
Обонятельный вектор: Обонятельный вектор | Психологические тренинги и курсы он-лайн. Системно-векторная психология
Обонятельный вектор | Психологические тренинги и курсы он-лайн. Системно-векторная психология
В носу есть рецепторы, которые отвечают за сознательное дифференцирование запахов: приятные — неприятные. А так называемый «нулевой нерв» различает не запахи, а феромоны. Феромоны — основа всего, происходящего между людьми. Через них происходят два важнейших процесса…
90 441349 20 Июня 2010 в 20:56
Автор публикации:
Мария Грибова, специалист в области международного права и международных отношений
Диана Кирсс, семейный врач
Характерные речевые обороты:
- В тихом омуте черти водятся…
- Бабушка надвое сказала
- Предупрежден—значит вооружен!
-
Не суй свой нос в чужой вопрос
Общая характеристика
| Численность | Менее 1 % |
| Архетип | Выжить во что бы то ни стало |
| Видовая роль | Стратегический разведчик, советник вождя, шаман |
| Цвет наибольшего комфорта | Фиолетовый (но носить предпочитает неприметный серый) |
| Геометрия наибольшего комфорта | Зигзаг |
| Место в квартели | Внутренняя часть квартели энергии, интроверт |
| Тип мышления | Интуитивное, невербальное, стратегическое |
Особенности психики
Раньше считалось, что человеческое обоняние — атавизм животной природы. Но именно благодаря ему происходит восприятие важнейшей информации: помимо сознательных запахов, он распознает неосознаваемые, так называемые феромоны.
Но именно благодаря ему происходит восприятие важнейшей информации: помимо сознательных запахов, он распознает неосознаваемые, так называемые феромоны.
От любого сенсора часть информации поступает в сознание, а часть, минуя сознание, попадает напрямую в бессознательное. Так, например, кожей я чувствую, как меня коснулся другой человек, но постоянного касания рубашки я не замечаю, не осознаю.
То же и с обонянием. В носу есть рецепторы, которые отвечают за сознательное дифференцирование запахов на приятные и неприятные. А так называемый «нулевой нерв» различает не запахи, а феромоны. Феромоны — основа всего, происходящего между людьми. Через них происходят два важнейших процесса: ранжирование (то есть выстраивание по рангам) у мужчин и возникновение влечения между мужчинами и женщинами. «Нулевой нерв» — это и есть эрогенная зона обонятельника.
Феромоны — это бессознательные запахи, которые говорят о человеке больше, чем он мог бы сказать о себе сам. Они — отражение наших эмоций, состояний, бессознательных желаний.
Так, ощущая запах, обонятельник «чует» и ваше состояние, и ваши мысли. Для обонятельника эти мысли «пахнут» хуже, чем самая вонючая помойка. Для него весь мир — источник всевозможных запахов, и среди них не существует приятных. Запах каждого состояния, каждой мысли выдает все несовершенство, все изъяны человеческой природы. Потому на его лице — маска постоянного отвращения. Рождаясь, обонятельник сразу попадает в эту постоянную «вонь». Он презирает людей. Его ощущение: «вы все ниже меня». Он не стремится к контакту с людьми.
Архетип обоняния — выжить во что бы то ни стало. Ключ к выживанию — обладание информацией. Обонятельник через восприятие феромонов получает уникальную, никому больше не доступную информацию. Это и дает ему возможность управлять, разделять, властвовать.
При этом никто ничего не знает о самом обонятельнике.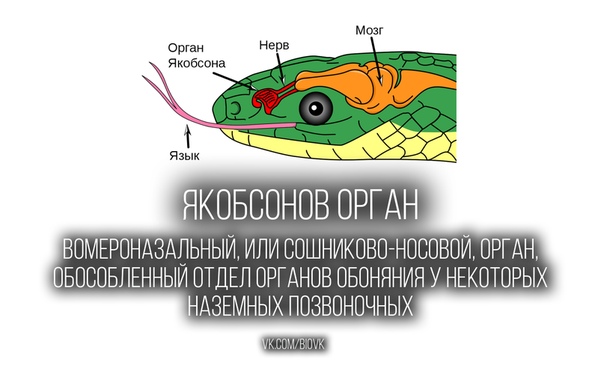 Он всегда в сторонке, но так, чтобы иметь все в поле зрения, и… поближе к выходу на случай, если придется спасаться бегством. Запахи обонятельника скрыты, чтобы его самого никто не мог «прочесть». Отсутствие запаха вызывает реакцию безотчетного страха у людей других векторов. Мы вроде видим человека, но его как будто бы и нет: не ощущая запаха, мы вообще не воспринимаем его как одушевленный объект. Таким образом обонятельник на бессознательном уровне влияет на других людей.
Он всегда в сторонке, но так, чтобы иметь все в поле зрения, и… поближе к выходу на случай, если придется спасаться бегством. Запахи обонятельника скрыты, чтобы его самого никто не мог «прочесть». Отсутствие запаха вызывает реакцию безотчетного страха у людей других векторов. Мы вроде видим человека, но его как будто бы и нет: не ощущая запаха, мы вообще не воспринимаем его как одушевленный объект. Таким образом обонятельник на бессознательном уровне влияет на других людей.
Обонятельник реализуется только через выживание стаи. Он презирает всех людишек, и в первую очередь — их вонючую глупость.
«Моя жизнь ничто, жизнь стаи все» — так мыслит уретральник. Обонятельника же люди не волнуют ни в каком смысле и ни в каком качестве, помимо одного: стая необходима ему для собственного выживания (никто не выживает поодиночке). Так в первобытной стае обонятельник становится советником вождя. Он занимается внутренней контрразведкой и внешней стратегической разведкой.
Внешняя разведка — разведка об опасностях на ландшафте. Когда зрительники посмотрят вокруг и скажут, что опасности нет, и когда звуковики послушают и скажут, что опасности нет, именно обонятельник заметит подвох. Он придет и скажет, что туда идти не надо. Непонятно почему, но это окажется так. Окажется, что там притаилась опасность.
Когда зрительники посмотрят вокруг и скажут, что опасности нет, и когда звуковики послушают и скажут, что опасности нет, именно обонятельник заметит подвох. Он придет и скажет, что туда идти не надо. Непонятно почему, но это окажется так. Окажется, что там притаилась опасность.
Внутренняя разведка — разведка об опасностях внутри стаи. Стая может выжить, только если каждый будет выполнять свою видовую роль. Поэтому обонятельник везде сует свой нос. Он следит за тем, чтобы все работали, все исполняли свою видовую роль, и избавляется от ненужных стае особей. Отсюда первобытный страх перед обонятельниками у зрительников, рационализируемый словами: «Нехороший человек, я это чувствую».
Между уретральным вождем и обонятельным советником существует бессознательный сговор. Уретральник — единственный, кто терпит обонятельника, потому что он предлагает вождю лучший путь для выживания на ландшафте. Обонятельник же становится советником вождя, потому что уретральник сохраняет его от ненависти стаи.
Задача выжить во что бы то ни стало не имеет перерыва ни на одну секунду, потому что если условно представить себе, что в природе существует перерыв для нее хотя бы на одну секунду в год, то именно в эту секунду все заканчивается.
Поэтому и эрогенная зона обонятельника, нос, никогда не отдыхает. Сон у обонятельников всегда поверхностный. Он просыпается в самую критическую минуту, сам не знает почему. Именно обонятельники становятся героями историй по типу «вышел посреди ночи в магазин, а в это время мой дом сгорел» или «решил задержаться дома и опоздал на самолет, который разбился».
У обонятельников уникальный интуитивный невербальный ум. У них нет ума в привычном понимании. Они не занимаются логическими построениями, не анализируют ситуацию, не пытаются ее представить, не пытаются точно разобрать, назвать. Вместо этого у обонятельников есть точнейшие, мощные, яркие ощущения, что и как нужно делать, основанные на воспринимаемом им феромонном фоне окружающих. Ощущения эти не складываются в слова и не проговариваются.
Никаких ключевых слов у обонятельника нет. Нет не только ключевых слов, даже точных названий запахов не существует. Ни в одном языке мира. Проговаривает же мысли обонятельника его младший брат по квартели энергии — оральник.
Поскольку задача выжить во что бы то ни стало важнее всех остальных, она не ограничивается ни нормами морали, ни нормами культуры, ни нормами нравственности и этики.
Обонятельник сохраняет исключительно живое вещество. Не культуру, не нравственность, а саму жизнь. И жизнь не каждого отдельного человека, а жизнь всей целостности людей. И потому на него не накладывается ни одно культурно-нравственное ограничение. Там, где обоняние будет ограничено, все живое умрет.
По этой же причине обонятельники не разделяют правду и ложь. Они единственные никогда не лгут, потому что просто не отличают правду и ложь.
В развитом состоянии обонятельники элементарно адаптированы к понятиям правды и лжи, добра и зла, но внутри себя они этого не ощущают. И исполняют свою видовую роль вне этих категорий. Иными словами, развитые обонятельники работают в областях, где понятия нравственности, правды и лжи не требуются, где важнее другое — обеспечение выживания стаи. Это шпионаж, политика, финансы и наука.
И исполняют свою видовую роль вне этих категорий. Иными словами, развитые обонятельники работают в областях, где понятия нравственности, правды и лжи не требуются, где важнее другое — обеспечение выживания стаи. Это шпионаж, политика, финансы и наука.
Сегодня внешняя разведка как таковая уходит в прошлое: сегодня мир глобален и космополитичен, границы между странами и народами становятся все более и более размыты, а всемирная паутина интернет-пространства и вовсе стирает все мыслимые границы. Но всем известны такие гениальные разведчики прошлого, как, например, Рудольф Абель, без работы которого СССР оказался бы беззащитным перед атомной бомбой.
Внешняя стратегическая разведка развилась в политику. Обонятельники сегодня выстраивают отношения нашей стаи с внешней стаей, основываясь на императиве выжить во что бы то ни стало. Занимая крупные политические посты, становясь министрами финансов, министрами иностранных дел, они договариваются между собой, проводя политику сдерживания. При этом они не основываются ни на каких ценностях нравственности и морали. «У Англии нет постоянных врагов и друзей, есть интересы ее короны».
При этом они не основываются ни на каких ценностях нравственности и морали. «У Англии нет постоянных врагов и друзей, есть интересы ее короны».
Все финансовые системы созданы исключительно обонятельниками и по сей день ими управляются. Среди известных финансистов: Джон Сноу, Генри Полсон, Алексей Кудрин, Александр Шохин.
Обонятельная разведка сегодня нацелена на угрозы со стороны микромира. Обонятельники — те ученые, которые заняты исследованием неизвестного в областях, где существуют риски для выживания людей. Это и называется в народе любопытством. Когда на это накладывается еще и соответствующее умение, то мы имеем дело с крайней любознательностью. Она делает из обонятельников гениальных ученых. Не тех, кто систематизирует готовые знания, а тех, кто делает колоссальные прорывы — такие, например, как изобретение пенициллина.
Неразвитых обонятельников считают большими мерзавцами. Большие интриганы, они иногда сами становятся жертвами своей интриги. Архетипичные обонятельники могут стать самыми страшными серийными убийцами и маньяками, выполняя свою видовую роль напрямую, то есть физически устраняя особей, не способных выполнять свои задачи в стае.
Склонный к меланхолии обонятельный ребенок не стремится к общению. Во дворе, в классе, в школе его все не любят. Он ощущает это как постоянную угрозу и, естественно, пытается от нее уйти, остаться дома под тем или иным предлогом. Если родители поддерживают такое поведение, они растят мерзавца и интригана с плохим будущим. Когда-нибудь он может сам стать жертвой своих интриг.
Нужно понимать, что по-настоящему заботиться о своем обонятельном ребенке означает выталкивать его в коллектив. Все во двор? И его во двор. Все в школу? И его в школу. Все на рыбалку? И его на рыбалку! Это — именно то, что ему нужно. Напрягая все свои способности, чтобы выжить в коллективе, не получить в свой эрогенный нос, обонятельник развивается. Так он учится выполнять свою видовую роль — выживать во что бы то ни стало.
Подробнее об особенностях психики людей с разными векторами, сценариях их взаимоотношений с окружающими можно узнать уже на бесплатных онлайн-лекциях «Системно-векторная психология» Юрия Бурлана. Регистрируйтесь здесь.
Регистрируйтесь здесь.
Корректор: Наталья Коновалова
Автор публикации:
Мария Грибова, специалист в области международного права и международных отношений
Диана Кирсс, семейный врач
Обонятельный вектор. | Блог Владимира Тычинского
Здравствуйте!
В этой статье рассмотрим последний из верхних векторов — обонятельный вектор.
Нередко в составе руководства больших компаний можно встретить человека, который, не обладая громкими титулами, имеет, тем не менее, фактическую власть мало чем уступающую власти “правящей верхушки”.
Эти люди не стремятся быть на виду, не пытаются тешить свои карьерные и финансовые амбиции, но при этом они всегда находятся рядом с “сильными мира сего”, а бывает, что и НАД ними.
“Серые кардиналы” наших дней, теневые стратеги и “тайные советники вождей” — все это они, представители одного из самых малочисленных векторов — обонятельного вектора! Наделенные чуть ли не животным чутьем (или если хотите сверх — интуицией) люди этого вектора способны “видеть” скрытые состояния других людей, их истинные желания, ощущения.
Визуальная диагностика присутствия обонятельного вектора
“Обонятельный” человек внутренне всегда стремится остаться незамеченным, “уйти в тень”, поэтому велика вероятность при первой встрече вообще его не запомнить или не обратить на него внимания. Манера не броско одеваться (преобладание серых и синих тонов), заурядный внешний вид, бедная мимика и жестикуляция “обонятельных” людей полностью соответствует этому стремлению.
Речь “обонятельника” зачастую невнятная, голос тихий, не отражающий эмоционального состояния говорящего. Людям этого вектора свойственна осторожная манера изъясняться недомолвками, как бы оставляя пути для отступления, в том случае, если разговор заходит в неприятное им русло.
Существует один внешний “индикатор”, который может выдать в человеке носителя обонятельного вектора… это слабо уловимое специфическое выражение лица, нечто среднее между высокомерием, презрением и ироничной полу — усмешкой.
“Обонятельники” единственные способны считывать эмоции людей, видеть их животную природу — отсюда выражение надменности, которое, впрочем, полностью отражает его внутреннюю сущность.
Взаимодействие с людьми обонятельного вектора
Для общения с людьми обонятельного вектора нужно знать о них следующие важную вещь: Попытаться обмануть человека этого вектора — дело заведомо проигрышное.
Неискренность они распознают лучше любого полиграфа, но никогда не выказывают того, что видят любого насквозь и даже наоборот, могут подыграть наивному “обманщику”. А такие игры с “обонятельником” ни к чему хорошему привести не могут.
Если среди Ваших бизнес — партнеров или клиентов есть люди обонятельного вектора будьте с ними искренны и честны. Если же это идет в разрез с Вашими интересами — лучше откажитесь от сотрудничества вообще, иначе гарантированно останетесь в проигрыше.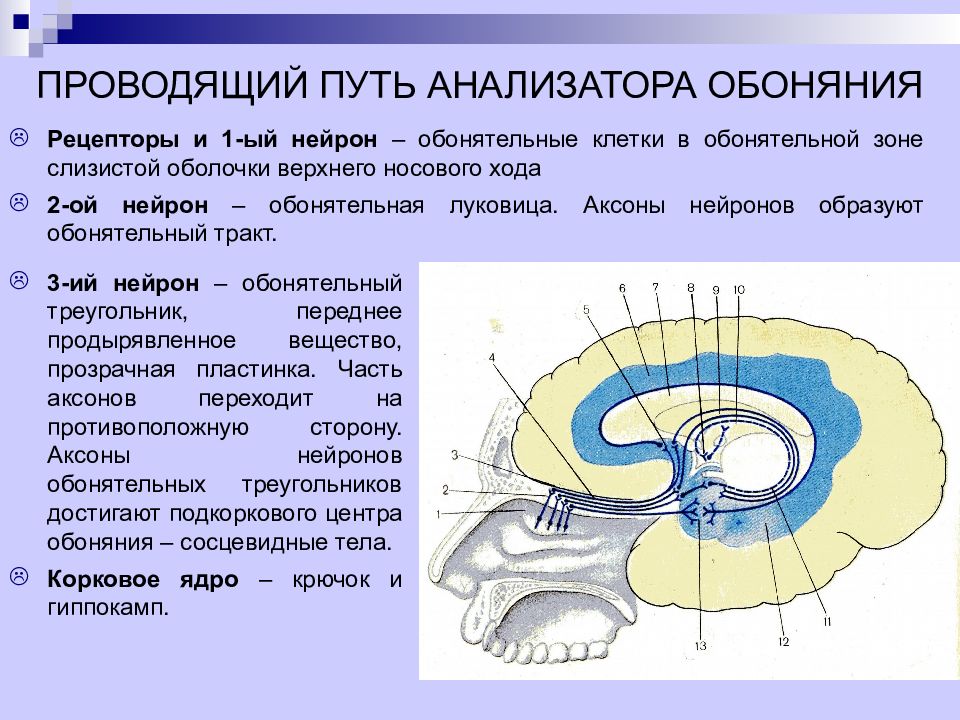
А вот иметь “обонятельника” среди подчиненных — большая удача для любого руководителя. Главное суметь его распознать. Идеальное применение способностей “обонятельника” — роль риск-менеджера, консультанта по кадровым вопросам, советника на бизнес -переговорах.
Мы рассмотрели все верхние вектора, а в следующий раз мы рассмотрим вектор, носители которого как никто другой расположены к ведению бизнеса. Это кожный вектор.
На сегодня всё. Будьте здоровы и благополучны!
Предыдущие записи:
Оральный вектор
Бизнес по законам природы
Системно-векторная психология
10 БЕСПЛАТНЫХ уроков
Фиолетовый вектор (обонятельный) — FAQ по реальности
Стратегический охранник.
Самый древний орган чувств. В стае они могут учуят проблему
задолго до приближения.
Сидит на высоком холме и нюхает воздух.
Чьё либо общество нюхать мешает. Фиолетовые поодаль.
«Парфюмер» Зюскинда – всё кругом воняет.
Приятный запах – свой. Иногда матери и отца.
Цвет, вкус и запах.
Цветов 15-20. Не названием. Вкусов 5-6.
А запахов? Их нет. О мире запахов невозможно договориться, у
каждого индивидуальный.
Ведомые чутьём.
Выбирают запах – и получают несмотря ни на что.
Интуитивные знания и решения. Советник вождя.
Реальная власть над стаей – у него. Сам не добывает себе
еду, питается соками стаи.
Уничтожит второго Фиолетового.
Жизнь среди запахов.
С трудом ездят в транспорте, там пахнут Красно-Чёрные. По
вестибюлю ходят зигзагами.
В метро не садятся – выше нос. Любят стоять у дверей, там
дует.
Привычки.
Роскошь не нужна. Достаточно комфорта.
В одежде фиолетового нет. Чаще серый.
Чихание – это удовольствие. Ковыряние в носу.
Парфюм, благовония и тп.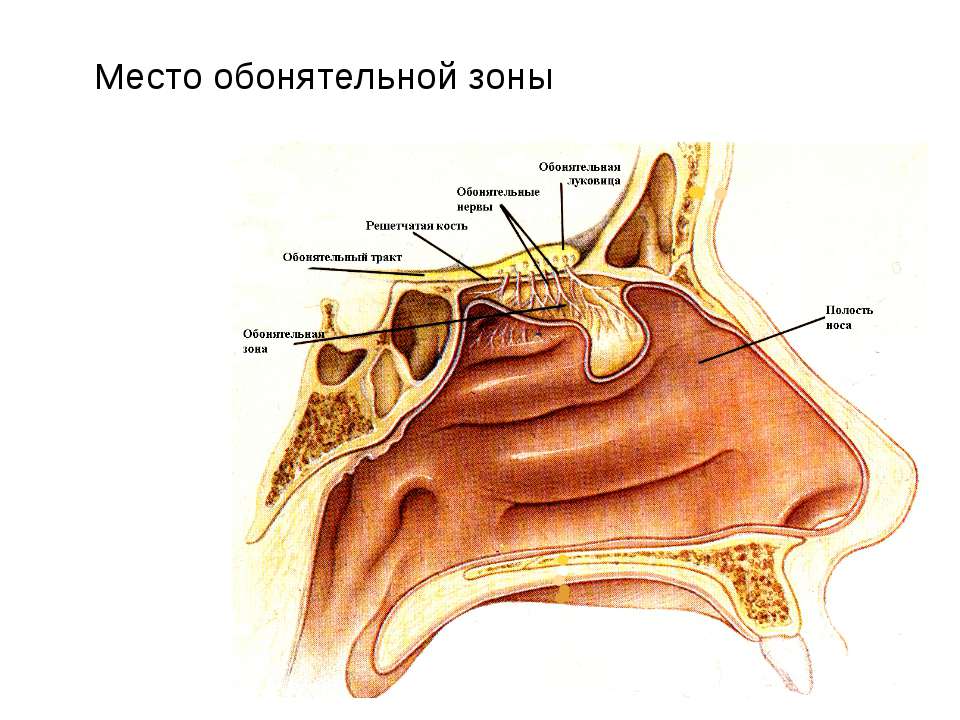 Дарить с осторожностью.
Дарить с осторожностью.
Многие, наоборот, принципиально не пользуются.
Сами пахнут очень слабо.
В гости ходить не любят, и к себе не зовут.
Перед гостями «принюхиваются», если что-то не так, могут
уйти домой.
Профессии и работа.
Парфюмеры, химики, метеорологи, психологи.
Сотрудники спецслужб, таможенники, экологи, поэты-пародисты,
писатели-сатирики, дипломаты, осведомители. Сомелье.
У фиолетовых врачей сильна диагностика.
Советник, помощник, стоящий за Красным лидером, которого он
не любит за запах власти, но который ему необходим. А красный ему доверяет,
потому что фиолетовый предвидит беды, в которые можно попасть без
раздумий.
Общение.
Не нравится «запах» — дел не ведём. Чует ложь.
Люди без запаха – опасны (по мнению Ф.)
Манипуляторы с детства (примитивно – ябеды)
Сами фиолетовые выходят сухими из воды.
Внешность и признаки.
Худощавы. Неприметная одежда.
Высокомерное и презрительное выражение.
Уголки рта чуть опущены, глаза чуть прищурены. Часто
приметный нос.
Иногда натягивают свитер на нос.
Заходя в дверь, оставляют её открытой – пути к отступлению,
создают запасные аэродромы.
Почерк неразборчивый, иногда сами не могут прочесть.
Принцип – не оставлять следов.
Не любят фотографироваться, давать автографы, подписывать
документы.
Юмор: острый, порой ядовитый.
Часто обнюхивает пищу, не пробует –а нюхает.
Редко занимаются спортом. Их спорт – шахматы по переписке.
Любовь и секс.
Потенциал невысокий, зависит от запаха партнёра.
Неприятный запах – нулевой потенциал.
А если ТОТ САМЫЙ ЗАПАХ – высокая половая мотивация.
Влюблённость до беспамятства. Можно издеваться, не уважать – никуда не уйдёт.
«Любимый» запах 1-2 раза в жизни. Часто предпочитают, чтоб
партнёр не пользовался парфюмом.
Характерен гостевой брак. Фиолетовые мужчины склонны
жениться на женщинах с подросшими детьми.
Кто не спрятался, тот не Фиолетовый.
Чувствует состояние окружающих. Умеет быть незаметным.
Точечными воздействиями направляет беседу (вплоть до поворота на 180).
Уходит от опасности заранее. (пример «ПОЖАР»)
Чуют кризисы, страхуются от неожиданностей.
Склонны нейтрально отвечать на тесты, чтоб не вычислить
никакой информации про них.
Нет ни картинки, ни внутреннего голоса – просто приходят
идеи.
Очень мнительные, много информации берут «из воздуха», постоянно
перестраховываются.
Людям не доверяют.
Здоровье.
Слабое. Часто заложен нос. Хронические насморки: аденоиды,
гаймориты итп – нужны, чтоб блокировать неприятные запахи.
Непринятые Ф+О – бронхиальная астма.
Мнительны по отношению к своему здоровью.
Характер в гармонии.
Приятен и инетересен. Один из двух самых интеллектуальных
векторов.
Тонкость мышления и интуиция.
Интересуется психологией и философией, задумываются о
смыслах и истине.
Ценители чая и кофе, разбирают сорта.
Воспоминания-запахи.
Характер в неврозе.
Уединение, немотивированная раздражительность, хроническое
недовольство всем вокруг. Часто человек не в курсе, что это из-за запаха.
Не найдя реализации себя – приспособленческий образ жизни.
«Принципиальная беспринципность»
Чихать я хотел на все
Злопамятность, мстительность.
Никогда не прощает обиду.
Неисправимый пессимизм.
Ненавидит всех вонючек, то есть всех.
И снова о запахе.
Бессознательно могут взять себе чужой запах.
Любят проветривать помещение (от чужих). Другие наоборот,
оставлять закрытым (свой запах)
Персонажи.
«История Адели Г.» — Адель «Горькая Луна» – Мими «Запах
женщины» — капитан Фаусто «Тайна третьей планеты» — Зелёный Кардинал Ришелье,
Парфюмер «Талантливый мистер Рипли» — Том Рипли «Леон» — Стэн Мюллер из «17
мгновений весны»
Связки в одном человеке.
Кр. + Ф – руководит как явно, так и скрытно. Хочет к людям,
но одновременно хочет в тень.
Ф. + Оранж. хорош для бизнеса и исследований: интуиция +
логика.
Различия.
Кор. Честность / Фиол. Изворотливость Кр. Честолюбие / Фиол.
Отчуждение
Ж. Жизнерадостность / Фиол. Замкнутость
З. Восторженность / Фиол. Настороженность
Ор. Логичность / Фиол. Алогичность
Обонятельный — советник вождя.
Функция: стратегия, выживание.
Предупрежден — значит вооружен!
Знание — сила! В тихом омуте черти водятся….
а) цвет – фиолетовый
б) геометрия – зигзаг
в) человек внутренней ЭНЕРГИИ, интроверт
г) тип мышления – интуитивно-стратегическое.
Внешнее:
Самый незаметный и неприметный человек в группе. Ничем не
выделяется, не бросается в глаза, не запоминается. Предпочитает серые цвета в
одежде. Лицо часто недовольное. Взгляд проницательный, колючий. Скошенный
подбородок. Заметный нос.
Внутреннее:
Слабая нервная система. Меланхолики. Вялые. Надменность.
Ехидство. Беспринципность. Коварство. Аморальность. Безнравственность.
Неуловимый интриган.
Обонятельный вектор | Что у нас сегодня? События дня с точки зрения Системно-векторной психологии
Согласно Системно-векторной психологии Юрия Бурлана существует 8 векторов. Вектор – это набор врожденных бессознательных желаний и психических свойств человека, определяющих его характер, поведение и тип мышления. Обонятельный вектор – самый редкий, им обладает менее 1% людей.
Невербальный интеллект
Люди с обонятельным вектором наделены невербальным интеллектом – они безошибочно, на уровне феромонов понимают, что происходит вокруг. Это происходит благодаря хорошо развитому вомероназальному органу – древнему анализатору, который улавливает бессознательные «запахи» влечения и ранжирования.
Обладая таким особенным видом мышления, обонятельник презирает всех вокруг, считает их ниже себя. Ведь поведение других людей, все их мысли можно так легко просчитать и прочитать, как открытую книгу. А самого обонятельника вычислить нельзя, он остается скрытым от других людей. Другие не чувствуют его феромонов, поэтому относятся к нему с опаской и подозрением. Именно обонятельник часто вызывает всеобщую неприязнь и страх.
Сам обонятельник безэмоционален, его естественное состояние – меланхолия. По этой причине другие люди, особенно эмоциональные зрительники, его отталкивают. Обонятельник всегда остается несколько в стороне от других людей, старается не привлекать к себе внимание. Интуитивно чувствует любую опасность, в помещении стремится занять место поближе к выходу. И все потому, что его природная задача – выжить любой ценой. А выживая сам, он обеспечивает сохранность всего коллектива.
Профессиональная реализация
Развитые обонятельники идут в науку, их интересуют прежде всего те направления, которые помогут противостоять угрозам выживания, – вирусология и микробиология, ядерная физика, психиатрия. Именно обонятельный А. Флеминг открыл пенициллин, тем сохранив жизнь многим людям.
Кроме того, обонятельники реализуют себя в политике или становятся шпионами, регулируют денежные потоки на должности крупного финансиста или министра финансов.
Системно-векторная психология . Ребенок-меланхолик: как его воспитывать?
Ребенок с обонятельным вектором – интроверт и меланхолик. От природы он очень спокойный и вялый. Как правило, имеет очень болезненную конституцию тела, но удивительно сильный, особенно, в руках. Такой ребенок не хочет выходить на улицу, общаться с другими детьми, зачастую он плохо ест или вообще кушать не хочет. Он предпочитает сидеть дома, при этом ничем особенным не занимаясь. Вывести его из себя практически невозможно, он не истерик и вообще никак не проявляет эмоциональные состояния, зачастую оставаясь абсолютно безучастным даже в самых сложных ситуациях… Что делать с таким ребенком? Как его воспитывать? Что из него может вырастить? Давайте для начала разберемся в причине меланхолии, и найдем более точные ответы о воспитании таких детей.
Дети бывают разные. Одни — солнечные лучики, которые смеются и плачут от малейшего изменения настроения. Другие — упорные и упертые, хоть делают все медленно, но до конца. Третьи — замкнутые и задумчивые, погруженные в свои мысли, часто задающие вопросы о мироздании своим родителям. Глядя на столь разнообразные психические характеристики разных детей, мы понимаем, что к каждому необходим свой, особенный подход, свое воспитание, свой метод. Только в таком случае ребенок откликнется, станет проявлять себя, развиваться.
Чтобы не навредить собственному ребенку, а помочь ему в становлении личности, очень важно понимать его психотип. Читайте о воспитании разных детей в статье «Как любить своего ребенка? Не Януш Корчак, а лучше…»
Ребенок-меланхолик: какой он?
Обонятельный человек и ребенок внешне всегда выглядит очень меланхолично и безэмоционально. Необходимо точно понимать, что имеется ввиду под этими словами. Ведь есть детки очень сдержанные, которые ограничивают себя, сдерживаются и не тратятся эмоционально наружу. Их тоже в некоторой степени можно назвать меланхоликами, но это будет неверно. Есть детки терпеливые, которые спокойно сидят и никуда не спешат, их надо подталкивать к работе, к переменам. Есть детки, поглощенные собственными мыслями, которые мало разговаривают и как будто постоянно в уме решают сложные задачи. Но все они — не меланхолики.
Обоняние же по-настоящему меланхолично, не сдержанно, не спокойно, не скрытно, а именно — настоящее олицетворение полного отсутствия эмоций. Он не боится, он просто не испытывает страха. Он не скрывает эмоции, их у него просто нет.
Зачастую мы, взрослые, родители и учителя, особенно при наличии зрительного или звукового вектора, такого ребенка характеризуем, как «слишком вялый», «подавленный», даже несколько «странный». Более того, у него «неприятное лицо», нет ни тени улыбки даже в самых смешных ситуациях, он часто морщит нос, трет его, а уголки губ опускаются вниз.
Конституция тела очень слабенькая, они меланхолики не только психологически, но и физически. У них часто скошенный подбородок, некрасивое, непривлекательное лицо. Бывает большой, выдающийся нос, с которым зачастую случаются проблемы: ребенок часто гнусавит, говорит в нос, хотя насморка у него нет. Хотя, конечно, это все неточные внешние характеристики и полагаться лишь на внешнее описание не стоит. Особенно, если учесть, что просто обонятельников не бывает, у этих людей есть хотя бы еще один «нижний вектор». А если к его векторальному набору прибавляется еще и зрение, то внешний облик человека меняется. Он умеет коммуницировать с людьми по-зрительному, и они зачастую воспринимают его именно так.
Еще одна особенность обонятельных детей — у них очень плохой аппетит. Они не могут кушать еду с резкими запахами и спокойно относятся разве что к безвкусным кашам. Вялый и неприязненный, обонятельный ребенок долгое время вообще может принимать пищу только от мамы, избегая кушать в школьной столовой или где-нибудь вовне дома. Даже складывается впечатление, что такой ребенок попросту боится есть. Часто мама приходит в ужас от такого поведения малыша: «Почему мой ребенок ничего не ест? Что делать с этой проблемой?». Его стараются запихнуть едой, но это не получается. У родителей наступает отчаянье, заставлять, насильственно кормить такого ребенка — это основная проблема мамы, особенно, в самом маленьком возрасте.
Обонятельник частенько уходит от прямого ответа и никогда от него не добьешься нормальной, связной речи.
В группе ребенок-флегматик, меланхолик всегда молчит, сидит поодаль или с краешку, недалеко от двери. Он как будто постоянно в тени и любое внимание к нему, отдается сморщенным лицом и неприязнью в глазах. Это некие серые мышки, которые никогда не выдают своего присутствия. Их очень часто не замечают, забывают. Вообще, он презирает окружающих, а окружающие отвечают ему взаимностью. Нет, его никогда не бьют, не обзывают, над ним не издеваются. И не ненавидят кого-то конкретно, слишком большая честь была бы конкретному Васе, если бы его лично ненавидели. Это общее состояние, ничего не имеющее общего со звуковой неприязнью «людишек» или анальной обидой с желанием отомстить.
У обонятельного ребенка нет верного друга, нет и заклятого врага. Он крутиться в толпе одногодок, не избегая их, но и не входя в крепкий контакт ни с кем. Разве что только с уретральным ребенком, если таковой есть в группе. Ведь именно он сможет осуществить защиту обонятельника. Так или иначе, а у ребенка-меланхолика есть только собственные интересы и ничто не помешает ему их реализовать, а люди вокруг — это лишь средство достижения.
В тоже время взгляд у них очень проницательный. У любого, кто посмотрит в его глаза, создается впечатление, что он как под гипнозом, а сам ребенок — вроде бы заглядывает в душу. Колючий, недобрый, пронизывающий взгляд. Мама может отмечать некоторую привязанность ребенка к себе, но в тоже время и прямо написанное на лице презрение, неприязнь.
Распространенное заблуждение
Очень часто родители, глядя на свое меланхоличное чудо, сильно печалятся. Что из него вырастет? Кем он может в жизни стать? Да вообще, что такой ребенок хочет, если даже от еды отказывается? Безэмоциональный, безынициативный, а значит, по аналогии, нам кажется, что и бесперспективный. Но как же мы глубоко заблуждаемся! Все как раз наоборот. Самый большой потенциал, самое большое будущее именно у обонятельных детей. Именно они более устойчивы к проблемам, именно они имеют больше возможностей, именно они управляются не собственными эмоциями и рационализациями, а настоящим чутьем жизни, интуицией.
Обонятельный вектор дарит человеку высшую степень желания сохранения собственного тела. Если другой ребенок лезет на дерево и может сорваться оттуда, поломав руки-ноги, то обонятельник никогда так не поступит. Не потому, что он боится, а потому что ему даже в голову не придет заниматься подобными глупостями. И ему совершенно безразлично общественное мнение. В то время, как все другие дети за компанию начинают потреблять алкоголь и курить, пробовать наркотики, обонятельний ребенок остается в стороне от такого саморазрушения. Он никогда не попадет в плохую историю, его не застанут за плохими делами.
Уже в раннем возрасте обонятельный ребенок научается примыкать к людям, наделенным властью или силой. Это умение в будущем, во взрослой жизни сослужит ему очень хорошую службу. Да, он не будет работать дворником или трактористом, он не будет в среде добрых и хороших, но он может стать крупным финансистом, хорошим политиком, великим ученым, врачом, который откроет лекарство от неизлечимой болезни, тайным разведчиком. Все профессии и реализации, где нужно исключительно владеть собой, иметь хорошие знакомства, уметь выкрутиться из самых сложных ситуаций, не поддаваться на эмоции — ему будут по плечу. Но только в развитом, реализованном состоянии.
При неправильном воспитании, стандартном подходе к обонятельному ребенку, при слишком большом неправильном давлении родителя на такого ребенка, или даже скорее, отсутствии давления, сверхзаботе, происходит его развитие в другую сферу. Когда обонятельный человек не реализован, он занимается самыми отвратительными делами, которые можно только себе представить. Тут он выдает из себя характеристики, которые звуковики и зрительники обозначают как «мерзавец, сволочь, подлец, человек без нравственности и морали».
Родителям такого ребенка-меланхолика надо понять, что обонятельный человек — это всегда значительная личность, и в позитивном и в негативном плане. Ему задано было стать великолепным финансистом, но может он стать и большим вором, обкрадающим банки, а то и государства. Ему задано было стать великим политиком, решающим судьбы народа, а станет великим злодеем, который полностью призирает все человеческие ценности, идет по головам и абсолютно не ценит никого из людей.
И на родителях лежит большая ответственность: кого они вырастят, тем он и будет.
Как правильно воспитывать детей с обонятельным вектором?
Легче, чем это может показаться. Обоняние — это врожденное правильное развитие и родителям, для того чтобы вырастить развитого обонятельника, необходимо меньше прилагать усилий, чем, например, в случае с звуковиком или зрительником. Вообще, обонятельный ребенок — это саморазвивающаяся личность и тут главное просто подтолкнуть его, направить в правильное русло, не мешать, а уж он сам научится правильно жить, выбирать себе компанию, найдет себе реализацию.
Неправильно будет запихивать в такого ребенка знания в огромном количестве. Люди с обонятельным вектором умные от природы, и для хорошей реализации в жизни им совершенно нет необходимости знать синусы или косинусы, пять иностранных языков или играть на скрипке вместе с фортепиано. Они принадлежат именно к тому разряду серых мышек в школе, которые учились едва-едва на тройки, а потом стали владельцами банков или заняли высокий пост на государственной службе уже в молодом возрасте. Потому что у них уже заданно врожденное сознание того, что действительно нужно в этой жизни, а что нет. И лучше как раз прислушиваться к ощущениям такого ребенка, чем идти наперекор ему. Они становятся великими финансистами без всякого образования. Они свои в этой системе, ощущают и чуют ее.
Дети с обонятельным вектором имеют свои, собственные отношения с запахами. У них обостренное обоняние и они буквально до тошноты не переваривают сильные запахи, от которых у них кружится голова, портиться настроение и сбиваются все ощущения.
Чем более чистым от ярких запахов будет окружающее его пространство, тем быстрее ребенок-меланхолик научится улавливать самые тонкие запахи, не заметные окружающим. Высшая форма развития обоняния — это умение по запахам определять настроение человека, его желания, свойства, качества. Ведь не секрет, что собаки могут в толпе легко определить плохого человека, негодяя, или например, выпившего, и облаять такового. Как они это делают? Именно по запаху, ведь плохой человек излучает плохой запах. И у человека с чувствительным носом может быть такой уровень развития, что он сможет различать запахи людей, понимая, кто несет угрозу, а кто — нормальный, положительный человек. Поэтому если в семье есть обонятельный ребенок, необходимо приложить все усилия, чтобы исключить любые химические и вообще сильные запахи.
Самое худшее, что могут сделать родители для своего обонятельного ребенка-меланхолика — это окружить его сверхзаботой, опекать его от проблем. Ведь именно проблемы — это то, что развивает такого ребенка, позволяет ему улучшать свои врожденные возможности правильно дифферинцировать людей. В принципе, ничто, ни побои, ни издевательства, не могут так повредить этому ребенку, как сверхзабота мамы, которая постоянно обнимает, целует, залюбливает — это настоящая трагедия.
Поэтому такому ребенку важно не сидеть дома, общаться с ровесниками, выходить в социум. Но проблема в том, что сам по себе обонятельник не хочет этого: он способен целыми днями сидеть в комнате, поглощенный собственной меланхолией и отсутствием эмоций. Заботливым родителям придется приложить огромные усилия, чтобы выгнать его в социум, во двор. Но это нужно сделать, ведь только там, среди людей он научится своим важным функциям и разовьется. В замкнутом пространстве, наедине с родителями, бабушками и дедушками, у него просто не будет объектов тренировки.
Внимание! Данная статья носит ознакомительный характер. Ребенок, кроме обонятельного вектора, может обладать еще одним или несколькими векторами и в его воспитании надо учитывать также и их. Если Вы хотите полностью и абсолютно понимать своего ребенка и знать, как его воспитывать, пройдите тренинг по системно-векторной психологии Юрия Бурлана. Чтобы попасть на бесплатные, вводные занятия, жмите на баннер внизу.
О воспитании детей с другими векторами читайте здесь. Если Вас интересует психология воспитания, обязательно оформите подписку в форме ниже.
Обонятельный вектор в лицах
Приглашаю прогуляться по импровизированной системно-векторной галлерее — знаменитые «обонятельники».
Абель — разведчик
Алан Гринспен — Председатель Федеральной Резервной Системы США
Арвид Карлсон — основоположник эры «антидепрессантов»
Герхард Домак — открытие Антибактериальные свойств протонзила, предшественник антибиотиков
Александр Флеминг — бактериолог, создатель пенициллина
Джордж Сорос — американский финансист, его называют финансовым спекулянтом, а также лоббистом наркотиков
Берия
Зоелик — Президент Всемирного Банка
Paul Wolfowitz — бывший президент всемирного банка, главный архитектор политики Буша в отношении Ирака
Harry Dexter White — отец Международного валютного фонда и Всемирного банка, заместитель министра финансов США
Познер
Dr.George Hitchings — разработал ряд лекарств против лейкимии, малярии итд. 1988 Премия нобеля
Кристина Лагард — бывшая министр финансов Франции
Sir James Whyte Black — ученый, открыл бэта-блокаторы, открытие, повлекшее появление противоязвенных препаратов
Зельман Абрахам Ваксман — открыл Стрептомицин, первый эффективный антибиотик против туберкулеза. Нобелевский лауреат
Henry Morgenthau — Министр финансов США, также архитектор плана Mass Starvation of Germans, 1945-1950 (Eisenhower’s Death Camps)
Источник: http://www.yburlan.ru/forum/
О том, что такое векторы, и в частности обонятельный вектор, и как они компонуют характер человека, можно почитать в библиотеке портала
и узнать на тренингах по системно-векторной психологии (расписание).
Векторная психология. Обонятельный вектор — мастер игры
Функция — устранение угроз выживанию, власть.
| квартель | корень | цвет | носители |
| энергия | ранжирование | фиолетовый | 1% |
Самый редко встречающийся в природе, самый непонятный и один из самых притягательных психотипов — это психотип мастера игры. Невероятно проницательные, осторожные, вкрадчивые люди, умеющие говорить с любым человеком на языке его мыслей и самых потаённых желаний. При полном внутреннем спокойствии, нулевой эмоциональной амплитуде, эти люди играют в самые масштабные политические игры, разыгрывают самые невероятные комбинации, плетут интриги и просто дурят честной люд по чём зря.
Единственным их настоящим желанием является получение власти и контроля над ситуацией, человеком или группой людей, в которой они оказались, но при этом не быть на виду у всех, оставаться в тени, — публичность им не особо интересна. Поэтому, чаще всего, такие люди стремятся стать советниками, заместителями руководителей, консультантами, сотрудниками службы HR, — то есть теми, кто влияет на многое, если не на всё, но несёт ограниченную ответственность.
Весь набор качеств характера и особенностей строения тела носителя этого психотипа работает на то, чтобы переиграть любого потенциального конкурента, а лучше — вообще любого человека, оказывать влияние на других, а самому не поддаваться ни чьему влиянию.
Обонятельный вектор. Плюсы.
-
Невероятная проницательность
по отношению к окружающим людям. Чтобы понять, насколько сильно это качество у мастера игры, нужно, конечно, понимать, как оно работает. Эта легендарная проницательность рождается вследствие полной сосредоточенности носителя этого психотипа на собеседнике, сосредоточенности настолько сильной, что внутри возникает то же самое ощущение, которое испытывает собеседник. Происходит буквально синхронизация своего состояния с состоянием другого человека. Ну и как возразить человеку, который стал воплощением тебя самого? -
Способность перевоплощаться
в разные роли в зависимости от обстановки. Это качество мы знаем по пословице «в политике нет друзей и врагов, в ней есть только деловые партнёры». Мастер игры, как никто другой, ощущает веяния времени, смены тенденций, и способен соотносить их с собственными целями. Однако, это только полдела, вторая часть мастерства — в каждой ситуации быть тем, кто сумеет извлечь из неё максимум: мало предложить хорошие условия сделки, намного важнее быть человеком, которого хочется иметь в качестве партнёра. В общем, реальные продажи начинаются не с продукта, а с продавца. -
Интуитивный интеллект
Это особая «чуйка жизни», которая ни на секунду не покидает носителя данного психотипа и позволяет ему играть на опережение. Вы слышали о случаях, когда люди не попадают на рейсы самолётов, или на корабли, которые потом терпят крушения? Помните, как Никола Тесла уговаривал своих друзей не садиться на Титаник? Это не частные непонятные случаи, это способность мастера игры, которую всегда использовали вожди, цари, короли, когда обращались к своим шаманам, магам и советникам за подсказкой: стоит ли начинать битву? Вступать ли в войну сейчас, или нужно немного подождать? Ну, вы же понимаете, кто на самом деле управляет, — тот, кто отдал приказ, или тот, кто посоветовал, когда надо отдать приказ?
Обонятельный вектор. Минусы.
-
Нелюбовь к людям,
если выражаться совсем уж мягко… к их желаниям, мыслям и целым судьбам, да и к человечеству в целом, чего уж там мелочиться. Нет, вы, конечно, никогда этого не почувствуете (ну, по крайней мере, пока не возникнет в этом необходимость, по мнению мастера игры). Он может быть с вами внимателен, может проявлять интерес к вашим переживаниям, и вам даже может показаться, что вы ему не безразличны. Важно только помнить, что этот интерес вызываете не вы, а желание изучить вас: чтобы было понятно, в какую игру нужно с вами играть. -
Гиперэгоизм,
игра всегда только в свои ворота. Носитель этого психотипа безразличен к интересам окружающих и учитывает эти интересы ровно настолько, насколько это выгодно для него самого. Вы уже знаете про отсутствие друзей и врагов: когда на кону собственная выгода, то, рано или поздно, любые близкие отношения встанут поперек этой выгоды, и придётся делать выбор. Мастер игры всегда выбирает лучшее решение, а потому, в любой ситуации, играет только за самого себя, и объединяется с другими в партнёрства по обстоятельствам. -
Неискренность, лживость, постоянная подмена фактов мнением
Мастер игры лидер по качествам со знаком «минус». Самый лучший способ не быть пойманным за язык — это предоставить говорить другим, никого ни в чём не убеждать и не спорить, а подкидывать информацию, которая способна повлиять на их решения. Вопрос только в том, какую же информацию подкинуть… Ответ нам даёт книга книг о военном деле «Искусство войны»: «Если ты хочешь мира — готовься к войне, если хочешь войны — показывай, что хочешь мира, если твои войска голодны — показывай, что сыты, если сыты — показывай, что голодны…» Думаю, принцип вы поняли. Возможно, для вас это покажется неприемлемо, а мастер игры так и живёт.
Обонятельный вектор. Качества.
В любви
В чём? В любви к себе мастер игры бесконечен в собственном самолюбовании, однако, никакой другой любви он не знает, так как она не совместима с внутренним состоянием войны.
В общении
Непонятный, пугающий, невероятно проницательный, догадливый, искусительный, непросчитываемый, интересный.
Мотивация
Власть.
Самооценка (относительно других психотипов)
Завышенная, «вы ниже меня».
Эмоциональность
Минимальная. Сильный эмоциональный стресс возможен, но очень быстро уступит место ровному меланхоличному состоянию.
В быту
Любит комфорт, но не роскошь, не демонстрирует свой достаток.
Творчество
Жизненная позиция
Активная, военная, очень жёсткая, с железной хваткой, но никогда не играет в открытую.
Главный подарок судьбы
Возможность влиять на других, а самому выскользнуть из-под влияния кого бы то ни было.
Ключ к развитию
Ставить цели выше и преодолевать больше трудностей.
Главная ловушка
Потерять бдительность от собственного успеха.
Источник силы
Интуиция.
Источник страха
Другие такие же (носители такого же вектора).
Отношение к переменам, тип восприятия нового
Изменения воспринимаются с удовольствием, как источник новых ощущений. Новизна является фактором возбуждения, источником возможностей для развития.
Факторы риска по здоровью
Заболевания носа и дыхательных путей, хронический насморк, аллергии, риск зрительной слепоты в старости, риск исчезновения волос на лице и голове к старости.
Обонятельный вектор. Развитость.
Как отличить более развитого носителя вектора от менее развитого?
Общее правило для определения развитости таково: более развитый человек принимает такие решения (и, соответственно, делает такие действия), в результате которых выигрывает большее количество человек; а менее развитый — такие решения (и действия), в результате которых выигрывает меньшее количество человек.
Таким образом, менее развитых мастеров игры мы можем наблюдать в роли магов, гадателей, шаманов, астрологов, шулеров, мошенников, организаторов сект, мелких манипуляторов, — личностей, которые используют свой дар проницательности с весьма сомнительной пользой для людей.
Профессии для максимальной реализации (наиболее развитые представители вектора): учёный-исследователь, химик, конструктор. Психолог, коуч, гипнотизёр. Модельер. Советник по безопасности, финансист, банкир, глава службы HR. Политик. Большой бизнесмен.
Обонятельный вектор. Реализованность.
Нереализованный вектор:
состояние, когда основные желания вектора не реализованы, человек хочет, но не имеет желаемого.
Нереализованный мастер игры выражен, чаще всего, созданием культа личности вокруг собственной персоны, подчёркиванием собственной уникальности на каждом шагу и претензиями на «избранность» себя как некоего особенного человека, не как все. Как понять, что есть проблема с реализованностью психотипа: если, по вашим представлениям, вы должны быть влиятельным человеком, но реальность такова, что ваши возможности малы, а круг вашего влияния узок.
Реализованный вектор
когда вы не ситуативно, а постоянно чувствуете себя хозяином положения, не подчиняетесь почти никому, а в сфере вашего влияния находится широкий круг людей. Вы чувствуете тенденции в изменениях общества и в настроении людей, и умеете оборачивать их в свою пользу. Вы умеете найти подход, ключики влияния, к любому человеку.
Мастера игры — люди с обонятельным вектором.
Семейство Рокфеллеров, семейство Ротшильдов, Иосиф Сталин, Владимир Путин, Борис Березовский, Александр Лукашенко, Сергей Лавров, Джорджио Армани, модельеры Дольче и Габбана, Сергей Брин, Карл Лагерфельд, Сидней Рейли, Феликс Дзержинский, Джордж Мартин, Джорж Сорос, Карлос Слим, Патрик Зюскинд, Николай Гоголь, Агата Кристи, Никола Тесла, Рупперт Мердок, Будда, большинство Римских Пап, пророк Мухаммед, Энштейн, Дмитрий Менделеев, Константин Циолковский, Казимир Малевич, Жанна Агузарова, мать Тереза, Владимир Познер, Борис Акунин, Ошо, Далай-Лама, Никита Хрущёв, Александр Друзь, Григорий Распутин.
Интраназальное введение нейротоксикантов животным: подтверждение гипотезы обонятельного вектора болезни Паркинсона
Причины болезни Паркинсона (БП) неизвестны, но есть свидетельства того, что воздействие факторов окружающей среды, включая ряд вирусов, токсинов, сельскохозяйственных химикатов, пищевых питательных веществ и металлов, в некоторых случаях связано с ее развитием. Наличие потери обоняния и патологическое поражение обонятельных путей на ранних стадиях болезни Паркинсона согласуются с утверждениями гипотезы обонятельного вектора.Эта гипотеза постулирует, что некоторые формы БП могут быть вызваны или катализироваться факторами окружающей среды, которые проникают в мозг через обонятельную слизистую оболочку. В этой статье мы представляем обзор доказательств, свидетельствующих о причастности ксенобиотиков к этиологии БП, и рассматриваем исследования на животных, в основном на грызунах, в которых токсиканты вводились в нос в попытке вызвать поведенческие или нейрохимические изменения, подобные тем, которые наблюдаются при БП. . Имеющиеся данные свидетельствуют о том, что этот путь воздействия приводит к весьма разным результатам в зависимости от задействованного ксенобиотика, истории воздействия, а также возраста и вида тестируемых животных.Некоторые соединения, такие как ротенон, паракват и 6-гидроксидофамин, обладают ограниченной способностью проникать в нигростриатальную дофаминергическую систему и повреждать ее интраназальным путем. Другие, такие как 1-метил-4-фенил-1,2,3,6-тетрагидропиридин (MPTP), легко проникают в мозг этим путем у некоторых видов и влияют на функцию нигростриатального пути. Интраназальное введение МРТР некоторым грызунам вызывает последовательность поведенческих и нейрохимических изменений, которые очень похожи на наблюдаемые при БП.По этой причине такая модель грызунов МРТР представляется экологически обоснованным средством оценки новых паллиативных методов лечения как моторных, так и немоторных симптомов БП. Однако необходимы дополнительные исследования этой и других экологически обоснованных моделей.
Обонятельная система человека при двух протеинопатиях: болезни Альцгеймера и болезни Паркинсона | Трансляционная нейродегенерация
Мастерс К.Л., Бейтман Р., Бленноу К., Роу К.С., Сперлинг Р.А., Каммингс Дж.Л.Болезнь Альцгеймера. Nat Rev Dis Primers. 2015; 1: 15056.
PubMed Статья Google Scholar
Поуэ В., Сеппи К., Таннер С.М., Холлидей Г.М., Брундин П., Фолькманн Дж., Шраг А.Е., Ланг А.Е. Болезнь Паркинсона. Nat Rev Dis Primers. 2017; 3: 17013.
PubMed Статья Google Scholar
Doty RL. Обонятельная дисфункция при нейродегенеративных заболеваниях: есть ли общий патологический субстрат? Lancet Neurol.2017; 16: 478–88.
PubMed Статья Google Scholar
Попытки Дж., Уокер Л., Джеллингер К.А. Поражение обонятельной луковицы при нейродегенеративных заболеваниях. Acta Neuropathol. 2014; 127: 459–75.
CAS PubMed Статья Google Scholar
Джакер М., Уокер Л.К. Самораспространение патогенных белковых агрегатов при нейродегенеративных заболеваниях. Природа.2013; 501: 45–51.
CAS PubMed PubMed Central Статья Google Scholar
Перри В.Х., Холмс К. Праймирование микроглии при нейродегенеративном заболевании. Nat Rev Neurol. 2014; 10: 217–24.
CAS PubMed PubMed Central Статья Google Scholar
van den Heuvel MP, Sporns O. Перекрестный беспорядочный коннектомный ландшафт дисконнектомности мозга.Nat Rev Neurosci. 2019; 20: 435–46.
PubMed Статья CAS Google Scholar
Nieuwenhuys R, Voogd J, van Huijzen C. Центральная нервная система человека. Амстердам: Спрингер; 2008.
Книга. Google Scholar
Pena-Melian A. Cabello-de la Rosa JP, Gallardo-Alcaniz MJ, Vaamonde-Gamo J, Relea-Calatayud F, Gonzalez-Lopez L, Villanueva-Anguita P, Flores-Cuadrado A, Saiz- Санчес Д., Мартинес-Маркос А. Черепная пара 0: Терминальный нерв.Анат Рек (Хобокен). 2019; 302: 394–404.
Артикул Google Scholar
Ubeda-Banon I, Pro-Sistiaga P, Mohedano-Moriano A, Saiz-Sanchez D, de la Rosa-Prieto C, Gutierrez-Castellanos N, Lanuza E, Martinez-Garcia F, Martinez-Marcos A Кладистический анализ обонятельной и вомероназальной систем. Фронт нейроанат. 2011; 5: 3.
PubMed PubMed Central Статья Google Scholar
Doty RL. Справочник по обонянию и вкусу. Нью-Йорк: Марсель Деккер; 2003.
Книга Google Scholar
Цена JL. Обонятельная система. В: Паксинос Г., редактор. Нервная система человека. Сан-Диего: Academic Press; 1990. стр. 979–98.
Глава Google Scholar
ван Хартевельт Т.Дж., Крингельбах М.Л.: Обонятельная система. В нервной системе человека.3-й. Май Дж. К., Паксинос Г. Сан-Диего: Academic Press; 2012: 1219–1238.
Глава Google Scholar
Мартинес-Маркос А. Об организации обонятельной и вомероназальной коры. Prog Neurobiol. 2009; 87: 21–30.
PubMed Статья Google Scholar
Mohedano-Moriano A, de la Rosa-Prieto C, Saiz-Sanchez D, Убеда-Banon I, Pro-Sistiaga P, de Moya-Pinilla M, Martinez-Marcos A.Центробежные теленцефальные афферентные связи с основными и вспомогательными обонятельными луковицами. Фронт нейроанат. 2012; 6:19.
PubMed PubMed Central Статья Google Scholar
Флорес-Куадрадо А., Саис-Санчес Д., Мохедано-Мориано А., Мартинес-Маркос А., Убеда-Банон I. Нейродегенерация и индукция контралатерального альфа-синуклеина после внутримозговых инъекций альфа-синуклеина в переднее обонятельное ядро модель мыши A53T с болезнью Паркинсона.Acta Neuropathol Commun. 2019; 7: 56.
PubMed PubMed Central Статья Google Scholar
Стандринг Анатомия С. Грея. Анатомические основы клинической практики. Сороковая изд. Лондон: Эльзевир; 2008.
Google Scholar
Doty RL. Обоняние при болезни Паркинсона и связанных с ней расстройствах. Neurobiol Dis. 2012; 46: 527–52.
PubMed Статья Google Scholar
Graziadei PP, Graziadei GA. Нейрогенез и регенерация нейронов обонятельной системы млекопитающих. I. Морфологические аспекты дифференцировки и структурной организации обонятельных сенсорных нейронов. J Neurocytol. 1979; 8: 1–18.
CAS PubMed Статья Google Scholar
Арнольд С.Е., Ли Э.Б., Моберг П.Дж., Штутцбах Л., Кази Х., Хан Л.Й., Ли В.М., Трояновски Дж.К. Бета-амилоид обонятельного эпителия и тау-патология парных спиральных нитей при болезни Альцгеймера.Энн Нейрол. 2010; 67: 462–9.
CAS PubMed PubMed Central Статья Google Scholar
Арнольд С.Е., Смутцер Г.С., Трояновский Д.К., Моберг П.Дж. Клеточная и молекулярная невропатология обонятельного эпителия и центральных обонятельных путей при болезни Альцгеймера и шизофрении. Ann N Y Acad Sci. 1998; 855: 762–75.
CAS PubMed Статья Google Scholar
Ли Дж. Х., Гёдерт М., Хилл В. Д., Ли В. М., Трояновский Дж. Тау-белки аномально экспрессируются в обонятельном эпителии пациентов с болезнью Альцгеймера и регулируются в спинном мозге плода человека. Exp Neurol. 1993; 121: 93–105.
CAS PubMed Статья Google Scholar
Сайто Ю., Шиоя А., Сано Т., Сумикура Х, Мурата М., Мураяма С. Патология тельца Леви включает обонятельные клетки при болезни Паркинсона и связанных с ней расстройствах.Mov Disord. 2016; 31: 135–8.
CAS PubMed Статья Google Scholar
Martinez-Marcos A, Sanudo JR. Черепные нервы: морфология и клиническое значение. Анат Рек (Хобокен). 2019; 302: 555–7.
Артикул Google Scholar
Мартинес-Маркос А., Санудо-младший. Черепные нервы: филогения и онтогенез. Анат Рек (Хобокен). 2019; 302: 378–80.
Артикул Google Scholar
Crosby EC, Humphrey T. Исследования конечного мозга позвоночных. II. Ядерная структура переднего обонятельного ядра, tuberculum olfactorium и миндалевидного комплекса у взрослого человека. J Comp Neurol. 1941; 74: 193–352.
Артикул Google Scholar
Смит Р.Л., Бейкер Х., Колстад К., Спенсер Д.Д., Грир, Калифорния. Локализация иммунореактивности тирозингидроксилазы и белка обонятельного маркера в обонятельной луковице человека и макака.Brain Res. 1991; 548: 140–8.
CAS PubMed Статья Google Scholar
Smith RL, Baker H, Greer CA. Иммуногистохимический анализ обонятельной луковицы человека. J Comp Neurol. 1993; 333: 519–30.
CAS PubMed Статья Google Scholar
Crespo C, Liberia T, Blasco-Ibanez JM, Nacher J, Varea E. Черепная пара I: обонятельный нерв.Анат Рек (Хобокен). 2019; 302: 405–27.
Артикул Google Scholar
Убеда-Банон I, Саис-Санчес Д., де ла Роса-Прието К. Мартинес-Маркос a: альфа-синуклеин в обонятельной системе при болезни Паркинсона: роль нейронных связей в распространении патологии. Функция структуры мозга. 2014; 219: 1513–26.
CAS PubMed Google Scholar
Ubeda-Banon I, Saiz-Sanchez D, de la Rosa-Prieto C, Argandona-Palacios L, Garcia-Munozguren S.Мартинес-Маркос a: альфа-синуклеинопатия в обонятельной системе человека при болезни Паркинсона: вовлечение кальций-связывающих белков и положительных по веществу P клеток. Acta Neuropathol. 2010. 119: 723–35.
CAS PubMed Статья Google Scholar
Mohedano-Moriano A, Martinez-Marcos A, Munoz M, Arroyo-Jimenez MM, Marcos P, Artacho-Perula E, Blaizot X, Insausti R. Взаимные связи между обонятельными структурами и корой верхнего рострального слоя височная борозда у обезьяны Macaca fascicularis.Eur J Neurosci. 2005; 22: 2503–18.
CAS PubMed Статья Google Scholar
Ом TG, Muller H, Braak E. Calbindin-D-28k-подобные иммунореактивные структуры в обонятельной луковице и переднем обонятельном ядре взрослого человека: распределение и типология клеток — частичная комплементарность с парвальбумином. Неврология. 1991; 42: 823–40.
CAS PubMed Статья Google Scholar
Саис-Санчес Д., Убеда-Банон I, де ла Роса-Прието С., Аргандона-Паласиос Л., Гарсия-Муньосгурен С., Инсаусти Р., Мартинес-Маркос А. Соматостатин, тау и бета-амилоид в переднем обонятельном ядре в Болезнь Альцгеймера. Exp Neurol. 2010; 223: 347–50.
CAS PubMed Статья Google Scholar
Убеда-Банон I, Флорес-Куадрадо А., Саис-Санчес Д., Мартинес-Маркос А. Дифференциальные эффекты болезни Паркинсона на подтипы интернейронов в переднем обонятельном ядре человека.Фронт нейроанат. 2017; 11: 113.
PubMed PubMed Central Статья CAS Google Scholar
Ом Т.Г., Браак Х. Изменения обонятельной луковицы при болезни Альцгеймера. Acta Neuropathol. 1987. 73: 365–9.
CAS PubMed Статья Google Scholar
Сэнгоку Р., Сайто Й., Икемура М., Хацута Х., Сакияма Й., Канемару К., Араи Т., Савабе М., Танака Н., Мотидзуки Х. и др.Частота и степень связанной с тельцами Леви альфа-синуклеинопатии у стареющих обонятельных луковиц человека. J Neuropathol Exp Neurol. 2008. 67: 1072–83.
PubMed Статья Google Scholar
Insausti R, Marcos P, Arroyo-Jimenez MM, Blaizot X, Martinez-Marcos A. Сравнительные аспекты обонятельной части энторинальной коры и ее проекции на гиппокамп у грызунов, нечеловеческих приматов и человека мозг. Brain Res Bull.2002; 57: 557–60.
CAS PubMed Статья Google Scholar
Ibarretxe-Bilbao N, Junque C, Marti MJ, Valldeoriola F, Vendrell P, Bargallo N, Zarei M, Tolosa E. Нарушение обоняния при болезни Паркинсона и аномалии белого вещества в центральных обонятельных областях: на основе вокселей исследование визуализации тензора диффузии. Mov Disord. 2010; 25: 1888–94.
PubMed Статья Google Scholar
Woodward MR, Dwyer MG, Bergsland N, Hagemeier J, Zivadinov R, Benedict RH, Szigeti K. Дефицит обонятельной идентификации предсказывает нарушение тракта белого вещества при болезни Альцгеймера. Психиатрия Res Neuroimaging. 2017; 266: 90–5.
PubMed PubMed Central Статья Google Scholar
Zald DH, Pardo JV. Функциональная нейровизуализация обонятельной системы человека. Int J Psychophysiol. 2000; 36: 165–81.
CAS PubMed Статья Google Scholar
Dourlen P, Kilinc D, Malmanche N, Chapuis J, Lambert JC. Новый генетический ландшафт болезни Альцгеймера: от амилоидного каскада до гипотезы генетически обусловленного синаптического сбоя? Acta Neuropathol; 2019.
Google Scholar
Genin E, Hannequin D, Wallon D, Sleegers K, Hiltunen M, Combarros O, Bullido MJ, Engelborghs S, De Deyn P, Berr C., et al. APOE и болезнь Альцгеймера: главный ген с полудоминантным наследованием.Мол Психиатрия. 2011; 16: 903–7.
CAS PubMed PubMed Central Статья Google Scholar
Scheltens P, Blennow K, Breteler MM, de Strooper B, Frisoni GB, Salloway S, Van der Flier WM. Болезнь Альцгеймера. Ланцет. 2016; 388: 505–17.
CAS PubMed Статья PubMed Central Google Scholar
Винтерс ХВ. Новые концепции болезни Альцгеймера.Анну Рев Патол. 2015; 10: 291–319.
CAS PubMed Статья Google Scholar
Tromp D, Dufour A, Lithfous S, Pebayle T, Despres O. Эпизодическая память при нормальном старении и болезни Альцгеймера: выводы из визуализации и поведенческих исследований. Aging Res Rev.2015; 24: 232–62.
CAS PubMed Статья Google Scholar
Goedert M, Spillantini MG.Век болезни Альцгеймера. Наука. 2006; 314: 777–81.
CAS PubMed Статья Google Scholar
Thal DR, Walter J, Saido TC, Fandrich M. Невропатология и биохимия Abeta и ее совокупностей при болезни Альцгеймера. Acta Neuropathol. 2015; 129: 167–82.
CAS PubMed Статья Google Scholar
Braak H, Alafuzoff I, Arzberger T, Kretzschmar H, Del Tredici K.Определение стадии нейрофибриллярной патологии, связанной с болезнью Альцгеймера, с использованием парафиновых срезов и иммуноцитохимии. Acta Neuropathol. 2006; 112: 389–404.
PubMed PubMed Central Статья Google Scholar
Postuma RB, Berg D, Stern M, Poewe W, Olanow CW, Oertel W, Obeso J, Marek K, Litvan I, Lang AE и др. Клинические диагностические критерии МДС болезни Паркинсона. Mov Disord. 2015; 30: 1591–601.
PubMed PubMed Central Статья Google Scholar
Polymeropoulos MH, Lavedan C, Leroy E, Ide SE, Dehejia A, Dutra A, Pike B, Root H, Rubenstein J, Boyer R, et al. Мутация в гене альфа-синуклеина идентифицирована в семьях с болезнью Паркинсона. Наука. 1997. 276: 2045–7.
CAS PubMed PubMed Central Статья Google Scholar
Kalia LV, Lang AE. Болезнь Паркинсона. Ланцет. 2015; 386: 896–912.
CAS PubMed PubMed Central Статья Google Scholar
Obeso JA, Stamelou M, Goetz CG, Poewe W, Lang AE, Weintraub D, Burn D, Halliday GM, Bezard E, Przedborski S и др. Прошлое, настоящее и будущее болезни Паркинсона: специальный очерк к 200-летию дрожащего паралича. Mov Disord. 2017; 32: 1264–310.
CAS PubMed PubMed Central Статья Google Scholar
Dickson DW, Braak H, Duda JE, Duyckaerts C, Gasser T., Halliday GM, Hardy J, Leverenz JB, Del Tredici K, Wszolek ZK, Litvan I.Невропатологическая оценка болезни Паркинсона: уточнение диагностических критериев. Lancet Neurol. 2009. 8: 1150–7.
CAS PubMed Статья Google Scholar
Годерт М., Спиллантини М.Г., Дель Тредичи К., Браак Х. 100 лет патологии Леви. Nat Rev Neurol. 2013; 9: 13–24.
CAS PubMed Статья Google Scholar
Spillantini MG, Schmidt ML, Lee VM, Trojanowski JQ, Jakes R, Goedert M.Альфа-синуклеин в тельцах Леви. Природа. 1997; 388: 839–40.
CAS PubMed PubMed Central Статья Google Scholar
Браак Х., Дель Тредичи К., Руб У, де Вос Р.А., Янсен Штер Э.Н., Браак Э. Стадия патологии головного мозга, связанной со спорадической болезнью Паркинсона. Neurobiol Aging. 2003. 24: 197–211.
PubMed Статья Google Scholar
Braak H, Bohl JR, Muller CM, Rub U, de Vos RA, Del Tredici K.Лекция Стэнли Фана 2005 г .: Пересмотр процедуры постановки патологии тельца включения, связанной со спорадической болезнью Паркинсона. Mov Disord. 2006 (21): 2042–51.
Толедо Дж. Б., Гопал П., Райбл К., Ирвин Д. Д., Бретчнайдер Дж., Седор С., Уэйтс К., Болуда С., Гроссман М., Ван Дирлин В. М. и др. Патологическое распределение альфа-синуклеина у субъектов с совпадающей патологией с тельцами Альцгеймера и Леви. Acta Neuropathol. 2016; 131: 393–409.
CAS PubMed Статья Google Scholar
Dubois B, Hampel H, Feldman HH, Scheltens P, Aisen P, Andrieu S, Bakardjian H, Benali H, Bertram L, Blennow K, et al. Доклиническая болезнь Альцгеймера: определение, естественное течение и диагностические критерии. Демент Альцгеймера. 2016; 12: 292–323.
PubMed PubMed Central Статья Google Scholar
Sheng C, Huang Y, Han Y. Рассечение продромальной болезни Альцгеймера. Front Biosci (Landmark Ed). 2018; 23: 1272–91.
CAS Статья Google Scholar
Сперлинг Р.А., Карлавиш Дж., Джонсон К.А. Доклиническая болезнь Альцгеймера — проблемы впереди. Nat Rev Neurol. 2013; 9: 54–8.
CAS PubMed Статья Google Scholar
Обесо Дж. А., Родригес-Ороз М. С., Гетц К. Г., Марин С., Кордауэр Дж. Х., Родригес М., Хирш Е. К., Фаррер М., Шапира А. Х., Халлидей Г. Недостающие части в головоломке о болезни Паркинсона.Nat Med. 2010. 16: 653–61.
CAS Статья PubMed Google Scholar
Hawkes CH. Продромальная фаза спорадической болезни Паркинсона: существует ли она и как долго длится она? Mov Disord. 2008; 23: 1799–807.
PubMed Статья Google Scholar
Postuma RB, Berg D. Продромальная болезнь Паркинсона: десятилетие в прошлом, десятилетие в будущем. Mov Disord.2019; 34: 665–75.
PubMed Статья Google Scholar
Schapira AHV, Chaudhuri KR, Jenner P. Немоторные особенности болезни Паркинсона. Nat Rev Neurosci. 2017; 18: 509.
CAS PubMed PubMed Central Статья Google Scholar
Berg D, Postuma RB, Adler CH, Bloem BR, Chan P, Dubois B., Gasser T., Goetz CG, Halliday G, Joseph L, et al.Критерии исследования МДС продромальной болезни Паркинсона. Mov Disord. 2015; 30: 1600–11.
PubMed Статья Google Scholar
Burns A. Может ли обонятельная дисфункция быть маркером ранней болезни Альцгеймера? Ланцет. 2000; 355: 84–5.
CAS PubMed Статья Google Scholar
Доти Р.Л., Димс Д.А., Стеллар С. Обонятельная дисфункция при паркинсонизме: общий дефицит, не связанный с неврологическими признаками, стадией заболевания или продолжительностью заболевания.Неврология. 1988; 38: 1237–44.
CAS PubMed Статья Google Scholar
Benarroch EE. Обонятельная система: функциональная организация и участие в нейродегенеративных заболеваниях. Неврология. 2010; 75: 1104–9.
PubMed Статья Google Scholar
Девананд Д.П., Ли С., Мэнли Дж., Эндрюс Х., Шупф Н., Доти Р.Л., Стерн Й., Заходне Л.Б., Луи Э.Д., Майё Р.Обонятельный дефицит предсказывает снижение когнитивных функций и деменцию Альцгеймера в городском сообществе. Неврология. 2015; 84: 182–9.
CAS PubMed PubMed Central Статья Google Scholar
Лафай-Маньян М.Э., Пуарье Дж., Этьен П., Трембле-Мерсье Дж., Френетт Дж., Роса-Нето П., Брейтнер Дж.С.С., Группа П-АР. Идентификация запаха как биомаркер доклинической БА у пожилых людей из группы риска. Неврология. 2017; 89: 327–35.
PubMed PubMed Central Статья Google Scholar
Робертс Р.О., Кристиансон Т.Дж., Кремерс В.К., Мильке М.М., Мачульда М.М., Василаки М., Альхурани Р.Э., Геда Ю.Е., Кнопман Д.С., Петерсен Р.С. Связь между обонятельной дисфункцией и амнестическими легкими когнитивными нарушениями и деменцией при болезни Альцгеймера. JAMA Neurol. 2016; 73: 93–101.
PubMed PubMed Central Статья Google Scholar
Reijs BLR, Ramakers I, Elias-Sonnenschein L, Teunissen CE, Koel-Simmelink M, Tsolaki M, Wahlund LO, Waldemar G, Hausner L, Johannsen P, et al.Связь идентификации запаха с маркерами болезни Альцгеймера в спинномозговой жидкости и познании. J. Alzheimers Dis. 2017; 60: 1025–34.
CAS PubMed Статья Google Scholar
Graves AB, Bowen JD, Rajaram L, McCormick WC, McCurry SM, Schellenberg GD, Larson EB. Нарушение обоняния как маркер снижения когнитивных функций: взаимодействие с аполипопротеином E эпсилон4-статус. Неврология. 1999; 53: 1480–7.
CAS PubMed Статья Google Scholar
Olofsson JK, Larsson M, Roa C, Wilson DA, Jonsson Laukka E. Взаимодействие между дефицитом идентификации запаха и APOE4 предсказывает 6-летнее снижение когнитивных функций у пожилых людей. Behav Genet. 2019.
Zhang J, Hao C, Jiang J, Feng Y, Chen X, Zheng Y, Liu J, Zhang Z, Long C, Yang L. Механизмы, лежащие в основе обонятельного дефицита у мышей с дефицитом аполипопротеина E: сосредоточиться на обонятельном эпителии и обонятельной луковице. Neurobiol Aging. 2018; 62: 20–33.
CAS PubMed Статья Google Scholar
Мерфи К. Обонятельные и другие сенсорные нарушения при болезни Альцгеймера. Nat Rev Neurol. 2019; 15: 11–24.
CAS PubMed Статья Google Scholar
Ю Х.С., Чон С., Чунг С.Дж., Юн М., Ли PH, Сон Й.Х., Эванс А.С., Е Б.С. Обонятельная дисфункция при когнитивных нарушениях, связанных с болезнью Альцгеймера и тельцами Леви. Демент Альцгеймера. 2018; 14: 1243–52.
PubMed Статья Google Scholar
Doty RL. Обонятельная дисфункция при болезни Паркинсона. Nat Rev Neurol. 2012; 8: 329–39.
CAS PubMed Статья Google Scholar
Понсен М.М., Стофферс Д., Буйдж Дж., Ван Эк-Смит Б.Л., Вольтерс Е., Берендсе Х.В. Идиопатическая гипосмия как доклинический признак болезни Паркинсона. Энн Нейрол. 2004; 56: 173–81.
PubMed Статья Google Scholar
Росс Г.В., Петрович Х., Эбботт Р.Д., Таннер С.М., Поппер Дж., Масаки К., Лаунер Л., Уайт Л.Р. Связь обонятельной дисфункции с риском развития болезни Паркинсона в будущем. Энн Нейрол. 2008; 63: 167–73.
PubMed Статья Google Scholar
Баба Т., Кикучи А., Хираяма К., Нишио Ю., Хосокай Ю., Канно С., Хасэгава Т., Сугено Н., Конно М., Сузуки К. и др. Тяжелая обонятельная дисфункция — продромальный симптом деменции, связанной с болезнью Паркинсона: трехлетнее продольное исследование.Мозг. 2012; 135: 161–9.
PubMed Статья Google Scholar
Морли Дж. Ф., Коэн А., Сильвейра-Морияма Л., Лис А. Дж., Уильямс Д. Р., Каценшлагер Р., Хоукс С., Штракс Дж. П., Вайнтрауб Д., Доти Р. Л., Дуда Дж. Э. Оптимизация обонятельного тестирования для диагностики болезни Паркинсона: анализ элементов теста идентификации запаха Университета Пенсильвании. NPJ Parkinsons Dis. 2018; 4: 2.
PubMed PubMed Central Статья Google Scholar
Duyckaerts C, Delatour B, Potier MC. Классификация и основные патологии болезни Альцгеймера. Acta Neuropathol. 2009. 118: 5–36.
CAS PubMed Статья Google Scholar
Пирсон Р.С., Эсири М.М., Хайорнс Р.В., Уилкок Г.К., Пауэлл Т.П. Анатомические корреляты распределения патологических изменений неокортекса при болезни Альцгеймера. Proc Natl Acad Sci U S. A. 1985; 82: 4531–4.
CAS PubMed PubMed Central Статья Google Scholar
Mrdjen D, Fox EJ, Bukhari SA, Montine KS, Bendall SC, Montine TJ. Основа клеточной и региональной уязвимости при болезни Альцгеймера. Acta Neuropathol. 2019; 138: 729–49.
PubMed Статья Google Scholar
Браак Х., Браак Э. Невропатологическая стадия изменений, связанных с болезнью Альцгеймера. Acta Neuropathol. 1991; 82: 239–59.
CAS PubMed PubMed Central Статья Google Scholar
Браак Х, Браак Э, Бол Дж, Братцке Х. Эволюция корковых поражений, связанных с болезнью Альцгеймера. J Neural Transm Suppl. 1998. 54: 97–106.
CAS PubMed Статья Google Scholar
Браак Х., Дель Тредичи К. Доклиническая фаза патологического процесса, лежащего в основе спорадической болезни Альцгеймера. Мозг. 2015; 138: 2814–33.
PubMed Статья Google Scholar
Esiri MM, Wilcock GK. Обонятельные луковицы при болезни Альцгеймера. J Neurol Neurosurg Psychiatry. 1984; 47: 56–60.
CAS PubMed PubMed Central Статья Google Scholar
Struble RG, Clark HB. Поражения обонятельной луковицы при болезни Альцгеймера. Neurobiol Aging. 1992; 13: 469–73.
CAS PubMed Статья Google Scholar
ter Laak HJ, Renkawek K, van Workum FP.Обонятельная луковица при болезни Альцгеймера: морфологическое исследование потери нейронов, клубков и старческих бляшек в связи с обонянием. Alzheimer Dis Assoc Disord. 1994; 8: 38–48.
PubMed Статья Google Scholar
Kovacs T, Cairns NJ. Lantos PL: отложение бета-амилоида и образование нейрофибриллярных клубков в обонятельной луковице при старении и болезни Альцгеймера. Neuropathol Appl Neurobiol. 1999; 25: 481–91.
CAS PubMed Статья Google Scholar
Kovacs T, Кэрнс, штат Нью-Джерси, Lantos PL. Обонятельные центры при болезни Альцгеймера: обонятельная луковица участвует на ранних стадиях болезни Браака. Нейроотчет. 2001; 12: 285–8.
CAS PubMed Статья Google Scholar
Tsuboi Y, Wszolek ZK, Graff-Radford NR, Cookson N, Dickson DW. Патология тау-белка в обонятельной луковице коррелирует со стадией Браака, патологией с тельцами Леви и аполипопротеином эпсилон4. Neuropathol Appl Neurobiol. 2003; 29: 503–10.
CAS PubMed Статья Google Scholar
Джеллингер К.А., Аттемс Дж. Патология болезни Альцгеймера в обонятельной луковице. Neuropathol Appl Neurobiol. 2005; 31: 203.
CAS PubMed Статья Google Scholar
Attems J, Jellinger KA. Патология обонятельного тау при болезни Альцгеймера и легких когнитивных нарушениях. Clin Neuropathol. 2006; 25: 265–71.
CAS PubMed Google Scholar
Саис-Санчес Д., Де ла Роса-Прието С., Убеда-Банон I, Мартинес-Маркос А. Интернейроны, тау и бета-амилоид в грушевидной коре при болезни Альцгеймера. Функция структуры мозга. 2015; 220: 2011–25.
CAS PubMed Статья Google Scholar
Saiz-Sanchez D, Flores-Cuadrado A, Ubeda-Banon I, de la Rosa-Prieto C, Martinez-Marcos A.Интернейроны обонятельной системы человека при болезни Альцгеймера. Exp Neurol. 2016; 276: 13–21.
CAS PubMed Статья Google Scholar
Del Tredici K, Rub U, De Vos RA, Bohl JR, Braak H. Где в головном мозге начинается патология болезни Паркинсона? J Neuropathol Exp Neurol. 2002; 61: 413–26.
PubMed Статья Google Scholar
Braak H, Muller CM, Rub U, Ackermann H, Bratzke H, de Vos RA, Del Tredici K.Патология, связанная со спорадической болезнью Паркинсона — чем она заканчивается? J Neural Transm Suppl. 2006: 89–97.
Daniel SE, Hawkes CH. Предварительная диагностика болезни Паркинсона по патологии обонятельной луковицы. Ланцет. 1992; 340: 186.
CAS PubMed Статья Google Scholar
Hoogland PV, van den Berg R, Huisman E. Неправильно проложенные обонятельные волокна и эктопические обонятельные клубочки у нормальных людей и у пациентов с болезнью Паркинсона и Альцгеймера.Neuropathol Appl Neurobiol. 2003; 29: 303–11.
CAS PubMed Статья Google Scholar
Хаббард П.С., Эсири М.М., Ридинг М., МакШейн Р., Надь З. Патология альфа-синуклеина в обонятельных путях у пациентов с деменцией. J Anat. 2007; 211: 117–24.
PubMed PubMed Central Статья Google Scholar
Силвейра-Морияма Л., Холтон Дж. Л., Кингсбери А., Эйлинг Х., Петри А., Стерлаччи В., Поу В., Майер Х., Лис А. Дж., Ревес Т.Региональные различия в степени тяжести патологии тельцов Леви в обонятельной коре. Neurosci Lett. 2009; 453: 77–80.
CAS PubMed Статья Google Scholar
Beach TG, White CL 3rd, Hladik CL, Sabbagh MN, Connor DJ, Shill HA, Sue LI, Sasse J, Bachalakuri J, Henry-Watson J, et al. Альфа-синуклеинопатия обонятельной луковицы обладает высокой специфичностью и чувствительностью к заболеваниям с тельцами Леви. Acta Neuropathol. 2009; 117: 169–74.
CAS PubMed Статья Google Scholar
Еллингер К.А. Альфа-синуклеинопатия обонятельной луковицы обладает высокой специфичностью и чувствительностью к заболеваниям с тельцами Леви. Acta Neuropathol. 2009; 117: 215–6, ответ автора 217–218.
PubMed Статья Google Scholar
Parkkinen L, Silveira-Moriyama L, Holton JL, Lees AJ, Revesz T. Может ли биопсия обонятельной луковицы быть оправданной для диагностики болезни Паркинсона? Комментарии на «Альфа-синуклеинопатия обонятельной луковицы обладает высокой специфичностью и чувствительностью к заболеваниям с тельцами Леви».Acta Neuropathol. 2009; 117: 213–4, ответ автора 217–218.
PubMed Статья Google Scholar
Табатон М., Монако С., Кордоне М.П., Колуччи М., Джакконе Дж., Тальявини Ф., Зануссо Г. Отложение прионов при обонятельной биопсии спорадической болезни Крейтцфельда-Якоба. Энн Нейрол. 2004; 55: 294–6.
PubMed Статья Google Scholar
Дуда Ю.Е. Патология обонятельной системы как модель нейродегенеративного заболевания Леви.J Neurol Sci. 2010. 289: 49–54.
CAS PubMed Статья Google Scholar
Kesslak JP, Nalcioglu O, Cotman CW. Количественная оценка магнитно-резонансного сканирования атрофии гиппокампа и парагиппокампа при болезни Альцгеймера. Неврология. 1991; 41: 51–4.
CAS PubMed Статья Google Scholar
Ди Паола M, Macaluso E, Carlesimo GA, Tomaiuolo F, Worsley KJ, Fadda L, Caltagirone C.Нарушение эпизодической памяти у пациентов с болезнью Альцгеймера коррелирует с атрофией энторинальной коры. Исследование морфометрии на основе вокселей. J Neurol. 2007. 254: 774–81.
PubMed Статья Google Scholar
Prestia A, Drago V, Rasser PE, Bonetti M, Thompson PM, Frisoni GB. Корковые изменения при начальной болезни Альцгеймера. J. Alzheimers Dis. 2010; 22: 1339–49.
PubMed Статья Google Scholar
Имабаяси Э., Мацуда Х., Табира Т., Арима К., Араки Н., Исии К., Ямасита Ф., Ивацубо Т. Японская нейровизуализация болезни Альцгеймера I: сравнение КТ головного мозга и МРТ для морфометрии болезни Альцгеймера на основе вокселей. Brain Behav. 2013; 3: 487–93.
PubMed PubMed Central Статья Google Scholar
Васавада М.М., Ван Дж., Эслингер П.Дж., Джилл Диджей, Сан Икс, Карунанаяка П., Ян QX. Дегенерация обонятельной коры при болезни Альцгеймера и легкие когнитивные нарушения.J. Alzheimers Dis. 2015; 45: 947–58.
PubMed Статья Google Scholar
Thomann PA, Dos Santos V, Toro P, Schonknecht P, Essig M, Schroder J. Уменьшение объема обонятельной луковицы и тракта при ранней стадии болезни Альцгеймера — исследование МРТ. Neurobiol Aging. 2009; 30: 838–41.
PubMed Статья Google Scholar
Thomann PA, Dos Santos V, Seidl U, Toro P, Essig M, Schroder J.Атрофия обонятельной луковицы и тракта на основе МРТ при легких когнитивных нарушениях и болезни Альцгеймера. J. Alzheimers Dis. 2009; 17: 213–21.
PubMed Статья Google Scholar
Сервелло А., Фиоретти А., Гуальди Дж., Ди Биази С., Питталис А., Соллаку С., Павачи С., Торторелла Ф., Фузетти М., Валенти М. и др. Обонятельная дисфункция, объем обонятельной луковицы и болезнь Альцгеймера: есть ли корреляция? Пилотное исследование 1. J. Alzheimers Dis.2015; 48: 395–402.
PubMed Статья Google Scholar
Vogels OJ. Болезнь Альцгеймера: стереология в поисках специфических маркеров. Neurobiol Aging. 1994; 15: 373–5 дискуссия 379–380.
CAS PubMed Статья Google Scholar
West MJ, Coleman PD, Flood DG, Troncoso JC. Различия в характере потери нейронов гиппокампа при нормальном старении и болезни Альцгеймера.Ланцет. 1994; 344: 769–72.
CAS PubMed Статья Google Scholar
Gomez-Isla T, Price JL, McKeel DW Jr, Morris JC, Growdon JH, Hyman BT. Глубокая потеря нейронов энторинальной коры второго слоя происходит при очень легкой форме болезни Альцгеймера. J Neurosci. 1996; 16: 4491–500.
CAS PubMed PubMed Central Статья Google Scholar
Прайс Дж.Л., Ко А.И., Уэйд М.Дж., Цу С.К., МакКил Д.В., Моррис Дж. С..Число нейронов в энторинальной коре и СА1 при доклинической болезни Альцгеймера. Arch Neurol. 2001; 58: 1395–402.
CAS PubMed Статья Google Scholar
Вест М.Дж., Кавас С.Х., Стюарт В.Ф., Рудоу Г.Л., Тронкосо Дж.С. Нейроны гиппокампа в доклинической болезни Альцгеймера. Neurobiol Aging. 2004. 25: 1205–12.
CAS PubMed Статья Google Scholar
Крил Дж., Ходжес Дж., Холлидей Г. Связь между объемом гиппокампа и потерей нейронов CA1 в головном мозге людей с болезнью Альцгеймера и без нее. Neurosci Lett. 2004; 361: 9–12.
CAS PubMed Статья Google Scholar
фон Гунтен А., Ковари Э., Ривара С.Б., Борас С., Хоф П.Р., Джаннакопулос П. Стереологический анализ патологии гиппокампа при болезни Альцгеймера у самых старых и пожилых людей: доказательства сохранения энторинальной коры и поля CA1.Exp Neurol. 2005; 193: 198–206.
Артикул CAS Google Scholar
фон Гунтен А., Ковари Э, Бюссьер Т., Ривара С.Б., Голд Г., Бурас С., Хоф П.Р., Джаннакопулос П. Когнитивное влияние нейрональной патологии в энторинальной коре и поле СА1 при болезни Альцгеймера. Neurobiol Aging. 2006; 27: 270–7.
Артикул CAS Google Scholar
Giannakopoulos P, von Gunten A, Kovari E, Gold G, Herrmann FR, Hof PR, Bouras C.Стереологический анализ нейропильных нитей в формировании гиппокампа: взаимосвязь с нейрональной патологией и познанием при болезни Альцгеймера. Neuropathol Appl Neurobiol. 2007; 33: 334–43.
CAS PubMed Статья Google Scholar
Mundinano IC, Caballero MC, Ordonez C, Hernandez M, DiCaudo C, Marcilla I, Erro ME, Tunon MT, Luquin MR. Увеличение количества дофаминергических клеток и белковых агрегатов в обонятельной луковице пациентов с нейродегенеративными расстройствами.Acta Neuropathol. 2011; 122: 61–74.
CAS PubMed Статья Google Scholar
Wattendorf E, Welge-Lussen A, Fiedler K, Bilecen D, Wolfensberger M, Fuhr P, Hummel T., Westermann B. Обонятельные нарушения предсказывают атрофию мозга при болезни Паркинсона. J Neurosci. 2009. 29: 15410–3.
CAS PubMed PubMed Central Статья Google Scholar
Wu X, Yu C, Fan F, Zhang K, Zhu C, Wu T, Li K, Chan P. Корреляция между прогрессирующими изменениями грушевидной коры и обонятельной функцией при ранней болезни Паркинсона. Eur Neurol. 2011; 66: 98–105.
PubMed Статья Google Scholar
Ли Дж. Э., Чо К. Х., Хэм Дж. Х., Сон С. К., Сон Ю. Х., Ли PH. Обонятельные функции действуют как когнитивный резерв у пациентов с болезнью Паркинсона, не страдающих деменцией. Паркинсонизм, связанный с расстройством.2014; 20: 186–91.
PubMed Статья Google Scholar
Lee EY, Eslinger PJ, Du G, Kong L, Lewis MM, Huang X. Обонятельная кортикальная атрофия связана с обонятельной дисфункцией при болезни Паркинсона. Mov Disord. 2014; 29: 1205–8.
PubMed PubMed Central Статья Google Scholar
Ille R, Wabnegger A, Schwingenschuh P, Katschnig-Winter P, Kogl-Wallner M, Wenzel K, Schienle A.Роль предрасположенности к отвращению при болезни Паркинсона: морфометрическое исследование на основе вокселей. J Int Neuropsychol Soc. 2015; 21: 314–7.
PubMed PubMed Central Статья Google Scholar
Эриксен Н., Старк А.К., Паккенберг Б. Возраст и смерть нейронов, связанная с болезнью Паркинсона, в компактной части черной субстанции. J Neural Transm Suppl. 2009: 203–13.
Старк А.К., Паккенберг Б. Гистологические изменения дофаминергической нигростриатальной системы при старении.Cell Tissue Res. 2004; 318: 81–92.
CAS PubMed Статья Google Scholar
Зайдель К., Мальке Дж., Сисванто С., Крюгер Р., Хайнсен Х., Обургер Дж., Бузроу М., Гринберг Л. Т., Вихт Х., Корф Х. В. и др. Патологии ствола мозга при болезни Паркинсона и деменции с тельцами Леви. Brain Pathol. 2015; 25: 121–35.
PubMed Статья Google Scholar
Jellinger KA. Невропатология спорадической болезни Паркинсона: оценка и изменение представлений. Mov Disord. 2012; 27: 8–30.
CAS PubMed Статья Google Scholar
Лис А.Дж., Харди Дж., Ревес Т. Болезнь Паркинсона. Ланцет. 2009; 373: 2055–66.
CAS PubMed Статья Google Scholar
Iacono D, Geraci-Erck M, Rabin ML, Adler CH, Serrano G, Beach TG, Kurlan R.Болезнь Паркинсона и случайная болезнь тельцов Леви: вопрос времени? Неврология. 2015; 85: 1670–9.
PubMed PubMed Central Статья Google Scholar
Чу Ю., Бухман А.С., Оланов К.В., Кордовер Дж. Х. Есть ли у субъектов с минимальными двигательными особенностями продромальная болезнь Паркинсона? Энн Нейрол. 2018; 83: 562–74.
CAS PubMed PubMed Central Статья Google Scholar
Педерсен К.М., Марнер Л., Паккенберг Х., Паккенберг Б. Отсутствие глобальной потери неокортикальных нейронов при болезни Паркинсона: количественное стереологическое исследование. Mov Disord. 2005; 20: 164–71.
PubMed Статья Google Scholar
Хардинг А.Дж., Стимсон Э., Хендерсон Дж.М., Холлидей Г.М. Клинические корреляты избирательной патологии миндалины больных болезнью Паркинсона. Мозг. 2002; 125: 2431–45.
PubMed Статья Google Scholar
Флорес-Куадрадо А., Убеда-Банон I, Саис-Санчес Д., де ла Роса-Прието С., Мартинес-Маркос А. Альфа-синуклеин гиппокампа и интернейроны при болезни Паркинсона: данные на моделях человека и мыши. Mov Disord. 2016; 31: 979–88.
CAS PubMed Статья Google Scholar
Джоелвинг Ф.К., Биллесков Р., Кристенсен Дж. Р., Вест М., Паккенберг Б. Число нейронов гиппокампа и глиальных клеток при болезни Паркинсона — стереологическое исследование.Гиппокамп. 2006; 16: 826–33.
CAS PubMed Статья Google Scholar
Пирс Р.К., Хоукс С.Е., Дэниел С.Е. Переднее обонятельное ядро при болезни Паркинсона. Mov Disord. 1995; 10: 283–7.
CAS PubMed Статья Google Scholar
Huisman E, Uylings HB, Hoogland PV. Увеличение количества дофаминергических клеток в обонятельной луковице на 100% может объяснить гипосмию при болезни Паркинсона.Mov Disord. 2004; 19: 687–92.
PubMed Статья Google Scholar
Huisman E, Uylings HB, Hoogland PV. Гендерные изменения в увеличении количества дофаминергических нейронов в обонятельной луковице пациентов с болезнью Паркинсона. Mov Disord. 2008; 23: 1407–13.
PubMed Статья Google Scholar
Олендер Т., Кейдар И., Пинто Дж. М., Татарский П., Алкелай А., Чиен М. С., Фишилевич С., Рестрепо Д., Мацунами Н., Гилад Ю., Ланцет Д.Обонятельный транскриптом человека. BMC Genomics. 2016; 17: 619.
PubMed PubMed Central Статья CAS Google Scholar
Фернандес-Иригойен Дж., Корралес Ф. Дж., Сантамария Э. Протеомный атлас обонятельной луковицы человека. J Proteome. 2012; 75: 4005–16.
CAS Статья Google Scholar
Lachen-Montes M, Gonzalez-Morales A, Schvartz D, Zelaya MV, Ausin K, Fernandez-Irigoyen J, Sanchez JC, Santamaria E.Прототип обонятельной луковицы различается в зависимости от спектра лобно-височной деменции. J Proteome. 2019; 201: 37–47.
CAS Статья Google Scholar
Фернандес-Иригойен Дж., Сантамария Э. Обонятельное протеотипирование: к просветлению нейродегенерации. Neural Regen Res. 2019; 14: 979–81.
PubMed PubMed Central Статья Google Scholar
Паломино-Алонсо М., Лахен-Монтес М., Гонсалес-Моралес А, Осин К., Перес-Медиавилла А, Фернандес-Иригойен Дж., Сантамария Е. Сетевые протеогеномики выявляют связанный со старением дисбаланс в обонятельном комплексе IkappaBalpha-NFkappa функциональность в модели мышей с болезнью Альцгеймера Tg2576. Int J Mol Sci. 2017; 18.
Lachen-Montes M, Gonzalez-Morales A, Palomino M, Ausin K, Gomez-Ochoa M, Zelaya MV, Ferrer I, Perez-Mediavilla A, Fernandez-Irigoyen J, Santamaria E.Молекулярные нарушения с ранним началом в обонятельной луковице мышей Tg2576: новое понимание динамики чувствительной к стрессу обонятельной киназы при болезни Альцгеймера. Front Aging Neurosci. 2019; 11: 141.
CAS PubMed PubMed Central Статья Google Scholar
Селайя М.В., Перес-Вальдеррама Э., де Морентин XM, Тунон Т., Феррер I, Люкин М.Р., Фернандес-Иригойен Дж., Сантамария Э. Динамика протеома обонятельной луковицы во время прогрессирования спорадической болезни Альцгеймера: определение общих и различные обонятельные мишени для сопутствующих патологий, связанных с болезнью Альцгеймера.Oncotarget. 2015; 6: 39437–56.
PubMed PubMed Central Статья Google Scholar
Lachen-Montes M, Gonzalez-Morales A, Zelaya MV, Perez-Valderrama E, Ausin K, Ferrer I, Fernandez-Irigoyen J, Santamaria E. нарушение комплекса запрета во время прогрессирования болезни Альцгеймера. Научный доклад 2017; 7: 9115.
PubMed PubMed Central Статья CAS Google Scholar
Лахен-Монтес М., Селайя М.В., Сегура В., Фернандес-Иригойен Дж., Сантамария Э. Прогрессивная модуляция транскриптома обонятельной луковицы человека во время эволюции болезни Альцгеймера: новое понимание обонятельной передачи сигналов при протеинопатиях. Oncotarget. 2017; 8: 69663–79.
PubMed PubMed Central Статья Google Scholar
Lachen-Montes M, Gonzalez-Morales A, Iloro I, Elortza F, Ferrer I, Gveric D, Fernandez-Irigoyen J, Santamaria E.Выявление обонятельного протеостатического расстройства при болезни Паркинсона с помощью профилирования в масштабе всего протеома. Neurobiol Aging. 2019; 73: 123–34.
CAS PubMed Статья Google Scholar
Лахен-Монтес М., Гонсалес-Моралес А., Фернандес-Иригойен Дж., Сантамария Э. Внедрение безметки количественной обонятельной протеомики для выявления кандидатов в биомаркеры спинномозговой жидкости при синуклеинопатиях. Методы Мол биол. 2019; 2044: 273–89.
CAS PubMed Статья Google Scholar
Tapia-Rojas C, Cabezas-Opazo F, Deaton CA, Vergara EH, Johnson GVW, Quintanilla RA. Все дело в тау. Prog Neurobiol. 2019; 175: 54–76.
CAS PubMed Статья Google Scholar
Riederer P, Berg D, Casadei N, Cheng F, Classen J, Dresel C, Jost W., Kruger R, Muller T. Reichmann H и др.: Альфа-синуклеин при болезни Паркинсона: причинная или случайная причина? J Neural Transm (Вена).2019; 126: 815–40.
Артикул Google Scholar
Уокер LC. Прионоподобные механизмы при болезни Альцгеймера. Handb Clin Neurol. 2018; 153: 303–19.
PubMed PubMed Central Статья Google Scholar
Angot E, Steiner JA, Hansen C, Li JY, Brundin P. Являются ли синуклеинопатии прионоподобными расстройствами? Lancet Neurol. 2010; 9: 1128–38.
PubMed Статья Google Scholar
Даннинг CJ, Reyes JF, Steiner JA, Brundin P. Может ли патология болезни Паркинсона передаваться от одного нейрона к другому? Prog Neurobiol. 2012; 97: 205–19.
CAS PubMed Статья Google Scholar
Olanow CW, Брундин П. Болезнь Паркинсона и альфа-синуклеин: является ли болезнь Паркинсона прионоподобным заболеванием? Mov Disord. 2013; 28: 31–40.
CAS PubMed Статья Google Scholar
Джордж С., Рей Н.Л., Райхенбах Н., Штайнер Я. Брундин П: альфа-синуклеин: бегун на длинные дистанции. Brain Pathol. 2013; 23: 350–7.
CAS PubMed PubMed Central Статья Google Scholar
Prymaczok NC, Riek R, Gerez J. Болезнь Паркинсона распространяется не только на слухи. Front Hum Neurosci. 2016; 10: 608.
PubMed PubMed Central Статья Google Scholar
Макканн Х., Картрайт Х., Халлидей GM. Невропатология распространения альфа-синуклеина и гипотеза браака. Mov Disord. 2016; 31: 152–60.
CAS PubMed Статья Google Scholar
Steiner JA, Quansah E, Brundin P. Концепция альфа-синуклеина как прионоподобного белка: десять лет спустя. Cell Tissue Res. 2018; 373: 161–73.
CAS PubMed PubMed Central Статья Google Scholar
Karpowicz RJ Jr, Trojanowski JQ, Lee VM. Передача семян альфа-синуклеина при нейродегенеративном заболевании: последние разработки. Lab Investig. 2019; 99: 971–81.
PubMed Статья Google Scholar
Hansen C, Li JY. Помимо передачи альфа-синуклеина: распространение патологии при болезни Паркинсона. Тенденции Мол Мед. 2012; 18: 248–55.
CAS PubMed Статья Google Scholar
Макин С. Патология: прионный принцип. Природа. 2016; 538: S13–6.
CAS PubMed Статья Google Scholar
Брундин П., Мелки Р. Прайнг в прионную гипотезу болезни Паркинсона. J Neurosci. 2017; 37: 9808–18.
CAS PubMed PubMed Central Статья Google Scholar
Surmeier DJ, Obeso JA, Halliday GM. Болезнь Паркинсона — это не просто прионное заболевание.J Neurosci. 2017; 37: 9799–807.
CAS PubMed PubMed Central Статья Google Scholar
Браак Х., Дель Тредичи К., Братцке Х., Хамм-Клемент Дж., Сандманн-Кейл Д., Руб У. Определение стадии патологии внутримозговых телец включения, связанной с идиопатической болезнью Паркинсона (доклиническая и клиническая стадии). J Neurol. 2002; 249 (Suppl 3 III /): 1–5.
Артикул Google Scholar
Браак Х., Дель Тредичи К. Потенциальные пути распространения аномального тау- и альфа-синуклеина при спорадических заболеваниях Альцгеймера и Паркинсона. Cold Spring Harb Perspect Biol. 2016; 8.
Браак Х., Дель Тредичи К. Патогенез болезни Альцгеймера: существует ли распространение от нейрона к нейрону? Acta Neuropathol. 2011; 121: 589–95.
CAS PubMed Статья Google Scholar
Браак Х, Руб У, Гай В.П., Дель Тредичи К.Идиопатическая болезнь Паркинсона: возможные пути, по которым уязвимые типы нейронов могут подвергаться нейроинвазии неизвестным патогеном. J Neural Transm (Вена). 2003. 110: 517–36.
CAS Статья Google Scholar
Дель Тредичи К., Браак Х. Патология Леви и нейродегенерация при премоторной болезни Паркинсона. Mov Disord. 2012; 27: 597–607.
PubMed Статья Google Scholar
Doty RL. Гипотеза обонятельного вектора нейродегенеративного заболевания: жизнеспособна ли она? Энн Нейрол. 2008; 63: 7–15.
PubMed Статья Google Scholar
Хоукс Ч., Дель Тредичи К., Браак Х. Болезнь Паркинсона: гипотеза двойного удара. Neuropathol Appl Neurobiol. 2007. 33: 599–614.
CAS PubMed PubMed Central Статья Google Scholar
Hawkes CH, Del Tredici K, Braak H. Болезнь Паркинсона: пересмотр теории двойного удара. Ann N Y Acad Sci. 2009; 1170: 615–22.
PubMed Статья Google Scholar
Боргхаммер П. Как начинается болезнь Паркинсона? Перспективы нейроанатомических путей, прионов и гистологии. Mov Disord. 2018; 33: 48–57.
PubMed Статья Google Scholar
Lionnet A, Leclair-Visonneau L, Neunlist M, Murayama S, Takao M, Adler CH, Derkinderen P, Beach TG. Болезнь Паркинсона начинается в кишечнике? Acta Neuropathol. 2018; 135: 1–12.
PubMed Статья Google Scholar
Dujardin S, Lecolle K, Caillierez R, Begard S, Zommer N, Lachaud C, Carrier S, Dufour N, Auregan G, Winderickx J, et al. Перенос тау-белка дикого типа от нейрона к нейрону через транс-синаптический механизм: актуальность для спорадических таупатий.Acta Neuropathol Commun. 2014; 2: 14.
PubMed PubMed Central Статья Google Scholar
Вильяр-Пике А, Лопес-да-Фонсека Т., Сант’Анна Р., Сего Е.М., Фонсека-Орнелас Л., Пинхо Р., Кариха А., Герхардт Е., Масараккья С., Абад Гонсалес Е. и др. Экологические и генетические факторы поддерживают диссоциацию между агрегацией альфа-синуклеина и токсичностью. Proc Natl Acad Sci U S. A. 2016; 113: E6506–15.
CAS PubMed PubMed Central Статья Google Scholar
Cavaliere F, Cerf L, Dehay B, Ramos-Gonzalez P, De Giorgi F, Bourdenx M, Bessede A, Obeso JA, Matute C, Ichas F, Bezard E. Нейротоксичность альфа-синуклеина in vitro и распространение среди нейронов и астроцитов с использованием Экстракты тельцов Леви из головного мозга при болезни Паркинсона. Neurobiol Dis. 2017; 103: 101–12.
CAS PubMed Статья Google Scholar
Рей Н.Л., Джордж С., Штайнер Дж. А., Мададж З., Лук К. К., Трояновски Дж. К., Ли В. М., Брундин П.Распространение агрегатов после инъекции фибрилл альфа-синуклеина в обонятельную луковицу связано с ранней потерей нейронов и снижается в долгосрочной перспективе. Acta Neuropathol. 2018; 135: 65–83.
CAS PubMed Статья Google Scholar
Палоп Дж. Дж., Чин Дж., Маке Л. Перспектива сетевой дисфункции при нейродегенеративных заболеваниях. Природа. 2006; 443: 768–73.
CAS PubMed Статья Google Scholar
Микконен М., Алафузов И., Тапиола Т., Сойнинен Х., Миеттинен Р. Подполевые и послойные изменения иммунореактивности парвальбумина, кальретинина и кальбиндина-D28K в энторинальной коре при болезни Альцгеймера. Неврология. 1999; 92: 515–32.
CAS PubMed Статья Google Scholar
Saiz-Sanchez D, Ubeda-Banon I, Flores-Cuadrado A, Gonzalez-Rodriguez M, Villar-Conde S, Astillero-Lopez V, Martinez-Marcos A.Соматостатин, обоняние и нейродегенерация. Front Neurosci. 2020; 14: 96.
Спорнс О., Тонони Г., Коттер Р. Коннектом человека: структурное описание человеческого мозга. PLoS Comput Biol. 2005; 1: е42.
PubMed PubMed Central Статья CAS Google Scholar
Fox MD. Сопоставление симптомов с сетями мозга с помощью человеческого Коннектома. N Engl J Med. 2018; 379: 2237–45.
CAS PubMed Статья Google Scholar
Мирс Д., Поллард HB. Сетевая наука и человеческий мозг: использование теории графов для понимания работы мозга и одного из его узлов — миндалевидного тела — в здоровье и болезнях. J Neurosci Res. 2016; 94: 590–605.
CAS PubMed Статья Google Scholar
Crossley NA, Mechelli A, Scott J, Carletti F, Fox PT, McGuire P, Bullmore ET. Хабы коннектома человека обычно участвуют в анатомии заболеваний головного мозга.Мозг. 2014; 137: 2382–95.
PubMed PubMed Central Статья Google Scholar
Рис К.Л., Уиллер Д.В., Гамильтон Д.Д., Уайт С.М., Комендантов А.О., Асколи Г.А. Теоретико-графический и мотивный анализ потенциала коннектома нейрона гиппокампа. eNeuro. 2016; 3.
Jagust W. Визуализация эволюции и патофизиологии болезни Альцгеймера. Nat Rev Neurosci. 2018; 19: 687–700.
CAS PubMed PubMed Central Статья Google Scholar
Chen XQ, Mobley WC. Патогенез болезни Альцгеймера: выводы из исследований молекулярной и клеточной биологии олигомерных видов Abeta и tau. Front Neurosci. 2019; 13: 659.
PubMed PubMed Central Статья Google Scholar
Нельсон П.Т., Абнер Э.Л., Патель Э., Андерсон С., Уилкок Д.М., Крисцио Р.Дж., Ван Элдик Л.Дж., Джича Г.А., Гал З., Нельсон Р.С. и др. Миндалевидное тело как очаг патологической неправильной укладки при нейродегенеративных заболеваниях.J Neuropathol Exp Neurol. 2018; 77: 2–20.
CAS PubMed Статья Google Scholar
Браак Х., Дель Тредичи К. Невропатологическая стадия патологии головного мозга при спорадической болезни Паркинсона: отделение пшеницы от плевел. J Park Dis. 2017; 7: S71–85.
Google Scholar
Флорес-Куадрадо А., Убеда-Банон И., Саис-Санчес Д. Мартинес-Маркос а: альфа-синуклеинопатия миндалины человека при болезни Паркинсона: дифференциальная уязвимость нейронов, экспрессирующих соматостатин и парвальбумин.J Neuropathol Exp Neurol. 2017; 76: 754–8.
CAS PubMed Статья Google Scholar
Рей Н.Л., Вессон Д.В., Брундин П. Обонятельная луковица как место входа для прионоподобного распространения при нейродегенеративных заболеваниях. Neurobiol Dis. 2018; 109: 226–48.
CAS PubMed Статья Google Scholar
Болезнь Паркинсона связана с дегенерацией обонятельной системы
Первым признаком болезни Паркинсона часто является нарушение обоняния.Это нейродегенеративное заболевание в первую очередь вызывает непоправимое повреждение нервных клеток в области мозга, участвующей в управлении движением. До сих пор неясно, как это влияет на обонятельную систему. Исследователи провели исследование, в котором сравнивали обонятельные луковицы людей с болезнью Паркинсона и без нее. Исследователи обнаружили, что общий объем, занимаемый функциональными единицами обонятельной луковицы — так называемыми клубочками — при болезни Паркинсона составляет лишь половину от объема, который у нормальных людей.Более того, при болезни Паркинсона изменяется распределение клубочков в обонятельной луковице.
Девять из десяти пациентов с болезнью Паркинсона страдают дефектами обоняния на ранних стадиях болезни — часто за годы до появления моторных симптомов, характерных для этой болезни. Двигательные симптомы вызваны потерей нервных клеток в области черной субстанции головного мозга, которая отвечает за управление движением. Причины этой гибели клеток еще полностью не выяснены, но, по-видимому, ключевую роль играют тельца Леви.Это включения внутри клеток, которые содержат неправильно свернутую, дефектную версию белка альфа-синуклеина. Тельца Леви обнаруживаются в обонятельной луковице до того, как они появляются в черной субстанции.
Гипотеза так называемого обонятельного вектора болезни Паркинсона предполагает, что факторы окружающей среды, такие как вирусы, тяжелые металлы или пестициды, являются факторами риска или даже причинами этого состояния. Никакая другая сенсорная система, кроме обонятельной, не находится в таком тесном контакте с внешней средой — вдыхаемым воздухом.Гипотеза утверждает, что возбудитель болезни попадает из носовой полости в обонятельную луковицу, где запускается болезнь Паркинсона и постепенно распространяется через другие части мозга.
Обонятельная луковица человека остается малоизученной. Исследования этой структуры мозга в решающей степени зависят от наличия нетронутых образцов, которые обычно берутся после смерти у доноров мозга. Неустойчивое расположение обонятельной луковицы под основной частью головного мозга и множество аксонов, которые соединяют ее со слизистой оболочкой обоняния, означают, что при сборе образцов необходимо приложить особые усилия для защиты морфологии обонятельной луковицы.
Исследователям удалось собрать обонятельные луковицы, пригодные для углубленного количественного исследования. В рамках масштабного проекта исследователи химически обработали посмертные обонятельные луковицы, разрезали тонкие срезы размером десять микрометров по всей длине и окрашивали срезы флуоресцентно меченными антителами. Помеченные срезы были затем отсканированы, а изображения реконструированы в 3D, что позволило провести количественный анализ всей обонятельной луковицы.
Поскольку клубочки обонятельной луковицы человека трудно однозначно подсчитать, исследователи предложили новый количественный параметр: глобальный объем вокселей клубочков.Определив этот новый параметр, исследователи сравнили значения между обонятельными луковицами в нормальном состоянии и при болезни Паркинсона и обнаружили, что он снизился более чем наполовину. Еще предстоит выяснить, является ли снижение результатом того, что у случаев болезни Паркинсона меньше или меньше клубочков, или же является результатом комбинации этих двух эффектов.
Кроме того, было изменено распределение клубочков. Обонятельные луковицы нормальных пациентов имели 70 процентов клубочкового компонента в нижней половине обонятельной луковицы, но обонятельные луковицы пациентов с болезнью Паркинсона содержали только 44 процента в нижней половине.Ученые также обнаружили, что чем больше тельцов Леви с агрегированным альфа-синуклеином, тем меньше гломерулярный компонент обонятельной луковицы.
Теперь вопрос в том, какой тип нейронов обонятельной луковицы поражается в первую очередь или в первую очередь при болезни Паркинсона. Затем исследователи хотели бы определить наиболее уязвимые нейроны обонятельной луковицы.
Статья: «Вентральный гломерулярный дефицит при болезни Паркинсона, выявленный при реконструкции всей обонятельной луковицы»
Перепечатано из материалов, предоставленных Max-Planck-Gesellschaft.
Болезнь Паркинсона связана с дегенерацией обонятельной системы — ScienceDaily
Первым симптомом болезни Паркинсона часто является нарушение обоняния. Это нейродегенеративное заболевание в первую очередь вызывает непоправимое повреждение нервных клеток в области мозга, участвующей в управлении движением. Как это влияет на обонятельную систему, неясно. Исследователи из отделения нейрогенетики Макса Планка во Франкфурте и Оклендского университета в Новой Зеландии провели исследование, в котором сравнивали обонятельные луковицы людей с болезнью Паркинсона и без нее.Исследователи обнаружили, что общий объем, занимаемый функциональными единицами обонятельной луковицы — так называемыми клубочками — в случаях болезни Паркинсона составляет лишь половину от того, что у нормальных людей. Более того, при болезни Паркинсона изменяется распределение клубочков в обонятельной луковице.
Девять из десяти пациентов с болезнью Паркинсона страдают дефектами обоняния на ранних стадиях болезни — часто за годы до появления моторных симптомов, характерных для этого заболевания.Двигательные симптомы вызваны потерей нервных клеток в области черной субстанции головного мозга, которая отвечает за управление движением. Причины этой гибели клеток еще полностью не выяснены, но, по-видимому, ключевую роль играют тельца Леви. Это включения внутри клеток, которые содержат неправильно свернутую, дефектную версию белка альфа-синуклеина. Тельца Леви обнаруживаются в обонятельной луковице до того, как они появляются в черной субстанции.
Так называемая гипотеза обонятельного вектора болезни Паркинсона предполагает, что факторы окружающей среды, такие как вирусы, тяжелые металлы или пестициды, являются факторами риска или даже причинами этого состояния.Никакая другая сенсорная система, кроме обонятельной, не находится в таком тесном контакте с внешней средой — вдыхаемым воздухом. Гипотеза утверждает, что возбудитель болезни попадает из носовой полости в обонятельную луковицу, где запускается болезнь Паркинсона и постепенно распространяется по другим частям мозга.
Требуются образцы интактных тканей
Обонятельная луковица человека остается малоизученной. Исследования этой структуры мозга в решающей степени зависят от наличия нетронутых образцов, которые обычно берутся после смерти у доноров мозга.Неврологический фонд Новой Зеландии Douglas Human Brain Bank в Окленде, Новая Зеландия, тесно сотрудничает с семьями пациентов, страдающих нейродегенеративными заболеваниями, для обеспечения этичного и эффективного сбора посмертных образцов мозга больных и здоровых. Неустойчивое расположение обонятельной луковицы под основной частью головного мозга и множество аксонов, которые соединяют ее со слизистой оболочкой обоняния, означают, что при сборе образцов необходимо приложить особые усилия для защиты морфологии обонятельной луковицы.
Исследователи из Новой Зеландии смогли собрать обонятельные луковицы, пригодные для углубленного количественного исследования. В рамках масштабного проекта исследователи химически обработали посмертные обонятельные луковицы, разрезали тонкие срезы размером десять микрометров по всей длине и окрашивали срезы флуоресцентно меченными антителами. Помеченные срезы были затем отсканированы во Франкфурте, а изображения реконструированы в 3D, что позволило провести количественный анализ всей обонятельной луковицы.
Новый количественный параметр
Поскольку клубочки обонятельной луковицы человека трудно однозначно подсчитать, исследователи предложили новый количественный параметр: глобальный объем вокселей клубочков.Это количество является суммой объема всех клубочков. Они образуются путем слияния аксонов обонятельных сенсорных нейронов, образующих синапсы с нейронами обонятельной луковицы. Определив этот новый параметр, исследователи сравнили значения между обонятельными луковицами в нормальных условиях и при болезни Паркинсона и обнаружили, что он был уменьшен более чем наполовину. Еще неизвестно, является ли снижение результатом того, что у случаев болезни Паркинсона меньше или меньше клубочков, или же является результатом комбинации этих двух эффектов.
Кроме того, было изменено распределение клубочков. Обонятельные луковицы нормальных пациентов имели 70 процентов клубочкового компонента в нижней половине обонятельной луковицы, но обонятельные луковицы пациентов с болезнью Паркинсона содержали только 44 процента в нижней половине. «Преимущественный дефицит клубочкового компонента в нижней половине обонятельной луковицы, рядом с обонятельной слизистой оболочкой, согласуется с гипотезой обонятельного вектора болезни Паркинсона», — утверждает Питер Момбертс, M.Доктор философии, руководитель исследовательского отдела нейрогенетики Макса Планка. Ученые также обнаружили, что чем больше тельцов Леви с агрегированным альфа-синуклеином, тем меньше гломерулярный компонент обонятельной луковицы. «Эта взаимосвязь может быть признаком того, что тельца Леви являются причиной уменьшения объема клубочков», — объясняет д-р Болек Запец, первый автор статьи.
Теперь вопрос в том, какой тип нейронов обонятельной луковицы поражается в первую очередь или в первую очередь при болезни Паркинсона.Затем исследователи хотели бы определить наиболее уязвимые нейроны обонятельной луковицы.
История Источник:
Материалы предоставлены Max-Planck-Gesellschaft . Примечание. Содержимое можно редактировать по стилю и длине.
Сравнительное вскрытие периферической обонятельной системы переносчиков болезни Шагаса Rhodnius prolixus и Rhodnius brethesi
Abstract
Американский трипаносомоз, или болезнь Шагаса, передается как домашними, так и лесными видами Triatominae, которые используют сенсорные сигналы для определения местонахождения своих позвоночных-хозяев.Среди них пахучие вещества, как было показано, играют ключевую роль. Предыдущая работа выявила морфологические различия сенсорного аппарата разных видов Triatomines, но на сегодняшний день сравнительное функциональное исследование обонятельной системы отсутствует. После исследования сенсиллы антенн с помощью сканирующей электронной микроскопии (СЭМ) мы сравнили обонятельные реакции Rhodnius prolixus и лесного Rhodnius brethesi , используя электрофизиологический подход. На записях электроантеннограмм (EAG) мы впервые показали, что антенна R . prolixus очень чувствителен к карбоновым кислотам, соединениям, обнаруженным в их среде обитания и в свободном пространстве их позвоночных-хозяев. Затем мы сравнили ответы от обонятельных сенсорных нейронов (OSN), размещенных в желобчатых сенсиллах обоих видов, поскольку они настроены на эти соединения с использованием записей одиночной сенсиллы (SSR). В R . prolixus , ответы SSR выявили более узкое настраиваемое дыхание, чем его лесной собрат, причем последний показал ответы на более широкий диапазон химических классов.Кроме того, мы наблюдали значительные различия между этими двумя видами в их реакции на определенные летучие вещества, такие как амилацетат и бутирилхлорид. Таким образом, близкородственный, но экологически дифференцированный R . prolixus и R . brethesi демонстрируют явные различия в своих обонятельных функциях. Учитывая продолжающееся быстрое разрушение естественной среды обитания лесных видов и вероятный сдвиг в сторону среды, созданной людьми, мы ожидаем, что наши результаты будут способствовать разработке эффективных стратегий борьбы с переносчиками болезней в будущем.
Сведения об авторе
По оценкам, восемь миллионов человек во всем мире инфицированы американским трипаносомозом, также известным как болезнь Шагаса, возбудителем которого является паразит Trypanosoma cruzi . Переносчиками болезни являются более сотни видов насекомых, принадлежащих к подсемейству Tritatomine, так как они распространяют T . cruzi через их feaces. Несколько исследований подчеркнули важность обоняния для поведения этих насекомых в поисках хозяина, что позволяет им определять местонахождение своих позвоночных-хозяев и получать жизненно важную кровяную пищу.Стратегии борьбы с переносчиками инфекции являются наиболее эффективной политикой борьбы с распространением болезни Шагаса триатомовыми насекомыми. Однако недавние изменения в естественной среде обитания этих насекомых ставят под сомнение эффективность этих стратегий, поскольку виды, которые до сих пор считались исключительно лесными, теперь часто встречаются в перидоместных районах. В этом контексте понимание того, как триатомы с разным распределением обнаруживают запахи, чтобы определить местонахождение своих хозяев и выбрать среду обитания, очень важно.В этом исследовании мы сравниваем обонятельную систему широко распространенного Rhodnius prolixus и лесного родственника Rhodnius brethesi на морфологическом и функциональном уровне. Мы обнаружили, что обнаружение летучих веществ-хозяев и среды обитания имеет много общего, но также обнаруживает явные различия между видами.
Образец цитирования: Campetella F, Ignell R, Beutel R, Hansson BS, Sachse S (2021) Сравнительное рассечение периферической обонятельной системы переносчиков болезни Шагаса Rhodnius prolixus и Rhodnius brethesi .PLoS Negl Trop Dis 15 (4): e0009098. https://doi.org/10.1371/journal.pntd.0009098
Редактор: Алессандра Апаресида Гварнери, Фонд Освальдо Крус Институт Рене Рашу, БРАЗИЛИЯ
Поступила: 16 декабря 2020 г .; Принята к печати: 24 марта 2021 г .; Опубликовано: 15 апреля 2021 г.
Авторские права: © 2021 Campetella et al. Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Доступность данных: Все соответствующие данные находятся в рукописи и ее файлах с вспомогательной информацией.
Финансирование: Эта работа финансировалась Обществом Макса Планка для F.C., B.S.H., S.S. Спонсоры не играли никакой роли в дизайне исследования, сборе и анализе данных, решении опубликовать или подготовке рукописи.
Конкурирующие интересы: Авторы заявили, что никаких конкурирующих интересов не существует.
Введение
Болезнь Шагаса или американский трипаносомоз, вызванная инфекцией простейшими Trypanosoma cruzi , является хроническим заболеванием, эндемичным в 21 латиноамериканской стране, где она серьезно поражает наиболее уязвимые слои населения.По оценкам, его распространенность в некоторых регионах может достигать 5%, а его ежегодное бремя расходов на здравоохранение составляет до 600 миллионов долларов [1]. Уже в 1905 г. было показано, что кровососущие насекомые, принадлежащие к подсемейству Triatominae (Heteroptera: Reduviidae), передают T . cruzi через фекалии. На сегодняшний день наиболее эффективными и успешными методами борьбы с распространением болезни Шагаса являются меры борьбы с переносчиками инфекции. Широкое использование пестицидов и обучение местных жителей способам выявления и уничтожения насекомых являются наиболее эффективными стратегиями на сегодняшний день [2].Однако с появлением устойчивых к пестицидам насекомых срочно необходимы новые стратегии управления.
Triatominae — плохо определенная и, возможно, парафилетическая группа настоящих хищных насекомых семейства Reduviidae [3]. Все 151 описанный вид, филогенетически сгруппированный в пять триб [4], способны передавать болезнь Шагаса [5]. Исходя из этого, некоторые виды, такие как Rhodnius prolixus и Triatoma infestans , считаются особенно важными с эпидемиологической точки зрения, поскольку они широко распространены в Южной Америке и часто встречаются, хотя и не исключительно, в домицилированных районах [6–6]. 10].Однако большинство видов трибы Rhodniini, которым R . Принадлежит prolixus , считается, что он имеет более ограниченное распространение в лесных районах. На территориях, где они обитают, триатомы часто находят убежище в пальмах [11] и, в зависимости от количества видов пальм, в которых они гнездятся, их можно классифицировать как убежища широкого профиля или специалистов [12–14]. В то время как R . Известно, что prolixus относится к первому типу, интересным примером особого лесного вида является Rhodnius brethesi , который до сих пор был обнаружен только на пальме Leopoldina piassaba [13, 14].Несмотря на интересный характер этих ассоциаций, исследования лесных видов были маргинальными, и большая часть исследований была сосредоточена на домицилированных видах. Однако по мере роста обезлесения и изменения климата [12, 15] лесные виды теряют свои естественные среды обитания и могут найти убежище в домашних и перидоместных районах [16], подвергая их жителей более высокому риску и становясь проблемой общественного здравоохранения. Таким образом, для разработки более эффективных стратегий борьбы с переносчиками болезней необходимо глубокое понимание различий и сходств между видами триатомов, имеющих разные требования к среде обитания.
Будучи активными ночью, триатомины используют физические и химические сигналы, чтобы найти своего хозяина [17, 18]. Несколько исследований подчеркнули важность обоняния для поиска хозяина у этих насекомых [19–24]. Наземные позвоночные, основные хозяева этих обязательных кроветворных насекомых, испускают сигнатуры запаха, которые могут состоять из до 1000 различных летучих веществ [25–27], многие из которых вырабатываются микрофлорой кожи [28]. Предыдущая работа показала, что Т . infestans и R . prolixus используют некоторые из этих летучих веществ, чтобы найти своих хозяев [20, 21, 23, 29–33]. В частности, углекислый газ, 1-октен-3-ол, ацетон, несколько аминов, а также карбоновые кислоты являются привлекательными подсказками для R . prolixus [21, 34, 35], обнаружение которых достигается с помощью специализированных обонятельных сенсилл в антенне [31, 33, 36–39]. Однако исследований, сравнивающих обонятельные реакции между разными видами триатомов, нет.
У насекомых о различиях в настройке обоняния сообщалось между видами одного и того же рода, а также между дикими и домашними насекомыми одного и того же вида [40–44]. Предыдущие исследования триатомов показали, что количество обонятельных сенсилл коррелирует со сложностью и количеством экотипов, в которых встречаются насекомые [45, 46]. Например, домашние виды со стабильной средой имеют меньшее количество хемосенсорных сенсилл, чем их лесные родственники [45–49]. Кроме того, пониженная экспрессия обонятельных связывающих белков (OBP) и хемосенсорных белков (CSP) обнаружена у домицилированных Triatoma brasiliensis по сравнению с сильватическими и перидоместическими.[50]. В этом исследовании мы предположили, что морфология и настройка обонятельной системы R . prolixus и R . brethesi отражают различное распределение местообитаний и потребности двух видов. Чтобы проверить это, мы использовали сравнительный подход к характеристике периферической обонятельной системы широко распространенного универсального препарата R . prolixus и лесной специалист R . brethesi на анатомо-функциональном уровне.
Материалы и методы
Разведение насекомых
Насекомых выращивали, как описано ранее [51, 52]. Взрослые самцы р . brethesi и R . В экспериментах использовали prolixus , голодавшие в течение 3–4 недель. Партии насекомых содержались в индивидуальных ящиках с циклом Свет: Темнота, установленным на 12:12 часов. Ящики помещали в камеру при 25 ° C и относительной влажности 60%. Каждое насекомое использовалось в начале скотофазы, поскольку было показано, что острота обоняния выше в этот момент времени [20, 53].
Было показано, что выращивание в лабораторных условиях оказывает видоспецифическое влияние на количество и распределение обонятельных и механосенсорных сенсилл [49]. Однако, согласно предыдущей работе, в случае R . prolixus этот эффект либо отсутствует, либо является умеренным [54]. В то время как в R . brethesi наблюдалось увеличение плотности механосенсорных сенсилл (щетинок) и уменьшение количества трихоидных и базиконных сенсилл у исследованных в лаборатории насекомых по сравнению с дикими [48], не было возможности включить образцы этот вид с поля.
SEM
Головы насекомых, включая усики, фиксировали 2,5% (об. / Об.) Глутаровым альдегидом в какодилатном буфере (pH 7,4) в течение 60 мин. После этого образцы промывали трижды по 10 мин какодилатным буфером и дегидратировали при возрастающих концентрациях этанола (30%, 50%, 70%, 90% и 100%) в течение 10 минут каждый. Впоследствии образцы были высушены до критической точки с использованием жидкого диоксида углерода и напылены золотом (приблизительно 2 нм) с использованием устройства для нанесения покрытия распылением SCD005 (BAL-TEC, Balzers, Лихтенштейн).Наконец, соответствующие поверхности были проанализированы с помощью сканирующего электронного микроскопа (SEM) LEO-1450 (Carl Zeiss NTS GmbH, Оберкохен, Германия), обеспечивающего вращающийся предметный столик для получения всестороннего изображения.
Запахи
Запахи были получены от Sigma-Aldrich, FLUKA, Aldrich с наивысшей доступной чистотой. Используемые соединения перечислены в таблицах S2 и S3 вместе с соответствующим используемым растворителем (парафиновое масло, CAS: 8012-95-1, Supelco, США; дистиллированная вода; или этанол, Sigma Aldrich, Германия), в котором каждый запах был разбавлен. .Для записи электроантеннограмм (EAG) использовалось 10% -ное разведение в парафиновом масле (Supelco, США), в то время как мы применяли все запахи в разбавлении 1% для одиночных записей сенсиллы (SSR). В записях EAG использовался углекислый газ сертифицированного качества, разбавленный воздухом до конечной концентрации 30%. Смесь запаха, используемая только в SSR, была создана путем смешивания всех соединений, перечисленных в таблице S3, в соотношении 1: 1, все при 1% разбавлении в парафиновом масле. Соединения в этой смеси, как известно, обнаруживаются пахучими рецепторами (OR) у других видов насекомых, и поэтому были разработаны для идентификации возможных OR, содержащихся в желобчатых сенсиллах штифта Rhodnius spp [55–58].
Применение запаха
Запахи, используемые в качестве стимулов, готовили в начале каждого экспериментального сеанса: аликвоту 10 мкл разбавленного запаха (см. Таблицы S2 и S3) пипеткой наносили на свежую фильтровальную бумагу (Ø = 1 см. 2 , Whatman, Dassel, Германия), который помещали внутрь стеклянной пипетки Пастера. Каждую загруженную фильтровальную бумагу использовали максимум 3 раза, чтобы обеспечить стабильную концентрацию во всех экспериментах. Легколетучие карбоновые кислоты и альдегиды загружались при каждом предъявлении стимула.Углекислый газ разбавляли незадолго до применения, и для его доставки использовался изготовленный на заказ шприц, который был подключен к автоматическому контроллеру стимулов, как описано ранее [59]. Контроллер стимула (Stimulus Controller CS-550.5, Syntech, Германия) использовали для доставки запахов к антенне насекомого через металлическую пипетку, расположенную на расстоянии менее 1,5 см (EAG) или 0,5 см (SSR) от антенны насекомого. Постоянный поток увлажненного воздуха в 1,0 л / мин -1 подавался к насекомому, в то время как каждый импульс запаха имел поток воздуха 0.5 л мин -1 , и был забуферен компенсирующим потоком воздуха той же величины.
Записи электроантеннограмм (EAG)
Антенну быстро перерезали между скапусом и ножкой и поместили между двумя металлическими электродами. На каждый конец антенны наносили проводящий гель (Spectra 360, Parker Laboratories, Fairfield, США). Электрод был подключен к аналого-цифровому преобразователю Syntech IDAC (Syntech). Регистрация проводилась с помощью Autospike32 с частотой дискретизации 2400 Гц.Хотя применение запахов было случайным, мы гарантировали, что контроль (парафиновое масло) будет применяться через регулярные промежутки времени. Во время проверки панели запаха мы наблюдали увеличение амплитуды отклика на контроль в зависимости от времени. Чтобы учесть это смещение, мы нормализовали каждую запись по следующей формуле, аналогично тому, как это делалось ранее [60]: с участием, где A n ( t ) — нормализованная реакция на заданный запаховый стимул n в данный момент времени t ; Z n — измеренная реакция на раздражитель запаха n , а C (t) — усредненная реакция растворителя в данный момент времени t , где — ближайшая реакция растворителя перед предъявлением стимула при t a и b ближайший отклик растворителя после предъявления стимула при t b .Вклад каждого из этих откликов на растворитель в усредненный отклик на растворитель учитывается с помощью коэффициента, где T = t b -t a .
Записи одиночной сенсиллы (SSR)
Насекомых помещали внутрь отрезанного пластикового наконечника объемом 5 мл (Eppendorf, Гамбург, Германия), который запечатывали стоматологическим воском (Erkodent, Pfalzgrafenweiler, Германия). Затем наконечник иммобилизовали на предметном стекле для микроскопии стоматологическим воском. Обе антенны приклеивали к покровному стеклу двусторонним скотчем.В качестве эталона использовали вольфрамовый электрод, вставленный в брюшную полость насекомого. Предварительные записи с серебряной проволокой в качестве электрода сравнения не показали улучшения отношения сигнал / шум. Препарат помещали под прямой микроскоп (BX51WI, Olympus, Гамбург, Германия), оснащенный воздушным объективом 50x (LMPlanFI 50x / 0,5, Olympus). Нейронная активность в виде цепочек спайков, исходящих от OSN, регистрировалась с помощью заостренного вольфрамового электрода, нацеленного на основание сенсиллы штифта с бороздками.Сигналы усиливали (Syntech Universal AC / DC Probe; Syntech) и отбирали 10 666,7 отсчетов с -1 через USB-IDAC (Syntech), подключенный к компьютеру. Спайки были извлечены с помощью программного обеспечения Autospike32 (Syntech). Ответы на запах от каждой сенсиллы рассчитывали как разницу в количестве импульсов за 0,5 с до и после появления стимула.
Ответ на каждый запах в записях SSR был рассчитан как изменение спайков s -1 при стимуляции запаха с использованием Autospike32.Отклик на растворитель вычитали из каждого измерения. Количество OSN, размещенных в каждой сенсилле в R . prolixus оценивается между 5 и 6 [29, 61]. Мы попытались подтвердить это наблюдение, используя полутонкий срез антенны, но, несмотря на наши усилия, не смогли однозначно определить количество сенсорных нейронов в сенсилле с бороздками (GP) у любого из двух видов. По этой причине мы решили определить каждую сенсиллу как отвечающую единицу, как это было сделано у других насекомых [62].
Последующий анализ был проведен в MATLAB (The MathWorks Inc, Натик, США), в котором была проведена агломеративная иерархическая кластеризация ответов сенсиллы с помощью евклидовой метрики и метода Уорда. Коэффициент несогласованности был рассчитан для каждого звена дендрограммы, как способ определения встречающихся в природе кластеров в данных [63, 64]. Глубина 4 и коэффициент отсечки 1,8 для R . prolixus и 1.0 для R . В расчетах было использовано бретелек .
Ответ каждого типа сенсиллы был принят как средний ответ отдельных сенсилл, принадлежащих к одному и тому же кластеру (т.е. тип сенсиллы). Затем средние ответы каждого типа сенсиллы были сгруппированы и усреднены по химическим классам. Затем эти ответы были нормализованы до максимального ответа в пределах каждого типа сенсиллы. Анализ главных компонентов (PCA, с алгоритмом разложения по сингулярным значениям (SVD)) был выполнен в MATLAB (The MathWorks Inc) с использованием усредненных масштабированных (от 0 до 1) и нормированных по z-баллу ответов для каждого типа сенсиллы и каждого вида.В качестве меры сходства мы применили односторонний ANOSIM, чтобы вычислить, представляют ли различные типы сенсиллы существенно разные классы [65].
Усредненные ответы были рассчитаны как среднее значение всех ответов сенсиллы на конкретный одорант. Для сравнения между химическими классами эти запахи были затем усреднены для каждого конкретного химического класса. Сравнение между видами проводили с использованием непарных двусторонних t-критериев Стьюдента (GraphPad Prism 8, Сан-Диего, США). Затем эти средние отклики были нормализованы к запаху, который вызывал самые высокие отклики у каждого вида, являющегося пропионовой кислотой для обоих видов, и вычисляли разреженность в течение жизни (S) каждой сенсиллы.Мы применили разреженность в течение жизни как меру ширины отклика каждой сенсиллы. Для его расчета использовалась следующая формула [66]: где S — разреженность в течение срока службы, N — количество испытанных запахов, а r j — реакция сенсиллы на любой заданный запах j , при этом r j ≥0 и S ∈ [ 0,1], где S = 0 соответствует случаю, в котором сенсилла одинаково реагирует на все пахучие вещества, а S = 1 соответствует случаю, когда сенсилла реагирует только на один запах из набора.
Результаты
Видовые морфологические различия антенны
Предыдущие исследования продемонстрировали, что паттерны сенсиллы у кроветворных насекомых, включая виды Triatominae, отражают специфические адаптации к разным хозяевам и средам обитания [67–70]. Путем сравнительного качественного и количественного анализа основного органа обоняния, антенны, мы оценили возможные морфологические различия между универсальным R . prolixus и лесной специалист R . brethesi , используя сканирующую электронную микроскопию (SEM) (рис. 1, S1 и S2). Качественный анализ морфологического рисунка сенсилл на антеннах обоих видов не выявил существенных различий. Для дальнейшего анализа сенсиллы были классифицированы в соответствии с исследованием Shanbhag et al. [71], как это было сделано ранее в триатомах [30, 33]. У обоих видов было обнаружено, что второй сегмент, или ножка, обогащен сенсиллами, которые, как описано, выполняют механосенсорную функцию [38-40]: сенсиллы трихоботриум, щетинки I и III (S1 и S2 Figs).Кроме того, в R показано, что пещерный орган сенсиллы имеет терморецептивную функцию. prolixus и T . infestans [72, 73] также был обнаружен в R . brethesi (рис. S2B). Однако идентифицировать ранее описанную орнаментированную пору [38] в R не удалось. brethesi , возможно, из-за угла ориентации нашего препарата. Примечательно, что в одном препарате наши микрофотографии показывают наличие двух ранее не описанных типов сенсиллы на ножке R . проликс . Один из них представляет собой сенсиллу, вставленную в ямку, с негибкой розеткой, без явных пор, размещенную внутри камеры в кутикуле усиков (S1B Рис.). Этот тип напоминает термочувствительную sensilla coeloconica [42], но его функция остается неизвестной. Второй тип сенсиллы напоминает целоконическую сенсиллу 3-го типа, характеризующуюся двумя порами в основании, описанную у других видов гемиптеранов (S1E Fig) [45].
Рис. 1. Морфология периферической обонятельной системы Rhodnius prolixus и Rhodnius brethesi .
(A, B) Световые микроскопические изображения всего тела R . prolixus и R . brethesi (левая панель). Указаны сегменты усиков: скапус (и), ножка (p), жгутик I (fI) и жгутик II (fII). Изображения головы (средняя панель) и соответствующих сегментов антенн (правая панель), полученные с помощью сканирующей электронной микроскопии (СЭМ). (C, D) Антенная сенсилла R . prolixus и R . brethesi на жгутике II: стрелками показаны (I) толстостенные (Tk) и тонкостенные (Th) трихоидные сенсиллы (также известные как трихоидные и базиконические сенсиллы) и щетинки (Br).(II) Толстостенная трихоидная сенсилла, (III) колокольчатая сенсилла и (IV) бороздчатая сенсилла штифта.
https://doi.org/10.1371/journal.pntd.0009098.g001
Стеночно-пористые сенсиллы с негибкими гнездами обнаружены на жгутике I и дистальной половине жгутика II у обоих видов. К ним относятся трихоидная и базиконная сенсиллы (также известные как толстостенные и тонкостенные трихоидные сенсиллы [74] соответственно), а также двустенные бороздчатые сенсиллы штифта (называемые базиконными сенсиллами в [74]) (рис. 1C). и 1D).Все эти типы сенсиллы имеют небольшие продольные бороздки, заполненные порами или щелями, указывающими на обонятельную функцию [30]. На одних и тех же сегментах усиков обоих видов мы также идентифицировали сенсиллу без пор с негибкой розеткой: колоколообразную сенсиллу [74, 75] (Рис. 1C и 1D).
Как показано в других исследованиях, количественные различия в количестве обонятельных сенсилл у видов потенциально отражают особые приспособления к их экологической нише. Таким образом, мы провели конфокальное сканирование автофлуоресценции антенн, залитых глицерином, чтобы количественно оценить плотность сенсиллы на флагелломере II, где предполагаемые обонятельные сенсиллы достигают наивысшей плотности, и ранее сообщалось о межвидовых различиях между триатомами [45] (рис. 2). .Наши результаты показывают, что плотность как базиконических, так и трихоидных сенсилл значительно выше у R . brethesi , чем в R . проликс . Напротив, плотность желобчатых сенсилл штифтов существенно не различалась между двумя видами.
Рис. 2. Плотность сенсиллы у Rhodnius prolixus и Rhodnius brethesi .
Плотность обонятельных сенсилл в жгутиках II для каждого вида оценивается как общее количество сенсилл для каждого типа по площади поверхности жгутиков, n = 3, непарный t-критерий: ** p <0.01. Данные представляют собой среднее значение ± стандартная ошибка среднего.
https://doi.org/10.1371/journal.pntd.0009098.g002
Ответы антенн у
Rhodnius prolixusПредыдущие исследования, нацеленные на определение реакции триатомов на запахи, были сосредоточены на T . infestans и сообщили о реакции на небольшое количество химических соединений, включая альдегиды, кислоты и амины [30, 32, 36]. Чтобы оценить, обнаруживаются ли антеннами триатомов другие химические классы, мы выполнили записи EAG в R . prolixus с использованием панели из 27 запахов, принадлежащих к различным химическим классам, которые, как ранее было показано, вызывают поведенческие реакции у триатоминов и других гематофагов (рис. 3 и таблицы S1 и S2). Значимые ответы (p <0,05) наблюдались для 33% испытанных запахов (t-критерий одного образца против нуля). Из них наиболее сильная реакция наблюдалась на уксусную кислоту, соединение, которое присутствует в триатомовых фекалиях и опосредует агрегацию [76], а затем на пропионовую кислоту, известный летучий хозяин, к которому относится T . infestans поведенчески активен [33]. Значительные реакции были также замечены на основной компонент феромона тревоги [77], изомасляную кислоту, соединение, которое также присутствует в летучих веществах хозяина [27], и на близкородственное соединение масляной кислоты. Взятые вместе, ответы на кислоты составили 44% от общего числа значимых ответов. Дополнительно R . prolixus показал значительную, хотя и меньшую, реакцию на другие летучие вещества хозяина, такие как циклогексанон, амилацетат и триметиламин.
Рис. 3. Антенные ответы от Rhodnius prolixus .
Экологически значимые одоранты (см. Таблицу S1) наносили на антенну насекомых, а вызванные реакции измеряли с помощью анализа электроантеннограмм. Ответы были нормализованы к их соответствующему отклику на растворитель (см. Методы). Закрашенные столбцы представляют собой статистически значимые ответы (t-критерий для одного образца против нуля: p <0,05, n = 7–10 для каждого испытанного запаха).
https: // doi.org / 10.1371 / journal.pntd.0009098.g003
Значительный ответ был зарегистрирован для 30% углекислого газа, хемосенсорного сигнала, который является привлекательным для T . infestans при более низких концентрациях [20]. Интересно, что мы также наблюдали значительную обонятельную реакцию на бутирилхлорид. Хотя предполагается, что это соединение действует как репеллент от насекомых, поскольку оно ингибирует активность сенсорных нейронов, обнаруживающих углекислый газ, у комаров [60], его функция и обнаружение у триатоминов до сих пор не изучены.
Реакция на запах в сенсилле с рифленым стержнем
Записи EAG R . prolixus продемонстрировал, что обонятельная система этих насекомых в основном реагирует на кислоты и амины. Эти соединения обычно встречаются в среде насекомых (таблица S1), и их роль в регулировании поведения, обусловленного запахом, оценивалась для некоторых видов триатоминов [19, 21, 22, 54]. У насекомых кислоты и амины обнаруживаются нейронами, размещенными в сенсиллах усиков с бороздками (GP) [78–80].Как показали наши морфологические исследования, этот тип сенсиллы присутствует в антеннах обоих видов Rhodnius с низкой плотностью, что делает его идеальной системой для оценки видоспецифичных различий в настройке обоняния.
Чтобы оценить настройку отдельных сенсилл GP, определяемых здесь как отвечающая единица, мы протестировали в общей сложности 38 запахов, из которых 17 были кислотами и 9 аминами, различающимися по длине углерода и разветвлению. Мы включили дополнительные летучие вещества (такие как индол и амилацетат), которые, как известно, присутствуют, но не исключительно, у позвоночных-хозяев или, как было ранее показано, обнаруживаются сенсиллами GP у других насекомых [25, 36].Кроме того, применялась специальная смесь OR (таблица S3), состоящая из соединений, обычно обнаруживаемых пахучими рецепторами (OR) у других видов [55–58]. Усредненные ответы сенсиллы показали, что R . brethesi обычно сильнее реагирует на запахи, чем R . prolixus (рис. 4А). Кроме того, значительная общая межвидовая разница была обнаружена для 58% одорантов. В то время как оба вида проявляли наиболее сильную реакцию на пропионовую кислоту, основные различия наблюдались для следующих соединений: масляная кислота, бензальдегид, валериановая кислота, 2-оксопропановая кислота, муравьиная кислота и для смеси OR с R . brethesi показывает более высокий отклик, чем R . prolixus во всех случаях. Бутирилхлорид был единственным соединением со значительно более высокой чувствительностью в R . проликс . Стимуляция пальмитиновой кислотой вызывала сильнейший тормозящий ответ у обоих видов. Когда ответы были нормализованы до максимальной реакции запаха (то есть пропионовой кислоты у обоих видов), значимые различия остались для пяти одорантов: бутиральдегид, бутирилхлорид, амилацетат, 3-метилиндол и для смеси OR.
Рис. 4. Ответы сенсиллы с бороздками на экологически значимые летучие вещества у Rhodnius prolixus и Rhodnius brethesi .
(A) Усредненная реакция запаха сенсиллы с рифленым стержнем из R . prolixus и R . человек . Ответы были нормализованы до максимального зарегистрированного ответа, которым была пропионовая кислота для обоих видов. (B) Круговые диаграммы представляют собой усредненные ответы согласно химическому классу для обоих видов.(C) Кривые настройки для каждого вида. По краям располагаются запахи, вызывающие самые слабые реакции. Порядок запахов у разных видов разный. Редкость продолжительности жизни ( S ) была рассчитана для каждого вида (см. Методы).
https://doi.org/10.1371/journal.pntd.0009098.g004
Rhodnius prolixus в среднем чаще реагировал на амины, чем R . brethesi составило 40% ответов в R . prolixus по сравнению с 24% в R . brethesi (рис. 4В). Напротив, R . brethesi сильнее реагировал на альдегиды, с 33% по сравнению с 16% в R . проликс . Ответы на кислоты в пределах каждого вида были сопоставимы и составляли 18% в R . prolixus и 22% в R . человек . Аналогичные результаты были получены для смешанной химической категории ( i . e ., «Other») с 18% в R . prolixus и 15% в R . человек . Наконец, усредненные ответы R . brethesi к сложным эфирам были немного выше, чем найденные для R . проликсус (от 14% до 8%).
Настройка отклика рифленого штифта sensilla
Для того, чтобы количественно оценить и сравнить ширину настройки сенсилл GP между обоими видами, мы построили кривые настройки для конкретных видов и определили разреженность продолжительности жизни (S) (рис. 4C).Редкость продолжительности жизни обычно рассчитывается для оценки того, насколько широко или узко настроены обонятельные рецепторы, в то время как в нашем случае это служит мерой настройки GP-сенсиллы. Этот анализ показывает, что R . prolixus действительно настроен на более узкий набор запахов, чем R . brethesi со значением S 0,5 для R . prolixus по сравнению с 0,35 в R . человек .
Затем мы задались вопросом, наблюдаются ли более сильные отклики для R . brethesi является результатом более высокой доли отдельных сенсилл, демонстрирующих ответы, вызванные возбуждающим запахом, или, что менее вероятно, снижение тормозных ответов сенсиллы в R . brethesi по сравнению с R . проликс . Таким образом, чтобы дополнительно охарактеризовать эти ответы, мы проанализировали отдельные комбинации запаха и сенсиллы. Поскольку каждая сенсилла GP была проверена с помощью комплексной панели запахов, состоящей в общей сложности из 38 запахов, наши данные SSR включали 950 комбинаций запаха и сенсиллы в R . prolixus и 380 комбинаций запаха и сенсиллы в R . человек . Пока в R . prolixus , только 31% этих комбинаций запах-сенсилла давали ответы> 15 спайков с — 1 выше реакции на растворитель, 60% — в R . brethesi (рис. 5). Это различие также соответствовало ответам с более высокими частотами пиков (> 50 пиков с -1 ): в R . prolixus , только 7% этих комбинаций давали ответы выше, чем 50 пиков на -1 над растворителем, в то время как в R . brethesi 26% комбинаций запаха и сенсиллы приводили к ответам> 50 спайков s -1 . Ответы выше 100 шипов s -1 были, как правило, редкими у обоих видов.
Рис. 5. Характеристики отклика сенсиллы рифленого стержня.
Процент комбинаций запаха и сенсиллы, активированных при указанной скорости срабатывания каждого одоранта. Общее количество комбинаций запаха и сенсиллы составило 950 для R . prolixus и 380 для R . человек . Одоранты отсортированы по оси абсцисс в соответствии с количеством сенсилл, которые они активируют. Ответы с частотой ниже -15 пиков / с считаются тормозящими.
https://doi.org/10.1371/journal.pntd.0009098.g005
Тормозные реакции были менее распространены, чем возбуждающие, и только 5% комбинаций запах-сенсилла были идентифицированы как тормозящие (<-15 спайков s -1 по сравнению с контролем с растворителем), в обоих R . prolixus и R . человек . Ингибирование нельзя было отнести к одному одоранту, поскольку 53% запахов в панели вызывали по меньшей мере одно подавление запаха-сенсиллы в R . prolixus , и 32% запахов приводили к ингибированию R . человек . Взятые вместе, наши данные показывают, что более сильные ответы наблюдались у в R . brethesi можно отнести к более высокой доле ответов, превышающей 15 шипов s -1 , а не к различию в ингибирующих ответах между этими видами.
Функциональная классификация сенсиллы штифта с рифлением
Для дальнейшего отнесения измеренных запаховых реакций к отдельным и функциональным подтипам сенсиллы GP в каждом из двух видов мы выполнили агломеративный иерархический кластерный анализ (рис. 6). Ответы могут быть сгруппированы в 4 группы у каждого вида, соответствующие предполагаемым функциональным типам сенсиллы, классифицированным как GP1-GP4. Следует отметить, что у обоих видов сенсиллы всех типов реагировали на масляную кислоту, а также на пропионовую кислоту.В частности, сильные ответы (т.е.> 50 спайков s -1 ) на кислоты были более выраженными в R . brethesi , причем все типы сенсиллы реагируют как минимум на 7 из 17 протестированных кислотных соединений. Основное различие между видами заключалось в реакции на нашу индивидуальную комбинацию ИЛИ. Пока на него откликнулась только одна сенсилла в R . prolixus (с> 50 шипами s -1 ), 50% сенсилл показали ответ на смесь R . человек .
Рис. 6. Функциональная классификация сенсилл рифленого штифта у Rhodnius prolixus и Rhodnius brethesi .
Цветовые ответы сенсиллы штифта с бороздками (флагелломера II) в R . prolixus (n = 25) и R . brethesi (n = 10) к панели запаха. В каждом случае дендрограмма представляет собой агломеративную иерархическую кластеризацию (метод Уорда, Евклидово расстояние) этих ответов.
https://doi.org/10.1371/journal.pntd.0009098.g006
Поскольку каждый из четырех предполагаемых типов сенсиллы реагировал на определенную комбинацию запахов (рис.6), мы предлагаем их в качестве диагностических запахов для каждого конкретного типа GP. (Рис. 7A). В R . prolixus , GP типа 1 (Rp-GP1), на долю которого приходится 40% зарегистрированных GP-сенсилл, реагирует преимущественно на амины триметиламин, аммиак и этиламин, о чем свидетельствуют средние ответы.Rp-GP2 содержит 16% GP-сенсиллы и лучше всего реагирует на пропионовую кислоту, триэтиламин, спермин, спермидин и бензальдегид. Rp-GP3 показывает самые высокие реакции на изоамиламин и бутирилхлорид и составляет 28% GP-сенсиллы, в то время как Rp-GP4, представляющий 16% сенсиллы, реагирует на аммиак, этиламин и бутирилхлорид.
Рис. 7. Профиль ответа функциональных подтипов сенсилл рифленого штифта.
(A) Цветовая кодировка средних ответов на выбранные диагностические запахи для каждого типа сенсиллы для каждого вида насекомых.Запахи были выбраны в качестве диагностических, когда они позволяли максимально разделять типы сенсилл. (B) Усредненные ответы для каждого химического класса и типа сенсиллы (левая панель), нормированные на максимальную реакцию внутри каждого типа сенсиллы (правая панель). (C) Усредненные ответы (n = 2–6) на кислоты и амины с указанной длиной углеродной цепи. В R . prolixus , GP1, GP3, GP4 показывают отрицательную корреляцию между длиной углерода и силой реакции на кислоты (корреляция Пирсона; Rp-GP1: r = -0.87, р = 0,0005; Rp-GP2: r = -0,59, p = 0,054; Rp-GP3: r = -0,75, p = 0,008; Rp-GP4: -0,61, p = 0,045, n = 11). В R . brethesi GP1 и GP2 показывают значительную отрицательную корреляцию между длиной углерода и силой реакции на кислоты (корреляция Пирсона; Rb-GP1: r = -0,89, p = 0,0002; Rb-GP2: r = -0,76, p = 0,006, n = 11). (D) Анализ главных компонентов (PCA) R . prolixus и R . brethesi типов сенсиллы. Для сенсиллярного пространства двух видов существенных различий не наблюдалось (ANOSIM, p = 0.32, Евклидова).
https://doi.org/10.1371/journal.pntd.0009098.g007
В R . brethesi , GP-сенсилла 1 типа (Rb-GP1) реагировала преимущественно на масляную кислоту и ингибировалась амилацетатом (фиг. 7A). Rb-GP2 с аналогичным профилем ответа на Rb-GP1 отличался от него ответами на амилацетат и 2-оксопропановую кислоту. Он также показал более высокую реакцию на изоамилацетат, масляную, валериановую и муравьиную кислоты, чем Rb-GP1. Тип Rb-GP3 показал высокие отклики на 2-оксопропановую кислоту и муравьиную кислоту.Наконец, введите GP4 из R . brethesi показал сильную реакцию на бензальдегид, аммиак и пропионовый альдегид. Rb-GP1 составлял 30%, Rb-GP2 20%, Rb-GP3 20% и Rb-GP4 30% от общего числа бороздчатых сенсилл штифтов, зарегистрированных у этого вида.
Настройка запаха на химические классы
Затем мы проанализировали, реагируют ли отдельные типы сенсиллы преимущественно на определенные химические классы (рис. 7B). В R . prolixus , Rp-GP1 сильнее всего реагировал на амины, Rp-GP2 — на альдегиды и в меньшей степени — на амины.Rp-GP3 не реагировал преимущественно на какой-либо класс химического запаха, при этом большинство ответов приходилось на бутирилхлорид, а Rp-GP4 демонстрировал самые сильные ответы на амины. В R . brethesi , все типы сенсиллы отвечали, по крайней мере, двум из протестированных химических классов. В то время как и Rb-GP1, и Rb-GP3 показали наиболее сильную реакцию на кислоты, Rb-GP3, но не Rb-GP1, дополнительно отреагировал на альдегиды. Rb-GP2 не реагировал на какой-либо конкретный класс запаха, причем его самые высокие ответы были показаны на смесь OR.Наконец, Rb-GP4 реагировал главным образом на альдегиды, а затем на амины.
Затем мы оценили, вызывают ли соединения запаха с определенной длиной углерода более сильные реакции у сенсиллы Rhodnius с бороздками, сосредоточив внимание на C1-C18 кислот и аминов (рис. 7C). В R . prolixus , мы наблюдали более высокие ответы для короткоцепочечных карбоновых кислот (C1-6 / 7), причем три типа сенсиллы показали отрицательную корреляцию между длиной углеродной цепи и силой ответа (корреляция Пирсона; Rp-GP1: r = -0 .87, р = 0,0005; Rp-GP2: r = -0,59, p = 0,054; Rp-GP3: r = -0,75, p = 0,008; Rp-GP4: -0,61, p = 0,045, n = 11). Что интересно, GP2 R . prolixus показал более слабые ответы на амины с короткой цепью, но более сильные ответы на амины с длинными цепями (C6-C10). Длина кислотной углеродной цепи также оказалась актуальной для R . brethesi , где он отрицательно коррелировал с интенсивностью ответа у 2 из 4 типов сенсиллы (корреляция Пирсона; Rb-GP1: r = -0.89, р = 0,0002; Rb-GP2: r = -0,76, p = 0,006, n = 11). Напротив, в случае аминов снижение активности с увеличением длины углерода наблюдалось в GP4 (корреляция Пирсона; r = -0,85; p = 0,016, n = 7) в R . prolixus , но не в других типах сенсиллы GP R . человек . Однако если сравнивать с R . prolixus , R . brethesi показал более сильные ответы на короткоцепочечные (C1-C5) амины ( R . проликсус : 19,63 ± 2,65, n = 125; Р . brethesi : 19,63 ± 2,65, n = 51; непарный t-критерий, p = 0,0005).
Наконец, мы обратились к сопоставимости описанных функциональных типов сенсилл между видами. Чтобы получить представление о сходстве между описанными типами GP, мы рассчитали евклидовы расстояния между типами сенсилл для двух видов (таблица S4). Усредненные значения ответов сначала были нормализованы по z-баллу (среднее значение = 0, стандартное отклонение = 1), чтобы гарантировать, что измеренное расстояние отражает несходство между шаблонами ответов, а не величину.Самая низкая дистанция показала сенсиллы GP2 в R . prolixus (Rp-GP2) и GP3 в R . brethesi (Rb-GP3; расстояние = 4,47). Пара Rp-GP4 и Rb-GP3 находилась на другом конце спектра, с наибольшим расстоянием (8,24). Мы обнаружили, что большинство (88%) комбинаций сенсилл находятся в диапазоне 6-8,3 единиц расстояния. Для дальнейшего изучения различий между двумя видами мы выполнили анализ главных компонентов (PCA), в котором 38-мерное пространство сенсиллы было уменьшено до более низких размеров (рис. 7D).Мы сосредоточились на первых двух компонентах, которые вместе объясняют 60% дисперсии. В то время как сенсиллы типа R . brethesi оказались более плотно сгруппированными, чем R . prolixus , расстояние между отдельными типами сенсилл у одного вида было больше, чем между видами (ANOSIM, R = 0,09, p = 0,32). Взятые вместе, эти результаты показывают, что подтипы сенсиллы не обязательно являются видоспецифичными, несмотря на то, что каждый демонстрирует различный запах, настраивая дыхание и реагируя на определенный набор лигандов.
Обсуждение
В данном исследовании морфологические и функциональные различия периферической обонятельной системы R . prolixus и R . Были оценены brethesi , два вида, различающиеся распространением и местом обитания убежища. Морфологические различия, выраженные в большей плотности базиконических и трихоидных сенсилл у R . brethesi по сравнению с R . prolixus , признак, ранее описанный для других триатомов [45, 46, 48, 69, 81].Корреляция между количеством обонятельных сенсилл и ареалом обитания была предложена для триатомов, а также для других видов кроветворных насекомых [68]. Однако для R это еще предстоит подтвердить. prolixus и R . brethesi , так как условия выращивания могут отрицательно повлиять на численность сенсиллы [48, 49]. Однако наши результаты интригуют, так как R . brethesi — специалист по беженцам, гнездящийся только на одном виде пальм в лесной среде, что предполагает, что этот вид может иметь другие потребности или использовать другую стратегию для обнаружения и различения запахов по сравнению с R . проликс .
Для оценки обонятельной функции R . Антенна prolixus мы первоначально использовали EAG-анализ с известными биологически активными соединениями, которые, как ранее было показано, участвуют во внутривидовой коммуникации и другом поведении, управляемом запахом. На удивление антенна R . prolixus отвечал только на ограниченное количество тестируемых соединений. Например, мы не наблюдали значительного ответа ни на 2-бутанон, ни на 3-метил-2-бутанол, соединения, которые, как известно, являются частью полового феромона [82].Мы предполагаем, что отсутствие антенного ответа, вероятно, является следствием небольшого количества специализированных нейронов, обнаруживающих эти соединения, и что ограниченная чувствительность анализа EAG не смогла обеспечить надежный сигнал. Однако вполне возможно, что эти химические вещества обнаруживаются другими органами, кроме антенны, поскольку несколько пахучих и ионтропных рецепторов (OR и IR), наряду с корецептором пахучих веществ orco, экспрессируются также в предплюснах, гениталиях и ростках [83] . Большинство запахов, вызывающих значительную антенную реакцию, были летучими веществами, характерными для запаха позвоночных (амниот), такими как уксусная кислота, пропионовая кислота, масляная кислота, изомасляная кислота, этилпируват, триметиламин и диоксид углерода.Все эти летучие соединения были обнаружены в свободном пространстве позвоночных, самцов и самок R . Было показано, что prolixus привлекает уксусную и изомасляную кислоты [21]. Эти соединения, однако, помимо того, что часто встречаются в секретах позвоночных-хозяев, также используются во внутривидовой коммуникации [84], подчеркивая важность сенсорной экономичности у этих насекомых [17].
У насекомых OR и IR отвечают за обнаружение летучих молекул.IRs считаются предковыми, поскольку они обнаруживаются у базальных насекомых и у их самых недавних филогенетических предшественников [85, 86]. Эти рецепторы, экспрессируемые в дендритах OSN, размещенных в желобчатых сенсиллах (т.е. сенсиллах с двойными стенками), выполняют консервативную функцию при обнаружении кислот и аминов у таксонов насекомых [33, 36]. Тем не менее, мы показываем, что насекомые-триатомы с разными требованиями к среде обитания и хозяевам демонстрируют различия в обонятельной настройке сенсилл GP. В то время как оба вида реагируют на кислоты и амины, различающиеся по разветвлению и длине углерода, R . prolixus , по-видимому, больше настроен на амины, чем его лесной собрат. Этот результат согласуется с предыдущим исследованием, показывающим, что R . prolixus привлекается к аминам, присутствующим в экскрементах позвоночных-хозяев, которые направляют их поиски хозяина [34].
Оба вида различались по своей реакции на определенные пахучие вещества. Р . prolixus показал значительно более сильную реакцию на бутирилхлорид, чем R . человек .Предполагается, что это соединение запаха имеет репеллентную функцию у комаров, подавляя активность нейронов, отвечающих на углекислый газ [60]. Требует подтверждения, выполняет ли бутирилхлорид аналогичную роль в триатомах. Интересно, что R . brethesi показал более высокую реакцию на амилацетат, соединение, обнаруженное в плодах [87, 88], 3-метилиндол, который содержится в кале и соцветиях в низких концентрациях [89–91], масляный альдегид и смесь OR. Предыдущие исследования показали, что соединения, присутствующие в этой смеси, обнаруживаются OR у других видов насекомых [55–58].Пока только одна сенсилла прощупывалась в R . prolixus ответили на эту смесь, половина из них ответила на R . человек . Это предполагает, что OR могут присутствовать в сенсиллах штифта с бороздками R . brethesi , но не R . prolixus , как, например, для пахучего рецептора OR35a, экспрессируемого целоконической сенсиллой Drosophila melanogaster [79]. В целом, эти различия могут отражать особую адаптацию к соответствующей среде.
Основываясь на их профилях запаховой реакции, мы идентифицировали четыре функциональных подтипа сенсиллы у каждого вида. Это контрастирует с исследованиями по T . infestans , где только три типа сенсиллы с бороздками были описаны для 5 личинок возраста [92]. Это несоответствие может быть вызвано несколькими причинами. Во-первых, возможно, что различия между видами триатомов больше, чем ожидалось, как предполагается, например, ультраструктурными исследованиями, демонстрирующими различное количество OSNs в сенсиллах GP триатомов [38, 61].Во-вторых, между T можно распознать различные модели поведения в ответ на пахучие вещества. infestans и R . prolixus [32]. В-третьих, мы регистрировали ответы взрослых особей, тогда как 5 личинок возраста были обследованы в случае Т . Инфестанс . Антенна R . prolixus претерпевает значительные изменения между 5 -м возрастом и взрослыми, с увеличением количества обонятельных сенсилл на жгутиках I и II [74], вероятно, связанных с внутривидовой коммуникацией или поведенческими потребностями.Эти изменения могут объяснять дополнительный подтип сенсиллы, наблюдаемый у взрослых особей Rhodnius . В-четвертых, и, наконец, в нашем скрининге мы зарегистрировали ответы от большего количества химических веществ, чем в предыдущих исследованиях [36], потенциально улучшая разрешение физиологических подтипов сенсиллы. Однако здесь следует отметить, что наше исследование не является окончательным, и записи с дополнительными соединениями могут помочь завершить текущую работу по классификации сенсилл в R . prolixus и R . человек . Кроме того, было бы интересно проанализировать характеристики доза-ответ, чтобы определить лучшие запаховые лиганды для различных типов сенсилл GP в будущих исследованиях.
Интересно, что R . brethesi продемонстрировал в целом более высокие и более широкие обонятельные реакции, чем R . prolixus , что отражено в средней реакции, настройке запаха сенсиллы и разреженности жизни в данных SSR, что предполагает сенсорную дифференциацию между этими видами.У насекомых количество обонятельных рецепторов и сложность экологической ниши, по-видимому, сильно коррелируют, причем количество OR увеличивается с увеличением сложности ниши. Например, в то время как у мухи цеце только 40–46 OR, у эусоциальных насекомых, таких как муравьи, более 350 OR [93–95]. Более того, у комаров было высказано предположение, что предпочтение хозяина объясняет различия в репертуаре хемосенсорных генов между видами-близнецами [96]. Примечательно, что в триатомах обонятельные связывающие белки (OBP) и хемосенсорные белки (CSP) присутствуют на более низких уровнях экспрессии у домашних насекомых — T . brasiliensis по сравнению с лесными и перидоместными [50]. При том, что R . prolixus не является исключительно домицилированным, и в связи с отсутствием в настоящее время данных о химических признаках, с которыми встречаются лесные виды триатомов в дикой природе, мы не можем окончательно определить, отражают ли наблюдаемые различия адаптацию двух видов Rhodnius к их специфическим условиям. среды обитания. Будущие эксперименты по сравнению диких и домицилированных особей R . prolixus необходимы, чтобы пролить свет на эту гипотезу. Важно отметить, что триатомины обрабатывают информацию об запахе в контексте других сенсорных сигналов [30]. Фактически рэнд . prolixus демонстрирует поразительную термочувствительность, а тепло является основным сигналом для этих насекомых, связанным с хозяином [23, 97–99]. Следовательно, в контексте наших результатов это могло быть R . prolixus в меньшей степени полагается на обонятельные сигналы, чем R . brethesi , аналогично тому, что ранее наблюдалось у других видов насекомых [100].
В заключение, наши результаты подтверждают предыдущие наблюдения фенотипической пластичности у рода Rhodnius . Мы демонстрируем, что виды различаются не только морфологией сенсорного оборудования, но и функционально с R . prolixus с явно сниженной обонятельной функцией. Вероятно, что состояние, обнаруженное у лесных видов, представляет состояние наследственного признака в подсемействе, тогда как производное сокращенное состояние может быть связано с изменениями в предпочтении среды обитания.В связи с продолжающимся быстрым разрушением природной среды [8] вполне вероятно, что по этому пути пойдет больше видов. Тщательный анализ различий и потенциальных сдвигов в сенсорном аппарате может оказаться полезным при разработке эффективных будущих стратегий борьбы с переносчиками болезней.
Дополнительная информация
S1 Рис. Сканирующая электронная микроскопия (СЭМ) сенсилл антенн
Rhodnius prolixus .Стрелки указывают: (A) сенсиллум trichobothrium (I) и щетину II (II), (B) сенсиллу типа «стержень в ямке», (C) щетину III, (D) орнаментированную пору и (E) целоконическую сенсиллу 3 типа, на ножке антенны.
https://doi.org/10.1371/journal.pntd.0009098.s001
(TIF)
S2 Рис. Сканирующая электронная микроскопия (СЭМ) антенных сенсилл
Rhodnius brethesi .Стрелки указывают (A) сенсиллум трихоботриум, (B) пещерный орган на ножке (I) и щетине III (II), (B ‘) деталь пещерного органа, (C) базиконный (также известный как тонкостенный трихоид) ) сенсилла, представляющая канал шелушения, (D) целоконическая сенсилла.
https://doi.org/10.1371/journal.pntd.0009098.s002
(TIF)
Благодарности
Мы благодарим Фейта Грабе, Катарину Шнееберг и Сандора Нитше за помощь в морфологическом анализе, Клаудио Лаццари и Марсело Лоренцо за полезные обсуждения, Гюнтера Шауба за постоянное предоставление насекомых и полезную информацию по обращению с насекомыми и уходу за ними и Норису Мели за помощь в селекции насекомых.
Ссылки
- 1. Перес-Молина Х.А., Молина И.Болезнь Шагаса. Ланцет. 2018; 391 (10115): 82–94.
- 2. Ямагата Ю., Накагава Дж. Контроль болезни Шагаса. Успехи паразитологии. 2006. 61: 129–65. pmid: 16735164
- 3. Барг, доктор медицины, Скофилд, С, Дюжарден, Дж. П. Классификация и филогения Triatominae. Американский трипаносомоз: Elsevier; 2010. с. 117–47.
- 4. Тартаротти Э., Азередо-Оливейра М., Церон С. Филогенетический подход к изучению триатомов (Triatominae, Heteroptera).Бразильский журнал биологии. 2006. 66 (2B): 703–8.
- 5. Justi SA, Galvão C. Эволюционное происхождение разнообразия переносчиков болезни Шагаса. Тенденции паразитологии. 2017; 33 (1): 42–52. pmid: 27986547
- 6. Гул Ф., Агилера Дж., Пинто Н., Вергара Д. Обновленное географическое распределение и экоэпидемиология триатомовой фауны (Reduviidae: Triatominae) в Колумбии. Биомедика. 2007. 27: 143–62. pmid: 18154255
- 7. Ангуло В.М., Эстебан Л., Луна КП.Attalea butyracea próximas a las viviendas como posible fuente de infestación domiciliaria por Rhodnius prolixus (Hemiptera: Reduviidae) en los Llanos Orientales de Colombia. Биомедика. 2012; 32 (2): 1–26.
- 8. Д’алессандро А., Баррето П., Саравиа Н., Баррето М. Эпидемиология Trypanosoma cruzi на восточных равнинах Колумбии. Американский журнал тропической медицины и гигиены. 1984. 33 (6): 1084–95. pmid: 6439062
- 9. Хашимото К., Скофилд С.Дж. Ликвидация Rhodnius prolixus в Центральной Америке.Паразиты и векторы. 2012; 5 (1): 1–10. pmid: 22357219
- 10. Рендон Л.М., Гуль Ф., Кордовес Дж. М., Эразо Д. Новые сценарии передачи Trypanosoma cruzi в регионе Ориноко в Колумбии. Memórias do Instituto Oswaldo Cruz. 2015; 110 (3): 283–8. pmid: 25830543
- 11. Galvão C, Justi SA. Обзор экологии Triatominae (Hemiptera: Reduviidae). Acta Tropica. 2015; 151: 116–25. pmid: 26086951
- 12. Ricardo-Silva AH, Lopes CM, Ramos LB, Marques WA, Mello CB, Duarte R и др.Корреляция между популяциями родниуса и присутствием пальм как факторов риска возникновения болезни Шагаса в регионе Амазонки, Бразилия. Acta tropica. 2012. 123 (3): 217–23. pmid: 22664419
- 13. Абад-Франч Ф., Лима М.М., Саркис О., Гургель-Гонсалвес Р., Санчес-Мартин М., Кальсада Дж. И др. О пальмах, клопах и болезни Шагаса в Америке. Acta tropica. 2015; 151: 126–41. pmid: 26196330
- 14. Mascarenhas BM. Tria tomineos da Amazonia: Sobre о среде обитания и algumas consideracoes comportamentais de Rhodnius brethesi matta, 1919 (Hemiptera: Reduviidae: Triatominae) na regiao do médio rio negro, Амазонас.1991.
- 15. Гомеш В.Х., Виейра И.К., Саломау Р.П., тер Стиге Х. Виды деревьев Амазонки, которым угрожают вырубка лесов и изменение климата. Изменение климата природы. 2019; 9 (7): 547–53.
- 16. Горла DE, Catala SS, Grilli MP. Efecto de la temperatura sobre la distribucion de Triatoma infestans y el riesgo de transmision vectorial de la enfermedad de Chagas en Argentina. Acta Toxicologica Argentina 5 (1): 36–9.
- 17. Lazzari CR. Глава 1. Ориентация на хозяев кроветворных насекомых.2009. с. 1–58.
- 18. Núñez JA. Ориентация на источники пищи и активность у Rhodnius prolixus Stål (Hemiptera: Reduviidae). Бюллетень энтомологических исследований. 1982. 72 (2): 253–62.
- 19. Баррозо РБ, Лаццари ЧР. Ориентационное поведение кровососущего клопа Triatoma infestans на короткоцепочечные жирные кислоты: синергетический эффект L-молочной кислоты и углекислого газа. Химические чувства. 2004. 29 (9): 833–41. pmid: 15574819
- 20. Баррозо РБ, Лаццари ЧР.Реакция кровососущего клопа Triatoma infestans на углекислый газ и другие запахи хозяина. Chem Senses. 2004. 29 (4): 319–29. Epub 2004/05/20. pmid: 15150145
- 21. Ортис М.И., Молина Дж. Предварительные доказательства привлекательности Rhodnius prolixus (Hemiptera: Triatominae) экстрактами запаха кожи человека. Acta tropica. 2010. 113 (2): 174–9. pmid: 19879853
- 22. Табарес М., Ортис М., Гонсалес М., Караццон С., Флорес М.Дж.В., Молина Дж. Поведенческие реакции Rhodnius prolixus на летучие органические соединения, выделяемые in vitro бактериями, выделенными из кожи лица человека.PLoS игнорирует тропические болезни. 2018; 12 (4): e0006423. pmid: 29684012
- 23. Guerenstein PG, Lazzari CR. Поиск хозяина: как триатомины получают и используют информацию для поиска крови. Acta Trop. 2009. 110 (2–3): 148–58. Epub 2008/11/06. pmid: 18983971
- 24. Гвидобальди Ф., Геренштейн П.Г. Синтетическая смесь хозяина и запаха, не содержащая CO2, которая притягивает и улавливает триатомины: влияние соотношений выделяемых одорантов. Журнал медицинской энтомологии. 2016; 53 (4): 770–5. pmid: 27252545
- 25.Bernier UR, Booth MM, Yost RA. Анализ излучений кожи человека с помощью газовой хроматографии / масс-спектрометрии. 1. Термическая десорбция аттрактантов комара желтой лихорадки (Aedes a egypti) с обработанных стеклянных бусинок. Аналитическая химия. 1999. 71 (1): 1–7. pmid: 92
- 26. Penn DJ, Oberzaucher E, Grammer K, Fischer G, Soini HA, Wiesler D, et al. Индивидуальные и гендерные отпечатки пальцев в запахе человеческого тела. Журнал интерфейса Королевского общества. 2007. 4 (13): 331–40. pmid: 17251141
- 27.Корк А., Парк К. Идентификация электрофизиологически активных соединений малярийного комара Anopheles gambiae в экстрактах человеческого пота. Медицинская и ветеринарная энтомология. 1996. 10 (3): 269–76. pmid: 8887339
- 28. Бракс М., Андерсон Р., Кнолс Б. Инфохимические вещества в выборе хозяина-комара: микрофлора кожи человека и паразиты Plasmodium. Паразитология сегодня. 1999. 15 (10): 409–13. pmid: 10481153
- 29. Wigglesworth V, Gillett J. Функция антенн у Rhodnius prolixus (Hemiptera) и механизм ориентации на хозяина.Журнал экспериментальной биологии. 1934; 11 (2): 120–39.
- 30. Баррозо РБ, Райзенман CE, Геренштейн П., Лаццари CR, Лоренцо MG. Взгляд изнутри на сенсорную биологию триатомов. J. Insect Physiol. 2017; 97: 3–19. Epub 2016/11/15. pmid: 27840287
- 31. Taneja J, Guerin PM. Аммиак привлекает гематофага Triatoma infestans: поведенческие и нейрофизиологические данные о нимфах. Журнал сравнительной физиологии A. 1997; 181 (1): 21–34.
- 32.Taneja J, Guerin PM. Ориентированные реакции трехатомных клопов Rhodnius prolixus и Triatoma infestans на запахи позвоночных в сервосфере. Журнал сравнительной физиологии A. 1995; 176 (4): 455–64.
- 33. Геренштейн П., Герен П. Обонятельные и поведенческие реакции кровососущего насекомого Triatoma infestans на запахи позвоночных-хозяев. Журнал экспериментальной биологии. 2001. 204 (3): 585–97. pmid: 11171309
- 34. Otálora-Luna F, Guerin PM.Амины позвоночных приводят к ресурсам триатомовых клопов. Журнал физиологии насекомых. 2014; 71: 52–60. pmid: 25260571
- 35. Оталора-Луна Ф, Герен П.М. Амины позвоночных животных направляют триатомовых клопов к ресурсам. J. Insect Physiol. 2014; 71: 52–60. Epub 2014/09/28. pmid: 25260571
- 36. Diehl PA, Vlimant M, Guerenstein P, Guerin PM. Ультраструктура и ответы рецепторных клеток сенсиллы усиков с бороздками Triatoma infestans (Hemiptera: Reduviidae).Строение и развитие членистоногих. 2003. 31 (4): 271–85.
- 37. Майер М. Ответ одиночной обонятельной клетки Triatoma infestans на дыхание человека. Природа. 1968. 220 (5170): 924–5. pmid: 4881008
- 38. Bernard J. électrophysiologique de récepteurs impliqués dans l’orientation vers l’hôte et dans l’acte hématophage chez un Hémiptère, Triatoma infestans 1974.
- 39. Reisenman CE. Голод — лучшая пряность: последствия голодания в ответах усиков кровососущего клопа Rhodnius prolixus.Журнал физиологии насекомых. 2014; 71: 8–13. pmid: 25280630
- 40. Макбрайд С.С., Байер Ф., Омонди А.Б., Спитцер С.А., Лютомия Дж., Санг Р. и др. Эволюция предпочтения комаров людьми, связанная с рецептором запаха. Природа. 2014. 515 (7526): 222–7. pmid: 253
- 41. Макбрайд К.С. Гены и запахи, лежащие в основе недавней эволюции предпочтения комаров людьми. Текущая биология. 2016; 26 (1): R41 – R6. pmid: 26766234
- 42. Ауэр Т.О., Халлаф М.А., Силберинг А.Ф., Заппиа Дж., Эллис К., Альварес-Оканья Р. и др.Эволюция обонятельных рецепторов и цепей способствует специализации хозяина. Природа. 2020; 579 (7799): 402–8. pmid: 32132713
- 43. Гулд Ф., Эсток М., Хиллиер Н.К., Пауэлл Б., Грут А.Т., Уорд С.М. и др. Сексуальная изоляция самцов моли объясняется одним феромонным ответом QTL, содержащим четыре рецепторных гена. Труды Национальной академии наук. 2010. 107 (19): 8660–5. pmid: 20404144
- 44. Чжао З., Макбрайд CS. Эволюция обонятельных цепей у насекомых. Журнал сравнительной физиологии А.2020; 206 (3): 353–67.
- 45. Catalá SS. Усиковые сенсиллы Triatominae (Hemiptera, Reduviidae): сравнительное исследование пяти родов. Международный журнал морфологии и эмбриологии насекомых. 1997. 26 (2): 67–73.
- 46. Catalá S, Dujardin JP. Структура сенсилл антенн указывает на географическую и экотопическую изменчивость среди популяций Triatoma infestans (Hemiptera: Reduviidae). Журнал медицинской энтомологии. 2001. 38 (3): 423–8. pmid: 11372968
- 47.Dujardin J, Bermudez H, Casini C, Schofield C, Tibayrenc M. Метрические различия между Silvatic и домашними Triatoma infestans (Heteroptera: Reduviidae) в Боливии. Журнал медицинской энтомологии. 1997. 34 (5): 544–51. pmid: 9379460
- 48. Souza AC, Catala S, Carbajal de la Fuente AL, Junqueira ACV. Фенотипическая изменчивость амазонских видов Rhodnius brethesi (Hemiptera: Reduviidae). J Med Entomol. 2017; 54 (4): 909–16. Epub 2017/04/28. pmid: 28449112
- 49.Catala SS, Maida D, Caro-Riaño H, Jaramillo N, Moreno J. Изменения, связанные с лабораторным выращиванием в структуре сенсилл усиков Triatoma infestans, Rhodnius prolixus и Rhodnius pallescens (Hemiptera, Reduviidae, Triatominae). Memórias do Instituto Oswaldo Cruz. 2004. 99 (1): 25–30. pmid: 15057343
- 50. Марчант А., Мугель Ф., Жакин-Жоли Э., Коста Дж., Алмейда С.Э., Гарри М. Недостаточная экспрессия хемосенсорных генов у домашних клопов вектора болезни Шагаса Triatoma brasiliensis.PLoS игнорирует тропические болезни. 2016; 10 (10): e0005067. pmid: 27792774
- 51. Balczun C, Siemanowski J, Pausch JK, Helling S, Marcus K, Stephan C и др. Кишечные аспартатные протеазы TiCatD и TiCatD2 гематофага Triatoma infestans (Reduviidae): характеристика последовательности, характер экспрессии и характеристика протеолитической активности. Биохимия насекомых и молекулярная биология. 2012; 42 (4): 240–50. pmid: 22210150
- 52. Штадлер А., Мейзер К. К., Шауб Г. А..«Живые шприцы»: использование гематофагов в качестве пробоотборников крови мелких и диких животных. Природа помогает: Springer; 2011. с. 243–71.
- 53.
Боден А, Баррозо РБ, Кутон Л, Лаццари ЧР. Временная модуляция и адаптивный контроль поведенческой реакции на запахи у Rhodnius prolixus. Журнал физиологии насекомых. 2008. 54 (9): 1343–8. pmid: 186
- 54. Ортис М.И., Суарес-Ривильяс А., Молина Дж. Поведенческие реакции на экстракты кожи человека и антеннальные фенотипы лесных первого дочернего поколения и долго выращиваемой лабораторной колонии Rhodnius prolixus.Memorias do Instituto Oswaldo Cruz. 2011; 106 (4): 461–6. pmid: 21739035
- 55. Кэри А.Ф., Ван Дж., Су Си-И, Цвибель Л.Дж., Карлсон-младший. Прием одоранта при малярийном комаре Anopheles gambiae. Природа. 2010. 464 (7285): 66–71. pmid: 20130575
- 56. Ван Г., Кэри А.Ф., Карлсон-младший, Цвибель Л.Дж. Молекулярные основы кодирования запаха москита Anopheles gambiae — переносчика малярии. Труды Национальной академии наук. 2010. 107 (9): 4418–23. pmid: 20160092
- 57.Де Брюйн М., Фостер К., Карлсон-младший. Кодирование запаха в антенне дрозофилы. Нейрон. 2001. 30 (2): 537–52. pmid: 11395013
- 58. Халлем Э.А., Карлсон-младший. Кодирование запахов рецепторным репертуаром. Клетка. 2006; 125 (1): 143–60. pmid: 16615896
- 59. Саксе С., Рюкерт Э., Келлер А., Окада Р., Танака Н.К., Ито К. и др. Зависимая от активности пластичность обонятельного контура. Нейрон. 2007. 56 (5): 838–50. pmid: 18054860
- 60. Tauxe GM, MacWilliam D, Boyle SM, Guda T, Ray A.Нацеливание на двойной детектор кожи и CO2 для изменения поиска хозяина-комара. Клетка. 2013; 155 (6): 1365–79. Epub 2013/12/10. pmid: 24315103
- 61. Чанка С. Ультраструктура сенсилл усиков клопа Rhodnius prolixus (Hemiptera, Reduviidae). Паразитология. 1980; 14 (6): 486. pmid: 7007992
- 62. Ozaki M, Wada-Katsumata A, Fujikawa K, Iwasaki M, Yokohari F, Satoji Y и др. Распознавание с помощью хемосенсорной сенсиллы муравейника и других, не соплеменников. Наука.2005. 309 (5732): 311–4. pmid: 15947139
- 63. Джейн Д.Д., Убес Д. Р.К .: Алгоритмы кластеризации данных. Серия расширенных справочных материалов Прентис-Холла; 1988.
- 64. Zahn CT. Теоретико-графические методы обнаружения и описания гештальт-кластеров. IEEE Transactions на компьютерах. 1971. 100 (1): 68–86.
- 65. Ø молотка, Харпер Д.А., Райан П.Д. ПРОШЛОЕ: программный пакет палеонтологической статистики для обучения и анализа данных. Palaeontologia electronica.2001; 4 (1): 9.
- 66. Grabe V, Baschwitz A, Dweck HK, Lavista-Llanos S, Hansson BS, Sachse S. Выяснение нейрональной архитектуры обонятельных клубочков в антеннальной доле дрозофилы. Сотовые отчеты. 2016; 16 (12): 3401–13. pmid: 27653699
- 67. Эрнандес Л., Абрахан Л., Морено М., Горла Д., Катала С. Фенотипическая изменчивость, связанная с геномными изменениями в основном переносчике болезни Шагаса в южной части Южной Америки. Acta Tropica. 2008. 106 (1): 60–7.pmid: 18328454
- 68. Сайед З. Химическая экология и обоняние у членистоногих-переносчиков болезней. Curr Opin Insect Sci. 2015; 10: 83–9. Epub 2015/08/01. pmid: 29588018
- 69. Родригес К.С., Крокко Л., Альтамирано А., Катала С. Изменения, связанные с полом, географическим населением и средой обитания, в фенотипе усиков Triatoma patagonica Del Ponte, 1929 (Hemiptera: Reduviidae). Acta tropica. 2013; 125 (2): 143–9. pmid: 23178218
- 70. Май-Конча I, Геренштейн П.Г., Рэмси Дж. М., Рохас Дж. К., Катала С.Антенный фенотип мексиканских гаплогрупп комплекса Triatoma dimidiata, переносчиков болезни Шагаса. Инфекция, генетика и эволюция. 2016; 40: 73–9. pmid: 268
- 71. Шанбхаг С., Мюллер Б., Штайнбрехт Р. Атлас обонятельных органов Drosophila melanogaster: 1. Типы, внешняя организация, иннервация и распределение обонятельных сенсилл. Международный журнал морфологии и эмбриологии насекомых. 1999. 28 (4): 377–97.
- 72. Catalá S. Пещерный орган Triatominae (Hemiptera, Reduviidae) под растровой электронной микроскопией.Memórias do Instituto Oswaldo Cruz. 1994. 89 (2): 275–7. pmid: 7885257
- 73. Lazzari CR, Wicklein M. Пещерный орган чувств в усиках клопов Triatominae. Memórias do Instituto Oswaldo Cruz. 1994. 89 (4): 643–8.
- 74. Catalá S, Schofield C. Антенные сенсиллы Родниуса. Журнал морфологии. 1994. 219 (2): 193–203. pmid: 29865357
- 75. Макивер С., Семицки Р. Тонкая структура предполагаемой термо- / гигросенсиллы усиков взрослого Rhodnius prolixus Stål (Hemiptera: Reduviidae).Журнал морфологии. 1985. 183 (1): 15–23. pmid: 29976013
- 76. Mota T, Vitta AC, Lorenzo-Figueiras AN, Barezani CP, Zani CL, Lazzari CR и др. Многовидовая приманка для переносчиков болезни Шагаса. PLoS игнорирует тропические болезни. 2014; 8 (2). pmid: 24587457
- 77. Guerenstein PG, Guerin PM. Сравнение летучих веществ, выделяемых взрослыми особями трех видов триатомов. Entomologia experimentalis et application. 2004. 111 (2): 151–5.
- 78. Питтс Р.Дж., Деррибери С.Л., Чжан З., Цвибель Л.Дж.Вариантные ионотропные рецепторы москита Anopheles gambiae, переносчика малярии, настроены на амины и карбоновые кислоты. Научные отчеты. 2017; 7 (1): 1–11. pmid: 28127051
- 79. Яо Калифорния, Игнелл Р., Карлсон-младший. Хемосенсорное кодирование нейронов целоконической сенсиллы антенны дрозофилы. Журнал неврологии. 2005. 25 (37): 8359–67. pmid: 16162917
- 80. Паппенбергер Б., Гейер М., Бекх Дж. Ответы обонятельных рецепторов усиков у комара желтой лихорадки Aedes aegypti на запахи человеческого тела.Обоняние при взаимодействии комара с хозяином. 1996: 254–66. pmid: 8894302
- 81. Carbajal De La Fuente A, Catalá S. Связь между рисунком сенсилл усиков и средой обитания у шести видов Triatominae. Memorias do Instituto Oswaldo Cruz. 2002. 97 (8): 1121–5. pmid: 12563477
- 82. Понтес Г.Б., Бохман Б., Унелиус К.Р., Лоренцо М.Г. Летучие метастернальные железы и половая коммуникация у трехатомного клопа, Rhodnius prolixus. Журнал химической экологии. 2008. 34 (4): 450–7.pmid: 18317844
- 83. Latorre-Estivalis JM, de Oliveira ES, Esteves BB, Guimarães LS, Ramos MN, Lorenzo MG. Паттерны экспрессии генов пахучих рецепторов в векторе болезни Шагаса. Биохимия насекомых и молекулярная биология. 2016; 69: 71–81. pmid: 26003917
- 84. Крус-Лопес Л., Мало Е., Рохас Дж., Морган Е. Химическая экология триатомовых ошибок: переносчики болезни Шагаса. Медицинская и ветеринарная энтомология. 2001. 15 (4): 351–7. pmid: 11776453
- 85.Ритц Р., Крозет В., Бентон Р. Ионотропные рецепторы (ИР): хемосенсорные ионотропные рецепторы глутамата у дрозофилы и за ее пределами. Биохимия насекомых и молекулярная биология. 2013; 43 (9): 888–97. pmid: 23459169
- 86. Croset V, Rytz R, Cummins SF, Budd A, Brawand D, Kaessmann H, et al. Древнее протостомическое происхождение хемосенсорных ионотропных рецепторов глутамата и эволюция вкуса и обоняния насекомых. PLoS генетика. 2010; 6 (8). pmid: 20808886
- 87. МАККАРТИ А.И., ПАЛМЕР Дж. К., Шоу С. П., АНДЕРСОН Э.Корреляция данных газовой хроматографии с характеристиками вкуса свежих плодов банана а. Журнал пищевой науки. 1963. 28 (4): 379–84.
- 88. Чжу Дж., Пак К-К, Бейкер ТК. Распознавание запахов перезрелого манго, привлекающих уксусных мух, Drosophila melanogaster. Журнал химической экологии. 2003. 29 (4): 899–909. pmid: 12775150
- 89. Дженсен М.Т., Кокс Р.П., Дженсен Б.Б. Производство 3-метилиндола (скатола) и индола смешанными популяциями фекальных бактерий свиней. Прикладная и экологическая микробиология.1995. 61 (8): 3180–4. pmid: 7487051
- 90. Смит Б.Н., Миз Б.Дж. Образование летучих аминов и скатола при цветении у некоторых видов арбузных лилий. Физиология растений. 1966. 41 (2): 343–7. pmid: 16656260
- 91. Борг-Карлсон A-K, Englund FO, Unelius CR. Диметилолигосульфиды, основные летучие вещества, выделяемые из Sauromatum guttatum и Phallus impudicus. Фитохимия. 1994. 35 (2): 321–3.
- 92. Guerenstein PG, Lazzari CR. 14. Роль обоняния в поиске хозяина клопами-триатомами.Обоняние во взаимодействиях вектор-хозяин. 2010; 2: 309.
- 93. Macharia R, Mireji P, Murungi E, Murilla G, Christoffels A, Aksoy S, et al. Полногеномный сравнительный анализ семейств хемосенсорных генов у пяти видов мухи цеце. PLoS игнорирует тропические болезни. 2016; 10 (2): e0004421. pmid: 26886411
- 94. Обиеро Г.Ф., Миреджи П.О., Ньянджом С.Р., Кристоффельс А., Робертсон Х.М., Масига Д.К. Пахучие и вкусовые рецепторы мухи цеце Glossina morsitans morsitans.PLoS Negl Trop Dis. 2014; 8 (4): e2663. pmid: 24763191
- 95. Чжоу X, Рокас А., Бергер С.Л., Либих Дж., Луч А., Цвибель Л.Дж. Эволюция хеморецепторов у перепончатокрылых и ее значение для эволюции эусоциальности. Биология и эволюция генома. 2015; 7 (8): 2407–16. pmid: 26272716
- 96. Rinker DC, Zhou X, Pitts RJ, Rokas A, Zwiebel LJ, Consortium A. Профили антенного транскриптома анофелиновых комаров показывают обонятельную специализацию человеческого хозяина у Anopheles gambiae.BMC genomics. 2013; 14 (1): 749. pmid: 24182346
- 97. Fresquet N, Lazzari CR. Реакция на тепло у Rhodnius prolixus: роль теплового фона. Журнал физиологии насекомых. 2011; 57 (10): 1446–9. pmid: 21806990
- 98. Шмитц Х., Треннер С., Хофманн М. Х., Блекманн Х. Способность Rhodnius prolixus (Hemiptera; Reduviidae) приближаться к источнику тепла исключительно с помощью инфракрасного излучения. Журнал физиологии насекомых. 2000. 46 (5): 745–51. pmid: 10742523
- 99.Lazzari CR. Ориентация на хозяев у кроветворных насекомых: интегративная перспектива. Успехи физиологии насекомых. 2009; 37: 1–58.
- 100. Кизи И. В., Грабе В., Грубер Л., Кёрте С., Обиеро Г. Ф., Болтон Г. и др. Обратное распределение ресурсов между зрением и обонянием у представителей рода Drosophila. Связь природы. 2019; 10 (1): 1–16. pmid: 30602773
Фиолетовый вектор — нос. Психология и ваши отверстия — Psy8
Стратегическая гвардия
Этот вектор, как и красный, легче рассматривать в контексте древнего отряда наших предков, в первую очередь потому, что способность воспринимать запахи является древнейшим из всех чувств, известных живым существам.Первые маленькие существа на планете, когда впервые появилась жизнь, не могли ни видеть, ни слышать. Вся информация, которой они располагали, была получена через их обоняние: они использовали обоняние, чтобы искать пищу и друг друга, защищаться от опасности и находить удобное место для жизни. Перенесемся на миллионы лет вперед, и сегодня у нас есть люди, которые живут так же.
В то время как люди с зеленым вектором (дневные сторожи) и с голубым вектором (ночные сторожа) обнаруживают опасность с хороших 3-5 километров, этого не всегда достаточно.Вот почему отряду нужен кто-то, называемый «стратегическая охрана», способный уловить опасность задолго до того, как она подойдет достаточно близко, чтобы стать угрозой.
Согласно науке, комары могут ощущать присутствие людей на расстоянии одиннадцати километров по запаху одной молекулы. Когда мы выходим в лес и через несколько часов к нам обращается их облако, мы не просто наткнулись на то место, где они уже гудели; столько времени им потребовалось, чтобы пролететь эти одиннадцать километров.
Возвращаясь к обсуждаемой теме, у таких людей преобладает их обоняние, и в нашей системе они сгруппированы в фиолетовый вектор.
Виктор Толкачев назвал это «обонятельным» вектором и описал его следующим образом:
Такие люди сидят на высоком холме и нюхают доносящийся до них воздух. Если бы они были со всеми у костра, где жарится мясо (а друг сидел рядом с ними после пары недель без купания), они не смогли бы выполнять свою работу. Вместо этого они чаще всего одиночки, которые сидят подальше от группы. Поскольку наличие людей поблизости означает, что они не могут выполнять свою основную функцию, люди с фиолетовым вектором ненавидят весь «вонючий» отряд.А кто главная «стерва»? Конечно, лидер с красным вектором, от которого пахнет феромонами, которые приходят с его силой. В результате люди с фиолетовым вектором не переносят альф.
Посмотреть по теме:
Предыдущая глава: VIII. Синий вектор — уши Следующая глава: X. Алфавитный список черт характера и внешних особенностейИспользование глубокого обучения для предсказания обонятельных свойств молекул
Автор: Александр Б. Вильчко, старший научный сотрудник Google ResearchЗапах — это чувство, которое разделяют невероятное количество живых организмов, и он играет решающую роль в том, как они анализируют мир и реагируют на него.У людей обоняние связано со способностью наслаждаться едой, а также может вызывать яркие воспоминания. Запах позволяет нам оценить все ароматы, которыми изобилует наша повседневная жизнь, будь то пресловутые розы, партия свежеиспеченного печенья или любимые духи. Тем не менее, несмотря на свою важность, исследователи машинного обучения не уделяют столько внимания запаху, как зрению и слуху.
Восприятие запаха у человека является результатом активации 400 различных типов обонятельных рецепторов (OR), экспрессируемых в 1 миллионе обонятельных сенсорных нейронов (OSN) в небольшом участке ткани, называемом обонятельным эпителием.Эти OSN отправляют сигналы в обонятельную луковицу, а затем в другие структуры мозга. Основываясь на аналогичных достижениях в области глубокого обучения для зрения и звука, должна быть возможность напрямую предсказать конечный сенсорный результат входной молекулы, даже не зная сложных деталей всех задействованных систем. Решение проблемы прогнозирования запаха поможет открыть новые синтетические одоранты, тем самым уменьшив экологическое воздействие сбора натуральных продуктов. Изучение полученных обонятельных моделей может даже привести к новому пониманию биологии запаха.
Небольшие молекулы одоранта являются основными строительными блоками ароматизаторов и ароматизаторов и, следовательно, представляют собой простейшую версию задачи прогнозирования запаха. Тем не менее, каждая молекула может иметь несколько дескрипторов запаха. Например, Ванилин имеет такие дескрипторы, как сладкий, ванильный, сливочный, и шоколадный , причем одни ноты более очевидны, чем другие. Таким образом, прогнозирование запаха также является проблемой классификации с несколькими метками.
В статье «Машинное обучение для запаха: изучение обобщенных перцептивных представлений малых молекул» мы используем графические нейронные сети (GNN), своего рода глубокую нейронную сеть, предназначенную для работы с графами в качестве входных данных, чтобы напрямую предсказывать дескрипторы запаха для отдельных молекул. , без использования каких-либо созданных вручную правил.Мы демонстрируем, что этот подход дает значительно более высокую эффективность в прогнозировании запаха по сравнению с существующими современными технологиями и является многообещающим направлением для будущих исследований.
Графические нейронные сети для прогнозирования запахов
Поскольку молекулы аналогичны графам, с атомами, образующими вершины, и связями, образующими ребра, GNN являются естественной моделью для их понимания. Но как перевести структуру молекулы в графическое представление? Первоначально каждый узел в графе представлен в виде вектора с использованием любой предпочтительной характеристики — идентичности атома, заряда атома и т. Д.Затем, в серии шагов передачи сообщений, каждый узел передает свое текущее значение вектора каждому из своих соседей. Затем функция обновления принимает набор отправленных ей векторов и генерирует обновленное значение вектора. Этот процесс можно повторять много раз, пока, наконец, все узлы графа не будут объединены в один вектор посредством суммирования или усреднения. Этот единственный вектор, представляющий всю молекулу, затем может быть передан в полностью связанную сеть в качестве изученной молекулярной характеристики.Эта сеть выводит прогноз для дескрипторов запаха, предоставленный экспертами по парфюмерии.
Каждый узел представлен как вектор, и каждая запись в векторе изначально кодирует некоторую информацию атомарного уровня. Для каждого узла мы смотрим на соседние узлы и собираем их информацию, которая затем преобразуется с помощью нейронной сети в новую информацию для центрированного узла.Эта процедура выполняется итеративно. Другие варианты GNN используют информацию на уровне ребер и графов.
Это представление ничего не знает о пространственном расположении атомов, и поэтому оно не может различать стереоизомеры, молекулы, состоящие из одних и тех же атомов, но в немного разных конфигурациях, которые могут пахнуть по-разному, такие как (R) — и (S) -карвон. .Тем не менее, мы обнаружили, что даже без разделения стереоизомеров на практике все еще можно довольно хорошо предсказать запах.Иллюстрация GNN для предсказания запаха. Мы переводим структуру молекул в графы, которые загружаются в слои GNN, чтобы лучше представить узлы. Эти узлы сокращаются до одного вектора и передаются в нейронную сеть, которая используется для прогнозирования нескольких дескрипторов запаха. Для предсказания запаха GNN последовательно демонстрируют улучшенную производительность по сравнению с предыдущими современными методами, такими как случайные леса, которые не кодируют структуру графа напрямую. Величина улучшения зависит от того, какой запах нужно предсказать.
Изучение модели и расширение ее на другие задачиПример эффективности GNN по дескрипторам запаха по сравнению с сильным исходным уровнем, как измерено с помощью оценки AUROC.Примеры описателей запаха выбираются случайным образом. Ближе к 1.0 — лучше. В большинстве случаев GNN значительно превосходят базовые показатели полевого стандарта, с аналогичной производительностью, наблюдаемой по сравнению с другими показателями (например, AUPRC, отзыв, точность).
Помимо прогнозирования дескрипторов запаха, GNN могут применяться к другим задачам обоняния. Например, возьмем случай классификации новых или усовершенствованных дескрипторов запаха с использованием только ограниченных данных.Для каждой молекулы мы извлекаем изученное представление из промежуточного уровня модели, которое оптимизировано для наших дескрипторов запаха, что мы называем «внедрением запаха». Это можно рассматривать как обонятельную версию цветового пространства, такую как RGB или CMYK. Чтобы увидеть, полезно ли это внедрение запаха для прогнозирования связанных, но разных задач, мы разработали эксперименты, которые проверяют наше выученное встраивание на связанных задачах, для которых оно не было изначально предназначено. Затем мы сравнили производительность нашего представления внедрения запаха с обычным хемоинформатическим представлением, которое кодирует структурную информацию молекулы, но не зависит от запаха, и обнаружили, что внедрение запаха обобщается для нескольких сложных новых задач, даже соответствующих современному уровню техники. на некоторых.
Работа на будущееДвухмерный снимок нашего пространства для встраивания с выделенными примерами запахов. Слева: Каждый запах группируется в своем собственном пространстве. Справа: Иерархический характер дескриптора запаха. Заштрихованные и контурные области вычисляются с оценкой плотности ядра вложений.
В сфере машинного обучения запах остается самым неуловимым из чувств, и мы рады продолжить небольшую часть, чтобы пролить свет на него с помощью дальнейших фундаментальных исследований.Возможности для будущих исследований многочисленны и касаются всего: от разработки новых обонятельных молекул, которые дешевле и более устойчиво производятся, до оцифровки ароматов или даже когда-нибудь дать тем, у кого нет обоняния, доступ к розам (и, к сожалению, яйца). Мы надеемся также привлечь внимание к этой проблеме большей части мира машинного обучения путем создания в конечном итоге высококачественных открытых наборов данных и обмена ими.Благодарности
Это раннее исследование является результатом работы и рекомендаций группы талантливых исследователей и инженеров Google Brain — Бенджамина Санчес-Ленгелинга, Дженнифер Вей, Брайана Ли, Эмили Рейф, Кэри Радебо, Макса Билески, Йони Халперн, Д.Скалли. Мы рады сотрудничать в этой работе с Ричардом Геркиным из АГУ и Аланом Аспуру-Гузиком из Университета Торонто. Мы, конечно, опираемся на огромный объем предыдущей работы, и особенно извлекли пользу из работы Джастина Гилмера, Джорджа Даля и других по фундаментальной методологии GNN, а также многих других работ в области нейробиологии, статистики и химии.