
Избирательность: Недопустимое название — Викисловарь
Селективность (избирательность) / КонсультантПлюс
11. Биоаналитическая методика должна обладать способностью дифференцировать анализируемое вещество и ВС от эндогенных компонентов матрицы и других компонентов образца. Селективность биоаналитической методики необходимо подтвердить, используя не менее 6 различных источников соответствующих холостых образцов, не содержащих анализируемого вещества (с экспериментальным подтверждением). В отношении редких разновидностей биологических образцов допустимо использовать меньшее количество источников. Отсутствие искажающего влияния компонентов холостого биологического образца, констатируется, как правило, если их сигнал по нижнему пределу количественного определения не превышает 20% для анализируемого вещества и 5% — для ВС.
В некоторых случаях может понадобиться исследование степени влияния метаболитов действующего вещества, а также продуктов деградации, образующихся при пробоподготовке, и одновременно применяемых лекарственных препаратов.
12. Если применимо (для нестабильных метаболитов, например, кислых метаболитов в эфире, нестабильных N-оксидов или глюкуронидов, соединений с лактонной структурой), необходимо оценить возможность обратного преобразования метаболита в исходное анализируемое вещество на различных этапах анализа (включая процедуры пробоподготовки или в извлечении для МС-анализа). Необходимо установить степень обратного преобразования и проанализировать его влияние на результаты исследования. На ранних этапах разработки нового химического соединения, пока его метаболизм еще не изучен, такую оценку осуществить невозможно. Тем не менее после получения в процессе разработки новых данных о метаболизме действующего вещества необходимо учитывать проблему обратного преобразования, что требует проведения частичной валидации.
В некоторых случаях достаточно сложно получить доступ к стандартным образцам исследуемых метаболитов. С другой стороны, обратное преобразование метаболита можно оценить, проводя повторный анализ активных образцов (образцов, содержащих анализируемые вещества, взятых от субъектов исследования или животных). Однако в этом случае нельзя исключить обратное преобразование в процессе пробоподготовки.
Открыть полный текст документа
Избирательная токсичность | справочник Пестициды.ru
О явлении избирательной токсичности говорят в медицине и животноводстве, однако чаще всего это понятие находит свое применение в растениеводстве, при определении свойств пестицидов, уничтожающих вредные организмы (бактерии, грибы, насекомые, сорняки и т.д.).
Избирательно токсичное вещество может оказывать действие тремя путями:
- Накапливаться во вредных организмах;
- Взаимодействовать с клеточными структурами, которые имеются только у вредного вида;
- Повреждать какую-нибудь жизненно важную химическую систему для вредного вида и не иметь большого значения для полезного растения.
 [2]
[2]
Для полезных растений, грибов, насекомых, человека и животных пестициды должны быть малотоксичны. В ряде случаев этого достичь очень трудно из-за схожей природы биохимико-физиологических процессов вредных и полезных организмов, или потому, что вредитель живет внутри защищаемого растения.[2]
В данном случае избирательная токсичность может определяться особенностями применения токсического действующего вещества, морфологии и поведения организмов, а также процессами проникновения пестицида, его превращения и выведения. Данную токсичность можно частично регулировать приготовлением специальных препаративных форм (микрокапсулированные суспензии, гранулы), направленного применения пестицидов на растения. [2]
Показатели селективности
Показатель селективности (ПС) или коэффициент избирательности (КИ) характеризуют, соответственно, степень выраженности селективности или избирательности. Показатель селективности определяется отношением среднетоксических доз (ЛД50):
ПС = ЛД50 одного организма / ЛД50 другого организма
Избирательность действия пестицида зависит от величины этого показателя. Во время разработки защитных мероприятий необходимо максимально сохранить энтомофагов, поэтому очень важно знать избирательность широко используемых пестицидов по отношению к наиболее распространенным в агроценозе энтомофагам. Для этого нужно определение отношения ЛД
Во время разработки защитных мероприятий необходимо максимально сохранить энтомофагов, поэтому очень важно знать избирательность широко используемых пестицидов по отношению к наиболее распространенным в агроценозе энтомофагам. Для этого нужно определение отношения ЛД
Препараты считаются для энтомофагов малоопасными, если они в течение десяти дней уменьшают их численность не выше, чем на 20%, умеренно опасными — на 20 – 50 %, опасными – более, чем на 50 % на протяжении 20 дней.[3]
Причины избирательности
Причинами избирательности служат биохимические и топографические факторы.
Биохимическая избирательность обусловлена способностью организмов детоксицировать действующее вещество или образовывать с ним неактивные комплексы (конъюганты) до того, как пестицид проникнет к месту действия. [3]
[3]
Например:
- гербицид Атразин (производное симмтриазина) быстро детоксицируется после поступления из почвы в корни кукурузы. Он превращается в гидроксиформу, вследствие чего не проникает в неизменном виде в хлоропласты, где реализуется его токсичность. Только этим обусловлена устойчивость кукурузы к данному препарату; [3]
- гербициды — производные феноксиуксусной кислоты (2,4-Д), связываясь с белками и некоторыми другими продуктами метаболизма, подвергаются иммобилизации в устойчивых растениях. Образовавшиеся конъюганты остаются в месте нанесения пестицида и не могут достичь меристематических тканей, где проявляется их токсичность;
- инсектицид Малатион является малотоксичным для теплокровных, поскольку в их организме он детоксицируется, превращаясь при этом в водорастворимые продукты, которые выводятся из организма. Но в организме насекомых он окисляется, и при этом образуется продукт еще более токсичный, чем действующее вещество Карбофоса.
 [3]
[3]
Топографическая избирательность
К примеру:
- устойчивость щитовки к инсектицидам объясняется тем, что она покрыта щитком, через который большинство препаратов не может проникнуть;
- древесница въедливая находится внутри одревесневших тканей, поэтому инсектициды на нее не попадают;
- ягодные кустарники и плодовые деревья имеют устойчивость ко многим гербицидам, потому что их корневая система залегает глубоко, куда гербициды почвенного действия не могут проникнуть.[3]
Избирательность пестицидов
Избирательная токсичность может определяться особенностями применения токсического действующего вещества, морфологии и поведения организмов, а также процессами проникновения пестицида, его превращения и выведения. Данную токсичность можно частично регулировать приготовлением специальных препаративных форм (микрокапсулированные суспензии, гранулы), направленного применения пестицидов на растения.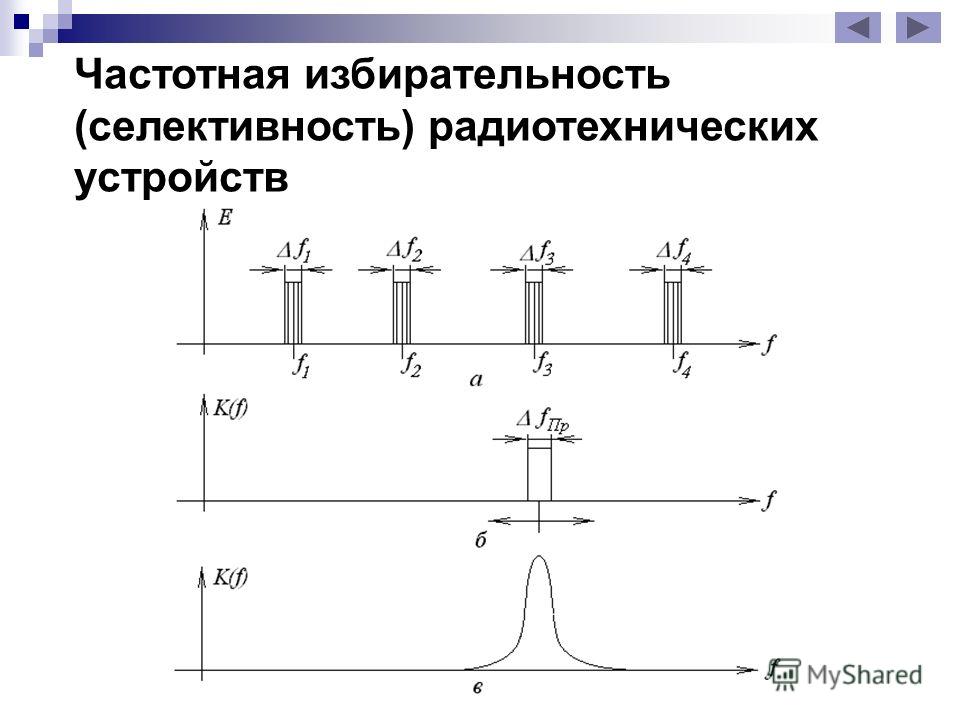
Сравнительно невысокая токсичность для человека некоторых фосфорорганических соединений (метатион, карбофос) обусловлена тем, что в организме млекопитающих эти вещества быстро разрушаются до нетоксичных веществ, а в организме насекомых этот процесс начинается с активации молекулы и идет медленнее.[1]
Полезные и вредные организмы имеют ряд цитологических различий, которые являются основой избирательности. Например, яды, которые действуют на нервную систему, токсичны для животных, но малотоксичны для растений, у которых ее нет. Вещества, разрушающие хлоропласты, для животных по понятным причинам практически не ядовиты.[1] Аналогичные примеры есть и при действии лекарственных препаратов на животных и человека. Например, большинство антибиотиков оказывает токсический эффект не только на микроорганизмы, но и на структуры нормальных клеток тела, и только средства из группы пенициллинов действуют исключительно на клеточные стенки бактерий, так как в организме человека и животных нет компонентов такого же строения.
Многие пестициды избирательно токсичны, поскольку воздействуют на биохимические процессы, жизненно важные или специфичные только для определенных организмов. К примеру, гербициды – производные мочевины и триазина и тиокарбонаты — малотоксичны для теплокровных животных и человека, поскольку избирательно нарушают процесс фотосинтеза, который присущ только растениям. Фосфорорганические соединения (инсектициды и акарициды) не угнетают развития и роста растений, так как действуют на процессы синаптической передачи нервных импульсов, которые растениям не присущи.[1]
Избирательность гербицидов
Для характеристики избирательного действия гербицидов применяют показатель селективности и индекс селективности. [2]
Показатель селективности представляет собой отношение показателя фитотоксичности одного препарата для разных видов растений. По нему можно установить, во сколько раз токсичность препарата больше для одного вида растения, чем для какого-то другого. Из двух сравниваемых объектов за первый принимается тот, у которого показатель больше, поэтому, чем сильнее показатель селективности превышает единицу, тем большей избирательностью характеризуется данный гербицид.[1][2]
Из двух сравниваемых объектов за первый принимается тот, у которого показатель больше, поэтому, чем сильнее показатель селективности превышает единицу, тем большей избирательностью характеризуется данный гербицид.[1][2]
Индекс селективности (ИС) — это отношение дозы, при использовании которой урожай снижается лишь незначительно, к дозе, уничтожающей большую часть сорных растений. Иными словами, эта величина показывает, во сколько раз доза, вызывающая достаточное снижение засоренности, меньше, чем доза, токсически действующая на культурные растения. Достаточно избирательным может быть признан препарат, который, поражая 80 и более процентов сорняков, не уничтожает или слабо угнетает (не больше 20 %) культурные растения.
Отношение доз, вызывающих 20%-ное снижение урожая культурных растений и 80%-ную гибель сорняков, условно принимают за единицу. Следовательно, чем больше единицы ЕС, тем более высокой избирательностью характеризуется гербицид. Пользуясь понятием ИС, можно характеризовать, насколько избирательность одного препарата больше или меньше избирательности другого. [2]
[2]
Близкие статьи
Ссылки:
Все статьи о токсикологии в разделе: Основы токсикологии
Статья составлена с использованием следующих материалов:
Литературные источники:
1.Ганиев М.М., Недорезков В.Д. Химические средства защиты растений. – М.: КолосС, 2006. – 248 с.
2.Груздев Г.С. Химическая защита растений. Под редакцией Г.С. Груздева — 3-е изд., перераб. и доп. — М.: Агропромиздат, 1987. — 415 с.: ил.
3.Зинченко В.А. Химическая защита растений: средства, технология и экологическая безопасность. – М.: «КолосС», 2012. – 127 с.
Свернуть Список всех источников| Тематика | Число статей |
| Аварийное восстановление | 369 |
| Авиационная медицина | 25.624 |
| Авиация | 90. 750 750 |
| Австралийское выражение | 9.067 |
| Австралия | 12 |
| Австрийское выражение | 21 |
| Австрия | 2 |
| Автоматика | 93.915 |
| Автоматическое регулирование | 983 |
| Автомобили | 66.038 |
| Авторское право | 244 |
| Агрономия | 7 |
| Агрохимия | 10.624 |
| Аддитивные технологии и 3D-печать | 153 |
| Административное деление | 28 |
| Административное право | 359 |
| Азартные игры | 965 |
| Айкидо | 4 |
| Аккумуляторы | 84 |
| Акридология | 4 |
| Акробатика | 3 |
| Активный отдых и экстремальный спорт | 5 |
| Акупунктура | 9 |
| Акустика раздел физики | 1. 712 712 |
| Акушерство | 459 |
| Албанский язык | 1 |
| Алгебра | 61 |
| Алжир | 7 |
| Алкалоиды | 132 |
| Аллергология | 164 |
| Альпинизм | 397 |
| Альтернативное урегулирование споров | 2.681 |
| Алюминиевая промышленность | 2.172 |
| Американская фондовая биржа | 13 |
| Американский вариант английского языка | 7 |
| Американский футбол | 48 |
| Американское выражение не вариант языка | 28.477 |
| Амфибии и рептилии | 6.029 |
| Анатомия | 11. 977 977 |
| Английский язык | 224 |
| Анестезиология | 256 |
| Антарктика | 186 |
| Антенны и волноводы | 8.741 |
| Антильские острова | 3 |
| Антимонопольное законодательство | 9 |
| Античность кроме мифологии | 443 |
| Антропология | 253 |
| Арабский язык | 661 |
| Арагон | 6 |
| Аргентина | 16 |
| Арго | 70 |
| Артиллерия | 6.940 |
| Архаизм | 1.353 |
| Археология | 1.180 |
| Архивное дело | 158 |
| Архитектура | 15. 262 262 |
| Астрология | 157 |
| Астрометрия | 29 |
| Астрономия | 7.876 |
| Астроспектроскопия | 8 |
| Астрофизика | 346 |
| Атомная и термоядерная энергетика | 13.426 |
| Аудиотехника | 13 |
| Аудит | 2.519 |
| Африка | 121 |
| Африканское выражение | 27 |
| Аэрогидродинамика | 17.514 |
| Аэродинамика | 245 |
| Аэропорты и управление водзушным движением | 197 |
| Аэрофотосъемка и топография | 29 |
| Базы данных | 1. 514 514 |
| Бактериология | 617 |
| Балет | 4 |
| Баллистика | 173 |
| Банки и банковское дело | 31.498 |
| Баскетбол | 711 |
| Бейсбол | 138 |
| Беларусь | 20 |
| Бельгийское выражение | 3 |
| Бережливое производство | 40 |
| Бетон | 164 |
| Библиография | 62 |
| Библиотечное дело | 208 |
| Библия | 2.818 |
| Бизнес | 73.455 |
| Бильярд | 414 |
| Биоакустика | 13 |
| Биогеография | 37 |
| Биология | 59. 850 850 |
| Биометрия | 98 |
| Бионика | 47 |
| Биотехнология | 3.724 |
| Биофизика | 218 |
| Биохимия | 5.880 |
| Биоэнергетика | 140 |
| Биржевой термин | 5.692 |
| Благотворительные организации | 31 |
| Бодибилдинг | 1 |
| Боевые искусства и единоборства | 17 |
| Боеприпасы | 13 |
| Бокс | 354 |
| Бондарное производство | 2 |
| Борьба | 113 |
| Борьба с вредителями | 324 |
| Борьба с коррупцией | 45 |
| Ботаника | 34. 868 868 |
| Бразилия | 16 |
| Британский вариант английского языка | 11 |
| Британское выражение не вариант языка | 4.710 |
| Бронетехника | 20.866 |
| Буддизм | 20 |
| Буквальное значение | 291 |
| Бурение | 21.058 |
| Бухгалтерский учет кроме аудита | 20.476 |
| Бытовая техника | 7.914 |
| Валютный рынок форекс | 39 |
| Вежливо | 21 |
| Вексельное право | 231 |
| Великобритания | 113 |
| Велосипеды кроме спорта | 1.810 |
| Велоспорт | 49 |
| Венгерский язык | 16 |
| Венерология | 27 |
| Венесуэла | 1 |
| Вентиляция | 319 |
| Верлан | 2 |
| Вертолёты | 244 |
| Ветеринария | 2. 925 925 |
| Ветроэнергетика | 5 |
| Взрывчатые вещества | 871 |
| Вибромониторинг | 361 |
| Видеозапись | 16 |
| Виноградарство | 191 |
| Виноделие | 1.029 |
| Вирусология | 693 |
| Внешняя политика | 1.102 |
| Внешняя торговля | 269 |
| Водные лыжи | 5 |
| Водные ресурсы | 523 |
| Водоснабжение | 3.323 |
| Военная авиация | 805 |
| Военно-морской флот | 1.491 |
| Военный жаргон | 1.484 |
| Военный термин | 307. 260 260 |
| Возвышенное выражение | 575 |
| Воздухоплавание | 812 |
| Волейбол | 20 |
| Волочение | 12 |
| Восклицание | 126 |
| Восточное выражение | 4 |
| Всемирная торговая организация | 224 |
| Вулканология | 113 |
| Вульгаризм | 315 |
| Выборы | 1.507 |
| Высокопарно | 321 |
| Высокочастотная электроника | 464 |
| Выставки | 132 |
| Вьетнамский язык | 6 |
| Вяжущие вещества | 1 |
| Гавайский | 29 |
| Газовые турбины | 3.346 |
| Газоперерабатывающие заводы | 4.972 |
| Галантерея | 314 |
| Гальванотехника | 48 |
| Гандбол | 5 |
| Гастроэнтерология | 374 |
| Гватемала | 1 |
| Гельминтология | 135 |
| Гематология | 1.152 |
| Геммология | 6 |
| Генеалогия | 24 |
| Генетика | 12.647 |
| Генная инженерия | 828 |
| Геоботаника | 12 |
| География | 15.256 |
| Геодезия | 1.510 |
| Геология | 67.969 |
| Геометрия | 368 |
| Геомеханика | 35 |
| Геоморфология | 187 |
| Геофизика | 16.893 |
| Геохимия | 165 |
| Геохронология | 27 |
| Геральдика | 326 |
| Германия | 56 |
| Герпетология вкл. с серпентологией | 219 |
| Гигиена | 196 |
| Гидравлика | 449 |
| Гидроакустика | 89 |
| Гидробиология | 2.753 |
| Гидрогеология | 187 |
| Гидрография | 683 |
| Гидрология | 10.001 |
| Гидрометрия | 66 |
| Гидромеханика | 79 |
| Гидропланы | 1 |
| Гидротехника | 217 |
| Гидроэлектростанции | 333 |
| Гимнастика | 65 |
| Гинекология | 1.203 |
| Гипсокартон и сис-мы сухого строительства | 1 |
| Гироскопы | 2.333 |
| Гистология | 412 |
| Гляциология | 110 |
| Голландский нидерландский язык | 35 |
| Голубиные гонки | 1 |
| Гольф | 110 |
| Гомеопатия | 35 |
| Гонки и автоспорт | 11 |
| Горное дело | 47.425 |
| Горные лыжи | 145 |
| Городская застройка | 17 |
| Горюче-смазочные материалы | 448 |
| ГОСТ | 1.342 |
| Гостиничное дело | 1.127 |
| Государственный аппарат и госуслуги | 59 |
| Гравиметрия | 34 |
| Гражданско-процессуальное право | 42 |
| Гражданское право | 211 |
| Грамматика | 2.162 |
| Гребной спорт | 34 |
| Греческий язык | 1.067 |
| Грубо | 2.398 |
| Грузовой транспорт | 67 |
| Гэльский шотландский язык | 1 |
| Дактилоскопия | 84 |
| Дамбы | 4 |
| Даосизм | 1 |
| Датский язык | 21 |
| Двигатели внутреннего сгорания | 617 |
| Дегустация | 26 |
| Деловая лексика | 1.032 |
| Делопроизводство | 62 |
| Демография | 282 |
| Дербетский диалект | 1 |
| Деревообработка | 6.584 |
| Дерматология | 579 |
| Детали машин | 823 |
| Детская речь | 376 |
| Дефектоскопия | 124 |
| Дзюдо | 10 |
| Диалектизм | 8.995 |
| Диетология | 42 |
| Дизайн | 45 |
| Дипломатия | 33.425 |
| Дистанционное зондирование Земли | 20 |
| Дистилляция | 134 |
| Договоры и контракты | 39 |
| Документооборот | 150 |
| Домашние животные | 162 |
| Доменное производство | 27 |
| Доминиканская Республика | 1 |
| Дорожное движение | 689 |
| Дорожное дело | 13.320 |
| Дорожное покрытие | 136 |
| Дорожное строительство | 386 |
| Дорожный знак | 49 |
| Дословно | 4 |
| Древнегреческая и древнеримская мифология | 696 |
| Древнегреческий язык | 117 |
| Древнееврейский язык | 23 |
| Европейский банк реконструкции и развития | 24.924 |
| Евросоюз | 1.233 |
| Египтология | 601 |
| Единицы измерений | 583 |
| Жаргон | 4.134 |
| Жаргон наркоманов | 3.341 |
| Железнодорожный термин | 33.619 |
| Жестяные изделия | 11 |
| Живопись | 591 |
| Животноводство | 7.531 |
| Журналистика терминология | 924 |
| Заболевания | 400 |
| Занятость | 399 |
| Звукозапись | 72 |
| Звукоподражание | 162 |
| Звукорежиссура | 9 |
| Здравоохранение | 1.805 |
| Землеведение | 9 |
| Зенитная артиллерия | 230 |
| Значение 1 | 2 |
| Значение 2 | 6 |
| Золотодобыча | 8.816 |
| Зоология | 8.542 |
| Зоотехния | 219 |
| Зубная имплантология | 4.505 |
| Зубчатые передачи | 936 |
| Иврит | 76 |
| Игрушки | 28 |
| Игры кроме спорта | 24 |
| Идиоматическое выражение | 15.168 |
| Идиш | 178 |
| Издательское дело | 647 |
| Измерительные приборы | 3.492 |
| Изоляция | 68 |
| ИКАО | 2 |
| Имена и фамилии | 4.750 |
| Иммиграция и гражданство | 56 |
| Иммунология | 19.225 |
| Имя | 3 |
| Имя собственное | 8.129 |
| Инвестиции | 5.112 |
| Индия | 57 |
| Индонезийское выражение | 16 |
| Инженерная геология | 295 |
| Инженерное дело | 100 |
| Иностранные дела | 3.195 |
| Инструменты | 1.056 |
| Интегральные схемы | 90 |
| Интернет | 6.500 |
| Информационная безопасность | 1.101 |
| Информационные технологии | 99.691 |
| Инфракрасная техника | 8 |
| Иран | 3 |
| Ирландский язык | 367 |
| Ирландское выражение | 6 |
| Ирония | 1.699 |
| Искусственный интеллект | 3.530 |
| Искусство | 3.175 |
| Ислам | 206 |
| Исландский язык | 12 |
| Испания | 2 |
| Испано-американский жаргон | 40 |
| Испанский язык | 308 |
| Исторические личности | 8 |
| История | 13.012 |
| Итальянский язык | 845 |
| Иудаизм | 16 |
| Ихтиология | 20.453 |
| Кабели и кабельное производство | 10.426 |
| Кадры | 1.737 |
| Казахстан | 20 |
| Калька | 22 |
| Каменные конструкции | 21 |
| Канада | 454 |
| Канадское выражение | 17 |
| Канализация и очистка сточных вод | 153 |
| Канцеляризм | 1.570 |
| Канцтовары | 10 |
| Карате | 12 |
| Карачаганак | 2.991 |
| Кардиология | 4.459 |
| Картография | 12.547 |
| Карточные игры | 1.149 |
| Карцинология | 33 |
| Карьерные работы | 103 |
| Каспий | 8.758 |
| Католицизм | 1.861 |
| Квантовая механика | 1.328 |
| Квантовая электроника | 120 |
| Керамика | 130 |
| Керамическая плитка | 9 |
| Кибернетика | 184 |
| Кинематограф | 10.151 |
| Киноосветительная аппаратура | 19 |
| Киносъёмочная аппаратура | 25 |
| Кинотехника | 90 |
| Кипр | 6 |
| Кирпич | 3 |
| Китай | 19 |
| Китайский язык | 796 |
| Классификация видов экон. деятельности | 289 |
| Классификация минералов | 5 |
| Климатология | 516 |
| Клинические исследования | 4.266 |
| Клише | 823 |
| Книжное/литературное выражение | 4.384 |
| Ковка | 15 |
| Кожевенная промышленность | 1.149 |
| Кокни рифмованный сленг | 2 |
| Коллекционирование | 5 |
| Коллоидная химия | 239 |
| Колумбия | 1 |
| Комиксы | 134 |
| Коммунальное хозяйство | 221 |
| Компьютерная графика | 690 |
| Компьютерная защита | 173 |
| Компьютерная томография | 21 |
| Компьютерные игры | 1.294 |
| Компьютерные сети | 17.474 |
| Компьютерный жаргон | 605 |
| Компьютеры | 22.502 |
| Конвертерное производство | 7 |
| Кондитерские изделия | 101 |
| Кондиционеры | 119 |
| Коневодство | 924 |
| Конный спорт | 281 |
| Консалтинг | 218 |
| Консервирование | 150 |
| Контекстуальное значение | 564 |
| Контроль качества и стандартизация | 14.176 |
| Конькобежный спорт | 14 |
| Кораблевождение | 1 |
| Коран | 4 |
| Корейский язык | 23 |
| Корма | 39 |
| Короткие текстовые сообщения | 11 |
| Корпоративное управление | 4.475 |
| Косметика и косметология | 1.779 |
| Космонавтика | 66.880 |
| Космос | 450 |
| Коста-Рика | 1 |
| Кофе | 22 |
| Красители | 255 |
| Красота и здоровье | 6 |
| Крахмально-паточная промышленность | 6 |
| Крикет | 1 |
| Криминалистика | 972 |
| Криминология | 7 |
| Криптография | 861 |
| Кристаллография | 674 |
| Куба | 1 |
| Кулинария | 10.621 |
| Культурология | 969 |
| Культы и прочие духовные практики | 1 |
| Кыргызстан | 30 |
| Лабораторное оборудование | 934 |
| Лазерная медицина | 946 |
| Лазеры | 2.437 |
| Лакокрасочные материалы | 509 |
| Ландшафтный дизайн | 68 |
| Ласкательно | 114 |
| Латиноамериканский сленг | 8 |
| Латиноамериканское выражение | 7 |
| Латынь | 3.096 |
| ЛГБТ | 41 |
| Легкая атлетика | 27 |
| Лесоводство | 39.215 |
| Лесозаготовка | 591 |
| Лесосплав | 66 |
| Лесохимия | 11 |
| Лимнология | 1 |
| Лингвистика | 15.948 |
| Линии электропередач | 15 |
| Литейное производство | 867 |
| Литература | 4.207 |
| Литология | 19 |
| Лифты | 143 |
| Логика | 643 |
| Логистика | 12.553 |
| Логопедия | 5 |
| Ложный друг переводчика | 7 |
| Лыжный спорт | 69 |
| Льдообразование | 267 |
| Магнетизм | 316 |
| Магнитная запись изображения | 5 |
| Магнитнорезонансная томография | 42 |
| Майкрософт | 25.979 |
| Макаров | 604.865 |
| Малайский язык | 15 |
| Малакология | 158 |
| Малярное дело | 98 |
| Маммология | 382 |
| Мануальная терапия и остеопатия | 1 |
| Маори | 197 |
| Маркетинг | 3.437 |
| Маркшейдерское дело | 9 |
| Марокко | 1 |
| Мартеновское производство | 11 |
| Масложировая промышленность | 43 |
| Математика | 124.309 |
| Математический анализ | 330 |
| Материаловедение | 2.053 |
| Машиностроение | 7.117 |
| Машины и механизмы | 909 |
| Мебель | 710 |
| Медико-биологические науки | 330 |
| Медицина | 251.095 |
| Медицинская техника | 5.065 |
| Международная торговля | 276 |
| Международное право | 1.078 |
| Международное частное право | 14 |
| Международные отношения | 1.109 |
| Международные перевозки | 433 |
| Международный валютный фонд | 10.600 |
| Мексиканское выражение | 17 |
| Мелиорация | 444 |
| Менеджмент | 3.880 |
| Местное название | 34 |
| Металловедение | 470 |
| Металлообработка | 64 |
| Металлургия | 47.851 |
| Метеорология | 7.869 |
| Метрология | 11.768 |
| Метрополитен и скоростной транспорт | 559 |
| Механика | 15.682 |
| Механика грунтов | 21 |
| Микология | 515 |
| Микробиология | 1.686 |
| Микроскопия | 377 |
| Микроэлектроника | 13.415 |
| Минералогия | 2.754 |
| Мифология | 1.435 |
| Млекопитающие | 9.396 |
| Мобильная и сотовая связь | 1.070 |
| Мода | 751 |
| Молдавский язык | 2 |
| Молекулярная биология | 2.567 |
| Молекулярная генетика | 853 |
| Моликпак | 2.428 |
| Молодёжный сленг | 91 |
| Молочное производство | 474 |
| Монтажное дело | 253 |
| Морское право | 18 |
| Морской термин | 98.194 |
| Морфология | 3 |
| Мостостроение | 2.082 |
| Мотоциклы | 268 |
| Мрачно | 10 |
| Музеи | 249 |
| Музыка | 11.200 |
| Музыкальные инструменты | 81 |
| Мультимедиа | 7 |
| Мультфильмы и мультипликация | 223 |
| Мучное производство | 69 |
| Мясное производство | 4.091 |
| Навигация | 414 |
| Надёжность | 61 |
| Название компании | 3 |
| Название лекарственного средства | 2.287 |
| Название организации | 4.006 |
| Название произведения | 11 |
| Названия учебных предметов | 105 |
| Налоги | 4.365 |
| Нанотехнологии | 56.809 |
| Напитки | 309 |
| Народное выражение | 185 |
| НАСА | 54 |
| Наследственное право | 66 |
| Насосы | 815 |
| Настольные игры | 11 |
| Настольный теннис | 144 |
| НАТО | 2.516 |
| Научно-исследовательская деятельность | 1.449 |
| Научный термин | 11.516 |
| Неаполитанское выражение | 1 |
| Небесная механика | 6 |
| Неврология | 1.457 |
| Негритянский жаргон | 158 |
| Недвижимость | 1.742 |
| Нейролингвистика | 6 |
| Нейронные сети | 653 |
| Нейропсихология | 99 |
| Нейрохирургия | 138 |
| Нелинейная оптика | 4 |
| Немецкий язык | 507 |
| Неодобрительно | 1.228 |
| Неологизм | 487 |
| Неорганическая химия | 841 |
| Непрерывная разливка | 5 |
| Нефрология | 174 |
| Нефтегазовая техника | 19.059 |
| Нефтеперерабатывающие заводы | 9.068 |
| Нефтепромысловый | 13.442 |
| Нефть | 95.314 |
| Нефть и газ | 60.203 |
| Нидерланды | 1 |
| Новозеландское выражение | 143 |
| Норвежский язык | 12 |
| Нотариальная практика | 10.526 |
| Нумизматика | 112 |
| Нью-Йоркская фондовая биржа | 9 |
| Обмотки | 9 |
| Обогащение полезных ископаемых | 708 |
| Обработка данных | 1.710 |
| Обработка кинофотоматериалов | 21 |
| Образное выражение | 4.251 |
| Образование | 12.905 |
| Обувь | 1.363 |
| Общая лексика | 1.512.414 |
| Общее право англосаксонская правовая система | 96 |
| Общественное питание | 1.557 |
| Общественные организации | 652 |
| Общественный транспорт | 17 |
| Обществоведение | 135 |
| Огнеупорные материалы | 154 |
| Одежда | 3.256 |
| Океанология океанография | 5.755 |
| Окна | 40 |
| Окружающая среда | 5.478 |
| Онкология | 3.038 |
| ООН Организация Объединенных Наций | 7.024 |
| Операционные системы | 224 |
| Оптика раздел физики | 1.573 |
| Оптическое волокно | 57 |
| Оптометрия | 4 |
| Организационно-правовые формы компаний | 91 |
| Организация производства | 1.151 |
| Органическая химия | 2.595 |
| Оргтехника | 605 |
| Орнитология | 16.866 |
| Ортопедия | 279 |
| Оружие и оружейное производство | 10.746 |
| Оружие массового поражения | 11.184 |
| Осветительные приборы кроме кино | 793 |
| Отопление | 263 |
| Официальный стиль | 2.944 |
| Офтальмология | 2.112 |
| Оффшоры | 15 |
| Охота и охотоведение | 996 |
| Охрана труда и техника безопасности | 2.678 |
| Ошибочное или неправильное | 106 |
| Паблик рилейшнз | 757 |
| Палеоботаника | 32 |
| Палеозоология | 2 |
| Палеонтология | 949 |
| Палинология | 114 |
| Панама | 4 |
| Паразитология | 147 |
| Парапланеризм | 3 |
| Парапсихология | 108 |
| Парикмахерское дело | 474 |
| Парусные суда | 55 |
| Парусный спорт | 18 |
| Парфюмерия | 13.318 |
| Паспорт безопасности вещества | 383 |
| Патенты | 16.934 |
| Патология | 405 |
| Педагогика | 17 |
| Педиатрия | 392 |
| Пенитенциарная система | 7 |
| Переключатели | 99 |
| Переносный смысл | 31.267 |
| Переплётное дело | 44 |
| Персидский язык фарси | 73 |
| Перу | 9 |
| Петанк | 7 |
| Петрография | 649 |
| Печатные платы | 374 |
| Пивоварение | 580 |
| Письменная речь | 10 |
| Пишущие машинки, машинопись | 6 |
| Пищевая промышленность | 23.286 |
| Плавание | 84 |
| Планирование | 346 |
| Пластмассы | 4.465 |
| Поговорка | 1.575 |
| Погрузочное оборудование | 366 |
| Подводное плавание | 982 |
| Подводные лодки | 425 |
| Пожарное дело и системы пожаротушения | 11.653 |
| Полезные ископаемые | 164 |
| Полиграфия | 31.712 |
| Полимеры | 29.549 |
| Полинезийское выражение | 4 |
| Политика | 26.206 |
| Политэкономия | 385 |
| Полицейский жаргон | 38 |
| Полиция | 2.321 |
| Полупроводники | 756 |
| Польский язык | 25 |
| Порошковая металлургия | 144 |
| Португальский язык | 43 |
| Пословица | 17.246 |
| Почвоведение | 993 |
| Почта | 474 |
| Почтительно | 13 |
| Пошив одежды и швейная промышленность | 1.169 |
| Поэзия терминология | 471 |
| Поэтический язык | 2.716 |
| Пояснительный вариант перевода | 739 |
| Права человека и правозащитная деят. | 26 |
| Правоохранительная деятельность | 349 |
| Православие | 3 |
| Прагматика | 15 |
| Превосходная степень | 22 |
| Презрительно | 998 |
| Пренебрежительно | 447 |
| Прессовое оборудование | 62 |
| Преступность | 370 |
| Приводы | 156 |
| Прикладная математика | 644 |
| Природные ресурсы и охрана природы | 58 |
| Программирование | 133.063 |
| Программное обеспечение | 3.436 |
| Проекторы | 6 |
| Проигрыватели виниловых дисков | 37 |
| Производственные помещения | 559 |
| Производство | 20.213 |
| Производство спирта | 281 |
| Производство электроэнергии | 21 |
| Прокат металлургия | 4.054 |
| Промышленная гигиена | 121 |
| Промышленность | 2.276 |
| Просторечие | 1.400 |
| Противовоздушная оборона | 204 |
| Протистология | 31 |
| Профессиональный жаргон | 1.240 |
| Профсоюзы | 2.561 |
| Процессуальное право | 106 |
| Прыжки в высоту | 1 |
| Прыжки на батуте | 1 |
| Прыжки с парашютом | 143 |
| Прыжки с трамплина | 12 |
| Прядение | 52 |
| Прямой и переносный смысл | 1.281 |
| Психиатрия | 4.668 |
| Психогигиена | 39 |
| Психолингвистика | 246 |
| Психология | 19.339 |
| Психопатология | 160 |
| Психотерапия | 1.031 |
| Психофизиология | 163 |
| Птицеводство | 396 |
| Публицистический стиль | 230 |
| Публичное право | 368 |
| Пульмонология | 619 |
| Пуэрто-риканский диалект испанского языка | 11 |
| Пчеловодство | 512 |
| Радио | 3.043 |
| Радиоастрономия | 32 |
| Радиобиология | 51 |
| Радиогеодезия | 12 |
| Радиолокация | 1.578 |
| Разговорная лексика | 148.765 |
| Ракетная техника | 1.456 |
| Распределение энергии | 4 |
| Расстройства речи | 5 |
| Растениеводство | 1.269 |
| Расходометрия | 205 |
| Расширение файла | 16 |
| Реактивные двигатели | 1 |
| Регби | 11 |
| Региональные выражения не варианты языка | 113 |
| Регулирование движения | 85 |
| Редко | 8.576 |
| Резиновая промышленность | 380 |
| Реклама | 37.311 |
| Релейная защита и автоматика | 1.115 |
| Религия | 36.778 |
| Рентгенография | 232 |
| Рентгенология | 628 |
| Риторика | 4.562 |
| Ритуал | 2 |
| Робототехника | 10.253 |
| Россия | 268 |
| Ругательство | 1.618 |
| Рудные месторождения | 37 |
| Рукоделие | 240 |
| Румынский язык | 7 |
| Русский язык | 322 |
| Рыбалка хобби | 241 |
| Рыбоводство | 10.750 |
| Рыболовство промысловое | 2.751 |
| Садоводство | 793 |
| Санитария | 224 |
| Санный спорт | 2 |
| Санскрит | 48 |
| Сантехника | 272 |
| Сарказм | 63 |
| Сахалин А | 1.140 |
| Сахалин Р | 4.233 |
| Сахалин Ю | 1.474 |
| Сахалин | 31.188 |
| Сахарное производство | 85 |
| Сварка | 4.381 |
| Связь | 7.969 |
| Северная Ирландия | 2 |
| Североамериканское выр. США, Канада | 34 |
| Седиментология | 1 |
| Сейсмология | 1.727 |
| Сейсмостойкость сооружений | 60 |
| Секс и психосексуальные субкультуры | 39 |
| Сексопатология | 258 |
| Селекция | 73 |
| Сельское хозяйство | 49.244 |
| Сенситометрия | 7 |
| Сестринское дело | 21 |
| Сигнализация | 197 |
| Силикатная промышленность | 11.106 |
| Силовая электроника | 168 |
| Синтоизм | 2 |
| Система наряд-допусков | 17 |
| Систематика организмов | 65 |
| Системы безопасности | 28.497 |
| Сказки | 162 |
| Скандинавская мифология | 123 |
| Скачки | 246 |
| Складское дело | 587 |
| Скорая медицинская помощь | 27 |
| Скульптура | 31 |
| Славянское выражение | 5 |
| Сленг | 63.553 |
| Слоистые пластики | 14 |
| Слуховые аппараты | 9 |
| Снабжение | 380 |
| Сниженный регистр | 549 |
| Сноуборд | 3 |
| Собаководство кинология | 1.549 |
| Собирательно | 2.162 |
| Советский термин или реалия | 931 |
| Современное выражение | 296 |
| Сокращение | 9.781 |
| Солнечная энергетика | 3.847 |
| Соматика | 238 |
| Сопротивление материалов | 215 |
| Социализм | 291 |
| Социальное обеспечение | 914 |
| Социальные сети | 248 |
| Социологический опрос | 11 |
| Социология | 6.017 |
| Союз-Аполлон | 3.098 |
| Спектроскопия | 1.363 |
| Спелеология | 2 |
| Специи | 51 |
| Спецслужбы и разведка | 2.173 |
| СПИД | 10 |
| Спичечное производство | 63 |
| Спорт | 21.676 |
| Спорттовары | 18 |
| Спутниковая связь | 51 |
| Средне-китайский | 16 |
| Средства индивидуальной защиты | 33 |
| Средства массовой информации | 14.459 |
| Станки | 605 |
| Старая орфография | 1 |
| Старомодное выходит из употребления | 28 |
| Старофранцузский | 3 |
| Статистика | 5.392 |
| Стеклоделие | 77 |
| Стеклотарная промышленность | 59 |
| Стерео | 8 |
| Стилистика | 95 |
| Стоматология | 26.701 |
| Стратиграфия | 58 |
| Страхование | 9.955 |
| Стрелковый спорт | 28 |
| Стрельба из лука | 28 |
| Строительная техника | 9 |
| Строительные конструкции | 992 |
| Строительные материалы | 1.867 |
| Строительство | 125.066 |
| Студенческая речь | 165 |
| Суда на воздушной подушке | 162 |
| Суда на подводных крыльях | 102 |
| Судебная лексика | 251 |
| Судебная медицина | 97 |
| Судостроение | 16.852 |
| Сухопутные силы | 70 |
| Сценарное мастерство | 11 |
| США | 1.449 |
| Сыроварение | 20 |
| Табачная промышленность | 452 |
| Табуированная обсценная лексика | 18.120 |
| Тавромахия | 1 |
| Тагмемика | 1 |
| Тайвань | 1 |
| Тайский язык | 12 |
| Таможенное дело | 936 |
| Танцы | 10 |
| Татарский язык | 3 |
| Театр | 2.559 |
| Текстильная промышленность | 46.827 |
| Тектоника | 108 |
| Телевидение | 3.879 |
| Телеграфия | 180 |
| Телекоммуникации | 90.620 |
| Телемеханика | 60 |
| Телефония | 1.586 |
| Тенгизшевройл | 7.532 |
| Теннис | 432 |
| Теория права | 65 |
| Тепличные технологии | 120 |
| Теплообменные аппараты | 189 |
| Теплопередача | 104 |
| Теплотехника | 14.693 |
| Теплоэнергетика | 66 |
| Тератология | 81 |
| Термин времен ГДР | 2 |
| Термодинамика | 86 |
| Техника | 545.536 |
| Типографика | 343 |
| Ткачество | 149 |
| Токсикология | 963 |
| Топография | 199 |
| Топология | 129 |
| Топоним | 272 |
| Торговая марка | 1.189 |
| Торговля | 3.813 |
| Торговый флот | 34 |
| Торпеды | 682 |
| Травматология | 219 |
| Трансплантология | 641 |
| Транспорт | 4.317 |
| Трансформаторы | 99 |
| Трибология | 371 |
| Трикотаж | 203 |
| Трубопроводная арматура | 176 |
| Трубопроводы | 4.802 |
| Трудовое право | 1.392 |
| Туннелестроение и проходческие работы | 32 |
| Турбины | 30 |
| Турецкий язык | 143 |
| Туризм | 3.716 |
| Турция | 1 |
| Тюремный жаргон | 321 |
| Тюркские языки | 11 |
| Тяжёлая атлетика | 12 |
| Увеличительно | 16 |
| Уголовное право | 1.956 |
| Уголовный жаргон | 303 |
| Уголь | 805 |
| Удобрения | 13 |
| Узкоплёночное кино | 4 |
| Украина | 57 |
| Украинский язык | 6 |
| Украинское выражение | 3 |
| Ультразвук | 14 |
| Уменьшительно | 461 |
| Университет | 826 |
| Уничижительно | 544 |
| Упаковка | 1.333 |
| Управление проектами | 1.227 |
| Управление рисками | 27 |
| Управление скважиной | 483 |
| Уровнеметрия | 154 |
| Урология | 607 |
| Уругвайский диалект испанского языка | 2 |
| Устаревшее | 37.773 |
| Устная речь | 39 |
| Утилизация отходов | 345 |
| Уфология | 68 |
| Уэльс | 9 |
| Фалеристика | 14 |
| Фамилия | 3 |
| Фамильярное выражение | 771 |
| Фантастика, фэнтези | 750 |
| Фармакология | 11.548 |
| Фармация | 5.985 |
| Федеральное бюро расследований | 14 |
| Фелинология | 4 |
| Ферментация | 4 |
| Фехтование | 97 |
| Фигурное катание | 193 |
| Физика металлов | 42 |
| Физика твёрдого тела | 249 |
| Физика | 10.086 |
| Физиология | 3.844 |
| Физиотерапия | 4 |
| Физическая химия | 979 |
| Филателия | 333 |
| Филиппины | 17 |
| Филология | 123 |
| Философия | 3.471 |
| Финансы | 24.766 |
| Финский язык | 53 |
| Фитопатология | 315 |
| Фольклор | 651 |
| Фонетика | 621 |
| Фортификация | 14 |
| Фотографическая запись звука | 1 |
| Фотография | 1.712 |
| Фотометрия | 2 |
| Фразеологизм | 10.219 |
| Французский язык | 2.209 |
| Фундаментостроение | 19 |
| Футбол | 2.394 |
| Хакерство | 39 |
| Хальцидология | 1 |
| Химическая номенклатура | 674 |
| Химическая промышленность | 720 |
| Химические волокна | 163 |
| Химические соединения | 967 |
| Химия | 66.066 |
| Хинди | 924 |
| Хирургия | 2.847 |
| Хлеб и хлебопечение | 342 |
| Хобби, увлечения, досуг | 194 |
| Хозйственное предпринимательское право | 135 |
| Хозяйственные общества и товарищества | 3 |
| Хоккей | 2.097 |
| Холодильная техника | 17.320 |
| Хореография | 37 |
| Христианство | 9.545 |
| Хроматография | 2.205 |
| Цветная металлургия | 157 |
| Цветоводство | 112 |
| Целлюлозно-бумажная промышленность | 2.115 |
| Цемент | 7.899 |
| Ценные бумаги | 1.024 |
| Центральная Америка | 2 |
| Церковный термин | 3.594 |
| Цинкование | 163 |
| Цирк | 69 |
| Цитаты, афоризмы и крылатые выражения | 2.029 |
| Цитогенетика | 40 |
| Цитология | 632 |
| Цифровая обработка звука | 14 |
| Цифровые валюты, криптовалюты, блокчейн | 60 |
| Часовое дело | 276 |
| Чаты и интернет-жаргон | 42 |
| Черчение | 190 |
| Чешский язык | 9 |
| Чили | 7 |
| Шахматы | 18.849 |
| Шведский язык | 7 |
| Швейцарское выражение | 47 |
| Школьное выражение | 591 |
| Шотландия | 593 |
| Шотландское выражение | 1.169 |
| Шоу-бизнес индустрия развлечений | 254 |
| Штамповка | 23 |
| Шутливо | 2.898 |
| Эволюция | 68 |
| Эвфемизм | 927 |
| Эзотерика | 204 |
| Эквадор | 1 |
| Экология | 43.305 |
| Эконометрика | 1.188 |
| Экономика | 132.655 |
| Экструзия | 29 |
| Электрические машины | 612 |
| Электричество | 2.117 |
| Электродвигатели | 17 |
| Электролиз | 4 |
| Электромедицина | 30 |
| Электрометаллургия | 32 |
| Электроника | 49.923 |
| Электронная почта | 140 |
| Электронная торговля | 17 |
| Электронно-лучевые трубки | 39 |
| Электротермия | 18 |
| Электротехника | 25.200 |
| Электротяга | 12 |
| Электрофорез | 48 |
| Электрохимия | 7.362 |
| Эмбриология | 361 |
| Эмоциональное выражение | 707 |
| Эндокринология | 332 |
| Энергетика | 60.579 |
| Энергосистемы | 4.752 |
| Энтомология | 14.606 |
| Эпидемиология | 217 |
| Эпистолярный жанр | 1 |
| Эскимосское выражение | 3 |
| Эсперанто | 7 |
| Эстонский язык | 1 |
| Этнография | 678 |
| Этнология | 1.011 |
| Этнопсихология | 10 |
| Этология | 187 |
| Ювелирное дело | 616 |
| Южная Америка | 29 |
| Южноафриканское выражение | 139 |
| Южнонидерландское выражение | 1 |
| Юридическая лексика | 121.253 |
| Ядерная физика | 2.464 |
| Ядерная химия | 50 |
| Ямайский английский | 67 |
| Япония | 6 |
| Японский язык | 242 |
| Яхтенный спорт | 2.198 |
| ASCII | 118 |
| Hi-Fi акустика | 919 |
| SAP технические термины | 7.504 |
| SAP финансы | 4.392 |
| SAP | 7.233 |
| Всего: | 7.848.729 |
ХиМиК.ru — ИЗБИРАТЕЛЬНОСТЬ АНАЛИЗА — Химическая энциклопедия
ИЗБИРАТЕЛЬНОСТЬ АНАЛИЗА (селективность), возможность метода или методики определять или обнаруживать искомый компонент (молекулы, ионы, функц. группы и т. д.) в присут. др. сопутствующих компонентов. Реагенты и хим. р-ции, обеспечивающие высокую избирательность анализа, наз. высокоизбирательными. Избирательность анализа характеризуют числом сопутствующих компонентов, к-рые не мешают определению искомого компонента, и макс. отношениями содержаний каждого из сопутствующих и искомого компонентов, при к-рых еще возможно надежное определение или обнаружение последнего (т. наз. факторами селективности). Мерой избирательности количеств. анализа служат обратные величины численных значений парциальной чувствительности gАК = (дgA)/(дCК), где gA — аналит. сигнал искомого компонента А при заданных содержаниях искомого и сопутствующих компонентов, СК — содержание сопутствующего компонента К. Избирательность многокомпонентной системы характеризуют матрицей парциальных чувствительностей (наз. также градуировочной матрицей данного способа анализа). Избирательность анализа зависит от избирательности отдельных стадий анализа и м. б. повышена маскированием мешающих компонентов или предварит. их отделением (концентрированием)с достаточно высоким коэф. разделения. В таких методах анализа, как полярография, эмиссионный спектральный анализ и др., избирательность анализа в конечной стадии определяется разрешающей способностью прибора, т. е. той миним. разностью между абсциссами сигналов искомого и сопутствующего компонентов на регистриреумой кривой (регистрограмме), при к-рой еще можно надежно обнаружить или измерить сигнал искомого компонента. Разрешающая способность прибора зависит от ширины сигнала. Наиб. высокой избирательностью анализа характеризуются методы многокомпонентного анализа — масс-спектрометрия, нейтронно-активац. анализ, газожидкостная хроматография и др.===
Исп. литература для статьи «ИЗБИРАТЕЛЬНОСТЬ АНАЛИЗА»: Данцер К., Тан Э., Мольх Д., Аналитика, пер. с нем., М., 1981; Чарыков А. К., «Ж. аналит. химии», 1984, т. 39, № 9, с. 1708-14. Б. Я. Каплан.
Страница «ИЗБИРАТЕЛЬНОСТЬ АНАЛИЗА» подготовлена по материалам химической энциклопедии.
Медийная избирательность – Новости – Научно-образовательный портал IQ – Национальный исследовательский университет «Высшая школа экономики»
Большое количество жертв в результате массовой трагедии может быть недостаточным фактором для того, чтобы событие стало темой номер один в СМИ. Значимую роль играет его включенность в информационную повестку. Это подтвердилось в результате исследования особенностей освещения в российских медиа трех трагедий, случившихся в один день в декабре 2016 года. Речь идет об убийстве российского посла в Турции, массовом отравлении «Боярышником» в Иркутске и террористической атаке в Берлине.
Теория повестки дня
В ходе исследования ученые использовали данные компании «Медиалогия», которая охватывает 40 тысяч русскоязычных СМИ – 1 974 газеты, 11 федеральных телеканалов, 34,905 онлайн изданий и 2,574 блогов. Также авторы проводили анализ освещения трагедий на данных базы печатной прессы «Интегрум». Полученные результаты отдельно отражены в статье, опубликованной в журнале «Мониторинг общественного мнения: экономические и социальные перемены».
В качестве основы исследователи опирались на теорию повестки дня (аgenda-setting theory). Ее суть заключается в том, что СМИ оказывают значительное влияние на общественность подбором того, что они освещают. То есть новости – это не просто отображение реальности, а сконструированная, отредактированная реальность. СМИ могут выделять определенные события, темы и проблемы, акцентируя на них внимание и заставляя тем самым аудиторию воспринимать их как чрезвычайно важные.
Два базисных предположения теории повестки дня лежат в основе большей части исследований:
1. Пресса и СМИ не отражают социальные проблемы: они конструируют их
2. Сконцентрированность СМИ на некоторых проблемах и предметах принуждает общественность рассматривать их как более важные по сравнению с другими.
Понятие повестки дня появилось в науке в конце 1960-х. Современные ученые используют также сетевую модель установления повестки дня (network agenda-setting model), допускающую возможность объединения разных повесток дня в «связки», когда обсуждение проблемы происходит в контексте других вопросов, связанных с ней тематически.
Почему «Боярышник» проиграл конкуренцию
Три трагических события – массовое отравление «Боярышником» (концентрат для принятия ванн) в Иркутске, убийство российского посла Андрея Карлова в Турции и теракт на рождественской ярмарке в Берлине произошли в один день – 19 декабря 2016 года. В результате отравления суррогатным алкоголем погибло 74 человека по данным на 23 декабря, в целом пострадавших – более 120. Это самое крупное массовое отравление на территории постсоветской России и бывших союзных республик. Теракт в Германии унес жизни 12 человек.
Логично было бы ожидать, что большее внимание СМИ привлечет событие с максимальным числом жертв, произошедшее на территории страны, считают авторы. Есть данные, которые позволяют делать такие предположения. Большее число жертв (и вообще наличие жертв) повышает внимание к трагедии. Об этом свидетельствует ряд исследований особенностей освещения террористических атак. Внимание со стороны СМИ к трагедиям может варьироваться под влиянием различных факторов. Интенсивность дискуссии оказывается более высокой при наличии жертв, а также в случае некоторых специфических типов террористических действий (например, атаки на борту самолетов), комментирует Анастасия Казун. Также, как отмечает исследователь, события в стране привлекают привлекают большее внимание, чем то, что происходит у соседей.
В случае терактов за рубежом на число сообщений в СМИ почти не влияет число жертв, важнее отношения между странами.
«В качестве факторов, определяющих число упоминаний страны в международных новостях, предлагается выделять характеристики страны (размер экономики, политическое влияние), ее взаимоотношения и связи с другими странами, а также уровень ее экономической и политической нестабильности.
К характеристикам страны, определяющим ее место в мировой новостной повестке, могут быть отнесены ее размер и влияние на политической арене. Так, значение имеют объемы экономики страны, ее военный потенциал и численность населения. Важным оказывается также наличие контактов, степень близости между странами. Так, например, наличие значительной географической или культурной дистанции будет снижать интерес к стране. Значение имеет и наличие двусторонних торговых отношений между странами: события в стране – торговом партнере будет освещаться с большей вероятностью, чем новости страны, торговые отношения с которой отсутствуют.
Впрочем, не все сводится к политическому влиянию и экономике страны, значение имеют и сами характеристики события. Исследователи отмечают, что СМИ в США с большей вероятностью обратят внимание на иностранные события, которые противоречат американским национальным ценностям».
Анастасия Казун
В ходе анализа исследователи сравнили степень внимания к трем трагедиям в разных типах СМИ. Так, например, за пять дней с 19 по 23 декабря тема убийства посла на Первом канале освещалась 54 минуты, теракт в Берлине около 15 минут, отравление алкоголем около 12 минут. В передаче «Воскресное время» 25 декабря турецкий сюжет занял около 20 минут, а темы берлинского теракта и алкогольного отравления по девять минут.
Похожая диспропорция с онлайн-изданиями. Авторы проанализировали 34,905 онлайн изданий. Выяснилось, что за те же дни убийству посла было посвящено 40,694 статей, отравлению 16,933, а теракту в Берлине 14,803. В печатных СМИ разброс не настолько сильный. И, как показали результаты анализа данных по базе «Интегрум», региональные газеты написали об иркутском инциденте наибольшее количество статей. Возможно, это связано с целевой аудиторией читателей, предполагают авторы.
Терроризм vs бедность и алкоголизм
Преступления в отношении дипломатов случаются редко. Последний раз российского посла убивали около 90 лет назад. Подобное событие касается не только России, но и всего мира, и оно связано с темой терроризма.
Напротив, гибель от алкогольной интоксикации – достаточно частое явление. Так за три квартала 2016 года в России было зафиксировано более 36 тысяч случаев алкогольного отравления, каждый четвертый из которых закончился летальным исходом.
То есть факт того, что инцидент с «Боярышником» в определенной мере остался «за кадром» на фоне других трагедий, может выглядеть как объяснимое явление. Тем не менее, согласно исследованиям, на степень внимания журналистов к тем или иным темам влияет множество факторов. Международная значимость – не главный, отмечают авторы. Например, лишение России права голоса в ПАСЕ было событием международного уровня, но не привлекло особого внимания СМИ и населения. Об этом, в том числе свидетельствуют данные социологических опросов.
Чтобы понять, почему алкогольное отравление проиграло «конкуренцию» в медиа, авторы проанализировали то, с какими особенностями освещались три трагедии. Подтвердилось, что характерной чертой дискуссии об убийстве посла оказалась ее встроенность в несколько других популярных сюжетов, среди которых и теракт в Берлине. Также тема соотносилась с вопросом действий России в Сирии и эпизодически упоминалась в дискуссии об ИГИЛ. Встречалась и совершенно неожиданная связка сюжета с открытием исторической сцены «Малого театра». Трагедия помешала В.Путину посетить торжественное мероприятие.
Отравление «Боярышником», напротив, освещалось как одиночное событие – без встроенности в актуальную повестку дня и, без связи с публикациями по проблеме бедности, вопросам социальной политики и алкоголизма. Основная дискуссия вокруг «Боярышника» сводилась к тому, «кто виноват и что делать». «Спецоперация» по поимке виновных в отравлении концентратом органически перекликалось со спецоперациями по поиску исполнителей теракта в Берлине и расследованием убийства посла. «Эти разные по своей сути трагедии подавались в рамках одного фрейма, при котором государство выступало в качестве сильной стороны, карающей виновных, а не слабой, допустившей трагедию», – отмечают авторы.
Объединение нескольких значимых проблем в ситуации с убийством посла можно интерпретировать, как стремление к их дополнительной проблематизации, отмечается в исследовании. Что касается отравления суррогатным алкоголем, то в повестке дня, традиционно обсуждаемой российскими медиа, по мнению авторов, не нашлось тем, которые можно было бы органически связать с отравлением алкоголем. СМИ мало и нечасто говорят о бедности, социальном неравенстве, алкоголизме. Эти вопросы в итоге не конструируются как проблемные, отмечают ученые
В ходе проведенного исследования модель сетевой повестки дня впервые применялась к дискуссиям в разных типах медиа на примере нескольких одновременных событий. До этого были работы, где сравнивалась сетевая повестка по одному вопросу в СМИ разных стран. Подобные исследования позволяют дополнить понимание того, как может работать государственная цензура, считают авторы. IQ
Авторы исследования:
Анастасия Казун, младший научный сотрудник Лаборатории экономико-социологических исследований НИУ ВШЭ Антон Казун, научный сотрудник Международного центра изучения институтов и развития НИУ ВШЭПодпишись на IQ.HSE
СМИ указали на избирательность «культуры отмены» и привели в пример Панарина — Газета.Ru
Из-за антикремлевских взглядов хоккеиста Артемия Панарина не многие на Западе знают о «скелетах» в его шкафу, пишет издание The Greanville Post.
«Когда речь идет о сексуальном насилии, «культура отмены» становится очень интенсивной. Движение «#MeToo» положило конец карьере многих знаменитостей-мужчин, которые, как утверждается, плохо обращались с женщинами. Многие утверждали, что они невиновны, но массы в социальных сетях действуют по правилу «верь женщинам», и само обвинение считается доказательством вины», — пишет издание.
В то же время можно заметить, что ключевым союзникам внешней политики США каким-то образом удается уворачиваться от пули за пулей. Прекрасный пример — звезда хоккея Артемий Панарин, считает автор публикации.
Панарин, например, не фанат других рас и публично заявил, что единственное, чего он никогда не сможет сделать, — это заняться сексом с чернокожей женщиной.
Как сообщала телекомпания NBC, он был вынужден извиниться за неприятие всех женщин определенной расы, но, как пишет The Greanville Post, его «темное прошлое» на этом не заканчивается.
«В 2011 году Панарина привлекли к уголовной ответственности в Латвии за нападение на девушку. Он несколько раз ударил 18-летнюю жертву за отказ заняться с ним сексом, а затем попытался изнасиловать ее. К счастью, его вовремя остановила полиция», — пишут «Дни.ру».
Андрей Назаров, еще одна звезда российского хоккея и бывший тренер Панарина, говорит, что единственная причина, по которой обвинения были сняты, — это взятки. Назаров заявил российским СМИ: «Я должен был спасти Артемия, иначе он был бы привлечен к уголовной ответственности по всей строгости закона… Артемия пришлось спасать за взятку».
«Знаменитости в Соединенных Штатах были «повержены», и их карьера была разрушена за гораздо меньшую цену. Однако, похоже, Панарин не опасается, что с ним такое случится. Вместо того, чтобы публично позорить его карьеру, он получил контракт на 81,5 млн долларов на 7 лет с хоккейной командой «Нью-Йорк Рейнджерс», без возмущения, без резких обвинений, только с похвалой за его антироссийскую позицию», — пишет издание.
4. Избирательность | RadioUniverse
Избирательность по соседнему каналу. Сигналы различных станций, достигающие приемной антенны, наводят в ней э. д. с. той или иной величины, и из всей массы этих сигналов приемник должен выделить только один — создаваемый нужной станцией. О том, как справляется приемник с этой задачей, судят по его избирательности, т. е. избирательность характеризует способность приемника выделять сигналы нужной станции и не пропускать сигналов других, мешающих приему, станций.
Выполнение этой задачи связано с значительными трудностями, так как количество радиостанций, работающих одновременно, очень велико. Выделять определенные станции приемник может только благодаря тому, что разные станции работают на разных частотах (на волнах разной длины). По международным соглашениям частоты радиовещательных станций должны отличаться одна от другой не менее чем на 10 кгц. Следовательно, приемник должен обладать способностью не пропускать сигналов, частота которых отличается более чем на 10 кгц от частоты его настройки, и по тому, насколько хорошо он задерживает сигналы этих станций, оценивают избирательность приемника. Этот показатель называют избирательностью по соседнему каналу.
От чего же зависит избирательность приемника и какими элементами его схемы она обеспечивается?
В современных радиовещательных приемниках выделение сигналов нужной частоты основано на использовании принципа резонанса; поэтому все избирательные способности приемника зависят от того, насколько хорошо выражены резонансные свойства его колебательных контуров. Решающее значение в этом имеют качество и количество этих контуров. Чем лучше колебательные контуры, т. е. чем меньше всякого рода потери в них, и чем больше контуров, встречает сигнал на пути своего прохождения в приемнике, тем лучше отсеиваются, отфильтровываются все помехи и тем лучше будет, следовательно, избирательность приемника.
Наглядное представление об избирательности дает резонансная характеристика приемника, которая показывает, как изменяется чувствительность приемника от частоты подводимого сигнала при неизменной настройке его на одну и ту же частоту. Наибольшая чувствительность будет, естественно, получаться при настройке приемника в резонанс на принимаемый сигнал. По мере того как частота сигнала
будет отходить от частоты настройки, чувствительности приемника будет уменьшаться, т. е. для получения на выходе приемника одной и той же постоянной звуковой мощности (или напряжения) необходимо подводить к его входу все большее и большее напряжение сигнала.
Резонансная характеристика может быть изображена графически в «иде кривой, у которой по горизонтальной оси отложена частота сигнала, а по вертикальной — напряжение на выходе приемника. Такая характеристика может быть названа также характеристикой избирательности.
На фиг. 2 изображены две резонансные характеристики — одна (кривая а) для приемника с худшей и другая (кривая б) для приемника с лучшей избирательностью.
В первом случае сигнал мешающей станции, частота которой отличается на 10 кгц от частоты принимаемой станции, создает на выходе приемника напряжение всего лишь в два раза меньшее, чем полезный сигнал. Это значит, что если обе станции будут воздействовать на антенну одинаково, то, настроившись на нужную нам станцию (fприн), мы одновременно будем слышать, хотя и значительно слабее, и передачу мешающей (соседней по частоте) станции.
Вторая кривая говорит о том, что сигналы мешающей станции будут ослаблены уже не в 2, а в 10 раз. Совершенно очевидно, что в этом случае действие помехи будет ощущаться значительно меньше. Это значит, что в первом случае чувствительность приемника для сигналов, отличающихся по частоте от принимаемого на 10 кгц, уменьшается в 2 раза, а во втором случае — в 10 раз.
По ГОСТ для избирательности установлены такие нормы: у приемников 1-го класса чувствительность при расстройке на 10 кгц должна уменьшаться не менее чем в 200 раз, у приемников 2-го класса — не менее чем в 20 раз и у приемников 3-го класса — не менее чем в 10 раз.
В приемниках супергетеродинного типа избирательность по соседнему каналу определяется главным образом контурами промежуточной частоты.
Избирательность по зеркальному каналу. Избирательность по соседнему каналу является параметром, одинаково важным для всех приемников независимо от того, по какому принципу они работают, по принципу прямого усиления или по супергетеродинному принципу. Но для супергетеродинных приемников необходимо учитывать еще помехи, обусловленные особенностями этого метода приема.
Дело в том, что в супергетеродине принимаемые сигналы любой станции, на которую настроен приемник, преобразуются в сигналы так называемой промежуточной частоты, которая остается неизменной независимо от частоты принимаемой станции. Этот процесс преобразования состоит в том, что на напряжение принятого сигнала накладывается напряжение другой частоты, создаваемое в самом приемнике вспомогательным гетеродином. В результате такого наложения двух колебаний разных частот и выделяются колебания новой промежуточной частоты. Эта частота равна разности частот накладываемых друг на друга колебаний.
Но в этом принципе таится скрытая опасность появления помехи. Из антенны в приемник могут попасть сигналы двух разных станций, каждая из которых отличается по частоте от гетеродина fгem на одну и туже величину, но частота одной из этих станций, например принимаемой станции fnpин, будет меньше, а частота другой — мешающей станции fмеш — больше частоты гетеродина, как это показано условно на фиг. 3. Сигналы этих станций, взаимодействуя с колебаниями гетеродина, будут создавать два вида колебаний на одной и той же промежуточной частоте. Пусть, например, частота сигнала принимаемой станции fсигн = 10 000 кгц, промежуточная частота fпром = 465 кгц и частота гетеродина fгет выше принимаемой на величину промежуточной частоты. Следовательно, fгет = fnpин + fnpом = 10 000 + 465 = 10465 кгц. Но такая же промежуточная частота fпром = 465 кгц получится и от мешающей станции, работающей на частоте fмеш = 10930 кгц, так как fмеш — fгет = 10 930 — 10 465 = 465 кгц. Эти полученные в приемнике колебания будут создавать сильные взаимные помехи в виде свистов и искажений, нарушающие нормальный прием.
Из фиг. 3 видно, что по отношению к гетеродину частоты мешающего и принимаемого сигналов расположены симметрично (одна из них представляет как бы зеркальное изображение другой), поэтому помехи такого рода называются зеркальными или симметричными.
Избавиться от подобной помехи можно только, не пропуская в приемник, к его преобразователю, сигналов зеркального канала. Эту задачу могут выполнить лишь контуры приемника, находящиеся между антенной и преобразовательной лампой и настроенные на частоту принимаемого сигнала.
Поэтому в отличие от избирательности по соседнему каналу, которую обеспечивают контуры промежуточной частоты, избирательность по зеркальному каналу обеспечивают контуры высокой частоты. Чем лучше качество высокочастотных контуров и чем больше их, тем сильнее ослабляются помехи по зеркальному каналу.
Избирательность по зеркальному каналу для одного и того же приемника оказывается различной на разных диапазонах; меньше всего ощущаются помехи этого рода на длинных волнах и сильнее всего — на коротких. Кроме того, они зависят от величины промежуточной частоты: у приемников с низкой промежуточной частотой (110—115 кгц) зеркальные помехи выражены сильнее, чем у приемников с более высокой промежуточной частотой (465 кгц).
Согласно ГОСТ ослабление зеркального канала у приемников 1-го класса должно быть не менее 1 000 раз на длинных, не менее 300 раз на средних и не менее 18 раз на коротких волнах; у приемников 2-го класса — не менее 60 раз на длинных, не менее 30 раз на средних и не менее 4 раз на коротких волнах; у приемников 3-го класса — не менее 20 раз на длинных и не менее 10 раз на средних волнах.
Ослабление сигналов с частотой, равной промежуточной. Другим источником помех, характерных только для приемников супергетеродинного типа, являются сигналы станций, частота которых равна промежуточной частоте приемника. Такие сигналы, проникая в приемник, приводят к результатам, сходным с описанными выше зеркальными помехами: внешняя помеха, взаимодействуя с колебаниями промежуточной частоты, создаваемыми в самом приемнике, приведет к появлению искажений и свистов, нарушающих нормальный прием.
Для борьбы с помехами этого рода на входе приемника включают так называемые фильтры-пробки — контуры, настроенные на промежуточную частоту, которые закрывают доступ в приемник сигналам на этой частоте. Чем тщательнее подобран фильтр, тем лучше устраняются помехи этого вида.
ГОСТ устанавливает нормы на ослабление помех и этого рода для приемников разных классов. Сигналы с частотой, равной промежуточной, должны ослабляться на пути от антенны к преобразователю в приемниках 1-го класса не менее чем в 100 раз, в приемниках 2-го класса — не менее чем в 50 раз и в приемниках 3-го класса — не менее чем в 10 раз.
Избирательность — обзор | ScienceDirect Topics
2.34.5.3 Повышенная селективность
В настоящее время селективность ферментов становится мощным активом ферментно-опосредованного асимметричного синтеза из-за растущей потребности фармацевтической промышленности в оптически чистых промежуточных соединениях [109].
В общем, селективность ферментов включает [113]:
- •
-
селективность субстрата — способность различать и воздействовать на подмножество соединений в более крупной группе химически родственных соединений;
- •
-
стереоселективность — способность действовать исключительно на один энантиомер или диастереомер;
- •
-
региоселективность — способность действовать исключительно на одно место в молекуле;
- •
-
Селективность функциональной группы — способность действовать на одну функциональную группу избирательно в присутствии других в равной степени реакционноспособных или более реакционноспособных функциональных групп, например, селективное ацилирование аминоспиртов [114].
Хотя резкое изменение селективности фермента с помощью генной инженерии было прекрасно продемонстрировано [115], есть также множество привлекательных примеров, в которых селективность фермента была изменена различными методами иммобилизации, например ковалентным связыванием, захватом и т. Д. и простая адсорбция. В нескольких крайних случаях было продемонстрировано, что неселективный фермент, такой как хлоропероксидаза, трансформировался в стереоселективный фермент после иммобилизации [116]; S -селективная липаза также была преобразована в R -селективную CR липазу путем ковалентной иммобилизации [117].
В целом селективность, на которую могут влиять методы иммобилизации, можно разделить на следующие категории в зависимости от источника эффекта:
- 1.
-
Селективность, контролируемая носителем
- •
-
селективность, контролируемая размером пор;
- •
-
селективность, управляемая диффузией; и
- •
-
селективность связывания, контролируемая химическим составом.
- 2.
-
избирательность, контролируемая конформацией
- •
-
избирательность, контролируемая микросредой и
- •
-
активная избирательность, контролируемая центром.
Влияние стерических затруднений на селективность фермента, например, карту продукта, наблюдали в 1970-х годах [118]. Было обнаружено, что на структуру продуктов расщепления белков, катализируемого субтилзином, иммобилизованным CPG, может влиять размер пор используемого носителя [119].Точно так же иммобилизованная аденозинтрифосфат (АТФ) дезаминаза, β-галактозидаза [121] и протеазы также имеют разные карты продуктов по сравнению с соответствующими нативными ферментами [120]. Ковалентно иммобилизованная на глиоксаль агарозе урокиназа имеет разную селективность [120]. α-Амилаза, иммобилизованная на диоксиде кремния [120] или ковалентно связанная с CNBr-активированной карбоксиметилцеллюлозой [118], дает продукты, состав которых отличается от состава природного фермента. Это во многом объяснялось тем, что размер пор, в которых расположены молекулы фермента, определяет доступность субстратов в зависимости от их размера.
Об энантиоселективности, контролируемой диффузией, недавно сообщалось после исследования энантиоселективности липазы CAL-B при переэтерификации в органических растворителях [123]. Впервые сообщалось, что диффузия может снижать энантиоселективность ферментов. Важным следствием этого открытия является то, что при скрининге фермента на разделение рацемических соединений важно гарантировать, что выбранный препарат фермента не имеет диффузионных ограничений; в противном случае реальный потенциал фермента может быть проигнорирован [122, 123].
Иммобилизация может не только изменить селективность (карту продукта или энантиоселективность), но наличие ограничений диффузии также может повлиять на селективность реакции между двумя реакциями, которые могут происходить параллельно в одной и той же реакционной системе. Одним из примеров является кинетически контролируемый синтез пептидов или β-лактамных антибиотиков, в котором один из реагентов, например сложный эфир или амид аминокислоты (или обычно называемый активным донором ацила), может быть интегрирован в желаемый продукт ( S ) или гидролизуется до нежелательной аминокислоты ( H ) [17].Таким образом, молярное соотношение S / H рассматривалось как критерий жизнеспособности соответствующего процесса [124].
Как и в случае селективности, контролируемой конформацией, часто бывает трудно отличить эффект микросреды от изменения конформации. Например, включение липазы Rhizopus miehei lipase (RML) в гелевое волокно из ацетата целлюлозы и TiO 2 улучшило селективность гидролиза 1,2-диацетоксипропана по сравнению с таковыми нативных ферментов [125] и энантиоселективность пегилированной липазы. Липаза Pseudomonas cepacia (PCL) была увеличена в три раза за счет захвата гранул геля из альгината кальция [126].В таких случаях липазы могут принимать конформацию, отличную от конформации нативных ферментов, из-за взаимодействия между носителем и ферментом (изменение конформации фермента) или из-за влияния микроокружения (градиент pH).
Влияние микросреды на селективность фермента, однако, было ясно продемонстрировано для 1,2-α-маннозидазы, для которой был применен метод двойной иммобилизации, адсорбция на фарфоровой глине или целлюлозе DE-52 с последующим захватом альгинатными шариками. использовал; Полученный спектр продуктов зависел от носителя, используемого для адсорбции перед захватом в альгинате натрия [127], предполагая, что конформация фермента могла в значительной степени определяться носителем во время адсорбции.Точно так же селективность субстрата декстрансукреазы, адсорбированной на DEAE-целлюлозе, отличалась от селективности нативного фермента [120].
Наиболее поразительно то, что недавно было обнаружено, что энантиоселективность липазы Candida rugosa (CRL), иммобилизованной на кремнеземе, активированном 2,4,6-трихлор-1,3,5-триазином, была примерно в 7 раз выше, чем у растворимого фермент, тогда как CLR, иммобилизованный на агарозе, активированной тозилатом, был только в 4 раза более селективным, чем нативный фермент [128], подразумевая, что химическая модификация фермента активными носителями также может влиять на селективность фермента.
Аналогичным образом, недавно было обнаружено, что на активность и селективность фермента также может влиять природа боковых связывающих функциональностей или функциональных групп, которые фактически не являются частями, участвующими в связывании. Например, энантиоселективность алкилсульфатазы, иммобилизованной на анионообменниках, таких как DEAE-сефадекс, TEAE-целлюлоза и Ecetola-целлюлоза, сильно различается в зависимости от боковых ионных групп. Иммобилизация алкилсульфатазы на эцетола-целлюлозе в несколько раз повысила селективность гидролиза втор-алкилсульфатов.Что касается того факта, что TEAE-целлюлоза и целлюлоза Ecetola различаются в основном спейсером, селективность иммобилизованного фермента в основном определяется боковой цепью и спейсером связывающих функций. Повышение селективности можно объяснить тем фактом, что заряженные группы могут быть способны приближаться к определенным отрицательно заряженным доменам или сайтам на поверхности белка, что приводит к эффекту ориентации (например, к активным сайтам) [129].
Селективность, контролируемая конформацией, также недавно наблюдалась для так называемых методов молекулярного импринтинга (MIT), которые основаны на гипотезе о том, что конформация, индуцированная лигандом, может быть заморожена физическими или химическими средствами, такими как лиофилизация или сшивание. или молекулярное ограничение.Одно из возможных объяснений состоит в том, что популяция некоторых конформеров ферментов увеличивается за счет используемых селекторов конформеров и, следовательно, селективность ферментов по отношению к некоторым субстратам может быть улучшена, как это показано на примере так называемых MIT [85].
При улучшении селективности ферментов путем иммобилизации важно обращать внимание на инженерию среды, потому что селективность, контролируемая микросредой, связана не только с выбранным носителем, но и с используемой средой. Иммобилизация фермента часто приводит к изменению оптимального pH или температуры.Таким образом, характеристики фермента, такие как активность и селективность, которые тесно связаны с pH и температурой, могут быть соответственно изменены. Оптимальный pH для селективности экспрессии также может отличаться от pH природного фермента; это было показано недавним исследованием растворения метилового эфира ( R, S ) -миндальной кислоты, катализируемого иммобилизованной липазой CRL [126]. В этом процессе степень повышения селективности сильно зависела от pH используемой реакционной среды.
В целом улучшение энантиоселективности ферментов путем иммобилизации может быть привлекательным из-за его простоты и универсальности, а также из-за того, что оно обычно устраняет необходимость в подробной структурной информации.
Как обсуждалось выше, иммобилизацию фермента можно рассматривать как процесс модификации. Неудивительно, что эффективность иммобилизованного фермента зависит от модификации (например, условий иммобилизации) природы модификатора (т.е.е., выбранные перевозчики). и природа ферментов (источник, чистота и штамм), которые необходимо изменить.
Что касается сходства между иммобилизацией фермента и химической модификацией [47, 48], многие методы и принципы, которые широко используются в химической модификации ферментов для повышения функциональности фермента, также могут быть использованы для улучшения характеристик носителя. -связанные иммобилизованные ферменты.
Например, стабилизация ферментов химической модификацией обычно может быть достигнута с помощью двух основных подходов, а именно ригидизации ферментного каркаса с использованием бифункционального сшивающего агента и инженерии микроокружения путем введения новых функциональных групп, которые способствуют развитию гидрофобное взаимодействие (за счет гидрофобизации поверхности фермента) или гидрофилизация поверхности фермента (за счет смягчения неблагоприятного гидрофобного взаимодействия) или образование новых солевых мостиков или водородных связей (за счет введения полярных групп) [130, 131].Точно так же эти два принципа все чаще применялись для улучшения характеристик фермента, например, стабильности, селективности и активности [30; и цитированные там ссылки].
Синонимы избирательности, антонимы избирательности | Тезаурус Мерриам-Вебстера
Тезаурус
Синонимы и антонимы слова
selectivity как в дискриминацияСинонимы и близкие синонимы для селективности
Антонимы и близкие антонимы для избирательности
См. Определение словаряПроизошла ошибка при настройке пользовательского файла cookie
Этот сайт использует файлы cookie для повышения производительности.Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка вашего браузера для приема файлов cookie
Существует множество причин, по которым cookie не может быть установлен правильно. Ниже приведены наиболее частые причины:
- В вашем браузере отключены файлы cookie. Вам необходимо сбросить настройки своего браузера, чтобы он принимал файлы cookie, или чтобы спросить вас, хотите ли вы принимать файлы cookie.
- Ваш браузер спрашивает вас, хотите ли вы принимать файлы cookie, и вы отказались.Чтобы принять файлы cookie с этого сайта, нажмите кнопку «Назад» и примите файлы cookie.
- Ваш браузер не поддерживает файлы cookie. Если вы подозреваете это, попробуйте другой браузер.
- Дата на вашем компьютере в прошлом. Если часы вашего компьютера показывают дату до 1 января 1970 г., браузер автоматически забудет файл cookie. Чтобы исправить это, установите правильное время и дату на своем компьютере.
- Вы установили приложение, которое отслеживает или блокирует установку файлов cookie.Вы должны отключить приложение при входе в систему или проконсультироваться с системным администратором.
Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу. Чтобы предоставить доступ без файлов cookie потребует, чтобы сайт создавал новый сеанс для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файле cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в cookie; никакая другая информация не фиксируется.
Как правило, в cookie-файлах может храниться только информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта. Например, сайт не может определить ваше имя электронной почты, пока вы не введете его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступа к остальной части вашего компьютера, и только сайт, который создал файл cookie, может его прочитать.
|
Определенные природные вещества в организме, такие как нейротрансмиттеры и гормоны, воздействуют на специфические рецепторы на поверхности клеток.Когда эти вещества связываются с рецептором в клетке, они стимулируют этот рецептор выполнять свою функцию, которая заключается в том, чтобы производить или подавлять определенное действие в клетке. Лекарства также могут нацеливаться на эти рецепторы и связываться с ними. Некоторые лекарства действуют как агонисты, стимулируя рецептор таким же образом, как и естественные вещества организма. Другие действуют как антагонисты, блокируя действие природного вещества на рецептор. Каждый тип рецептора имеет множество подтипов, и лекарства могут действовать на один или несколько подтипов рецепторов. |
|||
|
Препараты, нацеленные на рецептор |
|||
|
Адреналин и норадреналин |
Реакции «бей или беги»: сужение кровеносных сосудов кожи, пищеварительного тракта и мочевыводящих путей Расщепление глюкозы в печени (высвобождение энергии) Снижение активности желудка и кишечника Сокращение гладкой мускулатуры половых органов и мочевыводящих путей |
Агонист: метоксамин и фенилэфрин Антагонист: доксазозин, празозин, тамсулозин и теразозин |
|
|
Адреналин и норадреналин |
Снижение секреции инсулина, скопление тромбоцитов, сужение кровеносных сосудов кожи и кишечника и высвобождение норадреналина из нервов |
||
|
Адреналин и норадреналин |
Повышение частоты сердечных сокращений, силы сердечных сокращений и секреции ренина (гормона, контролирующего кровяное давление) |
Агонист: добутамин и изопротеренол Антагонист: бета-адреноблокаторы (используются для лечения гипертонии и болезней сердца), такие как атенолол и метопролол |
|
|
Адреналин и норадреналин |
Расширение гладкой мускулатуры кровеносных сосудов, дыхательных путей, пищеварительного тракта и мочевыводящих путей Распад гликогена в скелетных мышцах (высвобождение глюкозы для получения энергии) |
Агонист: альбутерол, изоэтарин и тербуталин |
|
|
Уменьшение частоты сердечных сокращений и силы сердечных сокращений Расширение кровеносных сосудов по всему телу Повышение активности желудка, кишечника, мочевого пузыря, слюнных, слезных и потовых желез |
Агонист: Бетанехол и карбахол Антагонист: атропин, ипратропий и скополамин |
||
|
Сокращение скелетных мышц |
Агонист: обычно не используется Антагонист: атракуриум, панкуроний и тубокурарин |
||
|
Производство аллергической реакции Сокращение мышц дыхательных путей и пищеварительного тракта Расширение мелких кровеносных сосудов |
Агонист: обычно не используется Антагонист: цетиризин, хлорфенирамин, клемастин, дифенгидрамин, фексофенадин и лоратадин |
||
|
Стимуляция желудочного секрета |
Агонист: обычно не используется Антагонист: Циметидин, фамотидин и низатидин |
||
|
Сужение кровеносных сосудов головного мозга Стимуляция активности (моторики) пищеварительного тракта Сокращение кровеносных сосудов Влияние на сон, память, сенсорное восприятие, регуляцию температуры, настроение, аппетит и секрецию гормонов |
Частичный агонист: Буспирон Агонист *: Сумитриптан и золмитриптан Антагонист: Метизергид и ондансетрон |
||
|
Участие в движении, настроении, мышлении, обучении и поиске вознаграждения Также увеличивает приток крови к почкам, что способствует увеличению выведения мочи. |
Агонист: прамипексол и ропинирол Антагонист: оланзапин и рисперидон |
||
Селективность в аналитической химии (Рекомендации ИЮПАК 2001 г.)
_Основные моменты теоретической и прикладной химии
_Селективность в аналитической химии (Рекомендации ИЮПАК 2001 г.)
Дж.Вессман, Р.И. Стефан, Дж. Ф. ван Стаден, К. Данцер, В. Линднер, Д. Т. Бернс, А. Файгель, Х. Мюллер
Чистая и прикладная химия, Vol. 73, No. 8, pp. 1381-1386 01)
Селективность — одно из ключевых свойств аналитической химии. Однако определения в рамках ИЮПАК были довольно расплывчатыми. В аналитическом химическом сообществе также имелось досадное совпадение терминов «селективность» и «специфичность», что сбивало с толку. В качестве средства правовой защиты в 1999 г. в отделе аналитической химии ИЮПАК был начат проект, который был завершен в Брисбене 01.
В итоговом документе говорится, что термин «селективность» развился параллельно с развитием более чувствительных и различающих методов и что в процессе распознавания используются несколько видов взаимодействий.
Селективность в методе достигается комбинацией нескольких этапов создания селективности, как показано на примере ЖХ-МС-МС (с селективностью разделения и обнаружения) и с помощью массивов датчиков, где вводится вычислительная селективность.
Избирательность можно выразить качественно разными способами, но, что наиболее важно, избирательность — это то, что можно оценивать в отличие от специфичности, которая является абсолютной.С другой стороны, вычислить степень селективности непросто, и было сделано много попыток. Поэтому подход к расчетам, полезный для практикующего аналитика, по-прежнему желателен.
Рекомендация IUPAC 01 гласит, что следует поощрять избирательность, а специфичность не поощрять, поскольку последняя неверна. Метод либо конкретен, либо нет, мало, если какие-то конкретные методы. С семантической точки зрения было получено выражение, что «избирательность — это состояние или качество тщательного выбора».
Рекомендуемое определение селективности: Селективность относится к степени, в которой метод может быть использован для определения определенных аналитов в смесях или матрицах без вмешательства со стороны других компонентов с аналогичным поведением.
Опубликовано в Интернете: 2009-09-01 Опубликовано в печати: 2002-01 © 2014 by Walter de Gruyter GmbH & Co. Информация о межбелковых взаимодействиях (например,г. аффинность связывания и эффективность) собирали для A 1 AR человека и A 2A AR. Общедоступные данные о биоактивности были взяты из ChEMBL версии 23 [16] и дополнены внутренними данными (см. Дополнительный файл 1). Поскольку соединения с рибозным каркасом часто связаны с агонистической активностью, а дицианопиридины — с частичным агонизмом, эти соединения были удалены для создания набора данных, ориентированного на антагонисты [17, 18]. Значения биологической активности были стандартизированы до pActivity (концептуально аналогичны значению pChEMBL, ансамблю из pK i / IC 50 / EC 50 / K d значений [19]) при одновременном объединении данных из разных лабораторий и анализы.Эти данные впоследствии были использованы для составления следующих наборов данных о соединениях: «Набор данных о биоактивности A 1 AR» (содержащий биоактивность соединений на A 1 AR), «Набор данных о биологической активности A 2A AR» (содержащий данные о биологической активности соединений на A 2A AR) и «A 1 AR / A 2A AR dataset» (биоактивность соединений протестирована как на A 1 AR, так и на A 2A AR). Последний набор данных включал информацию о селективности соединений.Соединения были названы «селективными», когда разница в активности между двумя белками была более чем 100-кратной (например, A 2A AR-селективным, когда pActivity A 1 AR = 6,5 и pActivity A 2A AR ≥ 8,5) . Набор данных A 1 AR / A 2A AR состоял из пяти классов: non-binder (pActivity A 1 AR и A 2A AR <6.5), A 1 AR-селективный (pActivity A 1 AR ≥ 6,5 и селективность ≥ 100 раз), A 2A AR-селективный (pActivity A 2A AR ≥ 6.5 и селективность ≥ 100 раз) и двойное связующее (pActivity A 1 AR и A 2A AR ≥ 6.5 и селективность ≤ десятикратная). Кроме того, соединения, биоактивность которых измерялась как для A 1 AR, так и для A 2A AR, но не подходили ни к одному из классов набора данных A 1 AR / A 2A AR, были названы «полу -селективные соединения. Соединения, у которых была измерена биоактивность только для одного рецептора и которые не присутствовали в наборе данных A 1 AR / A 2A AR, но были включены либо в набор данных биоактивности A 1 AR, либо в набор данных биоактивности A 2A AR, были называемые «одиночными точками».Распределение активности в разных наборах данных было сопоставимым (таблица 1) и нормально распределено как в A 1 AR, так и в A 2A AR (рис. 1). Следует отметить, что общее количество A 1 AR-селективных соединений примерно в три раза меньше, чем количество A 2A AR-селективных соединений (50 и 146 соединений соответственно). Распределение активности различных классов соединений для A 1 AR ( a ) и A 2A AR ( b ). Соединения из наборов данных биоактивности A 1 AR и A 2A AR, которые не принадлежали ни к одному из классов набора данных A 1 AR / A 2A AR, называются «одиночными точками» Несколько моделей QSAR были созданы для изучения селективности.Во-первых, селективность подтипа для A 1 AR и A 2A AR была смоделирована с использованием моделей классификации. Кроме того, моделировались неселективные соединения (двойные связующие) и несвязывающие. Были построены следующие четыре модели: модель 2-го класса (A 1 AR-селективная / A 2A AR-селективная), две модели 3-го класса (A 1 AR-селективная / A 2A AR- селективные / двойные ингибиторы с одной стороны, и A 1 AR-селективные / A 2A AR-селективные / несвязывающие агенты, с другой стороны, и модель 4-го класса (A 1 AR-селективные / A 2A AR-селективные / двойные / несвязывающие).Все модели были проверены с помощью пятикратной перекрестной проверки с использованием одних и тех же (химически сгруппированных) наборов тестов на итерацию для каждой модели (подробности см. В разделе «Методы»). Модель QSAR с двумя классами была наилучшей для прогнозирования селективности A 1 AR и A 2A AR (рабочая характеристика приемника (ROC) 0,87 ± 0,06 и коэффициент корреляции Мэтьюза (MCC) 0,40 ± 0,13). Добавление двойных и не связанных классов снизило производительность модели. ROC снизился до 0,76 ± 0,06 и 0.64 ± 0,09 для АО A 1 и до 0,62 ± 0,09 и 0,65 ± 0,09 для AR A 2A . Аналогично, MCC уменьшается до 0,22 ± 0,15 и 0,00 ± 0,07 для AR A 1 и до 0,20 ± 0,17 и 0,36 ± 0,12 для AR A 2A (таблица 2). Это указывает на то, что 100-кратно селективные соединения A 1 AR и A 2A AR достаточно химически отличаются друг от друга, чтобы их правильно предсказывала модель, и что они демонстрируют четкую взаимосвязь структура – активность.И наоборот, предполагается, что двойные и несвязывающие вещества имеют химическое сходство с селективными классами A 1 AR и A 2A AR, что усложняет модели дифференциацию между этими классами (рис. 2). Кроме того, чувствительность и прогностическая ценность положительного результата (PPV) были постоянно выше для соединений, селективных к A 2A , чем для соединений, селективных к A 1 , тогда как специфичность и прогностическая ценность отрицательного результата (NPV) были выше для A 1 AR-селективные соединения. Химическое сходство соединений классов селективности A 1 AR-селективный, A 2A AR-селективный, двойные и несвязывающие. Химическое сходство визуализируется с помощью t-SNE [20] на основе отпечатков пальцев FCFP4. a Используемые химические кластеры соединений: A 1 AR-селективный, A 2A AR-селективный, двойные связующие и несвязывающие. b Кластеры на основе химического сходства; каждая комбинация цвета и символа представляет собой уникальный кластер (всего 136 кластеров) Класс несвязующих веществ содержит соединения, неактивные по отношению к обоим рецепторам.Однако эти соединения не очень хорошо дифференцируются от активных классов (A 1 AR-, A 2A AR-селективный и двойной), что наблюдается по низким MCC (0,15 ± 0,06) и плохим ROC (0,57 ± 0,07). ) для не связующего класса. В следующем разделе описывается моделирование биоактивности A 1 AR и A 2A AR в попытке классифицировать несвязывающие вещества. Биоактивность соединений для A 1 AR и A 2A AR была смоделирована с помощью моделей классификации и регрессии.В классификационных моделях соединения с использованием заранее определенного порога (здесь pActivity ≥ 6.5) классифицируются как «активные», а соединения ниже этого порога называются «неактивными». Модель обучается этим классам активности, а также предоставляет класс активности для тестируемых соединений. Напротив, регрессионные модели обучаются не по классам, а по числовым значениям биологической активности. Результатом регрессионной модели является значение биоактивности, которое впоследствии может быть отнесено к классу активности.Как можно увидеть в Таблице 1, где показана средняя pActivity для наборов, это значение (pActivity 6.5) применимо для этих наборов данных и ранее также было показано, что это соответствующий порог, ведущий к сбалансированным классам [15]. Классификация биологической активности и регрессионные модели QSAR были обучены на наборе данных A 1 AR / A 2A AR, том же наборе данных, который использовался в моделях QSAR классификации селективности, описанных в предыдущем разделе. Кроме того, были добавлены полуселективные соединения для увеличения объема обучающих данных.Эти полуселективные соединения обладают экспериментальной активностью в отношении обоих рецепторов, но не соответствуют ни одному из четырех классов селективности (например, соединение с pActivity A 1 AR = 7,0, pActivity A 2A AR = 8,1). Однако для моделирования биоактивности класс селективности не имеет значения, и поэтому эти соединения теперь были включены для повышения производительности модели. Кроме того, отдельные модели QSAR были обучены на наборах данных биоактивности A 1 AR и A 2A AR.Наборы проверочных тестов были составлены на основе химических кластеров и биоактивности соединений; каждое подмножество содержало как активные, так и неактивные. Эти наборы для валидации не были равны вышеупомянутым наборам валидации (селективности), поскольку они использовались для другой цели: валидация моделей биоактивности вместо моделей селективности. Все модели биоактивности были проверены с использованием одних и тех же наборов тестов для перекрестной проверки, независимо от набора данных, который использовался при обучении (A 1 AR / A 2A AR набор данных, A 1 AR биоактивность или A 2A AR. биоактивность).Наборы данных о биоактивности A 1 AR и A 2A AR содержали больше точек данных, чем набор данных A 1 AR / A 2A AR, поскольку эти наборы также включали отдельные точки (биоактивность измерялась только для одного из двух рецепторов) . Единичные точки биоактивности были включены в обучение, но исключены из проверки, чтобы сохранить сопоставимость характеристик для различных моделей. Отдельные точки, которые принадлежали к тому же химическому кластеру, что и точки данных в тестовом наборе, также были исключены из обучения, чтобы предотвратить смещение.Модели регрессии показывают хорошее качество модели при обучении с высокими значениями R 2 (≥ 0,98) и низкими значениями RMSE (≤ 0,14). К сожалению, при применении к набору проверки производительность ниже, чем ожидалось, исходя из производительности обучения (вероятно, из-за сложного набора тестов, основанного на химической кластеризации). Тем не менее, судя о производительности модели по метрикам проверки классификации, можно сделать реалистичную оценку прогнозной производительности моделей. Средние характеристики всех моделей биоактивности (на основе пятикратной перекрестной проверки) были лучше для A 2A AR, чем для A 1 AR, при средней разнице ROC 0.12 (таблица 3). Кроме того, модели классификации работают хуже, чем модели регрессии, на что указывают их более низкие значения для обогащения (ROC) и MCC: средняя разница в ROC для A 1 AR = 0,20 и A 2A AR = 0,07, а также средняя разница в MCC для A 1 AR = 0,07 и A 2A AR = 0,03. Более того, MCC и ROC для моделей классификации биоактивности A 1 AR даже показали характеристики хуже, чем случайные (MCC <0 и ROC <0,5).Наиболее эффективные модели биоактивности были основаны на регрессии, которая достигла средней эффективности (оценка ROC 0,60–0,70) для прогнозирования биоактивности. Применение вышеуказанного Подход к модели биоактивности был протестирован при моделировании селективности соединений (т.е. моделирование аффинности соответствующих рецепторов и косвенное извлечение из этого селективности). Поскольку прогнозируемая биоактивность двух моделей биоактивности не коррелирует друг с другом, была проведена отдельная проверка, чтобы указать эффективность прогнозов селективности. Однако наборы перекрестной проверки моделей в таблице 3 были сгруппированы на основе биоактивности, а не классов селективности, поэтому модели биоактивности были переобучены с использованием разных составленных наборов перекрестной проверки, чтобы оправдать сравнение с обсуждавшейся позже моделью окна селективности.Были выбраны регрессионные модели биоактивности, поскольку они превосходили классификационные модели биоактивности. Более того, при моделировании селективности предпочтение было отдано регрессии, поскольку с помощью регрессии можно получить количественное значение селективности. Таким образом, модели регрессии биоактивности использовали для прогнозирования активности соединения для A 1 AR и A 2A AR. Модели были обучены на наборе данных A 1 AR / A 2A AR, включая дополнительные «полуселективные» соединения. Разница в прогнозируемой биоактивности для двух рецепторов рассчитывалась как значение селективности.Впоследствии классы селективности (A 1 AR-селективный, A 2A AR-селективный, двойной) были присвоены соединениям на основе предсказанной селективности в соответствии с теми же правилами категоризации, которые использовались при классификации ранее. Применение двух объединенных моделей биоактивности для определения селективности (двухступенчатая модель разности A 1 AR-A 2A AR) привело к моделям со средней производительностью. Хотя оценка ROC была приличной (0,75 ± 0,09 и 0,72 ± 0,15), MCC была низкой (0.19 ± 0,16 и 0,28 * ± 0,12, * одна неудачная проверка) как для A 1 AR, так и для A 2A AR, что указывает на то, что модели способны ранжировать соединения, но менее способны объяснить весь набор данных (Таблица 4 ). Продолжая наш подход единой модели для прогнозирования селективности, мы исследовали использование регрессии для модель окна избирательности, а не классификация.В отличие от двухэтапной разностной модели A 1 AR-A 2A AR (регрессия), единственная модель для прогнозирования селективности между A 1 AR и A 2A AR была основана на различии в аффинности. а не прогноз биологической активности и расчет результирующей селективности. Регрессионная модель была обучена непосредственно разнице в биоактивности для обоих рецепторов (pActivity A 1 AR — pActivity A 2A AR = окно селективности) и предсказывает количественную оценку степени селективности соединения (разница в pActivity ).Положительная оценка указывает на A 1 AR-селективность, отрицательная оценка A 2A AR-селективность, а оценка, близкая к нулю, указывает на двойное связывание. Модель оценивалась на основе показателя ROC и показателей классификации (MCC, чувствительность, специфичность, PPV и NPV). Правила классификации A 1 AR-, A 2A AR-селективных и двойных связующих были выведены из пороговых значений, применяемых в моделях классификации селективности: A 1 AR ≥ 100-кратная селективность равна разнице pActivity ≥ 2, A 2A AR ≥ 100-кратная селективность равна разнице pActivity ≤ — 2, а для двойных связующих (≤ 10-кратная селективность) разность pActivity ≥ -1 и ≤ 1. Модель регрессии окна селективности была обучена на тех же данных (набор данных A 1 AR / A 2A AR и полуселективные соединения), что и двухэтапная A 1 AR-A 2A AR разница описанная выше модель, в которой селективность вычиталась из двух отдельных моделей биоактивности. Окно селективности превзошло двухступенчатую разностную модель A 1 AR-A 2A AR с увеличенными значениями ROC для классов селективности A 1 AR- и A 2A AR-селективный (увеличение ROC 0.07–0.13) (таблица 4). На рисунке 3 показаны примеры соединений, которые были неправильно классифицированы с помощью двухэтапной разностной модели A 1 AR-A 2A AR, но были правильно предсказаны с использованием модели окна селективности. Сходство (Tanimoto FCFP4) между неверно предсказанными соединениями с помощью двухэтапной модели разности A 1 AR-A 2A AR составляло 0,25, тогда как сходство в пределах ошибочно предсказанных соединений с помощью модели окна селективности составляло 0,58. Это указывает на то, что двухэтапная модель различия A 1 AR-A 2A AR подвергается сомнению из-за предсказания селективности более разнообразных соединений, а модель окна селективности неэффективна на определенных химических каркасах.Наиболее часто неверно предсказываемый скаффолд моделью окна селективности был N — (2- (фуран-2-ил) -6- (1 H -пиразол-1-ил) пиримидин-4-ил) -2- феноксиацетамид (см. дополнительный файл 2). Химические структуры соединений с предсказаниями с помощью различных моделей селективности. Соединения были ошибочно предсказаны с помощью двухэтапной модели A 1 AR-A 2A AR и правильно предсказаны с помощью модели окна селективности. Прогнозы обозначены как: предсказанный A 1 AR-селективный (A 1 AR), A 2A AR-селективный (A 2A AR) и как двойное связующее (Dual) для лигандов CHEMBL260788 ( a ), CHEMBL3596506 ( b ) и CHEMBL201750 ( c ) Следует отметить, что для прогнозов A 1 AR и A 2A AR не всегда можно было рассчитать MCC и PPV при перекрестной проверке, что приводит к неудачным сверткам перекрестной проверки или итерациям.Это объясняется отсутствием истинных / ложных положительных результатов в конкретном периоде перекрестной проверки: PPV не может быть рассчитан, если нет положительных результатов, MCC не может быть рассчитан без PPV. Модель окна избирательности работает лучше, чем модель трехклассовой классификации избирательности, подтвержденная ранее (таблица 2). Было проверено, является ли эта повышенная производительность результатом увеличения объема данных, поскольку модель окна селективности включала дополнительные полуселективные соединения и несвязывающие вещества, которых в трехклассной модели классификации селективности обязательно не хватало, поскольку разница в аффинности была небольшой. достаточно, чтобы соответствовать отсечке классификации.Когда эти дополнительные точки данных были исключены из модели окна селективности, ROC упал с 0,78 до 0,67 (среднее по классам), что сопоставимо с ROC 0,65 трехклассовой модели классификации селективности. Это наблюдение четко подтверждает прямую связь между качеством модели и доступностью данных и показывает, что повышенная производительность модели окна избирательности объясняется дополнительными точками данных. Следовательно, при моделировании избирательности выгодно использовать непрерывные модели, поскольку в этом случае может быть включено больше данных.Помимо преимущества повышенной доступности данных, модели непрерывной селективности также предоставляют возможность вычислять коэффициент селективности, а не только класс. Это соотношение селективности указывает на степень селективности и, следовательно, не может только идентифицировать селективные соединения, но также может отличать высокоселективные соединения от слабо селективных соединений. Примечательно, что показатели, основанные на классификации (MCC, чувствительность, специфичность, PPV и NPV) для модели окна селективности (без несвязывающих веществ и полуселективных соединений) (таблица 4) ниже для A 1 AR -селективные соединения и двойные связующие, чем показатели модели 3-х классной классификации (оба обучены на одних и тех же данных) (таблица 2), тогда как оценки ROC сопоставимы или выше.Поэтому предсказания модели окна селективности сравнивались с экспериментально измеренными значениями селективности (рис. 4). Было обнаружено, что соединения, селективные к A 1 AR, имеют постоянно более низкие предсказания окна селективности, чем экспериментальные значения селективности. В результате меньшее количество соединений достигло порога селективной классификации A 1 AR, что резко снизило количество селективных положительных результатов A 1 AR. Из 50 A 1 AR-селективных соединений ни одно не достигло порогового значения.Из всех предсказаний только три соединения достигли порога селективности A 1 AR, которые были двойными связующими вместо соединений, селективных к A 1 AR. Этот недостаток предсказанных положительных результатов объясняет неудачные расчеты перекрестной проверки для класса A 1 AR-селективного. Взаимосвязь между экспериментальной селективностью и предсказанной селективностью. Прогнозируемые значения селективности показаны для модели окна селективности. A 1 AR-селективные пороги классификации показаны оранжевыми линиями (пунктирные = старый порог, сплошной = новый порог) селективность по A 1 AR-селективным соединениям.Соединения считались A 1 AR-селективными, когда окно селективности ≥ 0,5, A 2A AR-селективными, когда окно селективности ≤ — 2 (без изменений), и двойным связующим, когда окно селективности ≥ — 1 и < 0,5. При использовании новых пороговых значений значения метрик для прогнозов селективности AR A 1 улучшились: MCC 0,31 ± 0,10, чувствительность 0,45 ± 0,12, специфичность 0,91 ± 0,02, PPV 0,32 ± 0,06 и NPV 0,95 ± 0,01, что указывает на то, что пересмотренный порог улучшает категоризацию соединений, селективных к A 1 AR.Не все соединения, селективные к A 1 AR, были правильно отнесены к категории, но пороговое значение, оптимизированное post hoc, было сочтено адекватным, так как снижение порога селективности A 1 AR дополнительно повысило бы чувствительность (за счет классификации большего количества соединений как A 1 AR -селективный), но также снизит PPV. Здесь правильность прогнозов была важнее количества прогнозируемых активных соединений; следовательно, PPV был важнее чувствительности. Хотя модель окна селективности различает A 1 AR-, A 2A AR-селективные соединения и двойные связующие, модель не учитывает потенциальную неактивность соединений .Следовательно, не связующие не могут быть отфильтрованы с помощью этой модели. Поэтому был применен консенсусный подход статистического моделирования и стыковки на основе структуры для выявления и исключения несвязующих веществ. Модели регрессии биоактивности, описанные выше для A 1 AR и A 2A AR, были объединены с стыковкой набора данных A 1 AR / A 2A AR и полуселективных соединений в кристаллические структуры обоих белков. Прогнозы биоактивности для A 1 AR и A 2A AR и прогнозы окна селективности были получены для всего набора данных A 1 AR / A 2A AR и полуселективных соединений путем объединения прогнозов, сделанных во время Пятикратная перекрестная проверка ранее обученных регрессионных моделей.Соединения были состыкованы с кристаллическими структурами AR A 1 (PDB: 5UEN) [10] и A 2A AR (PDB: 5OLZ) [21], в результате чего были получены баллы стыковки для обоих рецепторов. Соединениям была присвоена отдельная метка биологической активности для A 1 AR и A 2A AR: соединения в A 1 AR были помечены как «активные», когда прогнозируемая pActivity ≥ 7 и оценка стыковки ≤ — 9. Соединения в A 2A AR были помечены как «активные», когда прогнозируемая pActivity ≥ 7 и оценка стыковки ≤ -10. Соединения с прогнозируемым окном селективности ≥ 0,5 или ≤ — 2, соответствующие A 1 AR- и A 2A AR-селективным, впоследствии были отфильтрованы с использованием фильтра согласованной биоактивности (рис. 5). PPV для соединений, селективных к A 1 AR, резко возрастает с 0,13 до 0,39, когда предсказания окна селективности были отфильтрованы с использованием фильтра согласованной биоактивности для A 1 AR. Фильтрация биоактивности A 2A AR также увеличивала PPV соединений, селективных к A 2A AR; здесь стыковочная и консенсусная фильтрация выполнялись одинаково хорошо (PPV A 2A AR-selected: 0.80). Положительная прогностическая ценность (PPV) соединений, которые, как предполагается, являются A 1 AR- или A 2A AR-селективными. PPV отображает количество экспериментально подтвержденных селективных соединений, деленное на общее количество предсказанных селективных соединений. PPV отображаются при применении различных фильтров: без фильтра биоактивности, статистической биоактивности, биоактивности на основе оценки стыковки и согласованной биоактивности (статистической биоактивности и стыковки на основе структуры) Несвязывающие вещества, которые не были удалены с использованием только статистического фильтра, но были отфильтрованы, когда использовался консенсусный подход, были проверены в кристаллической структуре A 1 AR.Некоторые не связывающие вещества (например, CHEMBL1800792) не адаптировались к благоприятной конформации при стыковке с A 1 AR (рис. 6). Более того, взаимодействие с карманным остатком Asn 6.55 (нумерация Баллестероса-Вайнштейна) часто не наблюдалось. Это важный отсутствующий элемент, поскольку взаимодействие с этим остатком, как было показано, важно для связывания лиганда с AR A 1 и AR A 2A [11, 22]. Однако некоторые не связывающие вещества смогли осуществить это взаимодействие (CHEMBL372580), но, тем не менее, имели оценку стыковки, которая не достигла установленного порога (оценка стыковки ≤ — 9).Положения несвязывающих веществ сравнивали с положениями соединения, селективного к A 1 AR (CHEMBL207824) и соединения, селективного к A 2A AR (CHEMBL371436). Оба селективных соединения адаптируют конформацию, которая способна взаимодействовать с остатком Asn 6.55 . Кроме того, позы также представляют собой те же ароматические взаимодействия (с Phe171 EL2 в A 1 AR и с Phe168 EL2 и His250 6.52 в A 2A AR), что и сокристаллизованные лиганды, и адаптируются. аналогичная ориентация каркаса.Наконец, позы селективных лигандов имеют благоприятные оценки стыковки (-10,30 и -10,91, соответственно), подтверждая, что эти соединения являются связующими для A 1 AR или A 2A AR. Пристыкованные позы соединений к их соответствующим целям. Позы двух несвязывающих веществ в A 1 AR (CHEMBL1800792 в a и CHEMBL372580 в b ), A 1 AR-селективного соединения (CHEMBL204780 в c ) и A 2A AR -селективное соединение (CHEMBL371436 в d ).Пристыкованные позы сравниваются с сокристаллизованными лигандами, показанными оранжевым цветом. Водородные связи между лигандами и Asn 6,55 показаны желтым Модель окна прогнозирующей селективности (обучена на A 1 AR / A 2A AR наборе данных и полуселективные соединения) была поставлена задача предсказать селективность соединений из набора внешней валидации. Этот набор содержал 1482 соединения, для которых величина биоактивности доза-ответ (K i / IC 50 / EC 50 / K d ) была известна по крайней мере для одного из двух рецепторов.Если для обоих рецепторов было доступно точное значение биоактивности, соединение классифицировалось в соответствии с предыдущими правилами, применявшимися в этом исследовании. Однако, если было известно точное значение биоактивности только для одного рецептора, использовалось менее точное измерение биоактивности (ингибирование как процентное смещение / эффективность / изменение) для определения неактивности отсутствующего рецептора. Низкая точность значений биоактивности делает эти данные менее подходящими для обучения модели количественной разнице между активностью на двух рецепторах, но подходящими для проверки классификации.Значение pChEMBL <4,5 или порог ингибирования ≤ 50% (при 10 мкМ) использовали для мечения неактивных соединений, тогда как значение pChEMBL ≥ 6,5 использовали для обозначения активных соединений. Модель окна селективности (см. Дополнительный файл 3) была применена к соединениям во внешнем наборе для проверки, предоставив им всем прогнозируемую оценку селективности и, впоследствии, класс селективности. Проверка охватывала все классы селективности: A 1 AR- и A 2A AR-селективные соединения, двойные связующие и несвязывающие.Поскольку модель окна селективности не имеет порога для несвязующих, несвязующие всегда считались либо истинными, либо ложноотрицательными (никогда ложными / истинно положительными). Без фильтрации неактивных веществ модель окна селективности показала среднее значение при прогнозировании A 1 AR-селективных соединений (ROC 0,75) и A 2A AR-селективных соединений (ROC 0,66) во внешнем наборе для проверки (таблица 5). Однако применение фильтра согласованной биологической активности привело к увеличению классификационного обогащения A 1 AR- и A 2A AR-селективных соединений.Хотя ROC для A 1 AR-селективных соединений снизился после применения фильтра биоактивности, PPV и, следовательно, доля истинных A 1 AR-селективных соединений по сравнению со всеми предсказанными соединениями A 1 AR увеличилась с 0,12 до 0,21. Кроме того, MCC немного увеличился с 0,13 до 0,18. Проверка соединений показала, что все несвязывающие вещества были удалены после фильтрации прогнозов окна селективности с помощью фильтра согласованной биоактивности. Таким образом, снижение ROC для AR-селективного класса A 1 было вызвано только наличием двойных связующих.Примечательно, что чувствительность для селективных соединений A 1 AR и A 2A AR составляла 1,00 (100%), тогда как чувствительность для двойных связующих составляла 0,00 (0%). Хотя двойные соединения присутствовали в наборе, который был отфильтрован с помощью модели окна селективности, эти соединения были ошибочно отнесены к категории A 1 AR- или A 2A AR-селективных. Прогнозируемые двойные связующие до фильтрации по биоактивности фактически не были связующими. Однако эти несвязывающие вещества были правильно отфильтрованы с использованием фильтра биоактивности, в результате чего класс двойных связующих веществ остался без соединений с положительным прогнозом.Обратите внимание, что результаты не указывают на двойное обогащение связующим, поскольку прогнозы биоактивности охватывают только соединения, которые, как предполагается, будут селективными по A 1 AR или A 2A AR. Селективность связывания коллоидных частиц является важной темой исследований в области направленной доставки лекарств .Обширная теоретическая работа показала, что высокая селективность может быть получена при использовании многовалентных слабых взаимодействий. Здесь мы предоставляем исчерпывающие экспериментальные доказательства с использованием частиц, покрытых ДНК. Сродство лиганд-рецептор варьируется путем изменения количества комплементарных оснований, показывая, что меньшее количество комплементарных оснований дает более высокую селективность связывания. Экспериментальные данные и соответствующее численное моделирование демонстрируют масштабное поведение между молекулярной плотностью, молекулярным сродством и результирующей плотностной селективностью межчастичного связывания.Эти результаты важны для разработки новых систем адресной доставки лекарств в виде наночастиц. Адресная доставка лекарств в значительной степени зависит от селективности связывания коллоидных частиц, транспортирующих груз. Обширная теоретическая работа показала, что для достижения высокой селективности для пороговой плотности рецепторов необходимы два фактора: многовалентность и слабые взаимодействия. Здесь мы изучаем модельную систему покрытых ДНК частиц с многовалентными и слабыми взаимодействиями, которые имитируют взаимодействия лиганд-рецептор между частицами и клетками.Используя эксперимент с оптомагнитным кластером, измеряют скорость агрегации частиц как функцию плотности лиганда и рецептора. Измеренные скорости агрегации показывают, что связывание становится более селективным для более коротких пар ДНК-лиганд-рецептор, доказывая, что поливалентные слабые взаимодействия приводят к повышенной селективности межчастичного связывания. Моделирование подтверждает экспериментальные данные и показывает роль диссоциации лиганд-рецептор в селективности слабого поливалентного связывания. Многовалентные взаимодействия повсеместно присутствуют в биологии, поскольку они обеспечивают сильные, обратимые и высокоселективные взаимодействия (1⇓ – 3).Множественные индивидуально слабые нековалентные взаимодействия кооперативно образуют высокоспецифичную поливалентную связь между биологическими объектами, которые могут иметь как молекулярные, так и клеточные размеры (4, 5). Эти принципы также используются в наномедицине для разработки наночастиц, нацеленных на определенные типы клеток, с целью улучшения диагностики и лечения (6–8). Нацеливание основано на идентификации сигнатур мембранных рецепторов, уникальных для клетки-мишени. Однако при многих заболеваниях не рецепторы, а уровни экспрессии рецепторов отличаются для больных клеток по сравнению со здоровыми клетками (9, 10).Следовательно, частицы для доставки лекарств должны проявлять специфичность рецепторной плотности, то есть частицы должны связываться как можно более избирательно с клетками с рецепторной плотностью выше определенной пороговой плотности (рис. 1 A ). Взаимодействие частица-частица имитирует взаимодействия ячейка-частица. ( A ) Многовалентное связывание покрытой лигандом частицы с рецепторами на клеточной мембране. ( B ) Частицы лиганда покрыты короткими конструкциями ДНК с одноцепочечным выступом, называемым лигандной ДНК.Рецепторные частицы покрыты короткими конструкциями ДНК с дополнительным одноцепочечным выступом, называемым рецепторной ДНК. Комплементарность выступа определяет силу взаимодействия лиганд-рецептор. Двухцепочечные нити ДНК-наполнители без одноцепочечных выступов вставляются для поддержания постоянной плотности поверхностного заряда. ( C ) Скорость агрегации измеряют как функцию плотности рецептора для постоянной плотности лиганда. Более слабые взаимодействия с меньшим количеством комплементарных нуклеотидов в одноцепочечном выступе рецепторной цепи вызывают более высокую селективность межчастичного связывания.( D ) Параметр селективности α рассчитывается из зависимости скорости агрегации от плотности рецептора. Слабые поливалентные взаимодействия обеспечивают повышенную селективность по сравнению с сильными поливалентными взаимодействиями. Группа Френкеля в Кембридже провела обширную теоретическую работу по теме специфичности рецепторной плотности. Они разработали статистическую механическую модель, которая описывает взаимодействие между покрытыми лигандом гостевыми частицами и покрытым рецептором хозяйским субстратом (11).Модель дает долю связанных гостевых частиц θ в состоянии равновесия как функцию плотности рецептора. Селективность связывания гость-хозяин количественно оценивается параметром селективности α, который меньше 1, если связывание имеет сублинейную зависимость от плотности рецептора, и больше 1, если зависимость быстрее линейной (суперселективность). Работы Френкеля и других групп предсказывают, что специфичность высокой плотности рецепторов может быть достигнута с помощью частиц, которые поливалентно взаимодействуют с клеточной мембраной (11–15).Интересно, что селективность увеличивается, когда константа равновесной ассоциации Ka отдельной пары лиганд-рецептор уменьшается (11, 16, 17). Другими словами, многие слабые связи более избирательны, чем одинарная сильная связь. Несколько экспериментальных исследований подтвердили часть теоретической работы Френкеля. Альбертацци и др. (18) показали суперселективность при самоорганизации супрамолекулярного полимера путем введения многовалентного связующего. Дубачева и др. (19) продемонстрировали суперселективное связывание для модельной системы связывания полимеров поливалентной гиалуроновой кислоты с функционализированной поверхностью.В этих экспериментах была доказана равновесная концепция суперселективности для связывания многовалентных полимеров. Однако в этих работах не изучались какие-либо кинетические аспекты, то есть как поливалентные системы изменяют свою кинетическую вероятность связывания в зависимости от числа рецепторов, а также не изучались свойства селективности биофункциональных коллоидных частиц. Здесь мы изучаем, как скорость межчастичной кинетики зависит от количества взаимодействующих рецепторов, имитируя взаимодействия частица-клетка (рис.1 В ). Коллоидные частицы суспендированы в растворе, где половина частиц покрыта лигандами с поверхностной плотностью σL, называемыми частицами лиганда (L), а другая половина частиц покрыта рецепторами с поверхностной плотностью σR, называемыми рецепторами. частицы (R). Мы используем модельную систему ДНК, в которой ДНК лиганда и ДНК рецептора состоят из одноцепочечных выступов, экспонированных из двухцепочечных фрагментов ДНК длиной 20 пар оснований. Когда две частицы (R = 250 нм) находятся в непосредственной близости, могут образовываться множественные связи лиганд-рецептор, в зависимости от плотности связующего.Изменяя длину одноцепочечных выступов, то есть изменяя количество комплементарных оснований, можно регулировать силу индивидуальных связей лиганд-рецептор. Чтобы количественно оценить связывание между лигандом и рецепторными частицами, мы измеряем скорость агрегации между частицами kagg, используя ранее разработанный эксперимент с оптомагнитным кластером (OMC) (20). Параметр kagg представляет скорость, с которой происходит биохимическая агрегация между частицами, для частиц, которые находятся в четко определенном состоянии близости.Вкратце, дисперсия суперпарамагнитных частиц подвергается воздействию внешнего магнитного поля. Магнитная сила притяжения между частицами приводит частицы в четко определенное проксимальное состояние, в котором могут образовываться межчастичные связи лиганд-рецептор. После фиксированного времени взаимодействия внешнее поле отключается. В этот момент несвязанные димеры частиц распадаются, и димеры, связанные по крайней мере одной связью лиганд-рецептор, остаются. Количество биохимически связанных димеров регистрируется как функция времени, что позволяет количественно оценить среднюю скорость агрегации между частицами kagg.Полное описание эксперимента OMC дано в SI Приложение , раздел S1. В этой статье мы экспериментально изучаем, как кинетика связывания частицы с частицами зависит от плотности лигандов, плотности рецепторов и силы их взаимодействия. Параметр селективности количественно оценивается для длины комплементарной ДНК в диапазоне от 15 пар оснований до всего лишь 5 пар оснований, то есть от сильных до очень слабых взаимодействий. Кроме того, представлена имитационная модель, которая поясняет, что повышенная селективность может быть получена только путем увеличения скорости диссоциации пар лиганд-рецептор.Статья завершается обсуждением того, как полученные результаты могут быть интерпретированы для приложений в адресной доставке лекарств. Суперпарамагнитные гранулы Ademtech Masterbeads, покрытые стрептавидином, были приобретены у Ademtech (диаметр 528 нм, коэффициент вариации 25%). Биотинилированные цепи ДНК были приобретены у IDT (полный список использованных последовательностей ДНК см. SI, приложение , раздел S2). Таблетки с фосфатно-солевым буфером (PBS), бычьим сывороточным альбумином (BSA, чистота> 98%), пробирки с биотином-Atto655 и Protein LoBind Eppendorf были получены от Sigma-Aldrich.Кюветы из боросиликатного стекла 3.3 с внутренними размерами 1,00 ± 0,05 мм × 1,00 ± 0,05 мм, внешними размерами 1,23 ± 0,05 мм × 1,23 ± 0,05 мм и длиной 20 ± 1 мм были получены от Hilgenberg GmbH. Частицы функционализировали аналогично тому, как это было представлено в предыдущей работе (21). Вкратце, покрытые стрептавидином Masterbeads Ademtech были функционализированы биотинилированным лигандом или цепями ДНК рецептора и цепями ДНК наполнителя путем последовательных стадий инкубации.Сначала 15 мкл исходного раствора частиц (10 мг / мл) смешивали с 285 мкл раствора лигандной ДНК или рецепторной ДНК в PBS и инкубировали в течение 60 мин в шейкере инкубатора (1200 об / мин, комнатная температура). Затем добавляли 2 мкл большого избытка ДНК-наполнителя для насыщения оставшихся стрептавидиновых групп ДНК и инкубировали в течение 60 мин в шейкере инкубатора (1200 об / мин, комнатная температура). Количество функционального лиганда ДНК или цепей ДНК рецептора на частице варьировалось на протяжении экспериментов.После второй стадии инкубации раствор частиц был промыт магнитом для удаления несвязанных цепей ДНК. Частицы повторно диспергировали в растворе BSA с концентрацией 10 мг / мл в PBS для подавления неспецифической агрегации частиц. Затем раствор частиц инкубировали в звуковой бане в течение 10 мин и раствор обрабатывали ультразвуком (10 × 0,5 с) для уменьшения количества фоновых кластеров в растворе. Для количественного определения количества нитей ДНК на покрытых стрептавидином шариках Ademtech Masterbeads был проведен непрямой флуоресцентный анализ супернатанта, аналогичный тому, что использовался в нашей предыдущей работе (21).Во-первых, биотиновая способность частиц была определена количественно путем связывания возрастающих количеств биотина-atto655 (b-atto655) на частицах в течение 60 мин в шейкере инкубатора (1200 об / мин, комнатная температура). Самая низкая концентрация b-atto655, при которой еще остается b-atto655 в супернатанте после инкубации, была определена количественно с использованием Thermo Fischer Fluoroskan Ascent FL (λex = 646 нм, λem = 679 нм, спектральная ширина 5 нм) ( SI Приложение , рис. S3 A ). Емкость b-atto655 на частицу была равна Н b-atto 655 = (7.3 ± 0,6) ⋅ 10 4 . Затем к частицам добавляли возрастающие количества биотинилированной ДНК лиганда или рецептора и инкубировали в течение 60 мин в шейкере инкубатора (1200 об / мин, комнатная температура). После этого этапа инкубации раствор частиц был промыт магнитом для удаления несвязанных цепей ДНК. Затем добавляли b-atto655 в концентрации, которая немного превышала емкость частиц b-atto655, после чего следовала стадия инкубации в течение 60 минут в шейкере инкубатора (1200 об / мин, комнатная температура).Частицы, которые не были полностью покрыты цепями ДНК, связывают часть b-atto655 в растворе, в то время как полностью покрытые частицы больше не связывают какой-либо b-atto655. После инкубации растворы частиц промывали магнитом и измеряли флуоресценцию супернатанта. Количество b-atto655 в супернатанте для определенной концентрации ДНК, Is.n. ([ДНК]), связано с количеством ДНК на частице, NDNA, согласно уравнению. 1 : NDNA = I0-Is.n. ([ДНК]) I0-Is.n. ([DNA] = 0) ⋅Nb-atto655.[1] Здесь I0 — интенсивность флуоресценции раствора b-atto655, добавляемого на втором этапе инкубации, а Is.n. ([DNA] = 0) — интенсивность флуоресценции супернатанта при добавлении нефункционализированного стрептавидина Ademtech Masterbeads. Для получения дополнительных сведений об анализе супернатанта см. SI Приложение , раздел S3. Мы исследуем, как связывание между коллоидными частицами зависит от плотности лиганда и рецептора, используя модельную систему ДНК на рис.1 В . Покрытые стрептавидином гранулы Ademtech Masterbeads были функционализированы с помощью трех различных биотинилированных конструкций ДНК: ДНК лиганда, ДНК рецептора и ДНК наполнителя. Нити лигандной ДНК состоят из двухцепочечного спейсера ДНК длиной 20 п.н. с одноцепочечным выступом длиной 15 нуклеотидов. Нити рецепторной ДНК также состоят из двухцепочечного спейсера длиной 20 п.н., но они имеют одноцепочечный выступ, который варьируется от 5 до 15 нуклеотидов. Наполнитель ДНК состоит только из двухцепочечного спейсера длиной 20 пар оснований и используется для насыщения частицы ДНК, так что поверхностный заряд частицы остается постоянным на протяжении всего эксперимента. Покрытые стрептавидином частицы последовательно инкубировали в течение 1 ч, сначала с определенной концентрацией лиганда или рецепторной ДНК для получения определенного покрытия лиганда или рецептора, а затем с избытком ДНК-наполнителя для насыщения поверхности частицы (полное описание процесса функционализации приведено в Материалы и методы ). Чтобы количественно определить количество функциональных цепей ДНК, которые связываются с частицами во время 1-часовой инкубации, был проведен анализ супернатанта (анализ супернатанта подробно описан в SI Приложение , раздел S3). SI Приложение , рис. S3 B показывает зависимость поверхностной плотности связанной ДНК как функцию от инкубированной поверхностной плотности ДНК, рассчитанную путем деления концентрации ДНК на концентрацию частиц. Измеренная кривая следует линейной зависимости до тех пор, пока не будет достигнуто плато. Поверхностная плотность ДНК этого плато представляет собой емкость ДНК частицы: σDNA, max = (2,2 ± 0,5) 104 мкм − 2. Полученное соотношение между покрытием инкубированной ДНК и покрытием связанной ДНК используется в оставшейся части этой статьи, так что плотность лиганда σL или плотность рецептора σR всегда представляют собой связанные поверхностные плотности. Для количественной оценки связывания между частицами лиганда и частицами рецептора их смешивали в соотношении 1: 1 и измеряли скорость агрегации с использованием эксперимента OMC (20). Фиг. 2 A показывает измеренную скорость агрегации как функцию плотности рецепторов для системы из 15 дополнительных пар оснований. Точка данных при нулевой плотности рецептора количественно определяет неспецифическое взаимодействие между частицами, покрытыми ДНК: kagg, ns = (5 ± 2) 10-3 с-1. При низких плотностях рецепторов (σR≈101 мкм − 2) в агрегации преобладают неспецифические взаимодействия между частицами.Для плотностей лигандов σL> 103 мкм − 2 и плотностей рецепторов σR ≈102 мкм − 2 измеренная скорость агрегации значительно превышает скорость неспецифической агрегации и увеличивается почти линейно с увеличением плотности рецептора. Скорость агрегации как функция плотности рецептора ( σ R ), плотности лиганда ( σ L ) и сродства лиганд-рецептор (п.о.): ( A ) 15 пар оснований, ( B ) 12 оснований, ( C ) 9 оснований, ( D ) 8 оснований, ( E ) 7 оснований и ( F ) 5 оснований комплементарен лиганду и рецептору. Для плотностей рецепторов σR> 103 мкм − 2 плато достигается при скорости агрегации немного выше kagg = 0,05 с − 1. Это самая высокая скорость агрегации, наблюдаемая для данной системы в эксперименте OMC. Половина магнитных димеров, которые образуются во время срабатывания, состоят из лигандной частицы и рецепторной частицы, что делает возможной специфическую гибридизацию ДНК. Другая половина магнитных димеров состоит либо из двух лигандных частиц, либо из двух рецепторных частиц, не допускающих специфического связывания. SI Приложение , рис. S4 показывает, что скорость неспецифической агрегации низкая и не зависит от плотности рецепторов или от длины одноцепочечного выступа. Рис. 2 B и C показывают аналогичные кривые для системы ДНК с комплементарностью 12 и 9 пар оснований соответственно. Измеренные кривые для всех плотностей лигандов напоминают кривые системы 15 п.н. Это можно понять из сходных скоростей ассоциации и медленной диссоциации относительно экспериментального времени.В эксперименте OMC димеры частиц формируются магнитным путем, и частицы удерживаются в непосредственной близости в течение приложенного времени взаимодействия. Из литературы известно, что скорость ассоциации kon коротких цепей ДНК в растворе очень слабо зависит от числа комплементарных оснований (22⇓ – 24). Таким образом, в эксперименте ожидается, что примерно такое же количество связей лиганд-рецептор будет образовано для систем из 15, 12 и 9 п.н. Затем поле выключают на время ожидания twait = 40 с, позволяя частицам повторно диспергироваться в растворе.После фазы ожидания измеряется количество связанных димеров частиц и рассчитывается скорость агрегации. Диссоциация димера во время фазы ожидания приводит к более низкой скорости агрегации. Однако во всех случаях (комплементарность 15, 12 или 9 пар оснований) время диссоциации одной молекулы (25–27) значительно превышает время ожидания 40 с, поэтому диссоциация димера в OMC не ожидается. эксперимент. Таким образом, мы объясняем сходство кривых на рис. 2 B и C для 15-, 12- и 9-нуклеотидной ДНК с их сходными скоростями ассоциации и медленной диссоциацией по отношению к экспериментальному времени. Результаты сильно различаются для систем с 8, 7 и 5 п.н. Фиг.2 D — F показывают скорость агрегации как функцию поверхностной плотности рецептора. Кривые резко зависят от плотности рецептора, что означает более высокий параметр селективности, чем для взаимодействий с высоким сродством (15/12/9 п.н.). Начало плотности рецепторов, то есть порог, при котором происходит агрегация, увеличивается с уменьшением плотности лиганда и с уменьшением количества комплементарных оснований.Это качественно согласуется с теоретическими предсказаниями Ванга и Дормидонтовой (17). Обратите внимание, что агрегация частиц наблюдается даже для взаимодействия 5 пар оснований, для которого время диссоциации одной молекулы (27) составляет около 1 с, что намного короче, чем время ожидания 40 с в эксперименте OMC. Тем не менее, агрегация частиц четко наблюдается при высоких плотностях рецепторов, что, следовательно, должно быть связано с наличием многовалентных связей между частицами в димере. Для количественной оценки и сравнения селективности связывания лиганд-рецептор для различной длины ДНК кривые скорости агрегации для плотности лиганда σL = (2.2 ± 0,5) 104 мкм − 2 для всех длин ДНК подобраны и нанесены на один график (рис. 3 A ). Измеренные кривые аппроксимируются с помощью сигмовидной кривой. Подробную информацию о фитинге можно найти в приложении SI , раздел S5. Повышенная селективность для слабых поливалентных взаимодействий. ( A ) Измеренные скорости агрегации как функция плотности рецептора для постоянной плотности лиганда σ L = (2,2 ± 0,5) 10 4 мкм -2 для всех сил взаимодействия.Слева от пунктирной линии в зоне взаимодействия в среднем присутствует менее одного рецептора; справа от пунктирной линии в среднем присутствует более одного рецептора. Данные с 15, 12 и 9 п.н. подбираются с помощью экспоненциальной функции, а данные с 8, 7 и 5 п.о. подбираются с помощью сигмоидальной функции. Подробную информацию о фитинге можно найти в приложении SI , раздел S4. ( B ) Расчетный параметр селективности с использованием уравнения. 2 , с параметрами подгонки, полученными из А.Слабые взаимодействия лиганд-рецептор (5/7/8 п.н.) обеспечивают повышенную селективность по сравнению с сильными взаимодействиями лиганд-рецептор (9/12/15 п.н.). На рис. 3 A серая пунктирная линия указывает плотность рецептора, выше которой возможно образование множественных связей. Оценка основана на радиусе частицы, длине связи лиганд-рецептор и расстоянии между частицами (для полного расчета см. SI Приложение , раздел S6). При плотностях рецепторов слева от пунктирной линии в зоне взаимодействия присутствует в среднем не более одного рецептора, который представляет собой область на рецепторной частице, которая находится на расстоянии от частицы лиганда не более длины связи (Lbond ~ 20 нм).Для длин ДНК, для которых одинарная связь стабильна во временных масштабах времени ожидания — 15, 12 и 9 п.н. — существует значительная специфическая агрегация при плотностях рецепторов в моновалентном режиме. Для более слабого взаимодействия — 8, 7 и 5 пар оснований — необходимы более высокие плотности рецепторов, чтобы обеспечить агрегацию частиц за счет поливалентного связывания. Параметр селективности α определяется Martinez-Veracoechea и Frenkel (11) как относительное изменение количества связанных частиц Nbound, образующихся в равновесии, как функция числа рецепторов на клетке nR: α = dln NbounddlnnR.В эксперименте OMC мы измерили скорость связывания между частицами kagg, которая представляет количество связанных димеров, деленное на среднее время взаимодействия, как функцию плотности рецепторов σR, которая представляет собой количество рецепторов, деленное на площадь частицы. Поскольку α описывает относительное изменение числа связанных частиц с числом рецепторов, мы можем умножить Nbound и nR на любую ненулевую константу, не изменяя α. Таким образом, мы можем заменить Nbound и nR на kagg 〈tint〉 и σRAparticle соответственно, чтобы получить следующее выражение: α = dlnNbounddlnnR = dln (kagg⋅ 〈tint〉) dln ( σR⋅Aчастица).[2] С помощью этого уравнения и подобранных кривых на рис. 3 A рассчитывается параметр селективности как функция плотности рецептора (рис. 3 B ). Заштрихованные полосы вокруг сплошных кривых получены из ошибок аппроксимации и представляют интервал неопределенности параметра селективности. Измеренные кривые агрегации имеют S-образную форму. При низких скоростях агрегации (в диапазоне kagg <0,01 с-1) в процессе агрегации преобладают неспецифические взаимодействия между частицами; здесь скорость не зависит от плотности рецепторов, и экспериментально определенная селективность равна нулю.Для увеличения плотности рецепторов параметр селективности α достигает 1 на изгибе кривой и возвращается к нулю, когда скорость агрегации достигает насыщения. Измеренная селективность для взаимодействий 15, 12 и 9 пар оснований находится в диапазоне от нуля до единицы, что означает, что измеренная скорость агрегации увеличивается не более чем линейно с плотностью рецептора. Напротив, селективности для взаимодействий 8, 7 и 5 пар оснований достигают почти значения 2, что означает, что измеренная скорость агрегации возрастает вплоть до квадратичной зависимости с плотностью рецептора.Это доказывает, что многовалентные слабые взаимодействия приводят к повышенной селективности межчастичного связывания. Для дальнейшего исследования экспериментально измеренной избирательности связывания было разработано кинетическое моделирование методом Монте-Карло, которое имитирует связывание частиц, индуцированное лигандом и рецептором. Цель моделирования — понять происхождение повышенной селективности для слабых поливалентных взаимодействий и оценить актуальность результатов для условий, выходящих за рамки эксперимента. В моделировании частицы имеют либо плотность лиганда σL, либо плотность рецептора σR. Во время такта срабатывания магнитного поля димеры частиц образуются с постоянной скоростью kdimmag. Здесь не учитываются истощение мономера и образование более крупных кластеров (см. Приложение SI, раздел S5 ссылки 20). Внутри магнитных димеров нетривиально моделировать расстояние от поверхности до поверхности частиц, потому что расстояние между частицами является результатом суммы нескольких межчастичных сил с разными знаками и диапазонами длин, например, сила Гамакера, электростатические силы, магнитная сила, энтропийные силы, такие как гидрофобный эффект и стерическое отталкивание, и сила истощения; кроме того, может иметь значение шероховатость поверхности (28).Чтобы сделать возможным расчет, исходя из основных уравнений силы, необходимо знать основные параметры частицы с высокой точностью, что не относится к частицам, используемым в этом исследовании. Поэтому необходим подход с ограниченным числом параметров, чтобы можно было провести сравнение с экспериментальными результатами. Из экспериментов мы знаем, что плотность ДНК на каждой частице одинакова, потому что ДНК лиганда и рецептора были дополнены ДНК наполнителя. Это означает, что заряд и стерически индуцированное межчастичное отталкивание могут быть аппроксимированы постоянными в экспериментах.Кроме того, магнитная сила может считаться постоянной, поскольку магнитное поле срабатывания было постоянным. Образование одномолекулярных связей может добавлять как притягивающие, так и отталкивающие вклады в межчастичный потенциал, например, из-за конечной длины связи (20 нм) и механической жесткости гибридизированной конструкции. Однако каждая гибридизированная конструкция имеет четыре точки шарнира, а именно два шарнира в точках прикрепления к частицам и два шарнира в начальных основаниях одноцепочечных выступов, что делает связи между частицами довольно гибкими.Кроме того, количество связей намного меньше, чем общее количество молекул ДНК в области межчастичного взаимодействия (по оценкам, около 1000, включая молекулы-наполнители ДНК), что ограничивает влияние конструкций на общий межчастичный потенциал. Таким образом, расстояние между частицами априори неизвестно, но мы предполагаем, что частицы имеют минимум своего потенциала взаимодействия на эффективном расстоянии между частицами Δ x , которое остается постоянным в течение времени срабатывания.Расстояние между частицами — это параметр, который определяет геометрическое перекрытие между молекулами лиганда и рецептора, поэтому интересно смоделировать, как скорость агрегации частиц будет масштабироваться с различными значениями расстояния между частицами. Мы моделируем частицы как сферы с радиусом R , из которых выступают молекулы лигандной или рецепторной ДНК. Вероятность того, что димер состоит из рецепторной частицы и лигандной частицы, составляет 50%. Каждый димер имеет индивидуальный оттенок времени взаимодействия, равный времени между моментом образования димера и окончанием времени срабатывания.В течение этого времени взаимодействия частицы в димере могут образовывать одну или несколько связей лиганд-рецептор, в зависимости от числа геометрически перекрывающихся молекул лиганда и рецептора и их эффективных скоростей ассоциации и диссоциации. Собственная скорость связывания kLR [мкм2 с-1] определяется как скорость, с которой лиганд-рецепторные связи образуются между частицей с единичной плотностью лиганда σL и частицей с единичной плотностью рецептора σR. Поскольку поверхности частиц изогнуты и взаимодействие лиганд-рецептор зависит от расстояния, kLR представляет собой среднюю скорость связывания по всем возможным расстояниям лиганд-рецептор.Максимальное расстояние, на котором может быть образована связь, — это длина связи лиганд-рецептор. Плотность связи лиганд-рецептор σLR может быть рассчитана численно в соответствии с формулой. 3 : dσLRdt = kLRσLσR-koffσLR. [3] Здесь koff [с-1] — скорость, с которой диссоциируют связи лиганд-рецептор. Чтобы рассчитать общее количество межчастичных связей, которые образуются во время взаимодействия, сначала численно рассчитывается плотность связи, а затем общее количество связей определяется с помощью уравнения. 4 : NLR = σLRAint. [4] Здесь Aint — это площадь взаимодействия на частицах, которая определяется как площадь на частицах, где расстояние между двумя частицами меньше, чем длина гибридизированной конструкции ДНК Lbond≈20 нм, так что могут образовываться связи ДНК – ДНК. Размер области взаимодействия зависит от расстояния между частицами Δx, потому что большее расстояние между частицами уменьшает область, в которой молекулы лиганда и рецептора перекрываются; более подробную информацию можно найти в приложении SI , рис.S6. Если за время взаимодействия образовалась хотя бы одна связь лиганд-рецептор, димер называется агрегированным. Впоследствии, в течение времени ожидания twait, уже сформированные связи лиганд-рецептор могут диссоциировать, но одновременно могут образовываться новые связи, поскольку частицы все еще удерживаются в непосредственной близости уже сформированными связями. Однако, когда все связи лиганд-рецептор в какой-то момент времени диссоциируют, димер больше не агрегируется и не может образовываться снова, поскольку оба критерия удержания частиц (за счет магнитного притяжения или за счет молекулярных связей) исчезают.По истечении времени ожидания остаются только димеры с сильными межчастичными связями, то есть димеры с эффективными временами диссоциации, превышающими время ожидания. Таким образом, параметр селективности является репрезентативным для этих прочных связей. Конечное количество связей лиганд-рецептор внутри димера рассчитывается по формулам. 3 и 4 . В конечном итоге определенная часть димеров агрегируется одной или несколькими связями лиганд-рецептор. Также принимаются во внимание неспецифические взаимодействия, включая скорость неспецифического взаимодействия kns [с-1].Из общей доли агрегированных димеров, скорость агрегации kagg рассчитывается в соответствии с SI Приложение , уравнение. С1 . Полный просмотр параметров моделирования приведен в приложении SI , раздел S7. Смоделированные кривые воспроизводят S-образную форму, как видно из измерений; скорость агрегации увеличивается, а кривые смещаются в сторону более низкой концентрации рецептора в случае уменьшения расстояния между частицами, увеличения собственной скорости связывания, увеличения плотности лиганда и уменьшения скорости диссоциации. Рис. 4 связывает результаты моделирования с экспериментально измеренными скоростями агрегации. Экспериментальные результаты для взаимодействий с высоким сродством (15/12/9 п.н.) очень похожи, и поэтому эти данные усреднены и представлены оранжевыми точками данных на рис. 4 A . Как упоминалось выше, диссоциацией молекул можно пренебречь из-за такого высокого сродства, поэтому скорость диссоциации может быть установлена равной нулю, а неизвестными параметрами моделирования являются Δx и kLR. Остальные параметры моделирования остаются постоянными ( SI Приложение , Таблица S8).Рис. 4 B, и SI Приложение , рис. S8 показывают, что существует несколько комбинаций Δx и kLR, для которых смоделированная скорость агрегации совпадает с измеренной скоростью агрегации. Небольшие расстояния между частицами (Δx≤8 нм) и высокие скорости связывания лиганд-рецептор (kLR≥10-5 мкм2⋅с-1) необходимы для согласования моделирования с экспериментом. Уменьшение расстояния между частицами ниже 8 нм (для высокого kLR) или увеличение скорости связывания лиганд-рецептор выше 10-5 мкм2 · с-1 (для низкого Δx) не изменяет моделируемую кривую.В этих диапазонах параметров одного лиганда и одного рецептора в области взаимодействия достаточно для образования связи за заданное время срабатывания, поэтому скорость связывания определяется исключительно плотностью молекул. По той же причине форма оранжевой кривой на рис. 4 A определяется исключительно вероятностью присутствия хотя бы одной рецепторной молекулы в области межчастичного взаимодействия. Результаты моделирования в сравнении с экспериментальными данными. ( A ) Точки экспериментальных данных, сопровождаемые смоделированными кривыми скорости агрегации для расстояния между частицами Δ x = 4 нм и скорости связывания лиганд-рецептор k LR = 10 −4 мкм 2 с −1 .Экспериментальные данные для 15-, 12- и 9-нуклеотидной ДНК усредняются, поскольку эти данные очень похожи. Смоделированные кривые скорости агрегации для других комбинаций Δ x и k LR показаны в приложении SI , рис. S8. ( B ) Тепловая карта, показывающая возможные комбинации Δ x и k LR , для которых моделирование воспроизводит экспериментальные данные. Малое межчастичное расстояние и высокая скорость связывания рецептора лиганда приводят к совпадению между моделированием и экспериментом.Красная точка показывает конкретную комбинацию Δ x и k LR для имитационных кривых в A. Далее экспериментальные данные для взаимодействия ДНК с 8, 7 и 5 п.н. воспроизведен с помощью моделирования. Для каждой подходящей комбинации Δx и kLR моделирование выполняется с изменяющейся ненулевой скоростью диссоциации. Рис. 4 A показывает смоделированные кривые скорости агрегации для Δx = 4 нм и kLR = 10-4 мкм2 · с-1 (красная точка на рис. 4 B ), сопровождаемые экспериментальными данными.Моделирование может воспроизводить измеренные данные для каждой длины ДНК. Это указывает на то, что повышенная селективность связывания при слабом сродстве может быть получена путем изменения только скорости диссоциации взаимодействия лиганд-рецептор. Таблица 1 дает обзор смоделированных скоростей диссоциации, которые приводят к совпадению между моделированием и экспериментом, для всех подходящих комбинаций Δx и kLR. Баланс между Δx и kLR определяет процесс ассоциации частиц. Увеличение расстояния между частицами Δx приводит к меньшему количеству взаимодействующих лигандов и рецепторов, но может быть компенсировано более высокой скоростью связывания лиганд-рецептор kLR.Диссоциация димеров частиц представляет собой баланс между, с одной стороны, количеством лигандов и рецепторов и их скоростью связывания (определяемой Δx и kLR), а с другой стороны, скоростью диссоциации отдельных связей лиганд-рецептор koff. Для определенного Δx возможны несколько значений kLR, которые варьируются в пределах порядков. Оказывается, что для 10-кратной скорости связывания соответствующая скорость диссоциации, которая следует из моделирования, также в 10 раз выше. Обзор результатов моделирования Эффективная константа сродства равновесия K∼a (единицы квадратных микрометров) может быть рассчитана по формуле. 5 для каждого моделирования в Таблице 1: K∼a = kLRkoff. [5] Рассчитанная эффективная константа сродства K∼a описывает баланс между ассоциацией и диссоциацией межчастичных связей. Данные таблицы 1 показывают, что K∼a практически не зависит от подгоночных параметров kLR и Δx. Основная причина заключается в том, что константа сродства связана с положением наклона S-образной кривой, то есть значением оси x точки перегиба кривой. Масштабирование констант сродства является ожидаемым: для более коротких длин ДНК полученная константа сродства уменьшается, что соответствует сдвигу кривой агрегации в сторону более высоких плотностей рецепторов. Полученные эффективные константы сродства K∼a определены в терминах поверхностей, но могут быть преобразованы в константы объемного сродства Ka и затем сравнены с константами объемного сродства, рассчитанными для последовательностей ДНК в растворе (подробности см. В приложении SI , раздел S9) . SI Приложение , таблица S9 показывает, что Ka, полученная из K∼a, слабее, чем константа объемного сродства, рассчитанная на основе последовательности ДНК. Например, для ДНК длиной 7 п.н. экспериментально полученное значение Ka = 4⋅101 M − 1 значительно ниже, чем рассчитанная константа сродства Ka = 8⋅107 M − 1.Можно выделить несколько факторов, которые могут быть ответственны за это более низкое сродство. Наиболее вероятная причина заключается в том, что пониженная молекулярная подвижность и доступность на поверхности дает гораздо меньшую разницу свободной энергии для гибридизации на поверхности, чем для гибридизации в растворе, как теоретически рассматривали Varilly et al. (29). Кроме того, гибридизации на поверхности может препятствовать отрицательный электростатический заряд, вызванный высокой плотностью молекул ДНК на частицах. Параметр селективности можно извлечь из результатов моделирования (таблица 1). Значения селективности увеличиваются с уменьшением длины ДНК. Это согласуется с экспериментальным наблюдением, согласно которому селективность увеличивается с уменьшением сродства лиганд-рецептор. Параметры селективности напоминают измеренные значения, показанные на фиг. 3 B для всех длин ДНК, за исключением ДНК длиной 5 пар оснований. Однако расхождение для ДНК длиной 5 п.н., вероятно, связано с ограниченными точками данных на переднем крае экспериментальной кривой. В этой статье мы измерили скорость агрегации покрытых ДНК частиц, чтобы имитировать связывание клеток с частицами. В эксперименте OMC частицы квазинепрерывно удерживаются в непосредственной близости (〈tint〉 = 10 с). Время взаимодействия достаточно велико для образования нескольких слабых связей лиганд-рецептор, которые вместе образуют стабильную связь между частицами. Однако, когда идентичные частицы свободно диспергировались в растворе, время сближения было бы намного короче. Если время близости короче, чем типичное время для образования единственной связи лиганд-рецептор, агрегация частиц очень маловероятна. Мы вычисляем время взаимодействия для частиц, находящихся в свободном состоянии в растворе, используя коэффициент диффузии частиц. Для частиц с радиусом R = 0,25 мкм в водном растворе с вязкостью η = 1 мПа⋅с типичное время, в течение которого частица диффундирует на расстояние, равное длине связи лиганд-рецептор Lbond = 0,02 мкм, рассчитывается по формуле. 6 : τprox = πηRLbond2kBT. [6] Это дает время приближения τprox = 0,1 мс. В эксперименте OMC такое короткое время срабатывания невозможно применить, но мы можем рассчитать количество связей лиганд-рецептор, которые будут образованы в течение этого времени близости, используя уравнения. 3 и 4 . Для самых высоких плотностей лигандов и рецепторов, которые были экспериментально оценены в этом исследовании, σL = σR = 2,2⋅104 мкм − 2, а типичная скорость ассоциации лиганд-рецептор в растворе составляет 105 М − 1 с − 1, что соответствует kLR = 10. −2 мкм2⋅с − 1, в сочетании со скоростью диссоциации koff = 103 с − 1, мы получаем в среднем пять связей на димер. Это означает, что во время столкновения отдельных частиц с частицами в принципе между частицами будет образовываться стабильная многовалентная связь, даже для связующих с низким сродством.Эти расчеты показывают, что сверхселективное связывание частиц также можно измерить для частиц, свободных в растворе, без применения каких-либо магнитных сил. Однако анализы без магнитной силы потребуют гораздо большего времени инкубации, чем анализы с магнитной силой (20). В этой статье изучалась селективность поливалентного связывания между покрытыми ДНК коллоидными частицами, количественно определяемая скоростью агрегации, наблюдаемой в эксперименте OMC. Скорость агрегации измеряли для широкого диапазона плотностей рецепторов и лигандов, показывая, как увеличение плотности приводит к увеличению скорости агрегации.Параметр селективности определяется как относительное увеличение количества димеров частиц в зависимости от плотности рецепторов на частицах, где процесс селективного связывания характеризуется параметром селективности, превышающим 1. Для рецепторов ДНК с различным количеством комплементарных оснований, от 15 до 5 пн были получены следующие параметры селективности: α15bp = 0,9 ± 01, α12bp = 0,7 ± 0,1, α9bp = 1,0 ± 0,2, α8bp = 1,7 ± 0,3, α7bp = 1,9 ± 0,5, α5bp = 1,8 ± 0,5. Эти результаты доказывают, что более слабое сродство лиганд-рецептор приводит к более высоким параметрам селективности.Этот экспериментальный набор данных демонстрирует масштабное поведение между молекулярным сродством и плотностной селективностью специфического связывания между коллоидными частицами. Кинетическое моделирование методом Монте-Карло было разработано на основе геометрического перекрытия между молекулами лиганда и рецептора на частицах, с расстоянием между частицами и внутренними скоростями связывания и развязывания в качестве параметров. Смоделированные кривые воспроизводят формы экспериментально измеренных кривых, и эффективные константы сродства к равновесному связыванию могут быть извлечены для различных длин гибридизации.Моделирование показывает, что повышенная селективность для более слабого сродства лиганд-рецептор может быть объяснена только увеличением скорости диссоциации лиганда и рецептора. Результаты моделирования показывают важность времени близости частиц для возможности образования многовалентных связей. Результаты описанных экспериментов и модели доказывают наличие сверхселективности в скорости кинетического связывания многовалентных взаимодействий частица-частица. Эта работа выходит за рамки исследований равновесия, поскольку позволяет получить представление о кинетических свойствах, которые имеют отношение к разработке систем доставки лекарств с точки зрения размера частиц, плотности лиганда и сродства лиганд-рецептор. Все данные исследования включены в документ и приложение SI . Авторы благодарят программу Peri-Operative IMPULS за финансовую поддержку. Вклад авторов: M.R.W.S., L.J.v.IJ. и M.W.J.P. спланированное исследование; M.R.W.S. проведенное исследование; M.R.W.S. проанализированные данные; и M.R.W.S., L.J.v.IJ. и M.W.J.P. написал газету. Авторы заявляют об отсутствии конкурирующей заинтересованности. Эта статья представляет собой прямое представление PNAS. См. В Интернете сопутствующее содержание, например комментарии. Эта статья содержит вспомогательную информацию в Интернете по адресу https://www.
Количественное прогнозирование селективности между аденозиновыми рецепторами A 1 и A 2A | Journal of Cheminformatics
Наборы данных
Моделирование A
1 AR / A 2A Селективность подтипа AR с использованием классификационных моделей QSAR Моделирование Биоактивность A
1 AR и A 2A AR с использованием моделей классификации и регрессии QSAR Моделирование A
1 AR / A 2A Подтип селективности AR с использованием регрессионных моделей Удаление несвязывающих веществ для повышения производительности
Проверка модели окна селективности на внешнем наборе
Многовалентные слабые взаимодействия повышают селективность связывания между частицами
Значимость
Abstract
Материалы и методы
Материалы.
Функционализация частиц.
Анализ супернатанта для количественной оценки покрытия стыковочной цепи ДНК.
Скорость агрегации как функция рецептора и плотности лиганда
Повышенная селективность для слабых поливалентных взаимодействий
Моделирование скорости агрегации
Заключение
Доступность данных.
Благодарности
Сноски