и организация митохондриального генома человека.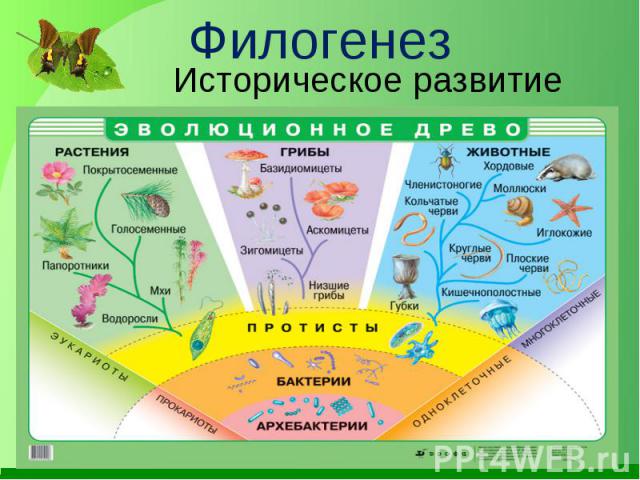 Природа. 1981; 290:457–465. doi: 10.1038/2
Природа. 1981; 290:457–465. doi: 10.1038/2
83. McBrearty S, Brooks AS. Революция, которой не было: новая интерпретация происхождения современного человеческого поведения. Джей Хам Эвол. 2000; 39: 453–563. doi: 10.1006/jhev.2000.0435. [PubMed] [CrossRef] [Google Scholar]
84. Berger LR, Hawks J, Dirks PHGM, Elliott M, Roberts EM. Homonaledi и плейстоценовая эволюция гоминидов в субэкваториальной Африке. электронная жизнь. 2017;6:e24234. doi: 10.7554/eLife.24234. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
85. Грюн Р. Прямое датирование окаменелостей человека. Годб Физ Антропол. 2006;49: 2–48. doi: 10.1002/ajpa.20516. [PubMed] [CrossRef] [Google Scholar]
86. Guo Y, Sun C, Luo L, Yang L, Han F, Tu H. 26 Al/ 10 Датировка захоронения ископаемого гоминина среднего плейстоцена Yiyuan место, провинция Шаньдун, Северный Китай. Национальная научная организация, 2019; 9:6961. doi: 10.1038/s41598-019-43401-5. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
doi: 10.1038/s41598-019-43401-5. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
87. Ferring R, Oms O, Agustí J, Berna F, Nioradze M, Shelia T, et al. Самые ранние поселения людей в Дманиси (Грузинский Кавказ) датируются 1,85–1,78 гг. Proc Natl Acad Sci USA. 2011;108:10432–10436. doi: 10.1073/pnas.1106638108. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
88. Лордкипанидзе Д., де Леон МСП, Маргвелашвили А., Рак Ю., Райтмайр Г.П., Векуа А., Золликофер П.Е. Полный череп из Дманиси, Джорджия, и эволюционная биология раннего Homo . Наука. 2013; 342:326–331. doi: 10.1126/science.1238484. [PubMed] [CrossRef] [Google Scholar]
89. Larick RR, Ciochon RL, Zaim Y, Sudijono, Suminto, Rizal Y, Aziz F, et al. Ранний плейстоцен 40 Ar/ 39 Ar возраст гомининов формации Бапанг, центральная Ява, Индонезия. Proc Natl Acad Sci. 2001;98:4866–4871. doi: 10.1073/pnas.081077298. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
90. Zaim Y, Ciochon RL, Polanski JM, Grine FE, Bettis EA, III, Rizal Y, et al. Новый Homo erectus возрастом 1,5 миллиона лет из Сангриана (Центральная Ява, Индонезия) J Hum Evol. 2011;61:363–376. doi: 10.1016/j.jhevol.2011.04.009. [PubMed] [CrossRef] [Google Scholar]
Zaim Y, Ciochon RL, Polanski JM, Grine FE, Bettis EA, III, Rizal Y, et al. Новый Homo erectus возрастом 1,5 миллиона лет из Сангриана (Центральная Ява, Индонезия) J Hum Evol. 2011;61:363–376. doi: 10.1016/j.jhevol.2011.04.009. [PubMed] [CrossRef] [Google Scholar]
91. Matsu’ura S, Kondo M, Danhara T, Sakata S, Iwano H, Hirata T, et al. Возрастной контроль данных о первом появлении яванского языка Homo erectus в районе Сангрии. Наука. 2020; 367: 210–214. [PubMed] [Google Scholar]
92. Hawks J, Elliott M, Schmid P, Churchill SE, de Ruiter DJ, Roberts EM, et al. Новые ископаемые останки Homo naledi из камеры Леседи, Южная Африка. электронная жизнь. 2017;6:24232. doi: 10.7554/eLife.24232. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
93. Dirks PHG, Roberts EM, Hilbert-Wolf H, Kramers JD, Hawks J, Dosseto A, et al. Возраст Homo naledi и связанные с ним отложения в пещере Восходящей звезды, Южная Африка. электронная жизнь. 2017;6:e24231. doi: 10.7554/eLife.24231. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
2017;6:e24231. doi: 10.7554/eLife.24231. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
94. Richter D, Grün R, Johannes-Boyau R, Steele TE, Amani F, Fernandes P, et al. Возраст окаменелостей гоминидов из Джебель-Ирхуд, Марокко, и происхождение среднего каменного века. Природа. 2017; 546: 293–296. doi: 10.1038/nature22335. [PubMed] [CrossRef] [Google Scholar]
95. Hublin JJ, Ben-Ncer A, Bailey SE, Freidline SE, Neubauer S, Skinner MM, et al. Новые окаменелости из Джебель-Ирхуда, Марокко, и панафриканское происхождение Человек разумный . Природа. 2017; 546: 289–292. doi: 10.1038/nature22336. [PubMed] [CrossRef] [Google Scholar]
96. Smith TM, Tafforeau P, Reid DJ, Grün R, Eggins S, Boutakiout M, Hublin JJ. Самые ранние свидетельства современной истории человеческой жизни в Северной Африке в начале г. Homo sapiens . Proc Natl Acad Sci U S A. 2007; 104:6128–6133. doi: 10.1073/pnas.0700747104. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
97. Harvati K, Röding C, Bosman AM, Karakostis FA, Grün R, Stringer C, et al. Окаменелости пещеры Апидима являются самыми ранними свидетельствами существования Homo sapiens в Евразии. Природа. 2019. 10.1038/s41586. [PubMed]
Harvati K, Röding C, Bosman AM, Karakostis FA, Grün R, Stringer C, et al. Окаменелости пещеры Апидима являются самыми ранними свидетельствами существования Homo sapiens в Евразии. Природа. 2019. 10.1038/s41586. [PubMed]
98. Росас А., Бастир М. Оценка затылочной кости позднего среднего плейстоцена из черепа Apidima 1 (Греция) L’anthropologie. 2020;124:10275. [Google Scholar]
99. Голуэй-Уитэм Дж., Стрингер К. Как эволюционировал Homo sapiens ? Наука. 2018; 360:1296–1298. doi: 10.1126/science.aat6659. [PubMed] [CrossRef] [Google Scholar]
100. Bae CJ, Douka K, Petraglia MD. О происхождении современного человека: взгляды Азии. Наука. 2018;358:eaai9067. doi: 10.1126/science.aai9067. [PubMed] [CrossRef] [Google Scholar]
101. Petit JR, Jouzel J, Raynaud D, Barkov NI, Barnola J-M, Basile I, et al. История климата и атмосферы за последние 420 000 лет из ледяного керна Восток, Антарктида. Природа. 1999; 399:429–436. дои: 10.1038/20859. [CrossRef] [Google Scholar]
102. Parrenin F, Barnola J-M, Beer J, Blunier T, Castellano E, Chappellaz, et al. Хронология EDC3 для ледяного керна EPICA Dome. Клим Прошлое. 2007; 3: 485–497. дои: 10.5194/ср-3-485-2007. [CrossRef] [Google Scholar]
Parrenin F, Barnola J-M, Beer J, Blunier T, Castellano E, Chappellaz, et al. Хронология EDC3 для ледяного керна EPICA Dome. Клим Прошлое. 2007; 3: 485–497. дои: 10.5194/ср-3-485-2007. [CrossRef] [Google Scholar]
103. Ланг Н., Вольф Э.В. Межледниковая и ледниковая изменчивость за последние 800 тыс. лет в морских, ледовых и наземных архивах. Клим Прошлое. 2011;7:361–380. doi: 10.5194/cp-7-361-2011. [CrossRef] [Google Scholar]
104. Ингман М., Кессманн Х., Паабо С., Гилленстен У. Изменчивость митохондриального генома и происхождение современного человека. Природа. 2000;408:708–713. doi: 10.1038/35047064. [PubMed] [CrossRef] [Google Scholar]
105. Hammer MF, Woerner AE, Mendez FL, Watkins JC, Wall JD. Генетические доказательства архаичной примеси в Африке. Proc Natl Acad Sci U S A. 2011;108:15123–15128. doi: 10.1073/pnas.1109300108. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
106. Lachance J, Vernot B, Elbers CC, Ferwerda B, Froment A, Bodo J-M, et al. Эволюционная история и адаптация на основе полных геномных последовательностей различных африканских охотников-собирателей. Клетка. 2012; 150:457–469. doi: 10.1016/j.cell.2012.07.009. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Эволюционная история и адаптация на основе полных геномных последовательностей различных африканских охотников-собирателей. Клетка. 2012; 150:457–469. doi: 10.1016/j.cell.2012.07.009. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
107. Hsieh PH, Woerner AE, Wall JD, Lachance J, Tishkoff SA, Gutekunst RN, Hammer MF. Анализ полногеномных данных на основе моделей раскрывает сложную эволюционную историю, включающую архаичную интрогрессию у центральноафриканских пигмеев. Геном Res. 2016;26:291–300. doi: 10.1101/gr.196634.115. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
108. Krause J, Orlando L, Serre D, Bence V, Prüfer K, Richards MP, et al. Неандертальцы в Средней Азии и Сибири. Природа. 2007; 449: 902–904. doi: 10.1038/nature06193. [PubMed] [CrossRef] [Google Scholar]
109. Reich D, Green RE, Kircher M, Krause J, Patterson N, Durand EY, Viola B, Briggs AW, Stenzel U, Johnson PLF, et al. Генетическая история архаичной группы гоминидов из Денисовой пещеры в Сибири.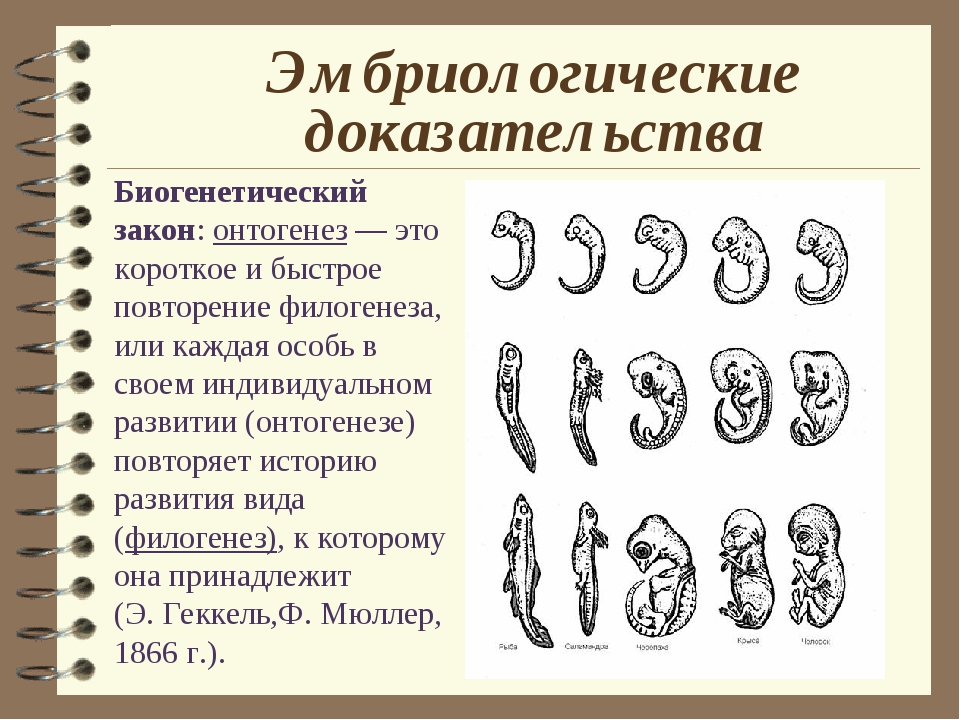 Природа. 2010; 468:1053–1060. дои: 10.1038/nature09710. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Природа. 2010; 468:1053–1060. дои: 10.1038/nature09710. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
110. Санкарараман С., Маллик С., Паттерсон Н., Рейх Д. Комбинированный ландшафт денисовского и неандертальского происхождения у современных людей. Карр Биол. 2016;26:1241–1247. doi: 10.1016/j.cub.2016.03.037. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
111. Vernot B, Tucci S, Kelso J, Schraiber JG, Wolf AB, Gittelman RM, Dannemann M, Grote S, McCoy RC, Norton H, et др. Раскопки ДНК неандертальцев и денисовцев из геномов меланезийцев. Наука. 2016; 352: 235–239. doi: 10.1126/science.aad9416. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
112. Тио Дж. Х., Леван А. Число хромосом человека. наследственность. 1956; 42: 1–6. doi: 10.1111/j.1601-5223.1956.tb03010.x. [CrossRef] [Google Scholar]
113. Арнасон А. 50 лет спустя – рассмотрение некоторых обстоятельств установления правильного числа хромосом у человека. наследственность. 2006; 143: 202–211. doi: 10.1111/j.2006.0018-0661.01963.x. [PubMed] [CrossRef] [Академия Google]
2006; 143: 202–211. doi: 10.1111/j.2006.0018-0661.01963.x. [PubMed] [CrossRef] [Академия Google]
114. Darlington CD, Haque A. Хромосомы обезьян и людей. Природа. 1955;175:32. [PubMed]
Филогенез от обезьяны к человеку | Происхождение разума: психологические взгляды на эволюцию гоминидов
Фильтр поиска панели навигации Oxford AcademicПроисхождение разума: психологические взгляды на эволюцию гоминидовКогнитивная психологияКнигиЖурналы Термин поиска мобильного микросайта
Закрыть
Фильтр поиска панели навигации Oxford AcademicПроисхождение разума: психологические взгляды на эволюцию гоминидовКогнитивная психологияКнигиЖурналы Термин поиска на микросайте
Расширенный поиск
-
Иконка Цитировать Цитировать
-
Разрешения
-
Делиться
- Твиттер
- Подробнее
Cite
Корбаллис, Майкл С. , 9 лет0003
, 9 лет0003
‘Филогения от обезьян до человека
,
в Майкл Корбаллис, и Стивен Э. Г. Ли (ред.)
,
. Сестра ;
Online Edn,
Oxford Academic
, 22 марта 2012 г.
), https://doi.org/10.1093/acprof:oso/9780192632593.003.0003,
. Выберите формат Выберите format.ris (Mendeley, Papers, Zotero).enw (EndNote).bibtex (BibTex).txt (Medlars, RefWorks)
Закрыть
Фильтр поиска панели навигации Oxford AcademicПроисхождение разума: психологические взгляды на эволюцию гоминидовКогнитивная психологияКнигиЖурналы Термин поиска мобильного микросайта
Закрыть
Фильтр поиска панели навигации Oxford AcademicПроисхождение разума: психологические взгляды на эволюцию гоминидовКогнитивная психологияКнигиЖурналы Термин поиска на микросайте
Advanced Search
Abstract
Сложности эволюции гоминидов поднимают много вопросов о том, когда именно могли возникнуть какие-либо психические свойства, которые могут быть присущи только человеку. В этой главе рассматриваются этапы эволюции от обезьян к обезьянам, от человекообразных обезьян к гоминидам и от гоминидов к людям. Каждая точка ветвления в этой эволюционной прогрессии является потенциальной точкой ментального разрыва. При описании различных видов в главе основное внимание уделяется физическим характеристикам. Во многих отношениях можно сказать, что эволюция характерных черт человека началась с появлением рода 9.0071 Человек . Около полумиллиона лет назад гоминиды стали существами с большим телом, большим мозгом и, возможно, с более сложной производственной технологией, чем можно предположить по каменным артефактам и довольно сложному языку. Кроме того, в главе обсуждается появление современного человека, известного как Homo sapiens .
В этой главе рассматриваются этапы эволюции от обезьян к обезьянам, от человекообразных обезьян к гоминидам и от гоминидов к людям. Каждая точка ветвления в этой эволюционной прогрессии является потенциальной точкой ментального разрыва. При описании различных видов в главе основное внимание уделяется физическим характеристикам. Во многих отношениях можно сказать, что эволюция характерных черт человека началась с появлением рода 9.0071 Человек . Около полумиллиона лет назад гоминиды стали существами с большим телом, большим мозгом и, возможно, с более сложной производственной технологией, чем можно предположить по каменным артефактам и довольно сложному языку. Кроме того, в главе обсуждается появление современного человека, известного как Homo sapiens .
Ключевые слова: эволюция, обезьяны, человекообразные обезьяны, гоминиды, люди, психическая прерывистость, физические характеристики, Homo, язык, Homo sapiens
Предмет
Когнитивная психология
В настоящее время у вас нет доступа к этой главе.
Войти
Получить помощь с доступом
Получить помощь с доступом
Доступ для учреждений
Доступ к контенту в Oxford Academic часто предоставляется посредством институциональных подписок и покупок. Если вы являетесь членом учреждения с активной учетной записью, вы можете получить доступ к контенту одним из следующих способов:
Доступ на основе IP
Как правило, доступ предоставляется через институциональную сеть к диапазону IP-адресов. Эта аутентификация происходит автоматически, и невозможно выйти из учетной записи с IP-аутентификацией.
Войдите через свое учреждение
Выберите этот вариант, чтобы получить удаленный доступ за пределами вашего учреждения. Технология Shibboleth/Open Athens используется для обеспечения единого входа между веб-сайтом вашего учебного заведения и Oxford Academic.
- Нажмите Войти через свое учреждение.
- Выберите свое учреждение из предоставленного списка, после чего вы перейдете на веб-сайт вашего учреждения для входа.
- Находясь на сайте учреждения, используйте учетные данные, предоставленные вашим учреждением. Не используйте личную учетную запись Oxford Academic.
- После успешного входа вы вернетесь в Oxford Academic.
Если вашего учреждения нет в списке или вы не можете войти на веб-сайт своего учреждения, обратитесь к своему библиотекарю или администратору.
Войти с помощью читательского билета
Введите номер своего читательского билета, чтобы войти в систему. Если вы не можете войти в систему, обратитесь к своему библиотекарю.
Члены общества
Доступ члена общества к журналу достигается одним из следующих способов:
Войти через сайт сообщества
Многие общества предлагают единый вход между веб-сайтом общества и Oxford Academic. Если вы видите «Войти через сайт сообщества» на панели входа в журнале:
Если вы видите «Войти через сайт сообщества» на панели входа в журнале:
- Щелкните Войти через сайт сообщества.
- При посещении сайта общества используйте учетные данные, предоставленные этим обществом. Не используйте личную учетную запись Oxford Academic.
- После успешного входа вы вернетесь в Oxford Academic.
Если у вас нет учетной записи сообщества или вы забыли свое имя пользователя или пароль, обратитесь в свое общество.
Войти с помощью личного кабинета
Некоторые общества используют личные аккаунты Oxford Academic для предоставления доступа своим членам. Смотри ниже.
Личный кабинет
Личную учетную запись можно использовать для получения оповещений по электронной почте, сохранения результатов поиска, покупки контента и активации подписок.
Некоторые общества используют личные аккаунты Oxford Academic для предоставления доступа своим членам.
Просмотр ваших зарегистрированных учетных записей
Щелкните значок учетной записи в правом верхнем углу, чтобы:
- Просмотр вашей личной учетной записи и доступ к функциям управления учетной записью.
- Просмотр институциональных учетных записей, предоставляющих доступ.
Выполнен вход, но нет доступа к содержимому
Oxford Academic предлагает широкий ассортимент продукции. Подписка учреждения может не распространяться на контент, к которому вы пытаетесь получить доступ. Если вы считаете, что у вас должен быть доступ к этому контенту, обратитесь к своему библиотекарю.
Ведение счетов организаций
Для библиотекарей и администраторов ваша личная учетная запись также предоставляет доступ к управлению институциональной учетной записью. Здесь вы найдете параметры для просмотра и активации подписок, управления институциональными настройками и параметрами доступа, доступа к статистике использования и т. д.
д.
Покупка
Наши книги можно приобрести по подписке или приобрести в библиотеках и учреждениях.
Информация о покупке
ПРАЙМ PubMed | Инверсия филогении человека: Homo покинул Африку как прямоходящий, вернулся как сапиенс сапиенс несмотря на раннее беспокойство по этому поводу среди филогенетиков с опытом, превышающим опыт Homo. Сомнительна и палеонтологическая поддержка гипотезы, обстоятельство, которое в свете расширения евразийских палеонтологических знаний стало акцентироваться в последние десятилетия.
РЕЗУЛЬТАТЫ
Направление эволюции в филогенетическом древе современного человека (Homo sapiens sapiens, Hss) было установлено, среди прочего, путем применения прогрессивного филогенетического анализа к образцам мтДНК, включавшим евразийских, лундских и африканских мбути, сан и Йоруба. Экспертиза определила африканские популяции как парафилетические, что поставило под угрозу OOAH. Этот вывод, который согласовывался с гипотезой вне Евразии, OOEH, был подтвержден интрогрессией мтДНК от Hss к Hsnn (неандертальцам), которая продемонстрировала временное и физическое сосуществование двух линий в Евразии. Результаты согласуются с палеонтологически установленным присутствием H. erectus в Евразии, евразийским расхождением между H. sapiens и H. antecessor ≈ 850 000 лет назад, расхождением Hs между Hss и Hsn (неандертальцы + денисовцы) ≈ 800 000 лет назад, интрогрессией мтДНК. из Hss в Hsnn * ≈ 500 000 лет назад и евразийское расхождение среди предков существующих Hss ≈ 250 000 лет назад при исходе мбути / сан в Африку.
Этот вывод, который согласовывался с гипотезой вне Евразии, OOEH, был подтвержден интрогрессией мтДНК от Hss к Hsnn (неандертальцам), которая продемонстрировала временное и физическое сосуществование двух линий в Евразии. Результаты согласуются с палеонтологически установленным присутствием H. erectus в Евразии, евразийским расхождением между H. sapiens и H. antecessor ≈ 850 000 лет назад, расхождением Hs между Hss и Hsn (неандертальцы + денисовцы) ≈ 800 000 лет назад, интрогрессией мтДНК. из Hss в Hsnn * ≈ 500 000 лет назад и евразийское расхождение среди предков существующих Hss ≈ 250 000 лет назад при исходе мбути / сан в Африку.
ВЫВОДЫ
Настоящее исследование показало, что Евразия была не приемником, а донором в эволюции Hss. Открытие того, что Homo покинул Африку как прямоходящий и вернулся как sapiens sapiens, представляет собой изменение в понимании эволюции Hs в сторону того, которое соответствует обширным евразийским записям палеонтологии и археологии Hs.
Authors+Show Affiliations
Árnason Ú
Кафедра нейрохирургии, Медицинский факультет, Лундский университет, Лунд, Швеция. [email protected].
[email protected].
Hallström B
Центр трансляционной геномики, медицинский факультет Лундского университета, Лунд, Швеция.
MeSH
AfricaAnimalsBiological EvolutionCytogenetic AnalysisDNA, MitochondrialEnvironmentEvolution, MolecularGene-Environment InteractionGenetics, PopulationHominidaeHuman GeneticsHumansPhylogeny
Pub Type(s)
Journal Article
Review
Language
eng
PubMed ID
33341120
Цитирование
Арнасон, Эльфур и Бьорн Халльстрем. «Поворот человеческой филогении: Homo покинул Африку как Erectus, вернулся как Sapiens Sapiens». Hereditas, vol. 157, нет. 1, 2020, с. 51.
Арнасон У., Халльстрём Б. Инверсия филогении человека: Homo покинул Африку как прямоходящий, вернулся как sapiens sapiens. Наследие . 2020;157(1):51.
Арнасон, Ю., и Халльстрем, Б. (2020). Инверсия филогении человека: Homo покинул Африку эректусом, вернулся как sapiens sapiens. Hereditas , 157 (1), 51. https://doi.org/10.1186/s41065-020-00163-9
Hereditas , 157 (1), 51. https://doi.org/10.1186/s41065-020-00163-9
Арнасон У, Халльстрем Б. Инверсия филогении человека: Homo покинул Африку как Erectus, Вернулся как Sapiens Sapiens. Наследственность. 19 декабря 2020 г .; 157 (1): 51. PubMed PMID: 33341120.
* Названия статей в формате цитирования AMA должны быть в регистре предложений
MLAAMAAPAVANCOUVER
TY — JOUR
T1 — Инверсия филогении человека: Homo покинул Африку как прямоходящий, вернулся как sapiens sapiens.
AU — Арнасон, Эльфур,
AU — Hallström, Björn,
Y1 — 2020/12/19/
PY — 30.10.2020/получено
PY — 27.11.2020/принято
PY — 20.12.2020/антрез
PY — 21.12.2020/опубликовано
PY — 31.08.2021/medline
KW — Эволюция человека
KW — Молекулярная филогенетика
кВт — ОООАХ
кВт — ОООЭХ
KW — Гипотеза выхода из Африки
KW — Гипотеза вне Евразии
кВт — ППА
KW — Палеонтология
KW — Прогрессивный филогенетический анализ
KW — мтДНК
KW — нуДНК
СП — 51
ЭП — 51
JF — Наследие
Джо — Наследница
ВЛ — 157
ИС — 1
N2 — ПРЕДПОСЫЛКИ: Гипотеза молекулярного происхождения из Африки, OOAH, считалась установленным фактом среди популяционных генетиков в течение примерно 25-30 лет, несмотря на раннее беспокойство по этому поводу среди филогенетиков, имеющих опыт не только Homo. Сомнительна и палеонтологическая поддержка гипотезы, обстоятельство, которое в свете расширения евразийских палеонтологических знаний стало акцентироваться в последние десятилетия. РЕЗУЛЬТАТЫ: Направление эволюции в филогенетическом древе современного человека (Homo sapiens sapiens, Hss) было установлено, среди прочего, путем применения прогрессивного филогенетического анализа к выборке мтДНК, которая включала евразийцев, лундов и африканцев мбути, сан и йоруба. Экспертиза определила африканские популяции как парафилетические, что поставило под угрозу OOAH. Этот вывод, который согласовывался с гипотезой вне Евразии, OOEH, был подтвержден интрогрессией мтДНК от Hss к Hsnn (неандертальцам), которая продемонстрировала временное и физическое сосуществование двух линий в Евразии. Результаты согласуются с палеонтологически установленным присутствием H. erectus в Евразии, евразийским расхождением между H. sapiens и H. antecessor ≈ 850 000 лет назад, расхождением Hs между Hss и Hsn (неандертальцы + денисовцы) ≈ 800 000 лет назад, интрогрессией мтДНК.
Сомнительна и палеонтологическая поддержка гипотезы, обстоятельство, которое в свете расширения евразийских палеонтологических знаний стало акцентироваться в последние десятилетия. РЕЗУЛЬТАТЫ: Направление эволюции в филогенетическом древе современного человека (Homo sapiens sapiens, Hss) было установлено, среди прочего, путем применения прогрессивного филогенетического анализа к выборке мтДНК, которая включала евразийцев, лундов и африканцев мбути, сан и йоруба. Экспертиза определила африканские популяции как парафилетические, что поставило под угрозу OOAH. Этот вывод, который согласовывался с гипотезой вне Евразии, OOEH, был подтвержден интрогрессией мтДНК от Hss к Hsnn (неандертальцам), которая продемонстрировала временное и физическое сосуществование двух линий в Евразии. Результаты согласуются с палеонтологически установленным присутствием H. erectus в Евразии, евразийским расхождением между H. sapiens и H. antecessor ≈ 850 000 лет назад, расхождением Hs между Hss и Hsn (неандертальцы + денисовцы) ≈ 800 000 лет назад, интрогрессией мтДНК. из Hss в Hsnn * ≈ 500 000 лет назад и евразийское расхождение среди предков существующих Hss ≈ 250 000 лет назад при исходе мбути / сан в Африку. ВЫВОДЫ: Настоящее исследование показало, что Евразия была не приемником, а донором в эволюции Hss. Открытие того, что Homo покинул Африку как прямоходящий и вернулся как sapiens sapiens, представляет собой изменение в понимании эволюции Hs в сторону того, которое соответствует обширным евразийским записям палеонтологии и археологии Hs.
СН — 1601-5223
UR — https://brain.unboundmedicine.com/medline/citation/33341120/the_reversal_of_human_phylogeny:_homo_left_africa_as_erectus_came_back_as_sapiens_sapiens_
ДБ — ПРАЙМ
ДП — Свободная медицина
ЭР —
из Hss в Hsnn * ≈ 500 000 лет назад и евразийское расхождение среди предков существующих Hss ≈ 250 000 лет назад при исходе мбути / сан в Африку. ВЫВОДЫ: Настоящее исследование показало, что Евразия была не приемником, а донором в эволюции Hss. Открытие того, что Homo покинул Африку как прямоходящий и вернулся как sapiens sapiens, представляет собой изменение в понимании эволюции Hs в сторону того, которое соответствует обширным евразийским записям палеонтологии и археологии Hs.
СН — 1601-5223
UR — https://brain.unboundmedicine.com/medline/citation/33341120/the_reversal_of_human_phylogeny:_homo_left_africa_as_erectus_came_back_as_sapiens_sapiens_
ДБ — ПРАЙМ
ДП — Свободная медицина
ЭР —
Эволюция человека — историческая геология
Кловис указывает из тайника Раммелс-Маске, Айова, СШАИзображение: Wikipedia, Billwhittaker, CC BY-SA 3.0 учащиеся должны уметь:
- Обсуждать Homo sapiens sapiens в более широком контексте эволюции гоминидов;
- Выделите некоторые критические разногласия в филогенетическом дереве человека;
- Обсудите некоторые особенности анатомии черепа гоминидов, которые можно использовать для сравнительного анализа генотипов между группами гоминидов;
- Опишите, как люди мигрировали из Африки, включая время и пути миграции;
- Опишите, как мы можем проследить эти миграции, используя отцовские и материнские гаплогруппы;
- Определите некоторые ключевые технологические инновации в ходе эволюции гоминидов, особенно среди анатомически современных людей и неандертальцев;
- Поймите, насколько эволюция человека очень похожа на эволюцию в других таксономических семействах организмов.

Эволюция людей началась 13,8 миллиардов лет назад с самого Большого Взрыва, который привел в движение силы и энергию, которые сделали нашу историю возможной (хотя и не неизбежной!). Созданные элементы и соединения, образовавшиеся после этого события, в конечном итоге сформируют нашу планету Земля. В какой-то момент спустя 4 миллиарда лет назад сформировались живые самовоспроизводящиеся материалы, которые станут нашими первыми предками. Чтобы это произошло, эти ранние формы жизни должны были использовать ресурсы из окружающей среды, чтобы выжить и процветать.
Так и с людьми сегодня. Вся жизнь извлекает энергию и питательные вещества из окружающей среды для дыхания, пищеварения и размножения. Наш вид и некоторые из наших предков подняли эту добычу на новый уровень. Они разработали инструменты. Эти инструменты сделали добычу ресурсов намного более эффективной, что привело к новым эволюционным давлениям, большему количеству приспособлений и, в конечном счете, к нам с вами.
В этом тематическом исследовании рассматривается развитие нашего вида. История длинная, так что это просто краткое изложение. В конечном счете, он служит для выделения нескольких аспектов человека. Подчеркиваются две нити эволюции гоминидов. Мы исследуем эволюцию использования инструментов среди гоминидов как способ изучения развития человеческого интеллекта и постоянно растущей способности нашего вида извлекать ресурсы из окружающей среды. Мы также исследуем некоторые особые анатомические особенности черепов гоминидов, чтобы продемонстрировать физиологическое развитие гоминидов.
Вот некоторые из ключевых нитей, которые переплетаются в этой главе:
- Как и другие организмы, анатомически современные люди развивались в соответствии с эволюционной теорией.
- Эволюцию гоминидов можно проследить с помощью окаменелостей и сравнительной анатомии.
- Как и другие организмы, люди извлекают энергию и изменяют окружающую среду, чтобы выжить.
- Человеческая изобретательность в обращении с инструментами зародилась глубоко в истории гоминидов и не уникальна для анатомически современных людей.

Если вы читаете об эволюции человека и области палеоантропологии, вы также столкнетесь с другими приматами, которых вы можете считать старыми друзьями ( например, ., шимпанзе), и группой необычных приматов, которые вымерли. Вы также изучите элементы области археологии, изучения материальной культуры. Эта последняя часть довольно уникальна для исследования эволюции гоминидов, поскольку нет никаких археологических данных о других организмах. Ближайший аналог археологических материалов в остальном живом мире — это ихнология, изучение следов окаменелостей. Археология делится на два подполя. Историческая археология исследует человеческую культуру после появления письменности. Доисторическая археология исследует все остальное. Археология обычно является подполем антропологии, изучения людей.
Палеоантропология — это слияние палеонтологии и антропологии, целью которого является понимание наших человеческих предков и связанной с ними культуры. Поле стремится к целостному исследованию. Это означает, что палеоантропологов интересуют биология человека, социальная структура, культурное развитие, материальная культура (археология), языковое развитие и многое другое.
Было много довольно известных палеоантропологов. Это связано с тем, что эта область, как правило, вызывает большой интерес у людей за пределами научного сообщества. Известные имена включают многих членов семьи Лики. Луи и Мэри Лики и их сын Ричард Лики, которые вместе со своей партнершей Мив Лики внесли большой вклад в то, что мы знаем о нашем общем африканском происхождении. Их работа в таких местах, как Олдувайское ущелье, Танзания и близлежащие окрестности, предоставила важные сведения о том, как люди развивались, мигрировали и связаны с нашими двоюродными братьями-приматами. Ян Таттерсолл, куратор Американского музея естественной истории в Нью-Йорке, работал над тем, чтобы глубже понять нашу связь с нынешними кузенами-приматами, а также над стандартизацией терминологии в этой области. Третьи, такие как Соня Харманд, посвятили свои исследования изучению культурного развития человека, о чем свидетельствует ее работа с каменными орудиями.
Палеоантропология — динамичная и очень молодая область. Новые взгляды на наше человеческое прошлое публикуются почти ежемесячно. Чтобы не отставать от этих изменений, обратитесь к двум конкретным местам, указанным в разделе «Дополнительная литература» в конце: программе эволюции человека Смитсоновского института и Институту происхождения человека в Университете штата Аризона.
Терминология филогении гоминидов
Погружаясь в изучение происхождения человека, вы столкнетесь с множеством запутанных терминов. Мы начнем с описания некоторых из них здесь.
Приматы. Приматы представляют собой большую группу социальных млекопитающих, возникших между 85 и 55 млн лет назад, вероятно, из мелких млекопитающих, живущих на деревьях. Общие характеристики приматов включают относительно большой мозг, остроту зрения, цветовое зрение, ловкие руки (хватательные с противопоставленными большими пальцами для захвата) и большой диапазон движений в их плечевых суставах. Приматы являются частью отряда Primata, который состоит из двух подотрядов, стрепсиринов и гаплоринов. Люди являются членами этого последнего подотряда.
Изображение: Wikimedia, Public Domain
Hominidae — гоминида — таксономическая семья человека. В просторечии его чаще называют «великими обезьянами». Термин «гоминид» иногда также используется в общем виде. В этом семействе в настоящее время насчитывается восемь существующих (живущих) видов, относящихся к четырем родам. К таким видам относятся мы, Homo sapiens , орангутаны (Pongo abelii, Pongo tapanuliensis, Pongo pygmaeus), Гориллы (Gorilla gorilla, Gorilla beringei), и шимпанзе (Pan troglodytes, Pan paniscus).
Homininae – гоминины являются нашим подсемейством и конкретно относятся к «африканским обезьянам». Это подсемейство разделено на два «племени» (этот термин не имеет отношения к тому, который используется для коренных человеческих групп): Hominini и Gorillini.
(Изображение: Wikimedia, By EoD — собственная работа, CC BY-SA 3.0)
Hominini — гоминини — это таксономическое племя, в которое входят наши человеческие предки и шимпанзе — всего два рода. Иногда шимпанзе ( Pan sp.) выделяют в собственное племя. В конечном счете, поиски общего человеческого предка между нашими близкими родственниками шимпанзе продолжаются примерно до 4-8 млн лет назад (Patterson et al. , 2006), но раскол, вероятно, произошел около 6 млн лет назад. Как только происходит это расщепление, филогенетическое древо полностью сосредоточено на линии наших предков, все из которых теперь вымерли, кроме нас. Эти роды включают Homo и австралопитек .
Модель филогении гоминини за последние 10 миллионов лет. Розовая линия для Gorillini отделяется около 8 млн лет назад, фиолетовая линия для Pan отделяется около 6 млн лет назад, а Homo и австралопитеки продолжаются до наших дней, представленные одиноким видом, Homo sapiens. Изображение: Wikimedia, Dbachmann — собственная работа, CC BY-SA 4.0
Я понял? Кто такие гоминиды?
Терминология черепа гоминидов
Ниже будет много обсуждений, использующих развитие черепа гоминидов для обсуждения эволюционных изменений гоминидов. Конечно, есть и другие анатомические особенности, которые можно использовать, когда есть более полные скелетные останки. Черепа и фрагменты черепа, вероятно, являются наиболее распространенными окаменелостями, поэтому они используются в данном тематическом исследовании. Ниже приводится краткое изложение некоторых основных частей анатомии черепа гоминидов.
Краткий обзор особенностей черепа гоминидов, сравнение черепов человека и австралопитека.Изображение: общественное достояние
Если мы подробно рассмотрим части черепа, некоторые из основных областей станут важными в истории человеческого развития. Первым из них является нижняя челюсть, или нижняя челюсть. Ключевое различие между обезьянами и людьми в этом признаке — размер. Обезьяны едят гораздо более жесткий волокнистый материал, чем люди, для пережевывания которых требуется более крепкая нижняя челюсть. Люди также разработали подчеркнутую структуру подбородка, чтобы помочь придать дополнительную силу тому, что является гораздо более слабой областью нашей нижней челюсти.
Изображение: Public Domain Стоматологический рисунок челюстей приматов, в том числе человека.
Изображение: Public Domain
Расположение зубов у человекообразных обезьян и гоминидов очень похоже. Зубной ряд включает моляры и премоляры для жевания, резцы для резания и клыки для разрывания. Зубы также являются одним из способов, которым обезьяны, в отличие от людей, могут проявлять половой диморфизм (различия между полами). У самцов обезьян клыки, как правило, намного больше, чем у самок. У более крупных собак нет никаких реальных преимуществ, кроме полового отбора, когда самки отдают предпочтение одному самцу из-за этой особенности перед другим. Хотя люди, безусловно, замечают зубы друг друга, партнеры не выбираются на основе размера их клыков. Еще одно важное различие между зубами обезьяны и человека заключается в толщине эмали, при этом эмаль человеческих зубов намного толще.
Изображение: общественное достояние
На верхней челюсти зубная аркада предоставляет дополнительные области, где мы можем искать физиологические изменения у гоминидов. Форма зубной аркады является очевидной отправной точкой, где у людей гораздо более V-образное расположение, а у обезьян — более U-образное. Во многом это связано с укорочением и уплощением лица в результате эволюционных изменений.
Другим большим отличием здесь является диастема, или промежуток между премолярами и резцами, где находятся большие клыки. У людей нет диастемы, в отличие от предков гоминидов.
Три другие особенности черепа связаны с жеванием или движением нижней челюсти. Это скуловая дуга, сагиттальный гребень и надбровная дуга.
Сагиттальный гребень был утерян гоминидами, поскольку потребность в жевании стала менее напряженной. Поскольку эти гребни были основной областью прикрепления височных мышц, контролирующих нижнюю челюсть, они могли быть весьма выраженными у очень ранних гоминидов.
Сагиттальный гребень на черепе примата указывает на наличие очень сильных челюстных мышц.Изображение: Public DomainСкуловая дуга.
Изображение: Public Domain
Скуловая дуга у более ранних приматов была намного больше, чем у людей. Это связано с потребностью в гораздо более толстых и крепких височных мышцах. Эти мышцы прикрепляются к нижней челюсти через скуловую дугу и позади нее, а затем прикрепляются к боковой поверхности черепа. Это мышцы, которые двигают нижнюю челюсть, и чем они крупнее, тем мощнее жевательная способность организма.
Надбровная дуга.Изображение: Public Domain
Надбровные дуги — одна из последних особенностей черепа, утраченных в ходе эволюции гоминидов, и у людей они больше не проявляются. Эта структура обеспечивала прочность лицевой части черепа. Поскольку мощные височные мышцы сильно напрягали лицо, когда они натягивали череп во время пережевывания пищи. Этот гребень предохранял глазницы от разрушения при таком напряжении.
Другие функции обеспечивают поддержку изменений в передвижении, в частности, эволюцию от ходьбы на четвереньках до в основном на задних лапах до полного двуногого или прямохождения на двух ногах. Сегодня люди двуногие. Особенности черепа, которые сопровождают это постепенное изменение положения, включают затылочную область и большое затылочное отверстие.
Затылочная область, где мышцы шеи прикрепляются к задней части черепа.Изображение: общественное достояние
Затылочная область (произносится как «нью-кул») расположена на основании ближе к задней части черепа. Это место, где мышцы шеи прикрепляются к черепу. У наших предков-приматов затылочная область была намного больше. Это связано с тем, что требуются гораздо более крупные мышцы, чтобы человек смотрел вперед, когда позвоночник расположен так далеко в черепе.
Изображение: Public Domain
Большое затылочное отверстие — это отверстие в основании черепа, куда входит спинной мозг. В зависимости от вида приматов большое затылочное отверстие расположено в точке опоры или точке равновесия, и поэтому его положение очень показательно для общего передвижения. У людей он находится ближе к передней части черепа, а у человекообразных обезьян, которые в основном передвигаются на четвереньках, ближе к задней части черепа.
Последним и, возможно, самым необычным изменением в черепе гоминидов является объем черепа. До рода Человек вид. эволюционирует, емкость черепа гоминида существенно не меняется. Однако у наших самых недавних предков он, безусловно, немного увеличивается. У Homo habilis , о котором пойдет речь ниже, объем черепа составляет всего 600 см 3 . У современного человека, напротив, это 1400 см 3 . Некоторые неандертальцы даже превышали это, их черепные объемы приближались к 1700 см 3 .
Изображение: Public Domain
Большой объем черепа развился, чтобы соответствовать растущему мозгу. У кого-то может возникнуть соблазн приписать этот больший мозг просто более высокому интеллекту, но это предмет больших споров. Хотя корреляция между двумя переменными, по-видимому, существует, она не обязательно является причинно-следственной, поскольку интеллект (как бы вы ни определяли этот термин) потенциально можно отнести к другим характеристикам мозга. Кроме того, большие размеры черепа также напрямую связаны с большими размерами тела и адаптацией к более холодному климату (что, безусловно, было необходимо у неандертальцев и современных людей). Среди гоминидов также существует некоторая степень полового диморфизма. В среднем у самцов, как правило, объем черепа несколько больше, чем у самок различных видов гоминидов. Это не проблема более высокого интеллекта, а скорее объем черепа, напрямую связанный с большим средним размером тела.
Это не означает, что интеллект не влияет на объем черепа. Скорее всего, физиология мозга контролирует это. В течение десятилетий предполагалось, что рост мозга у людей связан с своего рода усиливающей петлей обратной связи между интеллектом, технологическим развитием и охотой на мясо. В этом может быть доля правды, и в контексте обсуждения Homo habilis ниже связь между мясом и ростом мозга будет исследована более конкретно. В целом, по мере того, как гоминиды эволюционировали от строго вегетарианской диеты к поеданию костного мозга и непосредственно к охоте на дичь, за этим последовали объем черепа и рост мозга. Это было подчеркнуто технологическими разработками, которые сделали сбор мусора и охоту более эффективными. Простые деревянные орудия уступили место каменным скребкам. Они уступили место сложным наконечникам стрел и копий. Со временем люди научились плавить металлы. За медью последовали бронза и железо, у каждого из которых был свой «возраст», а теперь появился кремний. Является ли все это результатом большей емкости черепа и размера мозга? Возможно, что-то из этого так, но, безусловно, есть много других переменных, которые следует учитывать.
Я понял? Физиология черепа гоминидов
Как и все семьи, человеческая семья и ее предки наполнены интересными, странными персонажами с уникальными чертами и, в данном случае, следом важных эволюционных изменений и адаптаций. На изображение ниже можно ссылаться для контекста, когда мы движемся по линии предков человека, в данном случае справа налево (обратите внимание на вертикальную шкалу времени). Изображение ниже не включает первых четырех гоминидов, обсуждаемых ниже, все из которых предшествуют 9 веку.0071 Анамский австралопитек .
Текущее предварительное расположение предков человечества. Австралопитеки помещены здесь в контекст современных людей во времени. Изображение: Public Domain
Sahelanthropus tchadensis — 6-7 млн лет назад Sahelanthropus tchadensis, Toumai — TM 266-01-060-1, череп, вид 3/4
Изображение: Smithsonian Institution
Наш самый старый предок человека, непосредственно связанный с ним, Sahelanthropus tchadensis, восходит к современной африканской нации Чад около 6-7 млн лет назад. Ископаемые данные свидетельствуют о том, что они ходили прямо. Основным доказательством этого является более выдвинутое вперед большое затылочное отверстие, отверстие для спинного мозга в основании черепа. По мере того, как это отверстие эволюционно продвигается вперед, это указывает на то, что позвоночник перемещается в положение непосредственно под черепом, как это происходит сегодня у современных людей. Sahelanthropous tchadensis известен только по девяти черепным образцам (первый из которых был обнаружен Майклом Брюнетом в 2001 г.), поэтому ископаемые данные по многим другим видам очень ограничены. Мы также можем сказать, что из-за сильного износа зубов он, вероятно, был вегетарианцем.
Фрагмент черепа Sahelanthropus tchadensis. Обратите внимание на человеческие черты в расположенном вниз и вперед большом затылочном отверстии и обезьяноподобном выступающем рту, больших зубах, узком и маленьком черепном мозгу и ярко выраженном затылочном гребне над глазами. (Модель: Смитсоновский институт, лицензия Creative Commons)
Orrorin tugenensis и Ardipithecus kadabba – 5-6 млн лет назад
Об этих двух гоминидах известно мало. Оба происходят из Восточной Африки: Кении и Эфиопии соответственно. Судя по нескольким известным фрагментам черепа и ног, у них есть две общие черты с современными людьми. Существует один образец верхней бедренной кости Orrorin tugenensis , который демонстрирует такое же наращивание кости, типичное для двуногих. Зубной ряд или зубы также важны для обоих этих гоминидов. У обоих была толстая эмаль, что характерно для современных людей. Оба были размером с современного шимпанзе и, вероятно, питались вегетарианской диетой.
Ardipithecus ramidus – 4,4 млн лет назад
Первый частичный скелет этого организма, известного как «Арди», был представлен в 2009 г., хотя Арди известен с 1994 г. Этот гораздо более полный скелет содержит много важной информации. Решающим улучшением поведения с Арди, по-видимому, является всеядная диета. На это указывает его зубная эмаль, которая была умеренно толстой. Чем толще эмаль, тем лучше организм приспособлен к поеданию растений, поскольку они требуют большого количества дробления и измельчения, чтобы начать эту первую часть пищеварительного процесса. Мясо гораздо мягче. Другие характеристики скелета указывают на то, что практиковалось как лазание по деревьям, так и прямохождение. Учитывая, что эти образцы были найдены рядом с остатками фауны, указывающими на лесную среду, кажется разумным, что Арди не был обитателем открытой саванны. Скорее, они жили достаточно близко к лесам, где их древесная адаптация была бы полезной. Это поднимает старый спор в палеоантропологии — гипотеза о лесах и саваннах для древних людей. То есть то, что называется «гипотезой саванны», утверждает, что человеческое прямохождение развилось как прямой результат перехода наших предков из лесной среды в среду саванны, где древесный образ жизни больше не был выгоден. Арди, казалось бы, предполагает обратное.
Несколько слов о прямохождении
Нет никаких сомнений в том, что современные люди преимущественно двуногие, а наши предки-австралопитеки — нет. В промежутке между ходьбой на четвереньках и на двух ногах наблюдается четкий переход. Но почему?
Часть дороги Лаэтоли, сохранившаяся в вулканическом пепле. (Изображение: Fidelis T Masao Elgidius B Ichumbaki Marco Cherin Angelo Barili Giovanni Boschian Dawid A Iurino Sofia Menconero Jacopo Moggi-Cecchi Giorgio Manzicropping/editing by Dennis Pietras – части 2 рисунков из https://elifesciences.org/content/5/e19568 под лицензией CC BY 4.0) Преимущества прямохождения очень важны. Двуногие — это критическое развитие, которое позволяет свободно использовать руки для других вещей, кроме ходьбы или даже лазания. Внезапно вынашивать молодняк легче. Изготовление инструментов становится более возможным. Это также улучшает наше передвижение на большие расстояния, облегчая преодоление гораздо больших расстояний. Со способностью путешествовать на большие расстояния тесно связана способность охлаждаться, что значительно облегчается при прямохождении, когда тело не так сгорблено. Наконец, в условиях столь резкого изменения климата в Африке, от влажных тропических лесов до саванн, двуногие становятся стратегией выживания. Теперь можно видеть над травой.
У ходьбы на двух ногах есть и недостатки. Искривление нижней части позвоночника, которое делает возможным балансирование на двух ногах, также приводит к гораздо более сильным болям в пояснице. Движение к прямохождению привело к изменению положения пальцев задних ног, так что большой палец упирается в следующие два. Это делает «отталкивание» при ходьбе гораздо более эффективным, но делает лазание по деревьям, как у наших предков, очень неуклюжим. Наконец — и, возможно, это наиболее важно из-за его роли в репродукции — прямохождение работает с узким тазом. Этот узкий таз делает прямохождение очень эффективным, но значительно затрудняет роды, учитывая, что размер родовых путей — это часть того, чем жертвуют ради способности ходить прямо. Фактически, в большинстве современных человеческих родов голова слишком велика для родовых путей. У женщин это происходит за счет высвобождения гормона релаксина, который смягчает связки таза, позволяя различным костям создавать пространство, необходимое для рождения ребенка. Этому способствует податливость черепа младенца, костные пластины которого еще не срослись. Когда все вещи не работают вместе, как они эволюционировали, возникают осложнения при родах. Современная медицина нашла способы обойти такие ситуации, в том числе с помощью таких процедур, как кесарево сечение.
Несколько слов о прямохождении для бегунов на длинные дистанции
Двуногость гоминидов принесла с собой способность бегать. Бег был не так эффективен для наших предков-приматов. Было более эффективно использовать их древесные, цепкие навыки, чтобы лазить по деревьям. Почему? Что ж, если вы спасаетесь от хищников или даже гоняетесь за добычей в лесной среде, эти навыки весьма полезны.
Но что, если климат меняется, а вместе с ним и окружающая среда, так что влажные тропические леса уступают место саванне? Как вы адаптируетесь и сохраняете способность охотиться и убегать — или просто путешествовать на большие расстояния в поисках еды? Способность бегать становится важной. И не просто способность бегать, а бегать в течение длительных периодов времени. Начиная примерно со 2 млн лет назад, физиологическая способность бегать или даже рысь становится возможной благодаря улучшениям в структуре двуногого скелета. К ним относятся развитие более длинных ног и более коротких пальцев на ногах. Кроме того, большую роль сыграли большие ягодичные мышцы, меньшая талия и большая гибкость шеи. Стопа гоминида уже также развила более крупную пяточную кость (пяточную кость) и более длинные ахилловы сухожилия. Способность бегать также потребовала изменений в наших метаболических способностях. Эти изменения напрямую повлияли на наш мозг, потому что и бег, и мозговая активность потребляют большое количество глюкозы. Изменения в развитии мозга, происходящие во время бега, включают увеличение передней части гиппокампа, изменение, которое, вероятно, привело к улучшению памяти (Щулкин, 2016). Есть еще много примеров физиологических изменений, связанных с прямохождением, из которых мы можем почерпнуть в литературе. Но здесь определенно стоит отметить, что, хотя прямохождение само по себе принесло важные преимущества, так же и выносливость в сочетании с его эволюционным развитием для гоминидов.
Интересно, что иногда эволюционные приоритеты конфликтуют. В случае эволюции гоминидов преобладала способность к прямохождению. Это, несмотря на недостатки, возникшие при родах, демонстрирует, что прямохождение должно было быть более важной стратегией выживания в то время.
Australopithecus anamensis, A.
afarenesis, A. Africanus, A. garhi и A. Sediba – 1,5–4 млн лет назад Все эти пять видов австралопитеков вымерли, но дают важные сведения о развитии человеческих черт у предков.
Australopithecus anamensis – 4,2–3,8 млн лет назад
Обнаружен у озера Туркана, Кения, а также в Среднем Аваше, Эфиопия. Ископаемые свидетельства указывают на двуногость, особенно из-за человеческого положения голеностопного сустава. Но длинные предплечья также предполагают, что они лазили по деревьям. Как и Арди, этот образец, кажется, свидетельствует о все еще тесной связи с древесным образом жизни с развившимся двуногим. Его череп также имеет важные особенности, которые связывают его с людьми, такие как предисловие, выступающее из скуловой дуги или скулы. 9Однако 0071 A. anamensis по-прежнему сохраняет важные черты черепа обезьяны, такие как узкая черепная коробка и выступающее лицо.
Ископаемые кости анаменского австралопитека, выставленные в Королевском бельгийском институте естественных наук в Брюсселе. (Изображение: Ghedoghedo — собственная работа, CC BY-SA 4.0) «Люси» —
Australopithecus afarensis — 3,85–2,95 млн лет назад Реконструкция ископаемого скелета «Люси», Australopithecus afarensis.Изображение: 120, CC BY-SA 3.0, через Wikimedia Commons
Австралопитек afarensis наиболее известен тем, что является одним из наиболее полных скелетов раннего гоминида по прозвищу «Люси». Ископаемые свидетельства этого вида насчитывают более 300 особей, поэтому он достаточно хорошо изучен. Во многих отношениях этот вид представляет собой важный ориентир в эволюции человека, место, где у нас достаточно информации, чтобы уверенно смотреть вперед и назад, когда речь идет об общих чертах нашего собственного вида.
Существует довольно большая степень полового диморфизма у A. afarensis , прежде всего в разнице в среднем росте между взрослыми самцами (151 см) и самками (105 см). Из скелетов мы знаем, что, как и современные шимпанзе, они взрослели намного быстрее, чем современные люди. С культурной точки зрения это оставило бы им гораздо меньше времени для родительского руководства и социализации в течение этого короткого детства. Ян Таттерсалл (2012) даже предположил, что они проводили некоторое время, бродя по саванне стадами, как это иногда делают сегодня бабуины, а не жили небольшими племенами, такими как шимпанзе.
Этот вид по-прежнему сохранил многие обезьяноподобные черты, такие как плоский нос, выступающая вперед нижняя челюсть и меньший размер черепной коробки. Он также сохранил длинные тонкие руки и длинные пальцы, которые хорошо приспособлены для лазания по деревьям. Человеческие характеристики связаны с меньшими зубами и телом, которое полностью стояло на двух ногах и шло прямо. Это был в целом хорошо приспособленный вид, долгоживущий вид, существующий почти миллион лет.
Реконструкция «Люси» в Варшавском музее естественной истории.Изображение: Шалом – собственная работа, CC BY-SA 3.0
Обратите внимание на некоторые особенности черепа A. afarensis. По-прежнему есть обезьяноподобный выступающий рот и небольшая черепная коробка, но затылочный гребень над глазами гораздо менее выражен, а скуловая дуга (скула) расположена более вперед, как и большое затылочное отверстие на дне черепа. Зубной ряд не показан. (Модель: Смитсоновский институт, лицензия Creative Commons)
Я понял? Бипедализм
Африканский австралопитек — от 3,3 до 2,1 млн лет назад
A. africanus очень похож на своего непосредственного предка A. afarensis . Отличительные черты включают более округлый череп с большим мозгом и меньшими зубами. Судя по их зубам, считается, что они были вегетарианцами, в отличие от некоторых из их предков. Долгое время их считали хищниками. Это произошло не из-за обнаружения каменных орудий, а из-за того, что некоторые экземпляры были обнаружены рядом со сломанными орудиями животных. С тех пор считается, что они представляют хищничество животных на A. africanus , а не наоборот.
Тем не менее, эти особи были полностью двуногими и при этом были хорошо приспособлены к древесной жизни. Образцы были обнаружены в основном в Южной Африке, что может указывать на то, что этот вид был южным родственником A. afarensis.
Вверху: черепной образец A. africanus. Есть много человеческих черт, которые все еще развиваются, как описано в приведенных выше примерах, но большая мозговая коробка здесь, вероятно, является наиболее важным развитием.
Вверху: черепной образец A. africanus. Этот образец позволяет исследовать зубной ряд. Обратите внимание на отсутствие выступающего большого клыка, что указывало бы на более обезьяноподобный зубной ряд. Представленные здесь плоские коренные зубы свидетельствуют о вегетарианской диете. (Модель: Смитсоновский институт, лицензия Creative Commons)
Australopithecus garhi и A. sediba
Из всех образцов австралопитеков эти два последних вида малоизвестны, а также недолговечны. A. garhi известен лишь по нескольким фрагментам. Но, поскольку он относится к очень плохо задокументированному периоду эволюции человека, 3-2 млн лет назад, он очень важен. Несколько существующих фрагментов указывают на значительное увеличение длины двуногого шага и, следовательно, на более эффективную походку. Еще одним важным доказательством являются каменные орудия, найденные вместе с A. garhi . Один из первых задокументированных примеров использования инструментов древними людьми, фрагменты являются веским доказательством охоты как формы добычи пищи.
A. sediba известен по очень хорошо сохранившемуся и почти полному скелету, найденному в пещере Малапа в Южной Африке. Этот вид жил в то же время, когда род Homo впервые появляется в летописи окаменелостей. В хвостовой части австралопитеков A. sediba жил рядом с некоторыми из наших самых ранних прямых предков.
Этот подробный скелет раскрывает некоторые важные события. Изменения в тазе указывают на более прямохождение, различные черты зубов и широкая грудь больше похожи на человека, особенности бедренной кости и ног в целом указывают на то, что он проводил гораздо больше времени при ходьбе и делал это очень похоже на современных людей. гораздо больше, чем другие его предки австралопитеки. Производные черты его черепа также более человечны, такие как маленькие премоляры и коренные зубы, а черты лица менее выражены и более человечны.
Череп A. sebida. Обратите внимание на более человеческие черты, такие как большая черепная коробка, меньший выступ вперед нижней челюсти и более компактный зубной ряд. 3d Модель: Creative Commons, William Mitchell
Род
Homo — 2,4 мА Род Homo , по -видимому, отделился от Australopithecines где -то между Australopithecus Africanus и 1HECINOPITHECINES. Самый ранний прямой предок рода представлен Homo habilis , вид, наиболее известный по образцам, обнаруженным в некоторых частях восточной и южной Африки между 2,4 и 1,4 млн лет назад.
Очень важно отметить, что все ранние экземпляры рода Homo обнаружены в восточной Африке. Очень быстро после эволюции Homo erectus , где-то около 1,5 млн лет назад, предки человека обнаруживаются не только в Африке, но и в других частях мира. В этот момент началась человеческая диаспора.
Модель эволюции рода Homo за последние 2 миллиона лет (вертикальная ось). Быстрая экспансия H. sapiens «из Африки» показана в верхней части диаграммы с указанием примеси неандертальцев, денисовцев и неустановленных архаичных африканских гоминидов. Фиолетовым цветом показано позднее выживание устойчивых австралопитеков (парантропов) рядом с Homo до 1,2 млн лет назад.Изображение: Conquistador, пользователь: Dbachmann — обновленная версия файла: Homo-Stammbaum, версия Stringer-en.svg, CC BY-SA 4.0
Homo habilis — 2,4–1,4 млн лет назад
Наш самый ранний предок Homo , У Homo habilis была разнообразная диета, которая, по-видимому, была всеядной. Первоначально обнаруженные семьей Лики, в частности Джонатаном Лики, ранние образцы были найдены в Олдувайском ущелье, Танзания. Это первый представитель рода Homo , связанный с использованием каменных орудий труда из-за большого количества отщепов и инструментов, обнаруженных рядом с останками скелетов. Позже разрезанные и раздробленные кости животных также будут встречаться в контексте с 9.0071 H. habilis , что позволяет предположить, что мясо на каком-то уровне было важно для рациона.
Несколько слов о мясе и мозге
С 1950-х годов палеоантропологи связывают увеличенный объем черепа с увеличением содержания мяса в рационе предков человека. Доказательства, подтверждающие эту гипотезу, включают в себя то, что Homo habilis был известным производителем инструментов, и эти инструменты частично использовались для добычи мяса. Кроме того, вплоть до H. habilis объем черепа гоминидов увеличивался. Открытия, подобные этим, в древней Эфиопии, вокруг того места, которое тогда было довольно зеленым озером, интерпретировались как приток белка, который привел к крупным эволюционным и культурным достижениям в человеческой родословной. Да, мясоедение сделало нас умнее. Люди — единственные приматы, которые едят других крупных животных, отличных от представителей нашего вида (и каннибализм тоже хорошо известен). Следы разделки на костях животных, каменных орудиях и роде Homo все появляются в летописи окаменелостей примерно в одно и то же время.
Однако новое исследование предполагает, что, возможно, не мясо, а костный мозг и связанные с ним питательные жиры способствовали раннему развитию человеческого мозга и развитию инструментов (Thompson et al., 2019). Ранние люди, возможно, ели мясо, но употребление мяса на открытом воздухе было для наших предков опасным занятием, так как поблизости скрывались другие хищники и падальщики, а само мясо было трудно перемещать. И с таким количеством очень крупных млекопитающих более вероятно, что люди были одними из этих падальщиков, авантюрно ожидающих, пока крупные хищники обдирают добычу мяса, оставляя после себя кости. Затем кости можно было удалить и легко доставить в безопасное место для употребления в пищу. Кроме того, мясо быстро гниет, а костный мозг дольше остается свежим. Геологическая эпоха этого перехода, плиоцен, также была временем крупных экологических изменений, поскольку климат сильно охлаждался и становился все более изменчивым. Большие участки того, что раньше было тропическим лесом, превратились в пастбища и саванны. После 2 млн лет назад предки человека, вероятно, употребляли в пищу мясо в дополнение к костям, но весьма вероятно, что первыми были сбор костей и обработка их каменными орудиями. Вывод состоит в том, что мы ели мозг до того, как ели мясо.
Череп Homo habilis, в котором можно увидеть многие ранние человеческие, но все еще очень похожие на обезьяны черты. Человеческие черты включают более крупный череп, отсутствие выступающего клыка, более компактный зубной ряд из-за менее выступающего рта и направленное вниз/вперед большое затылочное отверстие. Над глазами еще есть очень выраженный затылочный валик. 3D-модель от BuluLulu на Sketchfab.
Затем осмотрите этот измельчитель из Олдувайского ущелья, Танзания. Возрастом 1,8 млн лет этот инструмент мог быть использован Homo habilis для доступа к костному мозгу. Инструменты такого типа часто были очень стилизованы, что свидетельствует об общей культуре, поскольку единообразие в изготовлении каменных орудий указывает не только на передачу культурных навыков и знаний, но также, возможно, на специализацию навыков в некоторой степени внутри этих ранних предков. человеческие популяции. Отщепы были удалены из более крупного ядра, чтобы создать острую кромку. Это лезвие можно было заострить до тех пор, пока оно не стало бесполезным.
Каменный скребок и древнее каменное орудие. (Модель: Смитсоновский институт, лицензия Creative Commons)
инструменты
Мы знаем, что предки человека — не единственные животные, использовавшие инструменты. Одним из кратких примеров за пределами нашей человеческой семьи являются шимпанзе. Джейн Гудолл хорошо задокументировала использование инструментов шимпанзе, живя среди сообществ шимпанзе. Поведение, которое она наблюдала, включало в себя сбор листовых веток, сдирание стеблей листьев и использование этих стеблей для «ловли» термитов и муравьев. С тех пор шимпанзе используют инструменты для колки орехов, а также затачивают палки для охоты на других млекопитающих. В последние тридцать лет гориллы и бонобо также наблюдались с использованием различных инструментов.
Орудиями пользуются не только приматы. Птиц, китов, морских выдр и даже различных хищников наблюдали с помощью различных инструментов. Учитывая все это, использование инструментов, безусловно, не свидетельствует о человеческом поведении, а является чем-то, что гоминиды, вероятно, унаследовали как наследственную адаптацию, но, похоже, это поведение, которое развилось независимо, развиваясь в нескольких направлениях.
Я понял? Интеллект
Homo erectus и Homo floresiensis – 1,89 млн лет назад до 50 тыс.
лет назад Homo erectus был хорошо известен с момента его первой находки окаменелостей в Индонезии в 1891 году. Это открытие фактически предшествовало всем открытиям предков человека в Африке. Первооткрыватель Юджин Дюбуа первоначально назвал вид Pithecanthropus erectus , или «прямоходящий человек-обезьяна», но в народе его называли «яванским человеком». За этим последовало множество открытий этого вида как в Китае (где его называли «пекинским человеком»), так и в Восточной Африке.
Homo erectus был вполне похож на человека по своим пропорциям. К этому моменту он потерял длинные руки и хватательные способности своих древесных предков. Эти черты уступили место современной человеческой походке и телосложению. H erectus был в среднем довольно высоким, в пределах 145-185 см (4,7-6 футов), с соответствующим диапазоном веса 40-68 кг (88-150 фунтов). Весьма вероятно, что они не только тратили большую часть своего времени на ходьбу, но и бег, вероятно, был важен для их передвижения, возможно, на большие расстояния. Молодь, похоже, росла быстро, как и другие обезьяны, но этот вид продемонстрировал доказательства того, что он, возможно, заботился о пожилых и более слабых особях. Homo erectus также ассоциируется с основным изобретением каменных орудий — ручным топором.
Череп человека прямоходящего. Обратите внимание на гораздо более человеческие характеристики, несмотря на все еще выраженный затылочный гребень. Выступ лица гораздо менее выражен, чем у его предков, а объем черепа значительно больше. (Модель: Смитсоновский институт, лицензия Creative Commons)
Вверху: ручной топор, датируемый 1,1 млн лет назад из Исампура, Индия и связанный с использованием Homo erectus. Подобные каменные орудия гораздо более продвинуты по дизайну и навыкам, необходимым для их создания, что указывает на развитие некоторой специализации в этом ремесле. (Модель: Смитсоновский институт, лицензия Creative Commons)
Выстукивание кремня или ручная обработка каменных инструментов было непростым делом. Это требовало высокоспециализированных и приобретенных навыков, особенно по мере того, как работа переходила от простых скребков к хорошо отесанным наконечникам копий. Это очень сложно. Видео ниже представляет собой отличное введение в древний набор навыков:
Homo erectus , безусловно, является самым долгоживущим видом Homo . Он прожил дольше, чем анатомически современные люди смогли просуществовать. Их высокое телосложение и большой мозг требовали очень большого количества энергии для функционирования, что делало мясоедение для легкоусвояемого белка ключевым ингредиентом их рациона. Развитие их каменных орудий определенно указывает на то, что обработка мяса является важной частью рациона. Еще одно развитие, на которое важно обратить внимание, — это использование огня. Вплоть до Homo erectus в человеческом происхождении не было никаких свидетельств целенаправленного разведения и использования огня. Сейчас остатки очагов находят в контексте. Хотя они почти наверняка использовались для приготовления пищи, они также, по-видимому, использовались для социального взаимодействия, тепла и отпугивания хищников.
Вымершим потомком Homo erectus является Homo floresiensis . По прозвищу «хоббит», он был обнаружен совсем недавно (2003 г.) на индонезийском острове Флорес. H. floresiensis также связан с использованием каменных орудий труда, но, похоже, он вымер уже 50 тыс. лет назад. Известные люди (а их всего несколько) имели рост всего 106 см (3,5 фута), крошечный мозг, большие зубы и множество необычных черт лица. К ним относятся плоские лбы и менее выраженные подбородки. Их ступни были очень большими для их ног. Несмотря на все это, есть свидетельства того, что они частично питались маленькими слонами, крупными грызунами, использовали огонь и изготавливали каменные орудия труда. На них могли охотиться большие вараны (например, драконы Комодо). Одна возможность для отличного роста и общей формы H. floresiensis is островная карликовость .
Homo heidelbergensis -1,3 млн лет назад 200 тыс. лет назад
В конце концов предки человека распространились за пределы Африки, Китая и Индонезии в Европу. Первый экземпляр Homo heidelbergensis был обнаружен недалеко от Гейдельберга, Германия, в 1908 году. Это первый вид предков человека, обитающий в более прохладном климате. Жить в Европе в разгар «Ледниковых периодов» эпохи плейстоцена было бы непросто, поэтому мы можем предположить потребность в покрытии тела, умение обращаться с огнем и другие приспособления к холодной погоде, включая охоту на крупных животных. Действительно, у нас этого много. Например, на территории Гешре Бенот Яаков в Израиле есть модифицированные огнем инструменты и дерево. H. hiedelbergensis также, вероятно, строил убежища. Доказательства этого приходят с сайта Терра Амата, Франция.
Доказательства того, что они охотились на крупную дичь, получены в результате анализа костей тех животных, у которых они были обнаружены вместе с останками H. heidelbergensis . К ним относятся следы бойни на оленях, лошадях, слонах, бегемотах и носорогах. Пример участка с такими особенностями, как раннее колющее копье, существует недалеко от Шёнингена, Германия.
Наконец, есть доказательства человеческого ритуала среди H. heidelbergensis недалеко от Атапуэрки, северная Испания. По-видимому, преднамеренное массовое захоронение 30 особей датируется 400 тыс. лет назад. Был ли это ритуал? Безусловно, в таком захоронении была какая-то цель. Эта яма, получившая прозвище Sima de los Huesos , или «Яма костей», также содержала свидетельства использования каменных орудий труда, поскольку в контексте был обнаружен ручной топор.
Череп H. heidelbergensis. Обратите внимание на человеческий вид этого черепа. Черепная коробка намного крупнее, а лицо более плоское. Скулы менее выражены. Тем не менее, есть довольно приподнятый затылочный гребень, который является более обезьяноподобным признаком, характерным для всех предков человека до этого момента. (Модель: Смитсоновский институт, лицензия Creative Commons)
Homo naledi – 335-236 тыс. лет назад
Останки Homo naledi до сих пор были обнаружены только в Южной Африке. H. naledi — родственники людей, которые никогда не покидали Африку. На месте обнаружения этого вида обнаружено 15 особей. Это один из крупнейших известных тайников ископаемых гоминидов. Таким образом, в этой небольшой популяции, включая как взрослых, так и молодых особей, имеется изрядное количество физиологической информации. Их останки были датированы возрастом от 335 000 до 236 000 лет.
Несмотря на явное сходство с другими видами рода Homo , ряд признаков Australopithecus также сохраняется. Это затруднило размещение H. naledi на филогенетическом дереве гоминидов.
Череп Homo naledi. В то время как лицо все еще сохраняет более обезьяноподобный выступ, а затылочный гребень все еще очень выражен, общий вид этого южноафриканского кузена современного человека имеет зубы, очень близкие к нашим, большой череп, передние скулы (скуловая дуга) и кажется, определенно был двуногим. 3D-модель ArtinScienceMA; используется с разрешения.
Неандертальцы
До сих пор было известно, что единственный вид Homo мигрировал из Африки, Homo erectus . У нас мало информации о судьбе этого вида, но мы знаем, что он вымер около 117 000 лет назад, задолго до того, как современные люди, Homo sapiens , столкнулись с такими же средами обитания в Азии. Таким образом, в настоящее время нет никаких доказательств того, что эти два вида когда-либо взаимодействовали.
Реконструкция головы и лица неандертальской женщины.Изображение: Wikimedia, By Bacon Cph CC BY 2.
5 Реконструкция пожилого неандертальца. Wikimedia, Pressebilder Neanderthal Museum, CC BY-SA 4.0
Ситуация с Homo sapiens neanderthalensis совершенно иная. Неандертальцы — подвид нашего собственного вида, и есть достаточно доказательств того, что они также произошли из Африки. Они покинули континент задолго до того, как наш вид, Homo sapiens sapiens , начал свою собственную миграцию (350 тыс. лет назад). Уехав, они мигрировали по всей Азии и в Европу. Учитывая эту временную шкалу, возможно, что они взаимодействовали с Homo erectus особей, обитавших в то время в Азии, хотя в настоящее время убедительных доказательств, подтверждающих это, нет. Недавняя информация свидетельствует о том, что древние близкие родственники неандертальцев, называемые «денисовцами», жили бок о бок и скрещивались с нашим видом на протяжении тысячелетий. Известные главным образом своими уникальными вариантами ДНК, обнаруженными у современных людей, они пережили своих неандертальских родственников как минимум на 10 000 лет, вымерли лишь 15 000 лет назад. Последнее задокументированное свидетельство сосуществования денисовцев и современных людей, установленное с помощью данных ДНК 30 тыс. лет назад, происходит из Новой Гвинеи (Jacobs et al., 2019).).
Изображение: Википедия, Джон Д. Крофт, CC BY-SA 3.0.
Современные люди и неандертальцы ненадолго взаимодействовали в Европе, прежде чем неандертальцы там тоже вымерли. Неизвестно, почему неандертальцы вымерли, хотя в качестве гипотез предлагались изменение климата и различная природа их взаимодействий между современными людьми. Некоторые примеры последнего включают скрещивание с людьми (ассимиляция в Homo sapiens sapiens 9).0072 населения) и врожденные проблемы, неблагоприятное положение человека в одних и тех же экосистемах, паразиты, патогены и насилие. Одно из последних известных занятий неандертальцев в Европе или Азии найдено в пещере на побережье Гибралтара, Испания. Остатки датируются 42 тыс. лет назад.
Изображение: Wikimedia, CC BY-SA 3.0
Верхняя часть черепа Homo sapiens neanderthalensis. Присутствуют все черты, присущие современным людям, за исключением более толстого затылочного гребня. (Модель: Смитсоновский институт, лицензия Creative Commons)
Нижняя челюсть неандертальца с зубами, которые очень соответствуют современным людям и смешанной всеядной диете. Сохранившиеся челюсти более редки у ископаемых гоминидов. Эта нижняя челюсть включена сюда как пример зубных рядов нижней челюсти у неандертальцев, поскольку они утратили любой ярко выраженный клык нижней челюсти, а также связанную с ним диастему. (Модель: Смитсоновский институт, лицензия Creative Commons)
Целую главу можно было бы посвятить тому, что мы знаем о неандертальцах. Что касается их культуры, мы знаем, что у них был разнообразный набор инструментов, они строили убежища, использовали огонь, носили одежду, охотились на крупную дичь (они были высшими хищниками), питались растениями, а также участвовали в ритуальных действиях. Этот последний пункт является наиболее важным. Ни один другой предшествующий гоминид не демонстрирует такой степени ритуализма, какой был известен среди неандертальцев, кроме современных людей. Прекрасным примером этого является включение того, что кажется подношением, например цветов для умерших, которые были похоронены.
Вместо длинного разоблачения неандертальцев в приведенном ниже видео представлена дополнительная информация, которая поможет нам понять нашу собственную человечность через призму их окаменелостей.
Также уникальным для неандертальцев является существование реальных прямых образцов ДНК нескольких людей из самых разных мест, включая Сибирь, Хорватию, Испанию и Германию. К 2009 году вся известная последовательность ДНК неандертальца была секвенирована Институтом эволюционной антропологии Макса Планка в рамках проекта «Геном неандертальца».
Современные люди
В этом тематическом исследовании задокументирована ваша и моя история. Это история наших предков-гоминидов в далеком прошлом. Полная история современного человека разнообразна, сложна и выходит за рамки данного тематического исследования. Однако следует отметить несколько важных моментов:
Как и все другие виды гоминидов, мы вышли из Африки. Эта миграция началась между 100 и 60 тыс. лет назад как вдоль Синайского полуострова в Египте, так и через то, что сейчас является Баб-эль-Мандебским проливом между Джибути и Йеменом. В момент миграции приближался последний ледниковый период, ледниковые щиты продвигались на север, а уровень моря был значительно ниже. Части Африки, покрытые тропическими лесами, уступили место саванне и даже пустыне в результате того, что иногда называют мегазасухой (Gibbons, 2007). По сути, наши предки, Homo sapiens sapiens , были вытеснены на более зеленые пастбища на восток, север и дальше на юг вглубь Африки.
Хотя имеется достаточно ископаемых свидетельств того, что люди покинули Африку в это время, а затем мигрировали по всей Азии, Европе и в Северную Америку (через Берингию уже 16,5 тыс. лет назад), трудно проследить эту миграцию в деталях. Тем не менее, многочисленные исследования пытались заполнить эти пробелы, используя ДНК, отслеживая митохондриальную ДНК (на материнской стороне) и через Y-хромосому (на отцовской стороне). Эти исследования проследили всех современных людей до «митохондриальной Евы», которая жила в Восточной Африке около 150 тыс. лет назад. На основе этой ДНК материнские гаплогруппы (лица, имеющие общего предка) можно проследить по всему миру, в том числе в Северной Америке. Все самки, даже члены групп, которые никогда не покидали Африку, имеют эту митохондриальную подпись.
Гипотетическая карта миграций человека на основе митохондриальной ДНК. Изображение: Wikimedia, Mauluccioni, CCBY-SA 3.0
Точно так же можно проследить отцовские гаплогруппы (используя Y-хромосому) по всему миру. В данном случае, однако, все мужчины, которые разделяют эту версию Y-хромосомы, происходят от одного человека, который, по-видимому, мигрировал из Африки около 60 000 лет назад, вероятно, выбрав для этого южный путь через Йемен. Важно отметить, что некоторые человеческие популяции, такие как представители народности !кунг в Намибии, Анголе и Ботсване, не имеют этой Y-хромосомы. Это потому, что они произошли от другой отцовской линии, которая отделилась до 60 000 лет, чтобы углубиться в западную Африку.
Карта ниже представляет собой слой ArcGIS Online, который иллюстрирует модели миграции различных материнских (митохондриальных) и отцовских (Y-хромосомных) гаплогрупп среди Homo sapiens sapiens , когда они мигрировали из Африки. Нажатие на поля включает и выключает различные гаплогруппы, и вы можете увидеть, куда члены этой группы мигрировали по всему миру:
Инструкции по использованию этой карты можно найти здесь.
Haplogroup.info, используется в рамках CCBY 3.0.
Изображение: Википедия, Д. Бахман, CC BY-SA 4.0.
Мы также можем проследить эту миграцию, особенно последние ее части, используя важное эволюционное и культурное развитие: язык. Подобно эволюционным изменениям среди изолированных скоплений видов, развитие языка происходит быстро, очень культурно специфично, эволюционно по своей природе и, таким образом, является очень полезным инструментом для антропологических и археологических исследований современной человеческой миграции и культуры. В мире существует большое разнообразие языковых групп, каждая из которых может быть прослежена лингвистами до нескольких основных языковых компонентов, которые в совокупности раскрывают аспекты того, что когда-то могло быть «родным языком» для этой основной языковой семьи.
Изображение: Википедия, выпускник, CC BY-SA 4.0 Схема распространения индоевропейского языка с ок. с 4000 по 1000 г. до н.э.
Изображение: Википедия, Джошуа Джонатан, CC BY-SA 4.0
Я понял? Гаплогруппы и миграция
Homo sapiens, Климат и природные ресурсы
Что вызвало всю эту миграцию? Homo sapiens sapiens , как и все другие животные, стремится есть, размножаться и выживать. В отличие от многих других животных, наш вид мигрировал способами, выходящим за рамки сезонных циклов. Мы смогли адаптироваться благодаря использованию инструментов, нашей диеты и погоне за ресурсами. Сегодня наш вид живет совсем иначе (в большинстве случаев), чем наши предки тысячи лет назад, и для большинства из нас мы живем совсем иначе, чем наши недавние предки всего столетие или даже несколько десятилетий назад. Чем вызваны эти изменения?
Значительная часть этого ответа связана с изменением климата. До того, как мы активно играли роль в изменении нашего климата, как мы делаем это сегодня, наш климат в значительной степени диктовал, где мы звонили домой. Изменение климата во многих случаях способствовало миграции. Несмотря на это, наши предки выдержали гораздо более холодный климат и проблемы с приспособляемостью, которые принес с собой этот климат. Однако технологические достижения произошли независимо от тех, которые обусловлены климатом. Хотя адаптация к более холодным регионам, безусловно, требовала обновления наших наборов инструментов, мы обновили эти наборы и для более теплого климата. Каменный век уступил место неолитическим технологиям (земледелие, животноводство). К этим новым неолитическим способам мы в конечном итоге добавим использование металлов, таких как медь, бронза, железо и, наконец, кремний (сегодня). Все эти инновации требуют дальнейшей добычи ресурсов. По сути, эта добыча ресурсов аналогична той, что практиковали наши предки-микробы более миллиарда лет назад. Как и в случае с более простыми организмами, все виды использования окружающей среды человеком были направлены на обеспечение нашего вида пищей, убежищем и возможностью размножаться.
В конце концов, мы животные.
Я понял? Люди и климат
Гиббонс, Энн (2007). Мегазасуха подкосила наших предков. Новости науки .
Джейкобс, Гай С., Худжашов, Георги, Сааг, Лаури, Кусума, Прадиптаджати, Дарусаллам, Челзи К., Лоусон, Дэниел Дж., Мондал, Маюх, Пагани, Лука, Рико, Франсуа-Ксавье, Стоункинг, Марк, Метспалу, Мейт, Судойо, Херавати, Лансинг, Дж. Стивен и Мюррей П. Кокс (2019). Множественные глубоко расходящиеся денисовские предки папуасов. Сотовый 177(4).
Паттерсон, Ник, Рихтер, Дэниел Дж., Гнерре, Санте, Ландер, Эрик С. и Дэвид Райх (2006). Генетические доказательства сложного видообразования человека и шимпанзе. Природа 441(7097).
Щулкин, Джей (2016). Эволюционная основа бега человека и его влияние на нервную функцию. Границы нейронауки 10:59.
Смитсоновский национальный музей естественной истории Хронология эволюции человека, по состоянию на 04.02.2021.
Инициатива Смитсоновского национального музея естественной истории «Происхождение человека», по состоянию на 04.02.2021.
Таттерсолл, Ян (2013). Хозяева планеты: В поисках нашего человеческого происхождения . Гриффин Святого Мартина.
- 1 Люди как вид
- 2 Палеоантропология
- 2,1 Терминология филогения гоминид
- 2,2 Терминология черепа
- 3 Основные моменты нашего семейного дерева: Australopithecina and Homo
- 3 Основные моменты нашего семейного дерева: Australopithecina и Homo
- 3
- 3
- .0308
- 3.2 Orrorin Tugenensis и Ardipithecus kadabba-5-6 MA
- 3.3 Ardipithecus ramidus-4,4 мА
- 3.3.1 Слово о бипеджизме
- 3.3.1.1. Слово о бипедализме. anamensis, A. afarenesis, A. Africanus, A. garhi, A. Sediba – 1,5–4 млн лет
- 3. 4.1 Australopithecus anamensis – 4,2–3,8 млн лет
- 3.4.2 «Люси» – Australopithecus afarensis – 3,85–2,95 млн лет
- 3.4.3 Африканский австралопитек – от 3,3 до 2,1 млн лет назад
- 3.4.4 Australopithecus Garhi и A. Sediba
- 3.
- 3.5 Род Homo-2,4 мА
- 3.5.1 Homo habilis-2,4-1,4 мА
- 3.5.1.1. Слово о мясе и браслетах
- 3,5,1,1,2 3,5,1,2 3,5,1, Инструменты
- 3.5.2. Homo erectus и Homo floresiensis-1,89 млн. До 50 Ka
- 3.5.3 Homo heidelbergensis-1,3 Ma-2008 Ka
- 3.5.4 Homo naledi-335-236 Ka
- .5.5.5.5.5.5.5.5.5.5.5.
- 3.5.6 Современные люди
- 3.5.1 Homo habilis-2,4-1,4 мА
- 3.6 Homo sapiens, климат и природные ресурсы
- 3.3.1.1. Слово о бипедализме. anamensis, A. afarenesis, A. Africanus, A. garhi, A. Sediba – 1,5–4 млн лет
- 4 Дополнительная литература
О филогении и онтогенезе гортани человека: морфологическое и функциональное исследование | JAMA Отоларингология – Хирургия головы и шеи
О филогении и онтогенезе гортани человека: морфологическое и функциональное исследование | JAMA Отоларингология – Хирургия головы и шеи | Сеть ДЖАМА [Перейти к навигации]Эта проблема
- Скачать PDF
- Полный текст
-
Поделиться
Твиттер Фейсбук Эл.
адрес
LinkedIn
- Процитировать это
- Разрешения
Артикул
Август 1972 г.
РОБЕРТ М. КОМОРН, MD
Арка Отоларингол. 1972;96(2):192-193. дои: 10.1001 / архотол.1972.007700
031
Полный текст
Эта статья доступна только в формате PDF. Загрузите PDF-файл, чтобы просмотреть статью, а также связанные с ней рисунки и таблицы.
Абстрактный
Доктор Винд явно потратил много времени и усилий на изучение филогенеза и онтогенеза человеческой гортани. Его книга, изначально являвшаяся докторской диссертацией, показывает развитие гортани человека от простого мышечного сфинктера до сложного органа, участвующего в защитной, дыхательной, глотательной и голосовой функциях.
Используемый подход заключается в том, чтобы проследить филогенетическое развитие, а затем сравнить и связать его с эмбриональным развитием гортани человека. Книга разделена на три части. Первый раздел посвящен терминологии и понятиям филогенеза. Он содержит краткий обзор современных теорий органической эволюции и показывает, как их можно применить как к изучению филогении человека в целом, так и к изучению гортани в частности. Доктор Винд указывает на трудности, присущие попыткам изучить органическую эволюцию органа из мягких тканей
Предварительный просмотр первой страницы Просмотреть большой
Полный текст
Добавить или изменить учреждение
- Кислотно-щелочное, электролиты, жидкости
- Наркологическая медицина
- Аллергия и клиническая иммунология
- Анестезиология
- Антикоагулянты
- Искусство и изображения в психиатрии
- Кровотечение и переливание
- Кардиология
- Уход за тяжелобольным пациентом
- Проблемы клинической электрокардиографии
- Клиническая задача
- Поддержка принятия клинических решений
- Клинические последствия базовой нейронауки
- Клиническая фармация и фармакология
- Дополнительная и альтернативная медицина
- Заявления о консенсусе
- Коронавирус (COVID-19)
- Медицина интенсивной терапии
- Культурная компетенция
- Стоматология
- Дерматология
- Диабет и эндокринология
- Интерпретация диагностических тестов
- Разнообразие, равенство и инклюзивность
- Разработка лекарств
- Электронные медицинские карты
- Неотложная медицинская помощь
- Конец жизни
- Гигиена окружающей среды
- Этика
- Пластическая хирургия лица
- Гастроэнтерология и гепатология
- Генетика и геномика
- Геномика и точное здоровье
- Гериатрия
- Глобальное здравоохранение
- Руководство по статистике и медицине
- Руководство
- Заболевания волос
- Модели оказания медицинской помощи
- Экономика здравоохранения, страхование, оплата
- Качество медицинской помощи
- Реформа здравоохранения
- Медицинская безопасность
- Медицинские работники
- Различия в состоянии здоровья
- Неравенства в отношении здоровья
- Информатика здравоохранения
- Политика здравоохранения
- Гематология
- История медицины
- Гуманитарные науки
- Гипертония
- Изображения в неврологии
- Наука внедрения
- Инфекционные болезни
- Инновации в оказании медицинской помощи
- Инфографика JAMA
- Право и медицина
- Ведущее изменение
- Чем меньше, тем лучше
- ЛГБТК
- Образ жизни
- Медицинское кодирование
- Медицинские приборы и оборудование
- Медицинское образование
- Медицинское образование и обучение
- Медицинские журналы и публикации
- Меланома
- Мобильное здравоохранение и телемедицина
- Оспа обезьян
- Нарративная медицина
- Нефрология
- Неврология
- Неврология и психиатрия
- Примечательные примечания
- Сестринское дело
- Питание
- Питание, Ожирение, Упражнения
- Ожирение
- Акушерство и гинекология
- Гигиена труда
- Онкология
- Офтальмологические изображения
- Офтальмология
- Ортопедия
- Отоларингология
- Лекарство от боли
- Патология и лабораторная медицина
- Уход за больными
- Информация для пациентов
- Педиатрия
- Повышение производительности
- Показатели эффективности
- Периоперационный уход и консультации
- Фармакоэкономика
- Фармакоэпидемиология
- Фармакогенетика
- Фармация и клиническая фармакология
- Физическая медицина и реабилитация
- Физиотерапия
- Врачебное руководство
- Поэзия
- Здоровье населения
- Профилактическая медицина
- Профессиональное благополучие
- Профессионализм
- Психиатрия и поведенческое здоровье
- Общественное здравоохранение
- Легочная медицина
- Радиология
- Регулирующие органы
- Исследования, методы, статистика
- Реанимация
- Ревматология
- Управление рисками
- Научные открытия и будущее медицины
- Совместное принятие решений и общение
- Медицина сна
- Спортивная медицина
- Трансплантация стволовых клеток
- Хирургия
- Хирургические инновации
- Хирургические жемчужины
- Обучаемый момент
- Технологии и финансы
- Искусство JAMA
- Искусство и медицина
- Рациональное клиническое обследование
- Табак и электронные сигареты
- Токсикология
- Травмы и травмы
- Приверженность лечению
- УЗИ
- Урология
- Руководство пользователя по медицинской литературе
- Вакцинация
- Венозная тромбоэмболия
- Здоровье ветеранов
- Насилие
- Женское здоровье
- Рабочий процесс и процесс
- Уход за ранами, инфекция, заживление
Сохранить настройки
Политика конфиденциальности | Условия использования
генетика — Онлайн филогенетическое дерево человеческих родословных
Задавать вопрос
Спросил
Изменено 3 года, 7 месяцев назад
Просмотрено 3к раз
$\begingroup$
Я ищу источник информации о разнообразии человеческих родословных и их взаимоотношениях.
С помощью быстрого поиска в Google легко найти этот тип дерева
Идеальный онлайн-ресурс будет…
- Содержит множество родословных, идущих к наиболее подробным родословным
- Содержит ссылки на Википедию или другой источник информации, который может помочь кому-то:
- определить группу
- имеют представление о своей культуре
- Содержат визуализацию своего текущего (или прошлого) географического ареала.
- Содержит визуализацию шаблонов миграции
- Поскольку мтДНК и Y-хромосома обычно могут показывать разные истории, было бы здорово, если бы ресурс различал их и выделял конкретные случаи, когда эти два дерева различаются.
Можете ли вы порекомендовать какие-нибудь онлайн-ресурсы?
Также приветствуется обзорная статья с красивыми графиками.
Связанный:
-
Сообщение Лучшее бесплатное и наиболее актуальное филогенетическое дерево в Интернете? порекомендуйте ресурсы для визуализации древа жизни.
-
Этот сайт предлагает удивительное дерево религий.
- генетика
- эволюция
- генетика человека
- база данных
- эволюция человека
$\endgroup$
4
$\begingroup$
Деревья для гаплогрупп
См.
- https://www.eupedia.com/genetics/phylogenetic_trees_Y-DNA_haplogroups.shtml
- https://www.eupedia.com/genetics/
- фокус на Европу: https://www.eupedia.com/europe/origins_haplogroups_europe.shtml
- и многие страницы, связанные с ним, такие как https://www.eupedia.com/europe/Haplogroup_J2_Y-DNA.shtml#subclades
- и так далее.
Подробнее о мтДНК (женская линия):
- https://en.wikipedia.org/wiki/Human_mitochondrial_DNA_haplogroup
- и множество ссылок в нем
Подробнее для Y-хромосомы (мужская линия):
- https://en. wikipedia.org/wiki/Human_Y-chromosome_DNA_haplogroup
- и множество ссылок в нем
- https://en.wikipedia.org/wiki/List_of_Y-chromosome_haplogroups_in_populations_of_the_world и ссылки в нем
- https://isogg.org/tree/index.html и связанные с ним Google Таблицы
- https://www.yfull.com/tree/
«Деревья» для популяций
Обратите внимание, что деревья предназначены для генов/гаплогрупп, тогда как такое «дерево» для популяций на самом деле также имеет поток генов через ветви. Представьте, что дерево мтДНК (женское происхождение) накладывается на немного другое дерево Y-ДНК (мужское происхождение).
География миграции
Миграция и географическая удаленность сыграли ключевую роль в появлении обособленных популяций. См.
- карты миграции по гаплогруппам: Миграция людей по гаплогруппам, Карта мира гаплогрупп Y-ДНК, Ранние миграции людей, Упрощенная интерактивная карта, И так далее.
- 3.3.1 Слово о бипеджизме