
Толерантность 10 класс презентация: Презентация классного часа «Урок толерантности» (10 класс)
Классный час по теме толерантность презентация | Классный час (10 класс) на тему:
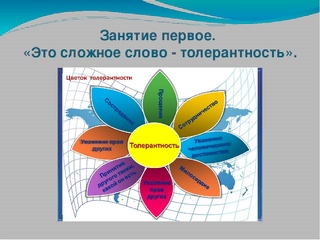
Слайд 1
МБОУ БРАТСКИЙ УВК Классный час в 10 классе « Уважая себя, уважай других». Понятие толерантности Классный руководитель: Аблаев Л.С 2016г.Слайд 2
Цель: Создать условия для формирования терпимости к различиям между людьми ( индивидуальным, различиям по полу, возрасту, социальному положению, национальности, расе, мировоззрению). Задачи: Способствовать выработке активного неприятия любых форм дискриминации. Познакомить учащихся с Декларацией принципов толерантности. Учащиеся должны осознать неодинаковость людей, научиться видеть различия между людьми и воспринимать это как положительный факт.
Слайд 3
Игра №1. Знакомство. « Звали, зовут, будут звать». « Когда я был маленьким, меня звали Вовочка, в школе меня зовут Володя , а когда я вырасту, меня будут звать Владимир Иванович .»
Слайд 4
Игра №2. Сходство и различия». «СХОДСТВА» и «РАЗЛИЧИЯ». Вопросы для обсуждения: Как влияют сходства и различия на жизнь людей? Национальность влияет на взаимоотношения людей? Лица каких рас и национальностей вам трудно различить? Какие религии вы знаете? Разные вероисповедания объединяют или разъединяют? Как сказывается на общении с другими людьми наличие физических недостатков у человека? Как общаются между собой старые и молодые?
Слайд 5
Выводы: О человеке судят по его профессии, отношениях в семье и окружающими. Выделяют особенности речи, умение ясно выражать свои мысли. Люди различны, различна их культура. Отчужденность, враждебность в непринятии другой культуры весьма распространенное явление в нашей жизни. Возникает из-за боязни неизвестного, ожидания враждебности со стороны другой группы, ощущения дискомфорта от вторжения «чужих» в привычную жизнь. Может быть, гораздо проще жить в мире, где все люди одинаковые?
Выделяют особенности речи, умение ясно выражать свои мысли. Люди различны, различна их культура. Отчужденность, враждебность в непринятии другой культуры весьма распространенное явление в нашей жизни. Возникает из-за боязни неизвестного, ожидания враждебности со стороны другой группы, ощущения дискомфорта от вторжения «чужих» в привычную жизнь. Может быть, гораздо проще жить в мире, где все люди одинаковые?
Слайд 6
Игра № 3. « Кто я?» Жизнь – театр, у каждого свои роли. Запишите на листочке: Кто я? Напишите свои жизненные роли: ученик, сын, внук, помощник, покупатель и др.
Слайд 7
Ситуация: Если вы попали мячом в стекло и разбили его. Что скажет мама? Что скажет соседка? Просто все воспринимают нас по — разному . А как мы себя ведем по отношению к чужим людям, родственникам? Поможем соседке донести тяжелую сумку? А если мама попросит пропылесосить комнату? Вывод: Если бы мы с большей долей терпения, уважения, предрасположенности, симпатии, понимания относились друг к другу, то и роли в жизни нам исполнять было бы проще.
Слайд 8
Секреты эфффективного общения 1. Говори так , чтобы тебя услышали. 2. Слушай так , чтобы ты понял. 3. Создавай доброжелательную атмосферу для общения. 4. Высказывай свои слова и чувства не унижая собеседника.
Слайд 9
Историческая справка В Х VIII – X I X веке во Франции жил князь Талейран Перигор . Он отличался тем, что и при Наполеоне и при короле Людовике оставался неизменно министром иностранных дел. Это был человек талантливый во многих областях, но более всего отличался умением учитывать настроение окружающих, уважительном к ним отношении, искал решение проблем способом, наименее ущемляющим интересы других людей. И при этом сохранял свои собственные принципы, стремясь к тому, чтобы управлять ситуацией, а не слепо подчиняться обстоятельствам.
Слайд 10
«Деклараци я принципов толерантности» генеральной конференции ЮНЕСКО от 16 ноября 1995 года, толерантность означает «уважение, принятие и правильное понимание богатого многообразия культур нашего мира, наших форм самовыражения и способов проявлений человеческой индивидуальности» «Толерантность» означает : на испанском языке способность признавать отличные от своих собственных идеи и мнения; на французском – отношение при котором допускается, что другие могут думать или действовать иначе, нежели ты сам; на английском – готовность быть терпимым, снисходительным; на китайском – позволять, принимать, быть по отношению к другим великодушным; на арабском – прощение, снисходительность, мягкость, милосердие, сострадание, благосклонность, терпение, расположенность к другим; на русском – способность терпеть что-то или кого-то (быть выдержанным, выносливым, стойким, уметь мириться с существованием чего-либо, кого-либо).
Слайд 11
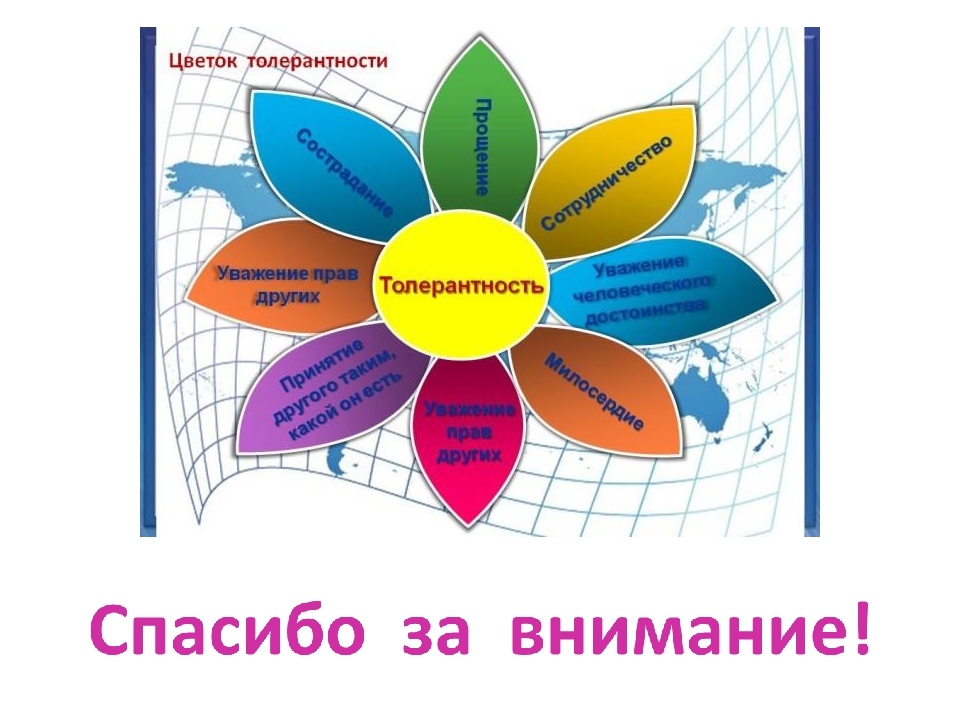
Толерантность Прощение Уважение прав других Сотрудничество, дух партнерства Уважение человеческого достоинства Милосердие Сострадание Терпимость к чужим мнениям, верованиям, поведению Принятие другого таким, какой он есть
Слайд 12
Какая цифра-? 6
Методические документы | Средняя общеобразовательная школа № 70 ИМЕНИ ГЕРОЯ СОВЕТСКОГО СОЮЗА ДМИТРИЯ МИРОШНИЧЕНКО
Классный час «Экскурсия по улице Красной» 06.01.2021
Презентация классного часа
Классный час «Народы Кубани» 30.12.2020
Презентация классного часа
Классный час «Есть такая профессия — Родину защищать» 18.02.2020
Сценарий классного часа
Презентация к классному часу
Классный час на тему «Коррупция в Российском обществе: образование» 14.11.2019
Сценарий классного часа
Презентация к классному часу
Классный час «Слава воину-победителю» 08.10.2019
Сценарий классного часа
Литературно-музыкальная композиция «День Победы» 08. 10.2019
10.2019
Сценарий мероприятия
Внеклассное мероприятие «Профилактика правонарушений среди несовершеннолетних» 27.09.2019
Сценарий мероприятия
- Классный час «Что такое экстремизм?» (9-11 класс)
- Классный час «Терроризм-угроза обществу»
- Классный час «Патриотизм без экстремизма»
- Классный час «Что такое толерантность?» (8-10 класс)
- Классный час «О толерантности»
- Классный час «Добро и зло. Твой нравственный выбор»
- Примеры тренировочных упражнений по формированию и развитию толерантности.
- Классный час «Этнические стереотипы и предубеждения»
- Тренировочное занятие для учащихся 9-х классов «Толерантность-путь к гражданскому обществу»
- Тренировочное занятие для учащихся 7-х классов в рамках проведения тренинга по выработке навыков толерантности.
- Урок правоведения в 10 классе. Деловая игра «Заседание Государственной Думы»
- Классный час «Поговорим о дружбе» (5 класс)
- Классный час «Как научиться дружить?» (начальная школа)
Скачать сборник
Классный час на тему «Толерантность-путь к миру»
МОУ Лесная СОШ
Классный час на тему
«Толерантность-путь к миру»
Подготовила:
Классный руководитель 8 б класса
Поспелова Е. Н.
Н.
Тверь 2018
Классный час по теме: «Толерантность – путь к миру»
Цель:
Познакомить учащихся с понятием «толерантность», его происхождением, значением и актуальностью его формирования, как нравственного качества личности.
Задачи:
Сформировать правильное представление о толерантном поведении;
Дать возможность учащимся оценить степень своей толерантности;
Развивать внимание и творческое мышление учащихся; развивать навыки социального поведения;
Воспитывать чувство коллективизма, сплочённости; способствовать развитию уважительного отношения между учащимися.
Оборудование: компьютер, экран, проектор, мультимедийная презентация, раздаточный материал.
Ход классного часа:
Теперь, когда мы научились
Летать по воздуху, как птицы,
Плавать под водой, как рыбы,
Нам не хватает только одного:
Научиться жить на земле, как люди.
Б.Шоу.
— Набирает обороты третье тысячелетие. Прогресс неумолимо движется вперёд. Техника пришла на службу человечеству. Казалось бы, жизнь должна стать размереннее, спокойнее. Но, к сожалению, мы всё чаще слышим слова: жертва насилия, беженец, теракт, война…
В сегодняшнем обществе происходит активный рост агрессивности, расширение зон конфликтов. Эти социальные явления особо затрагивают молодёжь, которой в силу возрастных особенностей свойственно стремление к простым и быстрым решениям социальных проблем. В последнее время в подростковой и молодёжной среде наблюдается катастрофический рост всевозможных форм асоциального поведения.
Очень уместны и точны в этой ситуации слова Бернарда Шоу, выбранные мною эпиграфом к нашему классному часу.
Упражнение “Салфетка”.
— На ваших столах лежат бумажные салфетки. Возьмите, пожалуйста, по одной и сверните её пополам любым способом. Оторвите с
любого края уголок. Сверните её ещё раз пополам и опять оторвите кусочек с любого уголка. Сверните в третий раз пополам и снова оторвите. А теперь разверните и продемонстрируйте то, что у вас получилось.
Сверните в третий раз пополам и снова оторвите. А теперь разверните и продемонстрируйте то, что у вас получилось.
— Вряд ли найдётся пара салфеток, на которых повторяется рисунок. Как вы думаете, почему мы получили такой результат? (Ответы)
— Послушайте, пожалуйста, китайскую притчу «Ладная семья».
Жила-была на свете семья. Она была не простая. Более 100 человек насчитывалось в этой
семье. И занимала эта семья целое село. Так и жили всей семьей и всем селом. Вы скажете: ну и что, мало ли больших семейств на свете. Но дело в том, что семья была
особая – мир и лад царили в этой семье и, стало быть, на селе. Ни ссор, ни ругани, ни, Боже упаси, драк и раздоров.
Дошел слух об этой семье до самого владыки страны. И он решил проверить, правду ли молвят люди. Прибыл он в село, и душа его возрадовалась: кругом чистота, красота, достаток и мир. Хорошо детям, спокойно старикам. Удивился владыка. Решил узнать, как жители села добились такого лада, пришел к главе семьи; расскажи, мол, как ты добиваешься такого согласия и мира в твоей семье. Тот взял лист бумаги и стал что-то писать. Писал долго – видно, не очень силен был в грамоте. Затем передал лист владыке. Тот взял бумаги и стал разбирать каракули старика. Разобрал с трудом и удивился. Три слова были начертаны на бумаге: любовь, прощение, терпение.
Тот взял лист бумаги и стал что-то писать. Писал долго – видно, не очень силен был в грамоте. Затем передал лист владыке. Тот взял бумаги и стал разбирать каракули старика. Разобрал с трудом и удивился. Три слова были начертаны на бумаге: любовь, прощение, терпение.
И в конце листа: сто раз любовь, сто раз прощение, сто раз терпение.
Почесал владыка, почесал, как водится, за ухом и спросил:
— И все?
— Да, — ответил старик, — это и есть основа жизни всякой хорошей семьи.
И подумав, добавил:
— И мира тоже.
— Какие три слова стали законом в семье, где царили мир и согласие?
Дети: любовь, терпение, прощение.
— Как вы думаете, о чём будем беседовать на занятии?
Дети: дружба, доброта, терпение, любовь, уважение…
— Всё то, о чём вы сейчас говорили, предполагали, можно объединить, назвать одним словом «толерантность».
— Девиз нашего классного часа звучит так:
Прийти вместе – это начало,
Остаться вместе – это развитие,
Работать вместе – это успех.
Игра «Волшебная рука
Участники обводят на листе свою руку. Предлагается написать на пальцах свои хорошие качества, а на ладони – то, что хотелось бы изменить в себе
Знакомство с понятием «толерантность».
Толерантность – (лат. tolerantia – терпение) – это способность терпеть что-то или кого-то.
Понятие «толерантность» для обычного российского сознания непривычно. Нам ближе наше российское слово – «терпимость».
Терпимость – способность и умение терпеть, быть терпеливым, мириться с чужим мнением.
Дать определение толерантности довольно-таки трудно из-за того, что в разных языках оно трактуется по-разному. И сейчас мы в этом убедимся.
Толерантность – способность признавать отличные от своих собственных идеи и мнения. (Испанский)
Толерантность – готовность быть терпимым, снисходительным. (Английский)
Толерантность – позволять, принимать, быть по отношению к другим великодушным. (Китайский)
Толерантность – прощение, снисходительность, мягкость, милосердие, сострадание, терпение. (Арабский)
(Арабский)
— Что общего в этих определениях?
Дети: умение жить в мире и согласии со всем миром.
«…Толерантность означает уважение, принятие и правильное понимание богатого многообразия культур нашего мира, наших форм самовыражения и способов проявлений человеческой индивидуальности. Ей способствуют знания, открытость, общение и свобода мысли, совести и убеждений. Толерантность – это свобода в многообразии. Это не только моральный долг, но и политическая, и правовая потребность. Толерантность – это добродетель, которая делает возможным достижение мира и способствует замене культуры войны культурой мира».
(«Декларация принципов толерантности», утвержденная резолюцией 5.61 Генеральной конференции ЮНЕСКО от 16 ноября 1995 года)
16 ноября жители многих стран мира отмечают Международный день толерантности или День терпимости. Этот праздник был учрежден в 1996 году по решению Генеральной Ассамблеи ООН.
Население планеты Земля составляет почти
7 миллиардов человек различных национальностей.
Символ толерантности — радужный флаг и сплетенные разноцветные руки
Задание «Соотнеси высказывание с заповедью».
«Наши сердца должны быть наполнены любовью к людям, нашим братьям и сестрам. Мы должны думать о них и помогать им, не ожидая ни малейшего вознаграждения». (возлюби ближнего своего)
«Гнев – зверообразная страсть по расположению духа, способная часто повторяться, жестокая и непреклонная по силе, служащая причиною убийств, союзница несчастия, помощница вреда и бесчестия». (не убий)
«Ничто притворное не может быть продолжительным». (не лги)
«Любовь к родителям — основа всех добродетелей». (почитай отца своего и матерь свою)
Воровство порождает лень и жадность. (не кради)
— Итак, быть толерантным, значит:
Уважать другого.
Любить ближнего своего.
Не злиться.
Быть добрым, терпимым.
Сострадать.
— Понятие «толерантность» изображено в виде цветка. Почему?
Почему?
Учитель: — цветок это красота мира, так и толерантная личность, совершая добрые
поступки, становится лучше, чище, светлее. От неё исходит добро, тепло. Вокруг неё всегда есть друзья, царит счастье. Толерантная личность понимает окружающих, всегда приходит им на помощь.
Задание «Толерантная и интолерантная личность».
Распределить в две колонки: I – черты, присущие толерантной личности, II – интолерантной личности.
Черты:
Терпимость
Бессердечность
Непонимание
Сострадание
Прощение
Вспыльчивость
Снисходительность
Милосердие
Злорадство
Лживость
Сотрудничество
Доверие
Раздражительность
Зависть
Пренебрежение
Умение владеть собой
Равнодушие
Чуткость
Агрессивность
Эгоизм
Альтруизм
Доброжелательность
Проверка качества выполнения задания. Таблица «Толерантная и интолерантная личность»
Таблица «Толерантная и интолерантная личность»
| Толерантная личность | Интолерантная личность |
| Терпимость | Бессердечность |
| Сострадание | Непонимание |
| Прощение | Вспыльчивость |
| Снисходительность | Злорадство |
| Милосердие | Лживость |
| Сотрудничество | Раздражительность |
| Доверие | Зависть |
| Умение владеть собой | Пренебрежение |
| Чуткость | Равнодушие |
| Альтруизм | Агрессивность |
| Доброжелательность | Эгоизм |
Решение проблемных ситуаций.
— Подумайте, а все ли мы обладаем качествами толерантной личности?
Дети: нет, ссоримся…
— Все ли можем спокойно выслушать друг друга? Поддержать в трудную минуту?
— Из-за чего между вами возникают конфликты со своими друзьями, братьями или сестрами?
— Предлагаю вам ситуации.
1.Покажите, как можно решить проблему мирным путем.
Вы играете в игру, а кто-то подходит и забирает ее, даже не спросив, закончили вы играть или нет.
— Как поступите в этой ситуации?
— Что можно сделать, чтобы сохранить спокойствие в конфликтной ситуации?
— Как можно избежать драки?
Дети: обсудить проблему и разойтись, сменить тему, отложить обсуждение до тех пор, пока оба не успокоятся.
2.Вы играете в игру, а один из участников команды не соблюдает правила.
— Как вы поступите в этой ситуации?
Задание составить памятку «Как себя вести в конфликтной ситуации».
А я хочу вам предложить памятку, которая научит вас, как правильно вести себя в конфликтной ситуации.
Памятка.
Сохраняй спокойствие, когда сердишься.
Владей собой, имей выдержку.
Имей терпение выслушать собеседника.
Спокойно объясни свою точку зрения.
Подумай, из-за чего возник спор.
Сказка Ольги Дунец «Молчание и молчание».
Однажды там, где нужно было слово, встретились молчание и молчание. За один шаг до надвигающейся беды узнали друг друга две горькие обиды. Взгляды их выражали одиночество, пустоту, и в них было что-то тоскливое.
Вдруг раздвинулась бездна и за спинами встали твердые скалы. Ужаснулись молчание и молчание. Они увидели конец своей дороги. Немые губы сжались и языки мучительно искали слово.
Их силы были на исходе. И осталось — не вместе жить, а вместе умереть. И потянулись руки навстречу, и слово родилось: «Прости!»
Беседа:
— Обижали ли вас когда-нибудь?
— Какими качествами должен обладать человек, умеющий прощать?
— Умеете ли вы прощать?
— А как вы думаете, прощение – удел слабых или сильных?
(Благословенны будьте, травы
И воды в зелени оправ!
Виновных нет: все люди правы,
Но больше всех – простивший прав!
(Звучит песня «Прости, поверь» в исполнении А. Варум.)
Варум.)
Задание « Объясни пословицу».
Есть слёзы – ( есть и совесть).
Упавшего не считай (за пропавшего).
Кайся, да опять (за старое не принимайся).
Тому тяжело, (кто помнит зло).
Злой человек, как уголь: (если не жжёт, то чернит).
— Когда нам наносят глубокую рану – мы никогда не исцелимся, пока не простим. Прощение никогда не изменит прошлое, но оно освободит будущее, сделает вас счастливыми!
Игра «Комплимент».
— Любой человек хочет, чтобы его любили и уважали, ценили и понимали, и очень часто нам просто необходимо слышать от окружающих людей добрые слова и пожелания. Ведь от этого улучшается настроение и мы готовы делиться радостью и помогать другим.
— Сделайте комплимент или похвалите своего одноклассника.
— Что чувствовал каждый из вас, когда о нём говорили?
— Поднимите руку те, кому были приятны сказанные слова?
— Кто был смущён? Почему комплименты были разными?
Продолжите фразу: НЕ ОЦЕНИВАЙТЕ ЛЮДЕЙ, А ЦЕНИТЕ!
— Все вы разные, и как из разных звеньев мозаики мы можем составить единое целое (картинку), так из вас разных можно собрать единый сплоченный коллектив.
— А что у вас общее?
Дети отвечают :
А) учёба;
Б) цель – получить образование;
В) класс, т.е коллектив;
Г) возрастные интересы.
— Невозможно нравится всем, невозможно любить всех, невозможно, чтобы все ребята класса были преданными друзьями…Потому что у нас разное воспитание, образование, интеллект, желание учиться, потребности, характер, вкусы, темперамент и т.д. Но мы можем жить в согласии друг с другом, ценить друг друга, принимать друг друга такими, какие есть, т.е быть толерантными.
Я пришел в этот мир
Не для того, чтобы оправдывать твои надежды,
Не для того, чтобы отвечать твоим интересам,
Не для того, чтобы соответствовать твоим ожиданиям.
И ты пришел в этот мир
Не для того, чтобы соответствовать моим ожиданиям,
Не для того, чтобы отвечать моим интересам,
Не для того, чтобы оправдывать мои надежды.
Потому что я – это я, а ты – это ты.
Но если мы встретились и поняли друг друга – то это прекрасно!
А если нет – ну что ж, печально.
Тест «Насколько я толерантен?»
-А сейчас мы с вами проведем тестирование и определим, насколько вы толерантны. Если вы выбираете первый ответ, загибайте палец.
1.Миша плохо одет…
Это неважно.
Ты подсмеиваешься над ним.
2. Петя питается не так, как ты, по причине своей религии…
Ты просишь, чтобы он объяснил это.
Ты говоришь, что он выглядит смешно.
3. Цвет кожи у Джо отличается от твоего…
Ты стремишься лучше узнать его.
Ты высказываешь по этому поводу обидные замечания.
4. Пожилая женщина медленно идет…
Ты помогаешь ей и придерживаешь дверь.
Ты отталкиваешь ее, чтобы обогнать.
5. На твоих глазах на кого – то нападают…
Ты пытаешься защитить его.
Ты делаешь вид, что ничего не замечаешь.
6. К тебе подходит ребенок – инвалид…
Ты естественным образом разговариваешь с ним.
Ты отходишь от него и не знаешь, что сказать.
Интерпретация результатов:
Загнул 6 пальцев? Прекрасно! Ты уверен в себе, можешь выразить свое мнение. И ты понял, что твоя свобода заканчивается там, где начинается свобода других.
Если от 3 до 5, то ты не очень толерантен. Ты недостаточно уверен в себе для того, чтобы делиться и выражать свое мнение, однако, ты добрый и со временем у тебя все получится.
Меньше трех? Ай – ай – ай! Ты совсем не толерантен! Если ты попытаешься лучше понять себя, какой ты есть, то сможешь стать по-настоящему счастливым!
Терпеть и верить:
Все на свете
Прекрасны –
Взрослые и дети,
Коты собаки и
Медведи,
И сослуживцы и соседи.
Терпимость —
Наш взаимный шанс,
Ведь кто-то также терпит нас.
Пит Хейн
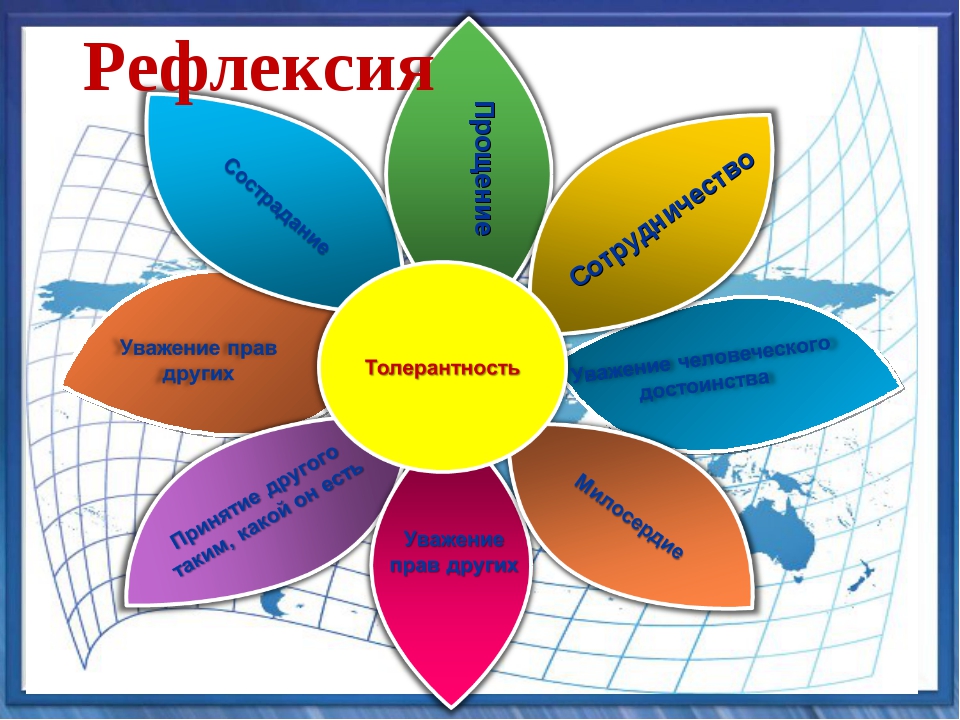
Рефлексия «Дерево толерантности» .Творческая работа.
— Вырастим вместе дерево Толерантности на нашей планете. Пусть наше дерево распустит листочки и зазеленеет. Крону дерева составим из листочков с вашими высказываниями на тему «Планета Толерантности». Возьмите каждый по листочку и напишите на них, что, по-вашему, надо сделать, наша школа стала пространством толерантности. Затем листочки приклейте на дерево.(на доске).
Возьмите каждый по листочку и напишите на них, что, по-вашему, надо сделать, наша школа стала пространством толерантности. Затем листочки приклейте на дерево.(на доске).
Мои пожелания:
Ребята, поступайте с другими так же, как хотите, чтобы они поступали с вами.
Будьте добрыми: любящими, внимательными, терпеливыми, заботливыми, милосердными, прощайте.
Будьте надёжными: честными, правдивыми, имейте чистое сердце, выполняйте свои обещания.
Будьте заботливыми: вежливыми, внимательными, любезными.
Будьте щедрыми: не жадными, бескорыстными, великодушными, готовыми помочь.
— И всё это вам поможет жить в мире.
Если каждый друг к другу будет терпим,
То вместе мы сделаем толерантным мир!!!
Мир из 100 человек. О толерантности учащимся 10-11 классов
1. Мир из 100 человек
О толерантностиучащимся 10-11 классов
2. Мир из 100 человек…
Если бы на всей земле жили только 100 человек, то 57 из них были бы выходцами из Азии, 21 – изЕвропы, 14 – со всего американского континента и 8 — из Африки.
 52 были бы женщинами, 48 –
52 были бы женщинами, 48 –мужчинами, 30 – белыми, 70 принадлежали бы к другим расам. Христианами были бы 30
жителей, мусульманами — 23, 47 исповедовали бы иные религии. Гетеросексуалами были бы 89
человек, 11 гомосексуалистами. 6 человек владели бы 59% всех ее богатств, и все они были бы
североамериканцами. 80 жили бы в крайней бедности. Читать не умели бы 70, 50 регулярно
недоедали. 1 человек каждый день умирал, 1 рождался. 1 человек имел бы высшее образование
и лишь 1 обладал компьютером.
Если вы проснетесь завтра утром скорее здоровым, чем больным, вам крупно повезло по
Если вы никогда не участвовали в военных битвах, не испытали мук голода, пыток, одиночества и
тюремного заточения, то вы счастливчик по сравнению с 500 млн. человек, которые все это
пережили.
Если вы можете исповедовать свою религию без риска преследования, ареста, пыток и смерти, то 3
млрд.
 ваших современников в мире не могут похвастаться тем же.
ваших современников в мире не могут похвастаться тем же.Если у вас есть еда в холодильнике, одежда и крыша над головой, то вы богаче 75% жителей земли.
Ну а уж если вы располагаете банковским счетом или, по крайней мере, у вас в бумажнике
завалялось немного наличности, то вы относитесь к 8% избранных преуспевающих землян.
Если ваши родители все еще живы и, более того, все еще не развелись, вы – редкий везунчик, каких
мало даже в Канаде и США.
Если вы самостоятельно прочитали эту информацию, это выгодно отличает вас от 2 млрд.
современников, которые не умеют читать.
3. Толерантность? Что это?
«Если я чем-то на тебя не похож, я этим вовсе неоскорбляю тебя, а, напротив, одаряю».
Антуан де Сент-Экзюпери
4. Цифры и факты:
В настоящее время только в России около 2 млн.детей с ограниченными возможностями здоровья.
Это 8% детей всего мира.
654 000 живут в неполной семье или являются
полными сиротами.
В России около 90000 глухонемых, что составляет
112 человек на 100000 жителей, в мире на 100000
жителей это соотношение колеблется от 18 человек
в Австралии до 380 в Аргентине.

5. Черты толерантного человека:
ВеликодушиеДружелюбность
Понимание
Отзывчивость
Мягкость
……
……
……
……
6. НЕ ТАКОЙ как все.
Что чувствует тот, кто не похож на остальных?7. Подводя итоги…
Кто такой толерантныйчеловек?
Зачем нужна толерантность?
Как научиться быть
толерантным?
школа 146 г.Челябинска — Методические материалы
Более подробная информация представлена на странице официального сайта Минпросвещения России.
Материалы онлайн-совещаний руководителей ГМО учителей-предметников
ОБУЧЕНИЕ С ИСПОЛЬЗОВАНИЕМ ДИСТАНЦИОННЫХ ТЕХНОЛОГИЙ»
РОССИЙСКИЕ ОБРАЗОВАТЕЛЬНЫЕ ПЛАТФОРМЫ с бесплатной регистрацией и с высоким уровнем качественного образовательного контента;
ОБЗОР ПРОГРАММНОГО ОБЕСПЕЧЕНИЯ, позволяющего организовывать и проводить онлайн-трансляции, вебинары, видеокурсы, тесты, опросы;
МЕТОДИЧЕСКИЕ РЕКОМЕНДАЦИИ по созданию «Наполняемых учебных курсов» в ГИС «Образование»;
ВИРТУАЛЬНАЯ ПРОГУЛКА по лучшим музеям мира.
О проведении мероприятий социально-экономической направленности
ГТО. Таблица нормативов» (2015 г.) (1-11 класс)
ГТО: Виды испытаний (1-4 класс)
Всероссийский урок «Готов к труду и обороне» (1-4 класс)
От ГТО к олимпийским медалям (1-11 класс)
Методические рекомендации к Единому уроку безопасности в сети Интернет
Видеоурок «Единый урок Интернет»
Дополнительный материал к уроку: «Развлечение и безопасность в сети Интернет», http://www.azbez.com/, «Безопасность в Интернете», «Базовые правила поведения в социальных сетях»
Методический материал «Электробезопасность»
Материал для родительского собрания (с презентацией) «Компьютерная зависимость детей: признаки, причины и последствия»
Начальная школа
Технологическая карта внеурочного занятия «Смотрю на мир глазами художника»/ Автор Пашнина Ю.Н.
Презентация внеурочного занятия «Смотрю на мир глазами художника»/ Автор Пашнина Ю.
Публикация на сайте «Копилка уроков» открытого внеклассного занятия «Дружба всего дороже»/ Автор Черненко Н.В.
Публикация на сайте «Копилка уроков» выступления на педсовете о профориентации в нач. школе «На пути к выбору своей мечты»/ Автор Черненко Н.В.
Глагол. Общее понятие (повторение). Конспект урока для 4 класса (формат doc), учитель Пашнина Ю.Н.
Работа со слабыми учениками на уроках, автор Абрамовских (формат ppt)
Интеллектуальное развитие младших школьников через использование на уроках словарно-орфографической работы, учитель Мещерякова Е.А. (формат 7z)
Развитие орфографической зоркости на уроках русского языка, учитель Абрамовских Г. Н. (формат doc)
Русский язык и литература
Программа элективного курса «Современная русская литература», автор Злодеева Ю.В. (>>> скачать, формат doc)
Средства формирования универсальных учебных действий учащихся 5-6 классов на уроках русского языка и литературы, учитель Злодеева Ю. В. (формат 7z)
В. (формат 7z)
Иностранный язык
Разработка внеклассного мероприятия и презентация для 7 класса «Своя игра» по английскому языку на тему «Англо-говорящие страны»./ Автор Товкалова О.А.
Урок+презентация по английскому языку для 9 класса «Поезда. Артикли с географическими названиями»/ Автор Товкалова О.А.
Контрольная работа по английскому языку на тему «Международный конкурс подростков» (7 класс), глава 1/ Автор: Товкалова О.А.
КОНСПЕКТ УРОКА АНГЛИЙСКОГО ЯЗЫКА В 9 КЛАССЕ НА ТЕМУ «ПРОБЛЕМЫ ВЫБОРА ПРОФЕССИИ ПОДРОСТКАМИ РОССИИ» (VII Всероссийский фестиваль методических разработок «Конспект урока» февраль-апрель 2016 год, электронный СМИ «Конференц-зал», электронный журнал)/ Автор: Товкалова О.А.
Методические разработки по немецкому и английскому языку. Автор Киселев В.А.
Разработка внеклассного мероприятия по немецкому языку «Пасха», автор Федотова А.А.(>>> скачать, формат zip)
Разработка урока для 7 класса «Образование в Британии», учитель Юдина Е.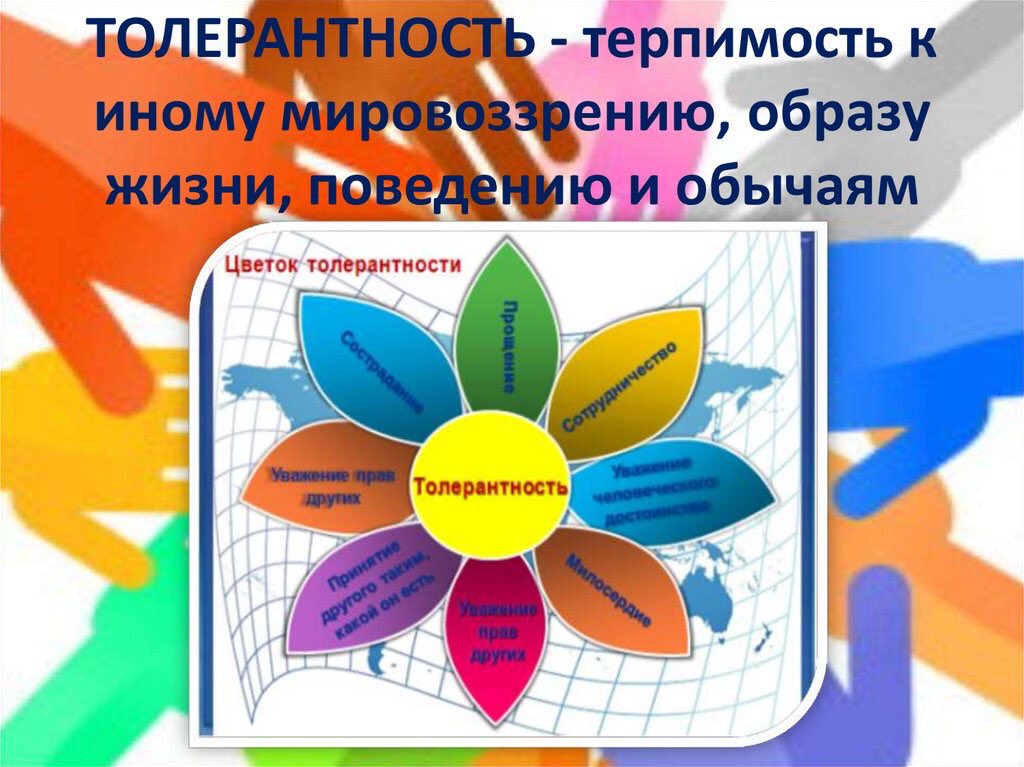 И (формат doc).
И (формат doc).
Формирование универсальных учебных действий обучающихся посредством применения метода проектов в обучении английскому языку, учитель Юдина Е.И.(формат doc).
Формирование социокультурной компетенции учащихся на уроках английского языка в средней школе, учитель Сударь А.С. (формат 7z)
Математика
Целенаправленное развитие математических способностей, автор Фахритдинова А.М. (формат doc)
Разработка мультимедийного урока по математике для 5 класса «Повторение за курс начальной школы. Путешествие в страну Математика», автор Бобылева Е.А.(>>> скачать, формат rar)
Презентация к уроку «Правильные выпуклые многогранники», 10 класс, автор Бобылева Е.А. (>>>скачать, формат ppt)
Разработка урока «Введение в стереометрию», 10 класс, автор Фахритдинова А.М. (>>>скачать, формат rar)
Тест по математике в режиме on-line для 6 класса (тест №1, тест №2 , тест №3 , тест №4), автор Бобылева Е. А.
А.
Тематические тесты для 7 класса в режиме on-line, автор Бобылева Е.А.:
Алгебра:
Работа №1 «Решение линейных уравнений»
Работа №2 «Решение задач с помощью уравнений»
Работа №3 «Функция»
Геометрия:
Работа №1 «Смежные и вертикальные углы»
Работа №2 «Первый признак равенства треугольников»
Информатика
Презентация к уроку «Программы и файлы», 5 класс, автор Бобылева Е.А. (>>>скачать, формат zip)
Презентация к уроку «Графический редактор Gimp», 9 класс, автор Бобылева Е.А. (>>>скачать, формат ppt)
Биология
Урок-игра по теме «Бактерии» (формат doc), автор Истомина Е.А.
Урок « Земноводные – необычные животные» (формат doc), автор Истомина Е.А.
Урок «Основы систематики растений» — 6 класс (формат doc), автор Истомина Е.А.
Урок по профилактике «Наркомания: проблемы и пути выхода» (формат doc), автор Истомина Е. А.
А.
Физика
Урок по теме «Оптические явления» — 8 класс , автор Шафикова Ф.З. (формат rar)
История
Обобщение опыта работы по теме «Организация спмостоятельной работы на уроках истории», учитель Самкова Н.А. (формат doc)
ОБЖ
Обобщение опыта работы «Использование игровых технологий на уроках ОБЖ», учитель Санатина О.А. (формат rtf)
Внеклассная работа
Классный час (презентация) «Георгий Дюперрон — родоначальник Российского футбола», автор Бобылева Е.А. (>>>скачать, формат rar)
Учебно-исследовательская деятельность актива школьного музея, типология творческих работ, автор Буханова Т.П.(формат doc)
Воспитание нравственности — интегральный компонент системы деятельности классного руководителя в начальной школе, автор Пашнина Ю.Н. (формат doc)
Социальное проектирование в деятельности спортивного клуба общеобразовательного учреждения, автор Панарина Р. И. (формат doc)
И. (формат doc)
Достижение учащимися личностных результатов обучения в ходе реализации программы воспитания и социализации обучающихся «Лестница успеха», автор Санатина О.А. (формат doc)
К социальному проектированию образовательного процесса в контексте требований Федерального Государственного образовательного стандарта начального общего образования, автор Шохова И.В. (формат doc)
Формирование толерантности на основе осознания себя как гражданина определенной культуры в рамках проекта«Ideal Welfare State: Social Programmes», автор Киселев В.А. (формат doc)
Развитие качеств лидера в условиях творческого коллектива дополнительного образования, руководитель танцевального коллектива «Сударушка» Белобородова Е.Ю. (формат doc)
Классный час для 5 класса «Пазлы одной картинки», автор Федотова А.А., школьный психолог (формат doc)
Организация и развитие ученического коллектива как среды, обеспечивающей развитие каждого ребенка, автор Федотова А.А., классный руководитель 5а класса (формат ppt)
Профилактика вредных привычек, учитель Истомина Е.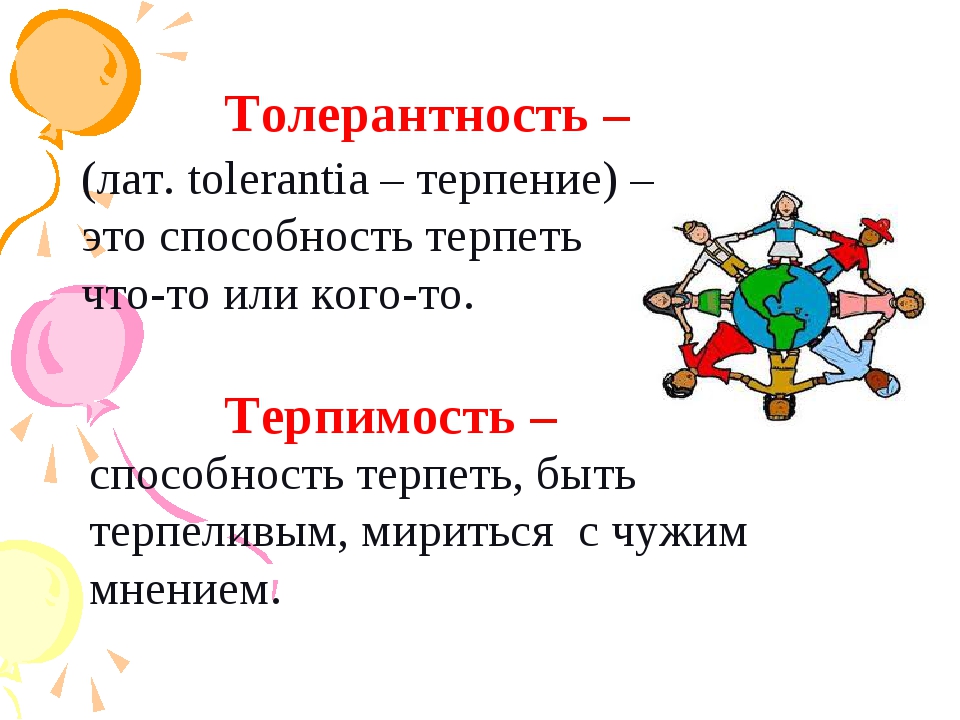 А.
А.
Легкая школа для трудных детей (формат ppt) Войкова Н.А., социальный педагог
Профилактика беспризорности (формат ppt), Войкова Н.А., социальный педагог
Холокост – преступление против человечества
27 января – Международный день памяти жертв Холокоста. В этот день традиционно проходят мемориальные мероприятия в память о более чем 6 млн евреев, ставших жертвами нацистов. Главной целью таких мероприятий является формирование мировоззрения людей, нацеленного на предупреждение и недопущение подобных событий в будущем, на борьбу с предрассудками и воспитание толерантности к традициям и культуре других народов. Холокост – страшный урок всему человечеству.
Предлагаем вашему вниманию материалы для проведения тематического занятия «Холокост – преступление против человечества».
Авторы: И. В. Гусенко, Т. В. Рубаник, Г. Р. Царева
Рекомендации по работе с презентацией тематического занятия «Холокост – преступление против человечества»
для обучающихся 9–11-х классов
Вариант проведения занятия [PDF] [DOCX]
Презентация [PDF] [PPTX]
Задания для обучающихся (рабочий лист) [PDF] [DOCX]
Цель: формирование у обучающихся нравственного и толерантного сознания на основе усвоения общечеловеческих ценностей.
Задачи:
- осмысление обучающимися нацистской политики Холокоста, её человеконенавистнической сущности и проявлений; а также событий, связанных с Холокостом, на территории СССР и стран Европы, с освободительной миссией Советского Союза, разгромившего войска Третьего рейха и уничтожившего нацизм;
- воспитание у обучающихся чувства патриотизма и гражданской ответственности;
- развитие умения аргументированно высказывать свою точку зрения;
- формирование гуманистических качеств личности.
Методический материал носит рекомендательный характер; учитель, принимая во внимание особенности каждого класса, может варьировать задания, их количество, менять этапы занятия.
27 января – Международный день памяти жертв Холокоста. Он установлен Генеральной Ассамблеей ООН 1 ноября 2005 г. (Резолюция № 60/7). Инициаторами принятия документа выступили Израиль, Канада, Австралия, Россия, Украина, США, а их соавторами — ещё более 90 государств.
Задание. Прочитайте текст Резолюции Генеральной Ассамблеи ООН и ответьте на вопросы.
С чем было связано принятие данной резолюции?
Как вы думаете, почему и сегодня, спустя более чем 70 лет после происходивших событий, эта тема продолжает быть важной?
27 января 1945 года Красная Армия освободила крупнейший нацистский лагерь смерти «Аушвиц» («Освенцим»).
Задание № 1 (рабочий лист). Прочитайте определение понятия «Холокост».
Подберите определения терминам, необходимым для обсуждения темы занятия. Занесите термины в таблицу.
Задание. Посмотрите видеофрагмент, ответьте на вопросы и выполните задания (задания 2, 3 в рабочем листе).
Вопросы:
1. Подумайте, почему политика нацистов по преследованию евреев характеризуется как государственная и последовательная.
2. В фильме звучат слова, которые еще в XIX веке произнес немецкий поэт Генрих Гейне: «Там, где сжигают книги, будут жечь людей». Какой исторический опыт привёл Гейне к такому утверждению? Почему это высказывание использовали авторы сюжета, который посвящён событиям XX века?
Какой исторический опыт привёл Гейне к такому утверждению? Почему это высказывание использовали авторы сюжета, который посвящён событиям XX века?
3. Что имели в виду авторы сюжета, говоря: «Привычный звук отбывающего поезда, спустя время, может изменить свое значение»?
Проанализируйте факты, посмотрите видеофрагмент и ответьте на вопросы.
Хронология Холокоста до начала Второй мировой войны
— 30 января 1933 г. – Гитлер назначен канцлером Германии – главой правительства.
— 22 марта 1933 г. – Создание лагеря Дахау.
— 1 апреля 1933 г. – Бойкот еврейских предприятий по всей Германии.
— 10 мая 1933 г. – Публичное сожжение произведений еврейских авторов и антинацистских книг.
— 14 июля 1933 г. – Начало принудительной стерилизации «неполноценных» граждан Германии (больных шизофренией, эпилепсией и т. д.).
— 15 сентября 1935 г. – Принятие антиеврейских Нюрнбергских законов.
— 29 марта 1936 г. – НСДАП получает 98 % голосов на выборах в рейхстаг.
— 19 июля 1937 г. – Создание концентрационного лагеря в Бухенвальде.
— 5 октября 1938 г. – Введение особых отметок «J» в паспорта евреев.
— 9 ноября 1938 г. – «Хрустальная ночь», массовые антиеврейские погромы по всей Германии.
— 24 января 1939 г. – Геринг создаёт «Центральную имперскую службу по делам еврейской эмиграции».
— 9 февраля 1939 г. – принятие антиеврейских законов в Италии.
Вопросы.
1. Почему события, происходившие в ноябре 1938 г., получили название «Хрустальная ночь»?
2. Нацисты использовали в своей терминологии понятия «всеобщее» и «окончательное решение еврейского вопроса». Предположите, что под этим имелось в виду.
3. Как вы думаете, были ли происходившие события только внутренним делом Германии?
4. Как проводившаяся западными государствами «политика умиротворения агрессора» влияла на действия нацистской Германии?
Систематическое уничтожение мирного еврейского населения нацистами началось сразу же после нападения гитлеровской Германии на Советский Союз. В нём участвовали 4 айнзатцгруппы СС: «А», «В», «С» и «D», приданные соответствующим группам войск вермахта, полицейские батальоны СС и тыловых частей вермахта, местные коллаборационисты, союзники нацистской Германии. Айнзатцгруппы уничтожали всех евреев в сельской местности, а также в городах, в зоне немецкой военной администрации (восточнее Днепра). Уничтожение зачастую проводилось в самих населённых пунктах на глазах у других жителей. Всемирную известность получило урочище Бабий Яр на окраине Киева, где 29–30 сентября 1941 г. было уничтожено несколько десятков тысяч евреев.
В нём участвовали 4 айнзатцгруппы СС: «А», «В», «С» и «D», приданные соответствующим группам войск вермахта, полицейские батальоны СС и тыловых частей вермахта, местные коллаборационисты, союзники нацистской Германии. Айнзатцгруппы уничтожали всех евреев в сельской местности, а также в городах, в зоне немецкой военной администрации (восточнее Днепра). Уничтожение зачастую проводилось в самих населённых пунктах на глазах у других жителей. Всемирную известность получило урочище Бабий Яр на окраине Киева, где 29–30 сентября 1941 г. было уничтожено несколько десятков тысяч евреев.
В зоне гражданской администрации было создано несколько сот гетто, самые крупные из которых в Минске, Каунасе и Вильнюсе просуществовали до середины 1943 г.
Выполните задание № 4 в рабочем листе.
Задание. Изучите карту, на которой отмечены самые крупные нацистские лагеря и места массового расстрела, и выполните задания.
Вопросы.
1. С какой целью нацисты создавали концентрационные лагеря?
С какой целью нацисты создавали концентрационные лагеря?
2. В каких странах располагались концентрационные лагеря?
3. Лагерь «Освенцим» был назван «фабрикой смерти». Как вы думаете, почему?
4. На входе нацистских концентрационных лагерей размещались лозунги, такие как «Каждому своё» (лагерь «Бухенвальд»), «Труд освобождает» (лагеря «Освенцим», «Терезин», «Заксенхаузен» и др.). Какой смысл вкладывали нацисты в эти фразы с точки зрения фашистской идеологии?
Задание № 5 в рабочем листе.
Изучите информацию о составе и категориях узников одного из концентрационных лагерей и ответьте на вопросы.
1. Подтверждают ли эти данные ваши предположения о том, какие народы стояли в списке нацистов первыми на уничтожение?
2. Ограничивался ли этот список только национальностью?
3. Что еще воспринималось нацистами как основание для уничтожения тех или иных категорий людей?
Символом еврейского Сопротивления стало восстание в Варшавском гетто, начавшееся 19 апреля 1943 года, первое городское восстание в оккупированной нацистами Европе. К его годовщине приурочен День памяти евреев – жертв нацизма и героев Сопротивления, отмечаемый ежегодно во всех еврейских общинах мира. Восстание продолжалось несколько недель, почти все его участники погибли с оружием в руках. Успешным было восстание и побег нескольких сот узников из лагеря смерти Собибор, организованное советским военнопленным евреем А. Печерским. Подпольные группы, организовавшие вооруженное сопротивление, а также побеги узников и снабжение партизан оружием и медикаментами, существовали в минском, каунасском, белостокском, виленском гетто.
К его годовщине приурочен День памяти евреев – жертв нацизма и героев Сопротивления, отмечаемый ежегодно во всех еврейских общинах мира. Восстание продолжалось несколько недель, почти все его участники погибли с оружием в руках. Успешным было восстание и побег нескольких сот узников из лагеря смерти Собибор, организованное советским военнопленным евреем А. Печерским. Подпольные группы, организовавшие вооруженное сопротивление, а также побеги узников и снабжение партизан оружием и медикаментами, существовали в минском, каунасском, белостокском, виленском гетто.
Вопрос: О чём свидетельствует факт выступлений против нацистов узников гетто и концентрационных лагерей?
Задание.
27 января 1945 г. Красная Армия освободила самый страшный лагерь смерти «Аушвиц» («Освенцим»). В освобождении лагеря и его филиалов принимало участие не менее 25 тысяч человек более 39 национальностей.
Пройдите по ссылке, изучите приведённую на сайте информацию и выполните задания. Рассмотрите карту, на которой отмечены биографии воинов-освободителей лагеря «Освенцим – Аушвиц».
Рассмотрите карту, на которой отмечены биографии воинов-освободителей лагеря «Освенцим – Аушвиц».
1. Как вы думаете, какую цель поставили перед собой создатели карты?
2. Расскажите об одном из героев-освободителей. Прочитайтеписьмо советского коменданта освобождённого Освенцима Г. Елисаветинского и телеграмму об освобождении лагеря.
3. Какие чувства, по вашему мнению, испытали советские солдаты, освобождавшие «Освенцим»?
Задание.
Прочитайте известную цитату из выступлений немецкого пастора Мартина Нимёллера:
«Когда нацисты хватали коммунистов, я молчал: я не был коммунистом. Когда они сажали социал-демократов, я молчал: я не был социал-демократом. Когда они хватали профсоюзных активистов, я молчал: я не был членом профсоюза. Когда они пришли за мной – уже некому было заступиться за меня».
1. Как вы думаете, оправдывал себя пастор Мартин Нимёллер или осуждал? Аргументируйте свою точку зрения.
2. Что, по вашему мнению, пастор Мартин Нимёллер хотел донести до своих слушателей?
Задание № 6 в рабочем листе.
Тему равнодушия и его последствий затрагивали в своих рассуждениях многие философы, писатели, общественные деятели. Прочитайте цитаты и ответьте на вопросы.
1. Какие из этих цитат наиболее созвучны мыслям пастора Нимёллера?
2. Согласны ли вы с позицией авторов?
3. Какие из высказываний, по вашему мнению, наиболее точно отражают пагубность безразличия, безучастности, невозмутимости по типу «оставьте меня в покое» к окружающим людям и происходящим событиям?
Несмотря на все ужасы тех условий, в которых находились люди на оккупированных территориях, многие не могли оставаться равнодушными. Это и русские семьи, выдававшие еврейских детей за своих, и польские крестьяне, укрывавшие раненых партизан, и немецкие офицеры, спасавшие узников концлагерей, и советские пленные, возглавляющие восстания в лагерях смерти. Имена многих из них неизвестны.
Задание № 7 в рабочем листе.
1. Что объединяет всех этих людей?
2. Как вы думаете, что было самым трудным для принятия решения помогать или нет?
3. Какие вопросы вы могли бы задать этим людям при встрече?
Какие вопросы вы могли бы задать этим людям при встрече?
Задание.
Посмотрите видеоряд и выполните задания. Ответьте на вопросы.
1. С какой целью по всему миру устанавливаются мемориалы жертвам Холокоста?
2. Согласны ли вы с утверждением немецкого писателя, антифашиста Арнольда Иоганна Бауэра?
«Память о Холокосте необходима, чтобы наши дети никогда не были жертвами, палачами или равнодушными наблюдателями». А.И. Бауэр
3. Как вы понимаете высказывание советского историка и философа Михаила Яковлевича Гефтера?
«Нет геноцида против «кого-то», геноцид всегда против всех». М. Я. Гефтер
Задание.
Приведите аргументы, подтверждающие высказывание «Холокост – преступление против человечества».
*Яд ва-Шем — израильский национальный мемориал Катастрофы (Холокоста) и Героизма, который находится в Иерусалиме.
Задание. Озвучьте виртуальную экскурсию, используя материалы занятия.
Дополнительные материалы
— ФГАОУ ДПО АПК и ППРО: «Методические рекомендации по организации и проведению тематических мероприятий, посвящённых Международному дню памяти жертв Холокоста (27 января)»
— Сайт «Центр и Фонд «Холокост»
— Сайт «Освободители»
— Электронная версия историко-документальной выставки «Холокост: уничтожение, освобождение, спасение»
— Классный час «Подвиг Александра Печерского»
| «КАК ВОСПИТАТЬ ТОЛЕРАНТНОГО ЧЕЛОВЕКА?»
Проблемой обсуждения с родителями является вопрос «Как воспитать толерантного человека?». Толерантность человека, выходящего в большую жизнь, является фактором социализации и в значительной мере определяет успешность жизненного пути человека. Именно поэтому важнейшей задачей является формирование у подрастающего поколения умения строить взаимоотношения в процессе взаимодействия с окружающими на основе сотрудничества и взаимопонимания, готовности принять других людей и их взгляды, привычки такими, какие они есть. Проблема, которая станет предметом нашего обсуждения, весьма актуальна «Как воспитать толерантного человека?» Необходимость ее решения продиктована тем, что толерантность человека, выходящего в большую жизнь, является фактором социализации и в значительной мере определяет успешность жизненного пути человека. Толерантность – понятие достаточно новое, это: Толерантность предполагает уважение к самобытному внутреннему миру ребенка, веру в победу доброго начала в межличностных отношениях, отказ от методов грубого понуждения и любых форм авторитаризма, позитивную лексику. Подросток осваивает социальные связи и роли в мире; становясь старше, он все больше ориентируется на ровесников и социальные институты. Родители, давая ему относительную свободу, должны быть надежным тылом для него, создавать у него чувство уверенности в трудной ситуации. Воспитывать толерантность в семье — значит прививать уважение, принятие и правильное понимание богатого многообразия культур нашего мира, наших форм самовыражения и способов проявлений человеческой индивидуальности. Тест для родителей «Пытаетесь ли Вы понять своего ребенка?» Из трех ответов на каждый вопрос выберите один, затем подсчитайте, Вы оценили наедине с собой свою собственную толерантность и, возможно, теперь измените характер воспитательных действий в отношении своего ребенка. Если ребенка постоянно критиковать, он учится ненавидеть. 2. Непоследовательность родителей в обучении детей правилам и нормам поведения. У детей формируется нравственный стержень поведения: сегодня родителям удобно говорить одно, и они же навязывают эту линию поведения детям, завтра им удобно говорить другое, и это другое тоже навязывается. 1. Учитесь слушать и слышать своего ребенка. Китайская притча «Ладная семья» Правила воспитания в духе толерантности Секрет толерантности Если ребенка постоянно критиковать, он учится ненавидеть. 1. Учитесь слушать и слышать своего ребенка. |
 Поэтому для начала необходимо самим относиться к своему ребенку толерантно. Во-первых, не обижать его. Во-вторых, выслушивать его мнение и считаться с ним. В-третьих, уметь прощать обиды и просить прощения у ребенка. Это самый сложный момент, но в то же время и самый важный, поскольку у детей обостренное чувство справедливости. В-четвертых, уметь договариваться без ссор и разрушительных конфликтов. Под словосочетанием «разрушительный конфликт» подразумевается противостояние, которое наносит «противнику» ущерб, моральный или физический. Разумеется, что в повседневной жизни невозможно избежать противоречий интересов, желаний и мнений. Задача – обратить конфликт в конструктивный, вызвав ребенка к разговору о возникших противоречиях и к совместному принятию компромиссных решений. В-пятых, нельзя унижать достоинство ребенка – игнорировать его, проявлять неуважение к его увлечениям и т.п. В-шестых, не стоит заставлять ребенка с помощью силы делать то, что хочется вам.
Поэтому для начала необходимо самим относиться к своему ребенку толерантно. Во-первых, не обижать его. Во-вторых, выслушивать его мнение и считаться с ним. В-третьих, уметь прощать обиды и просить прощения у ребенка. Это самый сложный момент, но в то же время и самый важный, поскольку у детей обостренное чувство справедливости. В-четвертых, уметь договариваться без ссор и разрушительных конфликтов. Под словосочетанием «разрушительный конфликт» подразумевается противостояние, которое наносит «противнику» ущерб, моральный или физический. Разумеется, что в повседневной жизни невозможно избежать противоречий интересов, желаний и мнений. Задача – обратить конфликт в конструктивный, вызвав ребенка к разговору о возникших противоречиях и к совместному принятию компромиссных решений. В-пятых, нельзя унижать достоинство ребенка – игнорировать его, проявлять неуважение к его увлечениям и т.п. В-шестых, не стоит заставлять ребенка с помощью силы делать то, что хочется вам.
 Именно она удовлетворяет все потребности малыша. Но, между тем, присутствие отца тоже имеет важное значение. Во-первых, именно он обеспечивает безопасность матери и ребенка и именно от его поведения зависит самочувствие матери. Во-вторых, ребенок смотрит на взаимоотношения родителей и то, что он видит, оказывает влияние на его будущую семью, дальнейшую жизнь и здоровье. Всем известно, что дочь надо воспитывать женственной и заложить в ней ценности материнства, а сыну следует привить чувство ответственности. Но самое главное – научить девочку благодарности, а мальчика заботливости. Ведь, если мальчик заботлив, то, став мужчиной, во имя достижения благополучия близких, он приложит все свои усилия, проявив и ум, и силу, и другие мужские качества. А благодарность – самое важное женское качество. Поскольку, если женщина благодарна, то хочется сделать для нее нечто большее, чем уже сделано и хочется ее радовать.
Именно она удовлетворяет все потребности малыша. Но, между тем, присутствие отца тоже имеет важное значение. Во-первых, именно он обеспечивает безопасность матери и ребенка и именно от его поведения зависит самочувствие матери. Во-вторых, ребенок смотрит на взаимоотношения родителей и то, что он видит, оказывает влияние на его будущую семью, дальнейшую жизнь и здоровье. Всем известно, что дочь надо воспитывать женственной и заложить в ней ценности материнства, а сыну следует привить чувство ответственности. Но самое главное – научить девочку благодарности, а мальчика заботливости. Ведь, если мальчик заботлив, то, став мужчиной, во имя достижения благополучия близких, он приложит все свои усилия, проявив и ум, и силу, и другие мужские качества. А благодарность – самое важное женское качество. Поскольку, если женщина благодарна, то хочется сделать для нее нечто большее, чем уже сделано и хочется ее радовать. Вопрос лишь в том, отложится это в сознании или в подсознании. Воспитывать ребенка личным примером сложнее всего, но в тоже время и наиболее эффективно. Для того чтобы дочь выросла женственной, матери необходимо самой быть такою и учить ее опрятности и женской работе по дому. Большую роль в жизни девочки играет и отец, поскольку от того, как он с ней общается, зависит то, каких взаимоотношений она будет искать с мужчинами, когда вырастет. Сына-подростка следует воспитывать в спокойной строгости. Разумеется, это не означает, что его не надо любить.
Вопрос лишь в том, отложится это в сознании или в подсознании. Воспитывать ребенка личным примером сложнее всего, но в тоже время и наиболее эффективно. Для того чтобы дочь выросла женственной, матери необходимо самой быть такою и учить ее опрятности и женской работе по дому. Большую роль в жизни девочки играет и отец, поскольку от того, как он с ней общается, зависит то, каких взаимоотношений она будет искать с мужчинами, когда вырастет. Сына-подростка следует воспитывать в спокойной строгости. Разумеется, это не означает, что его не надо любить.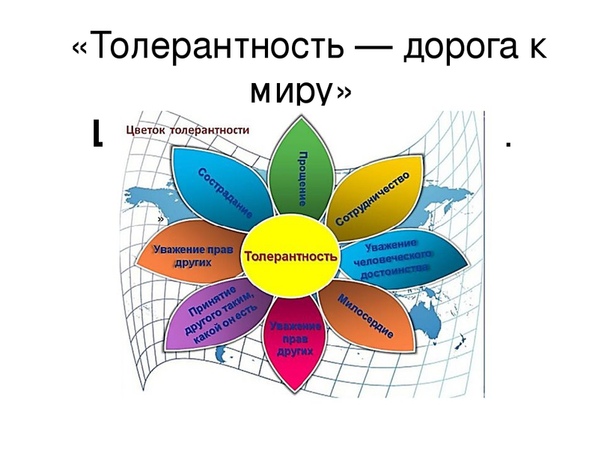 При в формировании толерантности этот принцип приобретает двойную значимость. Уважая и принимая позицию и мнение ребенка, при необходимости корректируя их, мы показываем ему пример толерантного отношения к человеку с иным взглядом на мир.
При в формировании толерантности этот принцип приобретает двойную значимость. Уважая и принимая позицию и мнение ребенка, при необходимости корректируя их, мы показываем ему пример толерантного отношения к человеку с иным взглядом на мир.
 Природа не терпит пустоты. Если мы не заполним его душу чем-то добрым, найдется кто-то другой, который заполнит ее дурным.
Природа не терпит пустоты. Если мы не заполним его душу чем-то добрым, найдется кто-то другой, который заполнит ее дурным.
 Поэтому в последнее время все чаще и чаще возникают разговоры о толерантном мире, т.е. мире без насилия и жестокости, мире, в котором самой главной ценностью является неповторимая и неприкосновенная человеческая личность. Но просто разбрасываться красивыми словами, конечно, недостаточно. Толерантность нужно воспитывать. Отсюда – ряд правил воспитания, связанные с темой нашего собрания:
Поэтому в последнее время все чаще и чаще возникают разговоры о толерантном мире, т.е. мире без насилия и жестокости, мире, в котором самой главной ценностью является неповторимая и неприкосновенная человеческая личность. Но просто разбрасываться красивыми словами, конечно, недостаточно. Толерантность нужно воспитывать. Отсюда – ряд правил воспитания, связанные с темой нашего собрания: А что же для этого нужно?
А что же для этого нужно? Три слова были начертаны на бумаге:
Три слова были начертаны на бумаге:

Классы международного допуска (IT) ISO 286-1 — 2010 (E) Таблица
Таблица классов международных допусков (IT) ISO 286-1
Меню стандартов механических допусков
Ссылка на таблицу классов международных допусков (IT) ISO 286. Этот класс механических допусков определяет допуски для соответствующих производственных процессов для заданного размера.
Связано: Формулы международных классов допуска:
Заявка:
Большинство производственных процессов имеют степень IT, установленную для спецификации.Уровни ИТ дают представление о типичных возможностях производственного процесса или о том, насколько точен он может, за исключением изготовления определенных функций или функций.
При проектировании детали и указании механических допусков инженеры или проектировщики определяют размер и расположение конкретной детали. С помощью указанной формулы International Tolerance Grade назначается IT-класс, необходимый или типичный для изготовления элемента детали. Например, для литья пластмасс под давлением определен IT-уровень 13, а для детали — IT-класс 5, при проектировании и проектировании следует рассмотреть альтернативный и более производительный процесс производства детали и элемента.
С помощью указанной формулы International Tolerance Grade назначается IT-класс, необходимый или типичный для изготовления элемента детали. Например, для литья пластмасс под давлением определен IT-уровень 13, а для детали — IT-класс 5, при проектировании и проектировании следует рассмотреть альтернативный и более производительный процесс производства детали и элемента.
| Номинал |
Классы стандартных допусков, мкм |
|||||||
|---|---|---|---|---|---|---|---|---|
| IT01 |
IT0 |
IT1 |
IT2 |
IT3 |
IT4 |
IT5 |
||
| Выше |
До |
мкм |
||||||
| — |
3 |
0,3 |
0,5 |
0,8 |
1,2 |
2 |
3 |
4 |
| 3 |
6 |
0,4 |
0,6 |
1 |
1,5 |
2,5 |
4 |
5 |
| 6 |
10 |
0,4 |
0,6 |
1 |
1,5 |
2,5 |
4 |
6 |
| 10 |
18 |
0,5 |
0,8 |
1,2 |
2 |
3 |
5 |
8 |
| 18 |
30 |
0,6 |
1 |
1,5 |
2,5 |
4 |
6 |
9 |
| 30 |
50 |
0,6 |
1 |
1,5 |
2,5 |
4 |
7 |
11 |
| 50 |
80 |
0,8 |
1,2 |
2 |
3 |
5 |
8 |
13 |
| 80 |
120 |
1 |
1,5 |
2,5 |
4 |
6 |
10 |
15 |
| 120 |
180 |
1,2 |
2 |
3,5 |
5 |
8 |
12 |
18 |
| 180 |
250 |
2 |
3 |
4,5 |
7 |
10 |
14 |
20 |
| 250 |
315 |
2,5 |
4 |
6 |
8 |
12 |
16 |
23 |
| 315 |
400 |
3 |
5 |
7 |
9 |
13 |
18 |
25 |
| 400 |
500 |
4 |
6 |
8 |
10 |
15 |
20 |
27 |
| 500 |
630 |
– |
– |
9 |
11 |
16 |
22 |
32 |
| 630 |
800 |
– |
– |
10 |
13 |
18 |
25 |
36 |
| 800 |
1 000 |
– |
– |
11 |
15 |
21 |
28 |
40 |
| 1 000 |
1 250 |
– |
– |
13 |
18 |
24 |
33 |
47 |
| 1 250 |
1 600 |
– |
– |
15 |
21 |
29 |
39 |
55 |
| 1 600 |
2 000 |
– |
– |
18 |
25 |
35 |
46 |
65 |
| 2 000 |
2 500 |
– |
– |
22 |
30 |
41 |
55 |
78 |
| 2 500 |
3 150 |
– |
– |
26 |
36 |
50 |
68 |
96 |
| Номинал |
Классы стандартных допусков мкм | ||||||
|---|---|---|---|---|---|---|---|
| IT6 |
IT7 |
IT8 |
IT9 |
IT10 |
IT11 |
||
| Выше |
До |
мкм | |||||
| — |
3 |
6 |
10 |
14 |
25 |
40 |
60 |
| 3 |
6 |
8 |
12 |
18 |
30 |
48 |
75 |
| 6 |
10 |
9 |
15 |
22 |
36 |
58 |
90 |
| 10 |
18 |
11 |
18 |
27 |
43 |
70 |
110 |
| 18 |
30 |
13 |
21 |
33 |
52 |
84 |
130 |
| 30 |
50 |
16 |
25 |
39 |
62 |
100 |
160 |
| 50 |
80 |
19 |
30 |
46 |
74 |
120 |
190 |
| 80 |
120 |
22 |
35 |
54 |
87 |
140 |
220 |
| 120 |
180 |
25 |
40 |
63 |
100 |
160 |
250 |
| 180 |
250 |
29 |
46 |
72 |
115 |
185 |
290 |
| 250 |
315 |
32 |
52 |
81 |
130 |
210 |
320 |
| 315 |
400 |
36 |
57 |
89 |
140 |
230 |
360 |
| 400 |
500 |
40 |
63 |
97 |
155 |
250 |
400 |
| 500 |
630 |
44 |
70 |
110 |
175 |
280 |
440 |
| 630 |
800 |
50 |
80 |
125 |
200 |
320 |
500 |
| 800 |
1 000 |
56 |
90 |
140 |
230 |
360 |
560 |
| 1 000 |
1 250 |
66 |
105 |
165 |
260 |
420 |
660 |
| 1 250 |
1 600 |
78 |
125 |
195 |
310 |
500 |
780 |
| 1 600 |
2 000 |
92 |
150 |
230 |
370 |
600 |
920 |
| 2 000 |
2 500 |
110 |
175 |
280 |
440 |
700 |
1 100 |
| 2 500 |
3 150 |
135 |
210 |
330 |
540 |
860 |
1 350 |
| Номинал |
Классы стандартных допусков мм |
|||||||
|---|---|---|---|---|---|---|---|---|
| IT12 |
IT13 |
IT14 |
IT15 |
IT16 |
IT117 |
IT18 |
||
| Выше |
До |
мм |
||||||
| — |
3 |
0,1 |
0,14 |
0,25 |
0,4 |
0,6 |
1 |
1,4 |
| 3 |
6 |
0,12 |
0,18 |
0,3 |
0,48 |
0,75 |
1,2 |
1,8 |
| 6 |
10 |
0,15 |
0,22 |
0,36 |
0,58 |
0,9 |
1,5 |
2,2 |
| 10 |
18 |
0,18 |
0,27 |
0,43 |
0,7 |
1,1 |
1,8 |
2,7 |
| 18 |
30 |
0,21 |
0,33 |
0,52 |
0,84 |
1,3 |
2,1 |
3,3 |
| 30 |
50 |
0,25 |
0,39 |
0,62 |
1 |
1,6 |
2,5 |
3,9 |
| 50 |
80 |
0,3 |
0,46 |
0,74 |
1,2 |
1,9 |
3 |
4,6 |
| 80 |
120 |
0,35 |
0,54 |
0,87 |
1,4 |
2,2 |
3,5 |
5,4 |
| 120 |
180 |
0,4 |
0,63 |
1 |
1,6 |
2,5 |
4 |
6,3 |
| 180 |
250 |
0,46 |
0,72 |
1,15 |
1,85 |
2,9 |
4,6 |
7,2 |
| 250 |
315 |
0,52 |
0,81 |
1,3 |
2,1 |
3,2 |
5,2 |
8,1 |
| 315 |
400 |
0,57 |
0,89 |
1,4 |
2,3 |
3,6 |
5,7 |
8,9 |
| 400 |
500 |
0,63 |
0,97 |
1,55 |
2,5 |
4 |
6,3 |
9,7 |
| 500 |
630 |
0,7 |
1,1 |
1,75 |
2,8 |
4,4 |
7 |
11 |
| 630 |
800 |
0,8 |
1,25 |
2 |
3,2 |
5 |
8 |
12,5 |
| 800 |
1 000 |
0,9 |
1,4 |
2,3 |
3,6 |
5,6 |
9 |
14 |
| 1 000 |
1 250 |
1,05 |
1,65 |
2,6 |
4,2 |
6,6 |
10,5 |
16,5 |
| 1 250 |
1 600 |
1,25 |
1,95 |
3,1 |
5 |
7,8 |
12,5 |
19,5 |
| 1 600 |
2 000 |
1,5 |
2,3 |
3,7 |
6 |
9,2 |
15 |
23 |
| 2 000 |
2 500 |
1,75 |
2,8 |
4,4 |
7 |
11 |
17,5 |
28 |
| 2 500 |
3 150 |
2,1 |
3,3 |
5,4 |
8,6 |
13,5 |
21 |
33 |
1) Значения для стандартных классов допуска IT01 и IT0 для основных размеров меньше или равных 500 мм приведены в IS0 286-1, приложение A, таблица 5.
2) Значения для стандартных классов допуска от IT1 до IT5 (включительно) для основных размеров более 500 мм включены для экспериментального использования.
3) Стандартные классы допуска от IT14 до IT18 (включительно) не должны использоваться для основных размеров меньше или равных 1 мм.
Связанные ресурсы:
Артикул:
ISO — Международная организация по стандартизации ISO 286-1 — 2010 (E)
|
окт.28 |
План урока: опрос SEL Слайд-дека: Нет Раздаточный материал: Нет |
План урока: опрос SEL Слайд-дека: Нет Раздаточный материал: Нет |
План урока: опрос SEL Слайд-дека: Нет Раздаточный материал: Нет |
План урока: опрос SEL Слайд-дека: Нет Раздаточный материал: Нет |
|
ноя. |
План урока: Урок по постановке целей G9.pdf Слайд-дека: Стенд для постановки целей G9 Раздаточный материал: G9-12 Рабочий лист постановки целей.docx |
План урока: Урок по постановке целей G10.pdf Slide Deck: G10 Goal Setting Deck Раздаточный материал: G9-12 Рабочий лист постановки целей.docx
|
План урока: Урок по постановке целей G11.pdf Slide Deck: G11 Установка целей Раздаточный материал: G9-12 Рабочий лист постановки целей.docx
|
План урока : Урок по постановке целей G12.pdf Slide Deck: G12 Goal Setting Deck Раздаточный материал: G9-12 Рабочий лист постановки целей.docx
|
|
24 ноября |
План урока : Урок по настройке портфолио G9.pdf Slide Deck: Настройка портфолио G9 Раздаточный материал: G9_Portfolio Setup Handout.pdf Годовые ожидания цифрового портфеля. |
План урока: G10 Урок баланса между школой и жизнью.pdf Слайд-колода: G10 School Life Balance Раздаточный материал: Раздаточный материал G10 о балансе школьной жизни.pdf |
План урока: G11 Mind Body Spirit.pdf Слайд-колода: G11 Mind Body Spirit Раздаточный материал: Раздаточный материал G11 Mind Body Spirit.docx |
План урока: G12 Financial Aid.pdf Слайд-колода: G12 Financial Aid Раздаточный материал: Раздаточный материал G12 по финансовой помощи.pdf |
| 9 декабря |
План урока: Gr. План урока 9 Study Skills Слайд-колода: G9 Study Skills Раздаточный материал: Gr. 9 Раздаточный материал по навыкам обучения |
План урока: G10 Portfolio 1 Expectations.pdf Slide Deck: Настройка портфолио G10 Раздаточный материал: Ежегодные ожидания цифрового портфеля.pdf |
План урока: G11 Portfolio 1 Expectations. Slide Deck: Настройка портфолио G11 Раздаточный материал: Ожидания от цифрового портфеля на год.pdf |
План урока: G12 Portfolio 1 Expectations.pdf Слайд-колода: Ожидания от портфеля G12 Раздаточный материал: Контрольный список цифрового портфолио Gr 12 и итоговые размышления.pdf Годовые ожидания цифрового портфеля |
| 23 декабря |
План урока: продолжение опроса SEL Слайд-дека: Нет Раздаточный материал: Нет |
План урока: продолжение опроса SEL Слайд-дека: Нет Раздаточный материал: Нет |
План урока: продолжение опроса SEL Слайд-дека: Нет Раздаточный материал: Нет |
План урока: продолжение опроса SEL Слайд-дека: Нет Раздаточный материал: Нет |
| Также декабрь.23 |
План урока : Урок по загрузке портфолио G9. Слайд-колода: Загрузки портфолио G9 Раздаточный материал: G9_Portfolio Setup Handout.pdf Годовые ожидания цифрового портфеля
|
План урока : Урок по загрузке портфолио G10.pdf Слайд-колода: G10 Загрузки портфолио Раздаточный материал: Ежегодные ожидания цифрового портфеля.pdf
|
План урока : загрузка портфолио G11.pdf Слайд-колода: G11 Загрузки портфолио Раздаточный материал: Ежегодные ожидания цифрового портфеля.pdf
|
План урока: загрузка портфолио G12.docx Слайд-колода: G12 Загрузки портфолио Раздаточный материал: Контрольный список для цифрового портфолио Gr 12 и итоговые размышления.pdf Годовые ожидания цифрового портфеля
|
| 6 января |
План урока: G9-12_ToleranceMicroaggressionsLesson.pdf Slide Deck: допуск G9-12 и микроагрессия Раздаточный материал: G9-12_ToleranceMicroaggressionsHandout. |
План урока: G9-12_ToleranceMicroaggressionsLesson.pdf Slide Deck: допуск G9-12 и микроагрессия Раздаточный материал: G9-12_ToleranceMicroaggressionsHandout.docx |
План урока: G9-12_ToleranceMicroaggressionsLesson.pdf Slide Deck: допуск G9-12 и микроагрессия Раздаточный материал: G9-12_ToleranceMicroaggressionsHandout.docx |
План урока: G9-12_ToleranceMicroaggressionsLesson.pdf Slide Deck: допуск G9-12 и микроагрессия Раздаточный материал: G9-12_ToleranceMicroaggressionsHandout.docx |
|
27 января |
План урока : Урок по загрузке портфолио G9.pdf Слайд-колода: Цифровое портфолио G9 загружено Раздаточный материал: G9_Portfolio Setup Handout.pdf Годовые ожидания цифрового портфеля |
План урока : Урок по загрузке портфолио G10.pdf Слайд-колода: Цифровое портфолио G10 загружено Раздаточный материал: Ежегодные ожидания цифрового портфеля. |
План урока: G11 Portfolio Uploads.pdf Слайд-колода: Цифровое портфолио G11 загружено Раздаточный материал: Ежегодные ожидания цифрового портфеля.pdf |
План урока: загрузка портфолио G12.docx Слайд-колода: Цифровое портфолио G12 загружено Раздаточный материал: Контрольный список цифрового портфолио Gr 12 и итоговые размышления.pdf Годовые ожидания цифрового портфеля |
|
12 февраля |
План урока: G9-12_CelebrationLesson.pdf Слайд-колода: G9 Celebration Раздаточный материал: G9-12_CelebrationHandout.pdf |
План урока: G9-12_CelebrationLesson.pdf Слайд-колода: G10 Celebration Раздаточный материал: G9-12_CelebrationHandout.pdf
|
План урока: G9-12_CelebrationLesson.pdf Слайд-колода: G11 Celebration Раздаточный материал: G9-12_CelebrationHandout.
|
План урока: G9-12_CelebrationLesson.pdf Слайд-колода: G12 Celebration Раздаточный материал: G9-12_CelebrationHandout.pdf
|
|
24 февраля` |
План урока : Урок по повторению целей G9-11.pdf Slide Deck: G9-11 Goal Review Раздаточные материалы: Форма академической рефлексии G9-11.docx G9-12 Рабочий лист постановки целей.docx |
План урока : Урок по повторению целей G9-11.pdf Slide Deck: G9-11 Goal Review Раздаточные материалы: G9-11 Форма для академических размышлений.docx G9-12 Рабочий лист постановки целей.docx
|
План урока : Урок по повторению целей G9-11.pdf Slide Deck: G9-11 Goal Review Раздаточные материалы: G9-11 Форма для академических размышлений.docx G9-11 Урок по обзору целей.pdf
|
пожилых людей примут участие в собрании старшего класса.
|
|
11 марта |
План урока: G9-12_SEL_BeThereLesson.pdf Слайд-колода: G9-12 Be There Ссылка на опрос: Исследование социальной осведомленности |
План урока: G9-12_SEL_BeThereLesson.pdf Слайд-колода: G9-12 Be There Ссылка на опрос: Исследование социальной осведомленности
|
План урока: G9-12_SEL_BeThereLesson.pdf Слайд-колода: G9-12 Be There Ссылка на опрос: Исследование социальной осведомленности
|
План урока: G9-12_SEL_BeThereLesson.pdf Слайд-колода: G9-12 Be There Ссылка на опрос: Исследование социальной осведомленности
|
|
26 марта |
Введение в карьерные возможности средней школы Уэстбрук |
Введение в карьерные возможности средней школы Уэстбрук
|
Введение в карьерные возможности средней школы Уэстбрук
|
План урока : загрузка портфолио G12. Слайд-колода: Цифровое портфолио G12 загружено Раздаточный материал: Контрольный список для цифрового портфолио Gr 12 и итоговые размышления.pdf Годовые ожидания цифрового портфеля
|
|
8 апреля |
Урок: Регистрация на курс Консультативные группыи удаленные учащиеся, присоединяйтесь к классной встрече Google Class of 2024. Раздаточные материалы: Программа обучения карьере.pdf PowerSchool registration.docx |
Урок: Регистрация на курс Консультативные группыи удаленные учащиеся, присоединяйтесь к классной встрече Google Class of 2023. Раздаточные материалы: Программа обучения по карьерной лестнице.pdf PowerSchool registration.docx |
Урок: Регистрация на курс Консультативные группыи удаленные учащиеся, присоединяйтесь к классной встрече Google Class of 2022. Раздаточные материалы: Программа обучения карьере.pdf PowerSchool registration.docx |
План урока: G12: вещи, которым не учили в старшей школе.pdf Слайд-колода: G12, чему не учили в старшей школе Раздаточные материалы: G12 Вещи, которым они не учили, в раздаточном материале HS.docx |
|
22 апреля |
Урок: Размышления и презентация портфолио G9-11.pdf Слайд-колода: отражение и презентация портфолио G9-11 Раздаточный материал: Ожидания от презентаций портфолио в 9–11 классах.pdf
|
Урок: Размышления и презентация портфолио G9-11.pdf Слайд-колода: отражение и презентация портфолио G9-11 Раздаточный материал: Ожидания от презентаций портфолио в 9–11 классах.pdf
|
Урок: Размышления и презентация портфолио G9-11.pdf Слайд-колода: отражение и презентация портфолио G9-11 Раздаточный материал: Ожидания от презентаций портфолио в 9–11 классах.pdf
|
Урок: Презентации портфолио G12.pdf Слайд-колода: Презентации портфолио G12 Раздаточный материал: Ожидания от презентаций портфолио для 12-го класса .pdf
|
|
6 мая |
Презентации портфолио |
Презентации портфолио |
Презентации портфолио |
Презентации портфолио |
|
20 мая |
Презентации портфолио |
Презентации портфолио |
Презентации портфолио |
Презентации портфолио |
 11
11  pdf
pdf  pdf
pdf  pdf
pdf  docx
docx  pdf
pdf  pdf
pdf  Чтобы получить ссылку на Google Meet, обратитесь к своему консультанту.
Чтобы получить ссылку на Google Meet, обратитесь к своему консультанту.  docx
docx Недостаточная презентация аутоантигена и нарушение толерантности в мышиной модели ревматоидного артрита
Arthritis Rheum.Авторская рукопись; доступно в PMC 2014 1 ноября 2014 г.
Опубликован в окончательной редакции как:
PMCID: PMC3
NIHMSID: NIHMS553318
, BS, 1 , MS, 1 , PhD, 1 , MD, 2 , PhD, 1 , MD, PhD, 2 and, PhD 1Jason Perera
1 Департамент медицины, Отдел ревматологии и Центр исследований волчанки и иммунологии им. Кнаппа, Университет Чикаго, Чикаго, Иллинойс
Сяо Лю
1 Департамент медицины, Отделение ревматологии и Центр исследований волчанки и иммунологии Кнаппа, Чикагский университет, Чикаго, Иллинойс
Yuzhen Zhou
1 Департамент медицины , Отделение ревматологии и Центр исследований волчанки и иммунологии Кнаппа, Чикагский университет, Чикаго, Иллинойс
Нора Э.Джозеф
2 Кафедра патологии, Чикагский университет, Чикаго, Иллинойс
Липинг Менг
1 Кафедра медицины, Отдел ревматологии и Центр исследований волчанки и иммунологии Кнаппа, Чикагский университет, Чикаго, Иллинойс
Джерролд Р. Тернер
2 Кафедра патологии, Чикагский университет, Чикаго, Иллинойс
Хаочу Хуанг
1 Кафедра медицины, Отдел ревматологии и Центр Кнаппа по волчанке и иммунологических исследованиях, Университет Чикаго, Чикаго, Иллинойс
1 Департамент медицины, Отделение ревматологии и Центр исследований волчанки и иммунологии Кнаппа, Чикагский университет, Чикаго, Иллинойс
2 Департамент патологии, Чикагский университет, Чикаго, Иллинойс
Адрес для корреспонденции и запросов на перепечатку: Haochu Huang, PhD, Департамент медицины, Отделение ревматологов. y, Центр исследований волчанки и иммунологии Кнаппа, Чикагский университет, Чикаго, штат Иллинойс 60637.ude.ogacihcu.dsb@gnauhh, тел .: 773-834-4482; Факс: 773-702-1576Джейсон Перера и Сяо Лю внесли равный вклад в эту работу.
Yuzhen Zhou, PhD (Текущий адрес: Медицинская школа Мичиганского университета, Анн-Арбор)
Окончательная отредактированная версия этой статьи издателя доступна бесплатно на сайте Arthritis Rheum См. Другие статьи в PMC, в которых цитируется опубликованная статья.Abstract
Цель
В модели ревматоидного артрита на мышах K / BxN Т-клетки, реактивные по отношению к аутоантигену глюкозо-6-фосфат-изомеразы (GPI), избегают отрицательного отбора, хотя экспрессия GPI является повсеместной.Мы стремились определить, может ли недостаточное представление GPI объяснить неудачу отрицательного отбора и развитие артрита.
Методы
Для увеличения антигенной презентации GPI мы создали трансгенных мышей, экспрессирующих мембраносвязанную форму GPI (mGPI), и скрестили их с мышами K / BxN. Моноклональные антитела, специфичные для альфа-цепи KRN TCR, были созданы, чтобы проследить судьбу GPI-специфических Т-клеток.
Результаты
трансгенных мышей mGPI продемонстрировали GPI более эффективно и показали резкое усиление отрицательного отбора и ингибирование артрита.Интересно, что отрицательный отбор тимуса у этих мышей оставался неполным, а ускользнувшие аутореактивные Т-клетки были анергическими на периферии, что позволяет предположить, что усиленная презентация антигена также вызывает периферическую толерантность. Несмотря на эту очевидную индукцию толерантности к GPI, у этих мышей действительно развилось хроническое истощение, характеризующееся воспалением толстой кишки с эпителиальной дисплазией, а также резким снижением количества Treg-клеток.
Заключение
Эти данные показывают, что недостаточная экспрессия или представление аутоантигена приводит к дефектам как центральной, так и периферической толерантности у мышей K / BxN.Это подтверждает идею о том, что недостаточный уровень аутоантигенов может лежать в основе развития аутоиммунитета.
ВВЕДЕНИЕ
Отрицательный отбор требует, чтобы аутоантигены были правильно доступны и эффективно представлены развивающимся тимоцитам (1, 2). Следовательно, уровни экспрессии и паттерны аутоантигенов могут влиять на эффективность клональной делеции (3, 4). Взаимосвязь между уровнями сывороточного белка и клональной делецией Т-клеток была исследована в нескольких экспериментальных системах.Концентрация лизоцима куриного яйца в сыворотке крови всего 0,1 нг / мл (10 −11 M) достаточна для удаления 3A9, но не 3L2, Т-лимфоцитов, специфичных к лизоциму куриного яйца (5). Напротив, делеция Т-клеток, специфичных для L-цепи Ig в качестве аутоантигена, требует концентрации в сыворотке более 100 мкг / мл (10 -6 M) (6). Таким образом, минимальный уровень экспрессии аутоантигена, необходимый для эффективного отрицательного отбора, сильно варьируется в зависимости от антигена и Т-клеточного рецептора (TCR), что, скорее всего, отражает присущие различия в способах получения этими аутоантигенами доступа к тимусу и процессинга. антиген-представляющими клетками тимуса (APC), а также сродством полученных пептидов к молекулам MHC и сродством этих комплексов пептид-MHC к их родственным TCR.
Хотя эти исследования предполагают связь между уровнями экспрессии и индукцией толерантности, не совсем понятно, способствует ли недостаточная экспрессия и представление аутоантигена дефектной толерантности Т-клеток и развитию аутоиммунитета. Более низкая восприимчивость к диабету 1 типа у людей связана с более высокими уровнями экспрессии инсулина в тимусе, предполагая, что более высокие уровни инсулина в тимусе могут способствовать отрицательному отбору инсулин-специфических Т-клеток (7).В соответствии с этой идеей, трансгенная сверхэкспрессия препроинсулина 2 существенно снижает возникновение и тяжесть диабета 1 типа у мышей с диабетом без ожирения (8).
Чтобы исследовать, как недостаточная презентация аутоантигена лежит в основе дефектной центральной толерантности и, в свою очередь, развития аутоиммунитета, мы использовали модель ревматоидного артрита на мышах K / BxN, вызванную дефектной толерантностью к самореактивному трансгенному TCR. Мыши K / BxN образуются, когда трансгенных мышей KRN TCR на фоне B6 (K / B) скрещивают со штаммом NOD (9).KRN TCR специфически распознает пептид глюкозо-6-фосфат изомеразы (GPI), представленный NOD-производной молекулой MHC II A g7 . Молодые животные K / BxN обнаруживают признаки клональной делеции в тимусе, однако значительное количество зрелых CD4 + Т-клеток наблюдается в возрасте 3-4 недель (9). Ускользнувшие Т-клетки KRN активируются и побуждают В-клетки производить высокие титры анти-GPI-антител, которые вызывают артрит в суставе, активируя каскад комплемента и клетки врожденной иммунной системы (10).
GPI — это повсеместный фермент, участвующий в гликолитическом пути. Важный вопрос заключается в том, как Т-клетки KRN, распознающие повсеместно распространенный белок, избегают ряда сложных механизмов, которые обычно обеспечивают толерантность к аутоантигенам. Пептиды, элюированные из IA g7 на В-клетках, включают пептиды из GPI (11, 12), однако специфический пептид GPI (282-294), который распознает KRN TCR, не входит в их число, что позволяет предположить, что GPI не эффективно процессируется и презентируется. к Т-клеткам KRN.В более раннем исследовании трансгенная экспрессия G7m, пептида, имитирующего GPI (282-294), показала массивную делецию в тимусе Т-клеток KRN и устранение Treg-клеток, но точную судьбу Т-клеток KRN невозможно было отследить из-за того, что отсутствие клонотипического антитела (13). Кроме того, мимотоп G7m стимулирует KRN T-клетки in vitro от 10 до 100 раз лучше, чем эндогенный пептид GPI (282-294). Хотя мимотоп, по-видимому, получает большую часть этого усиления от повышенного связывания с MHC, некоторые из контактных остатков TCR отличаются от нативного пептида GPI, что, возможно, может влиять на авидность TCR для измененного комплекса пептид-MHC.
В этом исследовании мы проверили гипотезу о том, что недостаточная обработка и представление GPI может объяснить отказ от негативного отбора и толерантности у мышей K / BxN за счет увеличения представления антигена эндогенного GPI. Мы создали трансгенную мышь, экспрессирующую мембраносвязанную форму GPI (называемую mGPI) и антитело, специфичное для альфа-цепи KRN TCR, для отслеживания трансгенных Т-клеток. Мы показали, что трансген mGPI приводит к более эффективной презентации пептида GPI, обширной делеции Т-клеток KRN в тимусе и ингибированию развития артрита.Несмотря на этот значительно усиленный отрицательный отбор, Т-клетки KRN все еще ускользают и накапливаются на периферии, но, в отличие от своих артритогенных аналогов, эти ускользнувшие Т-клетки поддерживаются в состоянии невосприимчивости к GPI. Эта невосприимчивость, по-видимому, не опосредована Treg-клетками, поскольку у трансгенных мышей mGPI вырабатывается гораздо меньше Treg-клеток тимуса и селезенки по сравнению с их нетрансгенными однопометниками. Кроме того, это снижение Treg-клеток коррелирует с развитием болезни истощения, характеризующейся воспалением толстой кишки и эпителиальной дисплазией высокой степени.Все вместе наши данные показывают, что недостаточная экспрессия и представление аутоантигенов может влиять как на центральную, так и на периферическую толерантность и может лежать в основе развития аутоиммунитета.
МАТЕРИАЛЫ И МЕТОДЫ
Получение трансгенных мышей mGPI
Лидерную последовательность из гена H-2K b амплифицировали с помощью ПЦР из плазмиды pODpCAGGS (14) и лигировали с полноразмерной кДНК GPI. Объединенный фрагмент клонировали в плазмиду pODpCAGGS с использованием Xba I для слияния с трансмембранной областью h3-D b .Фрагмент без векторных последовательностей использовали для создания трансгенных мышей B6 в центре трансгенных ядер Чикагского университета. Основатели были идентифицированы с помощью ПЦР хвостовой ДНК с использованием специфических праймеров. Все эксперименты были одобрены IACUC Чикагского университета.
Вестерн-блоттинг
Органы гомогенизировали в измельчителе тканей со стеклянными пестиками (Kontes) в 50 мМ Трис-HCl, pH 7,4, 150 мМ NaCl, 1% NP-40 с коктейлем ингибиторов протеазы (Roche). Лизаты центрифугировали при 12000 × g.Супернатант использовали для цитоплазматической фракции. Осадок (включая ядра) растворяли в 2% SDS, обрабатывали ультразвуком и использовали для мембранной фракции. GPI определяли с использованием сыворотки мышей K / BxN. Блоты удаляли и повторно зондировали антителом против тяжелой цепи клатрина (BD Transduction Laboratory, клон 23, № по каталогу 610500) и антителом против актина (Chemicon, № по каталогу MAB1501R) на мембранные и цитоплазматические фракции соответственно.
qRT-PCR
Органы гомогенизировали в Trizol с использованием гомогенизатора Даунса.После очистки РНК 1 мкг общей РНК использовали для синтеза кДНК с помощью обратной транскриптазы SmartScribe (Clontech).
Прямой (CCACTAACGGACTGATCAGCTTCATC) и обратный (AAGAGTCAGTGGACGGAGGA) праймеры были разработаны для специфической амплификации трансгена mGPI, а количественная ОТ-ПЦР была проведена с использованием SybrGreen PCR Mastermix (Applied Biosystems). Значения cT были нормализованы к стандартной кривой кДНК из образца толстой кишки, положительного по трансгену mGPI.
ELISA
Сывороточные титры IgG против GPI определяли с помощью ELISA.Планшеты покрывали 5 мкг / мл рекомбинантного GPI мыши. Серийные разведения образцов сыворотки определяли с помощью биотинилированного козьего антимышиного IgG (подклассы 1 + 2a + 2b + 3) Fcγ-специфических антител (Jackson ImmunoResearch) с последующим введением стрептавидина, конъюгированного с щелочной фосфатазой. Данные были аппроксимированы 4-параметрической кривой с использованием Prism (GraphPad). Титр определяется как разведение сыворотки, при котором ОП составляет 50% (точка перегиба) кривой.
Т-клеточная гибридома и анализ презентации антигена
Наивные KRN-трансгенные Т-клетки на фоне B6 инъецировали мышам TCRα — / — / B6xNOD F1 для их активации.Через одну неделю после инъекции Т-клетки CD4 + были очищены из селезенки хозяина и были непосредственно слиты с партнером слияния BWZ.36 (15). Гибриды Т-клеток субклонировали и проверяли на активность lacZ после культивирования с GPI-специфическими В-клетками в качестве APC. Клон G2 был выбран из-за его высокой активности lacZ и низкого фона.
Для анализов презентации антигена 1 × 10 5 клеток гибридомы KRN.G2 инкубировали со спленоцитами указанных мышей в течение 24 часов в 96-луночных планшетах. Клетки лизировали и измеряли общую активность lacZ с использованием хромогенного субстрата CPRG.
Первичный анализ пролиферации Т-клеток
Спленоциты метили CFSE и обогащали клетками CD4 + путем положительной селекции на магнитных колонках. 2,5 × 10 4 меченых спленоцитов CD4 + были смешаны с 2,5 × 10 5 спленоцитов стимулятора от мыши B6xNOD F1, которые были истощены по клеткам CD4 + и CD8 + путем отрицательной селекции на магнитных колонках. . Клетки культивировали в полной среде в течение 4,5 дней с градуированными концентрациями пептида GPI (282-294) и с 25 ед. / Мл человеческого IL-2 или без него (PeproTech).Клетки окрашивали анти-CD4, анти-KRNα и йодидом пропидия и анализировали на анализаторе FACSCanto (BD Bioscience).
Abs и проточный цитометрический анализ
Антитело против KRNα (клон 3-4G-B7) было получено путем иммунизации мышей B6 Т-клетками KRN, которые также экспрессируют связанный с мембраной овальбумин в качестве носителя для помощи Т-лимфоцитам (14) ( подробности будут описаны в другой рукописи). Коммерчески полученные mAb, использованные в этих исследованиях, включали: анти-CD4, анти-CD8α, анти-TCR Vβ6, анти-TCRβ BD Pharmingen).Обнаружение Treg осуществляли с помощью набора для окрашивания Foxp3 (eBiosciences), вкратце, клетки сначала окрашивали анти-CD4, анти-CD8 и анти-CD25 антителами, а затем внутриклеточно окрашивали анти-Foxp3.
Получение химерных мышей костного мозга
1–5 × 10 6 Истощенные Т-лимфоцитами клетки костного мозга вводили смертельно облученным мышам-хозяевам (1050 рад) с помощью внутривенного введения. инъекция через хвостовую вену. Мышей Chimera анализировали через два-три месяца после восстановления BM.
РЕЗУЛЬТАТЫ
Генерация трансгенных мышей mGPI
Мембранные белки процессируются и эффективно презентируются молекулами MHC класса II, а экспрессия пептида в мембраносвязанной форме усиливает его представление (16) (17). Поэтому мы адаптировали подход Ehst B.D. и другие. (14) для экспрессии мембраносвязанной формы GPI (mGPI) путем слияния сигнального пептида с N-концом GPI и мембранного домена с его C-концом. Слитый белок управляется повсеместно распространенным промотором куриного β-актина и энхансером CMV-IE ().Поскольку GPI является ферментом, участвующим в метаболизме глюкозы, и его сверхэкспрессия может иметь побочные эффекты, мы мутировали гистидин 388, который имеет решающее значение для ферментативной активности GPI (18), в глицин. Эта мутация не изменяет пептид GPI 282-294, распознаваемый Т-клетками KRN. Конструкцию сначала протестировали на культивируемых клетках 293 на экспрессию. Как показано на, экспрессия поверхностного GPI была легко обнаружена с помощью проточной цитометрии. Затем конструкцию использовали для создания трансгенных мышей на фоне C57BL6, и с помощью ПЦР идентифицировали одиннадцать основателей.Неожиданно нам не удалось обнаружить поверхностную экспрессию GPI ни у одного из основателей. Отсутствие поверхностной экспрессии GPI может быть связано с неправильной укладкой, нестабильной конфигурацией на поверхности клетки или ее быстрой интернализацией. Однако поверхностная экспрессия не требуется для эффективного процессинга и презентации антигена (16), а экспрессия mGPI действительно нацелена на GPI в мембранную фракцию клеток, не влияя на эндогенные цитоплазматические уровни GPI, как показано с помощью вестерн-блоттинга ().Одна линия трансгенных мышей (линия 5, обозначенная как mGPI) была использована в последующих исследованиях. Используя количественную ОТ-ПЦР, мы подтвердили, что экспрессия трансгена в этой линии обнаруживалась во всех исследованных органах ().
Экспрессия связанного с мембраной GPI увеличивает презентацию пептида GPI.A , Карта конструкции mGPI, используемой для создания трансгенных мышей. B , Экспрессия mGPI на клетках 293, временно трансфицированных конструкцией mGPI (темная линия) путем окрашивания mAb против GPI, по сравнению с контрольными клетками с ложной трансфекцией (пунктирная линия). C , Субклеточное распределение mGPI. Цитоплазматические и мембранные фракции селезенки и тимуса мышей mGPI + / g7 и mGPI — / g7 анализировали с помощью вестерн-блоттинга. Антитела против тяжелой цепи клатрина и антитела против бета-актина используются в качестве контроля нагрузки для мембранных и цитоплазматических фракций, соответственно. Расчетные молекулярные массы эндогенных GPI и mGPI составляют 62,8 кДа и 67,6 кДа соответственно. D , Способность спленоцитов мышей mGPI + / g7 представлять пептид GPI.1 × 10 5 клеток гибридомы KRN.G2 инкубировали со спленоцитами из mGPI + / A g7 (квадрат), mGPI — / A g7 (кружок) и анти-GPI BCR knock-in мышки (треугольник). Общую активность lacZ измеряли с использованием хромогенного субстрата CPRG. Три независимых эксперимента были обобщены после нормализации активности lacZ для внутреннего контроля. E, Обнаружение транскрипта mGPI в различных тканях. ПЦР-анализ в реальном времени выполняли с использованием праймеров, специфичных для трансгена, и значения cT нормализовали по стандартной кривой кДНК mGPI + / g7 толстой кишки.
Повышенная презентация пептида GPI у трансгенных мышей mGPI
Чтобы подтвердить, что сверхэкспрессия mGPI увеличивает презентацию пептида GPI, мы создали Т-клеточную гибридому (обозначенную как KRN.G2) с использованием трансгенных Т-клеток KRN и партнера слияния BWZ.36 несущий конструкцию NFAT-lacZ (15). При вовлечении TCR родственным MHC-пептидом продуцируется lacZ, обеспечивая удобное и чувствительное считывание по сравнению с измерениями продукции IL-2. Мышей mGPI сначала скрестили с B6.H-2 g7 конгенных мышей для введения молекулы I-A g7 (обозначаемой мышами mGPI + / g7), поскольку Т-клетки KRN специфичны для пептида GPI, представленного только I-A g7 . Клетки гибридомы KRN.G2 культивировали со спленоцитами мышей mGPI + / g7, трансген-отрицательных однопометников (mGPI — / g7) или в качестве положительного контроля спленоцитами мышей с нокаутом анти-GPI Ig на Rag — / — фон, поскольку они имеют гомогенную популяцию GPI-специфических B-клеток (19).Было обнаружено, что минимальное количество спленоцитов mGPI + / g7, необходимое для индукции lacZ, в 10-50 раз меньше, чем спленоцитов mGPI — / g7, что подтверждает усиленную презентацию пептида GPI у трансгенных мышей mGPI + / g7. ().
mGPI ингибирует развитие артрита
Затем мы скрестили мышей mGPI + / g7 с трансгенными мышами KRN TCR, чтобы проверить, как усиленное представление GPI влияет на развитие артрита. Полученных мышей mGPI + / K / g7 и контрольных однопометников mGPI — / K / g7 отслеживали на предмет возникновения и тяжести артрита после отлучения от груди.Как описано ранее (9), у мышей mGPI — / K / g7 артрит развился в возрасте 4–5 недель с резким началом отека суставов. Максимальный отек голеностопного сустава был достигнут в возрасте 6–8 недель с последующей деформацией голеностопного сустава. Напротив, 85% мышей mGPI + / K / g7 не имели признаков артрита. У оставшихся 15% мышей mGPI + / K / g7 развился мягкий артрит с отсроченным началом ().
Влияние трансгена mGPI на развитие артритаA , Толщину голеностопного сустава мышей mGPI + / K / g7 и контрольных однопометников mGPI — / K / g7 отслеживали, начиная примерно с 3-недельного возраста.Было показано среднее значение и стандартное отклонение в каждый момент времени (n = 35 для mGPI + / K / g7; n = 37 для mGPI — / K / g7). B , Сывороточные титры анти-GPI IgG мышей mGPI — / K / g7 и mGPI + / K / g7 (возраст 8–12 недель). Каждая точка представляет собой отдельную мышь. p <0,001, рассчитанное с помощью непарного t -теста.
Поскольку артрит опосредуется патогенными антителами против GPI, титры сывороточных антител против GPI IgG определяли в обеих группах мышей.Как показано на фигуре, у мышей mGPI + / K / g7 уровни антител IgG к GPI были примерно в 1000 раз ниже, чем у мышей mGPI — / K / g7. Таким образом, блокировка развития артрита у мышей mGPI + / K / g7 происходит до продукции анти-GPI-антител.
Отрицательный отбор Т-клеток KRN у трансгенных мышей mGPI
Профилактика продукции аутоантител и артрита у мышей mGPI + / K / g7 показала, что толерантность к GPI была восстановлена. Мы проанализировали Т-клеточные компартменты этих мышей с помощью проточной цитометрии.Как описано ранее, отрицательная селекция Т-клеток KRN у мышей K / BxN была неэффективной, и значительные популяции зрелых одноположительных клеток присутствуют в тимусе и периферических лимфоидных органах (9). Напротив, делеция тимоцитов у мышей mGPI + / K / g7 была намного более эффективной с 6-кратным снижением общей клеточности (). Анализ компартментов CD4 / CD8 показал почти полную делецию дважды положительных (DP) тимоцитов с относительно небольшим изменением компартмента двойных отрицательных (DN), что привело к резкому снижению отношения DP / DN ().Общее количество CD4 + одноположительных (SP) тимоцитов уменьшилось в 2 раза, хотя его процент увеличился. Несмотря на эффективную делецию тимоцитов, не было резкой разницы в общем количестве спленоцитов (1,48 × 10 8 ± 0,5 и 1,75 × 10 8 ± 0,7 для mGPI — / K / g7 и mGPI + / K / g7 соответственно) и процент CD4 + Т-клеток (). Анализ маркеров активации на Т-клетках CD4 + от обеих мышей показал повышенную долю клеток CD44 hi CD62 lo в соответствии с предыдущим столкновением с антигеном (данные не показаны).Повышенный фенотип отрицательного отбора не зависит от возраста животных, поскольку анализируемые мыши в возрасте двух недель показали очень похожие результаты ().
Выбор KRN T-клеток и использование цепей TCR, кодируемых трансгенамиA , Клетки, полученные из тимуса или селезенки трансгенных мышей mGPI + / K / g7 и mGPI — / K / g7 из однопометного потомства контрольную группу в возрасте от 8 до 12 недель окрашивали антителами к CD4 и CD8. B , соотношение дважды положительных и дважды отрицательных клеток в тимусе (верхний, n = 5) и общее количество тимоцитов (нижнее, n = 8) для мышей в возрасте от 8 до 12 недель. C, соотношение дважды положительных и дважды отрицательных клеток и общее количество тимоцитов для мышей в возрасте 2 недель (n = 6 для каждой группы) D , Клетки, полученные из тимуса и селезенки B6xNOD F1 (BxN), mGPI — / K / g7 контроль и mGPI + / K / g7 трансгенных мышей в возрасте 8–12 недель окрашивали антителом, специфичным к KRN TCRα (3-4G-B7), и анализировали с помощью проточной цитометрии. Показанные клетки сначала были привязаны к одиночным положительным клеткам CD4 + . D , общее количество CD4 + SP KRN T-клеток в тимусе (верхний, n = 5) и селезенке (нижний, n = 5), соответственно.Для всех графиков показаны среднее значение и стандартное отклонение. Весь статистический анализ выполнен с помощью непарного критерия Стьюдента t . * p <0,05, ** p <0,01, *** p <0,001.
Поскольку аллельное исключение для цепи TCRα не очень эффективно в Т-клетках, было неясно, действительно ли ускользающие CD4 + Т-клетки все еще экспрессируют рецептор, кодируемый трансгеном, или вместо этого экспрессируют рецепторы, продуцируемые эндогенными перестройками. Чтобы различить эти возможности, мы создали антитело (клон 3-4G-B7), специфичное для альфа-цепи KRN TCR (рукопись находится в стадии подготовки), и использовали его для отслеживания судьбы KRN-экспрессирующих Т-клеток, поскольку все клетки которые экспрессируют альфа-цепь KRN, также экспрессируют бета-цепь KRN.Как показано на фиг. 3-4G-B7, специфичен для Т-клеток KRN и не проявляет окрашивания Т-клеток дикого типа. Процент Т-клеток KRN в компартменте CD4SP в тимусе был немного снижен, но количество Т-клеток KRN уменьшилось в 3 раза, подтверждая усиление делеции Т-клеток KRN (). Аналогичные изменения наблюдались на периферии, где процентное содержание KRN T-клеток в CD4 + T-клетках уменьшилось на 40%, а количество KRN T-клеток уменьшилось в 3 раза (). Эти результаты показывают, что, несмотря на сокращение числа, все еще существует значительная популяция Т-клеток KRN на периферии, однако прекращение продукции антител и артрит указывают на то, что эти Т-клетки KRN функционально подавлены.
Мы подтвердили это путем измерения ответа Т-клеток на GPI (282-294) пептид in vitro . CD4 + Т-клетки были обогащены селезенками mGPI + / K / g7 и mGPI — / K / g7 и помечены CFSE. Эти меченые спленоциты смешивали с клетками-стимуляторами, несущими A g7 , в присутствии определенных количеств пептида GPI (282-294). KRN + CD4 + Т-клетки из mGPI — / K / g7 активно пролиферировали в ответ на пептид, в то время как Т-клетки из mGPI + / K / g7 — нет ().В некоторых моделях анергии Т-клеток добавление экзогенного ИЛ-2 может восстановить пролиферацию (20), однако мы не наблюдали какого-либо влияния на Т-клетки KRN + в культурах mGPI + / K / g7 с добавлением ИЛ. -2 (). Эти данные подтверждают идею о том, что ускользнувшие Т-клетки KRN поддерживаются в состоянии толерантности к GPI на периферии.
Пролиферация Т-клеток в ответ на GPI (282-294)2,5 × 10 4 CD4 + Т-клетки из mGPI — / K / A g7 и mGPI + / K / A Мышей g7 (возраст 2–5 месяцев) метили CFSE и совместно культивировали с 2.5 × 10 5 A g7 , несущие клетки-стимулятор в присутствии градуированного количества пептида GPI (282-294) (мкМ) и 25 Ед / мл IL-2 в течение 4,5 дней в 96-луночном планшете. Индивидуальные культуры анализировали с помощью проточной цитометрии после окрашивания антителом против CD4 и против KRN TCRα. A , Типичные графики FACS пролиферации в ответ на пептид. B , Количественная оценка распространения относительно нестимулированного контроля. Кратное изменение общего количества Т-клеток KRN в культуре в конце 4.Показано 5 дней относительно нестимулированного контроля. Все точки отражают среднее значение трех лунок со стандартным отклонением. Представитель трех независимых экспериментов.
Вклад APC, полученных из костного мозга, по сравнению с APC, полученными не из костного мозга эпителиальные клетки. Оба типа клеток могут эффективно опосредовать отрицательный отбор, если у них есть доступ к антигенам (21).Поскольку трансген mGPI управляется повсеместно распространенным промотором, мы сравнили относительный вклад APC, происходящих из BM, по сравнению с эпителиальными клетками тимуса для эффективной клональной делеции Т-клеток KRN, наблюдаемой у трансгенных мышей mGPI
+ / K / g7. Четыре типа мышей-химер BM были созданы с использованием доноров BM или хозяев, которые экспрессируют или не экспрессируют трансген mGPI (). Химера 4 служила отрицательным контролем, поскольку ни BM, ни хозяин не несли трансген mGPI. Химера 1 служила положительным контролем, поскольку и BM, и хозяева несли трансген mGPI.В химере 2 только BM несет трансген mGPI, а в химере 3 только хозяин несет трансген mGPI. Анализ количества общих тимоцитов и соотношения DP / DN показал, что либо APC, происходящие из BM, либо эпителиальные клетки тимуса могут опосредовать эффективную делецию KRN T-клеток, однако эпителиальные клетки тимуса были немного более эффективны, чем APC, полученные из BM (при сравнении химер 2 и 3). Эпителиальные клетки тимуса также индуцировали более резкое снижение соотношения DP / DN, предполагая, что делеция имеет место на более ранней стадии развития тимоцитов.Это согласуется с разной анатомической локализацией развивающихся тимоцитов. Клетки DP в основном локализованы в коре головного мозга, и они будут удалены кортикальными эпителиальными клетками тимуса, которые имеют достаточное количество комплекса пептид-MHC. Удаление APC, происходящих из BM, которые в основном находятся в мозговом веществе тимуса, будет происходить на более поздней стадии развития. Вклад BM по сравнению с APC, полученными не из BM, в делецию Т-клеток KRN.Мышей-химеры Crisscross BM получали с использованием доноров BM и хозяев, которые (+) или не (-) несут трансген mGPI.Хозяев смертельно облучали (1050 рад) перед восстановлением BM, и мышей анализировали через 3 месяца. Количество общих тимоцитов (верхний график) и соотношение DP / DN в тимусе (нижний график) были показаны для каждой комбинации BM и хозяина (n = 6–12 мышей на группу, * p <0,004, ** p < 0,0001 по непарному тесту t ).
Мыши mGPI
+ / K / g7 имеют дефектную продукцию Treg и развивают болезнь истощенияХотя у мышей mGPI + / K / g7 артрит не развился, измерение их массы тела с течением времени показало, что их рост был приостановлен рано ().К 2–3 месяцам они начали проявлять сгорбленную осанку, взъерошенный мех, были худыми и вялыми. Мышей, у которых развились самые тяжелые фенотипы, забивали в соответствии с рекомендациями Институционального комитета по уходу за животными и их использованию. Выживаемость мышей снижалась с возрастом, и только 35% мышей дожили до 5-месячного возраста (). Исследование нескольких органов у этих мышей не показало явного воспаления в печени, почках, легких, сердце и поджелудочной железе. Когда мы проанализировали желудочно-кишечный тракт, тонкий кишечник был гистологически нормальным, но толстый кишечник (слепая и толстая кишка) этих животных продемонстрировал активное хроническое воспаление от слабого до тяжелого у более чем 90% мышей в возрасте 2–5 месяцев ().У 5 из 11 мышей мы наблюдали дисплазию эпителия низкой степени и еще один случай дисплазии высокой степени с инвазией в подслизистую выстилку (данные не показаны). Напротив, мы не наблюдали воспаления толстой кишки или дисплазии у мышей, отрицательных по трансгену mGPI ().
Фенотип истощения и потеря Treg-клеток у трансгенных мышей mGPI + / K / g7A , Кривая выживаемости мышей mGPI + / K / g7 (n = 31) по сравнению с mGPI — / K / g7 (n = 15).Метки на кривой указывают, когда отдельные мыши были удалены и не завершили мониторинг. B , Вес тела мышей mGPI + / K / g7 (кружок), mGPI — / K / g7 (квадрат) и mGPI + / K / b (треугольник) с течением времени. C , толстая кишка мышей mGPI — / K / g7 с нормальной гистологией. D , толстая кишка мышей mGPI + / K / g7, показывающая активное и хроническое воспаление с архитектурным искажением крипт. Масштабная линейка равна 0,1 мм. E , Обнаружение Treg-клеток с помощью проточной цитометрии.Клетки получали из тимуса или селезенки мышей в возрасте 8–12 недель. Показаны среднее и стандартное отклонение процентного содержания Treg (n = 15). F , Общее количество клеток Foxp3 + CD25 + в тимусе (n = 15, p <0,001) и селезенке (n = 15, p <0,001). Показаны среднее и стандартное отклонение. Проведен непарный т -тест. G , Процент трансгенных Т-клеток KRN в компартменте Foxp3 + Treg, как определено с помощью антитела против цепи KRNα (n = 3). H, Пролиферация Т-клеток KRN из брыжеечных лимфатических узлов (MLN) в ответ на пептид GPI (282-294) в возрасте 6 месяцев. Выполняется, как описано в. Представитель трех независимых экспериментов.
Затем мы исследовали, были ли затронуты Treg-клетки у мышей mGPI + / K / g7. Как показано на фиг.4, процентное содержание Treg-клеток Foxp3 + CD25 + было значительно снижено в тимусе и селезенке мышей mGPI + / K / g7. Абсолютное количество Tregs уменьшилось 13.В 8 раз больше в тимусе и в 3,8 раза в селезенке (). Это снижение не было связано с неспособностью Т-клеток KRN + рекрутироваться в компартмент Treg, поскольку Treg-клетки KRN + наблюдаются с одинаковой частотой в обоих генотипах (). В соответствии с ролью Treg-клеток в подавлении аутоиммунитета кишечника во многих экспериментальных моделях (22) эти данные предполагают, что потеря Treg-клеток может быть причиной развития колита и истощения у мышей mGPI + / K / g7.
Учитывая неотзывчивую природу Т-клеток KRN в селезенке мышей mGPI + / K / g7, мы спросили, являются ли Т-клетки KRN анергическими в кишечнике. Мы измерили способность mGPI + / K / g7 Т-клеток из брыжеечного лимфатического узла (MLN) мышей в возрасте 6 месяцев реагировать на стимуляцию GPI (282-294) пептидом in vitro . В то время как KRN T-клетки из MLN mGPI — / K / g7 резко увеличивались по сравнению с их селезеночными аналогами, KRN T-клетки из MLN mGPI + / K / g7 оставались нечувствительными к пептиду ().
ОБСУЖДЕНИЕ
В этом исследовании мы исследовали, может ли недостаточная презентация аутоантигенов объяснить ускользание аутореактивных клеток и нарушение толерантности в случае повсеместного аутоантигена. Здесь мы показали, что усиленная презентация GPI действительно резко увеличивала делецию Т-клеток KRN на двойной положительной стадии в тимусе и предотвращала развитие артрита. Массивная делеция в тимусе также сопровождалась значительной потерей регуляторных Т-клеток и тяжелым хроническим воспалением в толстой кишке этих трансгенных животных.
Биохимические, структурные и функциональные анализы MHC II I-A g7 показывают, что его пептид-связывающие свойства играют важную роль в аутоиммунности (23). В частности, I-A g7 связывается со многими из своих лигандов в низком микромолярном диапазоне, который является слабым по сравнению с другими мышиными аллелями, такими как I-A k , которые связываются со своими пептидами в наномолярном диапазоне. Ускользание аутореактивных Т-клеток, специфичных для пептида β-цепи инсулина (9–23), объясняется слабым связыванием пептида с молекулой I-A g7 (обзор см. В (23)).Сравнение пептида GPI (SIALHVGFD) с пептидным связывающим мотивом I-A g7 (24) показывает присутствие предпочтительных аминокислот во многих положениях. Однако неспособность обнаружить этот пептид среди пептидов, элюированных из IA g7 на В-клетках, которые включают другие пептиды GPI, поддерживает модель, в которой слабое связывание GPI (282-294) с IA g7 позволяет избежать KRN T ячеек (11, 12). Наши результаты показывают, что более высокие уровни экспрессии могут компенсировать более низкое сродство пептида к MHC.Следовательно, эффективность индукции толерантности конкретной специфичности Т-клеток может зависеть от множества факторов, включая уровень экспрессии собственного белка, эффективность процессинга и презентации антигена, а также сродство пептида к MHC.
Уровни экспрессии аутоантигена могут влиять на дифференцировку Treg, хотя механизм не совсем понятен (обзор в (25, 26)). У мышей mGPI + / K / g7 повышенная презентация антигена приводит к уменьшению количества Treg-клеток.Наш результат согласуется с предыдущими исследованиями, в которых трансгенных мышей TCR, специфичных для HA, скрещивали с несколькими линиями мышей, экспрессирующих разные уровни HA (25). Было показано, что дифференцировка Treg зависит от узнавания собственного пептида (пептида HA), поскольку у мышей, экспрессирующих только трансген TCR, развивается небольшое количество Treg-клеток. Более высокие уровни экспрессии HA приводят к более эффективному удалению родственных Т-клеток, а среди клеток, которые избегают делеции, аналогичные пропорции приобретают экспрессию Foxp3.В конечном итоге более высокие уровни экспрессии HA приводят к снижению общего количества Treg-клеток.
Хотя механизм развития колита у мышей mGPI + / K / g7 здесь не рассматривался, наши результаты согласуются с хорошо установленной ролью регуляторных Т-клеток в поддержании гомеостаза кишечника (см. 22)). В модели колита с переносом Т-клеток адоптивно перенесенные наивные CD4 + Т-клетки вызывают воспаление кишечника у реципиентов SCID, и колит можно предотвратить с помощью ко-переносимых Treg-клеток.Колит у мышей mGPI + / K / g7 можно объяснить снижением продукции Treg наряду с потенциальным сужением специфичности Treg из-за экспрессии трансгена TCR. Также возможно, что Т-клетки KRN и экспрессия mGPI в толстой кишке играют роль в патогенезе колита. Было продемонстрировано, что эпителиальные клетки толстой кишки могут аберрантно экспрессировать молекулы MHC класса II в ответ на воспаление (27). Высокая экспрессия трансгена mGPI в толстой кишке может способствовать представлению GPI Т-клеткам KRN эпителиальными клетками толстой кишки или другими APC, тем самым обостряя заболевание.
Наши данные подтверждают, что центральные и периферические механизмы толерантности важны для предотвращения аутоиммунитета. Было подсчитано, что от половины до двух третей тимоцитов, отобранных положительно, впоследствии подвергаются отрицательному отбору (см. Обзор (28)). Однако центральный допуск не идеален или надежен. В нашей модели имеется множество Т-клеток KRN на периферии мышей mGPI + / K / g7, но они не вызывают заболевания. Как они отключены функционально? Поскольку количество Treg-клеток значительно снижено у мышей mGPI + / K / g7, Treg-клетки, по-видимому, не отвечают за восстановление толерантности.Анергия является одним из основных механизмов периферической толерантности, и in vivo анергия была описана для нескольких трансгенных TCR (обзор в (29)). Наша система напоминает эти модели с точки зрения невосприимчивости Т-клеток и неспособности экзогенного ИЛ-2 спасти пролиферацию. Исследования, посвященные влиянию уровня экспрессии антигена на индукцию анергии in vivo , показали, что трансген цитохрома c голубя способен одинаково хорошо толеризовать трансгенные Т-клетки как при высоком, так и при низком уровне экспрессии (30).Наша модель уникальна в двух аспектах. Во-первых, в нашей модели естественный уровень презентации приводит к активации, и только высокая презентация антигена приводит к толерантности. Во-вторых, развитие артрита обеспечивает функциональное считывание in vivo для решения судьбы Т-клеток. Возможно, что в нашей системе экспрессия mGPI влияет не только на силу антигенной стимуляции, но и на основной тип задействованных APC. У мышей mGPI + / K / g7 способность всех В-клеток представлять GPI повышена.Было показано, что нацеливание антигенов на В-клетки может индуцировать толерантность к Т-клеткам (31–33). Необходимы дальнейшие механистические исследования, и результаты могут пролить свет на то, как аутореактивные Т-клетки избегают периферических механизмов толерантности, чтобы управлять аутоиммунными ответами.
Благодарности
Мы благодарим доктора Марка Дженкинса за предоставление плазмиды pODpCAGGS; Доктора Дайан Матис и Кристоф Бенуа для трансгенных мышей KRN и конгенных мышей B6.h3 g7 ; Доктору Нилабху Шастри за предоставление BWZ.36 ячеек; Доктору Мартину Вейгерту за критическое прочтение рукописи; и основное предприятие по производству трансгенных мышей Чикагского университета для создания трансгенных мышей mGPI.
При поддержке NIH (грант NIAID R01-AI087645 доктору Хуангу). Джейсон Перера был частично поддержан грантом NIH / NIAID 2T32AI007090.
Список литературы
1. Goodnow CC. Многоэтапный патогенез аутоиммунного заболевания. Клетка. 2007. 130 (1): 25–35. [PubMed] [Google Scholar] 2. Хогквист К.А., Болдуин Т.А., Джеймсон СК. Центральная толерантность: обучение самоконтролю в тимусе.Nat Rev Immunol. 2005. 5 (10): 772–82. [PubMed] [Google Scholar] 3. Oehen SU, Ohashi PS, Bürki K, Hengartner H, Zinkernagel RM, Aichele P. Спасение тимоцитов и зрелых Т-клеток от клональной делеции из-за ограничения уровней экспрессии толерогенов. Клеточная иммунология. 1994. 158 (2): 342–52. [PubMed] [Google Scholar] 4. Аккараджу С., Хо В.Й., Леонг Д., Ханаан К., Дэвис М.М., Гуднау С.К. Диапазон толерантности Т-лимфоцитов CD4: частичная инактивация органоспецифического антигена позволяет проводить неразрушающий тиреоидит или инсулит. Иммунитет.1997. 7 (2): 255–71. [PubMed] [Google Scholar] 5. Харибхай Д., Энгл Д., Мейер М., Донермейер Д., Уайт Дж. М., Уильямс CB. Порог толерантности центральных Т-клеток к индуцибельному белку сыворотки. J Immunol. 2003. 170 (6): 3007–14. [PubMed] [Google Scholar] 7. Пульезе А., Зеллер М., Фернандес А., Зальцберг Л.Дж., Бартлетт Р.Дж., Рикорди С. и др. Ген инсулина транскрибируется в тимусе человека, и уровни транскрипции коррелируют с аллельными вариациями в локусе чувствительности INS VNTR-IDDM2 для диабета 1 типа. Нат Жене.1997. 15 (3): 293–7. [PubMed] [Google Scholar] 8. Jaeckel E, Lipes MA, von Boehmer H. Рецессивная толерантность к препроинсулину 2 снижает, но не отменяет диабет 1 типа. Nat Immunol. 2004. 5 (10): 1028–35. [PubMed] [Google Scholar] 9. Кускофф В., Корганов А.С., Дюшатель В., Деготт С., Бенуа С., Матис Д. Органоспецифическое заболевание, спровоцированное системным аутоиммунитетом. Клетка. 1996. 87 (5): 811–22. [PubMed] [Google Scholar] 10. Джи Х, Омура К., Махмуд У, Ли Д.М., Хофхус ФМА, Бокл С.А. и др. Артрит критически зависит от врожденной иммунной системы игроков.Иммунитет. 2002. 16 (2): 157–68. [PubMed] [Google Scholar] 11. Сури А., Видавский И., ван дер Дрифт К., Канагава О., Гросс М.Л., Унануэ Э.Р. В APC аутологичные пептиды, выбранные диабетогенной молекулой I-Ag7, уникальны и определяются аминокислотными изменениями в кармане P9. J Immunol. 2002. 168 (3): 1235–43. [PubMed] [Google Scholar] 12. Донгре А.Р., Коватс С., деРоос П., Маккормак А.Л., Накагава Т., Пахаркова-Вачкова В. и др. Презентация цитозольных белков in vivo MHC класса II, выявленная с помощью быстрой автоматизированной тандемной масс-спектрометрии и функционального анализа.Eur J Immunol. 2001. 31 (5): 1485–94. [PubMed] [Google Scholar] 13. Ши Ф.Ф., Мандик-Наяк Л., Випке Б.Т., Аллен П.М. Массивная делеция тимуса приводит к системному аутоиммунитету за счет устранения CD4 + CD25 + Т-регуляторных клеток. J Exp Med. 2004. 199 (3): 323–35. [Бесплатная статья PMC] [PubMed] [Google Scholar] 14. Ehst BD, Ingulli E, Jenkins MK. Разработка новой трансгенной мыши для изучения взаимодействий между CD4 и CD8 Т-клетками во время отторжения трансплантата. Am J Transplant. 2003. 3 (11): 1355–62. [PubMed] [Google Scholar] 15.Сандерсон С., Шастри Н. Индуцируемые LacZ, антиген / MHC-специфические Т-клеточные гибриды. Int Immunol. 1994. 6 (3): 369–76. [PubMed] [Google Scholar] 16. Брукс А., Хартли С., Кьер-Нильсен Л., Перера Дж., Гуднау С.С., Бастен А. и др. Ограниченная классом II презентация эндогенно производной иммунодоминантной Т-клеточной детерминанты лизоцима куриного яйца. Proc Natl Acad Sci U S. A. 1991; 88 (8): 3290–4. [Бесплатная статья PMC] [PubMed] [Google Scholar] 17. Уильямс С.Б., Видал К., Донермейер Д., Петерсон Д.А., Уайт Дж. М., Аллен П. М.. Экспрессия антагониста TCR in vivo: Т-клетки избегают центральной толерантности, но антагонизируют на периферии.J Immunol. 1998. 161 (1): 128–37. [PubMed] [Google Scholar] 18. Грахамсоломонс Дж., Циммерли Э., Бернс С., Кришнамурти Н., Свон М., Крингс С. и др. Кристаллическая структура мышиной фосфоглюкозоизомеразы при разрешении 1,6А и ее комплекса с глюкозо-6-фосфатом раскрывают каталитический механизм раскрытия сахарного кольца. Журнал молекулярной биологии. 2004. 342 (3): 847–60. [PubMed] [Google Scholar] 19. Хуанг Х., Кирни Дж. Ф., Грусби М. Дж., Бенойст С., Матис Д. Индукция толерантности в артритогенных В-клетках с рецепторами разного сродства к аутоантигену.Proc Natl Acad Sci USA. 2006. 103 (10): 3734–9. [Бесплатная статья PMC] [PubMed] [Google Scholar] 20. Беверли Б., Канг С.М., Ленардо М.Дж., Шварц Р.Х. Обращение клональной анергии Т-клеток in vitro путем стимуляции ИЛ-2. Int Immunol. 1992. 4 (6): 661–71. [PubMed] [Google Scholar] 21. Штокингер Б. Толерантность к Т-лимфоцитам: от делеции тимуса до механизмов периферического контроля. Adv Immunol. 1999; 71: 229–65. [PubMed] [Google Scholar] 22. Барнс М.Дж., Поури Ф. Регуляторные Т-клетки усиливают гомеостаз кишечника. Иммунитет. 2009. 31 (3): 401–11.[PubMed] [Google Scholar] 23. Сури А., Левисетти М.Г., Унануэ ER. Объясняют ли пептид-связывающие свойства молекул диабетогенного класса II аутореактивность? Curr Opin Immunol. 2008. 20 (1): 105–10. [Бесплатная статья PMC] [PubMed] [Google Scholar] 24. Чанг К.Ю., Сури А., Унануэ ER. Прогнозирование пептидов, связанных с молекулами гистосовместимости I-Ag7 класса II, с использованием нового алгоритма выравнивания с максимизацией ожидания. Протеомика. 2007. 7 (3): 367–77. [PubMed] [Google Scholar] 25. Simons DM, Picca CC, Oh S, Perng OA, Aitken M, Erikson J, et al.Как специфичность собственных пептидов влияет на развитие и функцию регуляторных Т-клеток. J Leukoc Biol. 2010. 88 (6): 1099–107. [Бесплатная статья PMC] [PubMed] [Google Scholar] 26. Wirnsberger G, Hinterberger M, Klein L. Регулирующая дифференцировка Т-клеток по сравнению с клональной делецией аутореактивных тимоцитов. Immunol Cell Biol. 2010. 89 (1): 45–53. [PubMed] [Google Scholar] 27. Kuhn R, Lohler J, Rennick D, Rajewsky K, Muller W. У мышей с дефицитом интерлейкина-10 развивается хронический энтероколит. Клетка. 1993. 75 (2): 263–74.[PubMed] [Google Scholar] 28. Матис Д., Бенуа С. Вернемся к центральной терпимости. Иммунитет. 2004. 20 (5): 509–16. [PubMed] [Google Scholar] 30. Сингх Нью-Джерси, Шварц Р. Сила стойкой антигенной стимуляции модулирует адаптивную толерантность периферических CD4 + Т-клеток. J Exp Med. 2003. 198 (7): 1107–17. [Бесплатная статья PMC] [PubMed] [Google Scholar] 31. Эйнон Э., Паркер, округ Колумбия. Малые В-клетки как антигенпрезентирующие клетки в индукции толерантности к растворимым белковым антигенам. J Exp Med. 1992. 175 (1): 131–8. [Бесплатная статья PMC] [PubMed] [Google Scholar] 32.Fuchs EJ, Matzinger P. B-клетки выключают первичные, но не Т-клетки памяти. Наука. 1992. 258 (5085): 1156–9. [PubMed] [Google Scholar] 33. Ян Дж., Мамула МДж. Толерантность к В- и Т-клеткам и аутоиммунитет у трансгенных мышей с аутоантителами. Int Immunol. 2002. 14 (8): 963–71. [PubMed] [Google Scholar] Диапазон допуска, формула фундаментального отклонения, формулы фундаментального отклонения, Iso 286, IT8 To IT16,
Формулы фундаментальных отклонений валов размером до 500-3150 мм
| ВАЛ | ОТВЕРСТИЯ | ФОРМУЛ ДЛЯ ОТКЛОНЕНИЯ В МИКРОН | ||||
| ОБОЗНАЧЕНИЕ ВАЛА | ОСНОВНОЕ ОТКЛОНЕНИЕ | ЗНАК | ОБОЗНАЧЕНИЕ ОТВЕРСТИЯ | ОСНОВНОЕ ОТКЛОНЕНИЕ | ЗНАК | Для «D» в мм |
| d | es | — | D | EI | + | 16D 0.44 |
| e | es | — | E | EI | + | 11D 0,41 |
| f | es | — | F | EI | + | 5.5D 0.41 |
| г | es | — | G | EI | + | 2.5D 0,34 |
| h | es | Без подписи | H | EI | Без подписи | 0 |
| js | ei | — | JS | ES | + | 0.5ITn |
| k | ei | + | K | ES | — | 0 |
| м | ei | + | M | ES | — | 0,024D + 12,6 |
| n | ei | + | N | ES | — | 0,04 + 21 |
| p | ei | + | P | ES | — | 0.072D + 37,8 |
| r | ei | + | R | ES | — | Среднее геометрическое значений «p» и «s» или «P» и «S» |
| s | ei | + | S | ES | — | IT7 + 0.4D |
| t | ei | + | T | ES | — | IT7 + 0.63D |
| u | ei | + | U | ES | — | IT7 + D |
Диапазон допуска ISO 286 от «IT8» до «IT16»
Указанный ниже диапазон допусков (от IT8 до IT16) в микрометрах = (10 -3) мм
| НОМИНАЛЬНЫЕ (ОСНОВНЫЕ) РАЗМЕРЫ (мм) | МЕЖДУНАРОДНЫЕ КЛАССЫ ДОПУСКА | ||||||||||||||||||
| БОЛЕЕ | ДО ВКЛ. | IT8 | IT9 | IT10 | IT11 | IT12 | IT13 | IT14 | IT15 | IT16 | |||||||||
| 0 | 3 | 14 | 25 | 40 | 60 | 100 | 140 | 250 | 400 | 600 | |||||||||
| 3 | 6 | 18 | 30 | 48 | 75 | 120 | 180 | 300 | 480 | 760 | |||||||||
| 6 | 10 | 27 | 36 | 68 | 90 | 60 | 220 | 360 | 580 | 900 | |||||||||
| 10 | 18 | 27 | 43 | 70 | 110 | 180 | 270 | 430 | 700 | 1100 | |||||||||
| 18 | 30 | 33 | 52 | 84 | 130 | 210 | 330 | 520 | 840 | 1300 | |||||||||
| 30 | 50 | 39 | 62 | 100 | 160 | 260 | 390 | 620 | 1000 | 1600 | |||||||||
| 50 | 80 | 46 | 74 | 120 | 190 | 300 | 460 | 740 | 1200 | 1900 | |||||||||
| 80 | 120 | 64 | 87 | 140 | 220 | 350 | 540 | 870 | 1400 | 2200 | |||||||||
| 120 | 180 | 60 | 100 | 160 | 250 | 400 | 630 | 1000 | 1600 | 2500 | |||||||||
| 180 | 250 | 72 | 115 | 185 | 290 | 460 | 720 | 1150 | 1850 | 2900 | |||||||||
| 250 | 315 | 81 | 130 | 210 | 320 | 620 | 810 | 1300 | 2100 | 3200 | |||||||||
| 315 | 400 | 89 | 140 | 230 | 360 | 570 | 890 | 1400 | 2300 | 3600 | |||||||||
| 400 | 500 | 97 | 155 | 250 | 400 | 630 | 970 | 1500 | 2500 | 4000 | |||||||||
| 500 | 630 | 110 | 175 | 280 | 440 | 700 | 1100 | 1750 | 2800 | 4400 | |||||||||
| 630 | 800 | 125 | 200 | 320 | 500 | 800 | 1250 | 2000 | 3200 | 6000 | |||||||||
| 800 | 1000 | 140 | 230 | 360 | 560 | 900 | 1400 | 2300 | 3600 | 5600 | |||||||||
| 1000 | 1250 | 165 | 260 | 420 | 660 | 1060 | 1650 | 2600 | 4200 | 6600 | |||||||||
| 1250 | 1600 | 195 | 310 | 500 | 780 | 1260 | 1950 | 3100 | 5000 | 7800 | |||||||||
| 1600 | 2000 | 230 | 370 | 600 | 920 | 1500 | 2300 | 3700 | 6000 | 920 | |||||||||
| 2000 | 2500 | 280 | 440 | 700 | 1100 | 1750 | 2800 | 4400 | 7000 | 11000 | |||||||||
| 2500 | 3150 | 330 | 540 | 860 | 1360 | 2100 | 3300 | 5400 | 8000 | 1350 | |||||||||
Расчет международных классов допуска:
Как упоминалось ранее, IT Grade имеет 18 стандартных допусков.Эти ИТ-классы имеют свои собственные числовые значения для каждой ступени диматора, приведенные в таблицах выше. IT-классы IT01 — IT5 являются наиболее точными и используются в основном для манометров. IT-классы IT6 — IT 16 менее точны и используются для несвязанных Габаритные размеры. Международный рейтинг можно рассчитать, как указано ниже;
T: Международный класс допуска в (мкм)
D: Средний геометрический размер в миллиметрах (мм)
ITG: IT-класс (положительное целое число)
Также T: международный Терпимость может проявляться как;
«i»: — единица / коэффициент стандартного допуска. «i» для значений от IT5 до IT 16 можно рассчитать по формуле ниже;
Допуск в (мкм). Для вычисления D;
где, D (мм) — среднее геометрическое нижнего и верхнего диаметров конкретный шаг диаметра, в пределах которого находится выбранный диаметр D. Стандартная единица допуска / коэффициент формула имеет параболическую связь с размером продукты. По мере увеличения размера допуск, в пределах которого деталь может быть производство тоже увеличивается.
Значения допусков по стандартам, соответствующие IT-классам IT01, IT0, IT1, см. Таблицу ниже
| МЕЖДУНАРОДНЫЕ КЛАССЫ ДОПУСКА | |||
| Значения в микронах | IT01 | IT0 | IT1 |
| Значения для D, мм | 0,3 + 0,008D, мм | 0,3 + 0,008D 0,012D | 0,8 + 0,020D |
Значения допусков по стандартам, соответствующие IT-классам от IT5 до IT16, см. Таблицу ниже
| МЕЖДУНАРОДНЫЕ СТЕПЕНИ ДОПУСКА | ||||||||||||||||||
| Значения в микронах | IT5 | IT6 | IT7 | IT8 | IT9 | IT10 | IT11 | IT12 | IT13 | IT14 | IT15 | IT16 | ||||||
| Значения для D в мм | 7i | 10i | 16i | 25i | 40i | 64i | 100i | 160i | 250i | 400i | 640i | 1000i | ||||||
ПРИМЕР:
Давайте выберем диаметр вала 60 мм и класс допуска H8f7; Расчет для Фундаментальное отклонение, допуски и предел размера отверстия и вала.Расчет стандартной единицы допуска;
Диапазон размеров для диаметра 60 мм будет 50-80 мм из таблицы ISO 286
D = √ 50 +80 мм = 63,25 мм
i = 10 -3 √ 50 +60 + 0,01×63,25 = 1,865 мкм или i = 0,001865 мм
Для вала f7 значение стандартной единицы «i» из таблицы;
Допуск = 16i = 16×1,865 = 0,030 мм
Для отверстия H8 стандартная единица измерения значения «i» из таблицы;
Допуск = 25i = 25×0.001865 = 0,046 мм
Поскольку конструкция основана на отверстии, основное отклонение отверстия составляет нуль. Фундаментальное отклонение вала;
= -5,5xD 0,41
= -5,5×63,25 0,41
= -30,114 мкм = 0,030 мм
Вал;
Верхний предел = основной размер — фундаментальное отклонение = 60-0,030 = 59,97 мм
Нижний предел = Верхний предел-допуск = 59,97-0,03 = 59,94 мм
Отверстие;
Нижний предел = Базовый размер = 60 мм
Верхний предел = нижний предел + допуск = 60 + 0.046 = 60,046 мм
Дендритные клетки, полученные из кожи, могут опосредовать делеционную толерантность к самореактивным Т-клеткам с ограничением класса I
Abstract
Кожно-дренирующие лимфатические узлы содержат ряд подмножеств дендритных клеток (ДК) различного происхождения. Некоторые из них являются мигрирующими, например происходящие из кожи эпидермальные клетки Лангерганса и отдельная подгруппа дермальных DC, тогда как другие являются лимфоидно-резидентными по своей природе, такие как CD8 + DC, обнаруженные во всех лимфоидных тканях.В этом исследовании мы изучаем подмножество DC-презентации кожного само-Ag мигрирующими и лимфоидно-резидентными DC, как в стабильном состоянии, так и в условиях локальной кожной инфекции. Мы показываем, что представление self-Ag ограничивается кожными мигрирующими DCs в обоих случаях. Устойчивое представление привело к делеционной толерантности Т-клеток, несмотря на то, что эти DC экспрессировали относительно зрелый фенотип, измеренный с помощью традиционных маркеров, таких как уровень экспрессии MHC класса II и CD86. Таким образом, собственный Ag может переноситься в дренирующие лимфатические узлы кожными ДК и там представляться этими же клетками для толеризации пула циркулирующих Т-клеток.
Внутри кожи есть две широкие популяции дендритных клеток (DC) 3 : клетки Лангерганса, обнаруженные в эпидермисе кожи, и отдельный пул DC в подлежащей дерме (1, 2, 3, 4, 5). Клетки Лангерганса и дермальные ДК представляют собой мигрирующие подмножества, которые мобилизуются инфекцией, травмой или воспалением кожи, и, как таковые, они долгое время считались ключевыми медиаторами Т-клеточного иммунитета во время кожной инфекции. Первоначально считалось, что кожный иммунитет является результатом скоординированного созревания этих клеток до иммуногенной формы в сочетании с представлением приобретенного кожей Ag.Однако теперь выясняется, что ситуация намного сложнее, чем описано в этой парадигме, с представлением, выходящим за пределы мигрирующей популяции DC (6, 7, 8, 9, 10). Хотя определенные патогены опосредуют активацию Т-клеток через кожных эмигрантов (8, 10), иммунитет к другим типам кожных инфекций, по-видимому, обусловлен исключительно лимфоидно-резидентными DC, а именно субпопуляцией CD8 + DC. Например, инфицирование цитопатическими вирусами, такими как вирусы простого герпеса и вирусы осповакцины, приводит к представлению, ограниченному классом I, которое почти исключительно обнаруживается в этой немигрирующей популяции.Более того, недавние данные свидетельствуют о том, что кожные ДК могут вызывать реакции толерогенного или регуляторного типа (11, 12). Это последнее наблюдение особенно интригует, потому что исследование дренирующих кожу лимфатических узлов (ЛУ) показывает, что мигрирующие ДК являются наиболее зрелыми, что определяется фенотипическими маркерами, такими как костимулирующие молекулы CD80 и CD86 (2, 13). Действительно, Майерова и соавт. (14) утверждали, что презентация экспрессируемого кожей собственного пептида мигрирующими ДК вызывает преимущественно аутоиммунный Т-клеточный ответ CD8 + как прямое следствие состояния созревания мигрирующей популяции, которое, как они полагают, возникает как следствие непрерывного воздействия окружающей среды. испытание.Принимая во внимание эти противоречивые представления о функции кожных ДК, мы повторно исследовали вопрос о подмножестве ДК представлений кожного собственного Ag. Наши результаты показывают, что, в отличие от того, что наблюдается при кожной инфекции цитопатическим вирусом, презентация собственного Ag происходит с помощью зрелых мигрирующих DCs как дермального, так и эпидермального происхождения, и эта презентация приводит к долговременной делеционной толерантности.
Материалы и методы
Мыши
C57BL / 6 (B6), B6.SJL-Ptprc a Pep3 b / BoyJ (B6.Ly5.1), B6.CH-2 bm-1 (bm1) (15), RIP.OVA high (16), K5m.OVA (17), мышей K5.mOVA × bm1, K5.mOVA × B6.Ly5.1, OT-I × B6.Ly5.1 и gBT-I × B6.Ly5.1 разводили и содержали в Отделении микробиологии. и иммунологии, Мельбурнский университет или Институт медицинских исследований Уолтера и Элизы Холл.
Получение и адаптивный перенос CFSE-меченных Т-клеток OT-I или gBT-I
T-клетки OT-I или gBT-I получали путем создания одноклеточной суспензии LN (подмышечных, плечевых, паховых, шейных и брыжеечных) и / или клеток селезенки от трансгенных мышей OT-I или gBT-I.В некоторых экспериментах очищали Т-клетки OT-I или gBT-I. Суспензии отдельных клеток инкубировали в течение 30 мин с заранее определенными оптимальными концентрациями следующих очищенных моноклональных антител: анти-Mac-1α (M1 / 70), анти-F4 / 80 (F4 / 80), антиэритроцитов (TER-119), анти-GR-1 (RB68C5), анти-IA / IE (M5114) и анти-CD4 (GK1.5). Затем клетки, покрытые антителом, удаляли путем инкубации с магнитными шариками, связанными с овечьим антителом к IgG крысы (Dynabeads: Dynal Biotech) или магнитными шариками с козьим антителом против IgG крысы (Qiagen). Чистоту определяли путем исключения клеток, окрашенных анти-CD8 и Vα2, пропидиум иодидом (PI).
Для маркировки CFSE Т-клетки OT-I или gBT-I ресуспендировали в PBS, содержащем 0,1% BSA (Sigma-Aldrich), и метили 2,5 мкМ CFSE (молекулярные зонды) в течение 10 мин при 37 ° C. Затем клетки дважды промывали HEPES MEM. Всего 10 6 CFSE-меченных OT-I Т-клеток (Ly5.1 + , CD8 + и Vα2 + ) были адоптивно перенесены с помощью внутривенного введения. инъекция мышам-реципиентам. Через 42 часа объединенные плечевые и паховые LN от отдельных мышей анализировали с помощью проточной цитометрии на пролиферацию Ly5.1 + CD8 + CFSE + PI — клеток. Для экспериментов in vivo, направленных на анализ пролиферации, использовали необогащенные CFSE-меченные OT-I Т-клетки из LN. Для экспериментов, включающих делецию in vivo и стимуляцию наивных CD8 + Т-клеток с помощью DC in vitro, использовали очищенные Т-клетки ОТ-I или gBT-I.
Вирусные инфекции
МышейK5.mOVA в возрасте 6–12 недель инокулировали 1 × 10 6 БОЕ HSV (штамм KOS HSV-1) с обеих сторон с помощью модели скарификации с боков, как описано ранее (18).Мышам K5.mOVA или C57BL / 6 в возрасте 6–12 недель прививали 500 БОЕ рекомбинантного вируса гриппа, несущего ограниченный MHC класс I OVA, инъецированный подкожно. в каждую заднюю лапу.
Проточно-цитометрический анализ экспансии Т-клеток OT-I
МышейK5.mOVA или C57BL / 6, которые получили 1 × 10 5 клеток LN OT-I × B6.Ly5.1, 4 неделями ранее инокулировали рекомбинантным вирусом гриппа, несущим OVA, ограниченный MHC класса I, путем инфицирования лапкой. Через семь дней после заражения мышей умерщвляли и удаляли их селезенки.Суспензии единичных клеток окрашивали анти-Ly5.1-FITC (A20), анти-Vα2-PE (B20.1) и анти-CD8α-аллофикоцианином (53-6,7). Окрашенные клеточные растворы анализировали проточной цитометрией с использованием ИП для исключения мертвых клеток.
Изоляция DC от LN
S.c. дренирующие ЛУ (плечевые, паховые и шейные) удаляли у мышей. LN разрезали на мелкие фрагменты и переваривали в течение 20 мин при комнатной температуре в растворе коллагеназы / ДНКазы (1 мг / мл коллагеназы типа II; Worthington Biochemicals) и 0.1% бычьей ДНКазы I поджелудочной железы II степени (Boehringer Mannheim) в мышиной тоничности RPMI 1640, содержащей 10% FCS, 50 мкМ 2-ME, 2 мМ l-глутамин, 100 Ед / мл пенициллина и 100 мкг / мл стрептомицина (полная среда RPMI ). Для разрушения комплексов Т-клетки-ДК к суспензии добавляли 0,1 М ЭДТА и расщепляли еще 5 мин при комнатной температуре. Все последующие процедуры проводились в сбалансированном солевом растворе / ЭДТА / 5% FCS. ДК очищали из суспензий одиночных клеток, как описано ранее (7). Вкратце, суспензии отдельных клеток инкубировали в течение 30 мин с заранее определенными оптимальными концентрациями следующих очищенных моноклональных антител: анти-CD3ε (KT3), анти-Thy-1 (T24 / 31.7), анти-CD19 (ID3) и антиэритроцит (TER-119). Затем клетки, покрытые антителом, удаляли путем инкубации с магнитными шариками, связанными с овечьим антителом против IgG крысы (Dynabeads; Dynal Biotech). Для очистки LN DC до их различных фенотипических популяций, DC были помечены анти-CD11c-FITC (N418), анти-CD205-PE (NLDC-145) и анти-CD8α-аллофикоцианином (53-6.7; BD Pharmingen), специфичными. mAb и отсортировали на проточном цитометре MoFlo (Cytomation).
ДК, экспрессирующие лангерин, были идентифицированы путем инкубации клеток в течение 30 минут с анти-CD16 / 32 (2.4G2) супернатант перед окрашиванием антителом против CD207 мыши (лангерин, 205C1; AbCys). Связанные с антителом клетки детектировали с использованием антимышиного IgM a -PE (AF6-78; BD Pharmingen). ИП использовали для исключения мертвых клеток из сортов.
Совместное культивирование субпопуляций DC с CFSE-меченными Т-клетками OT-I или gBT-I
Двукратные серийные разведения, начиная с 2,5 × 10 4 каждой популяции DC были совместно культивированы in vitro с 5 × 10 4 CFSE-меченных Т-клеток OT-I или gBT-I в 200 мкл полной среды RPMI в V -днижние планшеты для культивирования тканей (Costar).Пролиферацию измеряли как потерю окрашивания CFSE клеток CD8 + Ly5.1 + PI —, как определено с помощью проточного цитометрического анализа (FACSCalibur; BD Biosciences) после 60 часов культивирования.
Анализ делеции Т-клеток
Всего 5 × 10 6 Т-клеток OT-I (Ly5.1 + , CD8 + , Vα2 + ) были адоптивно перенесены в мышей-реципиентов, и через 6 недель мышей анализировали. Клетки собирали из подмышечных, плечевых, паховых, шейных и брыжеечных LN и селезенки каждой мыши-реципиента.Клетки окрашивали анти-Ly5.1-FITC (A20) и анти-CD8α-аллофикоцианином (53-6,7) вместе с анти-Vα2-PE (B20.1) или H-2K b -OVA 257 –264 -ПЭ тетрамер. Живые лимфоциты определяли по профилям прямого и бокового рассеяния вместе с исключением PI. Анализ проводили на проточном цитометре FACSCalibur. Всего для анализа было собрано 2 × 10 5 живых клеток. Количество клеток OT-I на животное подсчитывали с использованием известного количества небольших нефлуоресцентных шариков Sphero (BD Biosciences), добавленных к известному объему образца клеток.Следовательно, клеток на миллилитр = количество собранных клеток × (количество гранул в образце / количество собранных гранул) / объем образца (19).
Генерация химер костного мозга
Химерных мышей получали путем облучения мышей-реципиентов двумя дозами по 550 сГр с интервалом в 3 часа и восстанавливали 5 × 10 6 истощенных Т-клеток донорских клеток костного мозга. Клетки донорского костного мозга были истощены Т-клетками путем мечения клеточных суспензий следующими mAb: анти-CD4 (RL172), анти-CD8 (3.168) и анти-Thy-1 (J1J). Клетки, покрытые Ab, удаляли инкубацией с кроличьим комплементом в течение 30 минут при 37 ° C. На следующий день остаточные радиоустойчивые Т-клетки были истощены с помощью 100 мкл асцита Т24 (анти-Thy-1) внутрибрюшинно. Мышам давали возможность восстановиться в течение 8–10 недель перед использованием.
Выделение ДК из биопсий кожи
Дермальные DC и / или эпидермальные DC были выделены либо из цельной кожи, либо из эпидермальных слоев, соответственно, путем культивирования в присутствии 6Ckine (R&D Systems), как описано ранее (7).Мышей подрезали по бокам и депилировали с помощью Veet (Reckitt Benckiser). Снимали шкуру на всю толщину с области паха. S.c. ткань удаляли с помощью лезвия скальпеля, а кожу разрезали на мелкие кусочки. Кожу помещали (дермальной стороной вниз) в 1 мл полной среды RPMI, содержащей 0,1 мкг рекомбинантного мышиного 6Ckine, для стимуляции миграции DC. После 24 ч инкубации при 37 ° C эмигрантные клетки собирали и помещали при 4 ° C. Кожу переносили в свежую среду, содержащую 6Ckine, и инкубировали еще 24 ч при 37 ° C.Клетки, которые мигрировали в культуральную среду во время первой и второй инкубации, объединяли перед окрашиванием для анализа FACS.
Для DC, полученных из выползания эпидермального листа, кусочки кожи помещали дермальной стороной вниз в полную среду RPMI, содержащую 2,5 мг / мл диспазы II (Roche), в течение 90 мин при 37 ° C. Листы эпидермиса снимались с дермы и обрабатывались так же, как и при выползании по всей коже. Фенотип ДК, выделенных из кожи, анализировали путем окрашивания изолированных клеток различными комбинациями следующих антител: анти-CD45.2-FITC (104; BD Pharmingen), анти-CD205-PE (NLDC-45) или анти-IA b -PE (AF6-120.1; BD Pharmingen) и анти-CD11c-биотин (HL3; BD Pharmingen) или анти-H-2K b -биотин (5F1), за которым следует аллофикоцианин-стрептавидин (BD Pharmingen). Мертвые клетки были исключены с помощью ИП. Анализ проводили на проточном цитометре FACSCalibur.
Прямое выделение эпидермальных DC из биопсии кожи
Для прямого выделения эпидермальных DC кожу вырезали, как описано выше.Небольшие кусочки кожи помещали дермальной стороной вниз в полную среду RPMI, содержащую 2,5 мг / мл диспазы II, в течение 90 минут при 37 ° C. Листы эпидермиса были отделены от дермы и гомогенизированы с помощью лезвия скальпеля. Затем образцы переваривали в 3 мл коллагеназы (3 мг / мл коллагеназы типа III; Worthington Biochemicals) в мышином тонусе RPMI 1640, содержащем 2% FCS, 50 мкМ 2-ME, 2 мМ l-глутамин, 100 Ед / мл пенициллина и 100 мкг / мл стрептомицина) на мышь в течение 30 мин при 37 ° C. Образцы предварительно инкубировали с анти-CD16 / 32 (2.4G2 супернатант) для блокирования неспецифического связывания Ab. Все последующие процедуры выполнялись в сбалансированном солевом растворе / ЭДТА / 5% FCS. Образцы окрашивали анти-CD11c-биотином (HL3; BD Pharmingen) и анти-лангерином (205C1; AbCys), затем стрептавидином, конъюгированным с аллофикоцианином (BD Pharmingen) и антимышиным IgM a -PE (AF6-78; BD Фарминген). Другими использованными mAb были анти-I-A b -FITC (AF6-120.1; BD Pharmingen), анти-CD86-FITC (GL1) и анти-CD40-FITC (FGK45.5). ИП был включен для исключения мертвых клеток.Анализ проводили на проточном цитометре FACSCalibur.
Результаты
Дермальные ДК и клетки Лангерганса представляют собой кожный само-Ag
Azukizawa et al. (17) получили трансгенную мышь, экспрессирующую мембраносвязанную форму OVA под контролем кератинового промотора K5. У этой мыши экспрессия Ag была ограничена выбранными тканями, включая эпидермис кожи, и это привело к преимущественной пролиферации CFSE-меченных OVA-специфических Т-клеток OT-I в дренирующих кожу LNs (рис.1⇓ A ). Мы хотели изучить представление Ag подмножества постоянного тока в этих LN. Учитывая, что экспрессия трансгена ограничена эпидермисом кожи, мы сначала оценили, представляют ли DC, происходящие из этого местоположения, клетки Лангерганса, пептид OVA, ограниченный классом I. С этой целью клетки CD11c + из дренирующих кожу ЛУ были выделены на основе экспрессии маркера, специфичного для клеток Лангерганса, CD207 (лангерин (20)) и CD8 (рис. 1– B ). Последний был включен, потому что CD8 + DC могут представлять Ag после кожной вирусной инфекции (6, 7, 9).Очищенные популяции DC использовали для стимуляции CFSE-меченных OVA-специфических Т-клеток от трансгенной мыши OT-I (21) в анализе пролиферации in vitro. На рис. 1- C показано, что, хотя очищенное потомство клеток Лангерганса может эффективно представлять Ag, ограниченный классом I, активность в субпопуляции CD8 + DC невысока. Интересно, что некоторая презентация также наблюдается в популяции CD207 — CD8 — , которая состоит из ряда отдельных подмножеств DC, включая дермальные и миелоидные, немигрирующие резидентные DC.
РИСУНОК 1. Пролиферация Т-клетокOT-I ограничена подкожной клетчаткой. LN в мышах K5.mOVA и клетках Лангерганса представляют собой кожный самогенерируемый Ag. A . Всего 10 6 CFSE-меченных клеток OT-I × Ly5.1 адоптивно переносили реципиентам K5mOVA. Через 42 часа дренирующие кожу паховые и плечевые ЛУ, а также не дренирующие кожу брыжеечные ЛУ были проанализированы с помощью проточной цитометрии на пролиферирующие клетки Ly5.1 + CD8 + CFSE + PI — клеток. B , ДК были выделены из п / к. LN мышей K5mOVA. CD11c + DC были отсортированы методом проточной цитометрии на CD8 — CD207 high клетки Лангерганса, CD8 — CD207 — двойные отрицательные (DN) / дермальные DC и CD8 + CD207 — . C . Двукратные разведения очищенных субпопуляций DC культивировали совместно с 5 × 10 4 CFSE-меченных Т-клетками OT-I in vitro при 37 ° C. Через 60 часов культивирования CD8 + OT-I T-клетки анализировали с помощью проточной цитометрии на пролиферацию.Данные объединены из трех независимых экспериментов, показывающих среднее значение ± стандартная ошибка среднего. ДК, выделенные из нетрансгенных мышей C57BL / 6 (B6), не индуцировали пролиферацию CD8 + Т-клеток ОТ-I (данные не показаны).
Мы думали, что представленная популяция в подмножестве CD207 — CD8 — , показанном на фиг. 1⇑, будет преимущественно состоять из дермальных DC. Чтобы показать это, DC CD11c + были разделены на основе экспрессии CD205 (DEC205) и CD8. В этой ситуации различные CD8-отрицательные DC кожного происхождения можно дифференцировать на основе уровней экспрессии CD205 (2, 3).Потомство клеток Лангерганса экспрессирует наивысшие уровни этого маркера, что ясно видно при сравнении профиля ДК, происходящих в коже (которые содержат клетки Лангерганса CD205 high ) с таковыми из не дренирующих кожу ЛУ, таких как мезентериальные ЛУ (где Клетки Лангерганса отсутствуют) (рис. 2⇓ А ). Напротив, интерстициальные DC, такие как дермальные DC, дают рост популяции клеток CD205 int , обнаруживаемых в дренирующих кожу и не дренирующих кожу ЛУ. На рис. 2– B показано, что DC CD8 + представляют мало Ag, ограниченного классом I, что согласуется с результатами на рис.1⇑. Напротив, дермальные DC и клетки Лангерганса способствовали хорошей пролиферации Т-клеток OT-I. Чтобы дополнительно исключить, что презентация дермальных DC в предыдущих анализах не была частично связана с загрязнением клеток Лангерганса, мы показали, что дермальные DC, очищенные как CD207 — CD205 в клетках CD8 —, могут управлять пролиферацией OT-I ( Рис.2⇓ C ). Таким образом, обе популяции кожных ДК, кожные ДК и клетки Лангерганса, по-видимому, несут OVA из места его экспрессии для представления ограниченным классом I способом в дренирующих кожу LN.
РИСУНОК 2.Само-Ag кожного происхождения представлено DC кожного происхождения. A , DC были изолированы от п / к. или брыжеечные LN мышей K5mOVA. CD11c + DC были отсортированы методом проточной цитометрии на CD8 — CD205 high клетки Лангерганса, CD8 — CD205 внутри дермальных DC, CD8 — CD205 — CD158 и двойные отрицательные DC — CD205 внутри DC. B , Двукратные разведения очищенных субпопуляций DC совместно культивировали с 5 × 10 4 CFSE-меченых Т-клеток OT-I in vitro при 37 ° C.Через 60 часов культивирования CD8 + OT-I T-клетки анализировали с помощью проточной цитометрии на пролиферацию. Данные объединены из четырех независимых экспериментов, показывающих среднее значение ± стандартная ошибка среднего. C , CD11c + DC были разделены проточной цитометрией на CD8 — CD207 + клетки Лангерганса и CD207 — DC были дополнительно разделены на CD8 — CD205 в DC + в дерме. int DC. Двукратные серийные разведения очищенных субпопуляций DC (6.25 × 10 4 DC (показано) были совместно культивированы с 5 × 10 4 CFSE-меченых Т-клеток OT-I in vitro при 37 ° C. Через 60 часов культивирования CD8 + OT-I T-клетки анализировали с помощью проточной цитометрии на пролиферацию.
Радиоустойчивые кожные ДК могут управлять пролиферацией Т-клеток у трансгенных мышей с кожно-специфической экспрессией Ag
Известно, что предшественники клеток Лангергансаустойчивы к радиации (7, 22), хотя в начале этого исследования было неясно, в какой степени кожные ДК замещаются в этих химерах костного мозга.Мы решили эту проблему, сравнив мигрирующие ДК в культурах эксплантатов эпидермальных пластов (которые должны содержать исключительно клетки Лангерганса) и всей кожи (которые также имеют компонент ДК дермального происхождения) от химерных мышей костного мозга, различающихся экспрессией аллотипов CD45 (рис. 3). ⇓). Опять же, клетки Лангерганса можно отличить от кожных DC по более высокой экспрессии CD205 (2, 3). На рис. 3 показано, что клетки Лангерганса CD205 high , мигрирующие из эпидермальных слоев, почти полностью принадлежали хозяину (CD45.2) происхождение (рис. 3⇓ A ), подтверждающее их устойчивость к облучению. Напротив, большая часть мигрантов из цельной кожи имела донорское (CD45.1) происхождение (рис. 3⇓ B ). CD205 int дермальные DC в этих эмигрирующих популяциях подверглись замене на ~ 90%, в то время как большинство клеток с высоким содержанием CD205 имели происхождение от хозяина, что отражает их классификацию клеток Лангерганса. Это несколько более низкие уровни дермального замещения DC, чем недавно сообщалось Bogunovic et al.(23), которые обнаружили, что около 25% кожных ДК выживают после облучения. Все другие DC, не производные от кожи, в дренирующих LN происходят от хоста (данные не показаны). Таким образом, облученные химеры содержат популяцию в основном донорских ДК кожи и некожного происхождения по сравнению с клетками Лангерганса, происходящими от хозяина.
РИСУНОК 3.Различие между клетками Лангерганса и кожными DC с использованием химер, облученных костным мозгом. A , Эпидермальные листы, преимущественно содержащие клетки Лангерганса, или вся кожа, содержащая клетки Лангерганса и дермальные DC ( B ), выделенные из B6.Химерных мышей костного мозга Ly5.1 (CD45.1) → B6 (CD45.2) культивировали в течение 48 часов в полной среде, содержащей 6Ckine. Клетки, мигрирующие в среду, собирали в отдельные 24-часовые пулы и объединяли для окрашивания и анализа. Клетки окрашивали на предмет их экспрессии CD11c, CD205 и CD45.2 и процент ± стандартное отклонение клеток Лангерганса (CD11c + CD205 высокий ) или дермальных DC (CD11c + CD205 int. ), которые экспрессировали CD45. .2 был определен.
ВкладDC в стимуляцию Т-клеток in vivo можно оценить в контексте системы OVA Ag, используя химеры костного мозга с использованием донорских клеток от мыши bm1, которая несет молекулу класса I, неспособную представить детерминанту, распознаваемую ОТ. -I Т-клетки (24).Учитывая, что рис. 2⇑ показывает презентацию дермальными и происходящими из эпидермиса DCs и что значительная часть кожных DCs выживает после облучения (рис. 3⇑), можно было бы предсказать, что ограниченная классом I презентация будет сохраняться у химерных животных bm1 → K5mOVA. Проще говоря, у этих химер есть большая часть клеток Лангерганса и даже небольшая часть дермальных DCs, все еще способных представлять приобретенный кожей Ag. Однако изначально было описано, что именно эти химеры не обладают стимулирующей способностью in vivo (17).Следует отметить, что исходные данные показали, что у этих животных сохранилась некоторая стимулирующая способность, и не было принято во внимание, что эта остаточная активность могла быть результатом выживших радиоустойчивых клеток Лангерганса. Уровень остаточного представления показан здесь на рис. 4⇓. Видно, что животные bm1 → K5mOVA поддерживают гораздо более слабую пролиферацию перенесенных CFSE-меченных клеток OT-I (рис. 4⇓ A ), чем химерные животные K5mOVA → K5mOVA (рис. 4⇓ B ). Хотя стимуляция перенесенных клеток OT-I снижена в химерах bm1 → K5mOVA, она явно сильнее, чем в отрицательных контролях, таких как животные, не экспрессирующие Ag K5mOVA → B6 или bm1 → B6 (рис.4⇓, C и D ). Оглядываясь назад, следует ожидать ослабленной, но не отсутствующей стимуляции Т-клеток OT-I, потому что животные K5mOVA → K5mOVA имеют как кожные ДК, так и клетки Лангерганса, способные презентовать K b -ограниченный пептид OVA, тогда как презентация преимущественно ограничивается подмножество клеток Лангерганса в химерах bm1 → K5mOVA. В целом, CD207 + клетки Лангерганса составляют <20% всех кожных ДК в LN (где последний измеряется как популяция CD205 + CD8 — CD11c + ) (рис.4⇓ E ), что объясняет более слабую стимуляцию, наблюдаемую у этих последних химерных животных, где клетки Лангерганса и относительно небольшое количество радиорезистентных дермальных DC являются единственными клетками, способными к презентации Ag. Наконец, надежная стимуляция Т-клеток OT-I в K5mOVA. Мыши Langerin-DTR, у которых есть рецептор дифтерийного токсина, управляемый промотором лангерина, даже после удаления клеток Лангерганса обработкой дифтерийным токсином (данные не показаны), исключают клетки Лангерганса как единственные участники презентации Ag, происходящего из кожи.
РИСУНОК 4. Само-Ag, полученное из кожи, представлено радиационно-чувствительными и устойчивыми к излучению APC. В общей сложности 10 6 CFSE-меченных OT-I T-клеток были адоптивно перенесены мышам K5mOVA с трансплантацией костного мозга bm1 ( A ) или K5mOVA.Ly5.1 ( B ) и C57BL / 6 (B6). мышам, которым трансплантировали костный мозг K5mOVA.Ly5.1 ( C ) или bm1 ( D ). Через 42 часа объединенные паховые и плечевые ЛУ анализировали с помощью проточной цитометрии на пролиферирующие клетки CD8 + CFSE + . E , Проточный цитометрический анализ процента CD207 + клеток Лангерганса, которые составляют пул CD205 + CD8 — CD11c + кожных ДК в кожных ЛУ наивных C57BL / 6 (B6 ) мышей.
Радиоустойчивые APC могут управлять OVA-специфической делецией Т-клеток у трансгенных мышей K5-mOVA
ДК, происходящие из кожи, такие как клетки Лангерганса, которые мигрировали в ЛУ, считаются прототипическим подмножеством зрелых ДК (25), особенно в отличие от их прямых незрелых предшественников в периферических тканях (26).Это различие в статусе созревания было подтверждено показом того, что полученные из кожи клетки CD11c + CD207 + (клетки Лангерганса) в LN экспрессировали высокие уровни маркеров созревания MHC класса II и CD86 (рис. 5⇓), тогда как их ткани -производные аналоги, клетки CD11c + , CD207 + в эпидермисе, демонстрировали низкий уровень экспрессии этих маркеров созревания. Следует отметить, что, хотя это окрашивание было слабым, оно, тем не менее, явно превышало фон. Наконец, CD205 int CD8 — дермальные DC в дренирующих кожу LN также демонстрируют зрелый статус при оценке с использованием этих костимулирующих маркеров (данные не показаны).
РИСУНОК 5.Проточный цитометрический анализ CD11c + CD207 + Экспрессия MHC II (I-A b ) и CD86 на поверхности клеток Лангерганса, свежевыделенных из эпидермальных пластов (кожи) или подкожно. LN.
Устойчивое представление периферического само-Ag может привести к делеционной толерантности (27). Это также можно увидеть у трансгенных мышей K5.mOVA, у которых перенесенные Т-клетки OT-I в значительной степени делетируются в течение 6-недельного периода (фиг. 6– A ). Это похоже на устранение, наблюдаемое в RIP.У мышей OVA high ранее было показано, что они поддерживают делеционную толерантность (28) и контрастируют с увеличенной выживаемостью этих Т-клеток в контрольных группах, не экспрессирующих Ag C57BL / 6 (B6). Удаление Т-клеток также наблюдается у мышей B6 → K5.mOVA.bm1 (рис. 6⇓ B ), что согласуется с заменой большинства дермальных ДК хозяина, несущих K bm1 , на ДК из B6, способных представления толерогенного пептида. Что наиболее важно, делеция очевидна у химерных животных bm1 → K5.mOVA (рис.6- B ), где эпидермальные клетки Лангерганса явно имеют происхождение от хозяина (фиг. 6- C ) и, следовательно, способны представлять пептид OVA, ограниченный классом I. Наконец, перенесенные Т-клетки OT-I являются функционально толерантными у мышей K5.mOVA, поскольку их размножение серьезно нарушается при последующем заражении рекомбинантным вирусом гриппа, несущим пептидную детерминанту OVA (фиг. 6 — D ). В сочетании с приведенными выше данными о созревании мигрирующих DC, когда они достигают LN, эти результаты предполагают, что зрелые потомки DC, которые происходят из кожи, могут управлять делеционной толерантностью аутореактивных Т-клеток.
РИСУНОК 6.Презентация собственного Ag кожного происхождения посредством DC кожного происхождения индуцирует делецию OVA-специфических CD8 + Т-клеток. A , Всего 5 × 10 6 Т-клеток OT-I были адоптивно перенесены в мышей C57BL / 6 (B6) (10 животных), трансгенных мышей RIP-OVA high (10 животных), трансгенных мышей K5mOVA (10 животных) или следующие химерные мыши из костного мозга: B , B6 → B6 (9 животных), bm1 → B6 (8 животных), bm1 → K5mOVA (13 животных), B6 → K5mOVA (13 животных) и В6 → К5мОВА.bm1 (11 животных). Через 6 недель количество T-клеток OT-I в LN и селезенке определяли с помощью проточной цитометрии. Показаны индивидуумы (○) и средние значения (-) для каждой группы. C , Эпидермальные листы, выделенные из химерных мышей bm1 → K5mOVA, культивировали в течение 48 часов в полной среде, содержащей 6Ckine. Клетки, мигрирующие в среду, собирали в отдельные 24-часовые пулы и объединяли для окрашивания и анализа. Гейты генерировали на клетках Лангерганса CD11c + MHC class II + (I-A b ), и эти клетки анализировали на экспрессию H-2K b (черный).Процент ± стандартное отклонение клеток Лангерганса, которые являются H-2K b + , показан на левой гистограмме . Чтобы показать химеризм, лимфоциты крови этих мышей также были проанализированы на предмет их экспрессии H-2K b (черный), а процент ± стандартное отклонение лимфоцитов, которые были H-2K b- , показан на правой гистограмме . Пунктирная линия обозначает немаркированные элементы управления. D , Всего 1 × 10 5 Т-клеток OT-I было адоптивно перенесено либо в C57BL / 6, либо в K5.Мыши-реципиенты mOVA. Через четыре недели реципиентов иммунизировали 500 БОЕ рекомбинантного вируса гриппа-OVA (WSN-OVA), чтобы обеспечить клональную экспансию и обнаружение Т-клеток OT-I. Через семь дней после иммунизации спленоциты окрашивали антителами, специфичными для CD8, Vα2 и Ly5.1, и анализировали проточной цитометрией.
Кожная инфекция не изменяет характер представления субнабора DC кожного собственного Ag
Кожная инфекция цитопатическими вирусами, такими как HSV и вирус осповакцины, связана с CD8 + DC-представлением ограниченного класса I Ag (6, 7, 8, 9).Нас интересовало, может ли такая инфекция изменить представление субпопуляции DC кожного само-Ag. С этой целью мы инфицировали мышей K5.mOVA вирусом простого герпеса и исследовали презентацию подмножества DC как собственного Ag Т-клеткам OT-I, так и чужеродного пептида gB HSV соответствующим вирусспецифическим Т-клеткам из gBT-I. трансгенная мышь (29). На фиг. 7⇓ не показано никаких изменений в преобладании представления кожных ДК в случае пептида OVA. Напротив, все представления HSV Ag были ограничены подмножеством CD8 + DC, как было обнаружено в предыдущих исследованиях.Таким образом, инфекция не изменяет характер представления собственного Ag, который по своей сути отличается от представления пептида, производного от вируса.
РИСУНОК 7.Кожная инфекция HSV не изменяет характер представления субнабора DC кожного собственного Ag. Субпопуляции DC выделяли из плечевых LN мышей K5mOVA через 2 дня после инокуляции с фланга HSV. CD11c + DC разделяли проточной цитометрией на CD8 — CD205 high клетки Лангерганса, CD8 — CD205 в дермальных DC и CD8 + CD205 в DC.Двукратные серийные разведения очищенных субпопуляций DC (показано 6,25 × 10 4 DC) сокультивировали либо с 5 × 10 4 Т-клетками OT-I, меченными CFSE, либо с Т-клетками gBT-I in vitro при 37 ° C. Через 60 часов культивирования как Т-клетки CD8 + ОТ-I, так и Т-клетки gBT-I анализировали с помощью проточной цитометрии на пролиферацию. Этот эксперимент был проведен дважды с аналогичными результатами.
Обсуждение
Вопросы относительно роли клеток Лангерганса в иммунном надзоре возникли после наблюдения, что эти клетки, по-видимому, не действуют при некоторых кожных инфекциях (7, 10, 30, 31), а также не являются незаменимыми при контактной сенсибилизации (32, 33).Результаты, представленные здесь и в других источниках, доказывают, что эти происходящие из кожи ДК могут приобретать Ag, происходящий из периферических тканей, для окончательной перекрестной презентации ограниченным классом I Т-клеткам. Местоположение этого приобретения остается неопределенным, хотя незрелый статус клеток в коже в сочетании с исследованиями, показывающими, что клетки Лангерганса могут поглощать Ag для перекрестной презентации (34), предполагают, что это устойчивое состояние отражает перенос Ag между кератиноцитами и кожей. -резидентные ДЦ. Кожные DC могут также напрямую контактировать с кератиноцитами или, что более вероятно, могут приобретать Ag из мигрирующих клеток Лангерганса, когда они пересекают дерму на своем пути к дренирующим LN (25).
Интересно, что ограниченная классом I презентация кожно-экспрессируемого Ag явно не происходит в случае кожной инфекции вирусами, такими как HSV, когда наблюдается исключение кожной презентации DC. Это может указывать на то, что этот тип инфекционного агента инактивирует путь перекрестной презентации, например, путем созревания этих клеток (35), или что он инактивирует сами DC, например, путем прямого уничтожения инфицированных клеток (36, 37). Оба этих сценария все еще совместимы с устойчивым представлением собственного Ag перед лицом инфекции, что предположительно отражает приобретение и миграцию Ag до заражения.
Мы показываем, что DC-презентация собственного Ag через кожу ведет к делеционной толерантности. Предположения о роли кожных ДК в понижающей модуляции Т-клеток не беспрецедентны. Было показано, что удаление клеток Лангерганса приводит к усилению контактной сенсибилизации в некоторых (12), но не в других (32) (33) системах трансгенных мышей, а у людей с псориазом обнаружены дефекты миграции клеток Лангерганса (11), предполагая, что эти клетки могут при некоторых обстоятельствах играть регулирующую роль, подавляя иммунные ответы.Однако ДК кожного происхождения также связаны с определенными типами противовирусного иммунитета, особенно с теми, которые связаны с нецитопатическими инфекциями (8). Таким образом, чисто толерогенная роль этой популяции может оказаться маловероятной. Тем не менее, DC-опосредованная толерантность кожи в устойчивом состоянии является неожиданной, учитывая, что эти клетки кажутся зрелыми в дренирующих LN, что определяется такими маркерами, как MHC класса II и экспрессия дополнительных молекул. Особенно ярко это проявляется в случае клеток Лангерганса. Это прототипические мигрирующие DC, и предполагалось, что сопутствующее фенотипическое созревание действует как ключевой драйвер Т-клеточного иммунитета.Хотя есть соблазн предположить, что фенотипические изменения, такие как повышающая регуляция костимулирующих молекул, тесно связаны с постоянным воздействием факторов окружающей среды внутри кожи, они могут просто составлять часть конститутивного пути миграции и, как таковые, не зависеть от прямая активация постоянным током (38). Как следствие, миграция может быть связана с более непрямым созреванием, которое, в свою очередь, не преобразует DC в их иммуногенную форму. Было показано, что в то время как активация DC в результате вовлечения TLR напрямую приводит к эффективному иммунитету, непрямое созревание, опосредованное цитокинами, в этом отношении является недостаточным (39).Таким образом, миграция клеток Лангерганса в отсутствие какой-либо формы дополнительной иммуногенной стимуляции, по-видимому, приводит к явно зрелой популяции DC, которая, тем не менее, способна управлять делеционной толерантностью Т-клеток.
Учитывая наблюдаемую здесь толерантность, наши результаты, по-видимому, противоречат более ранней работе Mayerova et al. (14) и Shibaki et al. (40), которые показали, что презентация собственного Ag кожными ДК приводит к разрушению аутоиммунной ткани. Мы не смогли объяснить эти различия, хотя следует отметить, что, хотя здесь мы демонстрируем устранение перенесенных Т-клеток ОТ-I, эти Т-клетки также проявляли аутоагрессивное поведение на микроскопическом уровне (17).В самом деле, делеция и аутоиммунитет не обязательно исключают друг друга. Известно, что презентация собственного Ag толерогенными APC может приводить к различным уровням активации Т-клеток до их функционального устранения, от довольно приглушенных ответов (41) до полномасштабной индукции эффекторной функции (42). Что контролирует этот очевидный континуум, который варьируется от в значительной степени пролиферативного ответа Т-лимфоцитов, как показано здесь, до явно аутоагрессивного ответа, как показано у Kurts et al. (27) остается неопределенным.
Таким образом, мы сосредоточились на различии медиаторов презентации, которые действуют во время вирусной инфекции кожи, и установлении толерантности к кожному Ag. В наших исследованиях толерантность опосредуется мигрирующими DC, включая клетки Лангерганса, происходящие из эпидермального слоя, несущего экспрессию Ag. Более того, эта толерантность достигается, несмотря на то, что эти клетки кажутся фенотипически зрелыми, когда достигают дренирующих кожу ЛУ.
Благодарности
Мы благодарим Кена Филда из Центра проточной цитометрии Мельбурнского университета за техническую помощь и доктора Э.Клаервену Джонсу за критическое прочтение рукописи.
Раскрытие информации
У авторов отсутствует финансовый конфликт интересов.
Footnotes
-
Затраты на публикацию этой статьи были частично покрыты за счет оплаты страницы. Таким образом, данная статья должна быть помечена как реклама в соответствии с 18 U.S.C. Раздел 1734 исключительно для указания этого факта.
-
↵1 Эта работа была поддержана грантами Австралийского национального совета по здравоохранению и медицинским исследованиям, Wellcome Trust и Медицинского института Говарда Хьюза.
-
↵2 Запросы на переписку и перепечатку направляйте доктору Уильяму Р. Хиту, отдел иммунологии, Институт медицинских исследований Уолтера и Элизы Холл, Парквилл 3050, Австралия; Адрес электронной почты: heath {at} wehi.edu.au или д-р Фрэнсис Р. Карбоне, факультет микробиологии и иммунологии, Мельбурнский университет, Parkville 3010, Австралия; Адрес электронной почты: fcarbone {at} unimelb.edu.au
-
↵3 Сокращения, используемые в этой статье: DC, дендритная клетка; LN — лимфатический узел; PI, иодид пропидия.
- Получено 9 февраля 2007 г.
- Принято 17 июля 2007 г.
- Авторские права © 2007 Американской ассоциации иммунологов
Ссылки
- ↵
Анхуэре, Ф., П. Мартин, И. Ферреро, М. Л. Фрага, Г. М. дель Ойо, Н. Райт, К. Ардавин. 1999. Определение субпопуляций дендритных клеток, присутствующих в селезенке, пятнах Пейера, лимфатических узлах и коже мыши. Кровь 93: 590-598.
- ↵
Анри, С., Д. Времек, А. Камат, Дж. Уэйтман, С. Уильямс, К. Бенуа, К. Бернем, С. Селанд, Э. Хэндман, К. Шотмэн. 2001. Популяции дендритных клеток лимфатических узлов мышей. J. Immunol. 167: 741-748.
- ↵
Камат А. Т., С. Генри, Ф. Бэтти, Д. Ф. Жесткий, К. Шотмэн. 2002. Кинетика развития и продолжительность жизни дендритных клеток в лимфоидных органах мышей.Кровь 100: 1734-1741.
- ↵
Ruedl, C., P. Koebel, M. Bachmann, M. Hess, K. Karjalainen. 2000. Анатомическое происхождение дендритных клеток определяет продолжительность их жизни в периферических лимфатических узлах. J. Immunol. 165: 4910-4916.
- ↵
Salomon, B., J. L. Cohen, C. Masurier, D. Klatzmann. 1998. Три популяции дендритных клеток лимфатических узлов мыши с разным происхождением и динамикой.J. Immunol. 160: 708-717.
- ↵
Смит, К. М., Г. Т. Белз, Н. С. Уилсон, Дж. А. Вилладангос, К. Шотмэн, Ф. Р. Карбоне, В. Р. Хит. 2003. Передовые технологии: обычные дендритные клетки CD8α + предпочтительно участвуют в праймировании CTL после инфицирования подушечек стопы вирусом простого герпеса-1. J. Immunol. 170: 4437-4440.
- ↵
Аллан Р. С., К. М. Смит, Г. Т. Белз, А.Л. ван Линт, Л. М. Ваким, В. Р. Хит, Ф. Р. Карбоне. 2003. Эпидермальный вирусный иммунитет, индуцированный дендритными клетками CD8α + , но не клетками Лангерганса. Наука 301: 1925-1928.
- ↵
He, Y., J. Zhang, C. Donahue, L. D. Falo, Jr. 2006. Дендритные клетки кожного происхождения индуцируют мощный Т-клеточный иммунитет CD8 + при генетической иммунизации, опосредованной рекомбинантным лентивектором. Иммунитет 24: 643-656.
- ↵
Белз, Г.Т., К. М. Смит, Д. Эйхнер, К. Шотмэн, Г. Карупиа, Ф. Р. Карбоне, В. Р. Хит. 2004. Передний край: обычные дендритные клетки CD8α + обычно участвуют в формировании иммунитета CTL к вирусам. J. Immunol. 172: 1996-2000.
- ↵
Iwasaki, A .. 2003. Важность дендритных клеток CD11b + в активации CD4 + Т-клеток in vivo: с помощью интерлейкина 1. J. Exp. Med. 198: 185-190.
- ↵
Камбербэтч, М., М. Сингх, Р. Дж. Дирман, Х. С. Янг, И. Кимбер, К. Э. Гриффитс. 2006. Нарушение миграции клеток Лангерганса при псориазе. J. Exp. Med. 203: 953-960.
- ↵
Каплан Д. Х., М. К. Дженисон, С. Селанд, В. Д. Шломчик, М. Дж. Шломчик. 2005. У мышей с дефицитом эпидермальных клеток Лангерганса развивается повышенная контактная гиперчувствительность. Иммунитет 23: 611-620.
- ↵
Banchereau, J., Р. М. Штейнман. 1998. Дендритные клетки и контроль иммунитета. Nature 392: 245-252.
- ↵
Майерова Д., Парк Э. А., Бурш Л. С., Одумаде О. А., Хогквист К. А. 2004. Клетки Лангерганса активируют наивные аутоантиген-специфичные Т-клетки CD8 в устойчивом состоянии. Иммунитет 21: 391-400.
- ↵
Вайс, Э. Х., А. Меллор, Л. Голден, К. Фарнер, Э. Симпсон, Дж. Херст, Р. А.Флавелл. 1983. Структура мутантного гена H-2 предполагает, что генерация полиморфизма в генах H-2 может происходить в результате событий, подобных конверсии генов. Nature 301: 671-674.
- ↵
Куртс, К., У. Р. Хит, Ф. Р. Карбон, Дж. Эллисон, Дж. Ф. Миллер, Х. Косака. 1996. Конститутивная экзогенная презентация аутоантигенов, ограниченная классом I in vivo. J. Exp. Med. 184: 923-930.
- ↵
Азукидзава, Х., Х. Косака, С. Сано, В. Р. Хит, И. Такахаши, X. Х. Гао, Ю. Сумикава, М. Окабе, К. Йошикава, С. Итами. 2003. Индукция опосредованного Т-клетками кожного заболевания, специфичного для антигена, трансгенно экспрессируемого в кератиноцитах. Евро. J. Immunol. 33: 1879-1888.
- ↵
ван Линт, А., М. Айерс, А. Г. Брукс, Р. М. Коулз, В. Р. Хит, Ф. Р. Карбоун. 2004. Т-лимфоциты CD8 + , специфичные к вирусу простого герпеса, могут устранять установленные литические инфекции кожи и нервов и могут частично ограничивать раннее распространение вируса после кожной инокуляции.J. Immunol. 172: 392-397.
- ↵
Дэйви, Г. М., К. Куртс, Дж. Ф. Миллер, П. Булье, А. Штрассер, А. Г. Брукс, Ф. Р. Карбоун, В. Р. Хит. 2002. Периферическая делеция аутореактивных Т-лимфоцитов CD8 путем перекрестной презентации аутоантигена происходит посредством Bcl-2-ингибируемого пути, опосредованного Bim. J. Exp. Med. 196: 947-955.
- ↵
Валладо, Дж., О. Равель, К. Дезуттер-Дамбуянт, К.Мур, М. Клеймер, Ю. Лю, В. Дювер-Фрэнсис, К. Винсент, Д. Шмитт, Дж. Даву и др. 2000. Лангерин, новый лектин С-типа, специфичный для клеток Лангерганса, является эндоцитарным рецептором. что вызывает образование гранул Бирбека. Иммунитет 12: 71-81.
- ↵
Hogquist, K. A., S. C. Jameson, W. R. Heath, J. L. Howard, M. J. Bevan, F. R. Carbone. 1994. Пептиды-антагонисты Т-клеточного рецептора вызывают положительный отбор. Ячейка 76: 17-27.
- ↵
Мерад, М., М. Г. Манц, Х. Карсунки, А. Вейджерс, В. Петерс, И. Чаро, И. Л. Вайсман, Дж. Г. Цистер, Э. Г. Энглеман. 2002. Клетки Лангерганса обновляются в коже на протяжении всей жизни в устойчивых условиях. Nat. Иммунол. 3: 1135-1141.
- ↵
Богунович, М., Ф. Гинху, А. Вейджерс, М. Лубо, Л. М. Изола, Л. Лубрано, В. Найфельд, Р. Г. Фелпс, К. Гросскройц, Э.Scigliano, et al. 2006. Идентификация радиорезистентной и цикличной популяции дендритных клеток дермы у мышей и мужчин. J. Exp. Med. 203: 2627-2638.
- ↵
Николич-Зугич, Дж., Ф. Р. Карбоне. 1990. Влияние мутаций в бороздке связывания пептида MHC класса I на распознавание цитотоксическими Т-лимфоцитами детерминанты овальбумина, ограниченной K b . Евро. J. Immunol. 20: 2431-2437.
- ↵
Ларсен, К.П., Р. М. Штейнман, М. Витмер-Пак, Д. Ф. Ханкинс, П. Дж. Моррис, Дж. М. Остин. 1990. Миграция и созревание клеток Лангерганса в кожных трансплантатах и эксплантатах. J. Exp. Med. 172: 1483-1493.
- ↵
Шулер Г., Р. М. Штейнман. 1985. Мышиные эпидермальные клетки Лангерганса созревают в мощные иммуностимулирующие дендритные клетки in vitro. J. Exp. Med. 161: 526-546.
- ↵
Курц, К., Х. Косака, Ф. Р. Карбоне, Дж. Ф. Миллер, У. Р. Хит. 1997. Ограниченная классом I перекрестная презентация экзогенных аутоантигенов приводит к делеции аутореактивных CD8 + Т-клеток. J. Exp. Med. 186: 239-245.
- ↵
Куртс, К., Р. М. Сазерленд, Г. Дэйви, М. Ли, А. М. Лью, Э. Бланас, Ф. Р. Карбоун, Дж. Ф. Миллер, В. Р. Хит. 1999. Незнание Т-лимфоцитов CD8 или толерантность к островковым антигенам зависит от дозы антигена. Proc. Natl.Акад. Sci. США 96: 12703-12707.
- ↵
Mueller, S. N., W. Heath, J. D. McLain, F. R. Carbone, C. M. Jones. 2002. Характеристика двух линий трансгенных мышей TCR, специфичных для вируса простого герпеса. Иммунол. Клетка. Биол. 80: 156-163.
- ↵
Чжао, X., Э. Дик, К. Содерберг, М. Линехан, Д. Спеццано, Дж. Чжу, Д. М. Книпе, А. Ивасаки. 2003. Дендритные клетки подслизистой оболочки влагалища, но не клетки Лангерганса, индуцируют защитные Th2-реакции на вирус простого герпеса-2.J. Exp. Med. 197: 153-162.
- ↵
Филиппи, К., С. Хьюг, Дж. Казарет, В. Джулия, Н. Глайхенхаус, С. Уголини. 2003. CD4 + Поляризация Т-клеток у мышей модулируется штаммами, независимыми от основных комплексов гистосовместимости, различиями внутри дендритных клеток. J. Exp. Med. 198: 201-209.
- ↵
Киссенпфенниг, А., С. Анри, Б. Дюбуа, К. Лаплас-Биль, П.Perrin, N. Romani, C.H. Tripp, P. Douillard, L. Leserman, D. Kaiserlian, et al. 2005. Динамика и функция клеток Лангерганса in vivo: дермальные дендритные клетки колонизируют области лимфатических узлов, отличные от более медленно мигрирующих клеток Лангерганса. Иммунитет 22: 643-654.
- ↵
Беннет, К. Л., Э. ван Рейн, С. Юнг, К. Инаба, Р. М. Штейнман, М. Л. Капсенберг, Б. Э. Клаузен. 2005. Индуцируемое удаление клеток Лангерганса мыши уменьшается, но не устраняет контактную гиперчувствительность.J. Cell. Биол. 169: 569-576.
- ↵
Стойцнер П., К. Х. Трипп, А. Эберхарт, К. М. Прайс, Дж. Й. Юнг, Л. Бёрш, Ф. Рончезе, Н. Романи. 2006. Клетки Лангерганса перекрестно представляют антиген, полученный из кожи. Proc. Natl. Акад. Sci. США 103: 7783-7788.
- ↵
Уилсон, Н. С., Г. М. Беренс, Р. Дж. Ланди, К. М. Смит, Дж. Уэйтман, Л. Янг, С. П. Форхан, А. Маунт, Р. Дж.Steptoe, K. D. Shortman, et al. 2006. Системная активация дендритных клеток лигандами Toll-подобных рецепторов или малярийная инфекция ухудшают перекрестную презентацию и противовирусный иммунитет. Nat. Иммунол. 7: 165-172.
- ↵
Джонс, К. А., М. Фернандес, К. Херк, Л. Босняк, М. Миранда-Саксена, Р. А. Боадл, А. Каннингем. 2003. Вирус простого герпеса 2 типа вызывает быструю гибель клеток и функциональное нарушение дендритных клеток мышей in vitro.J. Virol. 77: 11139-11149.
- ↵
Босняк, Л., М. Миранда-Саксена, Д. М. Коэль, Р. А. Боадл, К. А. Джонс, А. Л. Каннингем. 2005. Инфекция дендритных клеток человека вирусом простого герпеса вызывает апоптоз и допускает перекрестную презентацию через неинфицированные дендритные клетки. J. Immunol. 174: 2220-2227.
- ↵
Heath, W. R., J. A. Villadangos. 2005. Никакое вождение без водительских прав.Nat. Иммунол. 6: 125-126.
- ↵
Sporri, R., C. Reis e Sousa. 2005. Медиаторы воспаления недостаточны для полной активации дендритных клеток и способствуют увеличению популяций CD4 + Т-клеток, лишенных вспомогательной функции. Nat. Иммунол. 6: 163-170.
- ↵
Шибаки А., А. Сато, Дж. К. Фогель, Ф. Миягава, С. И. Кац. 2004. Индукция РТПХ-подобного кожного заболевания путем пассивного переноса CD8 + трансгенных Т-клеток рецептора Т-клеток трансгенным мышам с кератином 14-овальбумина.J. Invest. Дерматол. 123: 109-115.
- ↵
Эрнандес, Дж., С. Аунг, В. Л. Редмонд, Л. А. Шерман. 2001. Фенотипический и функциональный анализ CD8 + Т-клеток, подвергающихся периферической делеции в ответ на перекрестную презентацию аутоантигена. J. Exp. Med. 194: 707-717.
- ↵
Хуанг, К. Т., Д. Л. Хусо, З. Лу, Т. Ван, Г. Чжоу, Э. П. Кеннеди, К. Г. Дрейк, Д. Дж.Морган, Л. А. Шерман, А. Д. Хиггинс и др. 2003. CD4 + Т-клетки проходят через эффекторную фазу в процессе индукции толерантности in vivo. J. Immunol. 170: 3945-3953.
Учащиеся десятого класса демонстрируют всесторонние знания и навыки в области здоровья и хорошего самочувствия. Их поведение отражает концептуальное понимание проблем, связанных с поддержанием хорошего здоровья.Они служат сообществу, применяя методы, способствующие укреплению здоровья, которые способствуют благополучию на протяжении всей жизни (самосознание, социальная осведомленность, самоуправление, ответственное принятие решений и навыки взаимоотношений).
Ресурсы по санитарному просвещению для всех 10-х классов
- Стандарты санитарного просвещения для десятого класса изучения слов / PDF
- Пешеходный переход десятого класса: поиск полезных уроков по Вирджинии SOL Word / PDF
- Ссылки на ресурсы с инструкциями по охране здоровья для десятого класса Word / PDF
Системы тела
- Пожалуйста, проверьте единицы для обучения системам тела.
Сообщество / Здоровье окружающей среды
- Урок 1 — 10 класс — Гигиена окружающей среды и антикризисное управление PDF
- Урок 2 — 10 класс — Исследование карьеры в области здравоохранения PDF
Психическое благополучие / социальные и эмоциональные навыки
- Урок 1 — 10 класс — Обзор уроков по социальным эмоциональным навыкам PDF
- Урок 2 — 10 класс — Cyber Tattoo PDF
- Урок 3 — 10 класс — Лицо психического заболевания PDF / PPT
- Урок 4 — 10 класс — Пройдите милю в обуви PDF / PPT
- Урок 5 — 10 класс — Как намухи довольствовались тем, кто они есть PDF / PPT
- Урок 6 — 10 класс — Приспособление и установление границ здорового образа жизни PDF / PPT
- Урок 7 — 10 класс — Будьте умны со смартфонами (и экранами) PDF / PPT
- Урок 8 — 10 класс — Предотвращение самоубийств: в поисках слов PDF / PPT
- Урок 9 — 10 класс — Повышение самооценки и собственного имиджа PDF / PPT / PPT / PPT
- Урок 10 — 10 класс — Журналы психического здоровья — Оценка PDF
- Урок 11 — 10 класс — Отражение выступления на TED — Оценка PDF
- Урок 12 — 10 класс — Коэффициент эмоционального интеллекта — Оценка PDF
- Урок 13 — 10 класс — Психическое заболевание «Акулья цистерна» — Оценка PDF
- Урок 14 — 10 класс — Кампания по повышению осведомленности в старших классах — Оценка PDF
- Урок 15 — 10 класс — Навыки эффективного общения — Оценка PDF
- Урок 16 — 10 класс — Каково быть вами? — Оценка PDF
Питание
- Урок 1 — 10 класс — Анализ привычек семьи в бакалейных товарах PDF / PPT
- Урок 2 — 10 класс — Покажи мне сахар PDF
- Урок 3 — 10 класс — Модные диеты PDF
- Урок 4 — 10 класс — Лучший повар PDF / PPT / PPT
- Урок 5 — 10 класс — Образ тела и расстройства пищевого поведения PDF
- Урок 6 — 10 класс — Оценка аквариума по питанию PDF
- Урок 7 — 10 класс — В защиту оценки размышлений о питании PDF
Физическое здоровье и профилактика заболеваний / укрепление здоровья
- Урок 1 — 10 класс — Медицинские осмотры и страховое покрытие PDF
- Урок 2 — 10 класс — Ресурсы по укреплению здоровья PDF
- Урок 3 — 10 класс — Рак яичка 101 Видео PDF / ВИДЕО
- Урок 4 — 10 класс — Проект по постановке целей PDF
- Урок 5 — 10 класс — Здоровый день. Подростковый путь PDF / PPT
- Урок 6 — 10 класс — Health Dice; Здоровые решения PDF / PPT
- Урок 7 — 10 класс — Сладкие сны PDF
- Урок 8 — 10 класс — Служебное обучение PDF
- Урок 9 — 10 класс — Герои здоровья PDF / PPT / PPT
- Урок 10 — 10 класс — Не останавливайте сердце, используя альт-PDF / PPT
- Урок 11 — 10 класс — Подразделение по вовлечению сообщества — Мое общественное здоровье PDF / PPT
- Урок 12 — 10 класс — Wellness Challenge PDF / PPT
- Урок 13 — 10 класс — Задача оздоровления: Take II PDF
- Урок 14 — 10 класс — Медицинское страхование PDF
- Урок 15 — 10 класс — Проект защиты общественного здоровья PDF
- Урок 16 — 10 класс — Лучший повар PDF
- Урок 17 — 10 класс — Реклама на радио — Оценка проекта подкаста PDF
- Урок 18 — 10 класс — От A до ZZZ — Урок сна PDF / PPT
- Урок 19 — 10 класс — Весело на солнце безопасно PDF / PPT
Безопасность / предотвращение травм
- Урок 1 — 10 класс — Научно-образовательный проект по предупреждению травм и насилия PDF
- Урок 2 — 10 класс — Безопасное вождение и предотвращение DUI PDF
- Урок 3 — 10 класс — Скорая помощь PDF
- Урок 4 — 10 класс — Континуум здоровья PDF / PPT
- Урок 5 — 10 класс — Первая помощь и сердечно-легочная реанимация в день стихийных бедствий PDF
- Урок 6 — 10 класс — Лучше непрочитанный, чем мертвый — Текстовые сообщения во время вождения PDF
Профилактика злоупотребления психоактивными веществами
- Урок 1–10 класс — Исследования в отношении алкоголя, табака, наркотиков и практика отказа PDF
- Урок 2 — 10 класс — Под давлением PDF / PPT
- Урок 3 — 10 класс — Смерть знаменитости PDF / PPT
- Урок 4 — 10 класс — DUI, Oh My! PDF
- Урок 5 — 10 класс — Объявление государственной службы (PSA) PDF / PPT / MOV
- Урок 6 — 10 класс — Жестокие сигареты PDF / PPT
- Урок 7 — 10 класс — Ролевые цели PDF
- Урок 8 — 10 класс — JUULs Not Cool PDF / PPT
- Урок 9 — 10 класс — Обсуждение родителей / опекунов — Пьянство PDF
- Урок 10 — 10 класс — Влияние ATOD на сообщество PDF / PPT
- Урок 11 — 10 класс — Навыки отказа от ATOD (алкоголь, табак и другие наркотики) PDF / PPT
- Урок 12 — 10 класс — Вы умнее руководителя отдела маркетинга? PDF / PPT
- Урок 13 — 10 класс — Ты дурак для JUUL? PDF
- Урок 14 — 10 класс — Не будь дураком, отложи JUUL PDF
Предотвращение насилия
- Урок 1 — 10 класс — Образовательные ресурсы по предотвращению насилия PDF
- Урок 2 — 10 класс — Все дело в утверждениях «Я» PDF
- Урок 3 — 10 класс — Отношения подростков PDF / PPT
- Урок 4 — 10 класс — Рэп о взаимоотношениях PDF / PPT
- Урок 5 — 10 класс — Отличный улов PDF
- Урок 6 — 10 класс — Реальность отношений PDF
- Урок 7 — 10 класс — Этапы навыков ведения переговоров PDF / PPT
- Урок 8 — 10 класс — Разрешение конфликтов PDF
- Урок 9 — 10 класс — Любовь не должна причинять боль PDF / PPT
- Урок 10 — 10 класс — Что делает отношения здоровыми? PDF / PPT
Учащиеся десятого класса владеют основными двигательными навыками и комбинациями навыков и компетентны в самостоятельно выбранных физических упражнениях, которыми они, вероятно, будут заниматься на протяжении всей жизни, включая занятия на свежем воздухе, занятия фитнесом, танцы и ритмические занятия, водные виды спорта, отдельные индивидуальные упражнения и сетку. / настенные и целевые игры.Они понимают и применяют концепции и принципы механики и анатомии в отношении движений человека и применяют концепции и принципы метаболической реакции организма на краткосрочную и долгосрочную физическую активность. Студенты — хорошие лидеры и хорошие последователи; они уважают других, предвидят и избегают небезопасных ситуаций, связанных с физической активностью. Они развивают способность понимать и предвидеть, как интересы и способности физической активности меняются в течение жизни. Учащиеся демонстрируют способность к физической активности на протяжении всей жизни, а также планируют, внедряют, самооценивают и изменяют личный план физической подготовки.Студенты готовы вести физически активный образ жизни.
Ресурсы по физическому воспитанию для всех 10-х классов
Развитие моторных навыков
- Блок 1 — 10 класс — Адаптированные удары ракетками pdf
- Блок 2 — 10 класс — Игры фрисби pdf
Анатомическая основа движения
Фитнес-планирование
- Блок 1 — 10 класс — Оценка фитнес-планирования 1 pdf
- Блок 2 — 10 класс — Оценка фитнес-планирования 2 pdf
- Блок 3 — 10 класс — Оценка фитнес-планирования 3 pdf
- Блок 4 — 10 класс — Оценка фитнес-планирования 4 pdf
- Блок 5 — 10 класс — Оценка фитнес-планирования 5 pdf
Каковы общие строительные допуски для легких деревянных конструкций?
Каковы общие строительные допуски для легких деревянных конструкций?
Строительные допуски — это средство управления такими элементами, как отвес вертикального элемента, размер и / или длина конструктивных элементов, а также общий план и расположение.Независимо от материала, из которого изготовлены конструктивные элементы здания, допуски часто контролируются производителями продукции и отделки, чтобы гарантировать эксплуатационные характеристики продукции. Приведенные ниже правила допусков типичны для всех конструкций с легким каркасом и не обязательно относятся только к деревянным каркасам.
Несмотря на то, что в Международном строительном кодексе (IBC) или ссылочных стандартах, таких как Национальная проектная спецификация® (NDS®) для деревянного строительства Американского совета по древесине, отсутствуют специальные требования к допускам на строительство легких каркасных деревянных конструкций, общепринятыми являются следующие руководящие указания.
Для вертикальных плоскостных допусков стен (т. Е. Отвесных поверхностей стен):
- В Руководстве по эксплуатационным характеристикам жилищного строительства предлагается максимальное отклонение от отвеса 3/8 дюйма на 32 дюйма по вертикали, в то время как в Справочнике строительных допусков говорится, что, возможно, потребуется затянуть его до 1/4 дюйма на 10 ° по окончании. такие как гипсокартон и штукатурка.
- Точно так же, United Applications Guide Specification (UFGS) предлагает допуск 1/4 дюйма на 8 дюймов для вертикальности стоек, когда используются такие отделки, как стеновая плита, штукатурка или керамическая плитка, уложенная в слой раствора.Для отделки, такой как керамика, закрепленная в сухом растворе, латексно-портландцементном растворе или органическом клее, UFGS предлагает допуск отвеса для стен 1/8 дюйма на 8 дюймов.
- Стандарты для жилищного и легкого коммерческого строительства предлагают предел допуска 1/4 дюйма на 32 дюйма по вертикали.
Для горизонтальных плоских стенок допуски (т. Е. Прямые поверхности стен):
- Справочник строительных допусков предлагает максимальное смещение 1/8 дюйма для соседних элементов каркаса, когда гипсокартонная плита прикрепляется непосредственно к ним.
- В Руководстве по эксплуатационным характеристикам жилищного строительства предлагается предел допуска 1/2 дюйма на 32 дюйма по горизонтали или 1/2 дюйма на 8 дюймов по вертикали для стен с изгибом.
- UFGS предлагает предел 1/4 дюйма на 8 дюймов для лицевой стороны элементов каркаса от истинной плоскости, когда используются такие отделки, как стеновая плита, штукатурка или керамика, укладываемая в раствор.
- В дополнение к этим документам многие производители отделочных материалов и / или отрасли промышленности устанавливают ограничения допусков для элементов каркаса, к которым они прикрепляются.Например, Gypsum Association заявляет в GA 216 и ASTM C-840, что при установке гипсокартона «поверхности крепления любого элемента каркаса не должны отклоняться более чем на 1/8 дюйма от плоскости поверхностей смежных элементов каркаса».
Для допусков положения или компоновки (т. Е. Расположения элементов):
- Справочник строительных допусков предлагает +/- 1/4 ″ для легких каркасных конструкций и +/- 1/8 ″ для тяжелых деревянных конструкций.
- Повторяя это, UFGS предлагает предел допуска 1/4 дюйма для расположения стен и перегородок относительно предполагаемого местоположения.UFGS также рекомендует допуск 1/4 дюйма на 8 дюймов для прямолинейности пластин и направляющих и истинной плоскости каркаса, когда используются такие отделки, как стеновая плита, штукатурка или керамика в растворе.
Несколько ссылок на допуски на строительство:
- Руководство по эффективности жилищного строительства , Национальная ассоциация жилищных строителей
- Спецификация Руководства по объединенным объектам, раздел 06 10 00: Грубые столярные изделия 3.4
- Справочник строительных допусков, Дэвид Кент Балласт.
- Стандарты жилищного строительства, Национальная ассоциация лицензионных агентств государственных подрядчиков
- Стандарты жилищного и легкого коммерческого строительства, RS означает
Хотя иногда их путают с допусками конструкции, особенно в постоянно нагруженных элементах, таких как балки перекрытия или балки крыши, базовые требования для проверки работоспособности, такие как отклонение элемента , указаны в коде .