
Функции сна: Сомнология: сон, его структура и функции; инсомния | Левин Я.И.
Использование функции «Режим сна» на iPhone для отслеживания сна
Задайте количество времени, которое вам необходимо тратить на сон каждую ночь, и приложение «Часы» будет напоминать вам, когда пора идти спать, и ставить будильник, чтобы будить вас.
Настройка функции «Режим сна»
При первоначальной настройке режима сна в приложении «Часы» вам будет задано несколько вопросов.
- Откройте приложение «Часы» и выберите вкладку «Режим сна».
- Нажмите «Приступить» и выберите настройки.

- Нажмите «Готово».
После установки времени отхода ко сну iPhone будет напоминать вам, когда пора идти спать, а в указанное время пробуждения будет звонить будильник.
Включение и отключение функции «Режим сна»
- Откройте приложение «Часы» и выберите вкладку «Режим сна».
- В разделе «Расписание» нажмите «Отход ко сну» или «Пробуждение».
- В верхнем правом углу включите или выключите функцию «Расписание сна».
Функция отключает будильник и напоминание об отходе ко сну.
Изменение времени и дней срабатывания будильника и напоминаний
- Откройте приложение «Часы» и выберите вкладку «Режим сна».
- В разделе «Расписание» нажмите «Отход ко сну» или «Пробуждение».
- Перетащите кнопку «Отход ко сну» или кнопку «Будильник» . По мере перетаскивания время в полях «Отход ко сну» и «Пробуждение» будет обновляться автоматически.

- В разделе «По каким дням» выберите дни срабатывания будильника и получения напоминаний. Отмеченные дни выделяются оранжевым цветом.
Отслеживание истории сна
- Откройте приложение «Часы» и выберите вкладку «Режим сна».
- Прокрутите экран вниз до раздела «Анализ», где показана недавняя история.
Если в предполагаемое время сна вы будете использовать iPhone, этот период будет засчитан в приложении как бодрствование. Если вы отложите срабатывание будильника, время сна обновится соответственно.
Данные, собираемые функцией «Режим сна», будут автоматически передаваться в приложение «Здоровье».
Изменение настроек
- Откройте приложение «Часы» и выберите вкладку «Режим сна».

- В верхнем левом углу нажмите «Параметры».
- Изменить можно следующие настройки.
- Время активации напоминания об отходе ко сну.
- Включите или выключите функцию «Отслеживать время сна». Функция дает более полное представление о вашем режиме сна, отслеживая моменты, когда вы ночью используете свой iPhone.
- Включение или выключение параметра «Не беспокоить во время сна». Во время сна яркость экрана блокировки уменьшается, отключаются звуковые сигналы о вызовах и оповещениях, которые приходят на заблокированное устройство, а уведомления регистрируются в истории.
- Мелодию для пробуждения на будильнике.
- Громкость будильника.
- Нажмите «Готово».
Просмотр истории в приложении «Здоровье»
Чтобы анализировать сон на ОС iOS 13, откройте приложение «Часы», выберите вкладку «Режим сна», затем нажмите «Показать еще в Здоровье».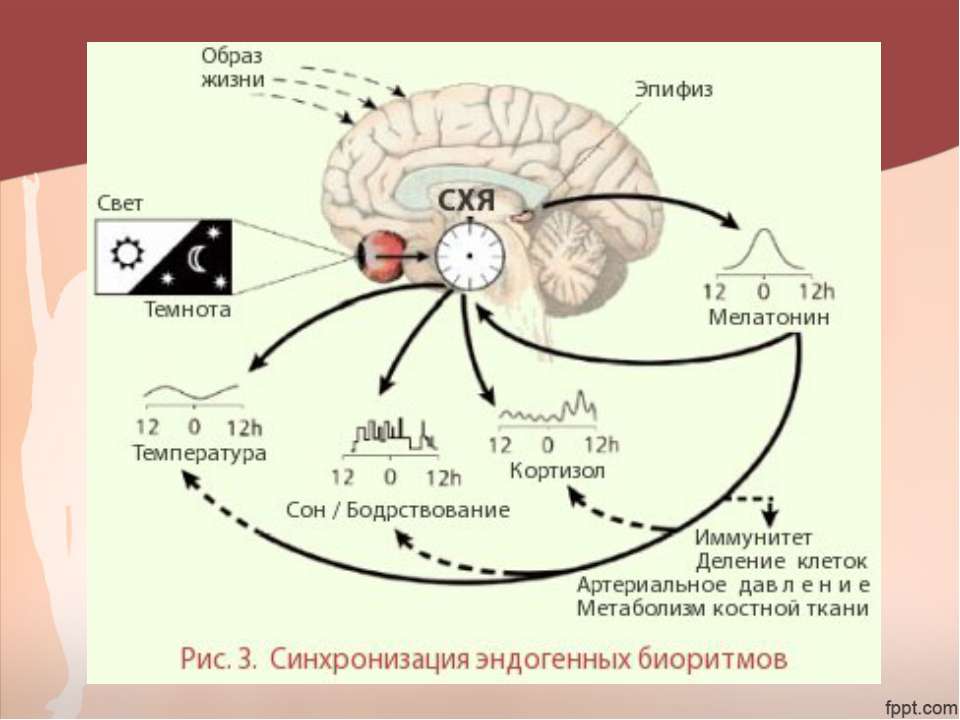 На графике «Анализ сна» отображается время, проведенное в постели или во сне.
На графике «Анализ сна» отображается время, проведенное в постели или во сне.
Функция «Режим сна» в приложении «Часы» отслеживает время, которое вы провели в постели, но не то, сколько вы спали или двигались. Часы Apple Watch также неспособны отслеживать показатели сна или время в постели.
Чтобы определить, сколько времени вы спите, попробуйте подключить к приложению «Здоровье» устройство для отслеживания сна. Также вы можете вручную указать время своего сна: откройте приложение «Часы», выберите вкладку «Режим сна», затем нажмите «Показать еще в Здоровье». В правом верхнем углу нажмите «Добавить данные». Узнайте больше о приложении «Здоровье».
Дата публикации:
Сон и когнитивные функции uMEDp
Представлены данные исследований влияния сна на функцию памяти. Разбираются механизмы, облегчающие консолидацию во сне. Описывается роль медленного и быстрого сна в обеспечении памяти различной модальности. На примере моделей депривации сна и таких расстройств сна, как синдром обструктивного апноэ сна и инсомния, разбирается влияние нарушения сна на функцию памяти и вероятность развития когнитивных расстройств. Оцениваются возможности улучшения когнитивных функций на фоне коррекции нарушений сна. Значение фаз сна для различных аспектов памятиВведение
Важнейшей функцией сна представляется обеспечение эффективной деятельности центральной нервной системы. Психическая активность – одна из главных форм такой деятельности. Когнитивные функции определяются как виды психической деятельности, которые направлены на познание человеком себя и окружающего мира (от лат.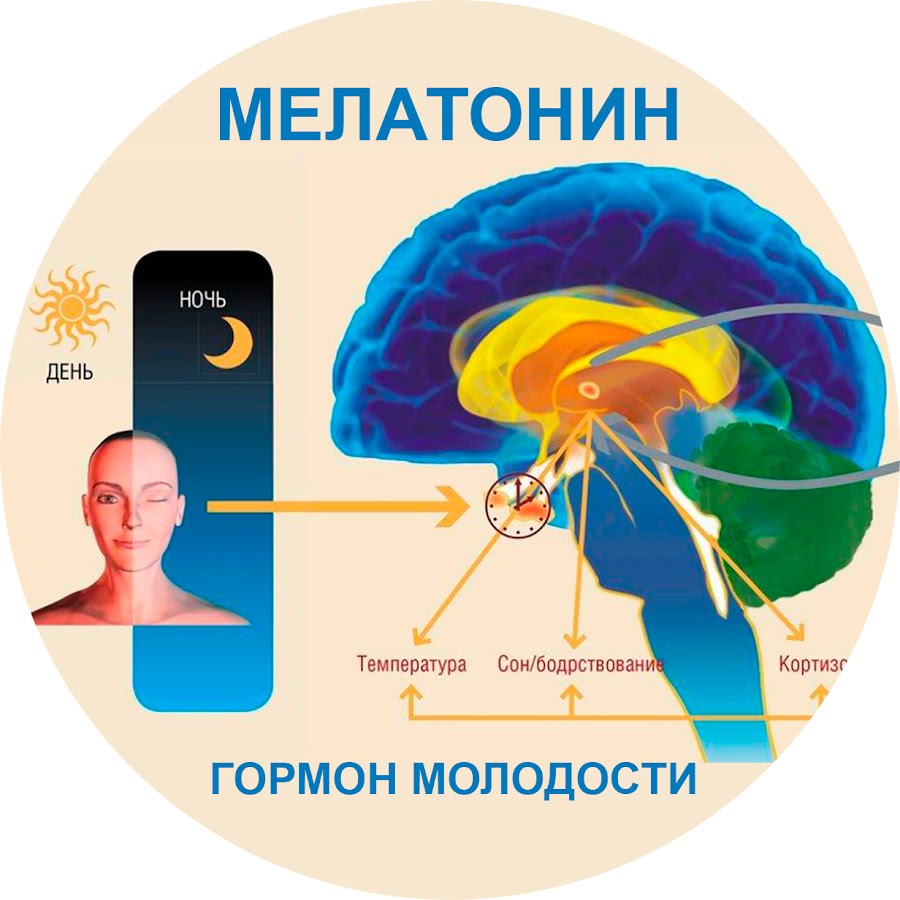
- восприятие информации – гнозис;
- обработка и анализ информации – так называемые исполнительные функции: произвольное внимание, обобщение, выявление сходств и различий, формально-логические операции, установление ассоциативных связей, вынесение умозаключений;
- запоминание и хранение информации – память;
- обмен информацией и построение и осуществление программы действий – так называемые экспрессивные функции: речь и навыки целенаправленной двигательной активности (праксис) [1].
В первом же опыте по полной депривации (лишению) сна человека, проведенном психологами из университета Айовы G.
Сон и память
Взаимоотношения важнейшей для обеспечения процесса познания когнитивной функции – памяти с состоянием сна представляются в настоящее время недостаточно изученными. Прежде всего это связано с гетерогенностью механизмов, используемых в процессе научения и запоминания.
Память – общее обозначение комплекса познавательных способностей и высших психических функций по накоплению, сохранению и воспроизведению знаний и навыков. Существует несколько классификаций памяти. По времени хранения выделяют ультракороткую (сенсорную), кратковременную и долговременную память. По степени осознанности – имплицитную (недекларативную, неосознаваемую) и эксплицитную (декларативную, произвольную или сознательную). Декларативная память представлена эпизодической (памятью на события), а также семантической памятью, содержащей знания, запечатленные средствами языка (семантический – относящийся к значению или смыслу слова). К недекларативной памяти относят процедурную (знание, как делать) и перцептуальную (память ощущений). В другом варианте этой дихотомии недекларативную память разделяют на навыки/привычки, прайминг (облегчение восприятия связанных стимулов) и условные рефлексы. Есть и другие классификации.
Существует несколько классификаций памяти. По времени хранения выделяют ультракороткую (сенсорную), кратковременную и долговременную память. По степени осознанности – имплицитную (недекларативную, неосознаваемую) и эксплицитную (декларативную, произвольную или сознательную). Декларативная память представлена эпизодической (памятью на события), а также семантической памятью, содержащей знания, запечатленные средствами языка (семантический – относящийся к значению или смыслу слова). К недекларативной памяти относят процедурную (знание, как делать) и перцептуальную (память ощущений). В другом варианте этой дихотомии недекларативную память разделяют на навыки/привычки, прайминг (облегчение восприятия связанных стимулов) и условные рефлексы. Есть и другие классификации.
По-видимому, первое упоминание о связи памяти со сном содержится в первой в истории монографии по сну «Философия сна» (1827) шотландского хирурга и философа R. Macnish, который описывает нарушение сна у людей с психическими заболеваниями, в частности при алкогольном делирии [3, цит. по изданию 1836 г.].
по изданию 1836 г.].
В уже упомянутом исследовании G. Patrick и J. Gilbert (1896) было обнаружено нарушение запоминания числовых последовательностей при полной депривации сна. О недостаточном качестве этой работы свидетельствует то, что при повторении этого исследования в 2008 г. психологами из Огайо T. Fuchs и J. Burgdorf в отношении запоминания были получены противоположные результаты: после депривации сна испытуемые успешно воспроизводили числовые последовательности в 100% случаев [4].
Тщательно спланированное исследование влияния сна на запоминание было проведено в 1924 г. психологами из Иллинойса J. Jenkins и K. Dallenbach [5]. Два студента-добровольца в течение двух месяцев жили в условиях лаборатории и пытались запоминать предъявляемые им бессмысленные утверждения. Через различные периоды времени (один, два, четыре, восемь часов), включавшие состояния сна или бодрствования, испытуемых просили воспроизвести материал. Уже через час отмечались достоверные отличия в количестве воспроизведенного материала в зависимости от функционального состояния – в случае, если ему предшествовал сон, воспроизводилось большее количество утверждений.
В дальнейшем многочисленные исследования, проводившиеся в конце XX–XXI вв., подтвердили активную роль сна в процессах консолидации памяти. Сон – неоднородное состояние, в нем выделяют две фазы – медленного (ФМС) и быстрого сна (ФБС), которые по-разному участвуют в процессах запоминания в зависимости от того, какая из модальностей памяти задействована [6].
Предполагают, что в ФМС происходят процессы консолидации декларативной и пространственной памяти, в то время как ФБС в большей степени связана с процессами недекларативной памяти. Это было продемонстрировано в опытах по селективному лишению одной из фаз сна. Воспроизведение парных слов оказалось лучшим в том случае, если после заучивания испытуемые имели возможность спать хотя бы половину времени сна, богатого ФМС. В то же время тест зеркального рисования, зависящий от состояния процедурной памяти, реагировал на количество сна, получаемого во второй половине, обычно богатой ФБС.
В то же время тест зеркального рисования, зависящий от состояния процедурной памяти, реагировал на количество сна, получаемого во второй половине, обычно богатой ФБС.
Существует и иная концепция, предполагающая, что процессы запоминания во сне идут последовательно. В любом случае признается важность сна для успешной консолидации полученной информации. При этом не подвергается сомнению и роль бодрствования: запоминание происходит даже в условиях полной депривации сна, однако оно тем успешнее, чем больше связано со сном.
Другое доказательство связи запоминания с состоянием сна – изменение структуры электроэнцефалографических феноменов, характерных для сна, после обучающей сессии. Наиболее типично последующее увеличение представленности ФБС и второй стадии медленного сна, а также увеличение мощности электроэнцефалографии в спектре веретен сна и тета-активности в быстром сне. В исследованиях с использованием функциональной магнитно-резонансной томографии отдельно и вместе с электроэнцефалографией показано, что изменения, наблюдаемые в мозге в процессе научения в бодрствовании, сохраняются и во время сна. Зависящая от предшествующего обучения реактивация гиппокампа обнаруживается в ФМС, при этом она связана с веретенами сна при тестах как на декларативную, так и на процедурную память. В ФБС нагрузочные тесты процедурной памяти сопровождаются увеличением активности в корковых зонах, вовлеченных в формирование навыка в бодрствовании. Таким образом, другой подход к изучению роли сна в обучении позволяет утверждать, что во время сна происходит повторное воспроизведение (replay) событий, имевших место в бодрствовании, а активность зон мозга, вовлеченных в эти процессы в бодрствовании, сохраняется и во время сна, проявляясь такими фазическими феноменами, как веретена сна. Кроме того, высказывается предположение о том, что обучение вызывает локальные синаптические изменения, которые выражаются усилением свойственной ФМС медленноволновой активности [6, 7].
Зависящая от предшествующего обучения реактивация гиппокампа обнаруживается в ФМС, при этом она связана с веретенами сна при тестах как на декларативную, так и на процедурную память. В ФБС нагрузочные тесты процедурной памяти сопровождаются увеличением активности в корковых зонах, вовлеченных в формирование навыка в бодрствовании. Таким образом, другой подход к изучению роли сна в обучении позволяет утверждать, что во время сна происходит повторное воспроизведение (replay) событий, имевших место в бодрствовании, а активность зон мозга, вовлеченных в эти процессы в бодрствовании, сохраняется и во время сна, проявляясь такими фазическими феноменами, как веретена сна. Кроме того, высказывается предположение о том, что обучение вызывает локальные синаптические изменения, которые выражаются усилением свойственной ФМС медленноволновой активности [6, 7].
Третий подход в изучении роли сна в процессах запоминания – стимуляционный. Установлено, что после заучивания звуков азбуки Морзе воспроизведение таких же звуков в период ФБС увеличивает объем запомненного материала по сравнению с контрольным заучиванием. Важно, чтобы звуковые стимулы во время ФБС подавались одновременно с такими его фазическими элементами, как понтогеникулоокципитальные спайки. В другом исследовании после заучивания на фоне специфического запаха этот же запах подавался испытуемым в ФМС, что также позволило увеличить запоминание. В другом исследовании было достигнуто улучшение декларативной памяти на фоне стимуляции в ФМС магнитным полем частотой 0,75 Гц, что соответствовало базовой частоте медленных волн. Природа этих явлений не ясна, однако результаты опытов подтверждают активную роль сна в процессе запоминания, причем эту функцию можно в определенных пределах улучшить [6].
Важно, чтобы звуковые стимулы во время ФБС подавались одновременно с такими его фазическими элементами, как понтогеникулоокципитальные спайки. В другом исследовании после заучивания на фоне специфического запаха этот же запах подавался испытуемым в ФМС, что также позволило увеличить запоминание. В другом исследовании было достигнуто улучшение декларативной памяти на фоне стимуляции в ФМС магнитным полем частотой 0,75 Гц, что соответствовало базовой частоте медленных волн. Природа этих явлений не ясна, однако результаты опытов подтверждают активную роль сна в процессе запоминания, причем эту функцию можно в определенных пределах улучшить [6].
Отдельно следует остановиться на значении сна в эмоциональном «обрамлении» памяти. Поговорка «утро вечера мудренее» как раз отражает протективную функцию сна, снимающего эмоциональную перегрузку, которая мешает адекватному мышлению. Показано, что в запоминании эмоционально значимого материала важную роль играет ФБС. Эмоционально значимые события лучше сохраняются в памяти после полной депривации сна. В гипотезе M. Walker и E. van der Helm «Сон, чтобы забыть, и сон, чтобы вспомнить» именно быстрому сну уделяется функция «разобщения» эмоционального наполнения события с его содержанием [8]. С каждым последующим циклом сна эта эмоциональная «шелуха» истончается, в итоге в долговременной памяти остается только представление о самом факте, а не о сопровождавших его эмоциях. Таким образом, мозг перерабатывает избыточную аффективную информацию, которая могла бы быть источником эмоциональных нарушений. Нейрохимической базой возможности такого разобщения авторы считают низкий катехоламинергический тонус, характерный именно для этой фазы сна. В качестве доказательства приводятся данные функциональной магнитно-резонансной томографии, демонстрирующие высокий уровень активности в области амигдалы на фоне депривации сна и сохранения в памяти большего количества текстов негативного содержания. Тем не менее роль ФМС в сохранении эмоциональной информации также обсуждается. Известно, что селективные воспоминания именно негативных аспектов заученных комплексных сцен коррелируют с ФМС и мощностью медленноволновой активности.
В гипотезе M. Walker и E. van der Helm «Сон, чтобы забыть, и сон, чтобы вспомнить» именно быстрому сну уделяется функция «разобщения» эмоционального наполнения события с его содержанием [8]. С каждым последующим циклом сна эта эмоциональная «шелуха» истончается, в итоге в долговременной памяти остается только представление о самом факте, а не о сопровождавших его эмоциях. Таким образом, мозг перерабатывает избыточную аффективную информацию, которая могла бы быть источником эмоциональных нарушений. Нейрохимической базой возможности такого разобщения авторы считают низкий катехоламинергический тонус, характерный именно для этой фазы сна. В качестве доказательства приводятся данные функциональной магнитно-резонансной томографии, демонстрирующие высокий уровень активности в области амигдалы на фоне депривации сна и сохранения в памяти большего количества текстов негативного содержания. Тем не менее роль ФМС в сохранении эмоциональной информации также обсуждается. Известно, что селективные воспоминания именно негативных аспектов заученных комплексных сцен коррелируют с ФМС и мощностью медленноволновой активности. А в исследовании 2018 г. было продемонстрировано, что добавление дневных засыпаний (содержащих преимущественно ФМС) существенно улучшает запоминание эмоциональных событий как негативного, так и позитивного содержания [9]. Суммарно представления об относительной важности фаз сна для различных аспектов памяти представлены в таблице.
А в исследовании 2018 г. было продемонстрировано, что добавление дневных засыпаний (содержащих преимущественно ФМС) существенно улучшает запоминание эмоциональных событий как негативного, так и позитивного содержания [9]. Суммарно представления об относительной важности фаз сна для различных аспектов памяти представлены в таблице.
Расстройства дыхания во сне и когнитивные нарушения
Наиболее изучены расстройства когнитивных функций при синдроме обструктивного апноэ сна (СОАС) – симптомокомплексе, развивающемся при множественных (индекс дыхательных расстройств ≥ 5 эпизодов за час сна) остановках дыхания обструктивного характера. Чаще всего СОАС проявляется храпом и дневной сонливостью. СОАС значительно распространен в популяции – по данным наиболее авторитетного Висконсинского когортного исследования, проводившегося в течение 20 лет, частота СОАС средней и тяжелой степени в общей популяции в 1988–1994 гг. составила 9%, а в период 2007–2010 гг. увеличилась до 13% [10].
увеличилась до 13% [10].
Интермиттирующая гипоксия и фрагментация ночного сна – основные патогенетические механизмы этого заболевания. СОАС отличается высокой коморбидностью с сердечно-сосудистыми и церебральными сосудистыми заболеваниями: при наличии этого синдрома риск развития инфаркта миокарда или инсульта увеличивается в два-три раза. К последствиям СОАС относят также фибрилляцию предсердий, инсулинорезистентность, увеличение риска развития опухолей. Данные последних исследований позволяют ассоциировать СОАС и с высоким риском развития нейродегенеративных заболеваний [11].
Когнитивные нарушения – типичные проявления этого заболевания. Многочисленные психологические исследования продемонстрировали нарушения внимания, бдительности, долговременной зрительной и слухоречевой памяти, пространственного и конструктивного праксиса и исполнительных функций. Нарушения памяти обнаруживались у 9% больных, от 2 до 25% имели проблемы с вниманием, а у 15–42% были установлены нарушения исполнительных функций [12].
Степень когнитивных нарушений коррелирует с тяжестью гипоксемии и выраженностью фрагментации сна больных СОАС. Имеются противоречивые данные о наличии или отсутствии у этих больных нарушений речевой функции и кратковременной памяти.
Особенно разрушительны нейрокогнитивные эффекты СОАС в детской популяции. У детей с апноэ ниже школьная успеваемость и показатели интеллекта по сравнению со сверстниками. Выявлена ассоциация степени ухудшения невербального интеллекта детей с СОАС со снижением медленноволновой активности в ФМС.
Данные психологических исследований подкреплены результатами нейровизуализации. Продемонстрировано, что у больных СОАС по сравнению с контрольной группой меньше объем серого вещества в различных областях головного мозга: медиальной префронтальной коре, передней поясной извилине, таламусе, задней латеральной теменной коре и парагиппокампальной области. Кроме этого, при СОАС наблюдается дисфункция белого вещества. Уменьшение соотношения N-ацетиласпартат-креатинина и холина/креатинина, как и абсолютной концентрации этих медиаторов в белом веществе лобных долей, подразумевает аксональную дисфункцию, что способствует разобщению префронтальной коры с другими регионами.
Уменьшение соотношения N-ацетиласпартат-креатинина и холина/креатинина, как и абсолютной концентрации этих медиаторов в белом веществе лобных долей, подразумевает аксональную дисфункцию, что способствует разобщению префронтальной коры с другими регионами.
Непосредственное нейротоксическое действие СОАС, увеличение риска развития острых сосудистых нарушений и ассоциированные иммунные процессы повышают при этом заболевании риск развития клинически оформленных когнитивных расстройств, вплоть до различных форм деменции.
В исследовании A. Spira и соавт. (2008) с участием 448 женщин факторами, ассоциированными с когнитивными нарушениями, были индекс дыхательных расстройств ≥ 30 эпизодов в час, минимальная сатурация ≤ 80% и число сопутствующих апноэ центрального характера. При этом у женщин, имевших аллель АПОе4, риск развития когнитивных нарушений был в пять раз выше. В следующем исследовании на том же материале было показано, что риск развития когнитивных нарушений у женщин с СОАС ассоциирован не с фрагментацией сна, а с интермиттирующей ночной гипоксией [11].
Представление о том, что СОАС может ускорять развитие процесса нейродегенерации, было поддержано работой R. Osorio и соавт. (2015). Ученые установили, что это состояние сопровождается более ранними сроками развития умеренных когнитивных нарушений и болезни Альцгеймера. В крупном исследовании тайваньских ученых пятилетнее наблюдение 1414 пациентов с СОАС и 7070 лиц контрольной группы выявило увеличение риска развития деменции при СОАС в 1,7 раза, причем этот эффект был гендерспецифичным – он наблюдался только у женщин [11]. Тип деменции, ассоциированной с СОАС, в этом исследовании не учитывался.
В контролируемом корейском исследовании, включавшем лиц старше 60 лет с умеренными когнитивными нарушениями, худшее качество сна и большая тяжесть СОАС были связаны с нарушениями речевой функции, что позволило предположить у пациентов лобно-подкорковую сосудистую деменцию. В подобном итальянском исследовании с расстройствами дыхания во сне в большей степени ассоциировалась деменция сосудистого характера, чем другие ее типы [11].
Методом выбора в лечении СОАС средней и тяжелой степени является СиПАП-терапия – создание во время сна постоянного положительного давления воздуха, подаваемого через маску на лице пациента. Метод широко используется с 1981 г. и доказал эффективность в устранении сердечно-сосудистых и гормональных нарушений, свойственных СОАС. Продемонстрировано, что проведение СиПАП-терапии сопровождается улучшением некоторых когнитивных доменов, в частности психомоторных функций, памяти и внимания. При этом были обнаружены и положительные структурные изменения в области лобной доли и гиппокампа. В другом исследовании на фоне СиПАП-терапии наблюдалось улучшение характеристик и белого вещества (уменьшение аксональной дисфункции) [12].
Положительное влияние СиПАП-терапии на когнитивные функции у больных СОАС подразумевает возможность применения этого метода даже при развившихся когнитивных расстройствах. В исследовании A. Troussière и соавт. (2014) у пациентов с болезнью Альцгеймера на фоне трехлетней СиПАП-терапии в меньшей степени ухудшились когнитивные функции по сравнению с группой больных, не проходивших лечение [13]. Такие же данные были получены R. Osorio и соавт. (2015) в когорте пожилых больных из исследования Alzheimer’s Disease Neuroimaging Initiative [14].
Такие же данные были получены R. Osorio и соавт. (2015) в когорте пожилых больных из исследования Alzheimer’s Disease Neuroimaging Initiative [14].
Инсомния и когнитивные нарушения
Инсомния представляет собой наиболее часто встречающееся расстройство сна с распространенностью в общей популяции, оценивающейся в 10%. В критерии диагноза инсомнии входят прежде всего «ночные» симптомы – трудности засыпания, частые пробуждения, ранние утренние пробуждения, ощущение некачественного, невосстанавливающего сна. К «дневным» проявлениям инсомнии относятся ощущение усталости, нарушение социального, семейного, производственного или учебного функционирования, расстройство настроения, дневная сонливость, снижение мотивации, энергичности, инициативности, подверженность ошибкам и несчастным случаям, беспокойство о своем сне и неудовлетворенность им. Когнитивные нарушения в форме нарушения внимания, сосредоточения или запоминания также входят в «дневные» критерии диагноза инсомнии. Кроме наличия одного из «ночных» и одного из «дневных» критериев для постановки диагноза инсомнии требуются достаточные условия и время для сна, а также частота нарушений сна не менее трех раз в неделю [15].
Кроме наличия одного из «ночных» и одного из «дневных» критериев для постановки диагноза инсомнии требуются достаточные условия и время для сна, а также частота нарушений сна не менее трех раз в неделю [15].
Острой или кратковременной называют инсомнию продолжительностью менее трех месяцев, а хронической – более трех месяцев. Такая дихотомия оправдана, поскольку наиболее часто инсомнические нарушения развиваются после воздействия стрессового фактора, связь с которым не может сохраняться дольше трех месяцев после прекращения воздействия. Если нарушение сна присутствует более длительное время, значит, приходят в действие поддерживающие психологические механизмы, такие как дисфункциональные убеждения или условнорефлекторное подкрепление.
Главным патофизиологическим механизмом формирования симптомокомплекса инсомнии считают гиперактивацию, которая может носить как врожденный, так и приобретенный характер. В нейровизуализационных исследованиях E. Nofzinger и соавт. (2004) показано, что у больных хронической инсомнией повышена активность структур «эмоционального мозга» – мезиотемпоральной коры и передней поясной извилины, в то время как в префронтальной коре наблюдается, наоборот, снижение активности [16].
(2004) показано, что у больных хронической инсомнией повышена активность структур «эмоционального мозга» – мезиотемпоральной коры и передней поясной извилины, в то время как в префронтальной коре наблюдается, наоборот, снижение активности [16].
E. Fortier-Brochu и соавт. (2010) в метаанализе обобщили результаты 24 исследований, в которых оценивались когнитивные функции пациентов с инсомнией. Авторы пришли к выводу о том, что при инсомнии ухудшается рабочая и эпизодическая память, перцептуальные процессы и некоторые аспекты исполнительных функций. Эти нарушения незначительно или умеренно выражены. При инсомнии в большей степени страдают функции, связанные с выполнением комплексных заданий, что подразумевает нарушение интегративных связей префронтальной коры с другими отделами мозга [17]. Эти изменения когнитивных функций похожи на таковые, полученные в экспериментальных условиях полной или частичной депривации сна. Однако в исследованиях, где результаты тестов сопоставлялись с данными полисомнографии, оказалось, что подобные нарушения могут присутствовать даже при сохранной структуре и достаточной продолжительности сна. Следовательно, необходимо искать другой, нежели чем депривационный, механизм развития когнитивных нарушений при инсомнии. Это может быть не только сокращение общего времени сна, но и изменение его микроструктуры (увеличение фрагментации) или базовых ритмов, необходимых для процессов консолидации, например основной частоты дельта-ритма [18].
Следовательно, необходимо искать другой, нежели чем депривационный, механизм развития когнитивных нарушений при инсомнии. Это может быть не только сокращение общего времени сна, но и изменение его микроструктуры (увеличение фрагментации) или базовых ритмов, необходимых для процессов консолидации, например основной частоты дельта-ритма [18].
Когнитивные нарушения при инсомнии, в частности в доменах памяти, подтверждаются данными нейровизуализационных исследований. Так, H. Noh и соавт. (2012) обнаружили связь между уменьшением объема гиппокампов и длительностью инсомнии, что коррелировало с ухудшением результатов тестов на вербальное и невербальное запоминание, внимание и лобные функции [19]. В морфометрическом исследовании E. Joo и соавт. (2014) продемонстрировано наличие у больных инсомнией атрофии зоны зубчатой извилины CA3-4-DG и аммонова рога, коррелирующих с показателями Питтсбургского индекса качества сна. Это, с точки зрения авторов, подразумевало нарушение нейрогенеза в условиях фрагментации сна и хронического стрессового состояния [20].
Когнитивные нарушения при инсомнии у здоровых людей с большей вероятностью переходят в клинически очерченные состояния (умеренные когнитивные нарушения или деменцию). В исследовании M. Jelicic и соавт. (2002) при трехлетнем наблюдении отмечалось существенное ускорение развития когнитивных нарушений при жалобах на нарушение сна у пожилых людей [21]. Такие же результаты были получены в более позднем исследовании KORA (2016): за три года частота когнитивных расстройств у пожилых людей, имевших нарушения поддержания сна, выросла на 33% [22].
Патофизиологической базой, объясняющей больший риск развития клинически явных когнитивных расстройств, особенно болезни Альцгеймера, при инсомнии, является сравнительно новая «очистительная» теория предназначения сна. В исследовании L. Xie и соавт. (2013) на оптогенетически модифицированных мышах показано, что при засыпании межнейрональные интерстициальные пространства расширяются и на 60% увеличивается клиренс бета-амилоида – балластного белка. Именно этот белок наиболее часто называется в качестве причины развития болезни Альцгеймера [23]. Постулируется, что нарушение сна приводит к сокращению его длительности и недостаточной очистке мозга от бета-амилоида. В недавних экспериментах было продемонстрировано, что даже одна ночь без сна увеличивает накопление бета-амилоида в гиппокампе на 5% [24].
Гипотетически коррекция инсомнии должна сопровождаться регрессом когнитивных нарушений. Однако возникает проблема негативного влияния снотворных: ГАМКергических препаратов, агонистов бензодиазепиновых рецепторов или центральных блокаторов рецепторов гистамина – на когнитивные функции. Например, у детей с эпилепсией на фоне длительной терапии ухудшались показатели интеллекта [25]. Препараты следующих поколений в меньшей степени влияли на когнитивные функции, что в основном зависело от периода полувыведения, поэтому пальма первенства перешла к бензодиазепиновым гипнотикам. Для обозначения негативных эффектов этих препаратов появились термины «когнитивная» и «поведенческая токсичность». Особенно негативно бензодиазепины влияют на когнитивные функции у пожилых людей, причем затрагиваются практически все модальности. Десятилетнее наблюдение S. Billioti de Gage и соавт. (2014) показало, что при постоянном употреблении бензодиазепинов риск развития болезни Альцгеймера у пожилых людей увеличивается на 31% [26].
Побочные эффекты, возникающие при приеме бензодиазепинов, привели к тому, что в качестве снотворных они были вытеснены небензодиазепиновыми агонистами ГАМКА-рецепторного комплекса – так называемыми Z-препаратами (зопиклоном, залеплоном, золпидемом). Эти средства обладают селективной аффинностью к альфа-1-субъединице ГАМКА-рецепторного комплекса, присутствующей преимущественно в структурах мозга, связанных со сном. Тем не менее даже такое селективное действие может негативно влиять на когнитивные функции. Прием Z-препаратов сопровождался удвоением риска дорожно-транспортных происшествий, а также приводил к уменьшению на следующий день функции внимания, памяти, психомоторной бдительности и равновесия [27].
Перспективным представляется применение при фармакотерапии инсомнии нового класса снотворных – двойных антагонистов орексиновых рецепторов, таких как суворексант. Действие этого препарата основывается на понижении активности одной из основных активирующих систем мозга – орексиновой. При этом реципрокно усиливается влияние сомногенных структур и обеспечивается улучшение засыпания и поддержания сна [27]. В настоящее время в России суворексант зарегистрирован, но в продаже отсутствует.
В качестве метода выбора в лечении хронической инсомнии международные клинические рекомендации предлагают использовать когнитивно-поведенческую терапию (КПТ-И) [28]. Подход направлен на устранение мешающих засыпанию ассоциаций и изменение поведения, связанного со сном. Он доказал эффективность в рандомизированных клинических исследованиях. КПТ-И включает в себя разнообразные техники – краткую КПТ-И, когнитивное реструктурирование, мультимодальную КПТ, релаксационный тренинг, контроль стимуляции, безоценочное осознание и т.д. Кратковременный эффект КПТ-И сравним с таковым снотворных препаратов, а долговременный эффект метода превышает таковой медикаментозных средств. Достигаемое улучшение сна сопровождается улучшением когнитивных функций, что наблюдается даже у лиц с имеющимися умеренными когнитивными нарушениями. В рандомизированном исследовании E. Cassidy-Eagle и соавт. (2018) результатом шестинедельной КПТ-И стало улучшение исполнительных функций у пожилых больных с умеренными когнитивными нарушениями [29]. Внедрение КПТ-И ограничивают недостаток обученных психологов для проведения сеансов и значительная трудоемкость метода. Нам известно лишь одно исследование с применением КПТ в российской популяции [30].
Ввиду ограничений для специфических фармакологических и нефармакологических подходов к лечению инсомнии, особенно в популяции пожилых пациентов и среди лиц, имеющих когнитивные нарушения, актуальным становится поиск методов фармакотерапии инсомнии, более щадящих в когнитивном отношении. Взгляд исследователей и клиницистов обращается к мелатонину – естественному хронобиотическому и снотворному агенту.
Мелатонин вырабатывается в организме человека преимущественно шишковидной железой (80% продукции). Его называют «гормоном темноты», поскольку большая часть (70%) секреции мелатонина приходится на темное время суток, подъем уровня мелатонина в крови начинается с 21:00–22:00. Мелатонин связывается со специфическими рецепторами двух типов, расположенными преимущественно в супрахиазменных ядрах гипоталамуса – «внутренних часах» организма млекопитающих. За счет этого осуществляется стабилизация циркадианного пейсмейкера, обеспечивающего ежесуточное изменение активности как самих «внутренних часов», так и вторичных осцилляторов в мозге и внутренних органах. Мелатонин обладает также собственным слабым снотворным эффектом [27]. Помимо влияния на хронобиологические ритмы мелатонин оказывает и другие действия, важнейшие из которых антиоксидантное и иммуномодулирующее.
Применение препаратов мелатонина сопровождается ускорением засыпания и увеличением общего времени сна. При этом они наиболее эффективны у людей старшего (после 55 лет) возраста. Экзогенный мелатонин разрушается на 90% при первом пассаже через печень, период его полувыведения не превышает 50 минут, что обусловливает важное свойство мелатонина как снотворного – отсутствие негативного влияния на когнитивную сферу (утреннего эффекта «последействия»). В отношении воздействия препаратов мелатонина на когнитивные функции имеются только косвенные данные – при использовании агониста мелатониновых рецепторов рамелтеона (не зарегистрирован в России) было подтверждено улучшение психомоторных функций на фоне улучшения сна [27].
В некоторых клинических руководствах, в частности Британской ассоциации психофармакологии [31], Российского общества сомнологов [32], препараты мелатонина рекомендованы для лечения инсомнии. В рекомендациях Американской академии медицины сна (2018) такие указания отсутствуют [33].
Один из препаратов мелатонина, доступный в России, – Соннован (ЗАО «Канонфарма продакшн»). Препарат рекомендуется назначать при нарушениях сна и десинхронозе. Для лечения инсомнии он принимается в дозе 3 мг (одна таблетка) за 30–40 минут перед сном [34].
Заключение
Подчеркнем основные положения, касающиеся значения полноценного сна в обеспечении функции памяти. Доказано, что сон необходим для обеспечения процессов запоминания. Медленный и быстрый сон играют различную роль в консолидации памяти в зависимости от ее модальности. Медленный сон в большей степени обеспечивает процессы эпизодической и семантической, а быстрый – процедурной и эмоциональной памяти. Нарушения сна в случае сокращения его времени или изменений микроструктуры оказывают негативное влияние на память. Коррекция этих нарушений сопровождается улучшением запоминания и даже позитивными морфологическими изменениями в головном мозге. Возможности фармакотерапии инсомнии применительно к сохранению когнитивных функций ограничены в связи с негативным влиянием на них большинства снотворных препаратов, что особенно важно для пожилых людей. По этой причине приобретают большее значение нефармакологические подходы, такие как КПТ-И, и прием адаптогенов, в частности препаратов мелатонина.
Suunto 5 — Функции — Отслеживание сна
Отслеживание сна
Хороший ночной сон — залог здоровья ума и тела. Часы можно использовать для отслеживания сна и его средней продолжительности.
Наденьте часы перед сном, чтобы Suunto 5 отследили показатели сна с помощью данных акселерометра.
Чтобы отследить сон:
- Перейдите к настройкам часов, прокрутите до пункта Сон и нажмите среднюю кнопку.
Включите функцию Отслеж. сна.
Задайте время отхода ко сну и пробуждения в соответствии с обычным распорядком.
Последний этап определяет период для сна. Этот период часы считают временем вашего сна (когда вы находитесь в постели) и учитывают отдельные его периоды в составе единого сна. Например, если вы встали ночью для того, чтобы выпить воды, весь последующий сон после этого часы учтут, как один период.
ПРИМЕЧАНИЕ:Если лечь спать до указанного периода сна и проснуться после него, то часы не сочтут эти отрезки единым периодом сна. Необходимо задавать период для сна по самому раннему возможному времени отхода ко сну и по самому последнему возможному времени пробуждения.
После включения отслеживания сна можно также задать целевой показатель сна. Обычно человеку нужно от 7 до 9 часов сна в день, хотя индивидуальная оптимальная продолжительность сна может отличаться от нормы.
Тенденции сна
После пробуждения активируется сводка сна. В сводку входит общая продолжительность сна, оценочное время бодрствования (когда определялось движение) и время глубокого сна (без движения).
Кроме сводки о конкретной ночи, можно также посмотреть общий тренд с анализом сна. Когда часы находятся в представлении циферблата, держите нижнюю правую кнопку нажатой до появления экрана Сон. В первом представлении последний сон сравнивается с целевым показателем сна.
Находясь на экране сна, нажмите среднюю кнопку, чтобы просмотреть средние данные о сне за последние семь дней. Нажмите нижнюю правую кнопку, чтобы просмотреть фактическую продолжительность сна в часах за последние семь дней.
Находясь на экране средней продолжительности сна, нажмите среднюю кнопку, чтобы просмотреть график средних значений ЧСС за последние семь дней.
Нажмите нижнюю правую кнопку, чтобы просмотреть фактические значения ЧСС за последние семь дней.
ПРИМЕЧАНИЕ:Находясь на экране Сон, держите нажатой среднюю кнопку, чтобы перейти к параметрам отслеживания сна.
ПРИМЕЧАНИЕ:Все измерения сна основаны только на движениях, поэтому являются приблизительными и могут не отражать реальных привычек сна.
Качество сна
Помимо продолжительности часы также оценивают качество сна, отслеживая изменение пульса во сне. Эти изменения показывают, насколько эффективно сон помогает вам отдохнуть и восстановиться. Качество сна отображается на шкале от 0 до 100 сводки сна, где 100 — самое высокое качество.
Измерение пульса во сне
Если на ночь не снимать часы, то можно получить дополнительные данные о частоте сердечных сокращений во время сна. Чтобы оптический датчик частоты сердечных сокращений работал в течение всей ночи, включите функцию ежедневного отслеживания ЧСС (см. Суточная ЧСС).
Автоматический режим «Не беспокоить»
Автоматическое включение режима «Не беспокоить» также можно использовать во время сна.
Suunto Spartan Sport — Функции
Отслеживание сна
Хороший сон ночью — залог здоровья ума и тела. Часы можно использовать для отслеживания сна и его средней продолжительности.
Во время сна часы Suunto Spartan Sport отслеживают сон на основе данных акселерометра.
Чтобы отследить сон:
- Перейдите в настройки часов, прокруткой перейдите к Сон и нажмите среднюю кнопку.
Включите функцию Отслеж. сна.
Задайте время отхода ко сну и пробуждения в соответствии с обычным распорядком.
Последний этап определяет период для сна. Часты используют этот период для определения состояния сна и учитывают все отдельные периоды сна как единый. Например, если ночью встать попить воды, весь последующий сон после этого часы все равно учтут как один период.
ПРИМЕЧАНИЕ:Если ложиться спать до указанного периода сна, а просыпаться после него, часы не будут учитывать эти отрезки как период сна. Необходимо задавать период для сна по самому раннему возможному времени отхода ко сну и по самому последнему возможному времени пробуждения.
После включения отслеживания сна, можно также задать целевой показатель сна. Обычно человеку нужно от 7 до 9 часов сна в день, хотя индивидуальная оптимальная продолжительность сна может отличаться от нормы.
Тренды сна
После пробуждения активируется сводка сна. В сводку входит общая продолжительность сна, оценочное время бодрствования (когда определялось движение) и время глубокого сна (без движения).
Кроме сводки о конкретной ночи можно также посмотреть общий тренд с анализом сна. Когда часы находятся в представлении циферблата, держите нижнюю правую кнопку нажатой до появления экрана Сон. В первом представлении последний сон сравнивается с целевым показателем сна.
Находясь на экране сна, проведите влево, чтобы просмотреть среднюю продолжительность сна за последние семь дней. Проведите вверх, чтобы просмотреть фактическую продолжительность сна в часах за последние семь дней.
Находясь на экране Сон, держите нажатой среднюю кнопку, чтобы перейти к параметрам отслеживания сна и сводке по последнему сну.
ПРИМЕЧАНИЕ:Все измерения сна основаны только на движениях, поэтому являются приблизительными и могут не отражать реальных привычек сна.
Автоматический режим «Не беспокоить»
Можно настроить режим «Не беспокоить», чтобы часы автоматически включали его во время сна.
Функции сна | Дистанционные уроки
14-Янв-2015 | Нет комментариев | Лолита Окольнова
Человек спит в среднем одну треть своей жизни
Т.е. если он прожил 75 лет, то 25 из них – спал!
Какие функции выполняет сон?
Автор статьи — Мария Щербакова
Сон – это прекрасное состояние дремы и грез, столь любимое многими из нас, без которого жизнь невозможна.
Крепкий, сладкий, здоровый сон – залог здоровья.
Известно, что если человеку не давать спать, то он погибает через 4-6 суток (очень жестокая пытка, кстати).
Сон — естественное физиологическое состояние организма, процесс пребывания в состоянии с минимальным уровнем активности головного мозга и сниженной реакцией на окружающий мир.
Однако, ученые доказали, что хотя сон и необходим человеку именно для отдыха мозга, но во время сна мозг человека работает еще активнее и продуктивнее, чем во время бодрствования, когда человек взаимодействует со внешним миром.
Очень интересны в своем изучении теории сна: функции сна, его значение для организма человека вообще и нервной системы в частности.
- первая достаточно старая, но забавная теория —
сон – это результат застоя крови в головном мозге человека, когда тот принимает горизонтальное положение
во время бодрствования в клетках организма человека накапливаются окисляющиеся вещества и сон возникает в результате дефицита кислорода
- и уже современная, научная — И.П. Павлов рассматривал сон как единый процесс с внутренним торможением — когда человек бодрствует, то торможение охватывает только отдельные участки его организма, тогда как другие активны.
Когда человек спит, торможение широко распространяется по коре больших полушарий, что обеспечивает восстановление мозга и других структур организма для дальнейшей деятельности.
В современной науке рассматривают три основные функции сна:
1. Энергетическая — во время сна организм восстанавливает потраченную за время бодрствования энергию и накапливает ее для будущей активности.
2. Информационная — во время сна уменьшается, ограничивается восприятие информации органами чувств.
Здесь же есть такая точка зрения, что во сне нуждаются не органы и клетки, а психические функции: восприятие, память, мышление.
Это происходит тогда, когда мозг человека переполнен информацией, и ему необходимо отключиться, что бы «переварить» и «записать» уже имеющуюся информацию мира.
3. Психическая функция сна — существует два типа бодрствования: спокойное и напряженное. Напряженное бодрствование необходимо для целенаправленных и координированных реакций. Так же и во время сна. Во время быстрой фазы — аналога напряженного бодрствования — работает лимбическая система, когда эмоции активны, а координированные реакции пассивны — именно в это время человек видит сны, может подергиваться, совершать какие-то движение, волноваться и т.д.
Медленный сон — мышечная активность снижается, снижается частота дыхания и пульса, замедляется обмен веществ, и понижается температура, глаза могут совершать медленные движения.
Сон – это прежде активный физиологический процесс, никак не пассивный.
Во время сна активные связи человека с окружающим миром прерваны, происходит процесс обработки информации, накопления ресурсов. Организм отдыхает и работает одновременно, отдыхает для нового дня, набирается сил, и работает по «переработке» уже прошедшего дня.
Ученые доказали, что в процессе сна закрепляется долговременная память, причем сознательная память (события и факты) закрепляется в фазе медленного сна, а бессознательная (эмоции, движения, мимика) в фазе быстрого сна. Так что учить экзамен в одну-две-три ночи, может и действенно, но не долговременно. Тут количество запомненной информации не переходит в качество. Как правило, человек все забывает через три дня. Гораздо эффективнее растянуть этот процесс на недели.
В заключение хочется отметить, что существуют негласные соревнования: кто дольше сможет не спать. В «Книге рекордов Гиннеса» есть достижение американца, который не спал 19 суток. Кто-то продержался еще дольше, кто-то чуть меньше, говорят есть даже люди, которые могут вообще не спать и наслаждаться жизнью при этом. Однако все это очень спорно.
Одно совершенно ясно – польза сна для функционирования организма человека неоспорима!
Когда человек долго не спит, повышается его раздражительность, растет общее недовольство миром, снижается скорость реакции, как двигательной, так и ментальной. Это все происходит в первые сутки, у кого-то быстрее, у кого-то чуть медленнее. Далее сознание начинается путаться, так как перестраивается гормональный фон человека, возможны галлюцинации и даже психоз. Если человек и дальше упорствует в не засыпании – тот тут два пути: или его «срубит» и он уснет, или он умрет.
Потребность во сне у всех разная, но сон, как залог здоровья ЦНС и всего организма в целом, нам необходим!
Еще на эту тему:
Обсуждение: «Функции сна»
(Правила комментирования)Мониторинг данных сна | HUAWEI поддержка россия
Сбор статистики сна
Если вы не снимаете умный браслет на ночь, он регистрирует данные сна и определяет статус вашего сна. Устройство автоматически регистрирует время засыпания и пробуждения, а также определяет фазу сна. Вы можете синхронизировать устройство с телефоном через приложение Здоровье (Huawei Health) и просматривать данные подробные сна этом приложении.- Если вы спите в течение дня менее 3 часов, то устройство регистрирует этот сон как дневной сон (перерыв на сон). Если дневной сон длился менее 30 минут или вы много двигались во время сна, часы могут не распознать сон и не записать данные сна.
- Данные сна на часах обновляются ежедневно в полночь. Архивные данные хранятся на ваших часах. Регулярно синхронизируйте данные с приложением Здоровье (Huawei Health), чтобы архивные данные не стирались.
HUAWEI TruSleep™
Откройте приложение Здоровье (Huawei Health), выберите раздел Устройства, затем имя вашего устройства, перейдите в раздел , а затем включите функцию мониторинга сна HUAWEI TruSleep™.
После включения функции мониторинга сна HUAWEI TruSleep™ ваши часы будут собирать данные сна, распознавать время засыпания, просыпания, распознавать фазу легкого, глубокого или быстрого сна, определять количество просыпаний и качество сна, чтобы предоставить анализ сна и помочь вам улучшить его качество.
- Если вы не двигаетесь, часы могут не распознать, что вы проснулись. В этом случае нажмите на кнопку питания, чтобы включить экран и выйти из режима сна.
- После включения функции HUAWEI TruSleep™ индикатор на задней панели будет гореть зеленым. Часы записывают данные активности с помощью акселерометра и используют функцию распознавания движения и алгоритмы сна, чтобы определять статус сна. HUAWEI TruSleep™ способствует более высокому энергопотреблению. Перед началом сна заряд батареи должен составлять не менее 30%.
- После включения режима Не беспокоить часы выключают трафик данных, функцию постоянного отображения экрана, уведомления, уведомления о вызовах и функцию поднятия запястья для активации, чтобы вас во время сна не беспокоили уведомления о сообщениях или входящих вызовах. Для включения этой функции нажмите на кнопку питания, откройте карточку Сон и проведите влево, чтобы включить режим Не беспокоить.
Просмотр статистики сна
Период сбора статистики составляет 24 часа: с 20:00 первого дня до 20:00 второго дня. Если вы спите 11 часов с 19:00 до 6:00, часы засчитают длительность сна до 20:00 первого дня, а также оставшееся время сна в течение второго дня.
Чтобы посмотреть данные сна, воспользуйтесь одним из следующих способов.
- Чтобы посмотреть данные сна на ваших часах за текущий день, нажмите на верхнюю кнопку, чтобы открыть список приложений. Выберите Сон и проведите вверх по экрану, чтобы посмотреть длительность ночного сна и количество прерываний на дневной сон.
- Чтобы посмотреть архивные данные сна в приложении Здоровье (Huawei Health), нажмите на карточку Сон на рабочем экране приложения, чтобы посмотреть данные сна за день, неделю, месяц или год.
Хронопатологические аспекты расстройств сна и когнитивных функций у детей с нарушениями зрения | Кельмансон
1. Хильдебрандт Г., Мозер М., Лехофер М. Хронобиология и хрономедицина: Пер. с нем. М: Арнебия 2006; 144. (Hildebrandt G., Moser M., Lehofer M. Chronobiology and chronomedicine: Translated from German. Moscow: Arnebia 2006; 144.)
2. Swaab D.F. The human hypothalamus : basic and clinical aspects. 1st ed. Amsterdam; Boston: Elsevier, 2003; 1:476; 2: 588.
3. Barinaga M. Circadian clock. How the brain’s clock gets daily enlightenment. Science 2002; 295: 5557: 955-957.
4. Dijk D.J., Archer S.N. Light, sleep, and circadian rhythms:
5. together again. PLoS biology 2009; 7: 6: el000145.
6. Thapan K., Arendt J., SkeneD.J. An action spectrum for mela-tonin suppression: evidence for a novel non-rod, non-cone photoreceptor system in humans. J Physiol 2001; 535: Pt 1: 261-267.
7. ReidKJ., Zee P.C. Circadian rhythm disorders. SeminNeurol 2009; 29: 4: 393-405.
8. Khalsa S.B., Jewett M.E., Cajochen C, Czeisler C.A. A phase response curve to single bright light pulses in human subjects. J Physiol 2003; 549: Pt 3: 945-952.
9. Valdez P., Ramirez С., Garcia A. Orcadian rhythms in cognitive pprocesses: implications for school learning. Mind Brain Educ 2014; 8: 4: 161-168.
10. BergerA., Posner M.I. Pathologies of brain attentional networks. Neuroscience andbiobehavioral reviews 2000; 24: 1: 3—5.
11. Sorter M., Givens В., Bruno J.P. The cognitive neuroscience of sustained attention: where top-down meets bottom-up. Brain research reviews 2001; 35: 2: 146-160.
12. Carrier J., Monk Т.Н. Orcadian rhythms of performance: new trends. Chronobiology international 2000; 17: 6: 719—732.
13. Кельмансон И.А. Интеллектуальные нарушения и расстройства сна у детей. Врач 2015; 1: 2—5. (Kelmanson LA. Inttelectual disorders and sleep dirturbances in children. Vrach3015; 1:2-5.)
14. Кельмансон И.А. Эмоциональные расстройства и расстройства поведения у детей, связанные с нарушениями сна. Рос вестн перинатол и педиат 2014; 59: 4: 32—40. (Kelmanson LA. Emotional disorders and behavioural disorders related to sleep disturbances in children. Ros vestn peri-natol ipediat 2014; 59: 4: 32-40.)
15. Rogers N.L., Dorrian J., Binges D.F. Sleep, waking and neu-robehavioural performance. Frontiers in bioscience 2003; 8: 1056-1067.
16. Wright K, Hull J.T., Hughes R.J. et al. Sleep and wakefulness out of phase with internal biological time impairs learning in humans. J Cognit Neurosci 2006; 18: 4: 508-521.
17. SaperC.B., Chou T.C., Scammell Т.Е. The sleep switch: hypo-thalamic control of sleep and wakefulness. Trends in neurosci-ences 2001; 24: 12: 726-731.
18. Deurveilher S., Semba К Indirect projections from the supra-chiasmatic nucleus to major arousal-promoting cell groups in rat: implications for the circadian control of behavioural state. Neurosci 2005; 130: 1: 165-183.
19. Tu J., Jhou T.C., SaperC.B. Identification of wake-active do-paminergic neurons in the ventral periaqueductal gray matter. J Neurosci 2006; 26: 1: 193-202.
20. Steriade M. The corticothalamic system in sleep. Frontiers in bioscience 2003; 8: d878-899.
21. Aston-Jones G. Brain structures and receptors involved in alertness. Sleep medicine 2005; 6: Suppl 1: S3—7.
22. Larkin J.E., Yokogawa T, Heller H.С. et al. Homeostatic regulation of sleep in arrhythmic Siberian hamsters. Am J Physiol Regulat Integrat Comparat Physiol 2004; 287: 1: R104-111.
23. Dijk D.J., Czeisler C.A. Paradoxical timing of the circadian rhythm of sleep propensity serves to consolidate sleep and wakefulness in humans. Neurosci letters 1994; 166: 1: 63—68.
24. Folkard S., Totterdell P., Minors D., Waterhouse J. Dissecting circadian performance rhythms: implications for shiftwork. Ergonomics 1993; 36: 1-3: 283-288.
25. Eckel-Mahan K.L., Storm D.R. Circadian rhythms and memory: not so simple as cogs and gears. EMBO reports 2009; 10: 6:584-591.
26. Swaab D.F. We are our brains : a neurobiography of the brain, from the womb to Alzheimer’s. New York: Spiegel&Grau, 2014; 417.
27. Belichenko P.V., Masliah E., Kleschevnikov A.M. et al. Syn-aptic structural abnormalities in the Ts65Dn mouse model of Down Syndrome. J Comparat Neurol 2004; 480: 3: 281-298.
28. Fernandez F, Morishita W., Zuniga E. et al. Pharmacotherapy for cognitive impairment in a mouse model of Down syndrome. Nat Neurosci 2007; 10: 4: 411-413.
29. Ruby N.F., Hwang C.E., Wessells C. et al. Hippocampal-de-pendent learning requires a functional circadian system. Proc NatAcadSci USA 2008; 105:40: 15593-15598.
30. Cohen R.A., Barnes H.J., Jenkins M., Albers H.E. Disruption of short-duration timing associated with damage to the supra-chiasmatic region of the hypothalamus. Neurology 1997; 48: 6:1533-1539.
31. Ohta H., Yamazaki S., McMahon D.G. Constant light desyn-chronizes mammalian clock neurons. Nat Neurosci 2005; 8: 3: 267-269.
32. Devon B.D., Goad E.H., Petri HE. et al. Circadian phase-shifted rats show normal acquisition but impaired long-term retention of place information in the water task. Neurobiol Learn Mem 2001; 75: 1:51-62.
33. Lament E.W., Robinson В., Stewart J., Amir S. The central and basolateral nuclei of the amygdala exhibit opposite diurnal rhythms of expression of the clock protein Period2. Proc Nat AcadSci USA 2005; 102: 11:4180-4184.
34. JilgA., Lesny S., PeruzkiN. et al. Temporal dynamics of mouse hippocampal clock gene expression support memory processing. Hippocampus 2010; 20: 3: 377-388.
35. Hampp G., Albrecht U. The circadian clock and mood-related behavior. Communicat Integrat Biol 2008; 1: 1: 1—3.
36. Van derZee E.A., Havekes R., BarfR.P. et al. Circadian time-place learning in mice depends on Cry genes. Curr Biol 2008; 18: 11: 844-848.
37. Guilding C, Piggins H.D. Challenging the omnipotence of the suprachiasmatic timekeeper: are circadian oscillators present throughout the mammalian brain? Eur J Neurosci 2007; 25: 11:3195-3216.
38. Hampp G., Ripperger J.A., Houben T. et al. Regulation of monoamine oxidase A by circadian-clock components implies clock influence on mood. Curr Biol 2008; 18: 9: 678-683.
39. Gonzalez M.M., Aston-Jones G. Light deprivation damages monoamine neurons and produces a depressive behavioral phenotype in rats. Proc Nat Acad Sci USA 2008; 105: 12: 4898-4903.
40. Monje F.J., Cabatic M., Divisch I. et al. Constant darkness induces IL-6-dependent depression-like behavior through the NF-kappaB signaling pathway. J Neurosci 2011; 31: 25: 9075-9083.
41. Lockley S.W., Dijk D.J., Kosti O. et al. Alertness, mood and performance rhythm disturbances associated with circadian sleep disorders in the blind. J Sleep Res 2008; 17: 2: 207-216.
42. American Academy of Sleep Medicine. The international classification of sleep disorders : diagnostic and coding manual. 2nd ed. Westchester, 111.: American Academy of Sleep Medicine, 2005; 297.
43. Sack R.L., Auckley D., Auger R.R. Circadian rhythm sleep disorders: part II, advanced sleep phase disorder, delayed sleep phase disorder, free-running disorder, and irregular sleep-wake rhythm. An American Academy of Sleep Medicine review. Sleep 2007; 30: 11: 1484-1501.
44. Shochat T, Martin J., MarlerM., Ancoli-Israel S. Illumination levels in nursing home patients: effects on sleep and activity rhythms. J Sleep Res 2000; 9: 4: 373-379.
45. Antoniadis E.A., Ко С.Н., Ralph M.R., McDonald R.J. Circadian rhythms, aging and memory. Behav Brain Res 2000; 114: 1-2: 221-233.
46. Gais S., Born J. Declarative memory consolidation: mechanisms acting during human sleep. Learn Memory 2004; 11:6: 679-685.
47. Cho К Chronic ‘jet lag’ produces temporal lobe atrophy and spatial cognitive deficits. Nat Neurosci 2001; 4: 6: 567—568.
48. Park H.J., Lee J.D., Kim E. Y et al. Morphological alterations in the congenital blind based on the analysis of cortical thickness and surface area. Neurolmage 2009; 47: 1: 98—106.
49. Lepore N., Shi Y, Lepore F. et al. Pattern of hippocampal shape and volume differences in blind subjects. Neurolmage 2009; 46: 4: 949-957.
50. Солнцева Л.И. Тифлопсихология детства. М: Полиграф сервис 2000; 250. (Solntseva L.I. Typhlopsychology of childhood. Moscow: Poligraph service 2000; 250.)
51. Zaidi F.H, Hull J.T., Peirson S.N. et al. Short-wavelength light sensitivity of circadian, pupillary, and visual awareness in humans lacking an outer retina. Curr Biol 2007; 17: 24: 2122-2128.
52. Shirani A., St Louis E.K. Illuminating rationale and uses for light therapy. JCSM2009; 5: 2: 155-163.
53. Gronfler C, Wright K.P., Kronauer R.E. et al. Efficacy of a single sequence of intermittent bright light pulses for delaying circadian phase in humans. Am J Physiol Endocrinol Metabol 2004; 287: 1: E174-181.
54. Yamadera H., Ito Т., Suzuki H. et al. Effects of bright light on cognitive and sleep-wake (circadian) rhythm disturbances in Alzheimer-type dementia. Psychiat Clinic Neurosci 2000; 54: 3: 352-353.
55. Lewy A.J., Bauer V.K., Hosier B.P. et al. Capturing the circadian rhythms of free-running blind people with 0.5 mg melatonin. Brain Res 2001; 918: 1-2: 96-100.
56. Lockley S. W., Skene D.J., James K. et al. Melatonin administration can entrain the free-running circadian system of blind subjects. J Endocrinol 2000; 164: 1: Rl-6.
57. Sack R.L., Brandes R.W., Kendall A.R., Lewy A.J. Entrain-ment of free-running circadian rhythms by melatonin in blind people. New Engl J Med 2000; 343: 15: 1070-1077.
58. Lewy A.J., Emens J.S., Lefler B.J. et al. Melatonin entrains free-running blind people according to a physiological dose-response curve. Chronobiol Intern 2005; 22: 6: 1093-1106.
59. Asayama K, Yamadera H., Ito T. et al. Double blind study of melatonin effects on the sleep-wake rhythm, cognitive and non-cognitive functions in Alzheimer type dementia. J Nippon Med School 2003; 70: 4: 334-341.
60. Riemersma-van derLekR.F., Swaab D.F., TwiskJ. et al. Effect of bright light and melatonin on cognitive and noncognitive function in elderly residents of group care facilities: a randomized controlled trial. JAMA 2008; 299: 22: 2642-2655.
61. Van Someren E.J.W., Riemersma R.F., Swaab D.F. Functional plasticity of the circadian timing system in old age: light exposure. In: M.A. Hofman, G.J. Boer, A.J. Holtmaat et al. (eds). Progress in Brain Research. Amsterdam: Elsevier Science B.V. 2002; 138: 205-231.
Изучение филогении для определения функции сна
Жданова И.В., Ван С.Ю., Леклер О.У. и Данилова Н.П. Мелатонин способствует состоянию сна у рыбок данио. Brain Res. 903 , 263–268 (2001).
CAS PubMed Google Scholar
Тоблер И. Влияние вынужденного передвижения на цикл покой – активности таракана. Behav. Brain Res. 8 , 351–360 (1983).
CAS PubMed Google Scholar
Райзен, Д. М. и др. Летаргус Caenorhabditis elegans состояние сна. Природа 451 , 569–572 (2008).
CAS PubMed Google Scholar
Форстер, А. П., Кришнан, Х. К., Чирелли, К. и Лайонс, Л. С. Характеристика сна у Aplysia californica . Sleep 37 , 1453–1463 (2014).
PubMed PubMed Central Google Scholar
Omond, S. et al. Отсутствие активности является никемерным, эндогенно генерируемым, гомеостатически регулируемым и модулируемым мелатонином у свободноживущего платигельминта плоского червя. Спящий режим 40 , zsx124 (2017).
Google Scholar
Нат, Р.D. et al. Медуза Cassiopea находится в состоянии сна. Curr. Биол. 27 , 2984–2990 (2017).
CAS PubMed PubMed Central Google Scholar
Уокер, Дж. М. и Бергер, Р. Дж. Сон как приспособление для сохранения энергии, функционально связанное с гибернацией и поверхностным оцепенением. Прог. Brain Res. 53 , 255–278 (1980).
CAS PubMed Google Scholar
Ту, Б. П. и Макнайт, С. Л. Метаболические циклы как основа биологических колебаний. Nat. Rev. Mol. Cell Biol. 7 , 696–701 (2006).
CAS PubMed Google Scholar
Ту, Б. П. и Макнайт, С. Л. Метаболический цикл дрожжей: понимание жизни эукариотической клетки. Колд Спринг Харб. Symp. Quant. Биол. 72 , 339–343 (2007).
CAS PubMed Google Scholar
Шмидт, М. Х. Функция распределения энергии сна: объединяющая теория сна, оцепенения и непрерывного бодрствования. Neurosci. Biobehav. Ред. 47 , 122–153 (2014).
PubMed Google Scholar
Бакстон, О. М. и др. Неблагоприятные метаболические последствия длительного ограничения сна в сочетании с нарушением циркадных ритмов у людей. Sci. Transl Med. 4 , 129ra43 (2012).
PubMed PubMed Central Google Scholar
Ван Каутер, Э., Шпигель, К., Тасали, Э. и Лепроулт, Р. Метаболические последствия нарушения сна и недосыпания. Sleep Med. 9 , S23 – S28 (2008).
PubMed PubMed Central Google Scholar
Недельчева А. В. и Шеер Ф. А. Метаболические эффекты нарушения сна, связь с ожирением и диабетом. Curr. Opin. Эндокринол. Диабет Ожирение. 21 , 293–298 (2014).
PubMed PubMed Central Google Scholar
Кэмпбелл С. и Тоблер И. Сон животных: обзор продолжительности сна в филогенезе. Neurosci. Biobehav. Ред. 8 , 269–300 (1984).
CAS PubMed Google Scholar
Franken, P., Chollet, D. & Tafti, M. Гомеостатическое регулирование потребности во сне находится под генетическим контролем. J. Neurosci. 21 , 2610–2621 (2001).
CAS PubMed Google Scholar
Шоу, П. Дж., Сирелли, К., Гринспен, Р. Дж. И Тонони, Г. Корреляты сна и бодрствования у Drosophila melanogaster . Наука 287 , 1834–1837 (2000).
CAS PubMed Google Scholar
Hendricks, J. C. et al. Отдых в Drosophila — состояние, подобное сну. Нейрон 25 , 129–138 (2000).
CAS PubMed Google Scholar
Тоблер И. И. и Нойнер-Йеле М. 24-часовой вариант бдительности у таракана Blaberus giganteus . J. Sleep Res. 1 , 231–239 (1992).
CAS PubMed Google Scholar
Кайзер В. и Штайнер-Кайзер Дж. Нейрональные корреляты сна, бодрствования и возбуждения у дневных насекомых. Nature 301 , 707–709 (1983).
CAS PubMed Google Scholar
Сингх Р. Н. и Салсстон Дж. Э. Некоторые наблюдения линьки у Caenorhabditis elegans . Nematologica 24 , 63–71 (1978).
Google Scholar
Cassada, R.C. & Russell, R.L. Дауэрларва, постэмбриональный вариант развития нематоды Caenorhabditis elegans . Dev. Биол. 46 , 326–342 (1975).
CAS PubMed Google Scholar
Singh, K., Ju, J. Y., Walsh, M. B., DiIorio, M. A. & Hart, A. C. Глубокая консервация генов, необходимая как для сна Drosphila melanogaster , так и Caenorhabditis elegans , играет роль дофаминергической передачи сигналов. Sleep 37 , 1439–1451 (2014).
PubMed PubMed Central Google Scholar
Шварц Дж., Левандровски И. и Брингманн Х. Снижение активности сенсорного нейрона во время сна у Caenorhabditis elegans . Curr. Биол. 21 , R983 – R984 (2011).
CAS PubMed Google Scholar
Nagy, S. et al. Гомеостаз сна C. elegans характеризуется двумя поведенческими и генетически различными механизмами. eLife 3 , e04380 (2014).
PubMed PubMed Central Google Scholar
Драйвер, Р. Дж., Лэмб, А. Л., Виннер, А. Дж. И Райзен, Д. М. DAF-16 / FOXO регулирует гомеостаз существенного поведения, подобного сну, во время личиночных переходов у C. elegans . Curr. Биол. 23 , 501–506 (2013).
CAS PubMed PubMed Central Google Scholar
Кайзер, М. С. и Бирон, Д. Сон и развитие генетически поддающихся лечению модельных организмов. Генетика 203 , 21–33 (2016).
CAS PubMed PubMed Central Google Scholar
Трояновский Н.Ф. и Райзен Д. М. Назовите это червячным сном. Trends Neurosci. 39 , 54–62 (2016).
CAS PubMed Google Scholar
Саттерли Р. А. Есть ли у медуз центральная нервная система? J. Exp. Биол. 214 , 1215–1223 (2011).
PubMed Google Scholar
Dzirasa, K. et al. Дофаминергический контроль состояний сна и бодрствования. J. Neurosci. 26 , 10577–10589 (2006).
CAS PubMed Google Scholar
Куме, К., Куме, С., Парк, С. К., Хирш, Дж. И Джексон, Ф. Р. Допамин является регулятором возбуждения у плодовой мухи. J. Neurosci. 25 , 7377–7384 (2005).
CAS PubMed Google Scholar
Андретич, Р., ван Суиндерен, Б.И Гринспен, Р. Дж. Допаминергическая модуляция возбуждения у Drosophila. Curr. Биол. 15 , 1165–1175 (2005).
CAS PubMed Google Scholar
Turek, M., Besseling, J., Spies, J.P., Konig, S. & Bringmann, H. Спецификация активных во сне нейронов и индукция сна требуют нейропептидов FLP-11 для системной индукции сна. eLife 5 , e12499 (2016).
PubMed PubMed Central Google Scholar
Nelson, M. D. et al. FMRFamide-подобные нейропептиды FLP-13 способствуют покою после теплового стресса у Caenorhabditis elegans . Curr. Биол. 24 , 2406–2410 (2014).
CAS PubMed PubMed Central Google Scholar
Нат, Р. Д., Чоу, Э. С., Ван, Х., Шварц, Э. М. и Штернберг, П. В. C. elegans вызванный стрессом сон возникает в результате коллективного действия нескольких нейропептидов. Curr. Биол. 26 , 2446–2455 (2016).
CAS PubMed PubMed Central Google Scholar
Shang, Y. et al. Короткий нейропептид F — это стимулирующий сон модулятор ингибирования. Нейрон 80 , 171–183 (2013).
CAS PubMed PubMed Central Google Scholar
Ленц, О., Сюн, Дж., Нельсон, М.D., Raizen, D. M. & Williams, J. A. Передача сигналов FMRFamide способствует сну, вызванному стрессом, у Drosophila. Brain Behav. Иммун. 47 , 141–148 (2015).
CAS PubMed Google Scholar
Lee, D. A. et al. Генетическая и нейрональная регуляция сна нейропептидом VF. eLife 6 , e25727 (2017).
PubMed PubMed Central Google Scholar
Deregnaucourt, S., Mitra, P.P., Feher, O., Pytte, C. & Tchernichovski, O. Как сон влияет на развитие пения птиц. Природа 433 , 710–716 (2005).
CAS PubMed Google Scholar
Хендрикс, Дж. К., Кирк, Д., Панкери, К., Миллер, М. С. и Пак, А. И. Модафинил поддерживает бодрствование у плодовой мухи Drosophila melanogaster . Sleep 26 , 139–146 (2003).
PubMed Google Scholar
Panckeri, K. A., Schotland, H. M., Pack, A. I. & Hendricks, J. C. Modafinil снижает гиперсонливость у английского бульдога, естественной животной модели нарушения дыхания во сне. Sleep 19 , 626–631 (1996).
CAS PubMed Google Scholar
Rihel, J. et al. Профилирование поведения рыбок данио связывает лекарства с биологическими мишенями и регуляцией покоя / бодрствования. Наука 327 , 348–351 (2010).
CAS PubMed PubMed Central Google Scholar
Foltenyi, K., Greenspan, R.J. & Newport, J. W. Активация EGFR и ERK с помощью ромбовидной передачи сигналов регулирует консолидацию и поддержание сна у Drosophila . Nat. Neurosci. 10 , 1160–1167 (2007).
CAS PubMed Google Scholar
Кушиката, Т., Фанг, Дж., Чен, З., Ван, Ю. и Крюгер, Дж. М. Эпидермальный фактор роста усиливает спонтанный сон у кроликов. г. J. Physiol. 275 , R509 – R514 (1998).
CAS PubMed Google Scholar
Kramer, A. et al. Регуляция ежедневной двигательной активности и сна с помощью передачи сигналов рецептора EGF гипоталамуса. Наука 294 , 2511–2515 (2001).
CAS PubMed Google Scholar
Van Buskirk, C. & Sternberg, P. W. Передача сигналов эпидермального фактора роста индуцирует поведенческое спокойствие у Caenorhabditis elegans . Nat. Neurosci. 10 , 1300–1307 (2007).
PubMed Google Scholar
Cirelli, C. & Tononi, G. Различия в экспрессии генов мозга между сном и бодрствованием, выявленные с помощью дифференциального отображения мРНК и технологии микрочипов кДНК. J. Sleep Res. 8 , S44 – S52 (1999).
Google Scholar
Naidoo, N., Giang, W., Galante, R.J. и Pack, A. I. Депривация сна вызывает ответ развернутого белка в коре головного мозга мыши. J. Neurochem. 92 , 1150–1157 (2005).
CAS PubMed Google Scholar
Джонс, С., Пфистер-Генскоу, М., Бенка, Р.М.& Cirelli, C. Молекулярные корреляты сна и бодрствования в мозгу белоголового воробья. J. Neurochem. 105 , 46–62 (2008).
CAS PubMed Google Scholar
Сандерс, Дж., Шольц, М., Мерутка, И. и Бирон, Д. Отчетливые развернутые белковые реакции смягчают или опосредуют эффекты несмертельной депривации сна C. elegans в различных тканях. BMC Biol. 15 , 67 (2017).
PubMed PubMed Central Google Scholar
Юргель, М. Э., Масек, П., ДиАнджело, Дж. И Кин, А. С. Генетическое вскрытие взаимодействий сна и метаболизма у плодовой мухи. J. Comp. Physiol. А 201 , 869–877 (2015).
CAS Google Scholar
Сегнет, Л., Галвин, Дж. Э., Сузуки, Ю., Готтшалк, Л. и Шоу, П. Дж. Стойкие дефекты кратковременной памяти после лишения сна в модели болезни Паркинсона Drosophila . Sleep 32 , 984–992 (2009).
PubMed PubMed Central Google Scholar
Сигел, Дж. М. Ключи к функциям сна млекопитающих. Природа 437 , 1264–1271 (2005).
CAS PubMed Google Scholar
Лямин, О., Пряслова, Дж., Ланс, В., Сигел, Дж. Поведение животных: непрерывная активность китообразных после рождения. Природа 435 , 1177 (2005).
CAS PubMed Google Scholar
Lesku, J. A. et al. Адаптивная потеря сна у полигинных грудных куликов. Наука 337 , 1654–1658 (2012).
CAS PubMed Google Scholar
Rattenborg, N.C. et al. Мигрирующая бессонница у бело-венценосного воробья ( Zonotrichia leucophrys gambelii ). PLOS Biol. 2 , E212 (2004).
PubMed PubMed Central Google Scholar
Rattenborg, N.C. et al. Доказательства того, что птицы спят в полете. Nat. Commun. 7 , 12468 (2016).
CAS PubMed PubMed Central Google Scholar
Мухаметов, Л.М. Однополушарный медленноволновый сон амазонского дельфина Inia geoffrensis. Neurosci. Lett. 79 , 128–132 (1987).
CAS PubMed Google Scholar
Сигел, Дж. М. Все ли животные спят? Trends Neurosci. 31 , 208–213 (2008).
CAS PubMed Google Scholar
Фонвьей, А.М., Райзинг Р., Спраул М., Ларсон Д. Э. и Равуссин Е. Взаимосвязь между стадиями сна и скоростью метаболизма у людей. г. J. Physiol. 267 , E732 – E737 (1994).
CAS PubMed Google Scholar
Бреббиа Д. Р. и Альтшулер К. З. Скорость потребления кислорода и электроэнцефалографическая стадия сна. Наука 150 , 1621–1623 (1965).
CAS PubMed Google Scholar
Демент В. и Клейтман Н. Циклические вариации ЭЭГ во время сна и их связь с движениями глаз, подвижностью тела и сновидениями. Электроэнцефалогр. Clin. Neurophysiol. 9 , 673–690 (1957).
CAS PubMed Google Scholar
Rechtschaffen, A. & Kales, A. (eds) Руководство по стандартизированной терминологии, методам и системе оценки стадий сна у людей (Национальный институт неврологических заболеваний и слепоты США, 1968).
Жуве М. и Мишель Ф. Электромиографические корреляции сна у кошек с хроническим декортикальным и мезэнцефалическим синдромом [французский язык]. C. R. Seances Soc. Биол. Fil. 153 , 422–425 (1959).
CAS PubMed Google Scholar
Шейн-Идельсон, М., Ондрачек, Дж. М., Лиав, Х. П., Рейтер, С. и Лоран, Г. Медленные волны, резкие волны, рябь и быстрый сон у спящих драконов. Наука 352 , 590–595 (2016).
CAS PubMed Google Scholar
Николс, А. Л., Эйхлер, Т., Латам, Р. и Циммер, М. Глобальное состояние мозга лежит в основе поведения сна C. elegans . Наука 356 , eaam6851 (2017).
PubMed Google Scholar
Yap, M. H. W. et al. Колебательная активность мозга в спонтанных и индуцированных стадиях сна у мух. Nat. Commun. 8 , 1815 (2017).
PubMed PubMed Central Google Scholar
Нитц, Д. А., ван Суиндерен, Б., Тонони, Г. и Гринспен, Р. Дж. Электрофизиологические корреляты покоя и активности у Drosophila melanogaster . Curr. Биол. 12 , 1934–1940 (2002).
CAS PubMed Google Scholar
Рамон, Ф., Эрнандес-Фалькон, Дж., Нгуен, Б. и Баллок, Т. Х. Медленный сон у раков. Proc. Natl Acad. Sci. США 101 , 11857–11861 (2004).
CAS PubMed Google Scholar
Trojanowski, N. F., Nelson, M. D., Flavell, S. W., Fang-Yen, C. & Raizen, D. M. Отчетливые механизмы лежат в основе покоя во время двух состояний, подобных сну Caenorhabditis elegans . J. Neurosci. 35 , 14571–14584 (2015).
CAS PubMed PubMed Central Google Scholar
ван Альфен, Б., Яп, М. Х., Киршенблат, Л., Коттлер, Б. и ван Свиндерен, Б. Динамическая стадия глубокого сна у дрозофил . J. Neurosci. 33 , 6917–6927 (2013).
PubMed Google Scholar
Блумберг, М.С., Коулман, К.М., Герт, А. И., Мак-Мюррей, Б. Пространственно-временная структура подергиваний во сне в фазе быстрого сна выявляет происхождение двигательной синергии в процессе развития. Curr. Биол. 23 , 2100–2109 (2013).
CAS PubMed Google Scholar
Дилли, Л. К., Вигдерман, А., Уильямс, К. Э. и Кайзер, М. С. Поведенческие и генетические особенности онтогенеза сна у Drosophila . Спящий режим 41 , zsy086 (2018).
Google Scholar
Хобсон, Дж. А. Сон зависит от мозга, от мозга и для мозга. Природа 437 , 1254–1256 (2005).
CAS PubMed Google Scholar
Сапер, К. Б., Скаммелл, Т. Э. и Лу, Дж. Гипоталамическая регуляция сна и циркадных ритмов. Природа 437 , 1257–1263 (2005).
CAS PubMed Google Scholar
Джойнер, У. Дж., Крокер, А., Уайт, Б. Х. и Сегал, А. Сон в Drosophila регулируется взрослыми грибовидными телами. Природа 441 , 757–760 (2006).
CAS PubMed Google Scholar
Брингманн, Х. Активные во сне нейроны: законсервированные двигатели сна. Генетика 208 , 1279–1289 (2018).
CAS PubMed PubMed Central Google Scholar
Лим, Дж. И Динджес, Д. Ф. Недосыпание и бдительное внимание. Ann. NY Acad. Sci. 1129 , 305–322 (2008).
PubMed Google Scholar
Киршенблат, Л. и ван Суиндерен, Б. Инь и янь сна и внимания. Trends Neurosci. 38 , 776–786 (2015).
CAS PubMed PubMed Central Google Scholar
Cirelli, C. & Tononi, G. Сон и синаптический гомеостаз. Sleep 38 , 161–162 (2015).
PubMed PubMed Central Google Scholar
Krueger, J. M. & Tononi, G. Сон в зависимости от местного употребления; синтез новой парадигмы. Curr. Вершина. Med. Chem. 11 , 2490–2492 (2011).
CAS PubMed PubMed Central Google Scholar
Вязовский В. В. и Харрис К. Д. Сон и отдельный нейрон: роль глобальных медленных колебаний в покое отдельных клеток. Nat. Rev. Neurosci. 14 , 443–451 (2013).
CAS PubMed PubMed Central Google Scholar
Крюгер, Дж. М., Хуанг, Ю. Х., Ректор, Д. М. и Байсс, Д. Дж. Сон: синхронность состояний малых сетей, управляемых активностью клеток. евро. J. Neurosci. 38 , 2199–2209 (2013).
PubMed PubMed Central Google Scholar
Сапер, К. Б., Фуллер, П. М., Педерсен, Н. П., Лу, Дж. И Скаммелл, Т. Е. Переключение состояния сна. Нейрон 68 , 1023–1042 (2010).
CAS PubMed PubMed Central Google Scholar
Hinard, V. et al. Ключевые электрофизиологические, молекулярные и метаболические признаки сна и бодрствования, выявленные в первичных корковых культурах. J. Neurosci. 32 , 12506–12517 (2012).
CAS PubMed Google Scholar
Jewett, K. A. et al. Фактор некроза опухоли усиливает состояние, подобное сну, а электрическая стимуляция вызывает состояние, подобное бодрствованию, в совместных культурах нейронов и глии. евро. J. Neurosci. 42 , 2078–2090 (2015).
PubMed PubMed Central Google Scholar
Laposky, A. et al. Делеция гена циркадных часов млекопитающих BMAL1 / Mop3 изменяет базовую архитектуру сна и реакцию на лишение сна. Sleep 28 , 395–409 (2005).
PubMed Google Scholar
Ehlen, J. C. et al. Функция Bmal1 в скелетных мышцах регулирует сон. eLife 6 , e26557 (2017).
PubMed PubMed Central Google Scholar
Уильямс, Дж. А., Сатьянараян, С., Хендрикс, Дж. К. и Сегал, А. Взаимодействие между сном и иммунным ответом у Drosophila : роль NFκB Relish. Sleep 30 , 389–400 (2007).
PubMed PubMed Central Google Scholar
Bennett, H. L. et al. Нормальные приступы сна не важны для выживания C. elegans , а FoxO важен для компенсирующих изменений во сне. BMC Neurosci. 19 , 10 (2018).
PubMed PubMed Central Google Scholar
Iannacone, M. J. et al. RF-амидный рецептор DMSR-1 регулирует вызванный стрессом сон у C. elegans . eLife 6 , e19837 (2017).
PubMed PubMed Central Google Scholar
Smith, C. L. et al. Новые типы клеток, нейросекреторные клетки и строение тела раннего расходящегося многоклеточного животного Trichoplax adhaerens . Curr. Биол. 24 , 1565–1572 (2014).
CAS PubMed PubMed Central Google Scholar
Сенаторе А., Риз Т. С. и Смит К. Л. Нейропептидергическая интеграция поведения в Trichoplax adhaerens , животном без синапсов. J. Exp. Биол. 220 , 3381–3390 (2017).
PubMed PubMed Central Google Scholar
Смит К. Л., Пивоварова Н. и Риз Т. С. Скоординированное пищевое поведение у Trichoplax , животного без синапсов. PLOS ONE 10 , e0136098 (2015).
PubMed PubMed Central Google Scholar
Varoqueaux, F. et al. Высокое разнообразие клеток и сложная пептидергическая передача сигналов лежат в основе поведения плакозоя. Curr. Биол. 28 , 3495–3501 (2018).
CAS PubMed Google Scholar
Sakarya, O. et al. Постсинаптический каркас у истоков животного мира. PLOS ONE 2 , e506 (2007).
PubMed PubMed Central Google Scholar
Никель М. Кинетика и ритм сокращений тела губки Tethya wilhelma (Porifera: Demospongiae). J. Exp. Биол. 207 , 4515–4524 (2004).
PubMed Google Scholar
Людеман, Д. А., Фаррар, Н., Риесго, А., Папс, Дж. И Лейс, С. П. Эволюционное происхождение ощущений у многоклеточных животных: функциональные доказательства нового сенсорного органа у губок. BMC Evol. Биол. 14 , 3 (2014).
PubMed PubMed Central Google Scholar
де Майран, Дж. Дж. Д. Histoire de l’Académie Royale des Sciences (Année 1729) 35–36 (Imprimerie Royale, 1731).
Hattori, A. et al. Идентификация мелатонина в растениях и его влияние на уровни мелатонина в плазме и связывание с рецепторами мелатонина у позвоночных. Biochem. Мол. Биол. Int. 35 , 627–634 (1995).
CAS PubMed Google Scholar
Арнао, М. Б. и Эрнандес-Руис, Дж. Функции мелатонина в растениях: обзор. J. Pineal Res. 59 , 133–150 (2015).
CAS PubMed Google Scholar
Поройко В.А. и др. Хроническое нарушение сна изменяет микробиоту кишечника, вызывает системное воспаление и воспаление жировой ткани и резистентность к инсулину у мышей. Sci. Отчет 6 , 35405 (2016).
CAS PubMed PubMed Central Google Scholar
Szuperak, M. et al. Состояние сна у личинок Drosophila необходимо для пролиферации нервных стволовых клеток. eLife 7 , e33220 (2018).
PubMed PubMed Central Google Scholar
Кайзер М. С., Юэ З. и Сегал А. Критический период сна для развития схемы ухаживания и поведения у Drosophila . Наука 344 , 269–274 (2014).
CAS PubMed PubMed Central Google Scholar
Роффварг, Х. П., Муцио, Дж.Н. и Демент, В. С. Онтогенетическое развитие цикла сна-сновидений человека. Наука 152 , 604–619 (1966).
CAS PubMed Google Scholar
Дэвис К. и Райзен Д. М. Механизм болезненного сна: уроки беспозвоночных. J. Physiol. 595 , 5415–5424 (2016).
Google Scholar
Prather, A. A., Janicki-Deverts, D., Hall, M. H. и Cohen, S. Поведенческая оценка сна и восприимчивости к простуде. Sleep 38 , 1353–1359 (2015).
PubMed PubMed Central Google Scholar
Куо, Т. Х. и Уильямс, Дж. А. Увеличенный сон способствует выживанию во время бактериальной инфекции у Drosophila . Sleep 37 , 1077–1086 (2014).
PubMed PubMed Central Google Scholar
Хилл, А. Дж., Мэнсфилд, Р., Лопес, Дж. М., Райзен, Д. М. и Ван Бускерк, С. Клеточный стресс вызывает защитное состояние, подобное сну, у C. elegans . Curr. Биол. 24 , 2399–2405 (2014).
CAS PubMed PubMed Central Google Scholar
Хуанг Т.К., Ту, Дж., Чоу, Т. Дж. И Чен, Т. Х. Циркадный ритм прокариот Synechococcus sp. РФ-1. Plant Physiol. 92 , 531–533 (1990).
CAS PubMed PubMed Central Google Scholar
Postgate, J. Nitrogen Fixation 3-е изд. (Cambridge Univ. Press, 1998).
Liu, Y. et al. Циркадная оркестровка экспрессии генов у цианобактерий. Genes Dev. 9 , 1469–1478 (1995).
CAS PubMed Google Scholar
Ту, Б. П., Кудлики, А., Ровицка, М. и Макнайт, С. Л. Логика метаболического цикла дрожжей: временная компартментализация клеточных процессов. Наука 310 , 1152–1158 (2005).
CAS PubMed Google Scholar
Чен, З., Odstrcil, E. A., Tu, B. P. и McKnight, S. L. Ограничение репликации ДНК восстановительной фазой метаболического цикла защищает целостность генома. Наука 316 , 1916–1919 (2007).
CAS PubMed Google Scholar
Xie, L. et al. Сон стимулирует выведение метаболитов из мозга взрослого человека. Наука 342 , 373–377 (2013).
CAS Google Scholar
Maret, S. et al. Homer1a — это основной молекулярный коррелят потери сна в мозге. Proc. Natl Acad. Sci. США 104 , 20090–20095 (2007).
CAS PubMed Google Scholar
Cirelli, C., Gutierrez, C. M. и Tononi, G. Обширные и дивергентные эффекты сна и бодрствования на экспрессию генов мозга. Neuron 41 , 35–43 (2004).
CAS Google Scholar
Anafi, R.C. et al. Сон предназначен не только для мозга: транскрипционные реакции на сон в периферических тканях. BMG Genomics 14 , 362 (2013).
CAS Google Scholar
Mackiewicz, M. et al. Биосинтез макромолекул: ключевая функция сна. Physiol. Геном. 31 , 441–457 (2007).
CAS Google Scholar
Thompson, C. L. et al. Молекулярные и анатомические признаки депривации сна в мозге мышей. Фронт. Neurosci. 4 , 165 (2010).
PubMed PubMed Central Google Scholar
Archer, S. N. et al. Несвоевременный сон нарушает циркадную регуляцию человеческого транскриптома. Proc. Natl Acad. Sci. США 111 , E682 – E691 (2014).
CAS PubMed Google Scholar
Balsalobre, A. et al. Сброс суточного времени в периферических тканях по сигналу глюкокортикоидов. Наука 289 , 2344–2347 (2000).
CAS PubMed Google Scholar
Каджимото, Дж., Мацумура, Р., Узел, К. и Акаши, М. Потенциальная роль инсулина поджелудочной железы в сбросе периферических часов человека. Genes Cells 23 , 393–399 (2018).
CAS PubMed Google Scholar
Хардман, Дж. А., Хаслам, И. С., Фарджо, Н., Фарджо, Б. и Паус, Р. Тироксин по-разному модулирует периферические часы: уроки из человеческого волосяного фолликула. PLOS ONE 10 , e0121878 (2015).
PubMed PubMed Central Google Scholar
Браун С.А., Замбрунн Г., Флери-Олела Ф., Прейтнер Н. и Шиблер У. Ритмы температуры тела млекопитающих могут поддерживать периферические циркадные часы. Curr. Биол. 12 , 1574–1583 (2002).
CAS PubMed Google Scholar
Франкен П. и Дейк Д. Дж. Гены циркадных часов и гомеостаз сна. евро. J. Neurosci. 29 , 1820–1829 (2009).
CAS PubMed Google Scholar
Дуркин, Дж. И Атон, С. Дж. Зависимая от сна потенциация в зрительной системе противоречит гипотезе синаптического гомеостаза. Sleep 39 , 155–159 (2016).
PubMed PubMed Central Google Scholar
Лю, З. В., Фарагуна, У., Сирелли, К., Тонони, Г. и Гао, X. Б. Прямые доказательства связанного с бодрствованием и связанного со сном снижения синаптической силы в коре головного мозга грызунов. J. Neurosci. 30 , 8671–8675 (2010).
CAS PubMed PubMed Central Google Scholar
Хенген, К. Б., Торрадо Пачеко, А., МакГрегор, Дж. Н., Ван Хузер, С. Д. и Турриджиано, Г. Г. Гомеостаз скорости возбуждения нейронов подавляется сном и стимулируется бодрствованием. Cell 165 , 180–191 (2016).
CAS PubMed PubMed Central Google Scholar
Тонони, Г. и Сирелли, К. Сон и цена пластичности: от синаптического и клеточного гомеостаза до консолидации и интеграции памяти. Neuron 81 , 12–34 (2014).
CAS PubMed PubMed Central Google Scholar
Райхле, М. Э. и Минтун, М. А. Работа мозга и визуализация мозга. Annu. Rev. Neurosci. 29 , 449–476 (2006).
CAS PubMed Google Scholar
Леску, Дж. А. и Ли, Л. М. Т. Происхождение сна: спокойные медузы — это спящие медузы. Curr. Биол. 27 , R1060 – R1062 (2017).
CAS PubMed Google Scholar
Weljie, A. M. et al. Щавелевая кислота и диацилглицерин 36: 3 являются межвидовыми маркерами недосыпания. Proc. Natl Acad. Sci. США 112 , 2569–2574 (2015).
CAS PubMed Google Scholar
Davies, S. K. et al. Влияние недосыпания на метаболом человека. Proc. Natl Acad. Sci. США 111 , 10761–10766 (2014).
CAS PubMed Google Scholar
Tu, B.P. et al. Циклические изменения метаболического состояния в течение жизни дрожжевой клетки. Proc. Natl Acad. Sci. США 104 , 16886–16891 (2007).
CAS PubMed Google Scholar
Юргель М.Э. и др.Ade2 функционирует в жировом теле Drosophila , способствуя засыпанию. G3 8 , 3385–3395 (2018).
PubMed Google Scholar
Thimgan, M. S., Suzuki, Y., Seugnet, L., Gottschalk, L. & Shaw, P. J. Гомолог перилипина, капля хранения липидов 2, регулирует гомеостаз сна и предотвращает нарушения обучения после потери сна. PLOS Biol. 8 , e1000466 (2010).
PubMed PubMed Central Google Scholar
Skora, S., Mende, F. & Zimmer, M. Дефицит энергии способствует состоянию сна в масштабе всего мозга, модулируемому сигналом инсулина в C. elegans . Cell Rep. 22 , 953–966 (2018).
CAS PubMed PubMed Central Google Scholar
Rechtschaffen, A., Bergmann, B.М., Эверсон, К. А., Кушида, С. А. и Гиллиланд, М. А. Депривация сна у крысы: X. Обобщение и обсуждение результатов. Sleep 12 , 68–87 (1989).
CAS PubMed Google Scholar
Шоу, П. Дж., Тонони, Г., Гринспен, Р. Дж. И Робинсон, Д. Ф. Гены реакции на стресс защищают от летальных последствий недосыпания у Drosophila . Nature 417 , 287–291 (2002).
CAS PubMed Google Scholar
Рехтшаффен А. Современные взгляды на функцию сна. Перспектива. Биол. Med. 41 , 359–390 (1998).
CAS PubMed Google Scholar
Эверсон, К. А. Клиническая оценка лейкоцитов в крови, цитокинов сыворотки и иммуноглобулинов сыворотки как реакции на лишение сна у лабораторных крыс. г. J. Physiol. Regul. Интегр. Комп. Physiol. 289 , R1054 – R1063 (2005).
CAS PubMed Google Scholar
Макхилл, А. В. и Райт, К. П. Младший. Роль сна и нарушения циркадного ритма в расходе энергии и в метаболической предрасположенности к ожирению и метаболическим заболеваниям человека. Obes. Ред. 18 , S15 – S24 (2017).
Google Scholar
Naidoo, N. et al. Старение и недосыпание вызывают развернутый белковый ответ в поджелудочной железе: последствия для метаболизма. Ячейка старения 13 , 131–141 (2014).
CAS PubMed Google Scholar
Рехтшаффен А., Гиллиланд М. А., Бергманн Б. М. и Винтер Дж. Б. Физиологические корреляты длительного недосыпания у крыс. Наука 221 , 182–184 (1983).
CAS PubMed Google Scholar
Уокер М. П. и Стикголд Р. Сон, память и пластичность. Annu. Rev. Psychol. 57 , 139–166 (2006).
PubMed Google Scholar
Бенингтон, Дж. Х. и Франк, М. Г. Клеточные и молекулярные связи между сном и синаптической пластичностью. Прог. Neurobiol. 69 , 71–101 (2003).
CAS PubMed Google Scholar
Zhang, J. et al. Длительное бодрствование: нарушение метаболизма и дегенерация нейронов голубого пятна. J. Neurosci. 34 , 4418–4431 (2014).
PubMed PubMed Central Google Scholar
Шпигель К., Лепроулт Р. и Ван Каутер Э. Влияние недосыпания на метаболические и эндокринные функции. Ланцет 354 , 1435–1439 (1999).
CAS PubMed Google Scholar
Имери, Л. и Опп, М. Р. Как (и почему) иммунная система заставляет нас спать. Nat. Rev. Neurosci. 10 , 199–210 (2009).
CAS PubMed PubMed Central Google Scholar
Dubowy, C. et al. Генетическая диссоциация ежедневного сна и сна после термогенетической депривации сна у Drosophila . Sleep 39 , 1083–1095 (2016).
PubMed PubMed Central Google Scholar
Seidner, G. et al. Идентификация нейронов с привилегированной ролью в гомеостазе сна у Drosophila melanogaster . Curr. Биол. 25 , 2928–2938 (2015).
CAS PubMed PubMed Central Google Scholar
Альваренга, Т. А., Андерсен, М. Л., Папале, Л. А., Антунес, И. Б. и Туфик, С. Влияние длительного ограничения пищи на характер сна у самцов крыс. Brain Res. 1057 , 49–56 (2005).
CAS PubMed Google Scholar
Slocumb, M. E. et al. Улучшенный сон — это эволюционно адаптивный ответ на стресс голодания у Drosophila . PLOS ONE 10 , e0131275 (2015).
PubMed PubMed Central Google Scholar
Дубуэ, Э. Р., Кин, А. К. и Боровски, Р. Л. Эволюционная конвергенция потери сна у популяций пещерных рыб. Curr. Биол. 21 , 671–676 (2011).
CAS PubMed Google Scholar
Keene, A.C. et al. Часы и цикл ограничивают потерю сна, вызванную голоданием, у Drosophila.Curr. Биол. 20 , 1209–1215 (2010).
CAS PubMed Google Scholar
Goetting, D. L., Soto, R. & Van Buskirk, C. Пищевая пластичность у Caenorhabditis elegans стресс-индуцированного сна опосредуется передачей сигналов TOR-FOXA и TGF-β. Genetics 209 , 1183–1195 (2018).
PubMed Google Scholar
Ramm, P. & Smith, C. T. Скорость синтеза церебрального белка связана с медленным сном у крыс. Physiol. Behav. 48 , 749–753 (1990).
CAS PubMed Google Scholar
Simor, A. et al. Краткосрочные и долгосрочные протеомные эффекты лишения сна на корковые и таламические синапсы. Мол. Клетка. Neurosci. 79 , 64–80 (2017).
CAS PubMed Google Scholar
Сигел, Дж. М. Сон рассматривается как состояние адаптивной бездеятельности. Nat. Rev. Neurosci. 10 , 747–753 (2009).
CAS PubMed Google Scholar
Почему мы вообще спим?
Голод и еда; Сонливость и сон
Хотя мы не часто задумываемся о том, почему мы спим, большинство из нас на каком-то уровне осознают, что сон заставляет нас чувствовать себя лучше. После хорошего ночного сна мы чувствуем себя более бодрыми, энергичными, счастливыми и способными функционировать.Однако тот факт, что сон заставляет нас чувствовать себя лучше, а отсутствие сна заставляет нас чувствовать себя хуже, только начинает объяснять, почему может быть необходим сон.
Один из способов подумать о функции сна — сравнить его с другой нашей жизнедеятельностью: едой. Голод — это защитный механизм, который эволюционировал, чтобы гарантировать, что мы потребляем питательные вещества, необходимые нашему организму для роста, восстановления тканей и правильного функционирования. И хотя относительно легко понять роль, которую играет еда — учитывая, что она включает в себя физическое потребление веществ, в которых нуждается наш организм, — еда и сон не такие разные, как может показаться.
Еда и сон регулируются мощными внутренними приводами. Отсутствие еды вызывает неприятное чувство голода, а отказ от сна вызывает непомерную сонливость. И так же, как еда уменьшает чувство голода и обеспечивает получение необходимых нам питательных веществ, сон снимает сонливость и гарантирует, что мы получаем необходимый нам сон. Тем не менее, остается вопрос: зачем нам вообще нужен сон? Есть ли у сна единственная основная функция или сон выполняет множество функций?
Вопрос без ответа?
Ученые исследовали вопрос, почему мы спим, с разных сторон.Они исследовали, например, что происходит, когда люди или другие животные лишаются сна. В других исследованиях они изучали режим сна у различных организмов, чтобы увидеть, могут ли сходства или различия между видами раскрыть что-то о функциях сна. Тем не менее, несмотря на десятилетия исследований и множество открытий, касающихся других аспектов сна, на вопрос, почему мы спим, было трудно ответить.
Отсутствие четкого ответа на этот сложный вопрос не означает, что это исследование было пустой тратой времени.Фактически, теперь мы знаем гораздо больше о функции сна, и ученые разработали несколько многообещающих теорий, объясняющих, почему мы спим. В свете собранных ими доказательств кажется вероятным, что ни одна теория никогда не будет доказана правильной. Вместо этого мы можем обнаружить, что сон объясняется двумя или более из этих объяснений. Есть надежда, что, лучше понимая, почему мы спим, мы научимся больше уважать функции сна и наслаждаться пользой для здоровья, которую он дает.
В этом эссе излагается несколько современных теорий о том, почему мы спим.Чтобы узнать о них больше, обязательно ознакомьтесь с функцией «Книжная полка» в конце этого эссе. Там вы найдете ссылки на статьи исследователей, изучающих этот увлекательный вопрос.
Теории, почему мы спим
Теория бездействия
Одна из самых ранних теорий сна, которую иногда называют адаптивной или эволюционной теорией, предполагает, что бездействие в ночное время является адаптацией, которая выполняет функцию выживания, удерживая организмы от вреда в те моменты, когда они особенно уязвимы.Теория предполагает, что животные, которые могли оставаться неподвижными и тихими в эти периоды уязвимости, имели преимущество перед другими животными, которые оставались активными. Эти животные не попали в аварию, например, во время работы в темноте, и не были убиты хищниками. Предположительно, благодаря естественному отбору эта поведенческая стратегия превратилась в то, что мы теперь называем сном.
Простой контраргумент этой теории состоит в том, что всегда безопаснее оставаться в сознании, чтобы иметь возможность отреагировать на чрезвычайную ситуацию (даже если он лежит ночью в темноте).Таким образом, отсутствие сознания и сон не дают никаких преимуществ, если безопасность имеет первостепенное значение.
Теория сохранения энергии
Хотя это может быть менее очевидным для людей, живущих в обществах с изобилием источников пищи, одним из самых сильных факторов естественного отбора является конкуренция и эффективное использование энергетических ресурсов. Теория энергосбережения предполагает, что основная функция сна состоит в том, чтобы уменьшить потребность человека в энергии и ее расход в течение части дня или ночи, особенно в те моменты, когда поиск пищи наименее эффективен.
Исследования показали, что во время сна энергетический обмен значительно снижается (на целых 10 процентов у людей и даже больше у других видов). Например, во время сна и температура тела, и потребность в калориях снижаются по сравнению с бодрствованием. Такие данные подтверждают предположение, что одна из основных функций сна — помогать организмам сохранять свои энергетические ресурсы. Многие ученые считают, что эта теория связана с теорией бездействия и является ее частью.
Восстановительные теории
Другое объяснение того, почему мы спим, основано на давнем убеждении, что сон каким-то образом служит для «восстановления» того, что потеряно в организме, пока мы бодрствуем.Сон дает телу возможность восстановиться и омолодиться. В последние годы эти идеи получили поддержку эмпирических данных, собранных в исследованиях на людях и животных. Самым поразительным из них является то, что животные, полностью лишенные сна, теряют всю иммунную функцию и умирают всего за несколько недель. Это также подтверждается данными о том, что многие из основных восстановительных функций организма, такие как рост мышц, восстановление тканей, синтез белка и высвобождение гормона роста, происходят в основном или в некоторых случаях только во время сна.
Другие омолаживающие аспекты сна относятся к мозгу и когнитивным функциям. Например, пока мы бодрствуем, нейроны мозга производят аденозин, побочный продукт деятельности клеток. Считается, что накопление аденозина в головном мозге является одним из факторов, который приводит к нашему ощущению усталости. (Между прочим, это чувство нейтрализуется употреблением кофеина, который блокирует действия аденозина в мозгу и держит нас в бдительности.) Ученые считают, что это накопление аденозина во время бодрствования может способствовать «влечению ко сну».«Пока мы бодрствуем, аденозин накапливается и остается на высоком уровне. Во время сна у тела есть шанс вывести аденозин из системы, и, как следствие, мы чувствуем себя более бодрыми, когда просыпаемся.
Теория пластичности мозга
Одно из самых последних и убедительных объяснений того, почему мы спим, основано на выводах о том, что сон коррелирует с изменениями в структуре и организации мозга. Это явление, известное как пластичность мозга, до конца не изучено, но его связь со сном имеет несколько важных последствий.Например, становится ясно, что сон играет решающую роль в развитии мозга у младенцев и детей младшего возраста. Младенцы проводят от 13 до 14 часов в день во сне, и примерно половину этого времени они проводят в фазе быстрого сна, стадии, на которой происходит большинство сновидений. Связь между сном и пластичностью мозга становится очевидной и у взрослых. Это видно по тому влиянию, которое сон и недосыпание оказывают на способность людей учиться и выполнять различные задачи.
Эта теория и роль сна в обучении более подробно рассматриваются в книге «Сон, обучение и память».
Хотя эти теории остаются недоказанными, наука добилась огромных успехов в обнаружении того, что происходит во время сна и какие механизмы в организме контролируют циклы сна и бодрствования, которые определяют нашу жизнь. Хотя это исследование не дает прямого ответа на вопрос «Почему мы спим?» он действительно создает основу для постановки этого вопроса в новом контексте и получения новых знаний об этой важной части жизни.
Чтобы узнать больше о том, почему мы спим, посмотрите видео «Почему сон имеет значение» и изучите последствия недостаточного сна.
к началу
Последнее обновление этого контента: 18 декабря 2007 г.
Время— Доступ ко времени и преобразования — документация Python 3.9.6
Этот модуль предоставляет различные функции, связанные со временем. Для связанных
функциональные возможности, см. также модули datetime и calendar .
Хотя этот модуль всегда доступен, не все функции доступны на всех платформах. Большинство функций Определенные в этом модуле функции библиотеки C вызывают одноименные функции.Это иногда может быть полезно ознакомиться с документацией платформы, потому что семантика этих функций варьируется в зависимости от платформы.
Пояснение некоторых терминов и условных обозначений необходимо.
-
Эпоха — точка, где начинается время, и платформа зависимый. Для Unix эпоха — 1 января 1970 г., 00:00:00 (UTC). Чтобы узнать, какая эпоха на данной платформе, посмотрите на
время.gmtime (0).
-
Термин секунда с эпохи относится к общему количеству секунд, прошедших с эпохи, обычно исключая високосные секунды.Високосные секунды исключаются из этого общего количества на всех POSIX-совместимые платформы.
-
Функции в этом модуле могут не обрабатывать дату и время, предшествующие эпохе или в далеком будущем. Точка отсечения в будущем определяется C библиотека; для 32-битных систем это обычно 2038.
-
Функция
strptime ()может анализировать 2-значные годы при заданном формате% yкод. При анализе 2-значных лет они преобразуются в соответствии с POSIX. и стандарты ISO C: значения 69–99 соответствуют 1969–1999, а значения 0–68 сопоставлены с 2000–2068 гг.
-
DST — переход на летнее время, изменение часового пояса (обычно) на единицу. час в течение части года. Правила перехода на летнее время волшебны (определяются местным законодательством) и может меняться из года в год. В библиотеке C есть таблица, содержащая локальные rules (часто для гибкости он читается из системного файла) и является единственным источник Истинной Мудрости в этом отношении.
-
Точность различных функций реального времени может быть ниже, чем предлагается единицы измерения, в которых выражается их значение или аргумент.Например. на большинстве Unix В системах часы «тикают» только 50 или 100 раз в секунду.
-
С другой стороны, точность
time ()иsleep ()лучше чем их эквиваленты в Unix: время выражается как числа с плавающей запятой,time ()возвращает наиболее точное доступное время (с использованием Unixgettimeofday ()где доступно) иsleep ()примет время с ненулевой дробью (для реализации этого используется Unixselect (), где имеется в наличии). -
Значение времени, возвращаемое функциями
gmtime (),localtime ()иstrptime ()и принимаетсяasctime (),mktime ()иstrftime (), представляет собой последовательность из 9 целых чисел. Возвращаемые значенияgmtime (),localtime ()иstrptime ()также предлагают атрибут имена для отдельных полей.Описание этих объектов см. В
struct_time.Изменено в версии 3.3: Тип
struct_timeбыл расширен для предоставленияtm_gmtoffиtm_zoneатрибутов, если платформа поддерживает соответствующиеstruct tmучастников.Изменено в версии 3.6: Атрибуты
struct_timetm_gmtoffиtm_zoneтеперь доступны на всех платформах. -
Используйте следующие функции для преобразования между представлениями времени:
Функции
-
время.asctime([ t ]) -
Преобразование кортежа или
struct_time, представляющего время, возвращаемоеgmtime ()илиlocaltime ()в строку следующего форма:'Вс, 20 июня 23:21:05 1993'. Поле дня состоит из двух символов. и заполняется пробелом, если день состоит из одной цифры, например:'среда, 9 июня 04:26:40 1993'.Если t не указано, текущее время, возвращенное функцией
localtime ()используется.Информация о локали не используетсяasctime ().Примечание
В отличие от одноименной функции C,
asctime ()не добавляет завершающий символ новой строки.
-
время.pthread_getcpuclockid( thread_id ) -
Возвращает clk_id специфичных для потока часов процессорного времени для указанного thread_id .
Используйте
threading.get_ident ()или идентификаторнарезки.Разбейтеобъектов, чтобы получить подходящее значение для thread_id .Предупреждение
Передача недопустимого или просроченного thread_id может привести к неопределенное поведение, такое как ошибка сегментации.
Доступность: Unix (см. Справочную страницу для pthread_getcpuclockid (3) для дальнейшая информация).
-
время.clock_getres( clk_id ) -
Возвращает разрешение (точность) указанных часов clk_id .Ссылаться на Константы идентификатора часов для списка допустимых значений для clk_id .
Доступность: Unix.
-
время.clock_gettime( clk_id ) → с плавающей точкой -
Возвращает время указанных часов clk_id . Ссылаться на Константы идентификатора часов для списка допустимых значений для clk_id .
Доступность: Unix.
-
время.clock_gettime_ns( clk_id ) → int -
Аналогично
clock_gettime (), но возвращает время в наносекундах.Доступность: Unix.
-
время.clock_settime( clk_id , time: float ) -
Установить время указанных часов clk_id . В настоящее время,
CLOCK_REALTIME— единственное допустимое значение для clk_id .Доступность: Unix.
-
время.clock_settime_ns( clk_id , время: int ) -
Аналогично
clock_settime (), но задает время в наносекундах.Доступность: Unix.
-
время.ctime([ сек ]) -
Преобразует время, выраженное в секундах с начала эпохи, в строку формы:
'Вс 20 июня 23:21:05 1993'по местному времени. Поле дня состоит из двух символов и заполняется пробелами, если день состоит из одной цифры, например:'среда, 9 июня 04:26:40 1993'.Если сек. не указано или
Нет, текущее время как возвращаетсяtime () используется.ctime (secs)эквивалентноasctime (местное время (сек)). Информация о локали не используетсяctime ().
-
время.get_clock_info( имя ) -
Получить информацию об указанных часах в виде объекта пространства имен. Поддерживаемые имена часов и соответствующие функции для чтения их значения являются:
Результат имеет следующие атрибуты:
-
регулируемый :
Истинный, если часы можно менять автоматически (например,г. по демон NTP) или вручную системным администратором,Falseв противном случае -
реализация : имя базовой функции C, используемой для получения значение часов. См. Возможные значения в разделе «Константы идентификатора часов».
-
монотонный :
Истинный, если часы не могут идти назад,Неверноиначе -
разрешение : разрешение часов в секундах (
float)
-
-
время.gmtime([ сек ]) -
Преобразует время, выраженное в секундах с начала эпохи, в
struct_timeв UTC, в котором флаг dst всегда равен нулю. Если сек не предоставлено илиНет, используется текущее время, возвращенное функциейtime (). Фракции секунды игнорируются. См. Выше описаниеstruct_timeобъект. См.calendar.timegm (), чтобы узнать обратное. функция.
-
время.по местному времени([ сек ]) -
Аналогично
gmtime (), но преобразуется в местное время. Если сек не предоставлено илиНет, используется текущее время, возвращенное функциейtime (). ДСТ Флаг устанавливается на1, когда летнее время применяется к заданному времени.
-
время.мктайм( т ) -
Это функция, обратная
localtime ().Его аргумент —struct_timeили полный 9-кортеж (поскольку необходим флаг dst; используйте-1как флаг dst, если он неизвестен), который выражает время в по местному времени , а не УНИВЕРСАЛЬНОЕ ГЛОБАЛЬНОЕ ВРЕМЯ. Он возвращает число с плавающей запятой для совместимости сtime (). Если входное значение не может быть представлено как действительное время, либоOverflowErrorилиValueError(что зависит от обнаружено ли недопустимое значение Python или базовыми библиотеками C).Самая ранняя дата, для которой он может создать время, зависит от платформы.
-
время.монотонный() → поплавок -
Возвращает значение (в долях секунды) монотонных часов, то есть часов это не может вернуться назад. На часы не влияют обновления системных часов. Контрольная точка возвращаемого значения не определена, поэтому только Разница между результатами двух звонков действительно.
Изменено в версии 3.5: Функция теперь всегда доступна и всегда доступна для всей системы.
-
время.monotonic_ns() → int -
Аналогично
monotonic (), но возвращает время в наносекундах.
-
время.perf_counter() → поплавок -
Возвращает значение (в долях секунды) счетчика производительности, т. Е. часы с наивысшим доступным разрешением для измерения короткой продолжительности.Это включает время, прошедшее во время сна, и является общесистемной. Ссылка точка возвращаемого значения не определена, так что только разница между результаты двух звонков действительны.
-
время.perf_counter_ns() → int -
Аналогично
perf_counter (), но возвращает время в наносекундах.
-
время.process_time() → с плавающей точкой -
Возвращает значение (в долях секунды) суммы системы и пользователя. Процессорное время текущего процесса.Не включает время, прошедшее в течение спать. Это процесс по определению. Ориентир возвращаемое значение не определено, поэтому только разница между результатами из двух звонков действительно.
-
время.process_time_ns() → интервал -
Аналогично
process_time (), но возвращает время в наносекундах.
-
время.спящий режим( сек ) -
Приостановить выполнение вызывающего потока на заданное количество секунд.Аргументом может быть число с плавающей запятой, чтобы указать более точный сон. время. Фактическое время приостановки может быть меньше запрошенного, потому что любое пойманный сигнал завершит
sleep ()после выполнения этого режим ловли сигналов. Кроме того, время приостановки может быть больше, чем запрашивается произвольной суммой из-за планирования другой деятельности в системе.Изменено в версии 3.5: функция теперь спит не менее секунд , даже если спящий режим прерван сигналом, кроме случаев, когда обработчик сигнала вызывает исключение (см. PEP 475 для обоснования).
-
время.strftime(формат [, t ]) -
Преобразование кортежа или
struct_time, представляющего время, возвращаемоеgmtime ()илиlocaltime ()в строку, указанную в формате аргумент. Если t не указано, текущее время, возвращенноеlocaltime ()используется. Формат должен быть строкой.ValueError— это повышается, если какое-либо поле в t выходит за пределы допустимого диапазона.0 — допустимый аргумент для любой позиции во временном кортеже; если это нормально недопустимое значение принудительно установлено на правильное.
Следующие директивы могут быть встроены в строку формата . Они показаны без необязательной ширины поля и спецификации точности и заменяются по указанным символам в
strftime ()результат:Директива
Значение
Примечания
% аСокращенное название дня недели для локали.
% AПолное название дня недели Locale.
% bСокращенное название месяца для локали.
% BПолное название месяца для локали.
% cСоответствующие дата и время локали представление.
% dДень месяца в виде десятичного числа [01,31].
% ВЧас (24-часовой формат) в виде десятичного числа [00,23].
% IЧас (12-часовой формат) в виде десятичного числа [01,12].
% jДень года в виде десятичного числа [001,366].
% мМесяц в виде десятичного числа [01,12].
% MМинуты в виде десятичного числа [00,59].
% пЛокальный эквивалент AM или PM.
(1)
% SСекунда в виде десятичного числа [00,61].
(2)
% UНомер недели в году (воскресенье как первое день недели) в виде десятичного числа [00,53]. Все дни нового года, предшествующие первому Воскресенье считается нулевой неделей.
(3)
% wДень недели в виде десятичного числа [0 (воскресенье), 6].
% ВтНомер недели в году (понедельник как первый день недели) в виде десятичного числа [00,53].Все дни нового года, предшествующие первому Понедельник считается нулевой неделей.
(3)
% xСоответствующее представление даты для локали.
% XСоответствующее временное представление локали.
% гГод без века в виде десятичного числа [00,99].
% ВыходГод с десятичным числом столетия.
% zСмещение часового пояса, указывающее положительное или отрицательная разница во времени с UTC / GMT форма + ЧЧММ или -ЧЧММ, где H представляет собой десятичную дробь цифры часа, а M представляет собой десятичную минуту цифры [-23: 59, +23: 59].
% ZНазвание часового пояса (без символов, если часовой пояс отсутствует существует).
%%Буквальный символ
'%'.Примечания:
-
При использовании с функцией
strptime ()директива% pвлияет только на поле выходного часа, если для синтаксического анализа часа используется директива% I. -
Диапазон действительно составляет от
0до61; значение60действительно в временные метки, представляющие дополнительные секунды, и поддерживается значение61по историческим причинам. -
При использовании с функцией
strptime (),% Uи% Wиспользуются только в расчеты, когда указаны день недели и год.
Вот пример формата дат, совместимого с указанным в RFC 2822 Стандарт электронной почты в Интернете.
>>> from time import gmtime, strftime >>> strftime ("% a,% d% b% Y% H:% M:% S +0000", gmtime ()) 'Чт, 28 июня 2001 14:17:15 +0000'Дополнительные директивы могут поддерживаться на определенных платформах, но только перечисленные здесь имеют значение, стандартизированное ANSI C.Чтобы увидеть полный набор кодов формата, поддерживаемых вашей платформой, обратитесь к strftime (3) документация.
На некоторых платформах необязательная спецификация ширины поля и точности может сразу после начального
"%"директивы в следующем порядке; это тоже не переносимо. Ширина поля обычно равна 2, за исключением% j, где это 3. -
-
время.strptime( строка [, формат ]) -
Разобрать строку, представляющую время в соответствии с форматом.Возвращаемое значение — это
struct_time, возвращенная функциейgmtime ()илипо местному времени ().Формат Параметр использует те же директивы, что и
strftime (); по умолчанию это"% a% b% d% H:% M:% S% Y", что соответствует форматирование, возвращаемое функциейctime (). Если строка не может быть проанализирована в соответствии с в формат , или если после синтаксического анализа у него есть избыточные данные,ValueErrorбудет поднятый.Значения по умолчанию, используемые для заполнения любых недостающих данных, когда больше точные значения не могут быть выведены:(1900, 1, 1, 0, 0, 0, 0, 1, -1). И строка, и формат должны быть строками.Например:
>>> время импорта >>> time.strptime («30 ноя 00», «% d% b% y») time.struct_time (tm_year = 2000, tm_mon = 11, tm_mday = 30, tm_hour = 0, tm_min = 0, tm_sec = 0, tm_wday = 3, tm_yday = 335, tm_isdst = -1)Поддержка директивы
% Zоснована на значениях, содержащихся вtznameи правда лидневной.Из-за этого он зависит от платформы за исключением признания UTC и GMT, которые всегда известны (и считаются быть часовыми поясами без перехода на летнее время).Поддерживаются только директивы, указанные в документации. Потому что
strftime ()реализовано для каждой платформы, иногда может предложить больше директив, чем перечисленные. Ноstrptime ()не зависит от какой-либо платформы. и, следовательно, не обязательно поддерживает все доступные директивы, которые не задокументировано как поддерживаемое.
-
класс
время.struct_time -
Тип последовательности значений времени, возвращаемый функцией
gmtime (),localtime ()иstrptime (). Это объект с именем Интерфейс кортежа: значения могут быть доступны по индексу и по имени атрибута. В присутствуют следующие значения:Индекс
Атрибут
Значения
0
tm_год(например, 1993)
1
тм_мондиапазон [1, 12]
2
tm_mdayдиапазон [1, 31]
3
tm_hourдиапазон [0, 23]
4
tm_minдиапазон [0, 59]
5
tm_secдиапазон [0, 61]; см. (2) в
strftime ()описание6
tm_wdayдиапазон [0, 6], понедельник 0
7
tm_ydayдиапазон [1, 366]
8
tm_isdst0, 1 или -1; см. ниже
НЕТ
тм_зонасокращение названия часового пояса
НЕТ
tm_gmtoffсмещение к востоку от UTC в секундах
Обратите внимание, что, в отличие от структуры C, значение месяца является диапазоном [1, 12], а не [0, 11].
При звонках на
mktime (),tm_isdstможет быть установлен в 1 при дневном свете. действует время экономии, а когда нет — 0. Значение -1 указывает, что это неизвестно и обычно приводит к заполнению правильного состояния.Когда кортеж с неправильной длиной передается функции, ожидающей
struct_timeили наличие элементов неправильного типа, aTypeErrorвозникает.
-
время.время() → поплавок -
Возвращает время в секундах с начала эпохи в виде числа с плавающей запятой. номер. Конкретная дата эпохи и обращение с дополнительные секунды зависят от платформы. В Windows и большинстве Unix-систем эта эпоха — 1 января 1970 г. 00:00:00 (UTC) и дополнительные секунды не учитываются во времени. в секундах с эпохи. Это обычно называют Время Unix. Чтобы узнать, какая эпоха на данной платформе, посмотрите на
gmtime (0).Обратите внимание, что даже если время всегда возвращается в виде числа с плавающей запятой число, не все системы обеспечивают точность времени лучше, чем 1 секунда.Хотя эта функция обычно возвращает неубывающие значения, она может возвращать более низкое значение, чем в предыдущем вызове, если системные часы были переведены назад между двумя звонками.
Число, возвращаемое функцией
time (), может быть преобразовано в более распространенное формат времени (например, год, месяц, день, час и т. д.) в UTC, передав его вgmtime ()или по местному времени, передав его функцииlocaltime ()функция. В обоих случаяхstruct_timeвозвращается объект, из которого компоненты календарной даты можно получить как атрибуты.
-
время.thread_time() → с плавающей запятой -
Возвращает значение (в долях секунды) суммы системы и пользователя. Процессорное время текущего потока. Не включает время, прошедшее в течение спать. По определению, он зависит от потока. Ориентир возвращаемое значение не определено, поэтому только разница между результатами двух вызовов в одном потоке.
Доступность: Windows, Linux, поддержка систем Unix
CLOCK_THREAD_CPUTIME_ID.
-
время.thread_time_ns() → интервал -
Аналогично
thread_time (), но возвращает время в наносекундах.
-
время.time_ns() → int -
Аналогично
time (), но возвращает время в виде целого числа наносекунд. с эпохи.
-
время.цсет() -
Сбросить правила преобразования времени, используемые библиотечными подпрограммами.Окружение переменная
TZопределяет, как это делается. Он также установит переменныеtzname(из переменной средыTZ),часовой пояс(без летнего времени секунд к западу от UTC),altzone(секунды DST к западу от UTC) идневной свет(до 0, если для этого часового пояса нет правил перехода на летнее время, или до ненулевое значение, если есть время, прошлое, настоящее или будущее, когда летнее время применяется).Доступность: Unix.
Примечание
Хотя во многих случаях изменение переменной среды
TZможет влияют на вывод таких функций, какlocaltime ()без вызоваtzset (), на такое поведение нельзя полагаться.Переменная среды
TZне должна содержать пробелов.Стандартный формат переменной среды
TZ(пробел добавлено для ясности):стандартное смещение [dst [смещение [, начало [/ время], конец [/ время]]]]
Где находятся компоненты:
-
stdиdst -
Три или более буквенно-цифровых символов, обозначающих сокращения часовых поясов. Это будет распространяется в time.tzname
-
смещение -
Смещение имеет вид:
± чч [: мм [: сс]].Это указывает значение добавлено местное время для прихода в UTC. Если перед ним стоит «-«, часовой пояс находится к востоку от нулевого меридиана; в противном случае это запад. Если не следует смещение dst предполагается, что летнее время на один час опережает стандартное время. -
начало [/ время], конец [/ время] -
Указывает, когда перейти на летнее время и обратно. Формат даты начала и окончания — одна из следующих:
-
J n -
Юлианский день n (1 <= n <= 365).Високосные дни не учитываются, поэтому в для всех лет 28 февраля - это 59-й день, а 1 марта - 60-й день.
-
n -
Юлианский день с отсчетом от нуля (0 <= n <= 365). Считаются високосные дни, и можно сослаться на 29 февраля.
-
М м . № . d -
d день (0 <= d <= 6) недели n месяца m года (1 <= n <= 5, 1 <= m <= 12, где неделя 5 означает «последний d день в месяц м », который может наступить либо в четвертом, либо в пятом месяце. неделя).Неделя 1 — это первая неделя, в которой наступает день d . День ноль — воскресенье.
времяимеет тот же формат, что исмещение, за исключением того, что нет ведущего знака (‘-‘ или ‘+’) разрешено. По умолчанию, если время не указано, 02:00:00. -
>>> os.environ ['TZ'] = 'EST + 05EDT, M4.1.0, M10.5.0' >>> time.tzset () >>> time.strftime ('% X% x% Z') '02: 07: 36 05.08.03 EDT ' >>> os.environ ['TZ'] = 'AEST-10AEDT-11, M10.5.0, M3.5.0 ' >>> time.tzset () >>> time.strftime ('% X% x% Z') '16: 08: 12 05.08.03 AEST 'Во многих системах Unix (включая * BSD, Linux, Solaris и Darwin) больше удобно использовать системную базу данных zoneinfo ( tzfile (5) ) для укажите правила часового пояса. Для этого установите среду
TZв путь к необходимому файлу данных о часовом поясе относительно корня база данных часовых поясов «zoneinfo» системы, обычно расположенная по адресу/ usr / share / zoneinfo.Например,«США / Восток»,«Австралия / Мельбурн»,«Египет»или«Европа / Амстердам».>>> os.environ ['TZ'] = 'США / Восток' >>> time.tzset () >>> time.tzname ('EST', 'EDT') >>> os.environ ['TZ'] = 'Египет' >>> time.tzset () >>> time.tzname ('EET', 'EEST') -
Константы идентификатора часов
Эти константы используются как параметры для clock_getres (), и
clock_gettime () .
-
время.ЧАСЫ_ВРЕМЯ ЗАГРУЗКИ -
Идентично
CLOCK_MONOTONIC, за исключением того, что включает любое время, которое система приостановлена.Это позволяет приложениям получать монотонные часы с поддержкой приостановки без иметь дело с осложнениями
CLOCK_REALTIME, которые могут имеют разрывы, если время изменяется с помощьюsettimeofday ()или аналогичный.Доступность: Linux 2.6.39 или новее.
-
время.ВЫСОКОЕ ЧАСЫ -
В ОС Solaris есть таймер
CLOCK_HIGHRES, который пытается использовать оптимальную аппаратный источник и может давать разрешение, близкое к наносекундному.CLOCK_HIGHRES— это нерегулируемые часы с высоким разрешением.Наличие: Solaris.
-
время.ЧАСЫ_MONOTONIC -
Часы, которые не могут быть установлены и представляют монотонное время, поскольку некоторая неопределенная отправная точка.
Доступность: Unix.
-
время.ЧАСЫ_MONOTONIC_RAW -
Аналогично
CLOCK_MONOTONIC, но обеспечивает доступ к необработанному аппаратное время, не зависящее от настроек NTP.Доступность: Linux 2.6.28 и новее, macOS 10.12 и новее.
-
время.CLOCK_PROCESS_CPUTIME_ID -
Таймер каждого процесса с высоким разрешением от ЦП.
Доступность: Unix.
-
время.CLOCK_PROF -
Таймер каждого процесса с высоким разрешением от ЦП.
Доступность: FreeBSD, NetBSD 7 или новее, OpenBSD.
-
время.ЧАСЫ_TAI -
Международное атомное время
В системе должна быть текущая таблица дополнительных секунд, чтобы правильный ответ. Программное обеспечение PTP или NTP может поддерживать таблицу секунды координации.
Доступность: Linux.
-
время.CLOCK_THREAD_CPUTIME_ID -
Часы процессора, зависящие от потока.
Доступность: Unix.
-
время.ЧАСЫ ВРЕМЕНИ -
Время, абсолютным значением которого является время, в течение которого система работала, а не приостановлено, обеспечивая точное измерение времени безотказной работы, как абсолютное, так и интервал.
Доступность: FreeBSD, OpenBSD 5.5 или новее.
-
время.ЧАСЫ_УПРЕЖДЕНИЕ_RAW -
Часы, которые монотонно увеличиваются, отслеживая время с момента произвольного точка, на которую не влияют настройки частоты или времени и не увеличивается при система спит.
Доступность: macOS 10.12 и новее.
Следующая константа — единственный параметр, который может быть отправлен в
clock_settime () .
-
время.ЧАСЫ_РЕАЛЬНОЕ ВРЕМЯ -
Часы реального времени в масштабе всей системы. Для установки этих часов требуются соответствующие привилегии.
Доступность: Unix.
Константы часового пояса
-
время.альтцона -
Смещение местного часового пояса летнего времени в секундах к западу от UTC, если он определен. Это отрицательно, если местный часовой пояс DST находится к востоку от UTC (как в Западной Европе, включая Великобританию). Используйте это, только если
дневной светотличен от нуля.См. Примечание ниже.
-
время.дневной свет -
Ненулевое значение, если указан часовой пояс DST. См. Примечание ниже.
-
время.часовой пояс -
Смещение местного (не летнего времени) часового пояса в секундах к западу от всемирного координированного времени (отрицательное в большая часть Западной Европы, положительная в США, ноль в Великобритании). См. Примечание ниже.
-
время.tzname -
Кортеж из двух строк: первая — это имя локального часового пояса без перехода на летнее время, второй — это название местного часового пояса летнего времени.Если часовой пояс DST не определен, вторую строку использовать не следует. См. Примечание ниже.
Примечание
Для указанных выше констант часового пояса ( altzone , daylight , timezone ,
и tzname ), значение определяется действующими правилами часового пояса
во время загрузки модуля или в последний раз вызывается tzset () и может быть некорректным
для времен в прошлом. Рекомендуется использовать tm_gmtoff и
tm_zone является результатом localtime () для получения информации о часовом поясе.
См. Также
- Модуль
datetime -
Более объектно-ориентированный интерфейс для дат и времени.
- Модуль
локаль -
Услуги интернационализации. Настройка локали влияет на интерпретацию многих спецификаторов формата в
strftime ()иstrptime (). - Модуль
Календарь -
Общие функции, связанные с календарем.
timegm ()— это инверсияgmtime ()из этого модуля.
Сноски
Изучение функции сна — ScienceDaily
Важен ли сон? Задайте этот вопрос недосыпающемуся новому родителю или ученику, который только что потянул «всю ночь», и ответ будет ворчливым: «Конечно!»
Но для исследователя сна вопрос о том, что представляет собой сон, настолько сложен, что ученые все еще пытаются определить основную функцию того, что мы делаем каждую ночь. Исследование, опубликованное на этой неделе в журнале PLoS Biology Кьярой Сирелли и Джулио Тонони, посвящено этому актуальному вопросу.
Поиск основной функции сна может показаться столь же неуловимым, как поиск мифологического феникса, говорит Сирелли, доцент психиатрии Школы медицины и общественного здравоохранения Университета Висконсина в Мэдисоне.
Некоторые ученые утверждают, что сон — это просто способ установить спокойное неподвижное состояние (покой), и сам по себе он не важен для млекопитающих и птиц. Это так называемая «нулевая гипотеза», которую Чирелли и Тонони отвергают.
«Мы не понимаем, зачем нужен сон, но он должен быть важен, потому что это делают все животные», — говорит Чирелли.
По ее словам, нет явных доказательств того, что какой-то вид животных не спит. Даже дельфин, которого иногда называют животным, которое не спит, потому что он постоянно движется, будет демонстрировать «однополушарный сон» с одним закрытым глазом, а половина его мозга показывает медленные волны, характерные для глубокого сна.
«Сам факт того, что дельфины развили замечательную специализацию … вместо того, чтобы просто полностью избавиться от сна, должен считаться доказательством того, что сон должен выполнять некоторые важные функции и не может быть устранен», — говорит Сирелли.
Она также утверждает, что сон строго регулируется мозгом, потому что за недосыпанием следует отскок, при котором лишенное сна животное либо спит дольше, либо проводит больше времени в более глубоком сне, характеризующемся большими медленными мозговыми волнами.
Доказано, что длительное лишение сна убивает крыс, мух и тараканов. Люди, страдающие генетической бессонницей, также могут умереть. В менее крайних случаях недосыпание влияет на когнитивные функции у животных, от мух до грызунов.Бодрствующие крысы будут участвовать в эпизодах «микросна», а недосыпающие люди склонны засыпать даже в самых опасных обстоятельствах.
Поскольку он универсален, строго регулируется и не может быть потерян без серьезного вреда, Сирелли утверждает, что сон должен иметь важную основную функцию. Но что?
«Сон может быть ценой, которую вы платите за то, чтобы на следующий день ваш мозг стал пластичным», — говорят Чирелли и Тонони.
Их гипотеза состоит в том, что сон позволяет мозгу перегруппироваться после тяжелого учебного дня, давая синапсам, которые в течение дня усиливаются, возможность ослабнуть до исходного уровня.Это важно, потому что мозг использует до 80 процентов своей энергии для поддержания синаптической активности.
Сон также может быть важен для закрепления новых воспоминаний и для того, чтобы позволить мозгу «забыть» случайные, неважные впечатления дня, так что на следующий день есть место для дальнейшего изучения. Возможно, поэтому мозговые волны так активны в определенные периоды сна.
«Хотя до сих пор нет единого мнения о том, почему животным нужен сон, может показаться, что поиск основной функции сна, особенно на клеточном уровне, по-прежнему стоит того», — заключает она.
История Источник:
Материалы предоставлены Публичной научной библиотекой . Примечание. Содержимое можно редактировать по стилю и длине.
нарушений сна | MedlinePlus
Что такое сон?
Сон — сложный биологический процесс. Пока вы спите, вы без сознания, но функции вашего мозга и тела по-прежнему активны. Они выполняют ряд важных дел, которые помогают вам оставаться здоровыми и работать с максимальной эффективностью.Поэтому, когда вы не высыпаетесь достаточно качественного сна, это не просто утомляет вас. Это может повлиять на ваше физическое и психическое здоровье, мышление и повседневное функционирование.
Что такое нарушения сна?
Расстройства сна — это состояния, которые нарушают ваш нормальный режим сна. Существует более 80 различных нарушений сна. Некоторые основные типы включают
- Бессонница — невозможность заснуть и уснуть. Это наиболее частое нарушение сна.
- Апноэ во сне — нарушение дыхания, при котором вы перестаете дышать на 10 секунд или более во время сна
- Синдром беспокойных ног (СБН) — ощущение покалывания или покалывания в ногах вместе с сильным желанием пошевелить ими
- Гиперсомния — неспособность бодрствовать в течение дня.Сюда входит нарколепсия, которая вызывает сильную дневную сонливость.
- Нарушения циркадного ритма — проблемы с циклом сна-бодрствования. Из-за них вы не можете заснуть и просыпаетесь в нужное время.
- Парасомния — необычные действия во время засыпания, сна или пробуждения ото сна, например ходьба, разговор или прием пищи
Некоторые люди, которые чувствуют усталость в течение дня, страдают настоящим нарушением сна. Но для других настоящая проблема — не давать достаточно времени для сна.Важно высыпаться каждую ночь. Количество необходимого вам сна зависит от нескольких факторов, в том числе от вашего возраста, образа жизни, состояния здоровья и от того, достаточно ли вы высыпались в последнее время. Большинству взрослых требуется около 7-8 часов каждую ночь.
Что вызывает нарушения сна?
Существуют разные причины нарушения сна, в том числе
Иногда причина неизвестна.
Есть также некоторые факторы, которые могут способствовать проблемам со сном, в том числе
- Кофеин и алкоголь
- Нерегулярный график, например, работа в ночную смену
- Старение.С возрастом люди часто меньше спят или проводят меньше времени в глубоком, спокойном сне. Их также легче разбудить.
Каковы симптомы нарушения сна?
Симптомы нарушения сна зависят от конкретного расстройства. Некоторые признаки того, что у вас может быть нарушение сна, включают:
- Каждую ночь вам требуется более 30 минут, чтобы заснуть
- Вы регулярно просыпаетесь несколько раз каждую ночь, а затем испытываете проблемы с засыпанием или просыпаетесь слишком рано утром
- Вы часто чувствуете сонливость в течение дня, часто спите или засыпаете в неподходящее время в течение дня
- Ваш партнер по постели говорит, что когда вы спите, вы громко храпите, фыркаете, задыхаетесь, издаете удушающие звуки или на короткое время перестаете дышать
- У вас мурашки, покалывания или ползания мурашек в ногах или руках, которые облегчаются от движения или массажа ими, особенно вечером и при попытке заснуть
- Ваш партнер по постели замечает, что ваши ноги или руки часто дергаются во время сна
- Вы переживаете яркие сказочные переживания во время сна или дремоты
- У вас бывают эпизоды внезапной мышечной слабости, когда вы злитесь или напуганы, или когда вы смеетесь
- Вы чувствуете, что не можете двигаться, когда впервые просыпаетесь
Как диагностируются нарушения сна?
Чтобы поставить диагноз, ваш лечащий врач будет использовать вашу историю болезни, историю сна и физический осмотр.Вы также можете пройти исследование сна (полисомнограмму). Наиболее распространенные типы исследований сна отслеживают и записывают данные о вашем теле в течение полной ночи сна. Данные включают
- Изменения мозговых волн
- Движения глаз
- Частота дыхания
- Артериальное давление
- Частота сердечных сокращений и электрическая активность сердца и других мышц
Другие типы исследований сна могут проверить, насколько быстро вы засыпаете во время дневного сна или можете ли вы бодрствовать и бодрствовать в течение дня.
Как лечить расстройство сна?
Лечение расстройств сна зависит от того, какое у вас расстройство. Они могут включать
- Привычки к правильному сну и другие изменения образа жизни, такие как здоровое питание и физические упражнения
- Когнитивно-поведенческая терапия или методы релаксации для уменьшения беспокойства о достаточном сне
- Аппарат CPAP (непрерывное положительное давление в дыхательных путях) для лечения апноэ во сне
- Светотерапия (утром)
- Лекарства, включая снотворное.Обычно медработники рекомендуют вам принимать снотворное в течение короткого периода времени.
- Натуральные продукты, например мелатонин. Эти продукты могут помочь некоторым людям, но обычно предназначены для краткосрочного использования. Обязательно проконсультируйтесь со своим врачом, прежде чем принимать какие-либо из них.
Что происходит в мозге во время сна?
Джон Пивер , директор Лаборатории системной нейробиологии Университета Торонто, и Брайан Дж. Мюррей , директор лаборатории сна в Центре медицинских наук Саннибрук, ответ:
Функция сна вводила в заблуждение ученых на протяжении тысячелетий, но современные исследования дают новые подсказки о том, что он делает для разума и тела.Сон восстанавливает энергию клеток организма, выводит отходы из мозга и поддерживает обучение и память. Он даже играет жизненно важную роль в регулировании настроения, аппетита и либидо.
Сон — неотъемлемая часть нашей жизни, и, как показывают исследования, он невероятно сложен. Мозг генерирует два разных типа сна: медленный сон (SWS), известный как глубокий сон, и быстрое движение глаз (REM), также называемый сном сновидений. Большая часть сна, который мы спим, относится к категории SWS, для которой характерны большие медленные мозговые волны, расслабленные мышцы и медленное, глубокое дыхание, которое может помочь мозгу и телу восстановить силы после долгого дня.
Когда мы засыпаем, мозг не просто отключается, о чем свидетельствует общая фраза «гаснет, как свет». Вместо этого серия тщательно спланированных событий поэтапно усыпляет мозг. Технически сон начинается в областях мозга, которые производят SWS. У ученых теперь есть конкретные доказательства того, что две группы клеток — вентролатеральное преоптическое ядро в гипоталамусе и парафациальная зона в стволе мозга — участвуют в инициации SWS. Когда эти клетки включаются, это вызывает потерю сознания.
После МС начинается быстрый сон. Это странный режим: мозг сновидца становится очень активным, в то время как мышцы тела парализованы, а дыхание и частота сердечных сокращений становятся неустойчивыми. Назначение быстрого сна остается биологической загадкой, несмотря на наше растущее понимание его биохимии и нейробиологии.
Мы действительно знаем, что небольшая группа клеток в стволе мозга, называемая ядром subcoeruleus, контролирует быстрый сон. Когда эти клетки повреждаются или заболевают, люди не испытывают мышечного паралича, связанного с быстрым сном, который может привести к поведенческому расстройству в фазе быстрого сна — серьезному состоянию, при котором пострадавшие жестоко разыгрывают свои сны.
Функция Python sleep () (с примерами)
Python имеет модуль с именем time, который предоставляет несколько полезных функций для обработки задач, связанных со временем. Одна из популярных среди них — sleep () .
Функция sleep () приостанавливает выполнение текущего потока на заданное количество секунд.
Пример 1: Python sleep ()
время импорта
print ("Распечатывается немедленно.")
time.sleep (2.4)
print ("Отпечатано после 2.4 секунды. ")
Вот как работает эта программа:
-
«Отпечатано немедленно»напечатано - Приостанавливает (задерживает) выполнение на 2,4 секунды.
-
«Отпечатано через 2,4 секунды» Распечатано.
Как видно из приведенного выше примера, sleep () принимает в качестве аргумента число с плавающей запятой.
До Python 3.5 фактическое время приостановки могло быть меньше аргумента, указанного для функции time () .
Начиная с Python 3.5 , время приостановки будет не менее указанного в секундах.
Пример 2: Python создает цифровые часы
время импорта
в то время как True:
localtime = время.localtime ()
result = time.strftime ("% I:% M:% S% p", местное время)
печать (результат)
время сна (1)
В приведенной выше программе мы вычислили и распечатали текущее местное время внутри бесконечного цикла while. Затем программа ждет 1 секунду. Опять же, текущее местное время вычисляется и печатается.Этот процесс продолжается.
Когда вы запустите программу, вывод будет примерно таким:
14:10:50 14:10:51 14:10:52 14:10:53 14:10:54 ... .. ...
Вот немного измененная лучшая версия вышеупомянутой программы.
время импорта
в то время как True:
localtime = время.localtime ()
result = time.strftime ("% I:% M:% S% p", местное время)
print (результат, end = "", flush = True)
print ("\ r", end = "", flush = True)
время сна (1)
Чтобы узнать больше, посетите цифровые часы в оболочке Python.
Многопоточность в Python
Прежде чем говорить о sleep () в многопоточных программах, давайте поговорим о процессах и потоках.
Компьютерная программа — это набор инструкций. Процесс — это выполнение этих инструкций.
Поток — это часть процесса. У процесса может быть один или несколько потоков.
Пример 3: Многопоточность Python
Все программы, перечисленные выше в этой статье, являются однопоточными.Вот пример многопоточной программы на Python.
импорт потоковой передачи
def print_hello_three_times ():
для i в диапазоне (3):
print ("Привет")
def print_hi_three_times ():
для i в диапазоне (3):
print ("Привет")
t1 = threading.Thread (цель = print_hello_three_times)
t2 = threading.Thread (цель = print_hi_three_times)
t1.start ()
t2.start ()
Когда вы запустите программу, вывод будет примерно таким:
Привет Привет Здравствуй Привет Здравствуй Здравствуй
Вышеупомянутая программа имеет два потока t1 и t2 .Эти потоки выполняются с использованием операторов t1.start () и t2.start () .
Обратите внимание, что t1 и t2 выполняются одновременно, и вы можете получить разные результаты.
Посетите эту страницу, чтобы узнать больше о многопоточности в Python.
time.sleep () в многопоточных программах
Функция sleep () приостанавливает выполнение текущего потока на заданное количество секунд.
В случае однопоточных программ sleep () приостанавливает выполнение потока и процесса.Однако в многопоточных программах функция приостанавливает поток, а не весь процесс.
Пример 4: sleep () в многопоточной программе
импорт резьбы
время импорта
def print_hello ():
для i в диапазоне (4):
time.sleep (0,5)
print ("Привет")
def print_hi ():
для i в диапазоне (4):
time.sleep (0,7)
print ("Привет")
t1 = threading.Thread (цель = print_hello)
t2 = threading.Thread (цель = print_hi)
t1.start ()
t2.start ()
Вышеупомянутая программа имеет два потока.Мы использовали time.sleep (0.5) и time.sleep (0.75) , чтобы приостановить выполнение этих двух потоков на 0,5 секунды и 0,7 секунды соответственно.