
У животного изображенного на рисунке отсутствует: Ваш браузер устарел
Чего нет(отсутствуе(ю)т) у данного животного,которое изображено на рисунке ? А)
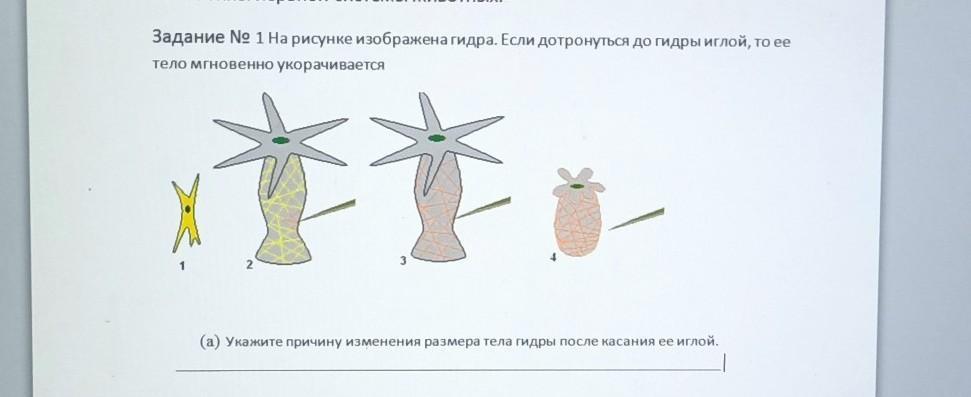
ЕТАПИ Користуючись підручником, заповніть таблицю. Етапи Етапи енергетичного обміну Другий — безкисневий, або анаеробне дихання Третій — кисневий, або … аеробне дихання Перший підготовчий 1. Вихідні речовини дихання 2. Умови реакцій 3. Кінцеві речовини розщеплення 4. Енергетичні перетворення 5. Сумарне рівняння
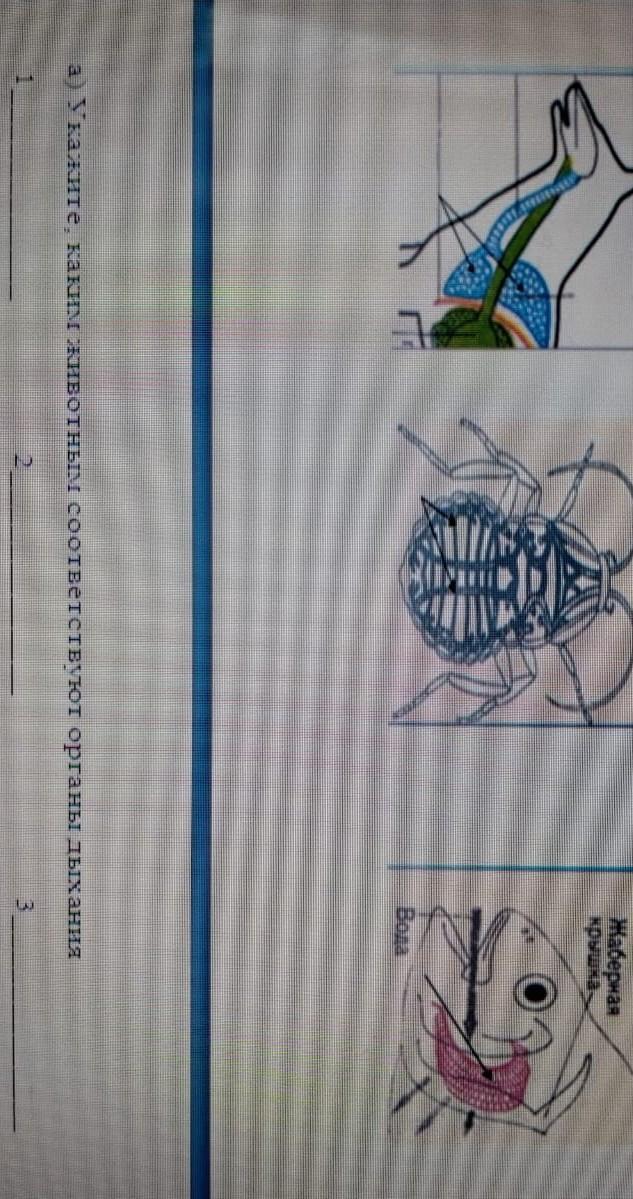
Помогите пожалуйста ДІЯЛЬНІСТЬ Самостійна робота з ілюстраціями Розгляньте ілюстрації та зіставте назви тварин із їх зобра- женнями: 1 – річковий рак … вузькопалий, 2 — блакитний кубинський рак, 3 — пальмовий злодій, 4 — камчатський краб, 5 – лангуст, 6 – мор- ський жолудь, 7 – омар, 8 — коропоїд. — В A Б Є Ж E
1. Порівняйте будову і функції рослинної та тваринної клітин. 2. Як будова клітинної мембрани відповідає її функціям? 3. Запропонуйте і обгрунтуйте гі … потезу, яка пояснює, як в клітині еукаріот з’явились пластиди і мітохондрії?
Де відбувається кисневе розщеплення біомолекул під час окисно-відновних реакцій?
вред наносяжий человеку нематоды
Задание 2.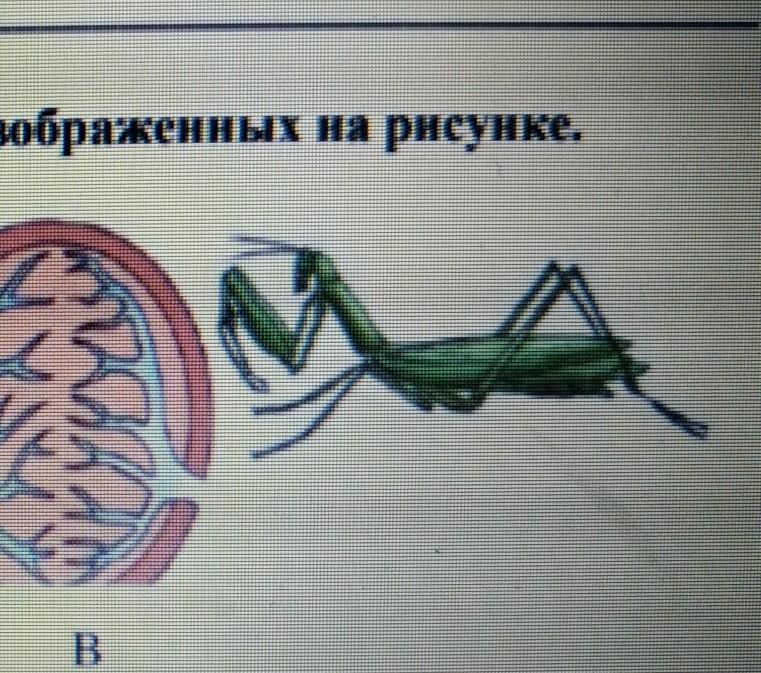 Определите основные функции каждой зоны корня. Дескрипторы: 1. Правильно определяет основную функции каждой зоны корня. Зоны корня функции
Определите основные функции каждой зоны корня. Дескрипторы: 1. Правильно определяет основную функции каждой зоны корня. Зоны корня функции
Тест по теме «Споровые растения»
1.Среди папоротникообразных преобладают жизненные формы:
а) древовидные,
б) травянистые многолетние,
в) травянистые о
…
днолетние.
2. Увеличению числа особей папоротника способствует их:
а) обитание под кронами деревьев,
б) размножение,
в) способность жить в слабо освещенных местах.
3. Что необходимо для размножения папоротникообразных:
а) свет, б) влага, в) тепло.
4. Папоротники имеют органы:
а) корень, клубень,
б) корень, стебель, листья,
в) листья, побеги.
5. Хвощ полевой – травянистое растение:
а) однолетнее, б) двухлетнее, в) многолетнее.
6. Какие из этих утверждений верны?
а) Папоротникообразные – исключительно небольшие травянистые растения
б) Папоротникообразные – высшие растения
в) Папоротникообразные – низшие растения
7. Что называют спорофитом?
а) Поколение, образующие споры
б) Поколение, развивающееся из споры
8.
Среди какого класса моллюсков преобладают организмы-фильтраторы? ДвустворчатыеМногостворчатыеГоловоногиеБрюхоногие
Помогите очень прошу
Помогите 1-5 вопросы зелёная рамочка .
Тренировочный вариант 1 ВПР 2021 по биологии 8 класс
Тренировочный вариант 1 ВПР 2021 по биологии 8 класс. Пробные варианты по биологии 8 класс ВПР 2021. ВСЕРОССИЙСКАЯ ПРОВЕРОЧНАЯ РАБОТА Биология 8 класс.
1.
Как называется раздел биологии, объектом изучения которого являются изображённые на фотографии объекты?
1) орнитология
2) териология
3) герпетология
4) зоология
Ответ.
Объясните свой ответ с использованием знаний об объектах изучения биологии.
Ответ.
2.
Рассмотрите изображённое на фотографии животное и опишите его, выполнив задания.
2.1. Укажите тип симметрии животного.
Ответ. _______________________________
2.2. Укажите среду обитания животного.
Ответ. _______________________________
2.3. Установите последовательность расположения систематических групп изображённого животного, начиная с самой крупной. Используйте слова и словосочетания из предложенного перечня. Запишите в таблицу цифры, под которыми они указаны.
Запишите в таблицу цифры, под которыми они указаны.
Список слов и словосочетаний:
1) Животные
2) Членистоногие
4) Чешуекрылые
5) Берёзовый шелкопряд
Ответ. Царство=?, Тип=?, Класс=?, Отряд=?, Вид=?
3.
Известно, что лось – жвачное парнокопытное животное, живёт в лесу и питается растительной пищей. Используя эти сведения, выберите из приведённого ниже списка три утверждения, относящиеся к описанию данных признаков этого животного.
Запишите в ответе цифры, соответствующие выбранным ответам.
1) Длина тела животного достигает 3 м при массе до 570 кг.
2) У животных растительная пища из желудка отрыгивается в рот и вторично пережёвывается.
3) Лоси летом питаются травами, зимой – побегами ив, осины, рябины, сосны.
4) Шерсть животного довольно жёсткая, имеет буровато-чёрный окрас.
5) Развиты третий и четвёртый пальцы, между которыми проходит ось ноги, что способствует быстрому передвижению.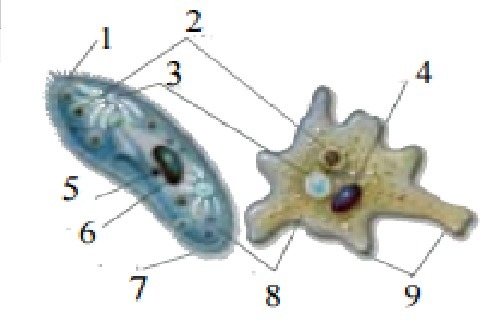
6) У лосей прекрасно развиты слух и обоняние, но очень плохое зрение.
Ответ: ?, ?, ?
4.
4.1. Определите тип питания организмов, приведённых в перечне. Запишите цифры, под которыми указаны организмы, в соответствующую ячейку таблицы.
Список организмов:
1) каштан конский
2) возбудитель чумы
3) малина обыкновенная
4) подберёзовик шахматный
5) мятлик луговой
6) шакал азиатский
Ответ: Автотрофный тип питания=? Гетеротрофный тип питания=?
4.2. Какой тип питания характерен для капустной белянки, изображённой на рисунке 1?
Ответ. ____________________________________
Обоснуйте свой ответ.
5.
Рассмотрите рисунок 2, на котором представлен цикл развития кошачьей двуустки, и ответьте на
вопросы.
5.1. Какой цифрой обозначен на рисунке второй промежуточный хозяин кошачьей двуустки?
Ответ.
5.2. Можно ли считать кошку промежуточным хозяином кошачьей двуустки? Ответ обоснуйте.
Ответ.
6.
В приведённой ниже таблице между позициями первого и второго столбцов имеется взаимосвязь.
Животное: ящерица, утка
Часть тела: чешуя, …
6.1. Какое понятие следует вписать на место пропуска в этой таблице?
1) трахея
2) лёгкое
3) копчиковая железа
4) воздушный мешок
Ответ:
6.2. Какую функцию выполняет чешуя у рыб?
Ответ.
7.
К какому классу относят животных, строение головного мозга которых показано на рисунке 3?
1) Земноводные
2) Пресмыкающиеся
3) Птицы
4) Млекопитающие
Ответ:
8.
8.1. Установите соответствие между характеристиками и типами беспозвоночных: к каждой позиции, данной в первом столбце, подберите соответствующую позицию из второго столбца.
ХАРАКТЕРИСТИКИ ТИПЫ
А) полость тела отсутствует
Б) мантия
В) нервная система лестничного типа
Г) наличие кровеносной системы
Д) сквозной кишечник
Е) наличие органов дыхания
БЕСПОЗВОНОЧНЫХ
1) Моллюски
2) Плоские черви
Ответ: А=?, Б=?, В=?, Г=?, Д=?, Е=?
8. 2. Приведите по три примера животных, относящихся к указанным типам. Запишите их
2. Приведите по три примера животных, относящихся к указанным типам. Запишите их
названия в таблицу.
Моллюски=?
Плоские черви=?
9.
Вставьте в текст пропущенные слова из предложенного перечня, используя для этого их цифровые обозначения. Впишите номера выбранных слов на места пропусков в тексте.
Особенности моллюсков
Моллюски – животные с ______(А) не разделённым на членики телом, имеющим наружную(-ный) ______(Б). У большинства моллюсков различают голову, ______(В) и мускульную ногу. К этим животным относят, например, ______(Г).
1) туловище
2) мягкий
3) грудь
4) хитиновый скелет
5) японский краб
6) твёрдый
7) раковина
8) виноградная улитка
Запишите в таблицу выбранные цифры под соответствующими буквами.
Ответ: А=?, Б=?, В=?, Г=?,
10.
10.1. Если у животного имеется глаз, изображённый на рисунке 4, то для этого животного, вероятнее всего, будут характерны
1) наличие крыльев
2) прямое развитие
3) одна пара усиков
4) замкнутая кровеносная система
5) лёгкие
Запишите в ответе цифры, под которыми указаны выбранные характеристики.
Ответ. _____________________________________
10.2. В описании животных зоологи часто употребляют термин «лёгкие». Что понимают под этим термином?
Ответ:
11.
Верны ли следующие суждения о земноводных?
Б. Передний мозг земноводных разделён на полушария.
1) верно только А
2) верно только Б
3) оба суждения верны
4) оба суждения неверны
Ответ:
12.
Изучите данные приведённой ниже таблицы и ответьте на вопросы.
Таблица
Состояние некоторых жизненных процессов у животных в период активности и во время спячки
У какого животного разница между пульсом в активном состоянии и во время спячки имеет минимальные значения?
Ответ. __________________________________________________________________________
К какому классу позвоночных относят животных, перечисленных в таблице?
Ответ. __________________________________________________________________________
У какого животного в период спячки наблюдается наибольшая потеря веса?
Ответ.
13.
Рассмотрите фотографию собаки породы немецкий дратхаар и выполните задания.
13.1. Выберите характеристики, соответствующие внешнему строению собаки, по следующему плану:
окрас шерсти, форма ушей, форма хвоста.
Запишите в таблицу выбранные цифры под соответствующими буквами.
Ответ. А=?, Б=?, В=?
13.2. Юлия решила выяснить, соответствует ли изображённая на фотографии собака породы немецкий дратхаар стандартам для использования её в целях чистопородного размножения в клубе собаководства. Помогите Юлии решить эту задачу, воспользовавшись фрагментом описания стандарта данной породы.
Стандарт породы немецкий дратхаар (фрагмент)
1. Окрас: однотонный (чёрный с сединой) или пятнистый.
2. Уши: посажены высоко, полустоячие.
3. Форма хвоста: прутом.
Сделайте заключение о соответствии изображённой на фотографии собаки указанным стандартам породы. Оцените возможность использования собаки этой породы для чистопородного размножения в клубе собаководства.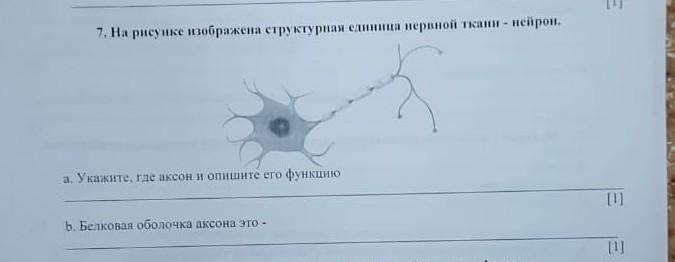
Ответ.
ОТВЕТЫ
1.
Правильный ответ должен содержать следующие элементы:
1) указание названия раздела биологии: 3 ИЛИ герпетология;
2) объяснение, например: герпетология – наука, изучающая земноводных (амфибий) и пресмыкающихся (рептилий).
ИЛИ На фотографии изображены земноводные (амфибии) и пресмыкающиеся (рептилии), а наука, которая изучает этих животных, называется герпетологией
2.
2.1 двусторонне-симметричное животное
2.2 наземно-воздушная
2.3 12345
2.4 В правильном ответе должно быть указано значение, например: берёзовый шелкопряд является вредителем леса.
ИЛИ Личинки берёзового шелкопряда объедают листья древесных лиственных пород
3.
235
4.
4.1 автотрофный тип питания – 135
гетеротрофный тип питания – 246
(в любой последовательности)
4.2 Правильный ответ должен содержать следующие элементы:
1) тип питания животного: гетеротрофный;
2) обоснование, например: у белянки отсутствуют хлоропласты, поэтому у неё гетеротрофный тип питания.
ИЛИ Капустная белянка питается готовыми органическими веществами, то есть имеет гетеротрофный тип питания.
ИЛИ Отсутствует фотосинтез
5.
5.1. 4
5.2. Правильный ответ должен содержать следующие элементы:
1) ответ на вопрос: нет, нельзя;
2) обоснование, например: в организме кошки обитает половозрелая особь, откладывающая яйца, а развития личиночных стадий не происходит.
Элементы ответа могут быть приведены в иных, близких по смыслу формулировках
6.
6.1. 3
6.2. Правильный ответ должен содержать указание функции, например: чешуя придаёт рыбам обтекаемую форму.
ИЛИ Чешуя выполняет защитную функцию.
ИЛИ Чешуя снижает сопротивление при движении в воде
7.
1
8.
8.1. 212111
8.2. Правильный ответ должен содержать по три примера беспозвоночных животных, относящихся к типам Моллюски и Плоские черви
9.
2718
10.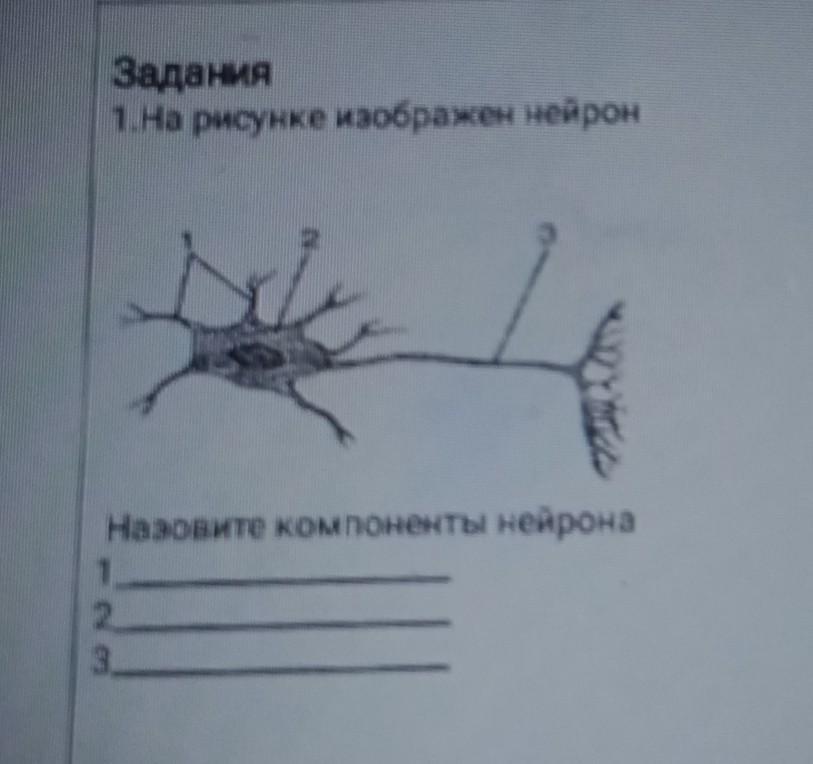
10.1. 13
(в любой последовательности)
10.2. Правильный ответ должен содержать объяснение термина, например: лёгкие – органы, в которых происходит газообмен;
ИЛИ органы, в которых кровь насыщается кислородом и отдаёт углекислый газ.
Объяснение термина может быть приведено в иной, близкой по смыслу формулировке
11.
2
12.
Правильный ответ должен содержать три элемента:
1) ёж обыкновенный;
2) Млекопитающие;
3) жёлтый суслик.
Элементы ответа могут быть приведены в иных, близких по смыслу формулировках
13.
13.1. 224
13.2. Правильный ответ должен содержать следующие элементы:
1) заключение, например: собака, изображённая на фотографии, соответствует стандартам породы по окрасу, форме хвоста и по форме ушей;
2) оценка, например: данная собака подходит для чистопородного размножения в клубе собаководства
Ответ 23 Три среды обитания
РАЗМЕЩЕНИЕ163. Дайте определение.
Дайте определение.
Среда обитания — это
- Ответ: Окружающая среда, в которой живут определенные организмы.
164. Подпишите на схеме три среды обитания организмов. Приведите примеры населяющих их животных.
165. С помощью учебника закончите заполнение таблицы
-
Ответ:
Условия жизни Наземно-воздушная среда Водная среда Почвенная среда Кислород Достаточно Не всегда достаточно Не всегда достаточно Вода Не всегда достаточно Достаточно Имеется Изменение температуры Значительные Незначительные Незначительные Свет Достаточно Чем глубже, тем меньше Отсутствует
166. Рассмотрите рисунок. Какие из изображенных животных обитают в почвенной среде? Подчеркните их синим карандашом.
Рассмотрите рисунок. Какие из изображенных животных обитают в почвенной среде? Подчеркните их синим карандашом.
- Ответ: Крот и змея.
167. В каких средах, по вашему мнению, обитают насекомые, ножки которых изображены на рисунке.Укажите стрелками. Объясните (устно), почему вы так думаете.
-
Ответ:
Можете ли вы назвать этих насекомых? Напишите названия под соответствующими номерами.
168. Рассмотрите рисунок. В чем сходство изображенных животных? Объясните, почему оно возникло.
- Ответ: Это хищные животные. Акула ведет полностью водный образ жизни, остальные — полуводные и являются млекопитающими. Их тело обтекаемо, что максимально приспособило их к водному образу жизни.
169. Укажите цифрами, какие организмы обитают в этих средах.
-
Ответ:
1) Почвенная среда
2) Наземно-воздушная среда
3) Водная среда
170. Приведите примеры, доказывающие что особенности строения тел животных связаны со средой их обитания и образом жизни.
- Ответ: У многих водных животных продолговатая форма тела. У кротов редуцированы глаза и лапы приспособлены для подземной жизни.
171. Вставьте пропущенные буквы.
- Ответ: ПочвЕнНая среда, слЕпыш, кАлиБри, гЕпарД.
Составьте 2 — 3 предложения с этими словами.
- Ответ: Калибри и гепард обитают в наземно-воздушном пространстве. А в почвенной среде обитают крот и слепыш.

систематических обзоров исследований на животных; Недостающее звено в трансляционных исследованиях?
Аннотация
Фон
Методологическое качество исследований на животных является важным фактором, затрудняющим перевод результатов исследований на животных в клинические условия. Систематические обзоры исследований на животных могут предоставить подходящий метод для оценки и, таким образом, улучшения их методологического качества.
Цели
Целями этого исследования были: 1) оценка риска систематических обзоров на животных и 2) изучение внутренней валидности первичных исследований на животных, включенных в эти систематические обзоры.
Источники данных
Мы систематически искали в Pubmed и Embase SR доклинических исследований на животных, опубликованных между 2005 и 2012 годами.
Результаты
В общей сложности 91 систематический обзор соответствовал нашим критериям включения. Риск систематической ошибки оценивался в 48 (52,7%) из 91 систематического обзора. Тридцать три (36,3%) СР предоставили достаточно информации для оценки внутренней валидности включенных исследований. Из оцененных первичных исследований 24,6% были рандомизированы, 14.6% заявили, что ослепили исследователя / опекуна, 23,9% ослепили оценку результатов и 23,1% сообщили о прекращении обучения.
Риск систематической ошибки оценивался в 48 (52,7%) из 91 систематического обзора. Тридцать три (36,3%) СР предоставили достаточно информации для оценки внутренней валидности включенных исследований. Из оцененных первичных исследований 24,6% были рандомизированы, 14.6% заявили, что ослепили исследователя / опекуна, 23,9% ослепили оценку результатов и 23,1% сообщили о прекращении обучения.
Выводы
Для улучшения перевода данных на животных в клиническую практику целесообразны систематические обзоры исследований на животных, но необходимо повысить внутреннюю валидность первичных исследований на животных. Кроме того, следует оценивать риск систематической ошибки с помощью систематических обзоров исследований на животных, чтобы получить представление о надежности имеющихся доказательств.
Образец цитирования: van Luijk J, Bakker B, Rovers MM, Ritskes-Hoitinga M, de Vries RBM, Leenaars M (2014) Систематические обзоры исследований на животных; Недостающее звено в трансляционных исследованиях? PLoS ONE 9 (3):
e89981. https://doi.org/10.1371/journal.pone.0089981
https://doi.org/10.1371/journal.pone.0089981
Редактор: Йенс Миннеруп, Мюнстерский университет, Германия
Поступила: 25 ноября 2013 г .; Принята к печати: 25 января 2014 г .; Опубликован: 26 марта 2014 г.
Авторские права: © 2014 van Luijk et al.Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Финансирование: У этих авторов нет поддержки или финансирования, чтобы сообщить о них.
Конкурирующие интересы: Авторы заявили об отсутствии конкурирующих интересов.
Введение
Большинство экспериментов на животных проводится в рамках доклинических исследований, т.е.грамм. проверить безопасность и эффективность новых методов лечения для улучшения здравоохранения. Однако перевод данных о животных в ситуацию с человеком оказался очень сложной задачей. На этот перевод влияют различные факторы, такие как биологические различия между видами, внутренняя валидность, различия в дизайне экспериментов между исследованиями на животных и клиническими испытаниями, недостаточная отчетность и систематическая ошибка публикации [1]. Систематические обзоры (SR) исследований на животных могут снизить некоторые проблемы при переводе данных на животных в клинические испытания, например, путем явной оценки внутренней валидности.СР пытаются идентифицировать, оценивать и синтезировать все эмпирические данные, которые соответствуют заранее определенным критериям приемлемости для ответа на заданный вопрос исследования. SR исследований на животных все еще довольно редки, но их количество, похоже, немного увеличивается [2] — [4]. Однако мало что известно о том, в какой степени доступные SR включают риск систематической оценки, в которой внутренняя валидность включенных первичных исследований на животных.
Однако перевод данных о животных в ситуацию с человеком оказался очень сложной задачей. На этот перевод влияют различные факторы, такие как биологические различия между видами, внутренняя валидность, различия в дизайне экспериментов между исследованиями на животных и клиническими испытаниями, недостаточная отчетность и систематическая ошибка публикации [1]. Систематические обзоры (SR) исследований на животных могут снизить некоторые проблемы при переводе данных на животных в клинические испытания, например, путем явной оценки внутренней валидности.СР пытаются идентифицировать, оценивать и синтезировать все эмпирические данные, которые соответствуют заранее определенным критериям приемлемости для ответа на заданный вопрос исследования. SR исследований на животных все еще довольно редки, но их количество, похоже, немного увеличивается [2] — [4]. Однако мало что известно о том, в какой степени доступные SR включают риск систематической оценки, в которой внутренняя валидность включенных первичных исследований на животных. Поэтому мы провели систематический обзор оценки риска систематической ошибки в СО исследований на животных.Впоследствии мы изучили внутреннюю валидность отдельных исследований, включенных в эти SR.
Поэтому мы провели систематический обзор оценки риска систематической ошибки в СО исследований на животных.Впоследствии мы изучили внутреннюю валидность отдельных исследований, включенных в эти SR.
Материалы и методы
Стратегия поиска
Чтобы найти все СО исследований на животных, опубликованные в период с 2005 по 2012 год, 28 января 2013 года была проведена следующая поисковая стратегия. Для выявления исследований на животных был проведен поиск в базах данных MEDLINE (платформа PubMed) и EMBASE (платформа OvidSP) с использованием «Животные». ‘фильтр для PubMed от Hooijmans et al., [5] и фильтр для EMBASE от De Vries et al., [6], [7] соответственно. Поскольку нас интересовали СР, мы использовали клинический запрос СР из PubMed, который мы адаптировали для Embase (см. S1).
Выбор исследования
Для целей данного исследования обзор был классифицирован как SR, когда были представлены по крайней мере все следующие элементы: 1) термин «систематический обзор» 2) поиск в базе данных (-ах) и 3) поисковые запросы.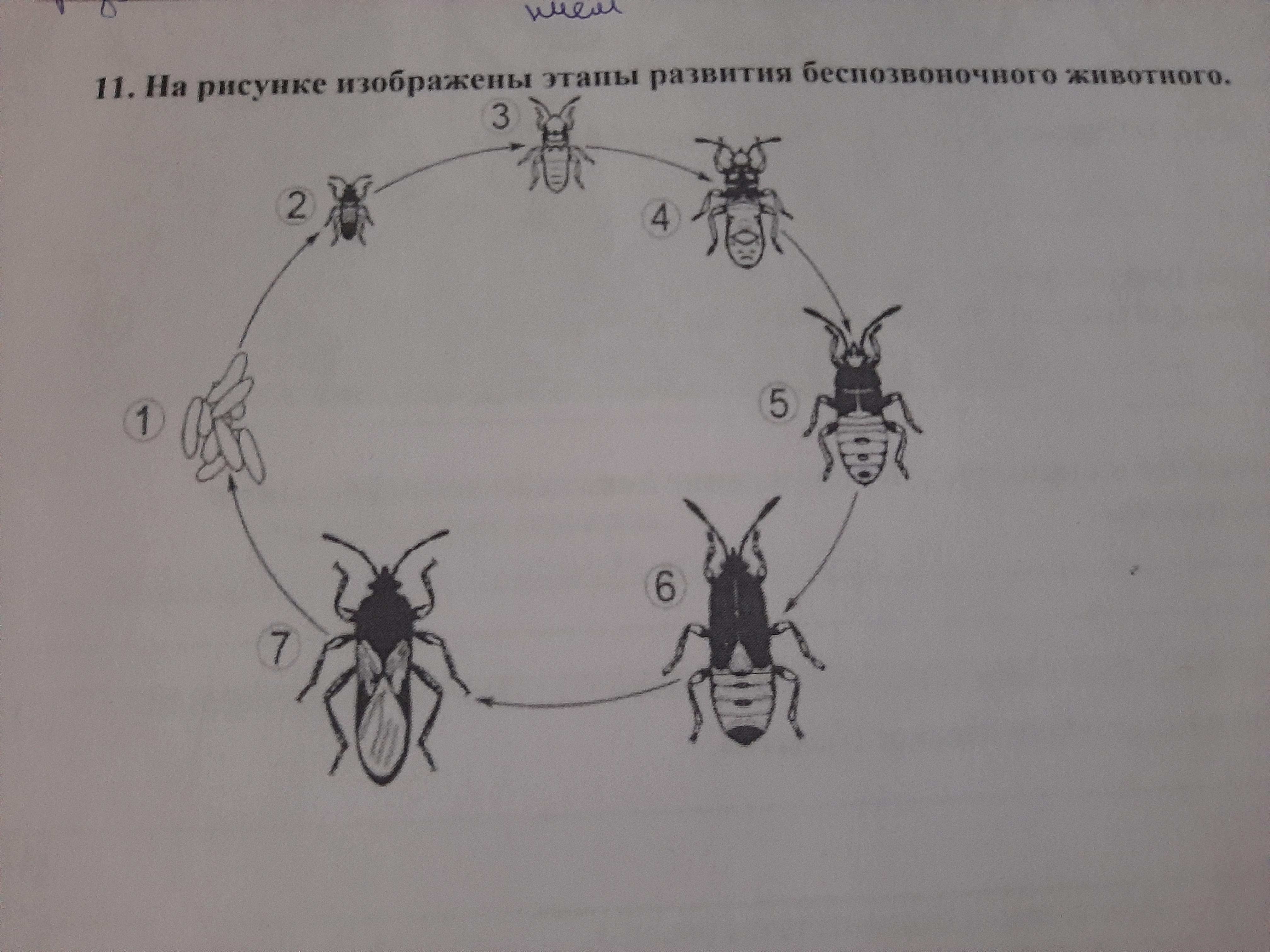 Отбор производился двумя независимыми наблюдателями, и разногласия разрешались путем обсуждения (JvL, BB, ML).Были включены только СР, нацеленные на информирование о здоровье человека путем обзора медицинских препаратов, таких как витаминные добавки или лечение стволовыми клетками. Медицинские устройства, такие как протезы и каркасы, и другие виды вмешательства, такие как кислород или тепло, были исключены. Мы также исключили SR, которые были написаны не на английском языке или которые не могли быть получены в полном тексте. Когда дополнительные данные были доступны в Интернете, они были получены.
Отбор производился двумя независимыми наблюдателями, и разногласия разрешались путем обсуждения (JvL, BB, ML).Были включены только СР, нацеленные на информирование о здоровье человека путем обзора медицинских препаратов, таких как витаминные добавки или лечение стволовыми клетками. Медицинские устройства, такие как протезы и каркасы, и другие виды вмешательства, такие как кислород или тепло, были исключены. Мы также исключили SR, которые были написаны не на английском языке или которые не могли быть получены в полном тексте. Когда дополнительные данные были доступны в Интернете, они были получены.
Процедура подсчета баллов SR
Данные как о характеристиках, так и о методах, используемых для оценки риска систематической ошибки в СР, были извлечены как минимум двумя независимыми рецензентами (JvL, BB, ML).В СР, где в СР были включены исследования как на животных, так и на людях, оценивались только данные на животных. Разногласия между рецензентами разрешались путем обсуждения, и при необходимости проводились консультации с третьим рецензентом.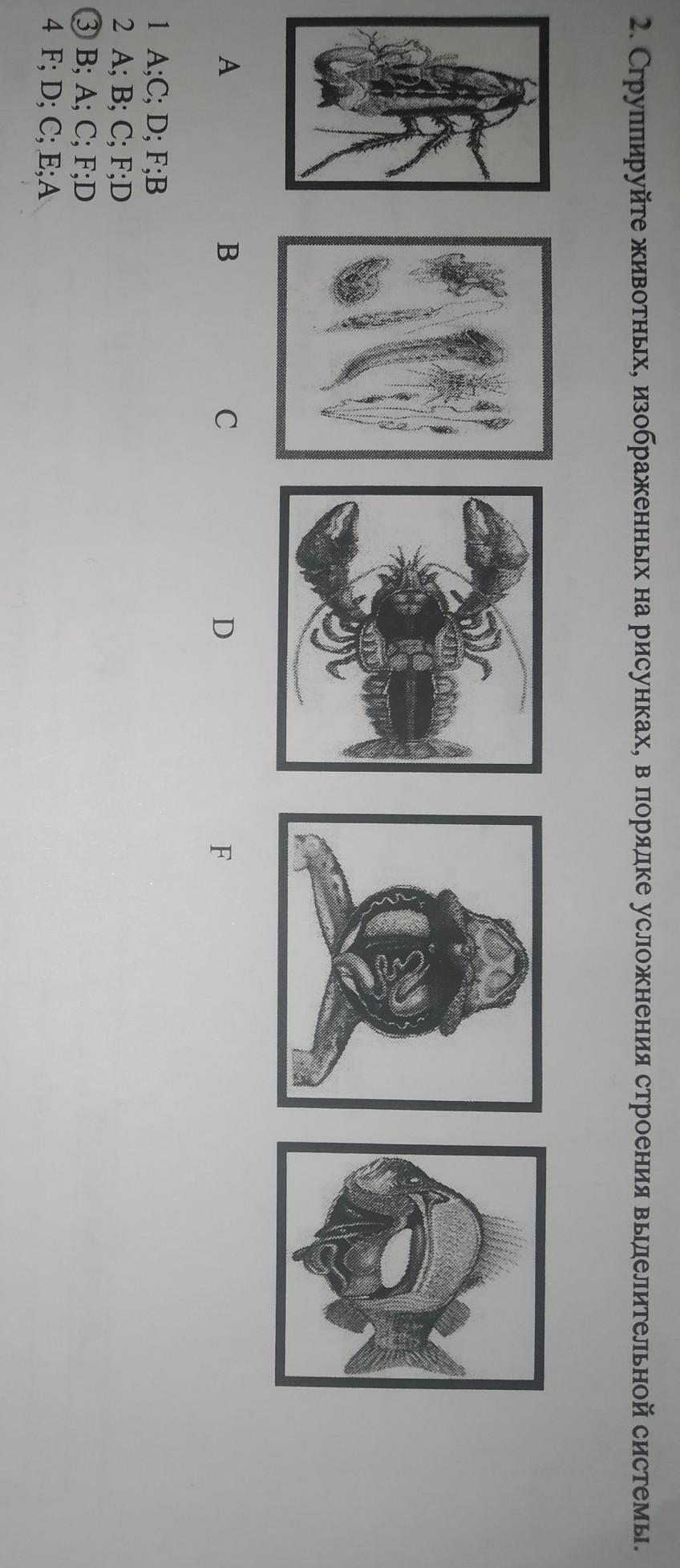
Оценка риска предвзятости.
Методология оценки качества различается в разных СО исследований на животных [8]. В рамках данного исследования мы сосредоточились на внутренней валидности первичных исследований. Поэтому мы определили оценку качества как оценку риска систематической ошибки.Чтобы соответствовать этому определению, оценка должна была включать по крайней мере один из следующих пунктов внутренней валидности: 1) рандомизированный дизайн исследования (систематическая ошибка отбора), 2) ослепление исследователя / лица, осуществляющего уход (систематическая ошибка производительности), 3) ослепление оценки результатов (обнаружение предвзятость) и 4) упоминание выбывших (предвзятость отсева).
SR характеристики.
Была извлечена дополнительная информация о характеристиках SR: 1) способ учета риска систематической ошибки в SR (например,грамм. проведение анализа подгрупп на основе качества, исключение исследований на основе качества или общего комментария / утверждения, относящегося к качеству исследования), 2) уровень детализации отчета о внутренней валидности (например, оценка по каждому пункту или сводка по качеству по каждому исследованию) и 3) область исследований СР.
Первичные исследования извлечения данных
SR, которые предоставили подробную информацию о необходимых элементах внутренней валидности, были использованы для оценки внутренней валидности включенных отдельных исследований.По SR данные были извлечены по: общему количеству включенных исследований и количеству исследований по каждому пункту (рандомизированный дизайн исследования, слепой исследователь / лицо, осуществляющее уход, слепая оценка результатов и выбывание).
Результаты
Поиск литературы и выбор SR
Мы определили 592 потенциально подходящие статьи, из которых 91 СП соответствовал нашим критериям включения. На рисунке 1 показано количество исследований, выявленных на каждом этапе процесса отбора. Полный список из 91 ссылки можно найти в другом месте (см. Таблицу S1 в файле S2).
Характеристики включенных систематических обзоров
Количество опубликованных систематических обзоров исследований на животных увеличилось за последние годы с 6 в 2005 году и 2, 6 и 12 в 2006, 2008 и 2010 годах до 32 в 2012 году, соответственно (Рисунок 2).
91 SR, включенный в этот обзор, охватывает широкий круг исследовательских тем. Большинство обзоров (n = 38; 41,8%) касаются неврологической темы, из которых 20 обзоров (22,0%) относятся к инсульту. Вторая по величине группа — эндокринология (n = 11; 12.1%). Другие темы включали сердечно-сосудистые заболевания, ортопедию, инфекционные заболевания, онкологию, фармакотоксикологию, стоматологию и гастроэнтерологию. Полный список тем и количество SR по каждой теме также можно найти в другом месте (см. Таблицу S1 в файле S2).
Оценка риска систематической ошибки
Риск систематической оценки систематических обзоров.
Почти половина СР (n = 43; 47,3%) не оценили ни один из пунктов риска предвзятости (диаграмма 3). В 48 обзорах (52,7%) были оценены один или несколько заранее определенных пунктов риска предвзятости.Тридцать три (36,3%) обзора также предоставили подробную информацию о результатах этой оценки по каждому отдельному исследованию.
Рисунок 3. Процент СР на количество набранных элементов внутренней валидности.
Процент СР на количество набранных элементов внутренней валидности.
Ноль пунктов на 47,3%, один пункт — все рандомизация, два пункта рандомизации и один уровень ослепления, три пункта рандомизации, ослепление лица, осуществляющего уход / следователь, и ослепление оценки результатов или рандомизации, один уровень ослепления и исключения) и все четыре позиции на 3,3%.
https://doi.org/10.1371/journal.pone.0089981.g003
На Рисунке 3 показано, что из 91 ПП только 3 (3,3%) оценили все 4 пункта внутренней валидности в своей оценке качества. Двадцать два СР (24,2%) оценили 3 пункта, из которых 17 СР (18,7%) не оценили отсев; остальные 5 (5,5%) не ослепляли воспитателя. Четырнадцать СР (15,4%) оценили два пункта, а именно рандомизацию и ослепление (из них 13 СР оценивали слепоту оценки результатов, в одном СР тип ослепления был неясным).Девять СР (9,9%) оценили только один пункт, который во всех случаях был рандомизированным.
Риск использования предвзятости в СР.

Из 48 СР, которые оценивали риск систематической ошибки включенных отдельных исследований, 45 (93,8%) указали на внутреннюю валидность первичных исследований в разделе результатов, обсуждения или заключения. Это означает, что три СР никоим образом не обсуждали результаты оценки риска систематической ошибки. В большинстве обзоров (n = 42; 87,5%) был сделан общий комментарий о качестве первичных исследований.В 25 СР (52,1%) качество первичного исследования использовалось как фактор в метаанализе (например, анализ подгрупп), а в трех СР качество исследования использовалось как критерий исключения (см. Таблицу S1 в файле S2).
Внутренняя действительность первичных исследований
Тридцать три SR, которые предоставили подробную информацию о риске оценки систематической ошибки, были использованы для оценки внутренней валидности включенных первичных исследований. Эти 33 SR включали в общей сложности 2280 первичных исследований (медиана 18, диапазон: 2–1152 первичных исследования). Большинство этих исследований были посвящены инсульту или другим неврологическим вопросам (см. Таблицы S1 и S2 в файле S2).
Большинство этих исследований были посвящены инсульту или другим неврологическим вопросам (см. Таблицы S1 и S2 в файле S2).
На рис. 4 представлен обзор оценок риска систематической ошибки в отдельных исследованиях на животных по каждому пункту (рандомизация, ослепление лица, осуществляющего уход / исследователь, ослепление оценки результатов и выбывание). Поскольку не во всех обзорах оценивались все четыре оцениваемых нами элемента (см. Рисунок 3), количество первичных исследований варьируется в зависимости от элемента на рисунке 4. Из 2280 включенных первичных исследований 562 (24,6%) были рандомизированы.Ослепление исследователя / опекуна оценивалось в 546 (23,9%) первичных исследованиях, из которых 80 (14,6%) фактически были слепыми. Ослепление оценки результатов проводилось в 2220 (97,4%) первичных исследованиях, из которых 530 (23,9%) действительно были слепыми. Прекращение учебы было оценено только в 78 (3,4%) первичных исследованиях, из которых 18 (23,1%) действительно сообщили о прекращении учебы. Одно исследование оценивало ослепление без указания типа ослепления. Поэтому данные этого исследования не были включены в наши результаты (см. Таблицы S1 и S2 в файле S2).
Одно исследование оценивало ослепление без указания типа ослепления. Поэтому данные этого исследования не были включены в наши результаты (см. Таблицы S1 и S2 в файле S2).
Обсуждение
Наши результаты показывают, что оценка методологического качества с помощью систематических обзоров исследований на животных довольно низкая. Половина из 91 оцененного СР не оценили критически риск систематической ошибки во включенных исследованиях. Кроме того, тридцать три обзора, которые действительно оценивали и сообщали о риске систематической ошибки, показали, что внутренняя валидность большинства индивидуальных исследований на животных также оставляет желать лучшего. Следовательно, существует реальный риск того, что результаты как отдельных исследований, так и последующих SR этих исследований будут предвзятыми.
Наши выводы о низком методологическом качестве СР согласуются с выводами Peters et al., Которые выявили ряд недостатков в проведении и составлении отчетов о СР и мета-анализах исследований на животных. Peters et al. предполагают, что инициативы по совершенствованию проведения и отчетности по первичным исследованиям на животных и СО исследований на животных должны идти рука об руку [2]. Низкая внутренняя валидность исследований на животных ранее была продемонстрирована Kilkenny et al. Из 271 публикации обследованных ими исследований на животных только 13% были рандомизированы, а 14% не учитывали оценку результатов [9].Мы обнаружили немного более высокий процент, а именно 24,6% рандомизации и 23,9% ослепления. Такой более высокий процент можно объяснить двумя факторами. Во-первых, наше исследование содержит относительно большое количество исследований инсульта. В течение последних десятилетий исследователи в области инсульта активно работали над рекомендациями и руководствами для доклинических исследований с целью улучшения эффективного перевода [10]. Во-вторых, в последние годы неуклонно растет общее понимание необходимости улучшения отчетов об исследованиях на животных.
Peters et al. предполагают, что инициативы по совершенствованию проведения и отчетности по первичным исследованиям на животных и СО исследований на животных должны идти рука об руку [2]. Низкая внутренняя валидность исследований на животных ранее была продемонстрирована Kilkenny et al. Из 271 публикации обследованных ими исследований на животных только 13% были рандомизированы, а 14% не учитывали оценку результатов [9].Мы обнаружили немного более высокий процент, а именно 24,6% рандомизации и 23,9% ослепления. Такой более высокий процент можно объяснить двумя факторами. Во-первых, наше исследование содержит относительно большое количество исследований инсульта. В течение последних десятилетий исследователи в области инсульта активно работали над рекомендациями и руководствами для доклинических исследований с целью улучшения эффективного перевода [10]. Во-вторых, в последние годы неуклонно растет общее понимание необходимости улучшения отчетов об исследованиях на животных.
Хотя как методологическое качество СО на животных, так и внутренняя валидность первичных исследований на животных изучались ранее, они изучались отдельно различными исследовательскими группами, а более поздние СО исследований на животных еще не принимались во внимание. Таким образом, главной сильной стороной нашего исследования является то, что, обновив и объединив эти оценки в одном исследовании, мы смогли получить более глубокое представление о текущем состоянии и уровне доступных доклинических данных.
Таким образом, главной сильной стороной нашего исследования является то, что, обновив и объединив эти оценки в одном исследовании, мы смогли получить более глубокое представление о текущем состоянии и уровне доступных доклинических данных.
Также следует обсудить некоторые потенциальные ограничения. Во-первых, мы ограничились одним типом SR, а именно SR исследований лекарственных препаратов на животных, что может помешать обобщению наших результатов на другие SR исследований на животных. Хотя мы исключили СР исследований на животных, которые не имеют прямого отношения к клиническим исследованиям, мы считаем вероятным, что СР последнего типа имеют более низкое методологическое качество, поскольку методология СР и меры по обеспечению внутренней валидности могут быть не так хорошо установлены, как в области, тесно связанные с клиническими исследованиями.Следовательно, наше ограничение могло вызвать переоценку методологического качества SR и внутренней валидности первичных исследований в целом. Во-вторых, нельзя исключить, что небольшая часть СР не оценивала определенные элементы внутренней валидности, потому что экспериментальный план включенных отдельных исследований не допускал риска систематической оценки (например, из-за отсутствия (независимого) контроля группы). В-третьих, некоторые индивидуальные исследования могли быть менее подвержены систематической ошибке, чем оценивали СР, из-за отсутствия (адекватных) , сообщающих о методах рандомизации и / или ослепления, которые они фактически использовали.В-четвертых, мы не исследовали, оценили ли СР адекватность метода рандомизации или ослепления. Неадекватная рандомизация и слепота в исследованиях на животных могут привести к завышению величины эффекта [11], [12] и, таким образом, могут оказаться ложными для других доклинических исследований или клинических испытаний. В принципе, это означает, что даже рандомизированные исследования могут быть предметом систематической ошибки, а именно, когда метод рандомизации не соответствует дизайну исследования.
Во-вторых, нельзя исключить, что небольшая часть СР не оценивала определенные элементы внутренней валидности, потому что экспериментальный план включенных отдельных исследований не допускал риска систематической оценки (например, из-за отсутствия (независимого) контроля группы). В-третьих, некоторые индивидуальные исследования могли быть менее подвержены систематической ошибке, чем оценивали СР, из-за отсутствия (адекватных) , сообщающих о методах рандомизации и / или ослепления, которые они фактически использовали.В-четвертых, мы не исследовали, оценили ли СР адекватность метода рандомизации или ослепления. Неадекватная рандомизация и слепота в исследованиях на животных могут привести к завышению величины эффекта [11], [12] и, таким образом, могут оказаться ложными для других доклинических исследований или клинических испытаний. В принципе, это означает, что даже рандомизированные исследования могут быть предметом систематической ошибки, а именно, когда метод рандомизации не соответствует дизайну исследования. Точно так же некоторые СР оценивали ослепление, но не указали уровень или тип ослепления.Однако до тех пор, пока отчеты об исследованиях на животных остаются скудными, этих ограничений вряд ли можно избежать.
Точно так же некоторые СР оценивали ослепление, но не указали уровень или тип ослепления.Однако до тех пор, пока отчеты об исследованиях на животных остаются скудными, этих ограничений вряд ли можно избежать.
Адекватная внутренняя валидность исследований на животных была описана как один из ключевых факторов для улучшения перевода результатов в исследования на людях [1]. СР могут быть полезным методом оценки и анализа (качества) имеющихся доказательств. Как указывалось ранее, СО по исследованиям на животных могут извлечь выгоду из использования руководящих принципов [2]. В настоящее время не существует стандартной процедуры проведения СО исследований на животных [8], [13].Это могло быть одной из причин, по которой так много СО животных не оценили ни один из элементов риска предвзятости. Здесь можно извлечь ценные уроки из руководств, используемых в клинических исследованиях, таких как утверждения CONSORT и PRISMA. Руководства по планированию, проведению и отчетности о первичных исследованиях на животных уже доступны [14], [15]. Несмотря на то, что рекомендации ARRIVE приняты многими журналами, влияние на стандарты публикации исследований на животных по-прежнему очень минимально. Таким образом, эффективное выполнение этих рекомендаций требует большего внимания [16].Как и образование по этому поводу. Хорошая образовательная стратегия, касающаяся как внутренней валидности исследований на животных, так и методологии SR, может помочь повысить осведомленность о текущем состоянии потенциально предвзятых данных о животных. Авторы, а также рецензенты и редакторы должны знать о потенциальном риске этого предубеждения в исследованиях на животных и о том, как его можно адекватно снизить, чтобы в конечном итоге провести высококачественные исследования с надежными результатами для здравоохранения.
Несмотря на то, что рекомендации ARRIVE приняты многими журналами, влияние на стандарты публикации исследований на животных по-прежнему очень минимально. Таким образом, эффективное выполнение этих рекомендаций требует большего внимания [16].Как и образование по этому поводу. Хорошая образовательная стратегия, касающаяся как внутренней валидности исследований на животных, так и методологии SR, может помочь повысить осведомленность о текущем состоянии потенциально предвзятых данных о животных. Авторы, а также рецензенты и редакторы должны знать о потенциальном риске этого предубеждения в исследованиях на животных и о том, как его можно адекватно снизить, чтобы в конечном итоге провести высококачественные исследования с надежными результатами для здравоохранения.
Выводы
Для улучшения перевода данных на животных в клиническую практику целесообразны систематические обзоры исследований на животных, но необходимо улучшить внутреннюю валидность отдельных исследований на животных. Кроме того, СР исследований на животных должны оценить риск систематической ошибки, чтобы получить представление о надежности имеющихся доказательств.
Кроме того, СР исследований на животных должны оценить риск систематической ошибки, чтобы получить представление о надежности имеющихся доказательств.
Благодарности
Авторы хотели бы поблагодарить Алису Тиллему, Ники ван Веггеля, Яна Виллема Вининка и Йоппе Тра за их работу по оптимизации и реализации стратегии поиска.
Вклад авторов
Задумал и спроектировал эксперименты: JVL BB MR RDV ML. Проведены эксперименты: JVL BB ML. Проанализированы данные: JVL BB ML.Предоставленные реагенты / материалы / инструменты анализа: MRH. Написал статью: JVL BB MR MRH RDV ML.
Ссылки
- 1. Hooijmans CR, Ritskes-Hoitinga M (2013) Прогресс в использовании систематических обзоров исследований на животных для улучшения трансляционных исследований. PLoS Med 10: e1001482.
- 2.
Петерс Дж. Л., Саттон А. Дж., Джонс Д. Р., Раштон Л., Абрамс К. Р. (2006) Систематический обзор систематических обзоров и метаанализов экспериментов на животных с руководящими указаниями по отчетности.
 J. Environ Sci Health B 41: 1245–1258.
J. Environ Sci Health B 41: 1245–1258. - 3. Mignini LE, Khan KS (2006) Методологическое качество систематических обзоров исследований на животных: обзор обзоров фундаментальных исследований. BMC Med Res Methodol 6: 10.
- 4. Кореваар Д.А., Хофт Л., тер Рит Г. (2011) Систематические обзоры и метаанализ доклинических исследований: систематическая ошибка публикации в экспериментах на лабораторных животных. Лаборатория Аним 45: 225–230.
- 5. Hooijmans CR, Tillema A, Leenaars M, Ritskes-Hoitinga M (2010) Повышение эффективности поиска с помощью поискового фильтра для поиска всех исследований экспериментов на животных в PubMed.Lab Anim 44: 170–175.
- 6. de Vries RB, Hooijmans CR, Tillema A, Leenaars M, Ritskes-Hoitinga M (2011) Фильтр поиска для увеличения поиска исследований на животных в Embase. Лаборатория Аним 45: 268–270.
- 7.
de Vries RB, Hooijmans CR, Tillema A, Leenaars M, Ritskes-Hoitinga M (2013) Письмо в редакцию.
 Lab Anim.
Lab Anim. - 8. Krauth D, Woodruff TJ, Bero L (2013) Инструменты для оценки риска систематической ошибки и другие методологические критерии опубликованных исследований на животных: систематический обзор.Environ Health Perspect 121: 985–992.
- 9. Килкенни С., Парсонс Н., Кадышевски Э., Фестинг М.Ф., Катхилл И.С. и др. (2009) Обзор качества экспериментального дизайна, статистического анализа и отчетности об исследованиях с использованием животных. PLoS ONE 4: e7824.
- 10. Академическая индустрия терапии инсульта R (1999) Рекомендации по стандартам, касающимся разработки доклинических нейропротективных и восстанавливающих лекарств. Ход 30: 2752–2758.
- 11. Macleod MR, Fisher M, O’Collins V, Sena ES, Dirnagl U, et al.(2009) Надлежащая лабораторная практика: предотвращение появления систематической ошибки на рабочем месте. Ход 40: e50–52.
- 12.
Bebarta V, Luyten D, Heard K (2003) Исследование на животных в медицине неотложной помощи: влияет ли использование рандомизации и ослепления на результаты? Acad Emerg Med 10: 684–687.

- 13. Henderson VC, Kimmelman J, Fergusson D, Grimshaw JM, Hackam DG (2013) Угрозы валидности при разработке и проведении доклинических исследований эффективности: систематический обзор руководящих принципов для экспериментов на животных in vivo.PLoS Med 10: e1001489.
- 14. Килкенни С., Браун В.Дж., Катхилл И.К., Эмерсон М., Альтман Д.Г. (2010) Улучшение отчетности по бионаучным исследованиям: рекомендации ARRIVE по отчетности об исследованиях на животных. PLoS Biol 8: e1000412.
- 15. Hooijmans CR, Leenaars M, Ritskes-Hoitinga M (2010) Контрольный список публикаций по золотому стандарту для улучшения качества исследований на животных, для полной интеграции трех принципов и для того, чтобы сделать систематические обзоры более осуществимыми. Альтернативная лаборатория Anim 38: 167–182.
- 16.
Бейкер Д., Лидстер К., Соттомайор А., Амор С. (2014) Два года спустя: журналы еще не соблюдают правила ARRIVE по стандартам отчетности для доклинических исследований на животных.
 PLoS Biol 12: e1001756.
PLoS Biol 12: e1001756.
Стволовые клетки планарии определяют личность отсутствующего глотки и запускают целевую регенерацию
Все три рецензента признали потенциальную значимость этой работы, но также разделили те же опасения по поводу размеров выборки, отсутствия биологических реплик и недостаточной информации о количественной оценке клеток.Учитывая интерес к рассматриваемому вопросу, если эти вопросы могут быть решены удовлетворительно, пересмотренный документ можно было бы рассматривать здесь как новое представление. Мы включили обзоры ниже и надеемся, что вы найдете их полезными.
Рецензент № 1:
Эта рукопись Бора и др. исследует, как стволовые клетки планарии реагируют на потерю определенного органа: глотки. Ранее предложенная «слепая к мишеням» модель регенерации планарии (LoCascio et al. 2017) постулировала, что стволовые клетки не реагируют напрямую на отсутствующие ткани, а скорее заменяют отсутствующие типы клеток на основе их нормальной скорости гомеостатического обновления.
Напротив, Bohr et al. предполагают, что планарные стволовые клетки могут ощущать и реагировать на потерю определенных недостающих тканей, используя глотку в качестве примера. Авторы заключают, что планарии могут использовать более одного режима регенерации, в зависимости от регенерируемой цели (глаз или глотка).
Вопрос, исследуемый в этой статье, имеет фундаментальное значение, и обеспечение альтернативной модели, с помощью которой планарные стволовые клетки регенерируют недостающие ткани, должно быть интересно широкому кругу читателей.К сожалению, в нынешнем виде рукопись представляет соблазнительные предварительные результаты, а не надежные экспериментальные наблюдения. В настоящее время рукопись имеет ограниченные размеры образцов и экспериментальные копии, что прискорбно. Поскольку в этой статье делается попытка опровергнуть ранее опубликованную модель, очень важно, чтобы данные были ясными и убедительными. В противном случае эти результаты можно было бы сразу отклонить без соответствующего обсуждения.
Если авторы смогут показать надежность и строгость своих результатов и решить основные проблемы, перечисленные ниже, эта рукопись внесет значительный вклад в наше понимание регенерации планарий.
Мы благодарим рецензента за признание важности вопросов, которые мы здесь рассмотрели. Однако мы хотели бы пояснить, что в наши намерения не входило опровержение ранее опубликованной модели. Эта путаница, вероятно, возникла из-за отсутствия ясности в нашем письме. В текущей версии мы явно тестируем «нецелевую» модель с включением EdU (рис. 1E, F и рис. 1 — добавление к рис. 1) и экспериментами по резекции с боков (рис. 2D). Наши данные показывают, что «нацеленная» модель является основным способом регенерации глотки, но «нецелевая» модель также действует сразу после ранения.Мы добавили в обсуждение целый раздел, чтобы читателям было ясно, что оба механизма, вероятно, работают во время регенерации.
1. На протяжении всей рукописи эксперименты либо не повторялись, либо количество биологических повторов не сообщалось.
В большинстве случаев оказывается, что эксперименты проводились только один раз (за исключением медикаментозного лечения). Количество биологических повторов и размеры образцов должны быть четко указаны, а данные из разных повторов должны быть представлены для рисунков 1D-G, 2B-D, 3C, 3E-F, 4B, 4D-H, 5C-D и 6A-E.
Мы согласны с тем, что рукопись страдала общим отсутствием экспериментальных копий и небольшими размерами выборки, как указано в обзоре выше. Принимая во внимание эти существенные критические замечания, мы повторили каждый эксперимент 2-3 раза, увеличили общий размер выборки и явно отметили различные биологические и технические повторы на графиках, где это возможно. Мы также включили таблицу с подробным описанием биологических и технических копий для каждой соответствующей фигуры.
2.Авторы недостаточно описывают свои методы визуализации и количественной оценки клеток (рисунки 1E, 1G, 2C-D, 3F, 4E-H, 5D, 6B). Размер территории, на которой собирались эти данные, неясен.
Показаны изображения с большим увеличением: это те области, которые были сняты? Если это так, их результаты могут быть искажены из-за выбора небольших областей интереса. В идеале авторы должны количественно определить более одного региона на животное. Кроме того, они не описывают глубину собранных z-стэков или то, как эти стеки были нормализованы / стандартизированы для разных условий.Все их выводы основаны на количественной оценке популяций предшественников в ответ на различные парадигмы ампутации или химическое лечение, поэтому стандарты визуализации и количественной оценки должны быть четко указаны.
Мы согласны с тем, что предыдущей версии рукописи не хватало ясности в отношении изображений и количественной оценки. Теперь мы рассмотрели эти важные критические замечания, четко описав их в подписях к рисункам и в разделе методов получения изображений, количественной оценки и статистического анализа (начиная со строки 759).Теперь мы явно указываем следующие стандартизированные параметры для данного эксперимента: 1) анализируемая область внутри животного, 2) измеренная площадь xy, 3) толщина (z-сечения) для данного эксперимента и 4) z-сечение, где количественная оценка началась.
3. Повсюду использовались неподходящие статистические тесты. Использование нескольких t-критериев увеличивает вероятность ошибки типа I и особенно проблематично, когда было выполнено до 7 сравнений! Авторам следует использовать односторонний дисперсионный анализ с множественными поправками для сравнения для всех экспериментов с более чем двумя группами.
Как было предложено, все рисунки, на которых сделаны множественные сравнения, теперь используют односторонний дисперсионный анализ для определения статистической значимости.
4. Рисунки 1D-E показывают, что после ампутации глотки, но не после ампутации головы, количество предшественников FoxA + piwi + глотки увеличивается. Эти данные страдают от проблем количественной оценки, отмеченных выше: способ количественной оценки данных описан недостаточно, были взяты только 3 точки данных (по одной на животное), эксперимент, по-видимому, проводился только один раз, и использовался неправильный статистический тест.
.Вместо того, чтобы сообщать количество подсчитанных клеток FoxA + piwi-1 +, авторы должны количественно определить общее количество дважды положительных клеток в процентах от piwi-1 + клеток, как было ранее опубликовано (Adler et al. 2014). Авторы также не уточняют, является ли изменение, наблюдаемое между «3 dpa phx» и «3 dpa phx» значительным, что является существенным моментом.
Мы рассмотрели проблемы, связанные с биологическими повторениями, размерами выборки и статистическим анализом, как описано в наших ответах на пункты 1 и 3.Мы также попытались указать, когда соответствующие различия не являются статистически значимыми, как в случае сравнения, о котором идет речь выше.
Что касается представления наших данных как абсолютного числа ячеек, а не как пропорции, рецензент прав, отмечая, что эта стратегия является отходом от нашей предыдущей статьи, где FoxA + piwi-1 + клетки были представлены в виде относительного процента (Adler et al. , 2014). Однако аналогичная количественная оценка глазных предшественников ovo + представлена в виде абсолютных чисел, а не в процентах (Lapan et al.и LoCascio et al.). Не имея четких стандартов количественной оценки в литературе, мы первоначально оценили обе стратегии, но обнаружили, что результаты очень похожи. Мы включили два рисунка параллельного сравнения этих двух стратегий количественной оценки для ключевых панелей из нашей рукописи (изображения ответа автора 1 и 2). Учитывая сложную динамику популяции стволовых клеток, мы считаем, что представление данных в виде относительных процентов упрощает наш анализ. Поскольку наш анализ предшественников первоначально сравнивает сценарии множественных травм, мы уверены, что изменения, которые мы видим в предшественниках, происходят независимо от общих реакций на травмы.Кроме того, представление данных как среднее абсолютное количество клеток в стандартизированной площади / толщине позволяет нам указывать отдельных животных и биологические копии на графиках, обеспечивая более прозрачное представление данных для читателей.
, 2014). Однако аналогичная количественная оценка глазных предшественников ovo + представлена в виде абсолютных чисел, а не в процентах (Lapan et al.и LoCascio et al.). Не имея четких стандартов количественной оценки в литературе, мы первоначально оценили обе стратегии, но обнаружили, что результаты очень похожи. Мы включили два рисунка параллельного сравнения этих двух стратегий количественной оценки для ключевых панелей из нашей рукописи (изображения ответа автора 1 и 2). Учитывая сложную динамику популяции стволовых клеток, мы считаем, что представление данных в виде относительных процентов упрощает наш анализ. Поскольку наш анализ предшественников первоначально сравнивает сценарии множественных травм, мы уверены, что изменения, которые мы видим в предшественниках, происходят независимо от общих реакций на травмы.Кроме того, представление данных как среднее абсолютное количество клеток в стандартизированной площади / толщине позволяет нам указывать отдельных животных и биологические копии на графиках, обеспечивая более прозрачное представление данных для читателей. Основываясь на этих критериях, мы сохранили нашу стратегию представления данных в виде абсолютных значений, наряду с обширными дополнениями к рукописи, разъясняющими, где и как были получены данные.
Основываясь на этих критериях, мы сохранили нашу стратегию представления данных в виде абсолютных значений, наряду с обширными дополнениями к рукописи, разъясняющими, где и как были получены данные.
Потеря глотки избирательно увеличивает количество предшественников глотки пропорционально стволовым клеткам.
(A) Доля клеток, дважды положительных по указанному маркеру-предшественнику и piwi-1 + относительно всех стволовых клеток piwi-1 + в области, обведенной пунктирными прямоугольниками на рисунках. В мультфильмах изображены разные условия ампутации. n ≥ 790 клеток на экспериментальную группу из 3 независимых экспериментов. (B) Среднее количество клеток FoxA + piwi-1 + у тех же животных и регионов, что и A.Те же данные, что и на рис. 2B, E рукописи. (C) Доля FoxA + piwi-1 + клеток в указанные моменты времени после ампутации глотки относительно всех стволовых клеток piwi-1 + в области, обведенной пунктирными прямоугольниками в A. n ≥ 631 клетка на экспериментальную группу из 3 независимых экспериментов. (D) Среднее количество клеток FoxA + piwi-1 + у тех же животных и областей, проанализированных как C. Те же данные, что и на рисунке 2C рукописи.Для всех графиков область размером 6000 мкм 2 в том же месте префарингеальной области была проанализирована на 20 z-срезах, которые представлены пунктирными прямоугольниками на A. Графики представляют пропорцию ± 95% доверительных интервалов (A, C) или среднее значение ± стандартное отклонение с символами = отдельные животные; формы различают биологические копии (B, D). *, p ≤ 0,05 **, p ≤ 0,01; ***, p ≤ 0,001; ****, p ≤ 0,0001, точный тест Фишера (A, C) или однофакторный дисперсионный анализ с тестом Тьюки (B, D).
2B, E рукописи. (C) Доля FoxA + piwi-1 + клеток в указанные моменты времени после ампутации глотки относительно всех стволовых клеток piwi-1 + в области, обведенной пунктирными прямоугольниками в A. n ≥ 631 клетка на экспериментальную группу из 3 независимых экспериментов. (D) Среднее количество клеток FoxA + piwi-1 + у тех же животных и областей, проанализированных как C. Те же данные, что и на рисунке 2C рукописи.Для всех графиков область размером 6000 мкм 2 в том же месте префарингеальной области была проанализирована на 20 z-срезах, которые представлены пунктирными прямоугольниками на A. Графики представляют пропорцию ± 95% доверительных интервалов (A, C) или среднее значение ± стандартное отклонение с символами = отдельные животные; формы различают биологические копии (B, D). *, p ≤ 0,05 **, p ≤ 0,01; ***, p ≤ 0,001; ****, p ≤ 0,0001, точный тест Фишера (A, C) или однофакторный дисперсионный анализ с тестом Тьюки (B, D).
Потеря ткани глотки избирательно увеличивает митотически активные предшественники глотки.
(A) Доля FoxA + h4P + клеток относительно всех стволовых клеток h4P + в указанные моменты времени после ампутации глотки или головы в области, обведенной пунктирными прямоугольниками в E. n ≥ 515 клеток на эксперимент группа из 2 независимых экспериментов. (B) Среднее количество клеток FoxA + h4P + , определенное количественно у тех же животных и регионов, что и A. Те же данные, что и на рис. 3C, D рукописи.(C) Доля клеток, дважды положительных по указанному маркеру предшественника и h4P + относительно всех стволовых клеток h4P + в области, обведенной пунктирными прямоугольниками в E. n ≥ 472 клеток на экспериментальную группу из 2 независимых экспериментов. (D) Среднее количество клеток, дважды положительных по указанному маркеру-предшественнику и количественно определенному h4P + у тех же животных и регионов, что и C. Те же данные, что и на рисунке 3E рукописи. (E) Мультфильмы, изображающие различные условия ампутации.Для A-D вся преглоточная область была проанализирована на 30 z-срезах, которые представлены пунктирными прямоугольниками. Графики представляют пропорцию ± 95% доверительные интервалы (A, C) или среднее ± SD с символами = отдельные животные; формы различают биологические копии (B, D). *, p ≤ 0,05 **, p ≤ 0,01; ***, р <0,001; ****, p ≤ 0,0001, точный тест Фишера (A, C) или однофакторный дисперсионный анализ с тестом Тьюки (B, D)
n ≥ 472 клеток на экспериментальную группу из 2 независимых экспериментов. (D) Среднее количество клеток, дважды положительных по указанному маркеру-предшественнику и количественно определенному h4P + у тех же животных и регионов, что и C. Те же данные, что и на рисунке 3E рукописи. (E) Мультфильмы, изображающие различные условия ампутации.Для A-D вся преглоточная область была проанализирована на 30 z-срезах, которые представлены пунктирными прямоугольниками. Графики представляют пропорцию ± 95% доверительные интервалы (A, C) или среднее ± SD с символами = отдельные животные; формы различают биологические копии (B, D). *, p ≤ 0,05 **, p ≤ 0,01; ***, р <0,001; ****, p ≤ 0,0001, точный тест Фишера (A, C) или однофакторный дисперсионный анализ с тестом Тьюки (B, D)
5. Рисунок 2D также страдает от неадекватных методов количественной оценки, описанных выше.В идеале клетки FoxA + следует количественно определять как процент наблюдаемых клеток h4P +.
Мы решили эту проблему, как описано в пунктах 1–4 выше.
6. Авторы используют термины «стволовые клетки», «предшественники», «стволовые клетки-предшественники» и «стволовые клетки-предшественники» в смешанной и запутанной форме на протяжении всей статьи. Например, в строках 174-175 авторы заявляют, что «пролиферация стволовых клеток FoxA + предшествует увеличению числа предшественников глотки». Это относится к клеткам FoxA + h4P + vs.FoxA + piwi-1 + клетки, но с той лишь разницей, что первые являются стволовыми клетками в акте митоза. Проводится ли различие? В другом месте статьи FoxA + piwi + клетки называются стволовыми клетками. Используемая терминология требует большей ясности и последовательности.
Что такое FoxA + PSC (строка 168)? Предполагают ли авторы, что FoxA является маркером плюрипотентности? Или авторы говорят, что tgs -1 имеет более широкий паттерн экспрессии и коэкспрессируется с маркерами предшественников? Если двойной FISH был выполнен с указанными маркерами-предшественниками (ovo, myoD, gata-4/5/6, six-1/2 и pax6), все ли они перекрывались с tgs -1 ? Эти эксперименты необходимо провести, чтобы сделать какие-либо заявления о экспрессии FoxA в контексте плюрипотентности.

Мы согласны с тем, что нашему языку, особенно в отношении терминологии, относящейся к стволовым клеткам, предшественникам и органам-предшественникам, недостает единообразия. Мы изменили язык следующим образом: 1) мы называем «стволовые клетки» piwi-1 + или клетки h4P + , 2) мы используем термин «органоспецифические предшественники» для обозначения к клеткам, дважды положительным по маркерам предшественников и h4P или piwi-1 , как обозначено на фиг. 2 и 3, и 3) мы удалили все примеры сбивающих с толку терминов «стволовые клетки-предшественники» и «стволовые клетки-предшественники».Кроме того, мы удалили данные tgs-1 и любые утверждения о плюрипотентных стволовых клетках, которые, по нашему мнению, были предварительными и неубедительными.
7. На рисунке 3С показано, как происходит регенерация глотки в течение такого длительного периода времени после лечения нокодазолом в течение 1-2 дней? Происходит ли регенерация вслепую, если это окно пропущено? Авторы должны повторить анализ предшественников FoxA + в более поздние моменты времени в этом состоянии и / или показать скорость включения BrdU в глотку с лечением нокодазолом и без него в этом окне.
Мы согласны с тем, что было бы полезно знать, как животные в конечном итоге регенерируют после возмущения предшественников глотки, и действуют ли нецелевые механизмы, чтобы медленно регенерировать этот орган. Однако мы изо всех сил пытались разработать эксперимент, чтобы четко определить эти параметры. Лечение нокодазолом в течение 1-2-дневного окна, вероятно, приводит к уничтожению предшественников глотки путем апоптоза. Поскольку предшественники не подвергаются митозу с такой же высокой скоростью у интактных животных, лечение нокодазолом не повлияет на них аналогичным образом, что затрудняет сравнение.Следовательно, введение BrdU во время этого окна потенциально может пометить совершенно разные популяции стволовых клеток.
8. Использование ингибитора PD на рисунке 4 проблематично. Не представлены данные, подтверждающие, что фосфорилирование ERK было ингибировано в этих экспериментах. Ссылки на предыдущее использование недостаточно.
Влияние PD на необрезанных животных WT в отношении клеток FoxA + не показано и является необходимым контролем. Чтобы ответить на вопросы о лекарственной специфичности, авторы должны подтвердить свои выводы вторым ингибитором передачи сигналов ERK, таким как U0126, который, как уже было показано, работает у планарий (Owlarn et al.2017).
Мы рассмотрели все проблемы, поднятые выше. 1) Все эксперименты с ингибитором MEK были повторены с UO126 со сравнимыми результатами (фиг. 5 — приложение к рисунку 1C-F, фиг. 5 — приложение к рисунку 3B-E, фиг. 6). 2) Вестерн-блоттинг подтверждает, что обработка ингибитором MEK в течение всего лишь 24 часов предотвращает фосфорилирование ERK (Рисунок 5 — рисунок в приложении 1E). 3) Теперь мы включаем данные, показывающие, что воздействие любого из ингибиторов блокирует регенерацию во фрагментах хвоста, указывая на то, что дефект регенерации глотки не является следствием неэффективного ингибирования (Рисунок 5 — рисунок в приложении 1F). 4) Теперь мы включаем количественную оценку FoxA + piwi-1 + у интактных животных после лечения PD и UO (Рисунок 5 — приложение к рисунку 2A).
4) Теперь мы включаем количественную оценку FoxA + piwi-1 + у интактных животных после лечения PD и UO (Рисунок 5 — приложение к рисунку 2A).
9. Заключение о том, что передача сигналов ERK регулирует дифференцировку, но не пролиферацию, является преждевременным (линия 249-264). Цифры 4E-H следует количественно определять как процентное соотношение клеток piwi-1 + и h4P +, особенно потому, что при лечении PD снижается общая пролиферация.
В текущей версии рукописи мы приложили все усилия, чтобы проанализировать роль передачи сигналов ERK в делении стволовых клеток по сравнению с дифференцировкой.Сейчас мы тщательно проверили график потребности в ERK при регенерации глотки. Основываясь на воздействии PD или UO (рис. 5C-H и рис. 5 — приложения к рисункам 1 и 3), наши данные теперь ясно показывают, что активность ERK не требуется до деления стволовых клеток, которое происходит через 1-2 дня после ампутации глотки. . Кроме того, хотя верно то, что воздействие PD и UO действительно снижает общее деление стволовых клеток, эти лекарственные препараты не влияют на деление предшественников глотки (Рисунок 5H и Рисунок 5 — приложение к рисунку 3D, 3E).Поскольку анализ клеток FoxA + h4P + и общего количества клеток h4P + проводится на одних и тех же животных, можно экстраполировать, что пропорциональные данные будут выглядеть одинаково. Фактически, уменьшение общего h4P увеличило бы относительное количество FoxA + h4P + клеток. Мы добавили текст, чтобы прояснить это в разделе «Результаты» (строки 426-429).
. Кроме того, хотя верно то, что воздействие PD и UO действительно снижает общее деление стволовых клеток, эти лекарственные препараты не влияют на деление предшественников глотки (Рисунок 5H и Рисунок 5 — приложение к рисунку 3D, 3E).Поскольку анализ клеток FoxA + h4P + и общего количества клеток h4P + проводится на одних и тех же животных, можно экстраполировать, что пропорциональные данные будут выглядеть одинаково. Фактически, уменьшение общего h4P увеличило бы относительное количество FoxA + h4P + клеток. Мы добавили текст, чтобы прояснить это в разделе «Результаты» (строки 426-429).
10. Использование фрагментов головы для сравнения регенерации глаза и регенерации глаза.регенерация глотки неуместна. Предыдущие исследования уже показали, что отсутствие глаз не обязательно для индукции амплификации ovo + предшественников (LoCascio et al. 2017). Таким образом, это не удивительный результат (строки 343-345), и авторы неверно цитируют предыдущие наблюдения (в более ранней работе фрагменты головы никогда не описывались).
Область, в которой были количественно определены ovo + клетки на Фигуре 6А, не обоснована или объяснена. Желтое поле помещено в медиальную область, где обычно не располагаются ovo + клетки.Авторы должны делать снимки внутри расположенных латерально потоков ovo +, которые были описаны ранее (Lapan and Reddien 2012; LoCascio et al.2017).
Мы согласны с тем, что этот эксперимент мало что добавил к общим результатам, и мы не смогли адекватно объяснить наше обоснование его включения. Он был удален из этой версии.
11. Вместо того, чтобы использовать фрагменты головы, авторам следует повторить эксперименты по резекции боковых сторон, показанные ранее (LoCascio et al.2017). Это предыдущее исследование показало, что увеличение инкорпорации BrdU в глотку происходило после резекции фланга, даже если глотка присутствовала. Этот результат мог быть 1) артефактом повышенного окрашивания BrdU из-за стимуляции пролиферации при повреждении, 2) вызванным непреднамеренным повреждением клеток, связанных с глоткой, или 3) ответом на потерю популяций предшественников FoxA +, которые окружают глотку.
а не потеря дифференцированного органа. У авторов есть возможность пересмотреть это опубликованное наблюдение путем количественной оценки реакции предшественников FoxA + во время резекции фланга +/- глотки.Без этих данных эта история неполна, и поэтому вывод о целевой реакции регенерации еще не убедителен.
Мы ценим предложение рецензента включить этот важный эксперимент. Теперь мы включаем количественную оценку FoxA + piwi-1 + клеток после резекции фланга (рис. 2D), что доказывает, что повреждения вне глотки не вызывают увеличения числа предшественников глотки. Кроме того, мы вводили EdU в разное время (0 или 24 часа) после ампутации головы или глотки (рис. 1E, F).Мы обнаружили, что клетки, образовавшиеся сразу (0 часов) после ампутации, широко включаются, подтверждая результат (LoCascio et al., 2017) о том, что резекция боковых сторон увеличивает маркировку BrdU в неповрежденном глотке. Важно отметить, что при введении EdU через 1 день после ампутации мы обнаружили, что клетки, образовавшиеся в это время, направлялись конкретно в регенерирующие органы, что дает доказательства, подтверждающие целевой механизм регенерации, который выборочно продуцирует предшественников отсутствующих тканей.
12.Отрицательные результаты о том, что пролиферация и передача сигналов ERK не обязательны для регенерации глаза на фиг. 6, слабы и неубедительны. Восстановленные глаза кажутся меньше; это должно быть определено количественно (количество PRN на глаз). Если маленькие фаринги, которые образуются на рисунках 3D и 4C, считаются вредоносным фенотипом, почему тот же стандарт не применяется к глазу? Кроме того, существующих глазных предшественников могло быть достаточно для регенерации глаз при лечении этими лекарственными препаратами. Более того, в 50% случаев глаза не восстанавливались после лечения нокодазолом.Не является ли более вероятным, что представленные наблюдения являются артефактами дозировки и времени? Как это повлияло на распространение? Эти наблюдения не соответствуют сделанным заявлениям.
Мы рассмотрели потенциальные проблемы с дозировкой и сроками, стандартизовав время воздействия для экспериментов с нокодазолом в течение 2 дней, увеличив количество животных и улучшив количественную оценку. Пролиферация и активность ERK необходимы только для регенерации глаза в контексте большой ампутации, но не только после резекции глаза (рис. 6).Что касается глотки, ламинин сильно экспрессируется в глотке и во рту и слабо в теле, где глотка прикрепляется. Даже несмотря на то, что через 7 дней после ампутации PD, UO или нокодазолом остается остаточное окрашивание ламинином (Рисунок 4D, 5D, Рисунок 5 — рисунок в приложении 1D), это вовсе не «глотка». Нормальная архитектура, которая проявляется в элементах управления ДМСО и на дополнительных фигурах, связанных с фигурами 4 и 5, полностью теряется после этих обработок. Мы разъяснили это в письменной форме (строки 290-296).И наоборот, регенерация глаз у животных, получавших лекарственные препараты, несомненно, более сопоставима с контролем (рис. 6В).
Пролиферация и активность ERK необходимы только для регенерации глаза в контексте большой ампутации, но не только после резекции глаза (рис. 6).Что касается глотки, ламинин сильно экспрессируется в глотке и во рту и слабо в теле, где глотка прикрепляется. Даже несмотря на то, что через 7 дней после ампутации PD, UO или нокодазолом остается остаточное окрашивание ламинином (Рисунок 4D, 5D, Рисунок 5 — рисунок в приложении 1D), это вовсе не «глотка». Нормальная архитектура, которая проявляется в элементах управления ДМСО и на дополнительных фигурах, связанных с фигурами 4 и 5, полностью теряется после этих обработок. Мы разъяснили это в письменной форме (строки 290-296).И наоборот, регенерация глаз у животных, получавших лекарственные препараты, несомненно, более сопоставима с контролем (рис. 6В).
Регенерация глаза опосредуется гомеостатическим обновлением, но остается неясным, достаточно ли существующих глазных предшественников для регенерации этих небольших структур после лечения лекарствами. Мы добавили в этот раздел текст, чтобы учесть эту возможность (строки 450-453).
Мы добавили в этот раздел текст, чтобы учесть эту возможность (строки 450-453).
13. Авторы утверждают, что ovo + клетки не являются пролиферативными (h4P +) даже в тех случаях, когда имеет место амплификация предшественников глаза (ампутация головы), но данные не показаны (строка 321).Они должны быть. Действительно, в предыдущих публикациях никогда не было показано, что ovo + клетки пролиферируют. Это может означать, что существуют пролиферирующие клетки-предшественники глаза, которые предшествуют экспрессии ovo. Авторам следует обсудить эту альтернативу.
Теперь мы включили изображения для клеток ovo + h4P + у интактных животных и после ампутации головы с обработкой или без обработки нокодазолом для обогащения клеток в митозе (Рисунок 3 — рисунок в приложении 3). Мы смогли обнаружить только делящиеся клетки ovo, , + после лечения нокодазолом, но не наблюдали какого-либо увеличения после ампутации головы по сравнению с контрольными группами, получавшими интактный нокодазол. Как было предложено, мы добавили обсуждение возможных объяснений в этот раздел (строки 265-267).
Как было предложено, мы добавили обсуждение возможных объяснений в этот раздел (строки 265-267).
14. Авт. Утверждают, что регенерация глаза не требует пролиферации или передачи сигналов ERK, но регенерация глотки требует. Этот вывод основан на общем наблюдении того, что глаза могут регенерироваться в присутствии нокодазола и PD (см. Пункт 12 выше). Эти данные грубые, а интерпретации неубедительны. Вместо этого их модель может быть непосредственно протестирована с помощью экспериментов с отслеживанием импульсов BrdU.Согласно модели авторов, можно было бы предсказать, что после ампутации глотки скорость включения клеток BrdU + в регенерирующую глотку должна быть выше, чем в неповрежденном контроле. И наоборот, скорость включения BrdU в регенерирующий глаз должна оставаться неизменной между травмированными (т.е. удаленными глазами) и контрольными животными (LoCascio et al.2017). После того, как авторы сделали вышеприведенное предсказание, у них есть возможность продемонстрировать эффекты нокодазола, PD и U0126 на включение BrdU в регенерирующем глазу по сравнению с глоткой после резекции глаза и ампутации глотки у одного и того же животного.
Таким образом, авт. Могут напрямую проверить потребность в пролиферации и / или передаче сигналов ERK в обеих тканях.
Мы согласны с рецензентом в том, что это важный эксперимент, и теперь мы рассмотрели различный вклад клеток EdU + либо в регенерирующий глотку, либо в мозг (рис. 1E, F). Однако мы зарезервировали фармакологические отклонения на будущее.
Рецензент № 2:
Бор, Широор и Адлер исследуют, как стволовые клетки реагируют на потерю определенных тканей у планарий.Популяция планарий стволовых клеток (необластов) распределена по всему телу планарий и включает плюрипотентные стволовые клетки и широкий спектр клеток-предшественников, коммитированных по клону. Как этот гетерогенный пул клеток ведет себя после травмы или ампутации, не совсем понятно. Открытие маркеров для маркировки потомства стволовых клеток открыло дверь для исследования реакции стволовых клеток на потерю ткани. Однако анатомия планарий затрудняет хирургическое удаление или повреждение определенных органов.
ИП этого исследования разработал метод удаления глотки путем «химической ампутации», чтобы изучить механизмы, лежащие в основе регенерации этого органа, без резкого нарушения или повреждения других тканей. Используя этот подход, в данной статье исследуется, как четко определенная популяция предшественников FoxA + реагирует на удаление глотки в ранние моменты времени во время регенерации. Их данные предполагают, что стволовые клетки способны обнаруживать потерю глотки и реагировать, генерируя значительно больше клеток, которым суждено стать глоткой, тогда как ампутация нефарингеальных тканей не оказывает очевидного влияния на динамику спецификации предшественников глотки.Кроме того, используя фармакологические методы лечения, авт. Показывают, что пролиферация клеток и передача сигналов ERK необходимы для размножения предшественников глотки и дифференцировки клеток. Напротив, другие типы клеток в глазу планарии, по-видимому, не нуждаются в пролиферации или передаче сигналов ERK, предполагая, что ответы стволовых клеток «нацелены на слепую», как предполагалось в предыдущем исследовании, а скорее настроены на конкретные отсутствующие ткани.

Эта работа может внести значительный вклад в эту область, продвигая наше понимание того, как популяция гетерогенных стволовых клеток планарий реагирует на потерю определенного органа.Однако отчет является предварительным в том виде, в котором он представлен. Похоже, что авторы провели множество экспериментов за один раз. Кроме того, описания методов недостаточно. Следовательно, прежде чем эту работу можно будет рассматривать для публикации, авторам необходимо в основном продемонстрировать воспроизводимость данных и надежность наблюдений.
1. Авторам необходимо повторить эксперименты, чтобы увеличить размер выборки для большинства экспериментов.
Мы решили эту проблему, как подробно описано в обзоре выше.Все эксперименты были повторены, увеличивая размеры выборки по всем направлениям. Мы также включили таблицу с подробным описанием биологических и технических копий для каждой соответствующей фигуры.
2. Подробности для визуализации и количественной оценки должны быть четко указаны в методах, а зарегистрированные числа клеток должны быть нормализованы, как необходимо для каждой серии экспериментов.

Мы решили эту проблему, как подробно описано в обзоре выше. Вкратце, методы построения изображения и количественной оценки теперь подробно описаны в разделе легенд и методов фигур, озаглавленном «Получение изображения, количественная оценка и статистический анализ» (начиная со строки 759).Анализируемые области были либо стандартной площади, либо нормализованы к площади, и для данного эксперимента конфокальной визуализации количество z-сечений оставалось неизменным.
3. Хотя авторы упоминают эксперименты по «отслеживанию клонов», они не проводят эксперименты по отслеживанию импульсов по аналогу ДНК для анализа временной прогрессии и пространственной локализации стволовых клеток в предшественниках FoxA + после удаления глотки. Авторы полагаются на окрашивание Ph4 в сочетании с FoxA и дополнительные эксперименты с использованием маркера плюрипотентных стволовых клеток tgs -1 (который был исследован только при 1 dpa).Не могли бы авторы пояснить, что, по их мнению, представляют собой стволовые клетки FoxA +? Это самообновляющиеся плюрипотентные стволовые клетки или предшественники с коммитом по клону? Могут ли авторы получить некоторое представление, отсканировав свои изображения клеток Ph4 +, экспрессирующих FoxA, явно претерпевающих метафазу? Являются ли дочерние клетки однородными с FoxA +, как это отражено в модели? По крайней мере, в одной из клеток, показанных в контрольных группах, обработанных нокодазолом, обе дочерние клетки экспрессируют FoxA (фиг.
3). Я предлагаю показать несколько изображений с большим увеличением, чтобы поддержать интерпретации / выводы.Другие утверждали (например, Ринк, глава 2 книги «Регенерация планарий: методы и протоколы»), что не каждая делящаяся клетка может быть долговременной самообновляющейся стволовой клеткой, и неизвестно, существуют ли временные амплифицирующие клетки или способствуют ли они регенерации у планарий. . Адлер и Санчес Альварадо (2015) обсуждают роль переходных состояний и то, как изменяются профили транскрипции в ответ на регенерацию. Мне было неясно, как авторы думают об этих клетках, основываясь на ограниченном количестве экспериментов и анализов, и есть несколько мест, где терминология непоследовательна, особенно в отношении пролиферирующих предшественников ovo + (P.14). Авторы должны быть ясны, и было бы полезно проиллюстрировать их модель на одном из ранних рисунков или включить ее в окончательную модель, в которой опущены tgs -1 из-за ограниченного количества экспериментов, проведенных с этим маркером генов.
(Рисунок 7).
Мы ценим предложение прояснить наш язык, связанный со стволовыми клетками, плюрипотентными стволовыми клетками и предшественниками. Чтобы обратиться к важному моменту, касающемуся временной прогрессии предшественников глотки, мы теперь включаем эксперименты с импульсной погоней по аналогу ДНК (EdU) для анализа продукции стволовых клеток, генерируемых либо сразу после ампутации, либо через 1 день, когда мы видим конкретное увеличение глотки. прародители.Этот эксперимент (рис. 1E, F) показывает, что стволовые клетки, полученные через 1 день после ампутации, выборочно включаются в регенерирующий зев. Как указано в пункте 4 обзора выше, мы сделали нашу терминологию во всей рукописи более последовательной.
Мы согласны с рецензентом в отношении сложности популяции стволовых клеток, которая включает как долгоживущие самообновляющиеся клетки, так и предшественники. Поскольку мы не могли сделать четких выводов на основе данных tgs-1 , мы удалили этот рисунок и любые утверждения о плюрипотентных стволовых клетках из этой версии рукописи. Хотя у нас нет данных, чтобы окончательно указать на эффективность предшественников FoxA + , теперь мы включаем изображения анафазных клеток, которые предполагают, что FoxA может сегрегировать на дочерние клетки как симметрично, так и асимметрично (Рисунок 3 — приложение к рисунку 1A) . Мы добавили раздел, посвященный этим вопросам, в обсуждение, потому что мы согласны с тем, что исследование этих вопросов важно (строки 511-528).
Хотя у нас нет данных, чтобы окончательно указать на эффективность предшественников FoxA + , теперь мы включаем изображения анафазных клеток, которые предполагают, что FoxA может сегрегировать на дочерние клетки как симметрично, так и асимметрично (Рисунок 3 — приложение к рисунку 1A) . Мы добавили раздел, посвященный этим вопросам, в обсуждение, потому что мы согласны с тем, что исследование этих вопросов важно (строки 511-528).
4. Глотка сложна, и нет данных для оценки возможного вклада других популяций предшественников.Я не думаю, что маловероятно, что предшественники FoxA + несут единоличную ответственность за реконструкцию глотки. Авт. Должны исследовать, как др. Популяции предшественников ведут себя во время процесса регенерации глотки, увеличивая временную шкалу анализа клеток предшественников. Это позволит выявить колебания в динамике предшественников по мере того, как животные регенерируют глотку или меняют свои пропорции после регенерации глотки. Например, могут ли авторы проверить, способны ли они обнаружить вклад нейральных предшественников в регенерацию глоточной нервной системы? И если да, то когда в процессе регенерации это происходит в контексте их изучения?
Рецензент прав, отмечая, что у нас до сих пор нет окончательных доказательств того, являются ли предшественники FoxA + единственным источником ткани глотки или генерируют только подмножество критических клеток.Глотка действительно сложный орган, и мы хотели бы знать, является ли FoxA главным регулятором их всех. Чтобы решить этот вопрос, мы уничтожили каждого предшественника, включенного в эту статью, и каждого предполагаемого предшественника глотки из литературы ( dd554 , meis , twist ), и ни один из них, похоже, не играет никакой роли в регенерации глотки (рис. 2 — дополнение к рисунку 3 и данные не показаны). Отсутствие детектируемого фенотипа этих маркеров заставляет нас неохотно делать выводы из их динамики во время регенерации глотки.
Рецензент № 3:
В этой рукописи Bohr et al. изучить, как система плюрипотентных стволовых клеток планарий реагирует на органоспецифические повреждения. Если и каким образом дифференциация определенных типов клеток динамически регулируется, является концептуально интересной проблемой для планарий и в целом при исследовании стволовых клеток. Авторы обращаются к этой проблеме, сравнивая ответ стволовых клеток при ампутации одного органа (глотки) и потере обширной ткани (декапитация).Их результаты показывают, что только удаление глотки запускает усиленную дифференцировку типов клеток глотки, в то время как потеря нефарингеальных тканей активизирует дифференцировку предшественников множества органов, но не глотки. Далее авт. Предполагают, что ограниченная во времени передача сигналов ERK является регуляторным компонентом дифференцировки типов глоточных клеток. Эти наблюдения важны еще и потому, что они контрастируют с ранее предложенной «целевой слепой» моделью (LoCascio et al., 2017), который утверждает, что дифференциация разных типов клеток находится в постоянных относительных пропорциях, а скорость делений стволовых клеток является глобальным регулятором скорости производства. Напротив, наблюдения Бора и др. предоставить дополнительные доказательства большей гибкости и специфичности в системе стволовых клеток планарии («целевое сознание») в смысле специфичных для клонов корректировок в скорости продукции определенных типов клеток.
Тем не менее, рукопись обычно страдает излишне узкой направленностью.Остаются важные вопросы относительно специфичности ответа стволовых клеток на ампутацию глотки, и во многих экспериментах отсутствуют важные элементы контроля (см. Ниже). Более того, авт. Упустили из виду, что «сознательный к мишени» ответ предшественников уже был продемонстрирован посредством избирательной пролиферации протонефридиального маркера, экспрессирующего необласты, в ответ на протонефридиальное повреждение с помощью RNAi (Vu et al., 2015). В результате текущая рукопись потребует значительных дополнительных экспериментов, чтобы консолидировать полученные результаты в достаточной степени для публикации в eLife.
Мы признательны рецензентам за признание нашей работы и их вдумчивую критику. В обновленную версию мы добавили новые элементы управления, включили важную ссылку Vu et al., 2015, и подробно обсудили конфликтующие модели регенерации (в нашей рукописи называемые «целевыми» и «нецелевыми»). Обращение к этой критике привело к значительно более ясной рукописи, которая помогает нам лучше понять механизмы, лежащие в основе регенерации.
1. Специфичность ответа стволовых клеток
Центральная посылка статьи — селективная амплификация предшественников глотки в ответ на ампутацию глотки. Этот вывод авторы делают на основании i) увеличения абсолютного количества клеток foxA + / piwi-1 + в определенной области, в то время как ii) уменьшение количества клеток не влияет на абсолютное количество клеток foxA + / piwi-1 + в та же площадь. Этот подход является недостаточной демонстрацией специфичности, так как известный феномен активации стволовых клеток, индуцированный раной, может также изменять абсолютное количество специфических подклассов необластов и может делать это зависимым от травмы образом.
Чтобы учесть это важное предостережение, авторам необходимо i) количественно определить ОТНОСИТЕЛЬНЫЕ пропорции клеток foxA + / piwi-1 + от общего количества клеток piwi (или от общего количества клеток h4P +) и ii) им необходимо включить другие предшественники органов в исходные данные. анализ. Последнее также имеет решающее значение, поскольку глотка — сложный орган, включающий потомков нескольких ветвей (например, мышцы, нейроны, эпидермис), и неясно, действительно ли клетки foxA + / piwi-1 + служат единственным источником всех составляющих линий. (как предполагают авторы), или если они предоставляют только подмножество глоточных клеток с лимитирующей ролью в сборке глотки (например,g., глоточная мышца). Перед лицом такой неопределенности, iii) количественная оценка включения новых клеток в глотку по сравнению с другими тканями с помощью мечения BrdU будет необходима для устранения этого предостережения и обеспечения глобальной перспективы специфичности ответа независимо от не полностью охарактеризованных маркерных генов.
Кроме того, необходимо решить или лучше задокументировать следующие проблемы экспериментального проектирования, в том числе:
• Авторы предоставляют недостаточную методологическую детализацию количественной оценки предшественников, даже несмотря на то, что вся рукопись основана на этом анализе.Каковы их критерии для оценки клетки piwi-1 + как дважды положительной для часто слабых и шумных меток клонов? Если проводилось «на глаз», использовалось ли двойное слепое подсчет очков? Были ли подсчитаны все ячейки в данном Z-стеке или только определенные плоскости? В последнем случае, по каким критериям были выбраны плоскости изображения для количественной оценки? Как были выбраны конкретные образцы из экспериментальной когорты для визуализации / количественной оценки? Хотя это не обязательно является уникальным недостатком этого конкретного исследования, эти моменты просто необходимо должным образом рассмотреть, чтобы строго поддерживать количественные различия между экспериментальными условиями (например,г., специфичность).
• Похоже, что авторы вообще не делают различия между техническими повторениями (например, несколько образцов в экспериментальной когорте) и биологическими повторениями (независимые экспериментальные группы). Это важно, потому что i) использование стандартной ошибки среднего (SEM), которую авторы используют повсюду, на самом деле не является подходящей мерой для одной биологической повторности с 3 животными — стандартное отклонение (SD) могло бы показаться более подходящим. соответствующая мера в этом контексте (SD).ii) количество червей, определенное количественно для каждого эксперимента, обычно невелико (n = 3 животных на рисунках 1, 3, 5, 6; n = 5 животных на остальных рисунках) с учетом наблюдаемой изменчивости данных (например, ~ 25-30 foxA + / piwi-1 + клеток в 3d после ампутации глотки на рисунке 1E по сравнению с 50 клетками на рисунке 1H). Точно так же для кинетических экспериментов, как на рис. 1H или 2C, просто крайне важно убедиться, что планки ошибок включают вариации в динамике ответа между множественными повторами из-за дрейфа базовой фракции h4P + -клеток или различной эффективности окрашивания (например.g., разные партии животных в разные дни), а не технические вариации только в одной экспериментальной когорте. Пожалуйста, устраните эти опасения, добавив больше образцов и подробное описание экспериментального плана.
Мы благодарим рецензента за выделение этих важных замечаний в нашей интерпретации наших данных. В пункте 2 введения к этому ответу и на рисунках 1 и 2 ответов автора мы включили параллельное сравнение абсолютной и относительной количественной оценки и показали, что для ключевых фигур в нашей рукописи две стратегии дают очень похожие результаты. полученные результаты.Кроме того, поскольку мы сравниваем различные парадигмы ранения, которые вызывают общие реакции раны, наши эксперименты находятся под внутренним контролем на предмет пролиферации, вызванной раной.
Рецензент прав, отмечая, что у нас до сих пор нет окончательных доказательств того, являются ли предшественники FoxA + единственным источником ткани глотки или генерируют только подмножество критических клеток (также рассмотрено в основной проблеме рецензента 4 выше). Чтобы решить этот вопрос, мы выбрали каждого предшественника, использованного в этой статье, и каждого предполагаемого предшественника глотки из литературы ( dd554 , meis , twist ).Ни один из них не вызывает дефектов регенерации глотки (Рисунок 2 — дополнение к рисунку 3 и данные не показаны). В текущую версию рукописи мы также включаем метку в режиме отслеживания импульсов с помощью EdU, чтобы продемонстрировать, что стволовые клетки действительно избирательно включаются в регенерирующий зев, в зависимости от того, когда они пролиферируют относительно удаления ткани (рис. 1E, F). Однако мы признаем оговорку, что FoxA может контролировать производство только подмножества клеток. Текущая работа, не включенная в этот документ, направлена на устранение этого недостатка.
Мы также исправили все недостатки, указанные рецензентом, связанные с повторениями и статистическим анализом в этой версии, как указано во введении к этому ответу, пункт 1. Вкратце, они включают: 1) уточнение и стандартизация методов визуализации и количественной оценки. , 2) различение биологических и технических повторов во всех экспериментах, 3) заменены полосы ошибок, представляющие SEM, на полосы ошибок, представляющие стандартное отклонение, и 4) увеличенное количество животных в каждом эксперименте.
2. Хронология регенерации глотки
Хронология регенерации глотки и связанные с ней события, которые представляют авторы, недостаточно подтверждены экспериментальными данными. Вывод в строке 215 «о том, что пролиферация в критическом окне через 1-2 дня после ампутации глотки дает популяцию предшественников, которые, вероятно, необходимы для регенерации глотки», основывается на i) диагнозе «пролиферативного пика стволовых клеток FoxA +, который происходит. после ампутации глотки (рис. 2С) »(строка 202).Однако вместо «пика пролиферации» на фиг. 2C показано широкое «плато пролиферации» стволовых клеток FoxA + между 6 часами и 3 днями после ампутации. Точно так же количественная оценка foxA + / h4P + после ампутации глотки на рисунке 4G также показывает отсутствие пика foxA + / h4P + от 1 до 2 дней после ампутации. Ii) связанные эксперименты с нокодазолом страдают от того факта, что авторы не оценили количественно влияние препарата на численность клеток foxA + / piwi-1 + во время интервалов лечения от 0 до 1 дня и 2-3 дня после ампутации глотки.Таким образом, авторы не могут исключить, что лечение нокодазолом может иметь аналогичные эффекты на численность клеток foxA + / piwi-1 + в течение 1-3 дней после ампутации, с более тяжелым фенотипом на уровне органов 1-2d. окно лечения, вызванное каким-либо другим действием препарата (например, дифференцировкой другого ограничивающего скорость типа клеток для регенерации глотки или, возможно, ингибированием прайминга нейрональной активности). Сходное беспокойство касается утверждения в строке 242: «… окно через 1-2 дня после ампутации, в котором активация передачи сигналов ERK важна для регенерации глотки.«Здесь: i) количественная оценка фенотипа конечной стадии лекарственного лечения в течение 1-2d временного интервала (восстановление способности к кормлению) отсутствует. Ii) Аналогичным образом, исследование последствий лечения PD на экспрессию foxA + в piwi- Клетки 1+ на панели 4D-H используют замачивание лекарством в течение 3 дней, однако соответствующий фенотип конечной стадии 3-дневного лекарственного лечения не показан. Iii) влияние ERK на регенерацию глотки является предварительным. изначально правильно введенный как «ингибитор MEK», впоследствии авторы переходят на фактически неправильное обозначение «ингибитор ERK» (например,г., строка 358). Кроме того, было бы желательно получить дополнительные экспериментальные доказательства предполагаемого ингибирования Erk как причины наблюдаемых фенотипов, чтобы строго поддержать вывод.
Эти предостережения необходимо принять во внимание, чтобы хронология биологии клетки оставалась частью этой рукописи.
Благодарим рецензента за содержательную критику. Хотя мы согласны с тем, что пик FoxA + h4P + может больше напоминать щитовой вулкан, чем Маттерхорн, независимо от терминологии, наблюдается очевидное и значительное увеличение пролиферации, которое происходит в течение двух дней после ампутации глотки.Этот вывод был подкреплен предложениями рецензентов увеличить размер выборки и изменить методы статистического анализа. Теперь мы включаем дополнительную характеристику FoxA + piwi-1 + предшественников глотки после лечения нокодазолом через 0-1 и 2-3 дня после ампутации глотки (Рисунок 4 — рисунок в приложении 2E), подтверждая наши утверждения о том, что пролиферация возникновение через 1-2 дня после ампутации имеет решающее значение для производства предшественников глотки. Кроме того, это 1-2-дневное окно совпадает со временем выборочного включения EdU + в регенерирующий глотку, подтверждая наш вывод о том, что это 1-2-дневное пролиферативное окно важно для продукции предшественников глотки и последующей регенерации.
Что касается экспериментов с ERK, мы рассмотрели все вопросы, поднятые рецензентом, расширив наши эксперименты с ингибиторами MEK для более тщательного тестирования, когда ERK требуется во время регенерации глотки. Это включает стандартизацию продолжительности лечения ингибитором MEK и выполнение множества дополнительных контролей, чтобы показать, что несколько ингибиторов MEK предотвращают фосфорилирование ERK и приводят к одинаковым результатам в регенерации. Чтобы подкрепить доказательствами, подтверждающими нашу временную шкалу, мы сделали следующее:
Теперь мы включаем данные для фенотипа конечной стадии после 1-2-дневного и 0-3-дневного воздействия PD (Рисунок 5 — рисунок в приложении 3C), которые показывают задержку регенерации.Мы также подвергали животных воздействию PD или альтернативного ингибитора MEK U0126 в течение 24-часовых интервалов от 0-1, 1-2 или 2-3 дней после ампутации глотки и анализировали количество предшественников глотки. Эти эксперименты показывают, что ингибирование ERK между 1-2 днями после ампутации специфически влияет на увеличение числа предшественников глотки (Рисунок 5F, Рисунок 5 — приложение к рисунку 3B). Вместе эти эксперименты укрепили наш аргумент, что ERK необходим для продукции предшественников глотки и регенерации глотки.
Мы изменили наш язык, чтобы называть лекарственные ингибиторы ингибиторами MEK, но продолжаем ссылаться на ингибирование ERK, поскольку применение ингибиторов MEK действительно подавляет активность ERK (Owlarn et al., 2017) и фосфорилирование, как показано в нашем вестерн-блоте ( Рисунок 5 — приложение к рисунку 1B).
3. Интеграция с существующей литературой
Авторам необходимо лучше интегрировать свои выводы с литературой. Во-первых, они должны процитировать результаты Vu et al., которые явно продемонстрировали специфическое увеличение фракционной численности piwi-1 + / протонефридиальных маркерных + клеток в ответ на РНКи-опосредованное повреждение протонефридий (Vu et al., 2015). Таким образом, это исследование уже демонстрирует основную точку зрения Бора и др., А именно, что система планарийных стволовых клеток способна обеспечивать «сознательную мишень» для обеспечения предшественников. По крайней мере, авторы должны считать эти результаты дополнительным доказательством своей модели. Еще один вывод, который они должны обсудить, — это демонстрация LoCascio et al.(LoCascio et al., 2017), что вырезы в боковых областях вызывают значительное увеличение включения клеток глотки по сравнению с исходным уровнем, несмотря на отсутствие повреждения глотки. Как авторы согласовывают несоответствие между этими данными и своими собственными? В общем, обсуждение значительно выиграет от более подробного сравнения между «целевой слепой» моделью и их данными, а также от более широкого взгляда на регуляцию гомеостаза стволовых клеток.
Мы благодарим рецензента за это предложение и приносим свои извинения за оплошность работы наших коллег.В пересмотренной версии мы более подробно описали и контекстуализировали наши выводы с существующей литературой. Мы добавили разделы во введении, обсуждении и рисунках, в которых конкретно рассматривается контраст между существующими моделями регенерации (которые в нашей рукописи называются целевыми и нецелевыми).
Мы также включили эксперименты, анализирующие FoxA + piwi-1 + клеток после резекции фланга (рис. 2D), но не наблюдали никакого увеличения.Мы согласовали этот вывод с существующей литературой, выполнив маркировку EdU после различных ампутаций, несколько похожих на те, что выполнялись в (LoCascio et al.2017), но в разное время. Эти эксперименты показывают, что целевой механизм направляет клетки к отсутствующим тканям через день после ампутации (рис. 1E, F), но что нецелевой механизм также способствует регенерации, возможно, если небольшое количество тканей было повреждено (например, резекция глаза). В первом разделе обсуждения подробно описаны эти две модели и то, как они могут быть использованы для регенерации планарий.
https://doi.org/10.7554/eLife.68830.sa2Свежий взгляд на старые зоонозы: чего нам не хватало в одном исследовании и образовании в области здравоохранения?
Пандемии COVID-19, в которых в настоящее время погибло более 100000 человек, напомнили, что здоровье человека неразрывно связано со здоровьем животных и окружающей среды. Большинство инфекционных заболеваний человека являются энзоотическими в популяциях животных, и передача является естественным продуктом их взаимоотношений…
Пандемии COVID-19, в которых в настоящее время погибло более 100000 человек, напомнили, что здоровье человека неразрывно связано со здоровьем животных и окружающей среды. Большинство инфекционных заболеваний человека являются энзоотическими в популяциях животных, и передача является естественным продуктом их взаимоотношений во взаимосвязанных средах. Возникающие зоонозы представляют собой растущую угрозу для здоровья населения мира, причинив за последние 20 лет экономический ущерб в сотни миллиардов долларов США.Однако наибольшее бремя для здоровья и средств к существованию человека несут эндемические зоонозы, которые представляют собой постоянные региональные проблемы здравоохранения во всем мире.
Борьба с этой группой болезней требует совместных межотраслевых усилий систем здравоохранения и здоровья человека и животных и мультидисциплинарного подхода, учитывающего сложности экосистем, в которых сосуществуют люди и животные, — идея, описываемая как перспектива единого здоровья.
Цель этого исследования — показать, насколько новые и эндемичные зоонозы становятся все более актуальными для мирового медицинского сообщества.В частности, эндемические зоонозы имеют большое значение как в промышленно развитых странах, так и во многих странах с низким и средним уровнем доходов, где они могут или не могут быть адекватно диагностированы или зарегистрированы, что вызывает неопределенность в официальных данных.
Особое внимание будет уделяться «забытым» зоонозным болезням, поражающим бедные и маргинализированные группы населения в условиях ограниченных ресурсов. Для этих болезней, бремя которых оказалось трудно оценить, практически невозможно оценить реальное воздействие на социальное благополучие и психическое здоровье затронутых сообществ, владельцев скота и их семей.В частности, в специальном выпуске содержится призыв к проведению исследований на стыке людей / животных / окружающей среды, чтобы лучше понять, влияют ли зоонозы на популяции в развивающихся или развитых странах, с особым упором на культурные обычаи и социальное воздействие.
Для эффективного решения очень сложных проблем, связанных с профилактикой зоонозов и борьбой с ними, необходимо внедрить междисциплинарный и трансдисциплинарный подход, включая медицину человека и ветеринарию, традиционное общественное здравоохранение, безопасность пищевых продуктов, санитарию и гигиену.Однако необходимо участие других областей, включая социальные науки, экономику, системы производства продуктов питания, устойчивые методы, управление отходами и здоровье экосистем.
Тема исследования будет направлена, в частности, на исследования, проводимые в рамках многосекторального сотрудничества, включая клиницистов, ученых в области общественного здравоохранения, экологов, ветеринаров и экономистов, для углубленного изучения причин зоонозных заболеваний и основных проблем, которые способствуют увеличению передачи патогенов среди людей. , дикая природа и домашний скот.Также приветствуются обзорные статьи по этим темам.
Руководство «Единое здоровье» очень необходимо для эффективной координации самых разных секторов и дисциплин, способных выявлять потенциальные риски, связанные с зоонозами, а также согласовывать и координировать действия по профилактике и контролю, а также выработку политики.
Таким образом, роль университетов имеет решающее значение, и многие из них уже участвуют в разработке инициатив и образовательных программ. Эти мероприятия проводятся для обеспечения разносторонних кадров здравоохранения, чтобы лучше подготовиться и реагировать на существующие и возникающие экологические и эпидемиологические кризисы.
Еще одной целью этого специального выпуска будет сбор материалов, которые будут стимулировать обсуждение роли академических кругов в образовании, исследованиях и передаче знаний «Единое здоровье» для укрепления и защиты здоровья человека, животных и окружающей среды.
Ключевые слова : зоонозы, социально-экономические последствия, общественное здравоохранение, образование, единое здоровье
Важное примечание : Все вклады в эту тему исследования должны быть в рамках раздела и журнала, в который они были отправлены, как это определено в их заявлениях о миссии.Frontiers оставляет за собой право направить рукопись за пределами области охвата в более подходящий раздел или журнал на любом этапе рецензирования.
Информация об утерянных и найденных вещах — Служба для животных
Вы скучаете по своему питомцу?
Щелкните ссылку ниже, чтобы увидеть фотографии животных, которые в настоящее время находятся в нашем приюте.
Мы стараемся сфотографировать каждое животное, но иногда не можем сделать снимок, например, если животное травмировано или агрессивно.
Мы также ведем список всех животных, которых забрали умершими.
Если вы скучаете по своему питомцу, единственный способ гарантировать, что вашего питомца здесь нет, — это как можно скорее лично посетить приют!
Найдите среди наших животных своего питомца
Моего питомца забрала служба контроля животных. Как мне его получить?
Изъятые животные будут возвращены лицу старше 18 лет, которое может предоставить документы, удостоверяющие личность, и владение животными.Взимается плата за выкуп; мы принимаем наличные, кредитные карты и чеки. Обратите внимание, что мы не можем указывать комиссию по телефону. Пожалуйста, принесите действующее действительное удостоверение личности с фотографией и одно или несколько из следующего, чтобы подтвердить право собственности:
- Место и дата, когда животное в последний раз видели;
- Возраст и пол животного;
- Актуальные фотографии питомца;
- Ветеринарные записи на домашнее животное, включая доказательства вакцинации и стерилизации;
- Информация о микрочипе
Что делать, если мое домашнее животное пропало?
НИКОГДА НЕ ПРИНИМАЙТЕ , что ваш питомец был украден или вернется сам.
ПОСЕТИТЕ местных приютов для животных в течение 24 часов после потери вашего питомца.
Попросите проверить питомники ISOLATION и QUARANTINE при посещении приютов. Также спросите о любом животном, которое могло быть подобрано мертвым.
POST Карты утерянных домашних животных или листовки на досках потерянных и найденных животных в местных приютах, ветеринарных клиниках и других общественных досках объявлений, если таковые имеются. Публикуйте сообщение в Юте о потерянных и найденных домашних животных на Facebook.Также разместите сообщение в приложении NextDoor и в Finding Rover.
СМОТРИТЕ для своего питомца днем И НОЧЬЮ .
ПОЗВОНИТЕ в ближайший ветеринарный офис, чтобы узнать, не попал ли ваш питомец как раненый бездомный.
МЕСТО объявлений о потерянных домашних животных в местных газетах и секретных разделах веб-сайтов в столбцах потерянных и найденных, таких как KSL и / или Craigslist.
ПРОВЕРИТЬ все укрытия в районе. Ваш питомец может оказаться на любом из них.
НЕ СДАВАЙТЕСЬ! Часто домашнее животное не попадает в приют в течение нескольких недель, а иногда и месяцев после того, как впервые потерялось.
Поддерживайте актуальность идентификационных меток и микрочипируйте своего питомца!
Приоритет животных для плотного генотипирования с целью вменения недостающих генотипов редко генотипированных животных | Genetics Selection Evolution
Data
Использовалась родословная свиней породы Ландрас, предоставленная NORSVIN AS [http://www.norsvin.no]. Всего насчитывалось 13 276 голов свиней, в том числе 619 производителей, всего 12 поколений.2500 недавно родившихся животных последнего поколения, все без потомков в наборе данных, были выбраны в качестве целевой популяции, по которой будет оцениваться условное исчисление. Таким образом, группы из свиней Т были отобраны из всей популяции на основании различных критериев плотного генотипирования (или полногеномного секвенирования). Успех вменения был измерен в целевой популяции, которая была редко генотипирована. Для оценки точности геномного отбора в целевой популяции 2000 животных из целевой популяции были случайным образом выбраны в качестве обучающей выборки, для которой были зарегистрированы как фенотипы, так и условные генотипы, а оставшиеся 500 были обработаны как проверочная выборка.Были оценены GEBV, и точность GEBV была оценена как корреляция между GEBV и истинной племенной ценностью.
Отбор животных для плотного генотипирования
Для отбора наборов свиней для плотного генотипирования из всей родословной свиней, включая целевую популяцию, использовалось семь методов. Предполагалось, что бюджет, необходимый для плотного генотипирования особей T , был доступен и что все целевые животные для тестирования вменения были редко генотипированы с плотностью D.Т был равен 100 или 200, а D был равен 50, 100 или 200 маркерам на Морган. Ниже описаны семь методов. Два из описанных ниже методов имеют оптимальные свойства для задачи, основанные на минимизации генетической дисперсии целевой популяции, обусловленной выбранным набором, с использованием либо отношений, полученных из матрицы отношений числителя Райта, A (MCA), либо матрицы геномных отношений. , G (MCG), полученный в результате разреженного генотипирования.Эти критерии минимизируют среднеквадратичную ошибку непредвзятого предиктора вмененного генотипа на основе полной информации о генотипе в выбранном наборе. Остальные пять методов являются эвристическими (KIN, REL, CON, SRS) или случайными (RAN).
Для метода MCA предполагается, что единственная доступная информация о родственных связях до плотного генотипирования получена из родословной. Матрица дисперсии-ковариации для подсчета эталонного аллеля в любом локусе родословной пропорциональна матрице отношений числителя Райта, A .Матрица вариации-ковариации, обусловленная отбором набора животных для плотного генотипирования (но до получения каких-либо генотипов), задается следующим образом: A11 * = A11 ‒ A12A22‒1A21, где жирный индекс 1 обозначает набор целевых животных, а жирный нижний индекс 2 обозначает набор животных с плотным генотипом. Так, например, A 11 представляет собой подматрицу родословных отношений между целевыми животными. Условные дисперсии, diagA11 *, представляют собой остаточные дисперсии, которые, как ожидается, сохранятся, если из выбранного набора будут получены плотные генотипы и использоваться для прогнозирования генотипов целевого набора.Следовательно, необходимо минимизировать именно diagA11 *, и использованная сводная статистика была , трассировка A11 *. Эта минимизация для отбора животных с плотным генотипом была проведена с использованием итеративной процедуры: сначала было выбрано животное, которое наиболее уменьшило след A11 *, то есть животное i , которое максимизировало след ( A 1 и A i 1 / A ii ), где A 1 и — вектор отношений животного i с целевым набором животных, A i 1 — его транспонирование, а A ii — это отношение животного и к самому себе.После выбора животного i вся матрица отношений была обусловлена генотипом животного i , A (1) = A — A : i A i : / A ii , где A : i — вектор отношений i со всеми особями в родословной. A (1) будет иметь нули в строке и столбце, соответствующих выбранному человеку i .Затем был выбран следующий человек (новый i ), чтобы максимизировать след A1i1Ai11 / Aii1, и отношения всей родословной были обусловлены тем, что этот второй человек даст A (2) , где A2 = A1 -A: i1Ai: 1 / Aii1. Последующий выбор продолжился с использованием A (2) . Эта итеративная процедура продолжалась до тех пор, пока не было отобрано T животных для плотного генотипирования. Точное решение потребует поиска по всем подмножествам размера T .Предлагаемый алгоритм является более быстрым в вычислительном отношении и предполагает, что набор, оптимальный для размера T , будет содержаться в наборе, оптимальном для размера T + 1.
Для метода MCG предполагалось, что разреженные генотипы целевые животные были доступны до выбора животных для плотного генотипирования. Это позволило заменить матрицу отношений числителя A матрицей геномных отношений G на основе разреженного генотипирования. G был построен с использованием метода FG, описанного в [8]: во-первых, вероятности генотипов были рассчитаны с помощью LDMIP [9], используя только варианты анализа сцепления, а матрица отношений G была рассчитана в соответствии с принципами [10], усреднение по всем позициям маркеров. Затем были отобраны животные-кандидаты, как для MCA, за исключением того, что A был заменен на G . Метод MCG был протестирован для каждой из плотностей разреженных маркеров (50, 100 и 200 маркеров на Морган), которые обозначены MCG-50, MCG-100 и MCG-200, соответственно.
Четыре из оставшихся методов являются эвристическими. Для KIN были отобраны T животных в родословной, которые имеют самые высокие средние коэффициенты родства с животными целевой популяции. Таким образом, выбранные T животных максимизируют 1 T A 12 1 , где 1 обозначает вектор-столбец подходящего размера из единиц. REL [11] отличается от KIN тем, что он выбирает животных T , которые максимизируют сумму вариативных коэффициентов регрессии T- количества аллелей для целевого набора на таковых для выбранного набора1TA22-1A211.(Эвристическое обоснование REL, приведенное в [11] для максимизации отношений между отобранными и целевыми животными, на самом деле больше подходит для KIN). Для SRS были отобраны особей T , которые имеют наибольшее количество сыновей и дочерей в родословной. Для CON были отобраны животные T , которые имели наибольший генетический вклад по происхождению в целевую популяцию, для которых вклады были получены от L в разложении LDL T матрицы A , как описано [12].Наконец, для RAN была отобрана случайная выборка из T животных из всей родословной свиней без замены.
Моделирование генотипов и фенотипов
Прямое моделирование [http://ihaiwtheoserv.umb.no/tools/xform/xform.tar.gz] использовалось для моделирования идеальных популяций, в которых мутации SNP накапливались в результате случайного спаривания поколений. спонтанными мутациями и рекомбинациями. Эффективные размеры ( N e ) идеальной популяции составляли 100 и 200, а частота мутаций составляла 10 — 8 на пару оснований на мейоз.После 10 000 поколений случайного спаривания смоделированные генотипы SNP последнего поколения были переданы через основателей родословной Ландрас в популяцию путем отбрасывания генов. Количество сгенерированных маркерных локусов было равно ~ 2200 локусов / Морган для N e = 100 и ~ 4900 локусов / Морган для N e = 200. Эти числа близки к ожидаемым результатам [13], который сообщил о 2120 локусах для N e = 100 и 4790 для N e = 200.Среди смоделированных локусов доли 0,47 и 0,52 имели частоту минорных аллелей (MAF) менее 0,05 для N e = 100 и 200 соответственно. Дальнейшее снижение затрат на вычисления было получено путем моделирования только одной хромосомы размером 1 Морган (M), что не повлияет на точность вменения.
Для проверки точности геномной оценки 30 аддитивных QTL были случайным образом выбраны из сегрегационных локусов, игнорируя MAF.Аллельные эффекты в QTL следовали распределению Лапласа со средним значением 0 и масштабным параметром 1. Затем индивидуальные фенотипы были смоделированы с наследуемостью 0,02 путем добавления условий ошибки NIID к племенной ценности индивидуума, которая была суммой аллельных эффектов в QTL. . Хотя на точность вменения генотипа не влияет количество смоделированных хромосом и наследуемость признака, это влияет на геномную оценку. Использование низкой наследуемости компенсирует небольшой размер генома 1 M, согласно концепции, согласно которой точность геномного предсказания определяется h 2 / Me , где Me — эффективное количество независимых сегментов в геном [14].
Вменение генотипа и оценка GEBV
Для моделирования разреженного генотипирования из всех доступных локусов, включая те, которые были отобраны для QTL, случайным образом были отобраны D = 50, 100 или 200 локусов. Минимальный MAF для выбранных маркерных локусов был равен 0, 0,05 или 0,10, и это применялось к отбору локусов, предположительно находящихся на «разреженном чипе», и к отбору локусов на «плотном чипе», который использовался на T животных, используемых для обучения, и, следовательно, по генотипам, подлежащим условному исчислению.Генотипы в невыбранных локусах в редком наборе затем были реконструированы с использованием (i) Beagle [15] или (ii) LDMIP [9], который, в отличие от Beagle, использует доступную информацию о родословных. Генотипы оценивали 0, 1 или 2, что соответствует количеству мутантных аллелей по сравнению с референсными аллелями. Частота ошибок рассчитывалась как частота неверных вмененных генотипов, усредненная по всем локусам и целевым животным. Корреляция между вмененными генотипами (с оценкой 0, 1 и 2) и истинным генотипом среди животных рассчитывалась для каждого локуса, а затем усреднялась по всем локусам.Было показано, что корреляция гораздо меньше зависит от MAF, чем от частоты ошибок [16].
Для оценки GEBV использовались два метода геномной селекции: MixP [17] и GBLUP (Геномное наилучшее линейное несмещенное предсказание) [13]. Сообщается, что GBLUP надежен при анализе реальных данных, но он меньше выигрывает от увеличения плотности маркеров и безразличен к архитектуре QTL. Метод MixP соответствует смеси двух нормальных распределений для эффектов SNP, аналогично BayesC [18], и, таким образом, пытается придать дополнительный вес важным маркерам и не придавать веса другим, сохраняя при этом затраты на вычисления на уровне, сопоставимом с GBLUP.Были зарегистрированы точности GEBV от двух методов, точность определялась как коэффициент корреляции между истинными смоделированными значениями разведения и GEBV.
Дизайн и репликация
Методы RAN, KIN, CON, SRS и MCA сравнивались для всех комбинаций трех значений D , трех пороговых значений MAF, двух значений T и двух значений Ne. В этом полном факторном дизайне все методы были воспроизведены 1000 раз, и вменение производилось с помощью Beagle.Для MCG, из-за некоторых вариаций между повторностями, вычислительная нагрузка была намного больше, и поэтому сравнения для MCG были ограничены всеми тремя значениями: D с Ne = 100, T = 200 и с MAF больше. чем 0,05. В этих случаях было выполнено минимум 400 повторов, что было достаточно большим, чтобы стандартные ошибки оставались небольшими по сравнению с эффектами. Сравнение метода REL с другими методами с использованием Beagle было ограничено 400 повторами T = 100, D = 100, MAF> 0.05 и Ne только = 100. Сравнение всех семи методов с использованием LDMIP для вменения, а не Beagle, также было ограничено 400 повторениями T = 100, D = 100, MAF ≥ 0,05 и Ne = 100.
Места, где пропавшие кошки были найдены (а) Кошачьим любопытством / клоуном …
Домашние кошки являются одними из самых популярных домашних животных. Из-за того, что они свободно перемещаются по природе, они могут потеряться и разлучиться со своими владельцами (Lord et al., 2007). То, что это реальная проблема, подтверждается наблюдением, согласно которому около 15% владельцев кошек сообщили о потере своего питомца за последние пять лет (Weiss et al., 2012). Разлучение со своим домашним животным может иметь сильное эмоциональное воздействие на владельцев, но также может обременять приюты для животных, где эти животные могут оказаться. Помимо того, что они свободно бродят, кошки теряются или не возвращаются домой по нескольким причинам. Частая причина — дорожно-транспортные происшествия (Wilson et al., 2017). Другие виды травм могут возникнуть в результате падений и нападений собак (Hernon et al., 2018). Глубокие изменения в их среде обитания (например, появление собаки) также иногда упоминаются как факторы риска того, что кошки не вернутся домой. Чтобы снизить риск потери кошки, можно предпринять несколько действий. Определенные меры могут помочь идентифицировать найденных кошек. Основными примерами таких мер являются микрочипирование и идентификационные метки (Lord et al., 2010; Weiss et al., 2016). Для поиска пропавшей кошки используются и другие методы, например поиск в районе, где животное видели в последний раз (Weiss et al., 2012). Социальные сети также используются для воссоединения владельцев с их потерянными домашними животными, как в небольших масштабах (например, распространение сообщений среди друзей), так и в специальных сообществах (например, в группах Facebook). Менее распространенная, но, возможно, очень эффективная стратегия — это локализация с помощью трекеров. Эти устройства обычно прикрепляются к ошейнику животного и обеспечивают местоположение животного с помощью ряда возможных средств, включая сигналы GPS. Однако есть и определенные недостатки. Трекеры могут быть довольно большими и часто требуют платной подписки.Кроме того, кошки могут застрять в ошейнике или потерять ошейники, к которым прикреплены трекеры, если эти ошейники имеют предохранительную разблокировку. Помимо этих мер, первым рефлексом должно быть начало осмотра и ухода за явно потерявшимися или ранеными кошками. Степень успеха воссоединения кошек с их хозяином не совсем ясна. Процент кошек, которые в конечном итоге обнаруживаются, колеблется в пределах 53-75% в литературе (Lord et al., 2007; Weiss et al., 2012; Huang et al., 2018). Можно ожидать, что этот процент сильно зависит от нескольких факторов, таких как используемые методы поиска и место, где пропало животное.Лучшее понимание степени успеха различных стратегий может стать началом для воссоединения большего числа владельцев со своими питомцами. Количество литературы о факторах, окружающих потерю кошек или стратегиях локализации, очень ограничено. Систематический обзор может предоставить структурированный обзор текущих знаний и указать направление для дальнейших исследований. Последующее когортное исследование могло бы выявить факторы риска, которые связаны с более высоким риском потери кошек, и определить наиболее эффективные методы поиска потерявшихся кошек.Из-за характера проблемы проспективные интервенционные исследования неосуществимы. Ссылки Hernon T, Gurney M, Gibson S. Ретроспективное исследование кошачьих пациентов с травмами, поступивших в специализированный центр. J Small Anim Pract. 2018; 59 (4): 243-7.
Отсутствуют 3R: исследования на животных, не представляющие научной ценности, неэтичны
Введение
Созданные Расселом и Берчем более 60 лет назад, 3R ( R eplacement, R eduction и R efinement) стали руководящими принципы этичного использования животных в исследованиях.1 Несмотря на всеобщее признание, продолжается дискуссия об их улучшении, внедрении и внедрении.2 Здесь мы утверждаем, что с их нынешним акцентом на благополучие животных, 3R не имеют важного этического аспекта. Исследования на животных этичны только в том случае, если они приносят пользу науке и обществу, а это измерение не отражено в нынешних трех принципах.
Отдельные исследовательские проекты имеют ценность только в том случае, если они позволяют получить знания, применяют надежные планы исследований и неизбирательно сообщают о своих результатах.Трудно оценить, будет ли исследовательский проект в конечном итоге способствовать инновациям в здравоохранении по нескольким причинам. Одна из причин заключается в том, что научные открытия могут проявиться годами. С другой стороны, об устойчивости можно судить на уровне исследовательского проекта. Если мы хотим лучше понять, какие вопросы исследований все еще недостаточно решены, нам нужно, чтобы отдельные проекты были доступны через реестры исследований на животных, открытые для общественности. Кроме того, только в случае проспективной регистрации протоколов мы можем идентифицировать выборочную отчетность о результатах исследования.
Мы утверждаем, что, хотя существующие 3R важны для обеспечения благополучия животных, при планировании, анализе и проведении исследований на животных необходимо учитывать научную ценность. Поэтому мы предлагаем добавить три дополнительных Rs, то есть R устойчивость, R регистрация и R eporting, к руководящим принципам этического использования животных в исследованиях (рисунок 1).
Рисунок 1Два основных принципа этики исследований на животных трансформируются в шесть практических руководящих принципов (6R).
Почему нам нужно дополнять фреймворк 3R сейчас?
За последние 5 лет несколько эмпирических исследований и экспертных анализов показали, что три проблемы ставят под угрозу ценность исследований на животных. Во-первых, в исследованиях на животных часто не хватает мер по снижению угроз достоверности, таких как предвзятость или отсутствие статистической мощности.3 4 Во-вторых, исследования на животных сталкиваются с существенной предвзятостью публикации, то есть нулевые и отрицательные результаты часто оказываются в ящике файлов.5 6 В-третьих, в публикации результатов часто не хватает важной информации, необходимой для критической оценки (например, информации о дизайне исследования или истощении животных).7 8 Эти проблемы отрицательно влияют на воспроизводимость исследований на животных 9 10 и актуальность исследований на животных для обоснования ранних исследований на людях 11 12 В целом, эти угрозы снижают ценность результатов исследований, потенциально приводя к неэффективному распределению государственных средств на нужды больных. -рекомендованные клинические исследования и ненужное использование и страдания экспериментальных животных.
Почему надежность, регистрация и отчетность?
Наш основной аргумент состоит в том, что существующие принципы 3R для исследований на животных, несмотря на их важность, ограничены из-за их односторонней ориентации на базовый этический принцип «благополучие животных».Им не хватает четкого и ориентированного на практику набора руководящих принципов, продвигающих второй базовый этический принцип «научная ценность». Кроме того, каждый из дополнительных принципов 3R (надежность, регистрация и отчетность) важен сам по себе и не может быть заменен двумя другими. Например, исследования на животных могут быть надежными, но сообщаться необъективно или иным образом некорректно. В качестве альтернативы о них можно надлежащим образом сообщать, но они не являются надежными. Оба сценария ставят под угрозу ценность исследования.Во времена, когда не сообщается примерно о 50% исследований на животных 13, только предварительная регистрация протоколов исследований на животных позволяет выявить предвзятые, отсроченные или незарегистрированные результаты. Наконец, этические рамки человеческих исследований уже затрагивают все три ценностных принципа по одним и тем же моральным причинам. Хельсинкская декларация, например, включает регистрацию (статья 35) и отчетность (статья 36) в качестве обязательных принципов14. Широко признанная система клинических исследований «Что делает клинические исследования этичными» из Эмануэля и др. подчеркивает надежность (научную обоснованность). ) как один из основных этических принципов.15
Как внедрить новые 3R в текущую практику?
Принцип отчетности относительно легко реализовать. Помимо стандартных рецензируемых журналов, новые форматы публикаций обеспечивают доступную отчетность обо всех типах результатов исследований, включая нулевые и отрицательные результаты, такие как серверы препринтов (например, bioRxiv), журналы открытого доступа (например, BMJ Open Science, PLoS One), журналы с обзором после публикации (например, f1000research) или репозиториями данных (например, Open Science Framework, Dryad, figshare).Соблюдение руководящих принципов отчетности, таких как ARRIVE, 7, также направлено на улучшение оценки и использования результатов исследования. Несколько ведущих спонсоров исследований, такие как Wellcome Trust, программа Horizon 2020 или Фонд Билла и Мелинды Гейтс совсем недавно подписали Совместное заявление ВОЗ и тем самым указали, что требования к отчетности должны быть частью решений о финансировании клинических испытаний16. финансирование отдельных исследований на животных могло бы обеспечить выполнение требования о своевременной и неизбирательной отчетности о результатах и оценке соблюдения.
Специальные инструменты для реализации принципа регистрации в исследованиях на животных, эквивалентные регистрам для исследований на людях (например, ClinicalTrials.gov), уже были запущены академическими инициативами (например, www.preclinicaltrials.eu) или совсем недавно правительственной организацией (www .animalstudyregistry.org). Эти платформы позволяют быструю регистрацию протокола с ограниченным вариантом в течение нескольких лет. Принцип регистрации повысит ценность исследований, но как это повлияет на эффективность исследований на животных? В недавнем исследовании эксперты из всех соответствующих групп заинтересованных сторон в исследованиях на животных выразили свое отношение к потенциальным сильным и слабым сторонам реестров исследований на животных.17 Некоторые выразили обеспокоенность тем, что регистрация исследований на животных может усугубить административное бремя и кражу идей. Другие подчеркнули противоположную точку зрения, согласно которой повышение прозрачности с помощью таких реестров может в конечном итоге сделать исследования на животных более эффективными.
Принцип устойчивости сложнее реализовать: как мы можем оценить надежность отдельных исследований на животных? Более конкретно: когда необходим расчет размера выборки или слепая оценка результатов? Как можно улучшить внешнюю и конструктивную валидность отдельных исследований? Недавние предложения экспертов о том, чтобы лучше различать исследовательские и подтверждающие исследования в исследованиях на животных, дали предварительные ответы.18 19 Первоначальное руководство по внедрению более систематической оценки устойчивости исследований на животных в стандартные процедуры обзора было недавно опубликовано Würbel.20 Würbel различает три аспекта достоверности (внутренняя, внешняя и конструктивная достоверность) и рекомендует оценивать каждый параметр в пределах вреда — анализ преимуществ для отдельных исследований на животных. Этим предложением он соответствует недавнему руководству Киммельмана о том, как оценивать достоверность исследований на животных в рамках процедур утверждения клинических испытаний фазы I / II.21 Оценка надежности отдельных исследований требует сложных суждений. Однако комиссии по этике исследований на животных уже требуют сложных суждений в отношении принципов благополучия, и во многих юрисдикциях уже принимают во внимание надежность исследования. Даже Рассел и Берч уже включили раздел «Планирование и анализ экспериментов» в главу, объясняющую принцип редукции.1 Они подчеркивают важность статистики для определения минимального количества животных, необходимого для эксперимента, и упоминают последовательный анализ и рандомизацию. как дополнительное средство для уменьшения неконтролируемой дисперсии.Однако в них не подчеркивается надежность или научная ценность как сам по себе принцип, и в них не упоминаются дальнейшие меры по повышению надежности, такие как слепая оценка результатов.
В соответствии с нашей рекомендацией добавить руководящие принципы научной ценности в этические рамки для исследований на животных, недавние мероприятия национальных центров для 3R, таких как Национальный центр улучшения замещения и сокращения животных в исследованиях (NC3R) или Немецкий центр защиты животных в исследованиях (Bf3R).Оба они уже продвигают новые принципы 3R для научной ценности несколькими способами. Пересмотренные руководящие принципы NC3Rs для исследований приматов, например, явно требуют надежности и отчетности22. Новый помощник по экспериментальному проектированию NC3Rs (EDA) не только поддерживает разработку надежных протоколов исследования, но также позволяет устанавливать временные метки для итоговых протоколов. Имея возможность сделать такие протоколы с отметками времени общедоступными, EDA упрощает предварительную регистрацию протоколов на добровольной основе.23 В январе 2019 года Bf3R запустила свой Реестр исследований на животных.Мы очень приветствуем эти недавние разработки, но хотим подчеркнуть, что они не вытекают напрямую из какого-либо из трех принципов защиты животных. Они имеют смысл только при рассмотрении научной ценности как дополнительного набора этических принципов.
«Румба рупий»?
В предыдущих разделах мы уже комментировали возможные контраргументы против введения дополнительного набора принципов 3R. Эти контраргументы напрямую касались актуальности или реализуемости регистрации, надежности или отчетности.Другой тип контраргумента более косвенный: имеет ли вообще смысл добавлять новые принципы R? В наших обсуждениях с коллегами и рецензентами были выдвинуты по крайней мере два аргумента: во-первых, в других статьях уже безуспешно предлагались новые R, такие как ответственность, воспроизводимость или строгость. Эти взносы не повлияли на исследования на животных, а, скорее, накалили румбу 24 рупий. Мы думаем, что прежние предложения о новых рупиях не увенчались успехом, потому что они были круглыми, слишком широкими или не содержали прямых указаний.Ответственность как принцип R явно циклична, так как не может указывать, как действовать ответственно. Воспроизводимость как принцип R не дает прямого указания. Это желаемая характеристика исследований на животных, которая сильно зависит от надежности и неизбирательной отчетности. Строгость как принцип R слишком широк, по крайней мере, в его нынешнем использовании. Строгость часто используется как синоним научного значения, поскольку она включает надежность, неизбирательную отчетность, а также может включать регистрацию.
Второй контраргумент против любой модификации структуры 3R основан на предположении, что текущая структура 3R является сильной концепцией, особенно потому, что она принята во всем мире.Добавление новых рупий несет в себе риск ослабить эту широко распространенную концепцию, что в конечном итоге приведет к более слабой защите благополучия животных. Однако мы не считаем правдоподобным полагать, что последовательный набор из трех новых руководящих принципов, которые все сосредоточены на дополнительном базовом принципе научной ценности, разбавит очень отчетливый базовый принцип благополучия животных. Напротив, мы утверждаем, что относительно узкая направленность нынешнего подхода 3R способствовала тому факту, что исследования на животных часто не имеют научной ценности.
Резюме
Исследования на животных этичны только тогда, когда они имеют научную и социальную ценность. Последние годы продемонстрировали, что ценность исследований на животных и, следовательно, их способность улучшать здоровье человека находятся под угрозой из-за отсутствия надежности и предвзятых или незарегистрированных результатов. Три этических принципа (устойчивость R , регистрация R и транспортировка R ) помогают сохранить ценность исследований на животных. Нынешняя широко распространенная этическая основа для исследований на животных (3R = R замена, R обучение и R улучшение) упускают этот ценностный аспект, поскольку сосредоточены исключительно на столь же важном аспекте благополучия животных.Мы рекомендуем дополнить текущую схему 3R (для благополучия животных) вторым набором 3R (для научной ценности). Регулирующие органы, советы по этике, ученые и спонсоры должны добавить надежность, регистрацию и отчетность к своим критериям при планировании, лицензировании или финансировании экспериментов на животных. В таких руководствах, как Базельская декларация, следует рассмотреть вопрос о том, чтобы сделать нормативную базу для исследований на животных более всеобъемлющей и согласованной.25 Национальным центрам 3R следует рассмотреть возможность пересмотра своей торговой марки и четкого рассмотрения этических соображений, лежащих в основе их недавней политики регистрации, надежности и отчетности.С этой целью требуется обсуждение и принятие решений с участием многих заинтересованных сторон, чтобы (1) концептуализировать особенности устойчивости, (2) разработать рамки, детализирующие обязательную информацию, которая регистрируется, а также допустимые периоды эмбарго, (3) уточнить требования к финансированию и утверждению, связанные с для отчетности о результатах и (4) определить соответствующие обязанности, компетенцию и законодательство для аудита соответствия 6R.