
Полушария головного мозга: Большие полушария головного мозга — урок. Биология, 8 класс.
Полушария головного мозга
Конечный мозг продольной щелью делится на два полушария, соединяющиеся друг с другом посредством системы спаек. Полушария большого мозга — наиболее прогрессивно развивающаяся у позвоночных животных часть головного мозга. В ходе эволюции позвоночных животных полушария становятся относительно и абсолютно все более крупными. Даже у таких примитивных плацентарных животных, как еж, они доминируют в головном мозге. Если общую массу мозга принять за 1000, то у слона на долю полушарий приходится 630/1000, а на долю мозгового ствола-370/1000, у кошки — соответственно 614/1000 и 386/1000, у узконосых обезьян (например, мартышек) — 709/1000 и 291/1000. Наконец, у человека полушария составляют 780/1000 общей массы головного мозга, а на все другие части мозга, в том числе мозжечок, — только 220/1000.
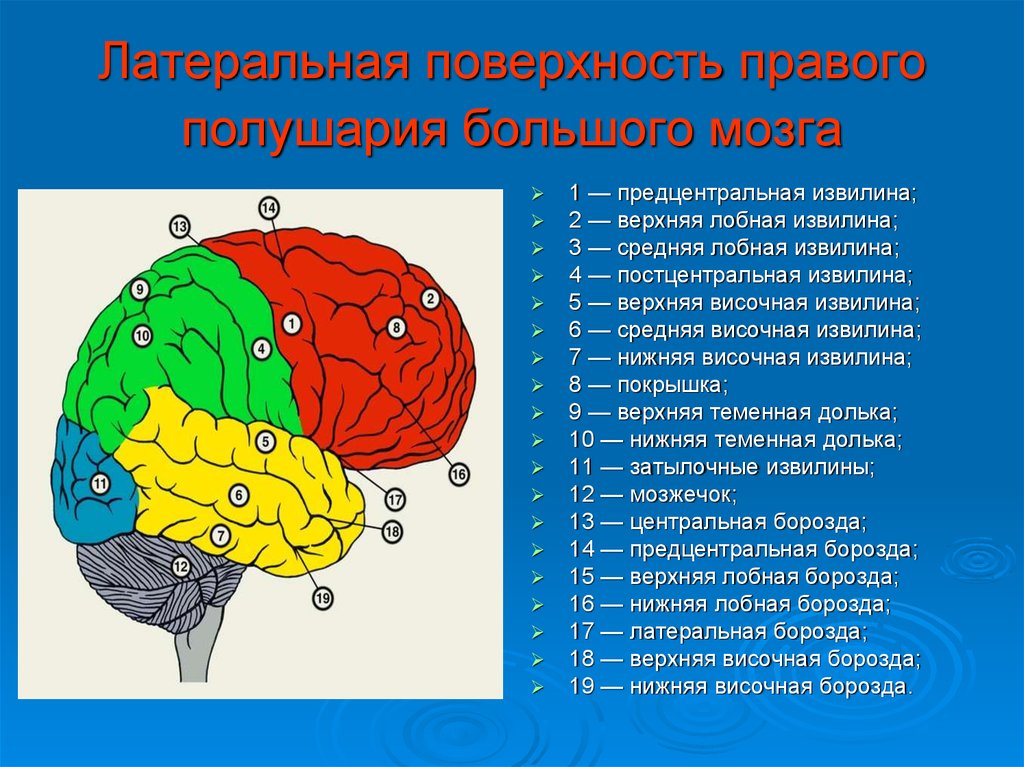
Каждое полушарие разделено
на 5 долей: лобную, височную, теменную,
затылочную и островковую. У человека
лобная доля полушария весит 450 г, теменная
— 251 г, височная и затылочная вместе — 383
г.
Полушария большого мозга имеют сложный рельеф, обусловленный наличием борозд и извилин. Поверхность полушарий покрыта серым веществом — корой большого мозга. Внутренние части полушарий состоят из белого вещества, в котором располагаются нервные ядра и боковые желудочки.
|
Кора большого мозга является наиболее дифференцированной и сложно устроенной нервной структурой. С корой связаны высшие формы отражения внешнего мира, все виды сознательной деятельности человека.
Образование борозд полушарий
начинается на 5-м месяце эмбрионального
развития. Первой образуется латеральная
(сильвиева) борозда, затем появляются
другие первичные борозды: центральная
(роландова), теменно-затылочная,
гиппокампальная, шпорная.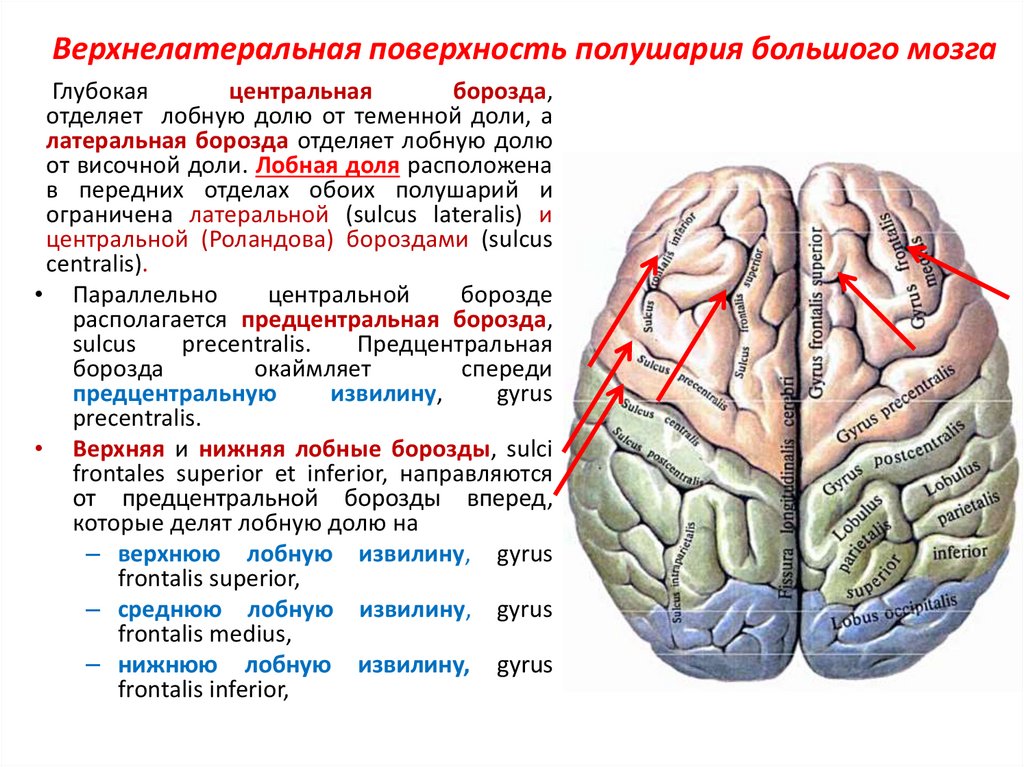
|
Площадь поверхности обоих
полушарий варьирует у взрослых людей
от 1469 до 1670 см2.
Из общей поверхности коры 2/3 находится
в глубине борозд и щелей, а 1/3 занимают
извилины и видимая поверхность полушарий.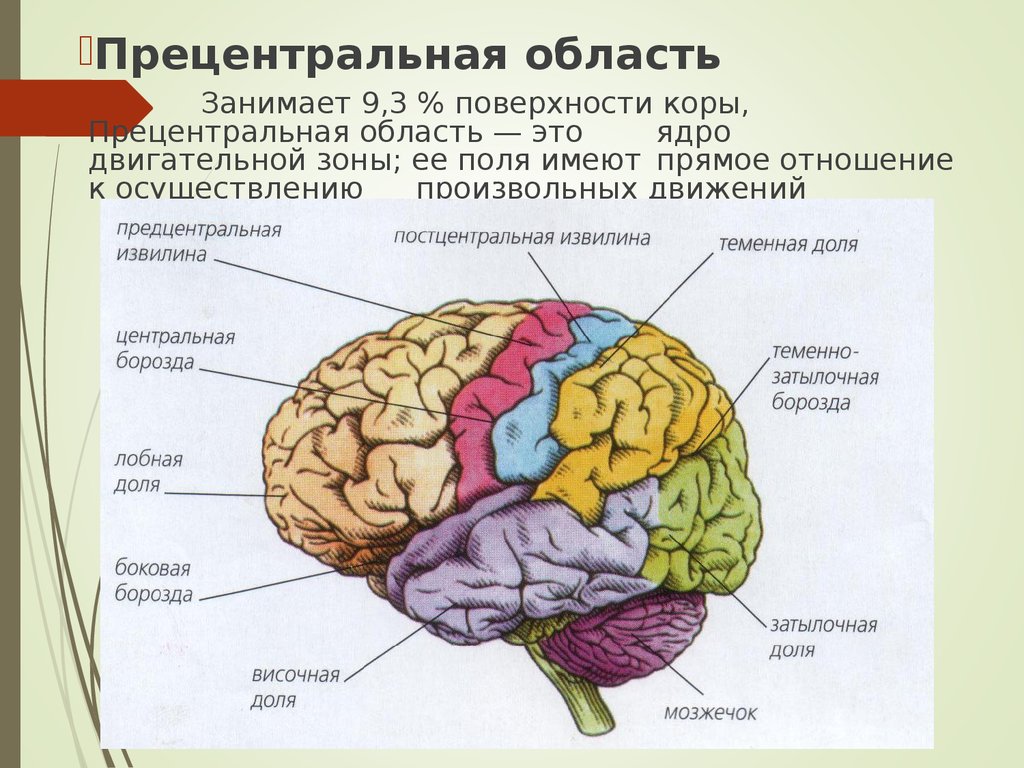 У человека толщина коры колеблется от
1.25 до 4 и даже до 6 мм. В глубине борозд
ширина коры уменьшается в 2-2.5 раза по
сравнению с вершиной извилины. Если
принять во внимание, что поверхность
коры в одном полушарии составляет в
среднем 800 см2,
а ее толщина равна в среднем 3 мм, то
объем коры составит 240 см3,
или 44% всего объема полушария. Количество
нейронов коры оценивается в 10-18 млрд.,
их суммарная масса составляет 21.5 г, а
объем — около 20 см3
(1:27 относительно
объема коры). Если считать, что отростки
одного нейрона имеют длину в среднем 4
см, то длина всех нервных волокон коры
превысит 400 000 км.
У человека толщина коры колеблется от
1.25 до 4 и даже до 6 мм. В глубине борозд
ширина коры уменьшается в 2-2.5 раза по
сравнению с вершиной извилины. Если
принять во внимание, что поверхность
коры в одном полушарии составляет в
среднем 800 см2,
а ее толщина равна в среднем 3 мм, то
объем коры составит 240 см3,
или 44% всего объема полушария. Количество
нейронов коры оценивается в 10-18 млрд.,
их суммарная масса составляет 21.5 г, а
объем — около 20 см3
(1:27 относительно
объема коры). Если считать, что отростки
одного нейрона имеют длину в среднем 4
см, то длина всех нервных волокон коры
превысит 400 000 км.
Учение о строении коры большого мозга, ее архитектонике, имеет несколько разделов. Нейроноархитектоника, или цитоархитектоника, изучает нейрональный состав коры, миелоархитектоника рассматривает ее волокнистое строение, глиоархитектоника занимается глиальными элементами, ангиоархитектоника — распределением в коре сосудов.
В филогенетическом отношении
выделяют древнюю (paleocortex) старую
(archeocortex) и новую (neocortex) кору.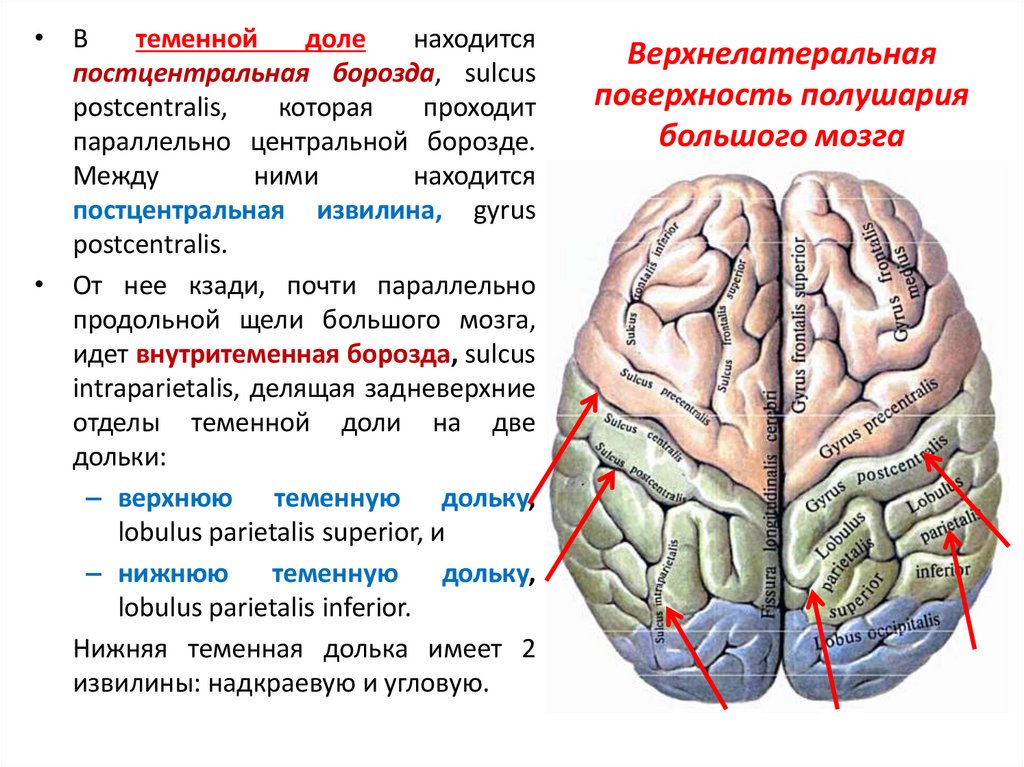 Древняя и
старая кора располагаются на медиальной
и базальной поверхности полушария. Их
окружают межуточные корковые формации,
выделенные под названием перипалеокортекса
и периархикортекса (mesocortex). На долю
древней коры приходится только 0.6% общей
поверхности коры, на долю старой — 2.2%,
на долю промежуточной — 1.6%. В совокупности
это составляет 4.4% поверхности полушария.
Остальные 95.6% поверхности занимает
новая кора.
Древняя и
старая кора располагаются на медиальной
и базальной поверхности полушария. Их
окружают межуточные корковые формации,
выделенные под названием перипалеокортекса
и периархикортекса (mesocortex). На долю
древней коры приходится только 0.6% общей
поверхности коры, на долю старой — 2.2%,
на долю промежуточной — 1.6%. В совокупности
это составляет 4.4% поверхности полушария.
Остальные 95.6% поверхности занимает
новая кора.
На основании эмбрионального
развития древняя, старая и межуточная
кора относятся к гетерогенетической
коре, которая не проходит стадии
шестислойного строения и в окончательном
виде имеет меньшее число слоев. Новая
кора определяется как гомогенетическая
кора. На 6-м месяце внутриутробного
развития ее зачаток подразделяется на
6 слоев. В дальнейшем шестислойность
коры может сохраняться. В таком случае
говорят о гомотипической коре. Если же
после шестислойной стадии количество
слоев увеличивается или уменьшается,
то такую кору называют гетеротипической.
Поверхностный слой
гомотипической гомогенетической коры
носит название молекулярной
пластинки. Она состоит
из сплетения нервных волокон и содержит
немногочисленные горизонтальные
нейроны. Толщина этого слоя – 0.15-0.20 мм.
Второй слой образует
наружная
зернистая пластинка толщиной
0.10-0.16 мм с густо расположенными малыми
зернистыми нейронами. В ней находятся
также малые пирамидные нейроны. Третий
слой называется наружной
пирамидной пластинкой,
его толщина – 0.8-1.0 мм. Для него характерно
наличие средних и больших пирамидных
нейронов с длинными аксонами. Глубже
лежит внутренняя
зернистая пластинка,
которая содержит малые зернистые и
звездчатые нейроны. Ширина этого слоя
– 0.12-0.30 мм. Пятый слой представлен
внутренней пирамидной
пластинкой толщиной
0.4-0.5 мм. Здесь находятся самые большие
пирамидные нейроны с аксонами, выходящими
из пределов коры. Шестой слой составляет
мультиформная пластинка,
в которой располагаются нейроны различной
формы. Ширина ее – 0.6-0.9 мм. Три наружных
слоя коры принято объединять под
названием главной наружной зоны, три
внутренних — под названием главной
внутренней зоны.
Ширина ее – 0.6-0.9 мм. Три наружных
слоя коры принято объединять под
названием главной наружной зоны, три
внутренних — под названием главной
внутренней зоны.
Гетеротипическая кора отличается от гомотипической тем, что внутренняя зернистая пластинка слабо выражена или отсутствует (агранулярная кора). Может отсутствовать внутренняя пирамидная пластинка. В других местах наружная пирамидная, внутренняя зернистая или внутренняя пирамидная пластинки бывают сильно развиты и в них выделяют подслои.
Функциональное значение
пластинок коры определяется их клеточным
составом и межнейрональными связями.
В молекулярной пластинке оканчиваются
волокна из других слоев коры и
противоположного полушария. Здесь
находятся разветвления верхушечных
дендритов пирамидных нейронов, на
которые переключаются импульсы,
приходящие от других корковых нейронов.
Наружная зернистая и наружная пирамидные
пластинки содержат в основном ассоциативные
нейроны, осуществляющие внутрикорковые
связи по горизонтали и вертикали.
Наряду с горизонтальной
организацией коры в форме пластинок в
настоящее время рассматривают принцип
вертикальной модульной организации
коры. Основу модулей составляют такие
конструктивные заготовки как колонки
нейронов и пучки их апикальных дендритов.
Принято считать, что в коре мозга
существуют две разновидности стабильных
генетически обусловленных объединений
нейронов: микро- и макроколонки.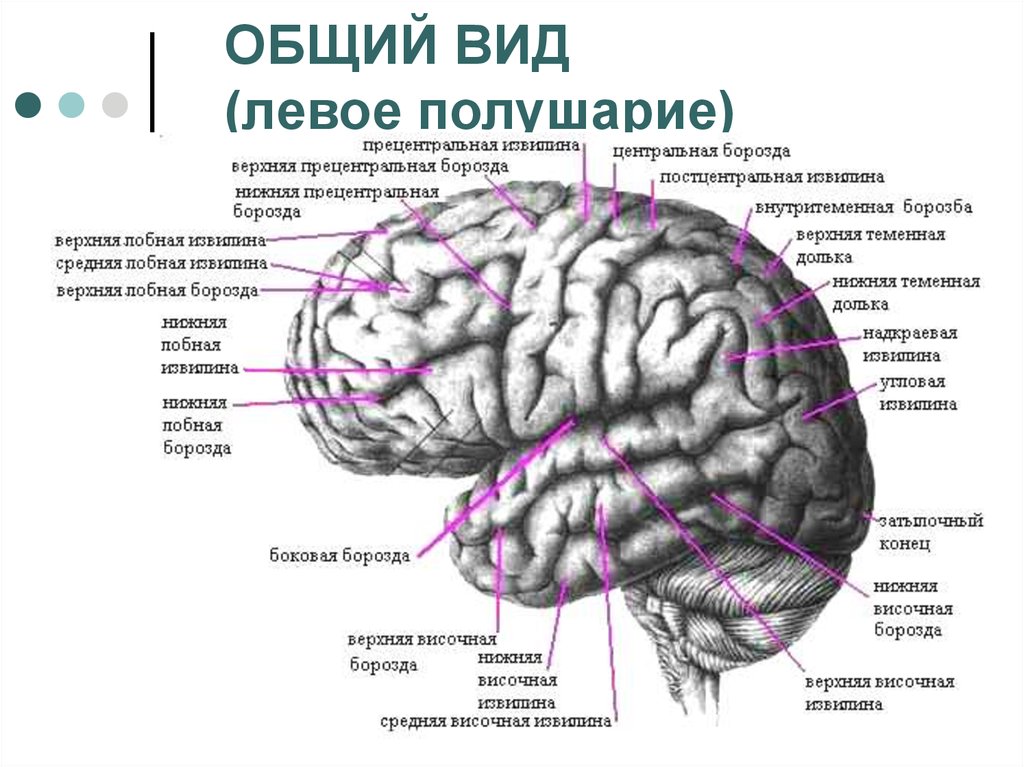
Микроколонка считаются
основной модульной субъединицей в коре.
Она представляет собой вертикально
ориентированный тяж клеток, состоящий
примерно из 110 нейронов и проходящий
через все пластинки коры. Корковые
колонки представляют собой модули,
единицы обработки информации, обладающие
собственным входом и выходом. Диаметр
колонок составляет около 30 мкм. Почти
во всех областях коры количество нейронов
в колонках относительно постоянно, и
только в корковых центрах зрения
количество нейронов в колонках больше.
Несколько сотен микроколонок объединяются
в более крупную структуру — макроколонку,
имеющую диаметр от 500 до 1000 мкм. Корковые
колонки окружены радиально расположенными
нервными волокнами и кровеносными
сосудами. Каждый такой модуль
рассматривается как фокус конвергенции
нескольких тысяч локальных, ассоциативных
и каллозальных волокон. Между корковыми
колонками и подкорковыми образованиями
существуют топографически упорядоченные
нервные связи, отдельным колонкам
соответствуют определенные группы
нейронов в базальных ядрах, таламусе,
коленчатых телах.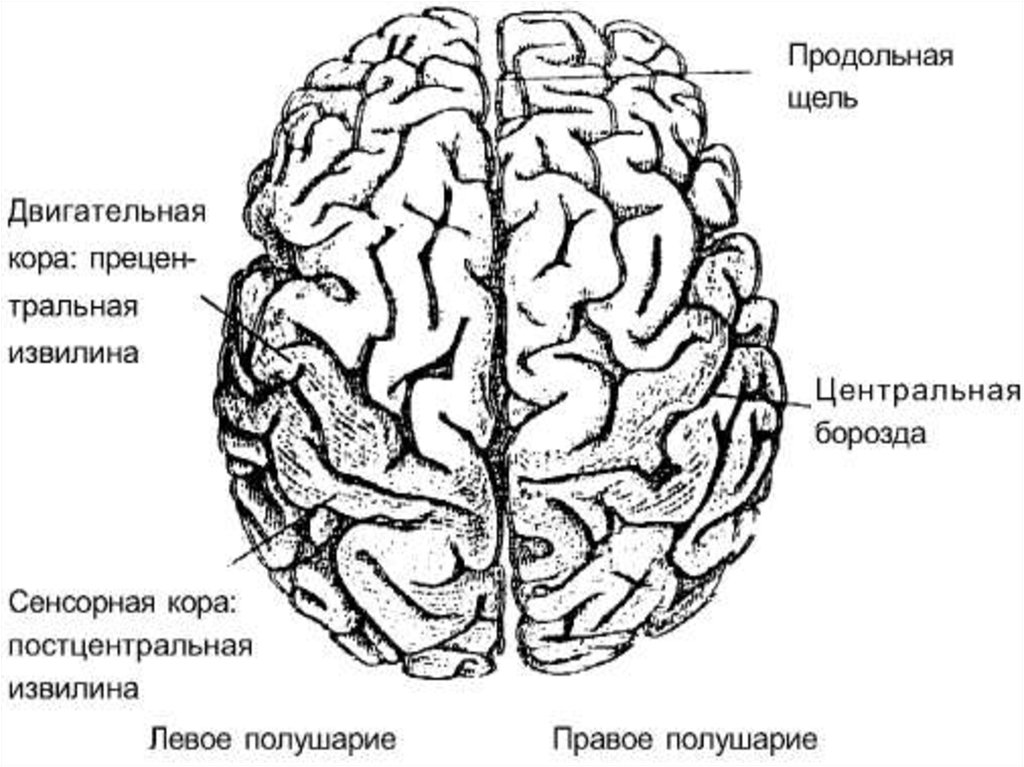
Наиболее простыми и константными объединениями элементов нейронов являются пучки дендритов. Вертикальные пучки дендритов выполняют, по-видимому, основную конструктивную роль в консолидации нейронов. Объединение нейронов различных микроколонок осуществляется, главным образом, аксонными терминалями релейных эфферентных волокон, а макроколонок — ассоциативными и каллозальными волокнами.
Отдельные дендриты в пучке могут на значительном протяжении непосредственно прилегать друг к другу, что создает благоприятные условия для реализации несинаптических влияний обмена ионами и метаболитами. В объединениях нейронов, образованных с помощью пучков дендритов, создаются структурные предпосылки как для дивергенции, так и для конвергенции синаптических импульсов.
С точки зрения миелоархитектоники
в коре выделяют радиальные и тангенциальные
нервные волокна. Первые вступают в кору
из белого вещества, или наоборот, выходят
из коры в белое вещество. Вторые
располагаются параллельно поверхности
коры и образуют на определенной глубине
сплетения, называемые полосками.
Различают полоски молекулярной пластинки,
наружной и внутренней зернистых
пластинок, внутренней пирамидной
пластинки. Волокна, проходящие в полосках,
связывают между собой нейроны соседних
корковых колонок. Число полосок в
различных полях коры неодинаково. В
зависимости от него различают
однополосковый, двухполосковый и
многополосковый тип коры. Особенно
хорошо выражены полоски в затылочной
доле, в зрительных полях (стриарная
кора).
Вторые
располагаются параллельно поверхности
коры и образуют на определенной глубине
сплетения, называемые полосками.
Различают полоски молекулярной пластинки,
наружной и внутренней зернистых
пластинок, внутренней пирамидной
пластинки. Волокна, проходящие в полосках,
связывают между собой нейроны соседних
корковых колонок. Число полосок в
различных полях коры неодинаково. В
зависимости от него различают
однополосковый, двухполосковый и
многополосковый тип коры. Особенно
хорошо выражены полоски в затылочной
доле, в зрительных полях (стриарная
кора).
|
В нервной системе особенно
ясно выступает принцип единства строения
и функции. В свое время И.П.Павлов
сформулировал применительно к нервной
системе принцип структурности, по
которому все мельчайшие детали строения
мозга имеют динамические (функциональное)
значение.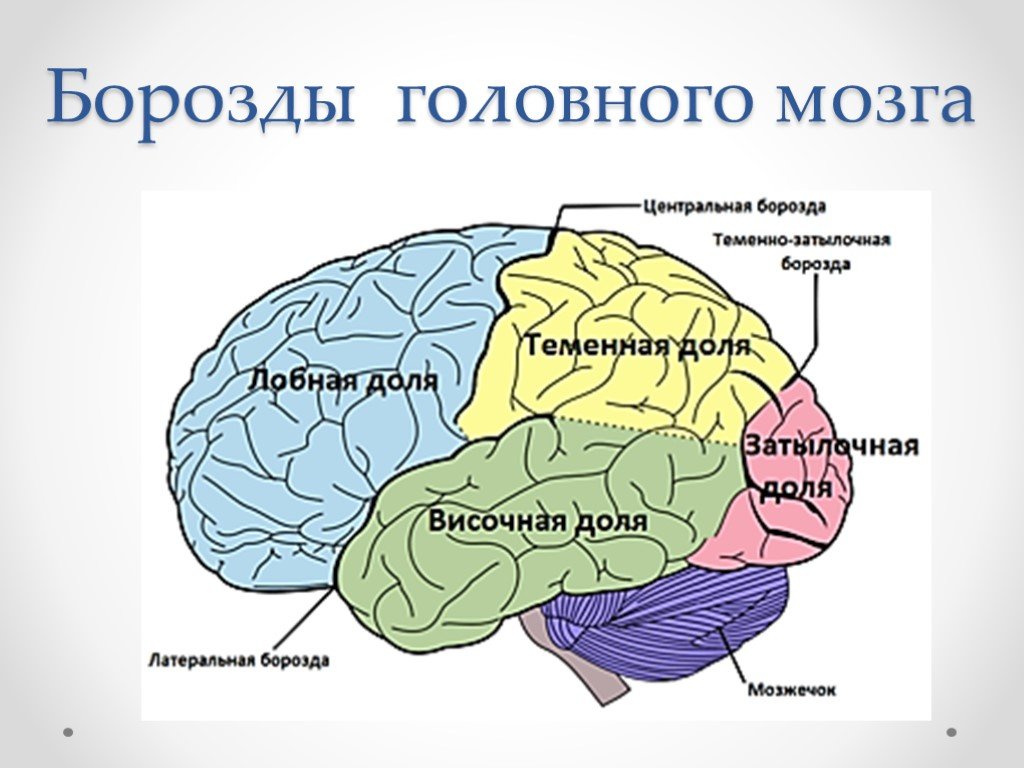 Исходя из этого, нужно признать,
что особенности архитектоники коры
большого мозга, различия в структуре
ее областей и полей связаны с их
функциональными отправлениями.
Исходя из этого, нужно признать,
что особенности архитектоники коры
большого мозга, различия в структуре
ее областей и полей связаны с их
функциональными отправлениями.
В структурно-функциональном отношении кора большого мозга может быть разделена на передний (лобная доля) и задний (затылочная, теменная и височная доли) отделы. Граница между ними проходит по центральной борозде. Задний отдел осуществляет восприятие афферентных сигналов. Расположенные здесь корковые поля неравноценны в функциональном отношении, и их можно разделить на первичные, вторичные и третичные.
Первичные поля коры
представляют собой четко отграниченные
участки, которые соответствуют центральным
частям анализаторов. В эти поля проходит
по специфическим проекционным афферентным
путям основная масса сигналов от органов
чувств. Первичные поля характеризуются
сильным развитием внутренней зернистой
пластинки. Первичные поля связаны с
реле-ядрами таламуса и ядрами коленчатых
тел.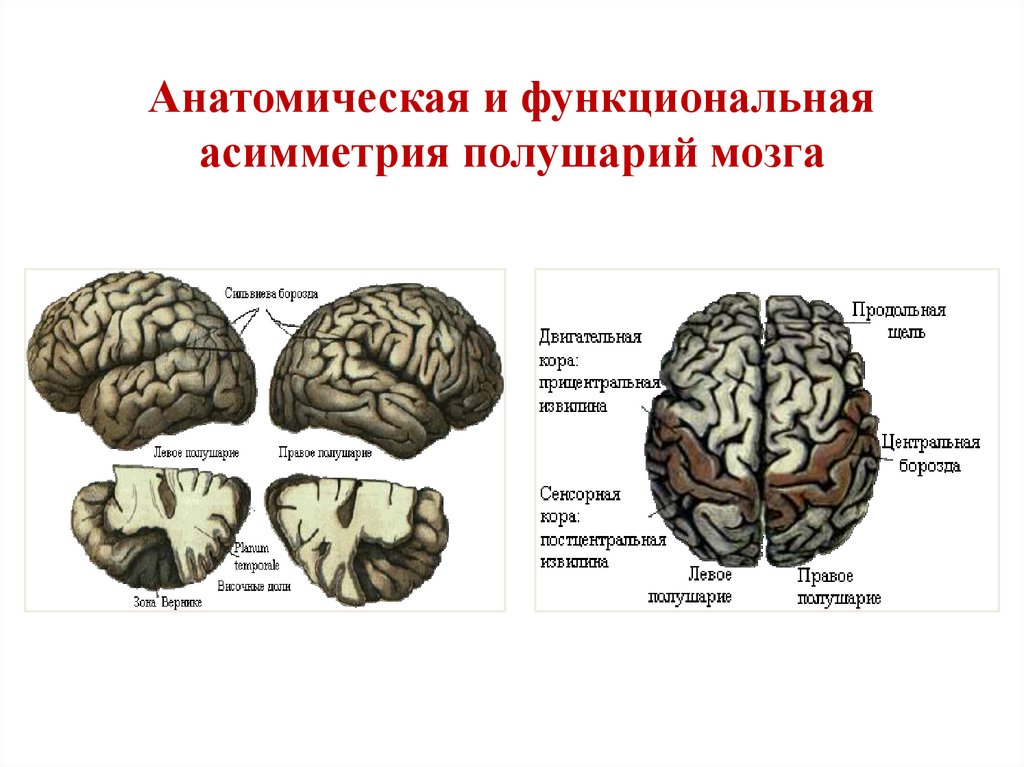 Они имеют экранную структуру и, как
правило, жесткую соматотопическую
проекцию, при которой отдельные участки
периферии проецируются в соответствующие
им участки коры. Повреждение первичных
полей коры сопровождается нарушением
непосредственного восприятия и тонкой
дифференцировки раздражений.
Они имеют экранную структуру и, как
правило, жесткую соматотопическую
проекцию, при которой отдельные участки
периферии проецируются в соответствующие
им участки коры. Повреждение первичных
полей коры сопровождается нарушением
непосредственного восприятия и тонкой
дифференцировки раздражений.
Представительство кожной
и сознательной проприоцептивной
чувствительности находится первичных
корковых полях (1, 2, 3), занимающих
постцентральную извилину. В каждом
полушарии имеется обратная проекция
поверхности противоположной половины
тела. В верхней части извилины находится
проекция нижней конечности, ниже
располагается проекция живота, груди,
еще ниже проецируется нижняя конечность.
Самую нижнюю часть постцентральной
извилины занимают зоны, связанные с
иннервацией головы и шеи, но проекция
частей лица является не обратной, а
прямой. Данные изучения колонковой
организации коры свидетельствуют о
том, что каждый сегмент тела (дерматом)
проецируется на кору в виде узкой
полоски, идущей спереди назад через все
цитоархитектонические поля постцентральной
коры; при этом к колонкам поля 1 подходят
афферентные волокна от кожи, к полю 2 —
от суставов и к полю 3 — от мышц.
Первичное зрительно поле (17) расположено на медиальной поверхности полушария вдоль шпорной борозды. Здесь хорошо развита внутренняя зернистая пластинка, которая подразделяется посредством белых полосок на 3 части. Корковые колонки образуют чередующиеся вертикальные пластинки для правого и левого глаза. Полагают, что нейроны глубоких слоев коры обладают свойствами «анализатора движения», а нейроны поверхностных слоев действуют как « анализатор формы».
Первичные слуховые поля (41, 42) локализуются в поперечных височных извилинах (Гешля) и заходят в верхнюю височную извилину. В этих полях представлены по порядку участки улитки, воспринимающие различные звуковые частоты. Деление на колонки выражено в слуховой коре наиболее отчетливо.
Первичные обонятельные поля находятся в археокортексе, покрывающем обонятельный треугольник, переднее продырявленное вещество, прозрачную перегородку и подмозолистое поле.
Первичное вкусовое поле
располагается, по мнению большинства
исследователей, в нижнем участке
постцентральной извилины, в глубине
латеральной борозды, и соответствует
проекции языка.
Корковый конец вестибулярного анализатора, по данным различных авторов, имеет представительство в средней височной извилине (поле 21), верхней теменной дольке, верхней височной извилине.
Представительство в коре внутренних органов изучено недостаточно, по-видимому, оно имеет более диффузный характер. Важная роль в регуляции функций внутренностей отводится лимбической области коры, в которую входит поясная и парагиппокампальная извилины, гиппокамп, прозрачная перегородка и подмозолистое поле. Лимбическая кора вместе с подкорковыми образованиями (миндалевидное тело, ядро поводков, ядра сосцевидных тел) составляет лимбическую систему, которая представляет субстрат эмоций и реакций, связанных с основными биологическими влечениями (голод, жажда, страх и т.д.).
Вторичные поля корыпримыкают к первичным полям. Их можно
рассматривать как периферические части
корковых анализаторов. Эти поля связаны
с ассоциативными ядрами таламуса.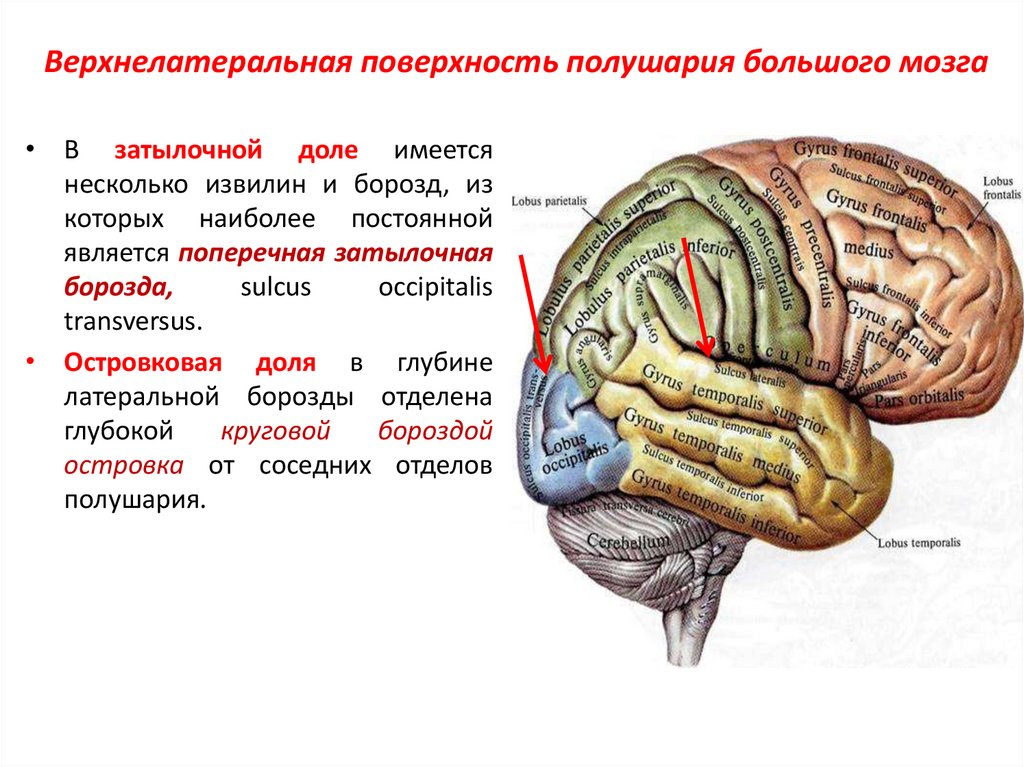 При
поражении вторичных полей сохраняются
элементарные ощущения, но нарушается
способность к более сложным восприятиям.
Вторичные поля не имеют четких границ,
в них не выражена соматотопическая
проекция.
При
поражении вторичных полей сохраняются
элементарные ощущения, но нарушается
способность к более сложным восприятиям.
Вторичные поля не имеют четких границ,
в них не выражена соматотопическая
проекция.
Вторичное поле общей чувствительности локализуется в верхней теменной дольке (поля 5, 7). Вторичные зрительные поля (18, 19) занимают медиальную поверхность затылочной доли и большую часть латеральной поверхности. Вторичное слуховое поле (22) находится в верхней и средней височных извилинах. Вторичные обонятельные и вкусовые поля локализуются в парагиппокампальной извилине и крючке (поля 28, 34).
Третичные поля коры
отличаются наиболее тонкой нейронной
структурой и преобладанием ассоциативных
элементов. Они занимают всю нижнюю
теменную дольку и часть верхней теменной
дольки, а также затылочно-височно-теменную
область. Эти поля связаны с задними
ядрами таламуса. В третичных полях
осуществляются наиболее сложные
взаимодействия анализаторов, лежащие
в основе познавательного процесса
(гнозия), формируются программы
целенаправленных действий (праксия).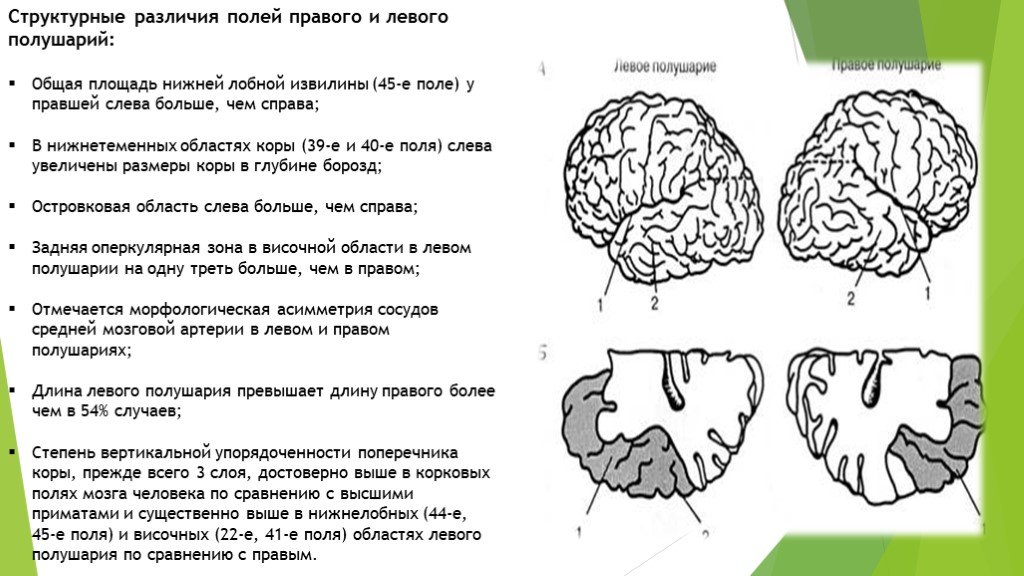
Кора височной доли имеет отношение к хранению и воспроизведению впечатлений. При электрическом раздражении некоторых точек височной коры наблюдаются своеобразные реакции в форме «вспышек пережитого» или ощущения «уже виденного». Полагают, что в коре височных долей создается нейронная запись потока сознания, она хранится неопределенно долгое время, но не может произвольно воспроизводиться, а «оживает» лишь при искусственной стимуляции и некоторых болезненных состояниях.
|
Передний отдел полушария
имеет отношение к организации действий
и также подразделяется на первичные,
вторичные и третичные корковые поля.
Первичное двигательной поле (4)
располагается в предцентральной
извилине.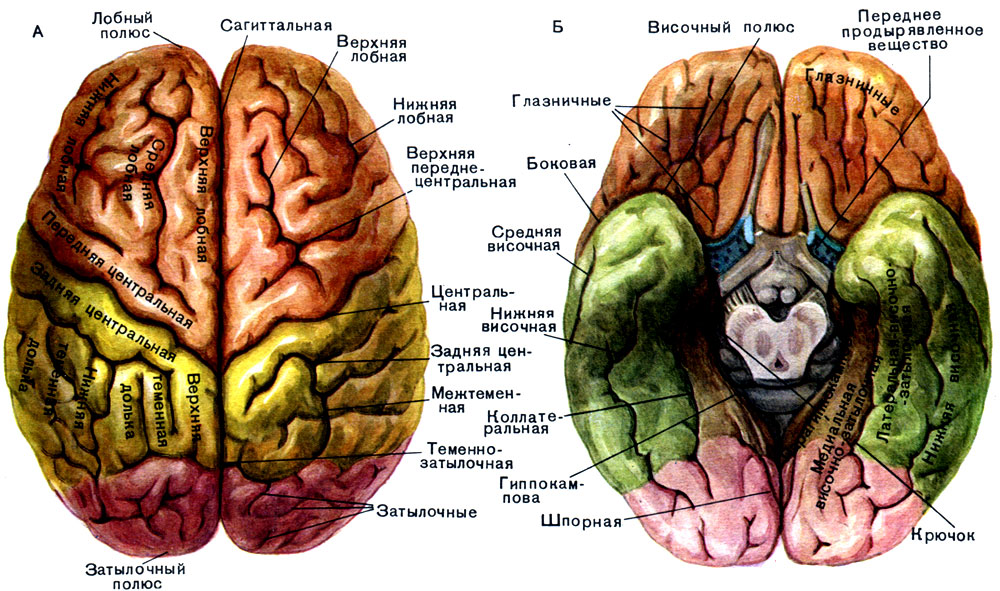 Здесь отсутствует внутренняя
зернистая пластинка (агранулярная кора)
и особенно сильно развита внутренняя
пирамидная пластинка с гигантскими
пирамидными нейронами Беца. Аксоны этих
нейронов образуют пирамидный путь. На
клетки Беца непосредственно переключаются
импульсы, поступающие из мозжечка через
центральное медиальное ядро таламуса.
В первичном двигательном поле вся
мускулатура тела представлена в обратной
проекции, как и кожный покров в
постцентральной извилине. Кора здесь
разделена на колонки, которые связаны
с определенными группами двигательных
нейронов спинного мозга и управляют
движением отдельных мышц или мышечных
групп.
Здесь отсутствует внутренняя
зернистая пластинка (агранулярная кора)
и особенно сильно развита внутренняя
пирамидная пластинка с гигантскими
пирамидными нейронами Беца. Аксоны этих
нейронов образуют пирамидный путь. На
клетки Беца непосредственно переключаются
импульсы, поступающие из мозжечка через
центральное медиальное ядро таламуса.
В первичном двигательном поле вся
мускулатура тела представлена в обратной
проекции, как и кожный покров в
постцентральной извилине. Кора здесь
разделена на колонки, которые связаны
с определенными группами двигательных
нейронов спинного мозга и управляют
движением отдельных мышц или мышечных
групп.
Вторичные двигательные
поля (6, 8) находятся кпереди от
предцентральной извилины. Они
характеризуются сильным развитием
наружной и внутренней пирамидных
пластинок, в которых преобладают большие
пирамидные нейроны. Во вторичные поля
передаются сигналы из мозжечка.
Эфферентные волокна идут отсюда к ядрам
полосатого тела. Таким образом, вторичные
двигательные поля имеют отношение к
экстрапирамидной системе, их функция
необходима для выполнения сложных
стереотипных двигательных актов.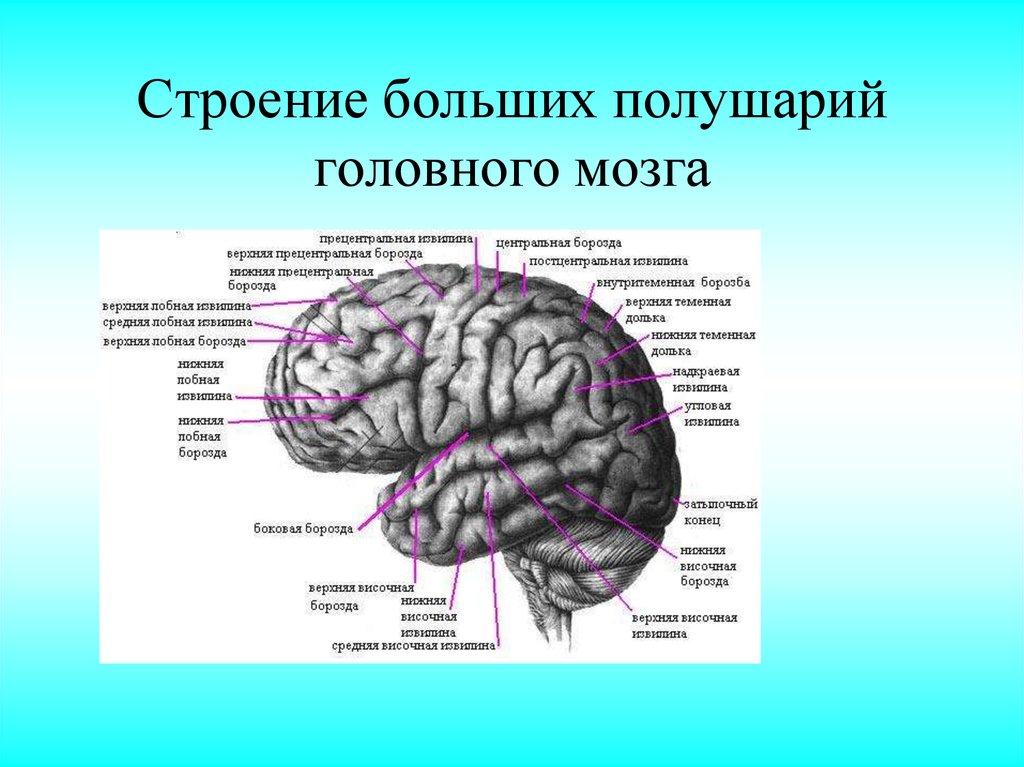 Первичные и вторичные двигательные
поля имеют богатые связи с задним отделом
полушария. Обратная связь между аппаратом
движения и корой осуществляется через
мозжечок, который воспринимает
проприоцептивные раздражения и после
соответствующей переработки передает
их в кору большого мозга.
Первичные и вторичные двигательные
поля имеют богатые связи с задним отделом
полушария. Обратная связь между аппаратом
движения и корой осуществляется через
мозжечок, который воспринимает
проприоцептивные раздражения и после
соответствующей переработки передает
их в кору большого мозга.
Третичные поля занимают большую часть лобной доли, на них приходится около 1/4 всей поверхности коры. Здесь хорошо выражена внутренняя зернистая пластинка, к нейронам которой идут волокна из медиальных ядер таламуса. Третичные поля лобной коры связаны с высшими формами целенаправленной деятельности и играют важную роль в социальном поведении. При их поражении не нарушается ощущение или движение, но человек становится пассивным, не может оценивать происходящие события и свое поведение, теряет способность к предвидению.
Важнейшую особенность
человека составляет членораздельная
речь. Академик И.П.Павлов относил речь
ко второй сигнальной системе, с помощью
которой происходит непрямое отражение
действительности.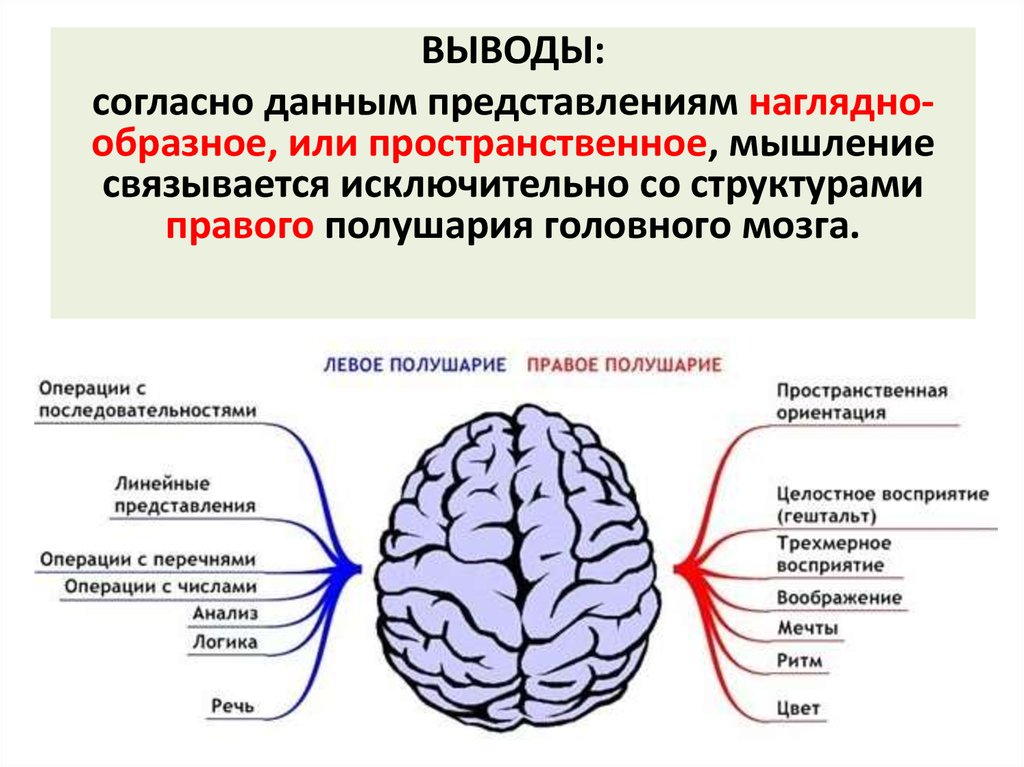 Речевые функции имеют
широкое представительство в коре
большого мозга. На основании данных,
полученных при электрическом раздражении
и удалении у больных различных участков
коры, выделены три корковых речевых
поля. Заднее речевое поле располагается
в затылочно-височно-теменной области,
захватывая все три височные, надкраевую
и угловую извилины. Это поле связано
преимущественно с восприятием и
пониманием речи и функционально является
ведущим. При его поражении всегда
наступает расстройство речи — афазия.
Переднее речевое поле лежит в задней
части нижней лобной извилины и
соответствует моторному центру речи
Брока. Верхнее, дополнительное, речевое
поле локализуется у верхнего края
полушария кпереди от предцентральной
извилины, при его поражении не всегда
наблюдаются расстройства речи. Речевые
поля, как другие части коры, связаны с
ядрами таламуса. Заднее поле связано с
задним ядром, верхнее поле — с латеральным
ядром, переднее поле — с медиальными
ядрами. Все речевые поля связаны
ассоциативными путями в единую
функциональную систему.
Речевые функции имеют
широкое представительство в коре
большого мозга. На основании данных,
полученных при электрическом раздражении
и удалении у больных различных участков
коры, выделены три корковых речевых
поля. Заднее речевое поле располагается
в затылочно-височно-теменной области,
захватывая все три височные, надкраевую
и угловую извилины. Это поле связано
преимущественно с восприятием и
пониманием речи и функционально является
ведущим. При его поражении всегда
наступает расстройство речи — афазия.
Переднее речевое поле лежит в задней
части нижней лобной извилины и
соответствует моторному центру речи
Брока. Верхнее, дополнительное, речевое
поле локализуется у верхнего края
полушария кпереди от предцентральной
извилины, при его поражении не всегда
наблюдаются расстройства речи. Речевые
поля, как другие части коры, связаны с
ядрами таламуса. Заднее поле связано с
задним ядром, верхнее поле — с латеральным
ядром, переднее поле — с медиальными
ядрами. Все речевые поля связаны
ассоциативными путями в единую
функциональную систему.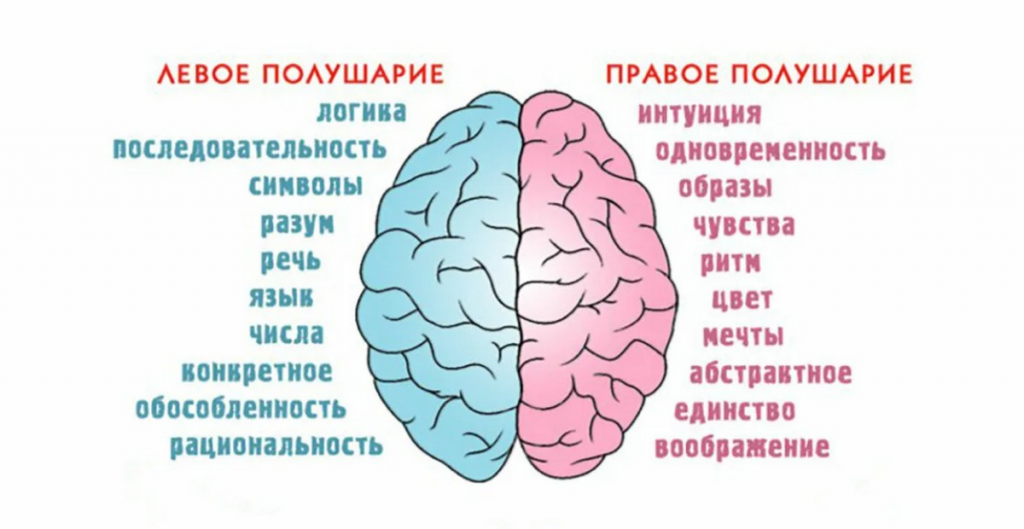
Особенностью речевых центров коры является их асимметрия. У большинства людей они локализуются в левом полушарии, которое является доминантным в отношении речи. Принято считать, что эта доминантность связана с праворукостью, и что у левшей речью управляет правое полушарие. В последнее время вопрос о функциональной асимметрии полушарий трактуется более широко. С левым полушарием связывают речь и абстрактное мышление, а с правым полушарием — пространственное представление, образное мышление, музыкальные способности.
|
Карта сайта
Карта сайта
|
|
 Открыл конференцию известный врач Рошаль Леонид Михайлович. Конференция посвящена 80-летию Дроздова Эдуарда Семеновича
Открыл конференцию известный врач Рошаль Леонид Михайлович. Конференция посвящена 80-летию Дроздова Эдуарда СеменовичаПолушария головного мозга: Топография | Кенхаб
Автор:
Лоренцо Крамби MBBS, BSc
•
Рецензент:
Димитриос Митилинаиос, доктор медицины, доктор философии
Последнее рассмотрение: 22 июля 2022 г.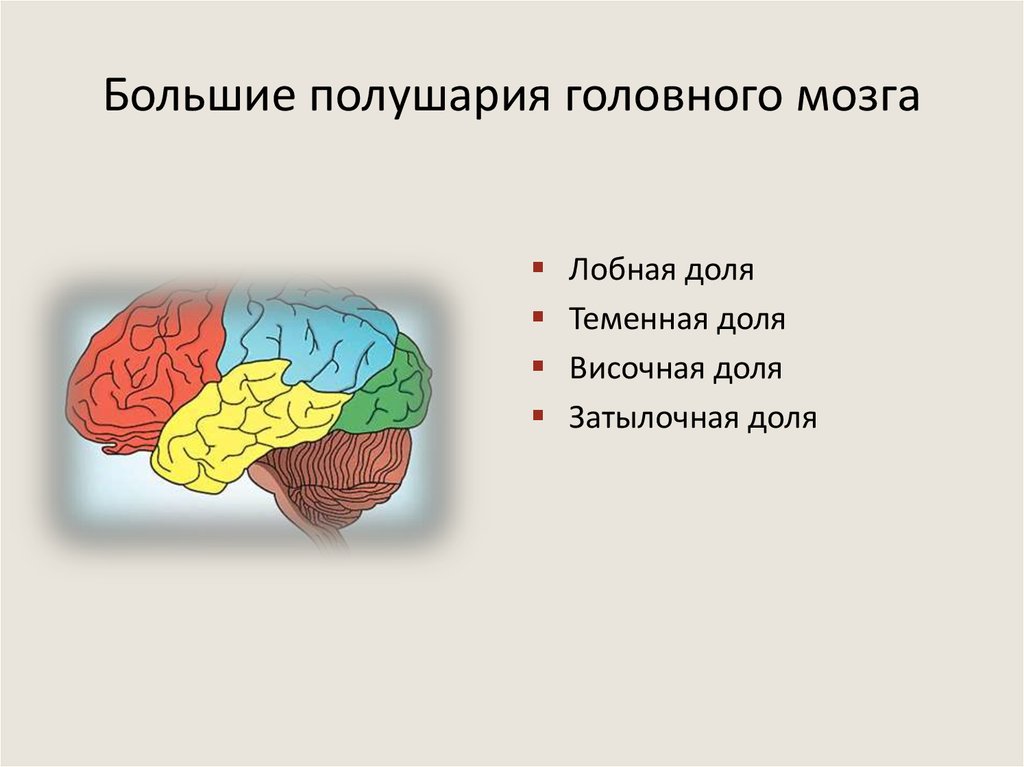
Время считывания: 11 минут
Головной мозг — самая большая часть головного мозга, охватывающая все три черепные ямки. Составные части можно классифицировать на основе их эмбриологического происхождения, функциональных ролей или их топографии. В этой статье мы рассмотрим организацию мозга по отношению к последнему. Кроме того, в статье головной мозг будет рассмотрен с разных точек зрения, так как некоторые структуры видны только под определенным углом.
Во-первых, головной мозг делится на два полушария — левое и правое — серпом большого мозга (нижняя проекция твердой мозговой оболочки, содержащая верхний и нижний сагиттальные синусы) вдоль продольной мозговой щели . Затем каждое полушарие можно разделить на долей , названных в соответствии с костями черепа, под которыми они расположены. Из этого правила есть одно исключение, о котором мы поговорим позже.
В процессе развития передний мозг претерпевает значительные складки.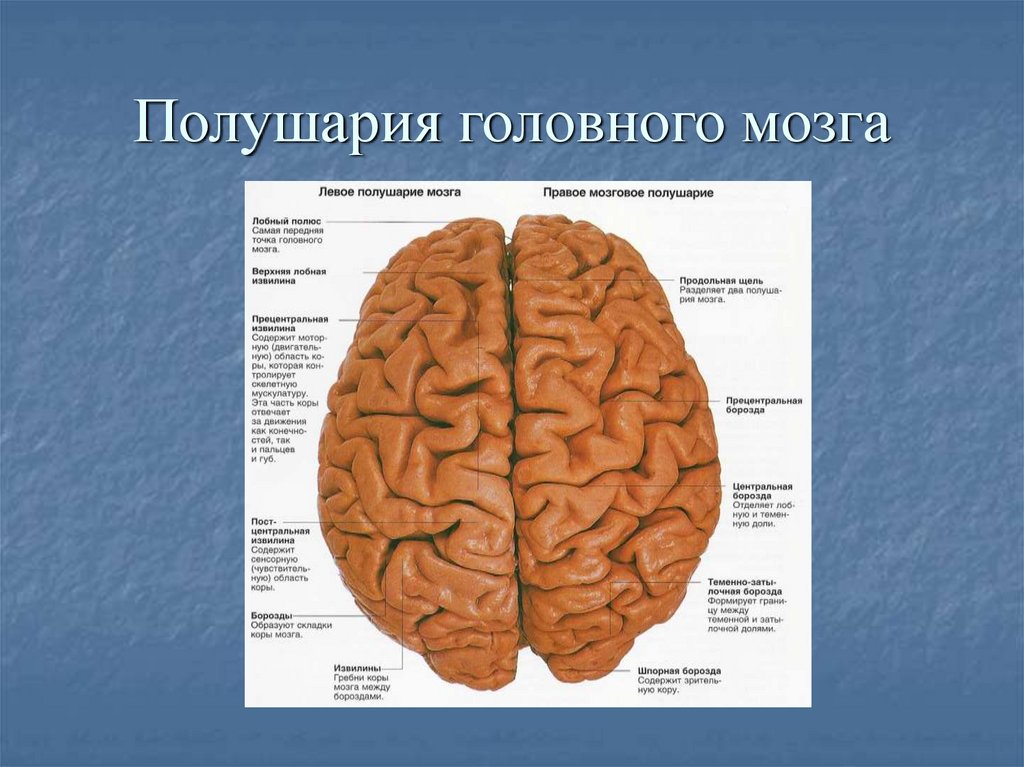 Это привело к тому, что поверхность коры головного мозга была организована в виде множества холмов и долин. Холмы известны как извилины (дольки) и долины борозды (или трещины ). Следует отметить, что мягкая мозговая оболочка (самый внутренний менингеальный слой) тесно связана с мозговой тканью и следует за ней в борозды.
Это привело к тому, что поверхность коры головного мозга была организована в виде множества холмов и долин. Холмы известны как извилины (дольки) и долины борозды (или трещины ). Следует отметить, что мягкая мозговая оболочка (самый внутренний менингеальный слой) тесно связана с мозговой тканью и следует за ней в борозды.
Наоборот, паутинная оболочка (средний менингеальный слой) покрывает извилины и покрывает борозды (не заходя в углубление).
Содержимое
-
Вид сбоку
- Бороздки и доли
- Лобная доля
- Височная доля
- Теменная доля
- Островок
- Затылочная доля
-
Медиальный вид
- Борозды и извилины
- Промежуточный мозг
-
Вид снизу
- Лобная доля
- Височная доля
-
Основные моменты
- Вид сбоку
- Медиальный вид
- Вид снизу
- Источники
+ Показать все
Поперечный срез головного мозга с выделением серпа головного мозгаВид сбоку
Бороздки и доли
Знание борозд помогает в идентификации различных долей. Самая большая борозда — это центральная борозда Роландо . Это срединная структура, которая отделяет лобной доли (расположенной спереди) от теменной доли .
Самая большая борозда — это центральная борозда Роландо . Это срединная структура, которая отделяет лобной доли (расположенной спереди) от теменной доли .
латеральная борозда Сильвиева (или сильвиева борозда) представляет собой нижнелатеральную борозду, которая отделяет височная доля от лобной и теменной долей (обе выше борозды).
Презатылочная вырезка представляет собой задненижнюю точку, которая разграничивает переход от височной доли к затылочной доле . Воображаемая линия от презатылочной вырезки до теменно-затылочной борозды также помогает определить границу между теменной и затылочной долями. Горизонтальная воображаемая линия, идущая от верхней части сильвиевой щели до пересечения с ранее описанной линией, также поможет обозначить переход от теменной доли к височной доле.
Лобная доля
Лобная доля занимает переднюю черепную ямку. Она состоит из четырех извилин — верхней, средней, нижней и прецентральной, которые отделены друг от друга верхней лобной, нижней лобной и предцентральной бороздами. верхняя лобная извилина расположена краниально по отношению к верхней лобной борозде .
Ниже этой борозды находится средняя лобная извилина , а каудально от нее находится нижняя лобная борозда и извилина . Все три извилины находятся впереди вертикальной прецентральной борозды и извилины (извилина находится непосредственно впереди центральной борозды Роландо).
нижняя лобная извилина далее подразделяется на три части передней и восходящей ветвями латеральной сильвиевой борозды. Кпереди от передней ветви находится pars orbitalis нижней лобной извилины.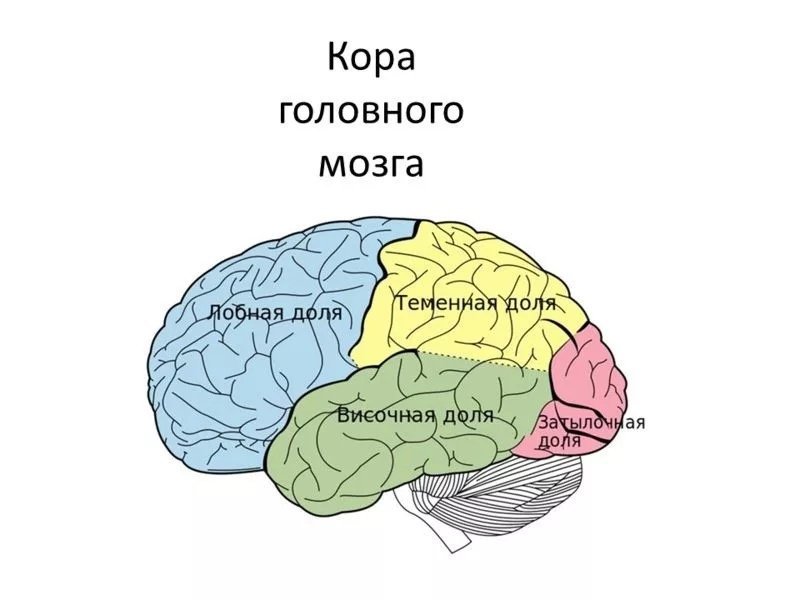 Между передней ветвью и восходящей ветвью находится треугольная часть . Наконец, кзади от восходящей ветви находится pars operculum , которая также ограничена сзади задней ветвью сильвиевой борозды .
Между передней ветвью и восходящей ветвью находится треугольная часть . Наконец, кзади от восходящей ветви находится pars operculum , которая также ограничена сзади задней ветвью сильвиевой борозды .
Височная доля
Височная доля находится в средней черепной ямке. Имеет три извилины и две борозды. Верхняя височная извилина расположена каудальнее Сильвиевой борозды, но краниальнее верхней височной борозды . средняя височная извилина лежит между верхней и нижней височными бороздами ; в то время как нижняя височная извилина ниже последней борозды.
Теменная доля
Теменная доля также имеет три извилины и две борозды. Центральная борозда Роландо является его передней границей постцентральной извилины . Кзади от постцентральной извилины находится постцентральная борозда .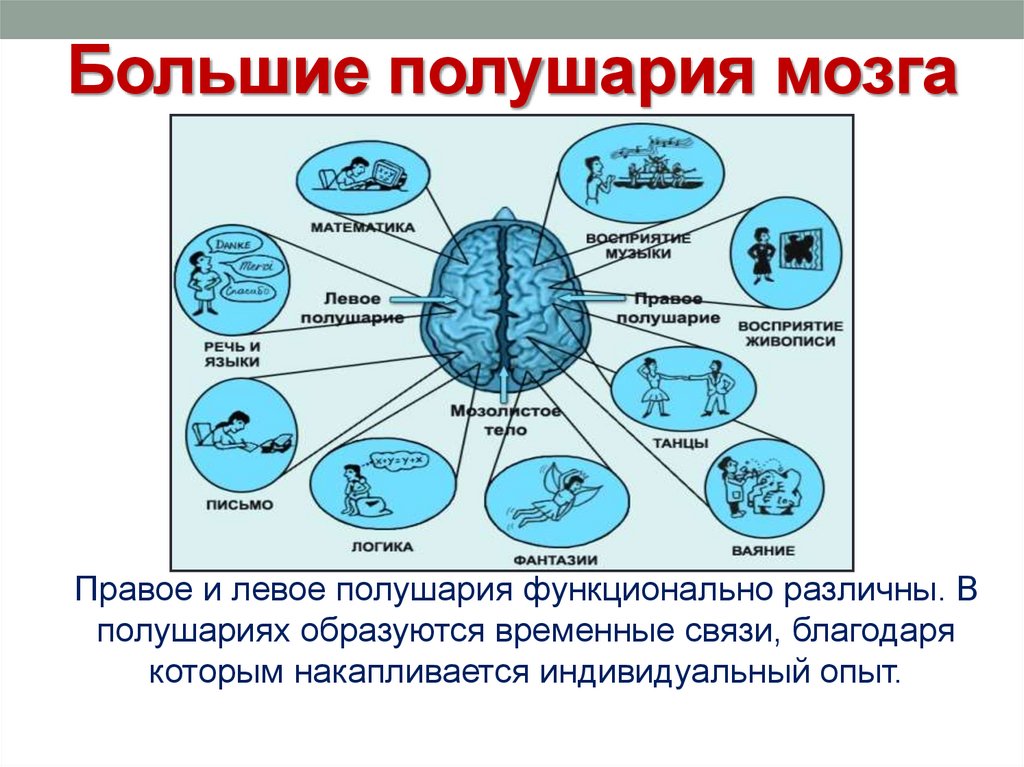 Внутритеменная борозда (примерно перпендикулярно постцентральной борозде) идет к затылочной доле с верхняя теменная долька вверху и нижняя теменная долька внизу. Последний далее подразделяется на надмаргинальную извилину спереди и угловую извилину сзади.
Внутритеменная борозда (примерно перпендикулярно постцентральной борозде) идет к затылочной доле с верхняя теменная долька вверху и нижняя теменная долька внизу. Последний далее подразделяется на надмаргинальную извилину спереди и угловую извилину сзади.
Постцентральная извилина теменной доли, прецентральная извилина лобной доли и парацентральная извилина (на медиальной поверхности головного мозга) вместе обозначаются как центральная доля головного мозга . Это не официальный термин, который можно найти в ТА, но в последнее время все больше и больше нейробиологов считают, что каждая из этих сенсомоторных областей коры имеет свои особенности и должна рассматриваться как отдельная доля. В Kenhub мы следуем более официальной терминологии, которая встречается в классических учебниках по нейроанатомии, и мы не описываем эту часть мозга как отдельную долю.
Инсула
Если верхний и нижний края сильвиевой щели отведены соответственно вверх и вниз, можно наблюдать пятую долю. Островок (или остров Рейля) представляет собой глубокую долю головного мозга, которая имеет две борозды и две извилины. Имеется короткая извилина , расположенная спереди, и длинная извилина сзади. Между обеими извилинами находится центральная борозда островка . Круговая борозда проходит вдоль медиальных отделов покрышек лобной, теменной, глазничной и височной долей.
Островок (или остров Рейля) представляет собой глубокую долю головного мозга, которая имеет две борозды и две извилины. Имеется короткая извилина , расположенная спереди, и длинная извилина сзади. Между обеими извилинами находится центральная борозда островка . Круговая борозда проходит вдоль медиальных отделов покрышек лобной, теменной, глазничной и височной долей.
Затылочная доля
Затылочная доля является самой задней из пяти долей. Она занимает заднюю черепную ямку и имеет в своем веществе несколько борозд. Имеется небольшая боковая проекция шпорной борозды , поперечной затылочной борозды и непостоянной полулунной борозды . Извилины затылочной доли лучше видны при медиальном осмотре головного мозга.
Медиальный вид
На сагиттальном срезе вдоль продольной мозговой щели открывается медиальная поверхность головного мозга.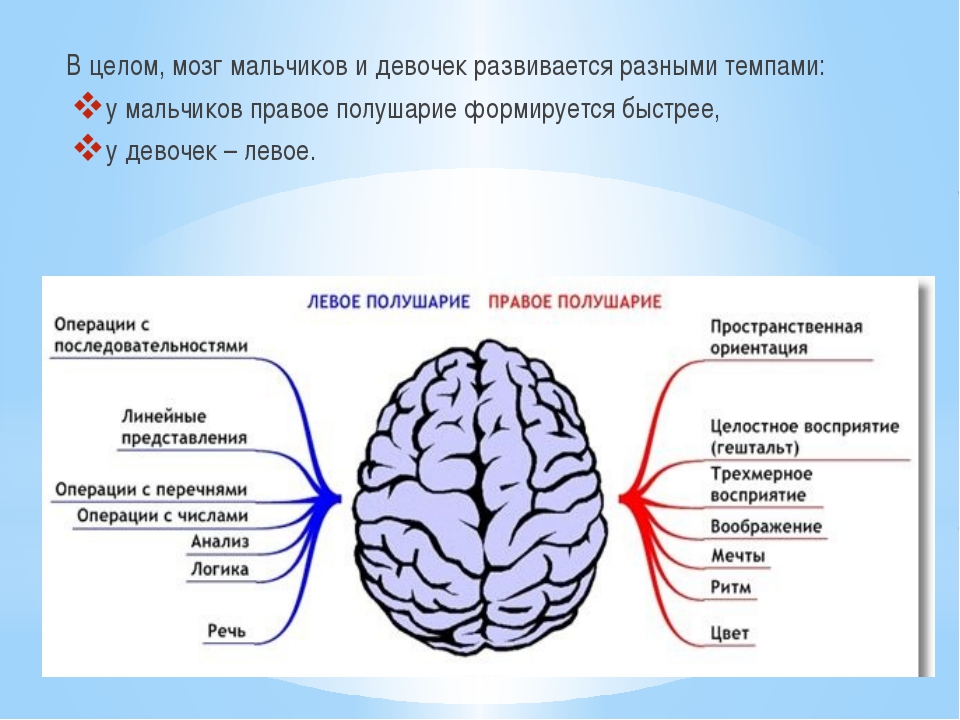 На этом ракурсе лучше видны дополнительные извилины и борозды ранее упомянутых долей.
На этом ракурсе лучше видны дополнительные извилины и борозды ранее упомянутых долей.
борозды и извилины
Книзу от колена мозолистого тела и кпереди от терминальной пластинки гипоталамуса находится подмозолистая извилина . поясная извилина начинается непосредственно перед парольфакторной областью (ниже колена мозолистого тела) и повторяет контур комиссурального тела и мозолистая борозда . Поясная борозда выше поясной извилины и отделяет ее от медиальной лобной извилины.
медиальная лобная извилина ограничена сзади парацентральной бороздой . Кзади от парацентральной дольки (которая находится за парацентральной бороздой и содержит небольшой медиальный выступ центральной борозды Роландо) поясная борозда идет вверх и становится маргинальной бороздой . Долька сразу позади маргинальной борозды — это предклинье . Сзади от cuneus отделяется теменно-затылочной бороздой.
Сзади от cuneus отделяется теменно-затылочной бороздой.
шпорная борозда проходит горизонтально от затылочного полюса до пересечения с теменно-затылочной бороздой непосредственно позади венозного соединения большой вены Галена и прямого синуса. Книзу шпорной борозды находится язычная извилина затылочной доли, которая упирается в мозговую поверхность намета мозжечка (примерно горизонтальная проекция твердой мозговой оболочки).
Промежуточный мозг
Медиальная проекция также показывает диэнцефальные структуры головного мозга. Можно наблюдать, как мозолистое тело изгибается в переднезаднем направлении, глубоко внутри головного мозга. Под ним находится septum pellucidum, соединяющая последний со сводом спереди.
Медиальная стенка таламуса и его межталамическая спайка расположены каудальнее свода и сосудистого сплетения третьего желудочка.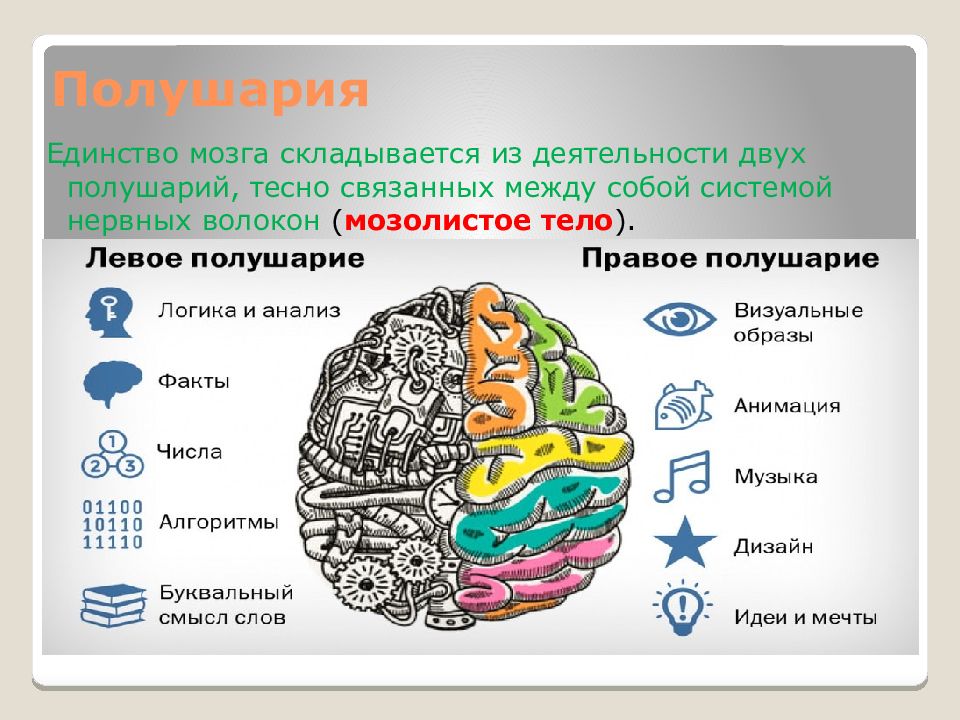 гипоталамическая борозда , отмечающая границу между таламусом и гипоталамусом , также видна в переднезаднем направлении от передней комиссуры до отверстия водопровода головного мозга. Ниже гипоталамуса находится перекрест зрительных нервов и гипофиз большого мозга . Наконец, позади таламуса находится эпиталамус (габенулярное ядро и шишковидная железа).
гипоталамическая борозда , отмечающая границу между таламусом и гипоталамусом , также видна в переднезаднем направлении от передней комиссуры до отверстия водопровода головного мозга. Ниже гипоталамуса находится перекрест зрительных нервов и гипофиз большого мозга . Наконец, позади таламуса находится эпиталамус (габенулярное ядро и шишковидная железа).
Нижняя проекция
Лобная доля
Нижний вид мозга показывает больше деталей лобных, височных и затылочных долей. На лобной доле (от медиальной к латеральной) имеется прямая извилина (прямая извилина) между продольной мозговой щелью и обонятельной бороздой . Обонятельная борозда, расположенная латеральнее прямой извилины, обеспечивает путь обонятельному тракту к месту назначения. Есть также две орбитальные борозды 9.0010 и две орбитальные извилины в нижней части лобной доли.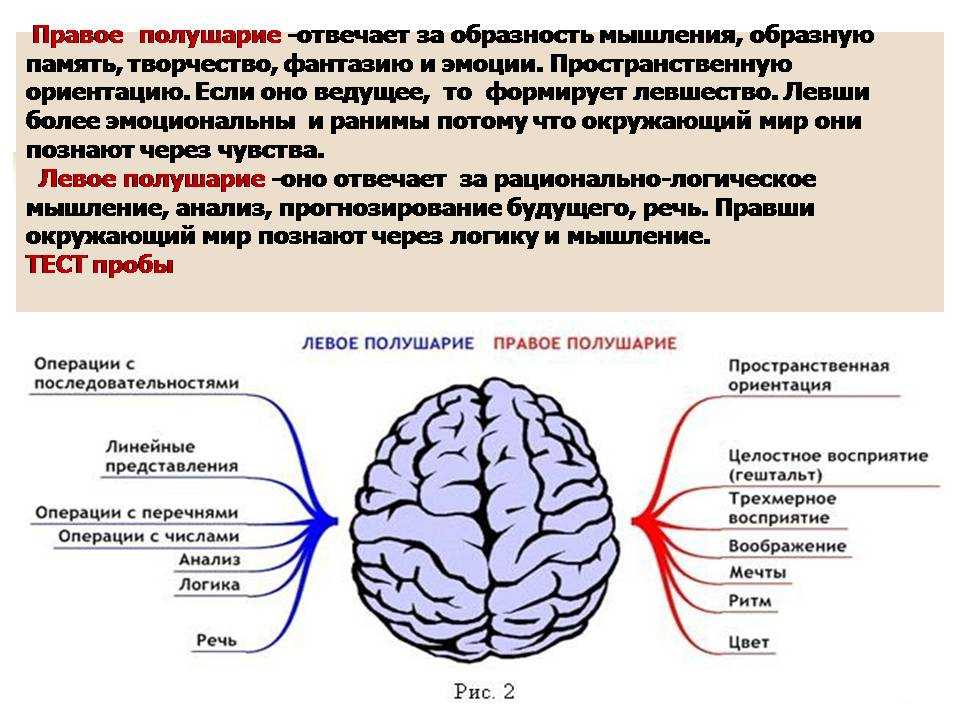
Височная доля
Ункус височной доли располагается латеральнее гипофиза большого мозга, бугра синего, сосцевидных тел и заднего продырявленного вещества (в межножковой ямке). Он также расположен кзади от переднего продырявленного вещества и кпереди от ножки большого мозга и латерального коленчатого тела таламуса. Прилегает к парагиппокампальной извилине. парагиппокампальная извилина латерально ограничена ринальной бороздой спереди и коллатеральной бороздой сзади. Он также продолжается сзади в виде медиальной затылочно-височной извилины . Коллатеральная борозда служит медиальной границей для латеральной затылочно-височной извилины , которая ограничена латерально затылочно-височной бороздой .
Основные моменты
Вид сбоку
- Центральная борозда Роландо
- Боковая сильвиева борозда
- Теменно-затылочная борозда
Лобная доля
- Верхняя/Средняя/Нижняя лобные извилины и борозды
- Прецентральная борозда и извилина
Височная доля
- Верхняя/средняя/нижняя височные извилины и борозды
Теменная доля
- Постцентральная борозда и извилина
- Внутритеменная борозда
- Верхняя/нижняя теменные дольки
- Супрамаргинальная извилина
- Угловая извилина
Островок
- Короткая извилина
- Длинная извилина
- Центральная борозда островка
- Круговая борозда
Затылочная доля
- Калькариновая борозда
- Поперечная затылочная борозда
- Полулунная борозда
Медиальный вид
- Подмозолистая извилина
- Поясная извилина
- Мозолистая борозда
- Поясная борозда
- Медиальная лобная извилина
- Краевая борозда
- Предклинье
- Кунеус
- Известковая борозда
- Язычная борозда
- Промежуточный мозг
Вид снизу
Лобная доля
- Извилина прямой мышцы живота
- Обонятельная борозда
- Орбитальные борозды и извилины
Височная доля
- Анкус
- Парагиппокампальная извилина
- Носовая борозда
- Медиальная затылочно-височная извилина
- Коллатеральная борозда
- Боковая затылочно-височная извилина
- Затылочно-височная борозда
Источники
Весь контент, публикуемый на Kenhub, проверяется экспертами в области медицины и анатомии.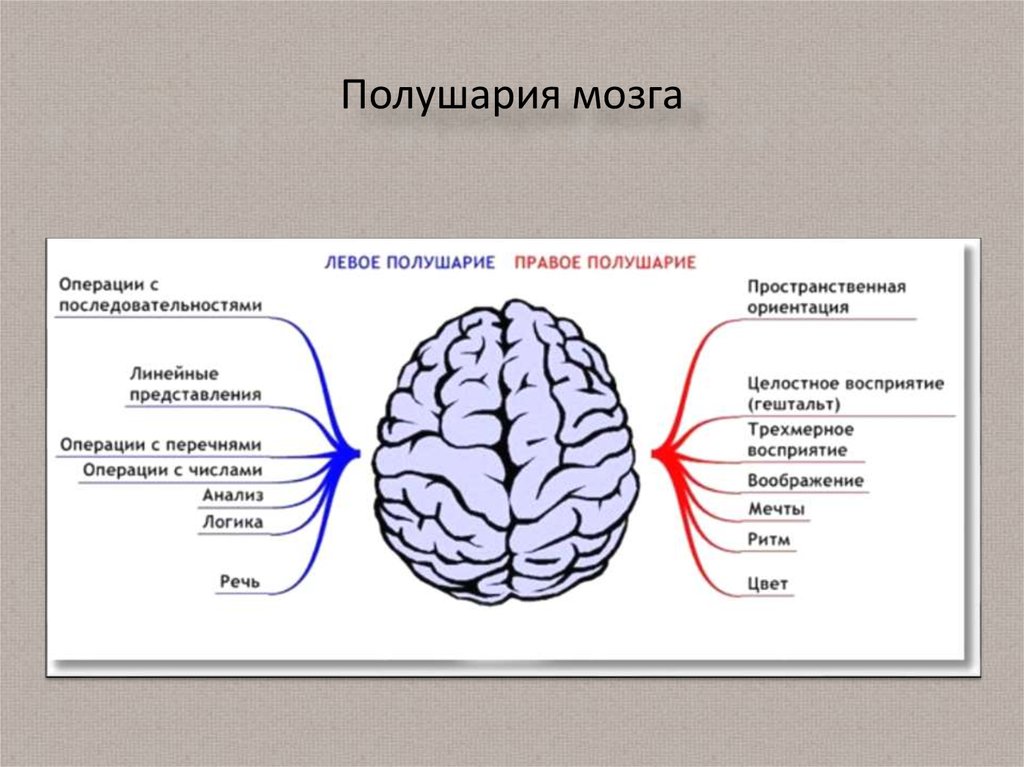 Информация, которую мы предоставляем, основана на научной литературе и рецензируемых исследованиях. Kenhub не дает медицинских консультаций. Вы можете узнать больше о наших стандартах создания и проверки контента, прочитав наши рекомендации по качеству контента.
Информация, которую мы предоставляем, основана на научной литературе и рецензируемых исследованиях. Kenhub не дает медицинских консультаций. Вы можете узнать больше о наших стандартах создания и проверки контента, прочитав наши рекомендации по качеству контента.
Каталожные номера:
- Неттер, Ф. Атлас анатомии человека. 6-е изд. Филадельфия, Пенсильвания: Elservier: Saunders, 2014 г., стр. 106–108.
- Снелл, Р. Клиническая нейроанатомия. 7-е изд. Филадельфия: Wolters Kluwer Health/Lippincott Williams & Wilkins, 2010, стр. 257–262.
Иллюстраторы:
- Поперечный срез головного мозга с выделением серпа головного мозга — Национальная медицинская библиотека
- Островковая доля (вид сбоку слева) — Пол Ким
- Промежуточный мозг (вид сзади) — Пол Ким
Топография полушарий головного мозга: хотите узнать об этом больше?
Наши увлекательные видеоролики, интерактивные викторины, подробные статьи и HD-атлас помогут вам быстрее достичь наилучших результатов.
Чем вы предпочитаете заниматься?
Видео викторины Оба
«Я бы честно сказал, что Kenhub сократил время моего обучения вдвое». – Читать далее. Ким Бенгочеа, Реджисский университет, Денвер
© Если не указано иное, все содержимое, включая иллюстрации, является исключительной собственностью Kenhub GmbH и защищено немецкими и международными законами об авторском праве. Все права защищены.Аномалии полушарий головного мозга — Педиатрия
By
Stephen J. Falchek
, MD, Детская больница Nemours/Alfred I. duPont
Последний обзор/редакция: декабрь 2018 г. | Изменено в сентябре 2022 г.
Посмотреть обучение пациентов
Тематические ресурсы
Полушария головного мозга могут быть большими, маленькими или асимметричными; извилины могут отсутствовать, быть необычно большими или множественными и маленькими.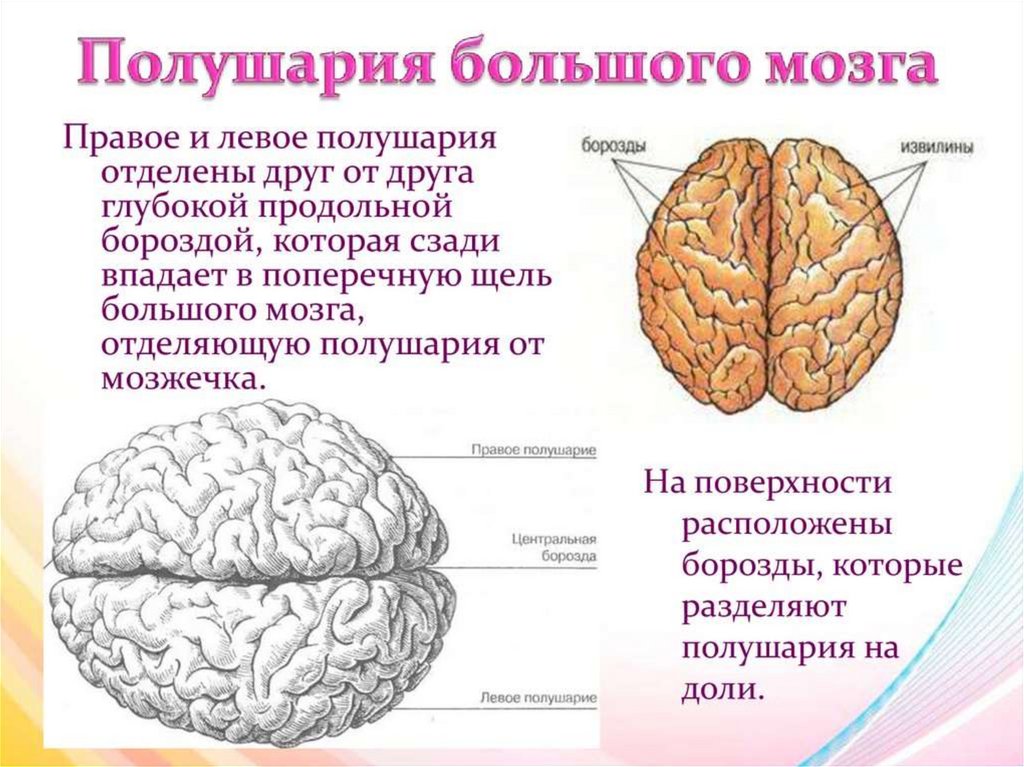
В дополнение к грубо видимым порокам развития микроскопические срезы нормально выглядящего мозга могут показать дезорганизацию нормального ламинарного расположения нейронов. Локализованные отложения серого вещества могут присутствовать в областях, обычно занятых только белым веществом (гетеротопное серое вещество).
Пороки развития полушарий головного мозга могут быть обусловлены генетическими или приобретенными причинами. Приобретенные причины включают инфекции (например, цитомегаловирус) и сосудистые нарушения, которые прерывают кровоснабжение развивающегося мозга.
Микроцефалия Микроцефалия Микроцефалия – окружность головы <2 стандартных отклонений ниже среднего значения для данного возраста. (См. также Введение в врожденные черепно-лицевые и скелетно-мышечные нарушения и Обзор врожденных... читать далее или макроцефалия Макроцефалия Макроцефалия (мегацефалия) — это окружность головы > 2 стандартных отклонений выше среднего для возраста ( 1, 2). (См. также Введение в Врожденные черепно-лицевые и скелетно-мышечные расстройства и. .. читать далее , умеренная или тяжелая двигательная и умственная отсталость Интеллектуальная инвалидность Интеллектуальная инвалидность характеризуется значительно ниже среднего умственного функционирования (часто выражается как коэффициент интеллекта <70-75) в сочетании с ограничениями адаптивных... читать далее , и эпилепсия Эпилептические припадки Припадок представляет собой аномальный, нерегулируемый электрический разряд, возникающий в корковом сером веществе головного мозга и временно прерывающий нормальную функцию мозга. проявления
.. читать далее , умеренная или тяжелая двигательная и умственная отсталость Интеллектуальная инвалидность Интеллектуальная инвалидность характеризуется значительно ниже среднего умственного функционирования (часто выражается как коэффициент интеллекта <70-75) в сочетании с ограничениями адаптивных... читать далее , и эпилепсия Эпилептические припадки Припадок представляет собой аномальный, нерегулируемый электрический разряд, возникающий в корковом сером веществе головного мозга и временно прерывающий нормальную функцию мозга. проявления
Лечение поддерживающее, включая при необходимости противосудорожные препараты.
Спектр голопрозэнцефалии возникает, когда эмбриональный прозэнцефалон не подвергается полной сегментации и дроблению.
Т 3 основных типа голопенсералии, в порядке снижения тяжести, являются
-
Alobar
-
Semilobar
-
LOUPAR
. Варена, обычно является FATLE.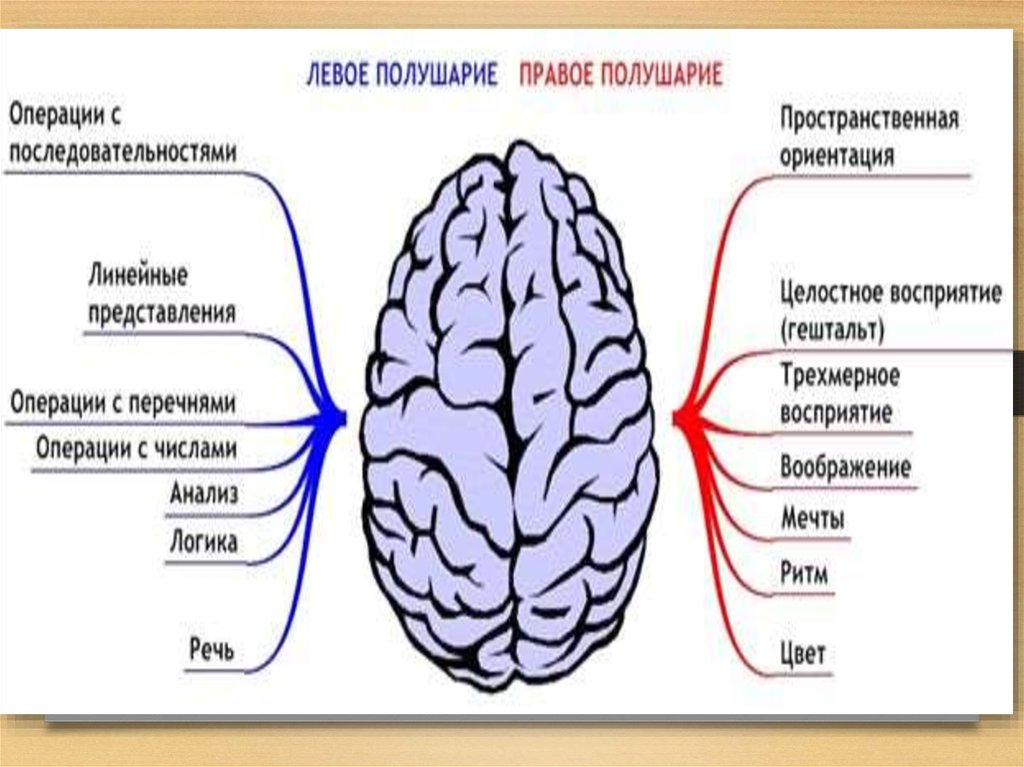 Он характеризуется полным отсутствием расщепления и наличием единственной полости желудочка без какой-либо перегородки.
Он характеризуется полным отсутствием расщепления и наличием единственной полости желудочка без какой-либо перегородки.
Полудолевая голопрозэнцефалия характеризуется частичным расщеплением на полушария сзади, но с сообщающейся единой желудочковой полостью спереди.
Долевая голопрозэнцефалия характеризуется отсутствием прозрачной перегородки (мембраны, разделяющей переднюю часть двух боковых желудочков), агенезией мозолистого тела, сращением передних рогов боковых желудочков и, возможно, слиянием поясной извилины.
Четвертый, редкий тип, называемый срединным межполушарным вариантом, характеризуется слиянием задней лобной и теменной долей, а также, возможно, таламуса, но с нормальной полушарной дифференцировкой в других местах.
Черепно-лицевые аномалии Введение во врожденные черепно-лицевые и мышечно-скелетные аномалии Черепно-лицевые и мышечно-скелетные аномалии распространены среди детей. Они могут затрагивать только одну специфическую локализацию (например, заячья губа, волчья пасть, косолапость) или быть частью синдрома множественных. .. читать далее, присутствующих при голопрозэнцефалиях, и их степень тяжести обычно отражает основную аномалию головного мозга. . В этот спектр входят анофтальмия. Анофтальмия. Глаза могут отсутствовать, быть деформированными или недоразвитыми при рождении, часто в сочетании с другими врожденными аномалиями и синдромами. (См. также Введение во врожденные черепно-лицевые и… читать далее или циклопию, деформированные или отсутствующие ноздри или полость носа, гипотелоризм Гипотелоризм Глаза могут отсутствовать, деформироваться или не полностью развиваться при рождении, часто в сочетании с другими врожденными аномалиями и синдромами. (См. См. также Введение в врожденную черепно-лицевую и… читать далее, расщепление губы и расщелины неба Расщелина губы и расщелины неба Расщелина рта и лица — это врожденный дефект, при котором губа, нёбо или и то, и другое не смыкаются в средней линии и остаются открытыми, образуя заячью губу и/или волчью пасть. Эти дефекты… читать дальше и центральный резец
.. читать далее, присутствующих при голопрозэнцефалиях, и их степень тяжести обычно отражает основную аномалию головного мозга. . В этот спектр входят анофтальмия. Анофтальмия. Глаза могут отсутствовать, быть деформированными или недоразвитыми при рождении, часто в сочетании с другими врожденными аномалиями и синдромами. (См. также Введение во врожденные черепно-лицевые и… читать далее или циклопию, деформированные или отсутствующие ноздри или полость носа, гипотелоризм Гипотелоризм Глаза могут отсутствовать, деформироваться или не полностью развиваться при рождении, часто в сочетании с другими врожденными аномалиями и синдромами. (См. См. также Введение в врожденную черепно-лицевую и… читать далее, расщепление губы и расщелины неба Расщелина губы и расщелины неба Расщелина рта и лица — это врожденный дефект, при котором губа, нёбо или и то, и другое не смыкаются в средней линии и остаются открытыми, образуя заячью губу и/или волчью пасть. Эти дефекты… читать дальше и центральный резец
Голопрозэнцефалии могут быть вызваны мутациями ряда генов, из которых известно > 14; среди них гены сигнального пути sonic hedgehog и их модификаторы (1 ссылка на голопрозэнцефалию).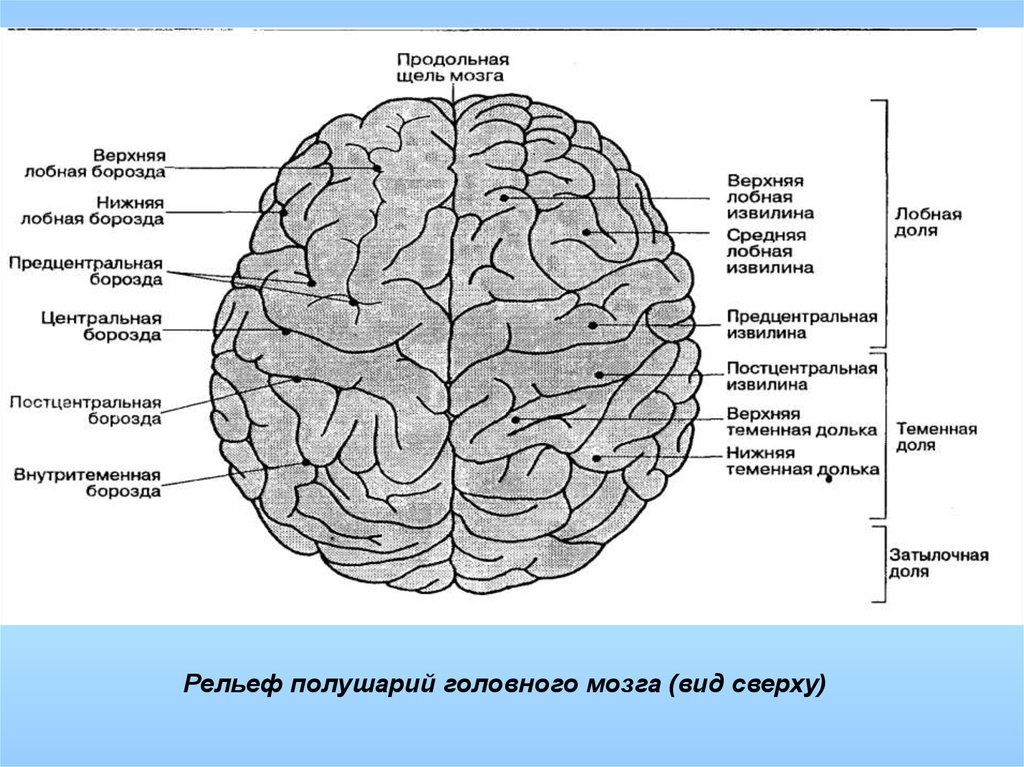 Полушария головного мозга могут быть большими, маленькими или асимметричными; извилины могут отсутствовать, быть необычно большими или множественными и маленькими. разделы… читать дальше ). Трисомия 13 Трисомия 13 Трисомия 13 вызывается дополнительной хромосомой 13 и вызывает аномальное развитие переднего мозга, средней части лица и глаз; тяжелая умственная отсталость; пороки сердца; и маленький размер при рождении. Диагноз… читать дальше и трисомия 18 Трисомия 18 Трисомия 18 вызывается лишней хромосомой 18 и обычно связана с умственной отсталостью, малым ростом при рождении и различными врожденными аномалиями, включая тяжелую микроцефалию, сердце… читать далее , как а также другие хромосомные делеции Синдромы хромосомных делеций Синдромы хромосомных делеций возникают в результате потери частей хромосом. Они могут вызывать тяжелые врожденные аномалии и значительную умственную и физическую инвалидность. Специфические хромосомные… читать далее и дупликации связаны с голопрозэнцефалией.
Полушария головного мозга могут быть большими, маленькими или асимметричными; извилины могут отсутствовать, быть необычно большими или множественными и маленькими. разделы… читать дальше ). Трисомия 13 Трисомия 13 Трисомия 13 вызывается дополнительной хромосомой 13 и вызывает аномальное развитие переднего мозга, средней части лица и глаз; тяжелая умственная отсталость; пороки сердца; и маленький размер при рождении. Диагноз… читать дальше и трисомия 18 Трисомия 18 Трисомия 18 вызывается лишней хромосомой 18 и обычно связана с умственной отсталостью, малым ростом при рождении и различными врожденными аномалиями, включая тяжелую микроцефалию, сердце… читать далее , как а также другие хромосомные делеции Синдромы хромосомных делеций Синдромы хромосомных делеций возникают в результате потери частей хромосом. Они могут вызывать тяжелые врожденные аномалии и значительную умственную и физическую инвалидность. Специфические хромосомные… читать далее и дупликации связаны с голопрозэнцефалией.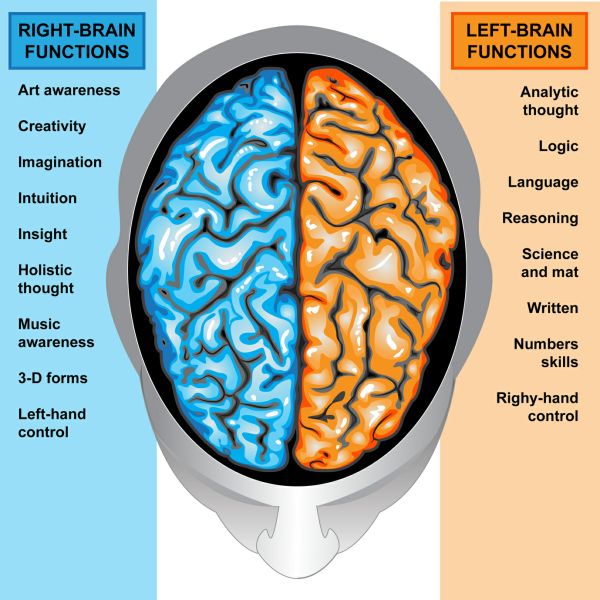
Плоды с тяжелым поражением могут погибнуть до рождения. Лечение голопрозэнцефалии поддерживающее.
-
1. Хонг М., Сривастава К., Ким С. и др.: BOC — ген-модификатор голопрозэнцефалии. Hum Mutat 38(11):1464–1470, 2017. doi: 10.1002/humu.23286. Epub 2017 Jul 21.
Лиссэнцефалия состоит из аномально толстой коры, уменьшенного или отсутствующего рисунка извилин на поверхности головного мозга, уменьшенной или аномальной слоистости коры головного мозга и часто диффузных гетеротопий нейронов.
Эта мальформация вызвана аномальной миграцией нейронов, процессом, при котором незрелые нейроны прикрепляются к радиальной глии и перемещаются из мест своего происхождения вблизи желудочка на поверхность мозга. Несколько дефектов одного гена Дефекты одного гена Генетические нарушения, определяемые одним геном (менделевские расстройства), легче всего анализировать и лучше всего понять. Если для проявления признака требуется только одна копия гена (один аллель).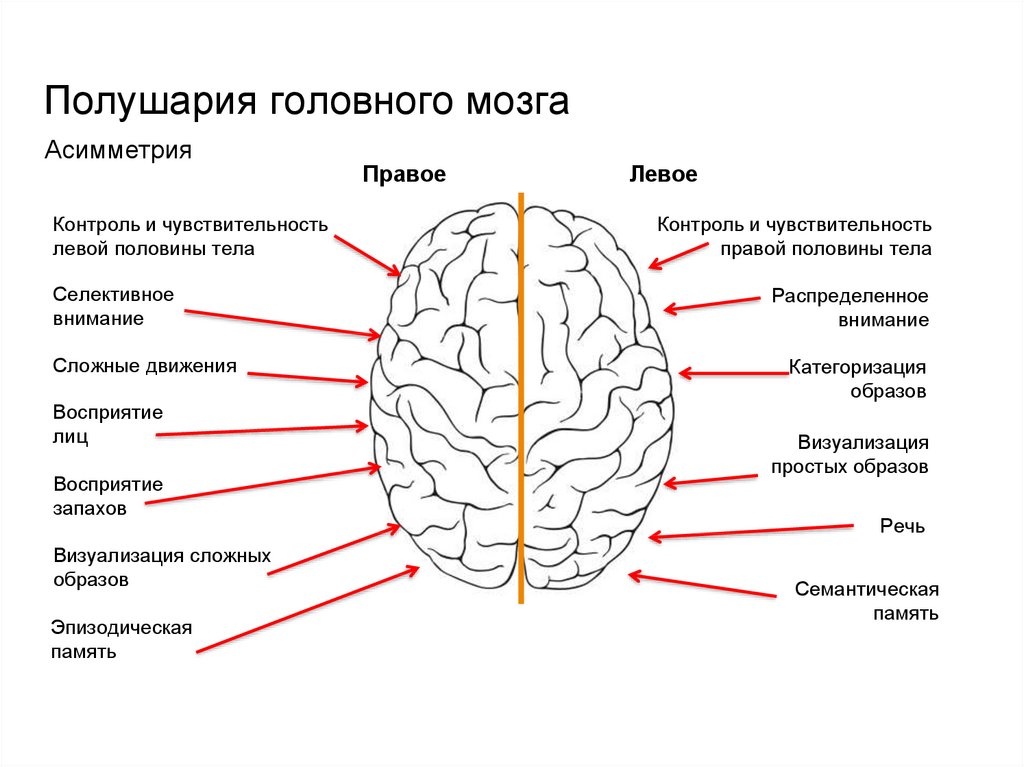 .. читать дальше может вызвать эту аномалию (например, ЛИС1 ). Один Х-сцепленный Х-сцепленный рецессивный Генетические нарушения, определяемые одним геном (менделевские расстройства), легче всего анализировать и лучше всего понять. Если для проявления признака требуется только одна копия гена (один аллель)… читать далее ген DCX вызывает семейную Х-сцепленную лиссэнцефалию у мужчин и другую, обычно более легкую миграционную аномалию у женщин, называемую гетеротопией подкорковых полос. При гетеротопии подкорковых полос широкая полоса эктопического серого вещества в подкорковом белом веществе напоминает «двойную кору» на МРТ.
.. читать дальше может вызвать эту аномалию (например, ЛИС1 ). Один Х-сцепленный Х-сцепленный рецессивный Генетические нарушения, определяемые одним геном (менделевские расстройства), легче всего анализировать и лучше всего понять. Если для проявления признака требуется только одна копия гена (один аллель)… читать далее ген DCX вызывает семейную Х-сцепленную лиссэнцефалию у мужчин и другую, обычно более легкую миграционную аномалию у женщин, называемую гетеротопией подкорковых полос. При гетеротопии подкорковых полос широкая полоса эктопического серого вещества в подкорковом белом веществе напоминает «двойную кору» на МРТ.
Пораженные младенцы почти всегда имеют значительную умственную отсталость Умственная отсталость Умственная отсталость характеризуется значительно ниже среднего умственного функционирования (часто выражается как коэффициент умственного развития < 70-75) в сочетании с ограничениями адаптивных. .. читать далее и судорогами (часто инфантильные спазмы Инфантильные Судороги Инфантильные спазмы — это приступы, характеризующиеся внезапным сгибанием рук, наклоном туловища вперед, разгибанием ног и гипсаритмией на электроэнцефалографии.Лечение адренокортикотропное… читать далее ).
.. читать далее и судорогами (часто инфантильные спазмы Инфантильные Судороги Инфантильные спазмы — это приступы, характеризующиеся внезапным сгибанием рук, наклоном туловища вперед, разгибанием ног и гипсаритмией на электроэнцефалографии.Лечение адренокортикотропное… читать далее ).
Лечение лиссэнцефалии поддерживающее; выживаемость зависит от тяжести припадка и наличия других осложнений, включая нарушение глотания, апноэ и трудности с отхождением секрета из ротоглотки.
Полимикрогирия, при которой извилины маленькие и избыточные, также связана с аномальной миграцией нейронов. Другие распространенные признаки включают упрощенное или отсутствующее расслоение коры в пораженных областях, гетеротопическое серое вещество, гипоплазию или отсутствие мозолистого тела и прозрачной перегородки, а также пороки развития ствола головного мозга и/или мозжечка. Структурные аномалии могут быть диффузными или очаговыми. Наиболее распространенной областью очагового поражения является перисильвиева щель (двусторонняя или односторонняя).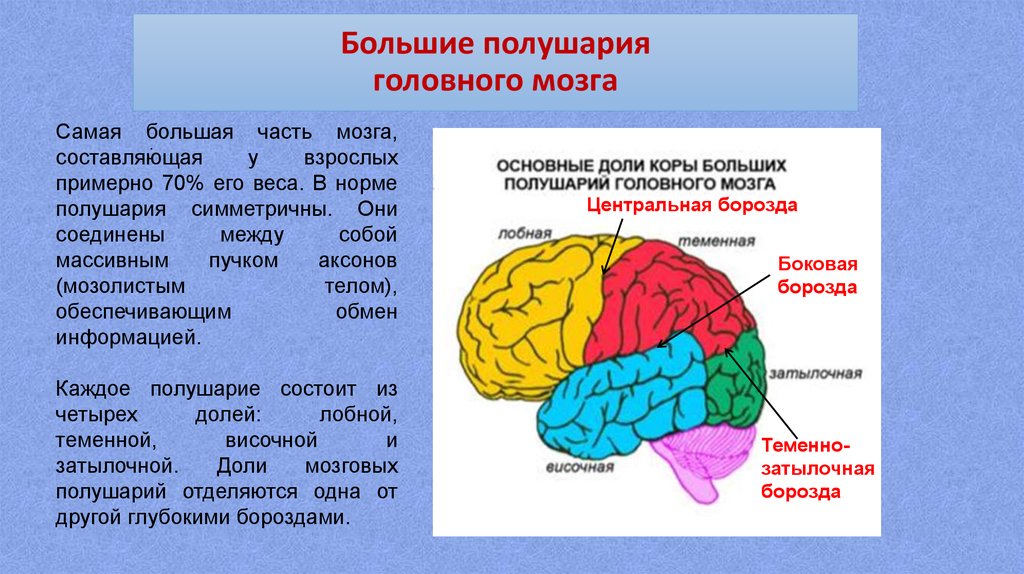
Полимикрогирия тесно связана с шизэнцефалией Шизэнцефалия Порэнцефалия представляет собой полость, которая может развиваться пренатально или постнатально в полушарии головного мозга. Полости часто сообщаются с желудочком, но могут быть и закрытыми (т.е. несообщающимися)… читать далее , в которых имеются аномальные щели, или расщелины, в полушариях головного мозга. Были идентифицированы многочисленные причины полимикрогирии, в том числе ряд мутаций одного гена (например, SRPX2 ) и первичная материнская инфекция цитомегаловирусом (т. е. при отсутствии у матери предшествующего иммунитета — см. Врожденная и перинатальная цитомегаловирусная инфекция (см. ЦМВ) Врожденная и перинатальная цитомегаловирусная инфекция (ЦМВ) Цитомегаловирусная инфекция может быть приобретена внутриутробно или перинатально и является наиболее распространенной врожденной вирусной инфекцией.Признаки при рождении, если они присутствуют, включают задержку внутриутробного развития, недоношенность.