
Оптико кинетические средства общения это: 2. Оптико — кинетические средства
|
 Женщины к тому же склонны на больший визуальный контакт, чем мужчины. Обычно люди избегают визуального контакта в ситуациях соперничества, чтобы этот контакт не был понят как выражение враждебности. Кроме того, мы склонны смотреть на говорящего больше, когда он находится на расстоянии: чем ближе мы к говорящему, тем больше избегаем визуального контакта. Обычно визуальный контакт помогает говорящему почувствовать, что он общается с Вами, и произвести благоприятное впечатление. Но пристальный взгляд обычно создает о нас неблагоприятное впечатление.
Женщины к тому же склонны на больший визуальный контакт, чем мужчины. Обычно люди избегают визуального контакта в ситуациях соперничества, чтобы этот контакт не был понят как выражение враждебности. Кроме того, мы склонны смотреть на говорящего больше, когда он находится на расстоянии: чем ближе мы к говорящему, тем больше избегаем визуального контакта. Обычно визуальный контакт помогает говорящему почувствовать, что он общается с Вами, и произвести благоприятное впечатление. Но пристальный взгляд обычно создает о нас неблагоприятное впечатление. краткое описание, особенности и интересные факты
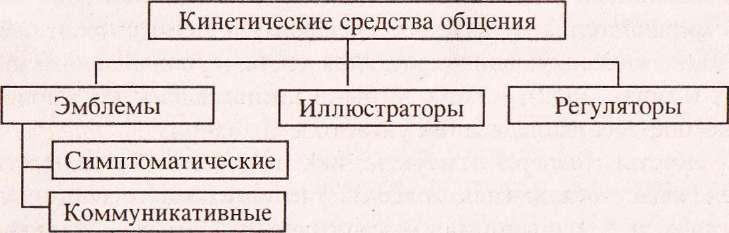
Кинетические средства общения представляют собой систему выражения эмоционального состояния собеседника. Чувства выражаются не словами, а посредством жестов и мимики. Как правило, люди сами за собой не замечают, как меняют положение тела, дотрагиваются до кончика носа или трут руками глаза. Но тот, кто умеет внимательно наблюдать и знает кратко кинетические средства общения, может делать соответствующие выводы. Очень часто оказывается, что люди говорят совсем не то, что думают. Многие используют слова, чтобы умело прятать свои настоящие чувства. Но чтобы снять эмоциональное напряжение, в ход идет невербальная система взаимодействия. Наука, которая изучает язык тела и жестов, называется кинетикой.
Чувства выражаются не словами, а посредством жестов и мимики. Как правило, люди сами за собой не замечают, как меняют положение тела, дотрагиваются до кончика носа или трут руками глаза. Но тот, кто умеет внимательно наблюдать и знает кратко кинетические средства общения, может делать соответствующие выводы. Очень часто оказывается, что люди говорят совсем не то, что думают. Многие используют слова, чтобы умело прятать свои настоящие чувства. Но чтобы снять эмоциональное напряжение, в ход идет невербальная система взаимодействия. Наука, которая изучает язык тела и жестов, называется кинетикой.
Изучая эти важные составляющие внеречевого общения, люди получают возможность наблюдать за поведением окружающих, анализировать их жесты и пантомимику, делать соответствующие выводы. Бывает так, что слова индивида расходятся с его мыслями и настроением. Многие предпочитают не выражать свои истинные чувства по той или иной причине, опасаясь насмешек и выраженного непонимания. Чаще всего они просто боятся негативной оценки общества, поэтому предпочитают молчать о своих истинных намерениях. Кинетические средства общения, их виды и характеристики будут рассмотрены в данной статье. Нужно обращать на них внимание, взаимодействуя с разными людьми.
Кинетические средства общения, их виды и характеристики будут рассмотрены в данной статье. Нужно обращать на них внимание, взаимодействуя с разными людьми.
Оптико-кинетическая подсистема
Представляет собой такие средства общения, которые влияют на понимание речи собеседника. Но даже если оппонент молчит и не произносит ни слова, мы все равно понимаем, какие чувства он испытывает по поводу той или иной ситуации. Оптико-кинетические средства общения – это те компоненты, которые складываются из позы и жестов. Люди подсознательно обращают на них внимание и делают соответствующие выводы. Мысли можно утаить от собеседника, но не выражение лица. Нас всегда будет выдавать тело.
Ни один человек не в состоянии полностью взять под контроль свои жесты. Оптико-кинетические средства общения – это довольно интересное направление в науке. Если всерьез заняться его изучением, то окажется, что можно научиться понимать друг друга без слов. В большинстве случаев люди даже до конца и не осознают, какие конкретные посылы передают друг другу, обмениваясь многозначительными фразами.
Внешний вид
Когда мы смотрим в глаза собеседнику, то обязательно представляем себе, что он чувствует в данную минуту. Задавая вопрос или ожидая ответа, можно наблюдать за психологическими реакциями, которые неизбежно проявляются в своем истинном свете. Внешний вид может рассказать о многом. Кинетические средства в деловом общении изучаются специально для того, чтобы определить настоящие чувства своего оппонента. Если человек пренебрегает своим внешним видом, то у него, как правило, низкая самооценка. Он бессознательно будет демонстрировать свое отношение к собственной личности через замкнутость, попытку одеваться как можно незаметнее. Внешний вид такого индивида часто отличается определенной неряшливостью. Бессознательно он стремится продемонстрировать людям, что не доверяет им, всячески отгораживается от них. Каждому народу и культуре свойственны свои определенные особенности. Например, испанцы и португальцы имеют привычку сильно жестикулировать и размахивать руками во время беседы. Оптико-кинетические средства общения англичан отличаются сдержанностью.
Оптико-кинетические средства общения англичан отличаются сдержанностью.
Мимика
Наблюдая за выражением лица, можно легко понять, что чувствует собеседник в данную минуту. Такое осознание, как правило, приходит интуитивно. Для этого достаточно просто внимательно наблюдать за тем, кто находится рядом. При этом не надо стремиться его перебивать или начинать в чем-то переубеждать. Мимика человека – вещь довольно конкретная. Она многое показывает сама, открывает в истинном свете. Оптико-кинетическая система невербальных средств общения – это язык, который невозможно подделать. Если в процессе разговора какое-то обстоятельство расстраивает собеседника, то он может не показать этого словами. Однако по выражению лица можно легко заметить произошедшую перемену: уголки губ опустятся вниз, человек начнет принимать позицию неудовольствия.
Многие предпочитают скрывать свои чувства и не выражать их вслух. Однако внимательный человек всегда заметит произошедшую перемену в психологическом состоянии. Мимика – кинетическое средство общения, которое сразу бросается в глаза. Общаясь с близким человеком, мы не можем не обращать внимания на эти явные сигналы его тела. Кинетические средства невербального общения помогают лучше понять чужую боль, настроение, прочувствовать общее эмоциональное расположение оппонента.
Мимика – кинетическое средство общения, которое сразу бросается в глаза. Общаясь с близким человеком, мы не можем не обращать внимания на эти явные сигналы его тела. Кинетические средства невербального общения помогают лучше понять чужую боль, настроение, прочувствовать общее эмоциональное расположение оппонента.
Позы и жесты
Даже если словесно человек никак не демонстрирует нам свое несогласие, это можно заметить по положению тела в пространстве. Неуверенность в себе заставляет занимать неудобную позицию, тесниться и испытывать всяческие ограничения. Поза человека при таких чувствах будет максимально закрытой, не располагающей к эффективному взаимодействию. Он просто не сможет находиться в радости и свободно вести себя. Чем больший внутренний дискомфорт человек испытывает, тем более неудобную для себя позу стремится занять.
Уверенная в себе личность держится раскованно. Такой человек много улыбается, не избегает смотреть в глаза своим собеседникам. Жесты демонстрируют расслабленность, комфорт и покой. В состоянии радости и морального удовлетворения индивид, как правило, перестает контролировать собственные движения. Он больше обращает внимание на других людей, следит за развитием предмета беседы. Можно сказать, что оптико-кинетические средства общения – презентация самого себя в социуме. А слова имеют уже второстепенное значение.
В состоянии радости и морального удовлетворения индивид, как правило, перестает контролировать собственные движения. Он больше обращает внимание на других людей, следит за развитием предмета беседы. Можно сказать, что оптико-кинетические средства общения – презентация самого себя в социуме. А слова имеют уже второстепенное значение.
Взгляд человека
Он имеет большое значение. По тому, как именно человек смотрит, его оценивают окружающие. Кинетические средства невербального общения могут о многом рассказать, помочь сделать определенные выводы.
Живой взгляд
Он обязательно привлекает к себе внимание, всячески располагает к дружескому участию. С доброжелательным человеком появляется желание общаться, поверять ему свои тайны и секреты. Живой взгляд выражает благосклонность, согласие, интерес. С его помощью индивид может передать собственное отношение к ситуации. Замечено, что, когда у человека доброе расположение, он внешне выглядит гораздо красивее и привлекательнее. В любом случае обращенный к людям взгляд лучше, чем хмурый и озлобленный. Чрезмерная погруженность в себя всегда отталкивает, заставляет переживать негативные чувства.
В любом случае обращенный к людям взгляд лучше, чем хмурый и озлобленный. Чрезмерная погруженность в себя всегда отталкивает, заставляет переживать негативные чувства.
Как правило, люди умеют с легкостью распознавать искренних и доброжелательных собеседников. Даже маленькие дети, которые только начинают свою жизнь, безошибочно определяют, кто к ним относится с большей теплотой и нежностью. Проникновенный взгляд во многом способствует установлению доверия. Улыбка обезоруживает, помогает начать разговор и направить беседу в нужное русло.
Безразличный взгляд
Вызывает отторжение у окружающих или такое же равнодушие в ответ. Безразличие – это то, что убивает любые искренние отношения. Человек, не выражающий никакого участия к окружающим, как правило, не вызывает ни доверия, ни теплоты в ответ. Равнодушие в конечном итоге приводит к изоляции и одиночеству. Такой индивид эмоционально никак не откликается на нужды других, а стремится решать только собственные проблемы. Он формирует в себе своеобразный вакуум, который не позволяет ему научиться понимать окружающих людей. Как правило, он ничего не делает для других просто так, без признания вторичной выгоды.
Как правило, он ничего не делает для других просто так, без признания вторичной выгоды.
Потухший взгляд
Как правило, выражает подавленность и боязнь действовать в том или ином направлении. Увидев человека с потухшим взором, мы сразу определяем его психологический настрой. Глядя на такое несчастное лицо, создается впечатление, что у него внутри кто-то выключил лампочки, столь необходимые для полноценной жизни. Поникшее настроение, общая подавленность препятствуют счастливому мироощущению, мешают начать воплощать в жизнь свои стремления и мечты. Такие люди, как правило, ни к чему не стремятся и не желают активно действовать для достижения желаемого результата.
Со стороны может показаться, что они абсолютно равнодушны ко всему происходящему, однако это не так. Просто есть что-то, определенная объективная причина, которая мешает осознавать себя самодостаточной и успешной личностью.
Задумчивый взгляд
Эмоциональная сосредоточенность заставляет индивида на время отрешиться от происходящих событий. Задумчивый взгляд может наблюдаться у человека в минуты сильной погруженности в себя. Кажется, что он внутри решает очень сложные проблемы, которые не могут быть выражены словесным образом. Когда мы уходим в себя, это сразу заметно окружающим. Такой индивид иногда перестает отвечать на вопросы, его лицо приобретает чрезмерно сосредоточенное выражение.
Задумчивый взгляд может наблюдаться у человека в минуты сильной погруженности в себя. Кажется, что он внутри решает очень сложные проблемы, которые не могут быть выражены словесным образом. Когда мы уходим в себя, это сразу заметно окружающим. Такой индивид иногда перестает отвечать на вопросы, его лицо приобретает чрезмерно сосредоточенное выражение.
Очень о многом может рассказать невербальное общение. Кинетические средства общения часто помогают сделать правильные выводы, глядя на человека. Ведь словами можно сказать о многом, однако не всегда они бывают искренними. Существует много примеров того, как люди умеют обманывать друг друга. Не обязательно это происходит с целью извлечения конкретной выгоды, просто ложь иногда оказывается удобной позицией. Люди с удовольствием выбирают для себя наименее проблемный вариант.
Паралингвистическая подсистема
Кинетические средства общения включают в себя также такие составляющие, как вокальные качества голоса, его диапазон, тональность, тембр. Все это имеет большое значение в процессе взаимодействия. Часто бывает так, что важно не то, что именно говорит человек, а именно сама интонация и тон. Люди обращают внимание прежде всего на эти параметры, а уже потом на произносимые слова. Именно так мы и решаем, приятен ли нам человек, хочется с ним дальше общаться или нет. Вот почему эти составляющие необходимо принимать во внимание.
Все это имеет большое значение в процессе взаимодействия. Часто бывает так, что важно не то, что именно говорит человек, а именно сама интонация и тон. Люди обращают внимание прежде всего на эти параметры, а уже потом на произносимые слова. Именно так мы и решаем, приятен ли нам человек, хочется с ним дальше общаться или нет. Вот почему эти составляющие необходимо принимать во внимание.
Особенности произношения
Все люди говорят по-разному. Хотя мы пользуемся одними и теми же фразами, однако произносим слова индивидуально. Кинетические средства общения – это то, что влияет на понимание собеседника. Тихая речь производит впечатление отсутствия инициативы, заторможенности, вялости. К такому индивиду приходится прислушиваться, чтобы понять, о чем он говорит, напрягать свой слуховой аппарат. Как правило, у окружающих подобная речь не вызывает особого восторга. Человеку становится довольно трудно что-то доказать или убедить в чем-либо окружающих. Чрезвычайно громкая речь тоже вызывает определенный дискомфорт у слушателей. Собеседник может волноваться, так как его начнет раздражать отрывистое звучание голоса.
Собеседник может волноваться, так как его начнет раздражать отрывистое звучание голоса.
Обычно люди, которые стремятся говорить слишком громко, имеют намерение навязывать окружающим свою волю. Они не хотят прислуживаться к советам извне, не стремятся подстраиваться под требования социума. Часто такие люди имеют выраженные лидерские качества. Они предпочитают демонстрировать свои наклонности ярко, быстро и безапелляционно. Лучше всего воспринимается на слух речь средней громкости: она не раздражает слух, не создает никакого негативного впечатления, не требует эмоционального напряжения.
Тональность и тембр голоса
У каждого из нас имеется своя манера произношения. Так человек получает возможность выражать свою индивидуальность. Высокий голос вызывает большее одобрение, чем низкий. Это обусловлено тем, что подсознательно люди стремятся к одобрению и всестороннему принятию. Каждый надеется оказаться услышанным в толпе, а не остаться наедине с самим собой, будучи охваченным безликой массой. Женские голоса воспринимаются слухом как более мягкие и ласковые, а мужские – твердыми и властными.
Женские голоса воспринимаются слухом как более мягкие и ласковые, а мужские – твердыми и властными.
Экстралингвистическая подсистема
Кинетические средства общения используются и в личном, и в деловом взаимодействии. Большой ошибкой было бы предполагать, что в условиях проведения различных научных конференций люди всецело концентрируются только на вербальной стороне. Конечно, большое значение имеют произносимые слова, однако немаловажную смысловую окраску приобретают такие составляющие как темп речи, наличие пауз или смеха во время монолога.
Темп речи
Люди, которые говорят слишком быстро, невольно привлекают к себе внимание. Они заставляют окружающих к ним прислушиваться и воспринимать произносимую речь серьезно. Как правило, они умеют уважать собственный труд, стараются открыто и доверительно преподносить информацию. Медленная речь, напротив, расслабляет, создает ощущение дремоты и монотонности. Если человек чрезмерно растягивает слова, то это настолько утомляет, что восприятие его голоса значительно усложняется. Хорошо воспринимается на слух средний темп речи, когда индивид произносит слова с одинаковой скоростью, максимально четко и внятно.
Хорошо воспринимается на слух средний темп речи, когда индивид произносит слова с одинаковой скоростью, максимально четко и внятно.
Наличие пауз
Когда приходится слушать длинную речь, очень важно, чтобы она не звучала монотонно. Однообразие невероятно утомляет слушателей, вызывает у них приступы зевоты. По этой причине, готовя длинный доклад, необходимо заранее включать туда небольшие паузы. Эти моменты помогут сконцентрировать внимание слушателей, помочь им в восприятии готового материала. Наличие пауз особенно необходимо тогда, когда тема сложная и требует дополнительной подготовки для восприятия информации. Лучше поделить свою речь на несколько блоков, тогда она будет лучше восприниматься и у слушателей не будет путаницы в головах.
Наличие смеха
К кинетическим средствам общения относятся также юмор и различные шутки. Эти элементы значительно облегчают восприятие сложного однообразного монолога. Когда речь оппонента сопровождается смехом, то его становится проще слушать. Юмор способствует формированию доверия и лучшему усвоению преподносимого материала. Чем больше человек старается улыбаться и шутить, тем легче с ним становится общаться.
Юмор способствует формированию доверия и лучшему усвоению преподносимого материала. Чем больше человек старается улыбаться и шутить, тем легче с ним становится общаться.
«Закрытая» поза
Оказывается, очень важным компонентом взаимодействия между людьми является то обстоятельство, насколько уверенно индивид умеет держаться. Важно, чтобы во время беседы он не демонстрировал окружающим так называемую «закрытую» позу: не рекомендуется скрещивать руки на груди, поворачиваться боком к собеседнику. Если человек поступает именно так, то он невольно транслирует в окружающий мир собственный страх, недоверие и неприятие. «Закрытая» поза может являться предвестником конфликта, когда личность по тем или иным соображениям опасается демонстрировать свои настоящие чувства.
«Открытая» поза
Она говорит о доброжелательном отношении. Как правило, человек свободно владеет собственным телом, никуда не спешит и не стремится в чем-то превосходить своих собеседников. «Открытая» поза привлекает к себе больше внимания, помогает сосредоточиться на предмете разговора, а не отвлекаться на собственное эмоциональное состояние.
Таким образом, кинетические средства общения помогают лучше воспринимать и усваивать информацию. Для эффективного взаимодействия между людьми одних только слов недостаточно. Мы воспринимаем посредством общения не только саму речь, но и считываем отношение к себе, обращаем внимание на интонацию, которая сопровождает высказывания.
Невербальная коммуникация у детей и подростков
Как М.Н. Ночевник справедливо отмечает, что «невозможно представить себе развитие человека, существование его как личности, его связь с обществом вне общения с другими людьми» (35, с. 37). Межличностное общение является необходимым условием человеческого существования, без которого невозможно полноценное формирование не только отдельных психологических функций, процессов и свойств человека, личности в целом и общества (социума). В связи с этим классики марксизма К. Маркс и Ф. Энгельс утверждали: «…действительное духовное богатство индивида зависит от богатства его действительных отношений» (35, с. 78). Исторический опыт и повседневная практика свидетельствуют, что полная изоляция человека от общества, удаление от общения с другими людьми, приводит к полной утрате человеческой личности, ее социальных характеристик (феномен «детей Маугли»).
Общение включает в себя все многообразие психических и материальных форм деятельности человека и является его жизненной потребностью (35, с.5). Коммуникация — это технический процесс обмена информацией между двумя или более лицами. Индивид — это отдельная личность, человек, принадлежащий к человеческому роду. Общение — это процесс обмена информацией между людьми. В нашей работе мы рассматриваем коммуникацию и общение как синонимы.
Использование невербальных средств общения уходит своими корнями в историю, когда Цицерон учил ораторов жестикулировать. На протяжении многих веков театральные актеры профессионально используют невербальные средства. С развитием технологий и появлением кинематографа невербальные средства стали ведущими средствами коммуникации в немых фильмах, а также были и остаются одними из важных средств в звуковых фильмах.
Современная школа, вовлекая ребенка в систему социального общения, вводит его в сферу регулируемых отношений с ближайшим социумом и позволяет ему усвоить нормы культурного сообщества. Одним из важнейших условий для полноценной социальной адаптации ребенка является правильный язык.
Анализ литературы по проблеме воспитания и обучения детей с нарушениями речи показал, что существуют различные методы профилактической работы, а также методические рекомендации по преодолению недоразвития невербальной коммуникации у детей с речевой патологией. Однако их недостаточная систематизация выявляет объективно существующее противоречие между признанием необходимости развития невербальной коммуникации и разработкой педагогической системы развития оптико-кинетических, паралингвистических и экстралингвистических средств общения у детей с речевой патологией.
К особенностям использования невербальных средств общения у детей 8-10 лет относятся: использование в межличностном общении большого количества жестов, заменяющих слова или усиливающих их значение, плохой контроль над использованием мимических движений, неумение менять темп и громкость речи в монологическом высказывании, недостаточная эмоциональная окраска речи в процессе обучения.
Общетеоретическая характеристика невербального компонента общения
Как пишет А.Пиз: «Кажется почти невероятным, что более чем за миллион лет эволюции человека невербальные аспекты коммуникации начали серьезно изучаться только с начала 60-х гг., а обществу стало известно об их существовании только после того, как Дж.Фаст опубликовал свою книгу в 1970 г.» (41, С.16). Эта книга обобщала исследования о невербальных аспектах общения, проведенные учеными-бихевиористами до 1970 г. Однако даже сегодня большинство людей все еще не знают о существовании языка телодвижений, несмотря на его важность в жизни. Вместе с тем, проблема восприятия и психологической интерпретации невербального поведения относится к проблемам с многовековой историей, о чем свидетельствуют данные, опубликованные в работах Е.А.Петровой, В.В.Мироненко, М.Битяновой. Анализ перечисленных источников позволяет сделать вывод, что к проблеме невербального общения в различное время обращались философы, психологи, медики, лингвисты, искусствоведы. На протяжении тысячелетий, по замечанию В.В.Мироненко, данная проблема обрастала научными и псевдонаучными фактами. Знания о невербальном поведении и его связях с внутренним миром человека регистрировались сначала в памятниках характерологического жанра — литературе афоризмов, моральных наставлений, позднее — в работах философов.
В средние века отдельные взгляды физиогномистов разделяли и в той или иной степени совершенствовали Ибн Сина и ряд ведущих алхимиков. В эпоху Возрождения — Иоанн Дунс Скот и Леонардо да Винчи, в XVI-XVIII столетиях — Френсис Бэкон и непревзойденный физиогномист, швейцарский священник, поэт и художник Иоганн Гаспар Лафатер. В своем сочинении «Искусство познавать людей по физиогномиям» он совершил практически первую научную попытку систематически исследовать выразительные движения.
В XVI в. выдающийся немецкий врач-хирург Парацельс также пользовался в своей практической деятельности мимическими критериями, хотя его идеи не получили сколько-нибудь широкого распространения.
В 1664 году Джон Баливер выпустил две книги, посвященные языку жестов человека: «Хирология, или Естественный язык жестов» и «Хирономия, или Искусство риторики рук». В этих книгах были составлены первые из известных в Европейских странах таблицы с систематизацией знаков выразительной жестикуляции (30, С.135).
Классическим примером осмысления роли и значения жестов, мимики, интонации в жизни людей можно считать труд Жан-Батиста Дюбо — французского философа-просветителя XVIII века «Критические размышления о поэзии и живописи», где автор приходит к мысли о том, что издавна существует наука «нема», которая, не размыкая уст, говорит руками и некоторыми телодвижениями. Наиболее влиятельной работой, по данным А.Пиза (42, С.17) к началу ХХ в. была работа Ч.Дарвина «Выражение эмоций у людей и животных», опубликованная в 1872 г. Она стимулировала современные исследования в области «языка тела», а многие идеи Ч.Дарвина и его наблюдения признаны сегодня исследователями всего мира.
Значительным шагом в описании жестов стали также работы известного французского режиссера Дельсарта. В России начала ХХ века система Дельсарта стала широко известной благодаря книге Сергея Волконского «Выразительный человек», где рассматривались как законы человеческого движения, так и их семиотическая классификация.
С 1900 по 1979 год на английском языке было издано шесть крупных монографий о жестах. Наибольший интерес во всем мире вызвали работы
Д.Эфрона, М.Кричли, Ч.Морриса. Серьезным исследованием «боди лэнгвиджа» с конца 70-х гг. начал заниматься автсралийский ученый Аллан Пиз , который является признанным знатоком психологии человеческого общения и автором методики обучения основам коммуникации.
Интерес к невербальным средствам общения в ХХ столетии столь возрос, что выделилась особая область исследований — кинесика (от англ. Kinesics, греч. — kinesis). Основы изучения этой области психологии заложены в 50-х гг. работами шведского ученого Р.Бердвистелла.
Современные представители данной научной области (Дж.Фаст, Дж.Ниренберг) раскрывают закономерности влияния невербальных способов общения на установление контактов и взаимопонимания между партнерами.
Под кинесикой принято понимать зрительно воспринимаемый диапазон движений, выполняющих экспрессивно-регулятивную функцию в общении. Кинесика — это не только «язык тела» (жесты, мимика, позы, взгляд), но также манера одеваться, причесываться. К кинесике помимо названных движений относятся также такие движения, которые связаны с использованием предмета: хлопанье дверью, поскрипывание стулом, почерк. Как видно, кинесика — понятие, использующееся для обозначения различных движений чеоловека, но чаще всего при изучении движений рук и лица.
Понятие невербальной коммуникации в психолингвистической литературе
В психолингвистической литературе различают вербальные и невербальные средства общения. Психологический словарь определяет невербальное общение (от латинского — nonverbal) как невербальные средства общения и взаимодействия с собеседником.
Невербальные средства общения изучаются на протяжении многих лет следующими науками:
Кинетика, изучающая внешние проявления человеческих чувств и эмоций. Изучаются такие проявления, как мимика (движение лицевых мышц), жесты (жестикуляционные движения отдельных частей тела), пантомима (двигательные движения тела: поза, поза, поклоны, походка).
Такесика — прикосновения в ситуации общения: рукопожатие, поцелуи, прикосновения, поглаживания, отталкивания и другие.
Проксемика — расположение людей в пространстве во время общения. При контакте с человеком существуют следующие зоны дистанции:
А) Интимная зона (15-45 см), в эту зону допускаются только близкие, хорошо знакомые люди, для этой зоны характерно доверие, мягкий голос, тактильный контакт, прикосновения. Исследования показывают, что проникновение в интимную зону вызывает определенные физиологические изменения в организме: усиленное сердцебиение, повышенное выделение адреналина, прилив крови к голове и т.д.
Преждевременное проникновение в интимную зону в процессе общения всегда воспринимается собеседником как посягательство на его неприкосновенность.
B) личная или приватная зона (45-120 см) — для непринужденных бесед с друзьями и коллегами, предполагает только визуальный и зрительный контакт между партнерами, поддерживает беседу;
в) социальная зона (120-400 см) обычно наблюдается на официальных встречах в офисах, лекционных залах и других рабочих местах, как правило, с людьми, которых мало кто знает;
D) публичная зона (более 400 см) предполагает общение с большой группой людей — в лекционном зале, на собрании и т.д.
К средствам невербальной коммуникации относятся:
Выражение лица, поза, взгляд собеседника. Вы можете укрепить. Дополните или опровергните значение фразы. Выражение лица — движение лицевых мышц, отражающее внутреннее эмоциональное состояние, может дать правдивую информацию о том, что испытывает человек. Выражение лица дает нам более 70% информации, то есть глаза, взгляд, лицо человека могут рассказать больше, чем произнесенные слова. Так, на основании многих исследований замечено, что человек пытается скрыть свою информацию (или лжет), если его глаза встречаются с глазами партнера менее 1/3 времени разговора. Лоб, брови, рот, глаза, нос, подбородок — эти части лица выражают основные человеческие эмоции: Печаль, гнев, радость Удивление, страх, отвращение, счастье, интерес, грусть и т.д. Положительные эмоции легче всего распознать: Радость, Любовь, Удивление и негативные эмоции — самые сложные для восприятия человеком: Печаль, гнев, отвращение. Важно отметить, что брови и губы несут основную когнитивную нагрузку в ситуации распознавания истинных чувств человека.
Жесты как средство невербальной коммуникации могут быть повседневными, то есть иметь фиксированное значение, или экспрессивными, то есть служить для того, чтобы сделать речь более выразительной. Таким образом, жесты в общении несут много информации, в языке жестов, как и в речи, есть слова, предложения.
Дистанция, с которой общаются собеседники, зависит от культурных, национальных традиций, а также от степени доверия к собеседнику.
Мы можем сделать вывод, что все в нашем невербальном общении, что может быть преодолено и отменено с помощью очевидных фактов опыта.
Мы не просто поддаемся потоку впечатлений, передаваемых языком мимики и жестов, но активно наблюдаем, то есть настраиваем свои органы чувств на максимально точное восприятие взаимодействий. Например, когда мы смотрим на сложный объект, мы фиксируем оба глаза и направляем их так, чтобы четко видеть точку, которая привлекла наше внимание, то есть так, чтобы ее изображение попадало на поверхность обоих глаз. Затем мы исследуем все точки объекта, которые заслуживают нашего внимания.
Проблемы обучения младших школьников невербальному общению
Развитие невербального общения невозможно без обогащения впечатлений младшего школьника, без его знакомства с широким кругом предметов и явлений окружающего мира, без овладения действиями с вещами. На этом основано владение значениями слов.
Было бы неправильно думать, что развитие невербальных средств общения важно только для умственного развития ребенка. Он играет важную роль в развитии первых моральных оценок. Эти оценки усваиваются через речь и эмоциональное поведение взрослых. По мнению К.Д. Ушинского, невербальное общение совершенно необходимо для того, чтобы у ребенка сформировалось иное отношение к хорошему и плохому, красивому и безобразному. Из этого вырастают настоящие нравственные чувства и идеи зрелого человека.
На каждом шагу дети сталкиваются с тем, что взрослые по-разному относятся к тем или иным явлениям и оценивают их определенными словами, мимикой и жестами. Даже первые слова-оценки «хорошо» и «плохо» сопровождаются интонацией голоса, мимикой и жестами, которые усиливают эффект от этих слов. Вначале дети подражают взрослым и говорят только о том, что они хорошо знают и что прочно укоренилось в их еще небольшом жизненном опыте. Но постепенно они начинают расширять диапазон своих суждений.
Развитие невербального общения имеет особое значение для усвоения ребенком правил поведения. Ведь почти все требования взрослых выражаются либо словом «нельзя», либо словом «нельзя», сопровождаемым эмоционально окрашенными жестами. Взрослые также вводят слово «можно», и ребенок начинает использовать это слово, чтобы спросить разрешения сделать что-то.
Значительный скачок в общем развитии ребенка, связанный с овладением невербальным общением, особенно заметен в школьном возрасте. Ребенок уже лучше понимает язык других людей, и это открывает большие возможности для появления совершенно новых способов общения со взрослыми и сверстниками. Взрослый должен при любой возможности стараться чаще напоминать ученику о необходимости считаться с другими людьми. Молодой ученик довольно легко «заражается» чувствами других людей. Учитывая его легко поддающееся влиянию поведение, родители и учителя должны обращать внимание на нюансы его собственного поведения.
Здесь важно отметить важность создания положительных эмоций у ребенка и устранения отрицательных. Преобладают ли эти или другие эмоции у младшего школьника, во многом зависит от того, как организована его жизнь и какие чувства вызывает поведение старших. Часто бывает, что с интересами ученика родители не особо считаются, разговаривают с ним раздраженно, находят повод лишний раз упрекнуть его, позволяют старшим братьям или сестрам проявлять «власть» над младшими по возрасту. Стоит ли удивляться, что ребенок не весел, не спокоен в своем поведении, что он не проявляет сострадания к другим, когда им плохо!
Большое значение в младшем школьном возрасте имеет общение со сверстниками. С самого раннего возраста родители должны всячески поддерживать дружеские отношения ребенка друг с другом. В совместной работе учащихся по воспитанию можно наглядно увидеть проявление невербального общения, ведь нужно считаться с интересами друг друга, уступать, делиться с товарищем, помогать ему. Младшие школьники начинают проявлять сострадание к другим.
Что делать, когда дети ссорятся? Споры не должны оставаться без внимания. Взрослый должен разобраться с причинами споров и справедливо разрешить возникшее недоразумение.
В то же время в некоторых работах мы видим, что младшие школьники могут подражать друг другу и учиться помогать друг другу, проявлять эмпатические чувства под умелым руководством взрослых, они чувствуют потребность общаться друг с другом, используя невербальные средства общения, и это способствует развитию невербальной коммуникации.
Для того чтобы младшие школьники научились различным невербальным средствам общения, с ними можно выполнять различные упражнения в классе.
Современные учебные программы и учебники по обучению невербальному общению для учащихся начальной школы
В современной начальной школе существуют интегрированные курсы «Знакомство с окружающим миром», «Естествознание», «Литература мирового искусства», «Занимательный русский язык», в которых рассматриваются вопросы вербального и невербального общения. Учитывая специфику курсов и возрастные особенности младших школьников, допускается включение этих дисциплин в учебный план как отдельного предмета и в меньшем объеме, чем это указано в неизменной части базисного учебного плана. В этом случае содержание этих курсов дополняется изучением других предметов (чтение, изобразительное искусство и т.д.).
В программе «Воспитание музыкальной культуры учащихся» (Шмагина Т.С.) основная цель — показать возможность углубления основных положений программы формирования музыкальной культуры учащихся в средней школе за счет включения невербальной коммуникации. Конечно, знакомство с содержанием программных уроков можно рассматривать как проблемные ситуации, использовать нетрадиционные приемы и методы работы, конкретные логические вопросы, вовлекать учащихся в дискуссию, тем самым всегда поддерживается музыкальная деятельность учителя и учащихся.
Обучение младших школьников невербальным средствам общения особенно ярко проявляется на уроках музыки, так как — это постоянное творческое общение между музыкой — учителем — детьми, между композитором — исполнителем — слушателем.
В современных учебных программах начальной школы художественные категории составляют основу для определения основных понятий обучения. Таким образом, художественно-образовательная идея урока является первичной, а тема четверти — вторичной. Вместо привычных методических терминов (тип урока, план, конспект и структура урока) вводятся категории жанр, сценарий, драматургия и режиссура.
Художественно-педагогическая идея становится не только идеей для одного урока, но и может служить своеобразным подходом к усвоению тематического материала на всю учебную четверть.
Программа по русскому языку Рамзаевой Т.Г. «Уроки русского языка» дает не только методический комментарий к блокам уроков, но и является помимо конкретного материала «Для учителя», здесь мы предлагаем также литературу, которая даст практическую помощь в разработке уроков по развитию неязыковых средств общения младших школьников.
Интересно описать уроки творчески работающих учителей. Личная интонация, творческая свобода, художественная новизна — все, что отличает творческого человека. Будь то просто мини-диалог учителя с детьми или искренняя любовь учителя к теме, интересное содержание?
Каждый вопрос, жест, выражение лица должны быть мотивированы учителем. Вопрос ради вопроса не вписывается в современное общество. Человек, задающий вопрос, должен быть заинтересован в ответе, выражать его невербально и действительно хотеть услышать ответ. И ярче всего это проявляется в школе, на уроках искусства и творчества.
На странице курсовые работы по психологии вы найдете много готовых тем для курсовых по предмету «Психология».
Читайте дополнительные лекции:
- Виды психических процессов: ощущение
- Методология психологии
- Механизмы переработки представлений в воображаемые образы — Сущность и виды воображения
- Основные стратегии совладания и формы поведения человека при внезапной утрате близкого человека
- Проблема детских страхов
- Роль клинического психолога в организации мероприятий первичной профилактики
- Формирование психодиагностической тактики в работе медицинского психолога с пациентом
- Кризисы профессионального становления
- Гендерные различия в формировании и проявлении профессионального стресса
- Рисуночные тесты, их значение для психодиагностики нарушений развития
Волновая кинетика случайных волоконных лазеров
Волновое кинетическое уравнение для волоконных лазеров
Кинетическая теория описывает среднюю (макроскопическую) вероятностную эволюцию сложной системы. Исходя из основных микроскопических динамических уравнений, управляющих взаимодействием элементарных составляющих (например, частиц или волн), кинетическое описание эффективно уменьшает большое количество степеней свободы в исходной нелинейной системе, предполагая некоторые предположения о статистике флуктуаций.В контексте волновой системы кинетическое уравнение описывает изменение величин, которые являются средними за периоды времени, превышающие периоды волн. В этом случае волны с разными частотами приобретают разные фазы, что позволяет рассматривать их при усреднении как примерно независимые. В настоящее время волновой кинетический подход (известный в некоторых контекстах как волновая турбулентность 4 ) используется в целом ряде физических приложений, от конденсата Бозе – Эйнштейна до астрофизики 5 .
В традиционной волновой кинетике начальный (произвольный) волновой спектр F ω постепенно эволюционирует через каскады многочисленных слабых нелинейных взаимодействий в сторону статистически устойчивого состояния, как показано на рис.1а. Эволюция от начального к асимптотически стационарному спектру определяется волновым кинетическим уравнением 4 . Долговременное асимптотическое состояние устойчиво как глобально, так и локально, что означает, что статистические свойства не меняются при любом произвольном сдвиге во времени. Это достигается, когда внешняя закачка энергии в систему и сброс энергии из системы не зависят от времени.
Рисунок 1. Волновая кинетика в активных циклических системах.( a ) В классической волновой кинетике исходный волновой спектр постепенно эволюционирует до статистически стационарного волнового спектра, когда накачка / сброс энергии однородна в течение времени эволюции.Эволюция регулируется волновым кинетическим уравнением. Глобальный статистически стационарный волновой спектр также является локальным стационарным решением, то есть не изменяется при сдвиге во времени на любое произвольное значение. ( b ) В активных циклических системах накачка / сброс энергии действует периодическим образом, что приводит к циклической динамике и двухмасштабной эволюции спектра волн. Когда накачка / сброс энергии изменяется в пределах цикла, спектр волн локально нестационарен, демонстрируя сильные изменения в пределах каждого цикла.Эта эволюция регулируется уравнением кинетики волн с локальной накачкой (3). В то же время спектр развивается постепенно, от цикла к циклу, подобно классической волновой кинетике. Если общая накачка в цикле равна сбросу энергии, система приближается к глобальному стационарному решению. ( c ) В случайном волоконном лазере оптическая накачка распределяется по волокну, а демпфирование происходит на концах волокна, где выходит излучение. Каждый проход оптического волокна составляет один цикл.Спектр генерации сильно изменяется во время эволюции в каждом цикле из-за оптического усиления. Случайная распределенная обратная связь связывает оптический спектр в последующих циклах. Поскольку усиление равно потерям в лазере, оптический спектр должен быть идентичным в разных циклах. Таким образом, глобальное стационарное решение действительно существует.
В волоконном лазере излучение претерпевает сильные периодические (циклические) изменения вдоль резонатора из-за комбинированного действия усиления, потерь, дисперсии и нелинейности, наложенного на медленную эволюцию от одного обхода к другому.Глобальное статистическое равновесие все еще может существовать в терминах отображения Пуанкаре 20 как состояние, которое статистически воспроизводится после каждого кругового обхода. Однако, в отличие от случая систем, однородных по эволюционной переменной, локального статистически устойчивого состояния не существует, а это означает, что спектр волн существенно изменяется при любом произвольном сдвиге во времени. В результате в лазерных системах действительно существуют два масштаба по координате эволюции — быстрая эволюция в пределах каждого кругового обхода и медленная эволюция от цикла к циклу.Это приводит к неравномерной спиралевидной эволюции, которая отличается от монотонной релаксации в классической волновой кинетике, как схематически изображено на рис. 1b. Важно, чтобы амплитуды волн могли в достаточной степени увеличиваться или уменьшаться во время быстрой эволюции в течение каждого обхода в зависимости от того, в какой момент в резонаторе лазера преобладает накачка или сброс. Таким образом, классический волновой кинетический подход нельзя напрямую применять к типичным волоконным лазерам, поскольку необходимо учитывать сильную локальную (квазипериодическую) динамику волнового спектра.
Чтобы иметь дело с активными циклическими системами, мы выводим волновое кинетическое уравнение с накачкой, определяющее эволюцию спектра быстрой и сильной волны в оба конца, в дополнение к « стандартному » волновому кинетическому уравнению, определяющему эволюцию спектра медленных и инкрементальных волн из цикла на цикл.
Далее мы кратко опишем вывод кинетического уравнения для циклической активной волновой системы, имеющей множество волн со случайной фазой, нелинейно взаимодействующих посредством кубической нелинейности, которая считается слабой.Мы рассматриваем квадратичный закон дисперсии, характерный для оптики и спектрально узких возбуждений. Система подвергается периодической перекачке и сбросу энергии, повторяющимся циклически, и развивается в течение каждого периода и от цикла к циклу (рис. 1b).
Начнем с общего динамического уравнения, описывающего эволюцию комплексного поля огибающей ψ по эволюционной координате z (которая, например, может быть расстоянием распространения внутри полости волокна) в дисперсионной нелинейной среде 21 :
Здесь t обозначает время, а z — координата эволюции внутри цикла.Для волновой системы коэффициенты β и γ описывают, соответственно, дисперсию и нелинейность бегущей волны с интенсивностью, пропорциональной, линейный оператор описывает накачку / сброс энергии в / из системы. В частном случае оптики уравнение (1) представляет собой обобщенное нелинейное уравнение Шредингера, которое описывает распространение света в одномерной среде, γ связано с нелинейностью Керра и является оператором, описывающим оптическое спектрально-зависимое усиление. .Динамические модели, основанные на нелинейном уравнении Шредингера (1), широко используются для описания лазерного излучения в волоконных лазерах (см. Ссылки 22, 23), а также в других оптических системах, характеризующихся сильным влиянием шума и стохастичности 24,25 , 26 .
Консервативная часть уравнения (1), то есть одномерная NLSE, полностью интегрируема и обладает бесконечным числом интегралов движения 27 . С точки зрения кинетики это уравнение представляет собой очень частный случай.Как было указано Захаровым 27 , существует широкий класс интегрируемых уравнений, приводящих к тривиальной кинетике, то есть кинетики вообще нет, и одномерный НУШ принадлежит к этому классу. Отклонения от интегрируемости в уравнении (1) задаются членом с и имеют решающее значение, поскольку; действительно, эти отклонения определяют нетривиальную кинетику волновой системы. Мы строим общую схему, позволяющую вывести кинетическое уравнение для таких почти интегрируемых систем (см. Дополнительное примечание 1 и дополнительный рис.1), который можно использовать во множестве других физических приложений.
Волновая кинетическая теория имеет дело с парной корреляционной функцией
здесь угловые скобки обозначают усреднение на z больше, чем длина дисперсии, где Δ — характерная ширина волнового спектра, определяемая как преобразование Фурье одноточечной корреляции. функция F ( z , z , t ): F ω ( z ) = ∫ dt e iωt F ( z , т ).Далее нас интересует кинетика, то есть эволюция волнового спектра. Обратите внимание, что в оптике F ω ( z ) соответствует непосредственно измеренному оптическому спектру.
Затем мы выводим волновое кинетическое уравнение для корреляционной функции F ω , используя отклонение уравнения (1) от интегрируемого NLSE. Мы используем стандартное предположение, что поле ψ состоит из множества волн со случайными фазами.Эти волны взаимодействуют посредством небольшой нелинейности, которая является источником их случайности. Подробные технические детали того, как выводится уравнение локальной волновой кинетики с использованием диаграмм, можно найти в дополнительных примечаниях 2 и 3.
Полученное волновое кинетическое уравнение выглядит так:
Здесь g ( ω , z ) — коэффициент усиления, зависящий от частоты, — коэффициент дисперсии, g a = g ( ω , z ) + g ( ω 2 , z ) + g ( ω 3 , z ) — g ( ω 1 , z ), g b = g ( ω , z ) + g ( ω 1 , z ) + g ( ω 3 , z ) — g ( ω 2 , z ) , г c = г ( ω 1900 16, z ) + g ( ω 2 , z ) + g ( ω 3 , z ) — g ( ω , z ) ).Уравнение выводится при двойном разделении масштабов, то есть мы рассматриваем случай сильной дисперсии по спектру по сравнению с усилением, и мы предполагаем, что само усиление намного больше, чем изменение усиления по ширине спектра. Отметим, что уравнение (3) описывает волновую кинетику в довольно общем классе активных циклических систем, не ограничиваясь квадратичным дисперсионным соотношением: различные законы дисперсии могут быть легко включены в (3) путем соответствующего изменения множителя Φ.
Существует несколько принципиальных отличий волнового кинетического уравнения (3) для активной циклической системы от классического волнового кинетического уравнения.Прежде всего, поскольку накачка и сброс энергии неоднородны по координате эволюции z в пределах цикла, спектр волн демонстрирует сильные изменения во время эволюции внутри цикла, поэтому локальное стационарное статистически устойчивое состояние не существует. Формально это означает, что нельзя приравнять производную по эволюционной координате z к нулю и найти таким образом стационарное решение, подобное классической волновой кинетике.
Традиционное волновое кинетическое уравнение для консервативной гамильтоновой системы может быть получено из локального волнового кинетического уравнения (3) в пределе нулевого усиления.В этом случае связанные с усилением лоренцевы члены в правой части уравнения (3) превращаются в дельта-функции, обеспечивая сохранение энергии и давая классическое кинетическое волновое уравнение. Однако классическое кинетическое уравнение, полученное из NLSE, тривиально в том смысле, что правая часть обращается в нуль из-за интегрируемости одномерного NLSE 27 . Другими словами, в интегрируемой системе на основе NLSE спектр вообще не эволюционирует. Это не относится к уравнению кинетики локальной волны (3).С формальной точки зрения, волновое взаимодействие однородно в течение медленного времени эволюции 4 в классической волновой кинетике. Однако в активных циклических системах эти взаимодействия опосредованы неоднородным усилением (см. Лоренцевы факторы из-за усиления в уравнении (3)), что приводит к тому, что взаимодействие эффективно только в течение конечного интервала эволюционной координаты. Обратите внимание, что уравнение (3) не сохраняет импульс, потому что накачка и разгрузка изменяются по эволюционной координате z .Мы хотели бы также отметить, что интересная модификация обычного кинетического уравнения волновой турбулентности была изучена в статье Ascheri et al . 28 , где интегрируемость исходной модели NLSE была нарушена из-за поперечной пространственной неоднородности показателя преломления.
Нелинейная кинетическая теория оптического спектра лазера
Теперь применим формализм волновой кинетики в активных циклических системах к оптической системе и впервые получим, насколько нам известно, нелинейную кинетическую теорию оптический спектр лазера.Вообще говоря, классическая волновая кинетика не может быть применена для описания оптического спектра лазера, поскольку лазеры представляют собой активные циклические системы с двухуровневой эволюцией. Первые шаги к кинетическому рассмотрению были сделаны в экспериментальных работах 29,30 , в которых была обозначена проблема. Сначала для конкретной системы был проведен эвристический анализ, и в нем были представлены убедительные экспериментальные доказательства необходимости новой строгой нелинейной кинетической теории 29,30 .
Здесь локальное волновое кинетическое уравнение (3) позволяет нам сформулировать нелинейную кинетическую теорию лазерного спектра, обобщающую знаменитую линейную кинетическую теорию Шавлоу и Таунса 31 . Мы сосредоточимся на частном, хотя и очень интересном случае случайного волоконного лазера, работающего через обратную связь по рэлеевскому рассеянию 19 . До сих пор, кроме прямого численного моделирования на основе NLSE 32 , не существовало теории, ни динамической, ни кинетической, описывающей оптический спектр случайного волоконного лазера, что было дополнительной мотивацией для выбора этой конкретной системы в качестве теста. кровать нашей общей теории.
Рассмотрим случайный волоконный лазер длиной L , который представляет собой длинный участок стандартного телекоммуникационного волокна с оптической накачкой. Слабая случайная распределенная обратная связь вызвана рэлеевским рассеянием в сердцевине волокна на неоднородностях показателя преломления, а рамановское усиление индуцируется излучением накачки, что в совокупности приводит к случайной генерации 19 . Оптический спектр лазера состоит из множества волн случайных амплитуд и фаз, которые взаимодействуют посредством нелинейности Керра.Фактически, резонатор случайного волоконного лазера прямо соответствует концепции активных циклических систем, рис. 1в. Действительно, существуют две встречные генерирующие волны, которые идентичны при условии симметричной оптической накачки. Распространение каждой волны от одного конца оптического волокна к другому определяется динамическим уравнением (1). Мы рассматриваем каждый проход оптического волокна как один кинетический цикл. Во время распространения оптический спектр подвержен сильным изменениям.Действительно, оптическое усиление распределено по всему волокну, и интенсивность генерации в начальной точке каждого цикла, при z = 0, очень мала, поскольку почти вся оптическая мощность излучается с конца волокна. Следовательно, необходимо применять кинетическое уравнение локальной волны (3).
Мы сосредотачиваемся на статистическом равновесии, соответствующем среднему спектру лазера, который достигается в течение многих циклов. Формально наличие стационарного лазерного спектра эквивалентно следующему граничному условию на корреляционную функцию:, где — коэффициент случайного отражения выходного элемента связи лазера.Принимая локальное кинетическое уравнение (3) и граничные условия, используя теорию возмущений для небольшой нелинейности и после некоторых расчетов, мы выводим кинетическое уравнение, непосредственно определяющее стационарный лазерный спектр F ω , ( Дополнительное примечание 4 и Дополнительное уравнение 43), являющиеся всего лишь модифицированной версией локального кинетического уравнения (3). В зависимости от величины дисперсии спектр волны, то есть спектр генерации случайного волоконного лазера, имеет различную форму (рис.2а, и разная скорость передачи энергии от центра спектра к крыльям спектра (рис. 2б). Отметим, что в случае длинного рамановского волоконного лазера с обычным линейным резонатором подобная форма спектра гиперболического секущего наблюдалась и вводилась феноменологическим образом в предельном случае, когда дисперсия намного больше, чем нелинейность 29 .
Рисунок 2: Статистически стационарные решения локального волнового кинетического уравнения.Спектры численно вычисляются из уравнения кинетики локальной волны (3) в частном случае случайного волоконного лазера в качестве примера Дополнительного уравнения 43.( a ) Спектр волны в зависимости от отношения усиления / дисперсии λ = 2 г / β Δ 2 , где Δ — ширина оптического спектра. ( b ) Наклон спектра волны θ при различных параметрах усиления / дисперсии λ . Наклон определяется из аппроксимации крыла волнового спектра экспоненциальной функцией exp ( −θx ). Пунктирной линией показан наклон волнового спектра в случае спектра волны секущей гиперболической формы.
Чтобы проверить предсказания нашей нелинейной кинетической теории о спектре лазера, мы разработали случайный волоконный лазер, работающий в той же области параметров, что и в теории (см. Методы и дополнительное примечание 5 и дополнительный рис.2). Спектр лазера был измерен и показан на рис. 3а. Ниже порога наблюдается усиленный спектр спонтанного излучения, соответствующий профилю усиления. Выше порога генерации случайный спектр волоконного лазера демонстрирует нелинейное кинетическое уширение. Форма спектра прекрасно описывается нашей теорией (рис. 3а). Кроме того, теория описывает закон уширения спектра лазера от мощности накачки (рис. 3б (красная линия)).
Рис. 3. Нелинейное кинетическое описание случайного оптического спектра волоконного лазера.( a ) Оптический спектр, измеренный экспериментально: около порога генерации (синяя кривая, мощность лазера = 0,025 Вт), немного выше порога генерации (зеленая кривая, 0,2 Вт) и значительно выше порога генерации (красная кривая, 1,5 W). Оптический спектр, предсказанный с помощью дополнительного уравнения 43 кинетического уравнения локальной волны, для мощности лазера 1,5 Вт показан красной пунктирной линией ( b ). Ширина спектра как функция выходной мощности лазера в теории и эксперименте.Экспериментальные данные показаны черными кружками. Прогноз расширения спектра из нелинейной кинетической теории на основе уравнения локальной волновой кинетики (3) показан синей пунктирной линией. Прогноз для сужения спектра на основе модифицированного дополнительного уравнения 47 линейной кинетической теории Шавлоу – Таунса показан пунктирной зеленой линией. Красная линия — это сумма нелинейного и линейного вкладов. На вставке — сужение спектра вблизи порога в логарифмическом масштабе.
Помимо нелинейного кинетического вклада в спектр волн, хорошо выраженного при высокой мощности, существует также линейный кинетический вклад, доминирующий при малых мощностях.Действительно, из основополагающей работы Шавлова и Таунса 31 хорошо известно, что спектр лазера демонстрирует сужение спектра, в то время как мощность накачки увеличивается выше порога генерации. Спектральное сужение наблюдается и в спектре случайного волоконного лазера (см. Рис. 3b. Однако подход Шавлова и Таунса 31 может быть использован только для лазерных резонаторов, имеющих четко определенные моды резонатора, что не относится к случайный волоконный лазер, имеющий безмодовый, то есть непрерывный спектр.Чтобы справиться с этим, мы модифицируем подход Шавлоу – Таунса и выводим уравнение, описывающее сужение спектра случайного волоконного лазера при малой мощности (см. Дополнительный рисунок 3, дополнительное примечание 6 и дополнительное уравнение 45). Его решение, дополнительное уравнение 47, хорошо описывает экспериментально наблюдаемое сужение оптического спектра вблизи порога генерации (рис. 3b, вставка).
Обратите внимание, что на самом деле Дополнительное уравнение 45 является линейным кинетическим уравнением, действующим при более низких мощностях, где можно пренебречь нелинейными взаимодействиями между различными спектральными компонентами.В этом смысле локальное волновое кинетическое уравнение, уравнение (3), описывающее нелинейный спектр, расширяющееся выше порога генерации, является расширением уравнения Шавлоу – Таунса на случай взаимодействия нелинейных мод. Реальный оптический спектр имеет как линейный, так и нелинейный вклад. Сумма линейных и нелинейных членов хорошо описывает экспериментально измеренную ширину спектра лазера во всем диапазоне мощностей (рис. 3б). Небольшая остаточная разница между теорией и экспериментальными данными может быть объяснена влиянием эффекта кросс-фазовой модуляции (XPM), вызванного волной накачки.Оценка дает значение наведенной XPM спектральной ширины 0,2 нм при малой мощности.
Оптическая мощность сигнала — обзор
3.1.1 Измерение относительной интенсивности шума (RIN)
В полупроводниковых лазерах, работающих выше порогового значения, хотя стимулированное излучение доминирует в процессе излучения, все же существует небольшой процент фотонов, которые генерируются спонтанное излучение. Причиной возникновения шума интенсивности оптического излучения в полупроводниковом лазере являются эти фотоны спонтанного излучения.В результате, когда полупроводниковый лазер работает в непрерывном режиме, эффект спонтанного излучения заставляет и плотность носителей, и плотность фотонов колебаться вокруг своих равновесных значений [1].
Обычно шум интенсивности в лазерном диоде, вызванный спонтанным излучением, представляет собой широкополосный шум; однако этот шум может быть дополнительно усилен резонансом релаксационных колебаний в резонаторе лазера, который изменяет спектральную плотность шума. Как показано на рисунке 3.1.1, каждое отдельное событие спонтанного увеличения плотности носителей увеличивает оптическое усиление лазерной среды и, таким образом, увеличивает плотность фотонов в резонаторе лазера.Однако эта повышенная плотность фотонов будет потреблять больше носителей в резонаторе, создавая насыщение усиления в среде; в результате плотность фотонов будет уменьшаться из-за уменьшения оптического усиления. Это, в свою очередь, увеличит плотность несущих из-за уменьшения эффекта насыщения. Этот резонансный процесс является сильным на определенной частоте Ω R , которая определяется оптическим усилением G , дифференциальным усилением dG / dN и плотностью фотонов P в лазерном резонаторе как
Рис. 3.1.1. Объяснение релаксационного колебательного процесса в полупроводниковом лазере.
(3.1.1) ΩR2 = G⋅P⋅dGdN
Из-за релаксационных колебаний интенсивность шума полупроводникового лазера зависит от частоты. Нормированная спектральная плотность шума интенсивности может быть описана следующим выражением:
(3.1.2) H (Ω) ∝ | ΩR2 + BΩ2jΩ (jΩ + γ) + ΩR2 | 2
, где B и γ — параметры затухания. в зависимости от конкретной конструкции лазера и состояния смещения. На рисунке 3.1.2 показаны примеры нормированной спектральной плотности шума интенсивности с тремя различными параметрами затухания.Частота релаксационных колебаний, использованная на этом рисунке, равна ΩR = 2π × 10 ГГц. На частотах, намного превышающих частоту релаксационных колебаний, динамическая связь между плотностью фотонов и плотностью носителей является слабой, и поэтому шум интенсивности становится менее зависимым от частоты.
Рисунок 3.1.2. Спектральная плотность шума нормированной интенсивности с частотой релаксационных колебаний 10 ГГц.
Помимо релаксационных колебаний, внешняя оптическая обратная связь с полупроводниковыми лазерами также может изменять спектральное распределение шума интенсивности [2, 3].Это может быть вызвано внешними оптическими отражателями, такими как разъемы, или другими оптическими устройствами, которые образуют внешние полости с помощью выходного зеркала полупроводникового лазера. Природа резонанса во внешней полости может быть достаточно сильной, чтобы вызвать чрезмерную флуктуацию плотности носителей в лазере и привести к сильной флуктуации оптической мощности. Отличительной особенностью шума интенсивности, вызванного оптической обратной связью, является то, что он имеет четко определенные периодические резонансные пики в частотной области, а период равен Δf = c / (2nL), где L — оптическая длина внешнего резонатора, c — скорость света, а n — показатель преломления внешней полости.
Другой источник шума оптической интенсивности является результатом преобразования частотного шума в интенсивность шума. В оптической системе нестабильность частоты в лазерном источнике может быть преобразована в шум интенсивности посредством частотно-зависимой передаточной функции, а также хроматической дисперсии. Например, резонаторы Фабри-Перо могут быть образованы несколькими отражателями (например, соединителями) в оптической системе, которые вносят частотно-зависимые характеристики передачи. Как показано на рисунке 3.1.3, когда частота лазера колеблется, эффективность передачи изменяется, вызывая колебания интенсивности.
Рисунок 3.1.3. Иллюстрация преобразования частотного шума в интенсивность шума посредством частотно-зависимой передачи.
Кроме того, если система передачи является дисперсионной, разные частотные компоненты будут приходить на детектор в разное время; интерференция между ними также вызывает преобразование частотного шума в интенсивность шума [4].
Как правило, интенсивность шума оптического излучения в оптической системе линейно пропорциональна оптической мощности сигнала; поэтому удобнее нормировать интенсивность шума на полную оптическую мощность.Относительная интенсивность шума ( RIN ) определяется как отношение между спектральной плотностью мощности шума и полной мощностью. На рисунке 3.1.4 показана блок-схема системы измерения RIN, которая состоит из широкополосного фотодетектора, электрического предусилителя и анализатора радиочастотного спектра.
Рисунок 3.1.4. Блок-схема системы для измерения RIN лазера.
Для определения характеристики RIN измеряемый лазерный диод работает в непрерывном режиме. Очевидно, что без шума интенсивности в лазере можно было бы увидеть только постоянную составляющую на анализаторе радиочастотного спектра.Из-за шума интенсивности флуктуация оптической мощности приводит к флуктуации фототока, и спектральная плотность этой флуктуации фототока измеряется анализатором радиочастотного спектра. Из-за принципа квадратичного детектирования электрическая мощность P ele , генерируемая на выходе фотодиода, пропорциональна квадрату принятой оптической мощности P opt , то есть Pele∝Popt2. Следовательно, RIN можно определить как отношение между спектральной плотностью мощности шума и мощностью сигнала в электрической области ,
(3.1.3) RIN (ω) = F {〈(Popt-Popt, ave) 2〉 Popt, ave2}
, где P opt, ave — средняя оптическая мощность, а F — преобразование Фурье. ℜ2 〈(Popt-Popt, ave) 2〉 — это среднеквадратичная флуктуация интенсивности, а его преобразование Фурье — это спектральная плотность мощности шума в электрической области, где ℜ — чувствительность детектора. ℜPopt — это фототок сигнала, а ℜ2Popt2 — электрическая мощность сигнала. Следовательно, определение RIN также можно просто выразить как
(3.1.4) RIN = SP (ω) ℜ2Popt, ave2
, где S P (ω) — измеренная спектральная плотность мощности электрического шума на анализаторе радиочастотного спектра. Фактически, если пренебречь шумом детектора, значение чувствительности фотодиода не влияет на результат измерения RIN, потому что и номинатор, и диноминтор уравнения 3.1.4 пропорциональны этой чувствительности.
Очевидно, спектральная плотность мощности шума S P (ω) увеличивается с увеличением квадрата средней оптической мощности Popt, ave2.RIN — удобный способ характеризовать качество оптического сигнала, и на результат не влияют погрешности оптического затухания в оптических системах; поэтому абсолютная калибровка чувствительности фотоприемника не требуется. Единица измерения RIN — [ Гц -1 ] или [ дБ / Гц ] в качестве относительной меры.
На практике RIN является удобным параметром для расчета характеристик оптической системы. Например, в аналоговой системе, если RIN является единственным источником шума, отношение сигнал / шум (в электрической области) можно легко определить с помощью
(3.1.5) SNR = m221∫0BRIN (f) df
, где m — индекс модуляции, а B — ширина полосы приемника.
Полупроводниковые лазеры хорошего качества обычно имеют уровни RIN ниже –155 дБ / Гц, и обычно сложно точно измерить RIN на таком низком уровне. Это требует, чтобы измерительная система имела гораздо более низкий уровень шума. Учитывая тепловой и квантовый шум в фотодиоде и тепловой шум в электрическом предусилителе, измеренное значение RIN можно выразить как
(3.1.6) RINmeasure = Sp + σshot2 + σth3 + σamp2ℜ2Popt2 = RINlaser + RINerror
, где
(3.1.7) RINerror = 2qℜPopt + kT (FAGA + FSA − 1) / GAℜ2Popt2 + 4kT / RL2 9ℜ2Popt из-за ошибки измерения 9ℜ2Po инструментальный шум. Где σshot2 = 2qℜPopt — спектральная плотность мощности дробового шума, σth3 = 4kT / RL — спектральная плотность мощности теплового шума на фотодиоде, а σamp2 = kT (FAGA + FSA − 1) / GA — эквивалентная спектральная плотность мощности шума, вносимая электрический предусилитель и анализатор ВЧ спектра. F A — коэффициент шума электрического предусилителя, F SA — коэффициент шума входного каскада анализатора спектра, а G A — коэффициент усиления предварительного усилителя.
На рисунке 3.1.5 показана ошибка измерения RIN, оцененная с ℜ = 0,75 A / W , R L = 50 Ом, T = 300 K , F A = F SA = 3 дБ и G A = 30 дБ.Погрешность измерения уменьшается с увеличением оптической мощности сигнала, и в конечном итоге квантовый шум становится преобладающим шумом в области высокой мощности. В этом конкретном случае для достижения уровня ошибки 180 дБ / Гц (который считается намного ниже, чем фактическое значение RIN лазера, равное –155 дБм / Гц), оптическая мощность сигнала должна быть выше 0 дБмВт.
Рисунок 3.1.5. Ошибка RIN в зависимости от мощности входного оптического сигнала.
Пример 3.1
Рассмотрим систему измерения RIN, показанную на рисунке 3.1.4. Полоса разрешения анализатора радиочастотного спектра установлена на 3 МГц. Когда лазерный источник находится под смещением постоянного тока при рабочем токе I B = 50 мА, измеренный уровень шума анализатора спектра составляет –80 дБмВт на частоте 1 ГГц. Затем лазерный источник модулируется синусоидой на f m = 1 МГц, в которой ток инжекции колеблется от порогового значения до 50 мА. Анализатор спектра измеряет очень узкий пик на частоте 1 МГц с амплитудой –5 дБмВт.Найдите значение RIN этого лазера на частоте 1 ГГц.
Решение
Обычно показание уровня мощности на анализаторе ВЧ спектра представляет собой мощность ВЧ в пределах разрешающей способности полосы пропускания. Следовательно, показание –80 дБмВт на анализаторе спектра в этом случае означает спектральную плотность мощности S p = –80 дБм / 3 МГц = –144,78 дБм / Гц.
Когда лазер имеет аналоговую модуляцию с индексом m , среднеквадратичная электрическая мощность, измеренная на анализаторе спектра, составляет –5 дБмВт.Поскольку это синусоидальная модуляция, спектральная полоса пропускания на частоте модуляции должна быть намного уже, чем полоса разрешения 3 МГц. Следовательно, фактическая полная электрическая мощность, преобразованная из оптического сигнала, должна составлять приблизительно –2 дБмВт (при индексе модуляции м = 1, эффективная мощность сигнала равна 1/2). Следовательно, RIN этого лазера при смещении 50 мА должно быть RIN = –144,78 — (–2) = 142,78 дБм / Гц.
ГЛАВА 1: ВВЕДЕНИЕ В ДАТЧИКИ | Расширение видения сенсорных материалов
, что чувствительные элементы должны быть линейными и бесшумными; однако при анализе конструкции сенсорной системы необходимо учитывать стоимость добавленной электроники.
Возможные преимущества концепции интеллектуального датчика:
-
меньшее обслуживание;
-
сокращено время простоя;
-
более высокая надежность;
-
отказоустойчивых систем;
-
адаптируемость для самокалибровки и компенсации;
-
более низкая стоимость;
-
меньший вес;
-
На
меньше соединений между несколькими датчиками и системами управления; и
-
менее сложная системная архитектура.
Эти преимущества интеллектуальных датчиков зависят от области применения. Распределение обработки сигналов по большой сенсорной системе, безусловно, оправдано для многих приложений, так что каждый сенсор имеет свою собственную калибровку, диагностику неисправностей, обработку сигналов и связь, тем самым создавая иерархическую систему. Инновации в сенсорной технологии, как правило, позволили объединить большее количество сенсоров в сеть или разработать более точные сенсоры или включить калибровку на кристалле.В целом новые технологии способствовали повышению производительности за счет повышения эффективности и точности распределения информации и снижения общих затрат. Однако эти улучшения производительности были достигнуты за счет увеличения сложности отдельных сенсорных систем. В настоящее время практическая полезность интеллектуальных датчиков, по-видимому, ограничивается приложениями, требующими очень большого количества датчиков.
РЕЗЮМЕ
Область сенсорной техники чрезвычайно широка, и ее дальнейшее развитие будет включать взаимодействие практически всех научных и технических дисциплин.Основные определения и терминология в этой главе были представлены для обеспечения некоторой последовательности в обсуждениях приложений и технологий датчиков, поскольку в определениях и классификациях датчиков существует значительная неоднозначность. В оставшейся части настоящего отчета используется система классификации датчиков, основанная на измеряемой величине или первичной входной переменной. Комитет признает, что альтернативные системы сенсорной таксономии могут быть полезны в определенных обстоятельствах, но для целей настоящего исследования вышеупомянутая схема была принята как наиболее практичный вариант.Чтобы ускорить внедрение появляющихся сенсорных материалов в новые приложения, критически важно, чтобы сообщество сенсорных материалов могло легко определять потребности в зондировании и нацеливаться на те физические явления, которые могут ощущаться материалами-кандидатами.
Определения терминов «датчик», «сенсорный элемент» и «сенсорная система», приведенные выше, были приняты комитетом для облегчения последовательного и последовательного анализа сенсорных технологий. Многие современные «сенсоры» на самом деле являются сенсорными системами, включающими в себя некоторую форму обработки сигналов.Интеграция функций датчиков в систему «черный ящик», в которой техническая сложность эффективно скрыта от пользователя, является растущей тенденцией в разработке датчиков. Особый интерес представляет концепция интеллектуального зондирования, которая создает новые возможности для использования новых материалов в датчиках. , например, сняв ограничение на то, что сенсорные элементы должны быть линейными и бесшумными (хотя рентабельность такого подхода будет зависеть от области применения). Поскольку современные сенсоры включают в себя гораздо больше, чем просто трансдукционный материал, существует множество возможностей для внедрения новые материалы в сенсорных системах, хотя в этом отчете основное внимание уделяется материалам преобразователей.
ССЫЛКИ
Gimzewski, J.K., C. Gerber, and E. Meyer. 1994. Наблюдения за химической реакцией с помощью микромеханического сенсора. Письма по химической физике 217 (5/6): 589.
Göpel, W., J. Hesse, J.N. Земель, ред. 1989. Датчики: всесторонний обзор, Vol. 1. Нью-Йорк: ВЧ.
Инструментальное общество Америки. 1975. Номенклатура и терминология электрических преобразователей. Стандарт ANSI MC6.1. Парк Исследований Треугольника, Северная Каролина: Инструментальное общество Америки.
Лев, К.С. 1969. Преобразователи — проблемы и перспективы. IEEE Transactions по промышленной электронике 16 (1): 2–5.
Миддлхук С., Д.Дж.У. Ноорлаг. 1982. Трехмерное изображение входных и выходных преобразователей. Датчики и исполнительные механизмы 2 (1): 29–41.
Датчики. 1992. 1993 Руководство покупателя. Датчики: журнал машинного восприятия 9 (12).
Кинетика взаимодействия малых молекул с мембранными белками в отдельных клетках, измеренная с помощью механической амплификации
Sci Adv.2015 окт; 1 (9): e1500633.
, 1, 2 , 1 , 1, 2 , 1 , 3, * и 1, 2, 3, * Ян Гуань1 Центр биоэлектроники и биосенсоров, Институт биодизайна, Университет штата Аризона, Темпе, Аризона 85287, США.
2 Школа электротехнической, компьютерной и энергетической инженерии, Университет штата Аризона, Темпе, Аризона 85287, США.
Xiaonan Shan
1 Центр биоэлектроники и биосенсоров, Институт биодизайна, Университет штата Аризона, Темпе, Аризона 85287, США.
Фенни Чжан
1 Центр биоэлектроники и биосенсоров, Институт биодизайна, Университет штата Аризона, Темпе, Аризона 85287, США.
2 Школа электротехнической, компьютерной и энергетической инженерии, Университет штата Аризона, Темпе, Аризона 85287, США.
Shaopeng Wang
1 Центр биоэлектроники и биосенсоров, Институт биодизайна, Университет штата Аризона, Темпе, AZ 85287, США.
Hong-Yuan Chen
3 Государственная ключевая лаборатория аналитической химии для наук о жизни, Школа химии и химической инженерии, Нанкинский университет, Нанкин 210093, Китай.
Нунцзянь Тао
1 Центр биоэлектроники и биосенсоров, Институт биодизайна, Университет штата Аризона, Темпе, Аризона 85287, США.
2 Школа электротехнической, компьютерной и энергетической инженерии, Университет штата Аризона, Темпе, Аризона 85287, США.
3 Государственная ключевая лаборатория аналитической химии для наук о жизни, Школа химии и химической инженерии, Нанкинский университет, Нанкин 210093, Китай.
1 Центр биоэлектроники и биосенсоров, Институт биодизайна, Университет штата Аризона, Темпе, Аризона 85287, США.
2 Школа электротехнической, компьютерной и энергетической инженерии, Университет штата Аризона, Темпе, Аризона 85287, США.
3 Государственная ключевая лаборатория аналитической химии для наук о жизни, Школа химии и химической инженерии, Нанкинский университет, Нанкин 210093, Китай.
Поступила 18 мая 2015 г .; Принято, 2015 г. 14 сентября.
Это статья в открытом доступе, распространяемая в соответствии с условиями некоммерческой лицензии Creative Commons Attribution, которая разрешает использование, распространение и воспроизведение на любом носителе, при условии, что в результате будет использовано , а не для коммерческих целей. преимущество и при условии, что оригинальная работа процитирована должным образом. Эта статья цитировалась в других статьях в PMC.- Дополнительные материалы
-
Дополнительные материалы к этой статье доступны по адресу http: // advances.sciencemag.org/cgi/content/full/1/9/e1500633/DC1
Взаимодействие WGA и гликопротеинов в живой клетке
Рис. S1. Взаимодействие АЗП и гликопротеинов в живой клетке.
Взаимодействие антител против EGFR с EGFR в фиксированных клетках A431
Рис. S2. Взаимодействие антител против EGFR с EGFR в фиксированных клетках A431.
Отрицательный контроль взаимодействия нАХР и ацетилхолина
Рис. S3. Отрицательный контроль взаимодействия нАХР и ацетилхолина.
иммунофлуоресценция нАХР
Рис.S4. Фазово-контрастные и иммунофлуоресцентные изображения nAChR-положительных и nAChR-отрицательных клеток.
Спектр мощности шума движения края соты фиксированной соты CP-D
Рис. S5. Спектр мощности шума движения края ячейки фиксированной ячейки CP-D.
Деформация мембраны, вызванная молекулярным связыванием, вдоль краев клеток
Рис. S6. Деформация мембран по краям клеток, вызванная молекулярным связыванием.
Калибровка интенсивности дифференциальной визуализации и движения края ячейки
Рис.S7. Калибровка дифференциального обнаружения движения края ячейки.
Дифференциальный метод обнаружения
Рис. S8. Схема, иллюстрирующая процедуру метода дифференциального обнаружения.
Статистический анализ взаимодействий WGA и гликопротеинов на фиксированных клетках CP-D
Таблица S1. Кинетика связывания между WGA и гликопротеином на разных фиксированных клетках CP-D.
Статистический анализ взаимодействий WGA и гликопротеинов на живых клетках CP-D
Таблица S2.Кинетика связывания между WGA и гликопротеином на разных живых клетках CP-D.
Статистический анализ взаимодействия ацетилхолина и nAChR на фиксированных клетках SH-EP1-hα4β2.
Таблица S3. Кинетика связывания ацетилхолина и nAChR на разных фиксированных клетках SH-EP1-hα4β2.
Ссылка ( 43 )
Резюме
Измерение взаимодействий малых молекул с мембранными белками в отдельных клетках имеет решающее значение для понимания многих клеточных процессов и для скрининга лекарств.Однако создание такой возможности было сложной задачей. Мы показываем, что молекулярные взаимодействия с мембранными белками вызывают механическую деформацию клеточной мембраны, а мониторинг деформации в реальном времени с субнанометровым разрешением позволяет количественно анализировать кинетику взаимодействия малых молекул с мембранными белками в отдельных клетках. Эта новая стратегия обеспечивает механическое усиление малых сигналов связывания, что позволяет обнаруживать взаимодействия малых молекул с мембранными белками.Эта возможность вместе с пространственным разрешением также позволяет изучать гетерогенную природу клеток путем анализа изменчивости кинетики взаимодействия между разными клетками и между различными областями одной клетки.
Ключевые слова: измерение взаимодействия малых молекул с мембранными белками in situ, одиночная клетка, обнаружение без меток, измерение кинетики, оптическая визуализация, механическое усиление
ВВЕДЕНИЕ
Достижения в структурной биологии привели к экспоненциальному росту числа мембранных белков с определенной трехмерной (3D) структурой ( 1 ).Однако для понимания клеточных функций мембранных белков также необходимо определить кинетику взаимодействия мембранных белков с различными молекулами. Это связано с тем, что клетки выполняют множество функций, включая связь, посредством взаимодействия своих мембранных белков с молекулами внеклеточной среды. Количественная оценка взаимодействий мембранных белков с молекулами также имеет решающее значение для открытия и проверки лекарств, поскольку большинство лекарственных мишеней являются мембранными белками ( 2 , 3 ).Однако измерение взаимодействий молекул с мембранными белками в естественной липидной среде было сложной задачей.
Традиционный метод изучения молекулярных взаимодействий использует радиоактивные или флуоресцентные метки. Эти анализы конечных точек не предоставляют кинетических констант, которые необходимы для количественной оценки взаимодействий мембранных рецепторов, относящихся к биологическим функциям ( 4 , 5 ). Для определения кинетической информации текущая практика включает извлечение мембранных белков из клеток, очистку их от экстрактов, иммобилизацию очищенных белков на твердой поверхности, а затем воздействие на них лиганда для кинетического исследования ( 6 ).Эти процедуры не только трудоемки, но и подвержены изменению естественных функций мембранных белков, особенно интегральных мембранных белков, которые постоянно прикреплены к мембране. Кроме того, изоляция мембранных белков из их естественной клеточной среды препятствует изучению аллостерического эффекта в молекулярных взаимодействиях ( 7 ) и изучению гетерогенной природы клеток ( 8 , 9 ). Измерение кинетики связывания мембранных белков на клетках ( 10 ) или липидных бислоях ( 11 , 12 ) было продемонстрировано с помощью поверхностного плазмонного резонанса (SPR) и SPR с новыми наноструктурами, такими как наночастицы и нанопоры.Однако ограничение этих технологий состоит в том, что сигнал обнаружения уменьшается с увеличением массы молекулы, что приводит к трудностям в обнаружении небольших молекул, которые играют важную роль в клеточных функциях и представляют собой подавляющее большинство существующих лекарств.
Здесь мы сообщаем о наблюдении механической деформации клеток при взаимодействии белков клеточной мембраны с молекулами внеклеточной среды и демонстрируем анализ взаимодействий в отдельных клетках в реальном времени путем анализа механической деформации с субнанометровым разрешением.Были разработаны различные методы измерения деформации клеточной мембраны, в том числе оптический пинцет ( 13 , 14 ), количественная фазовая визуализация ( 15 , 16 ) и атомно-силовая микроскопия (АСМ) ( 17 — ). 19 ). Наши измерения основаны на методе дифференциального обнаружения, который обеспечивает субнанометрическую точность для отслеживания деформации края ячейки. Используя эту возможность, мы отслеживали кинетику взаимодействий как больших, так и малых молекул с мембранными белками, включая гликопротеины и ионные каналы в интактных клетках (фиксированных или живых), и получили кинетические константы связывания.Для больших молекул кинетические константы согласуются с константами, полученными с помощью метода плазмонной визуализации. Для малых молекул настоящий метод представляет собой первое кинетическое измерение, и прямое сравнение с другими методами невозможно, но константы равновесия, извлеченные из настоящего метода, согласуются с константами, полученными с помощью анализов радиоактивного мечения конечной точки. Возможность визуализации позволила нам выявить изменчивость от клетки к клетке и вариабельность от региона к региону в пределах одной и той же клетки.
РЕЗУЛЬТАТЫ
Принцип и экспериментальная установка для измерения механической деформации клеточной мембраны, связанной со связыванием молекул с мембранными белками, показаны на рис. Механическая деформация ожидается, потому что закон термодинамики предсказывает, что, когда молекулы связываются с поверхностью, поверхностное натяжение изменяется, что приводит к механическому отклику в клеточной мембране (). Согласно термодинамике, поверхностная концентрация (Γ) молекул, связанных на поверхности мембраны, равна
при постоянных температуре и давлении, где γ — поверхностное натяжение, а μ — химический потенциал молекул.Для идеальных растворов химический потенциал связан с объемной концентрацией, c , согласно
, где R — газовая постоянная, а T — температура. Из уравнений. 1 и 2, при данной концентрации аналита молекулярное связывание прямо пропорционально изменению поверхностного натяжения, и, таким образом, молекулярные взаимодействия с мембранными белками могут быть определены путем измерения механической деформации в мембране (). Обратите внимание, что согласно формуле.1, обнаруженная здесь механическая деформация не зависит от размера молекулы, поэтому метод в принципе работает как для больших, так и для малых молекул. Мы вернемся к этому в Обсуждении.
Обнаружение молекулярных взаимодействий с мембранными белками в клетках посредством механической амплификации.( A ) Схематическое изображение экспериментальной установки на основе инвертированного фазово-контрастного микроскопа с объективом 40 × фаза 2. ( B ) Дифференциальное оптическое обнаружение для точного отслеживания изменений краев клеток, вызванных взаимодействием анализируемого вещества и рецептора.( C ) Схема типичной кривой связывания, определенной по движению края ячейки. ( D ) Среднеквадратическое значение фиксированного изменения края ячейки составляет 0,46 нм. ( E ) Иллюстрация края ячейки изменяется во времени во время процесса связывания, где i, ii и iii соответствуют этапам, отмеченным в (C). Синий и красный прямоугольники на (B) и (E) — это области интереса для дифференциального обнаружения.
Для обнаружения связывания небольшого количества молекул очень важно иметь возможность измерять небольшие механические деформации клеточной мембраны.Хотя в принципе АСМ можно использовать для измерения деформации клеток ( 17 — 19 ), мы разработали простой метод, отслеживая движение края клетки с помощью оптического микроскопа с использованием алгоритма дифференциального обнаружения. По сравнению с АСМ, это неинвазивный и быстрый метод, позволяющий измерять несколько клеток одновременно. Алгоритм дифференциального обнаружения позволил нам достичь предела обнаружения движения края ячейки 0,5 нм (размер атома) с миллисекундным временным разрешением [ограничено только частотой кадров камеры устройства с зарядовой связью (ПЗС)].
показывает схематическую иллюстрацию экспериментальной установки на основе инвертированного оптического микроскопа. Край клетки четко виден с помощью обычного фазово-контрастного изображения, как показано на. Прямоугольная область интереса (ROI) определяется таким образом, что край ячейки проходит через центр прямоугольника, разделяя ROI на две равные половины: одна половина находится внутри ячейки, а вторая половина находится за пределами ячейки. Обозначим интенсивности двух половин как A и B .Если клетка расширяется после молекулярного связывания (), тогда A уменьшается, а B увеличивается (). Мы измеряем дифференциальную интенсивность изображения ( A — B ) / ( A + B ) и используем его для определения движения края ячейки в каждом месте. Связь между перемещением края ячейки и ( A — B ) / ( A + B ) калибруется с использованием процедуры, описанной в дополнительных материалах. Следуя за движением края ячейки, мы получаем кинетику связывания, как показано на.
Мы называем этот метод отслеживания края дифференциальным оптическим обнаружением. Дифференциальное оптическое обнаружение вычитает общий шум в оптической системе, обеспечивая тем самым превосходный предел обнаружения. отображает уровень шума с течением времени, показывая стандартное отклонение 0,46 нм. Хотя в настоящей работе мы фокусируемся на фазово-контрастном изображении, этот алгоритм оптического обнаружения может быть легко применен к другим режимам оптического изображения, таким как светлое поле ( 20 ) и визуализация SPR ( 21 , 22 ).
Чтобы продемонстрировать способность метода обнаружения и количественной оценки взаимодействий молекул с мембранными белками, мы сначала изучили молекулярное связывание с гликопротеинами. Гликопротеины представляют собой наиболее распространенные мембранные белки с сахарными группами, простирающимися во внеклеточное пространство клеток, что имеет решающее значение для распознавания клетками и коммуникации с различными сигнальными молекулами посредством специфических взаимодействий ( 23 ). Важный пример специфических взаимодействий — между лектинами (белками, которые связываются и распознают определенные сахарные структуры) и гликопротеинами на клеточных мембранах.Мы изучили агглютинин зародышей пшеницы (WGA; молекулярная масса 36 кДа), лектин, который может специфически распознавать N -ацетилглюкозамин (GlcNAc) и группы сиаловой кислоты на клетках CP-D (CP-18821), полученных из пищевода Барретта.
(A и B) показывает фазово-контрастные изображения клеток CP-D, прикрепленных к предметному стеклу. Измерение проводили, сначала пропуская 1 × фосфатно-солевой буферный раствор (PBS) над клетками при скорости потока 350 мкл / мин в течение 30 с для получения базовой линии. Затем в течение 90 с вводили WGA в 1 × PBS, чтобы обеспечить связывание (ассоциацию) WGA с гликопротеинами на поверхности клетки.Во время процесса ассоциации край клетки перемещался наружу, как показано на C и D. После процесса ассоциации раствор WGA был переключен на 1 × PBS, чтобы позволить связанному WGA отделиться от клеток. (C и D) показывает, что край клетки вернулся в исходное положение во время процесса диссоциации. Сообщалось о механических ответах клеточных мембран на связывание WGA с использованием микропипеток ( 24 , 25 ), что подтверждает настоящее наблюдение. Путем глобального сопоставления данных с кинетикой первого порядка были найдены константа скорости ассоциации ( k на ), константа скорости диссоциации ( k от ) и константа диссоциации ( K D ). быть 1.09 ± 0,02 × 10 5 M −1 с −1 , 2.20 ± 0,01 × 10 −3 с −1 и 19,0 ± 0,5 нМ соответственно. Кинетика связывания WGA и гликопротеина на большем количестве клеток показана в таблице S1. Связывание WGA с гликопротеинами на живых клетках также было обнаружено с помощью того же метода (рис. S1 и таблица S2). Результаты как для фиксированных, так и для живых клеток согласуются с результатами, полученными с помощью метода плазмонной визуализации, недавно опубликованного нами ( 10 ). Другой пример — связывание антитела против рецептора эпидермального фактора роста (EGFR) (молекулярная масса 134 кДа) с EGFR на клетках A431 (рис.S2), который описан в разделе S-2 дополнительных материалов. В отличие от метода плазмонной визуализации, который плохо подходит для измерения малых молекул ( 26 , 27 ), настоящее механическое усиление может обнаруживать как большие, так и маленькие молекулы.
Взаимодействия крупных молекул: взаимодействие WGA с гликопротеинами.( A и B ) Фазово-контрастные изображения фиксированных клеток CP-D со связыванием WGA [0,56 мкМ (20 мкг / мл) (A) и 0.14 мкМ (5 мкг / мл) (B)]. Белыми стрелками отмечены анализируемые ячейки. ( C и D ) Усредненное перемещение края клетки по всей клетке (черные точки) и общая аппроксимация (красные кривые) для 0,56 мкМ (20 мкг / мл) (C) и 0,14 мкМ (5 мкг / мл) ( Г) WGA соответственно. Шкала 10 мкм.
Чтобы продемонстрировать способность настоящего метода к обнаружению связывания малых молекул, мы исследовали связывание ацетилхолина (молекулярная масса 146,2 дальтон) с никотиновыми ацетилхолиновыми рецепторами (nAChR) с использованием сконструированных клеток SH-EP1, которые экспрессировали человеческие рецепторы α4β2 (hα4β2).nAChR являются одними из наиболее изученных мембранных рецепторов из-за их критической роли в нейротрансмиссии и никотиновой зависимости ( 28 , 29 ). Определение кинетики связывания нейромедиаторов, таких как ацетилхолин, с nAChR в нейронах важно для базовой нейробиологии и для клинической оценки никотиновой зависимости ( 30 ). Аффинность связывания нейротрансмиттер-рецептор, указанная в учебниках и литературе, была получена с помощью метода радиоактивного мечения, который требует лизиса нейронов ( 31 ) и инкубации с нейротрансмиттерами и, таким образом, не может измерить аффинность связывания каждой отдельной клетки.Поскольку радиоактивные или другие методы мечения являются анализами конечной точки, они не обеспечивают кинетических констант.
Показано изображение клеток SH-EP1-hα4β2, где белой стрелкой отмечена анализируемая клетка. Буфер (1 × PBS) сначала вводили для протекания через ячейку в течение 25 с, а затем буфер заменяли раствором ацетилхолина в 1 × PBS. После объединения раствор ацетилхолина при каждой концентрации снова переключали на 1 × PBS, чтобы обеспечить диссоциацию. Вышеуказанная процедура была повторена для различных концентраций ацетилхолина.Как показано на фиг.4, край клетки расширяется во время фазы ассоциации и сокращается во время фазы диссоциации. также показывает, что степень размножения клеток во время процесса ассоциации увеличивается с концентрацией ацетилхолина, что ожидается для кинетики связывания первого порядка. Константы скорости ассоциации ( k на ) и диссоциации ( k от ) оказались равными 1,20 ± 0,01 × 10 −6 M −1 s −1 и 1,96 ± 0.02 × 10 -2 с -1 , соответственно, которые представляют собой первое прямое измерение кинетических констант для связывания нейромедиатора с nAChR в интактных клетках. Из значений k на и k на константа равновесной диссоциации ( K D = k off / k на ) была определена как 16,4 ± 0,2 нМ. Построив график зависимости реакции равновесия от концентраций ацетилхолина (), константа равновесия ( K D ) оказалась равной ~ 26 нМ, что согласуется с величиной, полученной с помощью кинетических измерений.Поскольку это первое кинетическое измерение связывания ацетилхолина с nAChR, мы не можем сравнивать наши результаты с другими эталонными технологиями или предшествующими данными. Однако определенная здесь константа равновесной диссоциации согласуется со средним значением K i , определенным с помощью анализов связывания радиолигандов, которые включали центрифугирование и образование осадка клеток ( 31 , 32 ).
Взаимодействия малых молекул: взаимодействие ацетилхолина с nAChR в клетках.( A ) Фазово-контрастное изображение фиксированных трансфицированных hα4β2 клеток SH-EP1, где белой стрелкой отмечена анализируемая клетка. Масштабная линейка 20 мкм. ( B ) Усредненное перемещение края клетки по всей клетке (черные точки) и общие результаты подгонки (красные кривые) в процессе связывания для ацетилхолина различных концентраций (снизу вверх: 1, 5, 20 и 100 нМ) . ( C ) Движение края клетки при равновесии в зависимости от концентрации ацетилхолина. Константа равновесия ( K D ) была определена как ~ 26 нМ путем аппроксимации данных с помощью изотермы Ленгмюра.
В качестве контрольного эксперимента мы провели измерение с фиксированными клетками SH-EP1 дикого типа, которые не имеют nAChR, экспрессируемых на поверхности клетки, и не наблюдали деформации клеточной мембраны (рис. S3). Уровни экспрессии nAChR в сконструированных клетках и клетках дикого типа были подтверждены с помощью иммунофлуоресцентной визуализации, которая показала высокий охват nAChR в клетках SH-EP1-hα4β2 и небольшой охват в клетках SH-EP1 дикого типа (рис. S4). . Уровень экспрессии nAChR составлял 110 nAChR / мкм 2 на трансфицированных клетках SH-EP1-hα4β2 ( 31 ).Эти результаты продемонстрировали, что механическая деформация сконструированных клеток SH-EP1 действительно была вызвана специфическим связыванием ацетилхолина с экспрессируемыми nAChR.
Настоящий метод не только позволяет количественно оценить кинетику связывания низкомолекулярных взаимодействий с мембранными белками в отдельных клетках, но также может исследовать межклеточные вариации кинетики связывания. (A и C) показаны фазово-контрастные изображения клеток SH-EP1-hα4β2, где числа в кружках отмечают анализируемые клетки.Клетки 1 и 2 культивировали на одном предметном стекле, а клетки 3 и 4 — на втором предметном стекле. Ответы этих клеток на 100 нМ ацетилхолина показаны в, B и D, и соответствующие кинетические константы приведены в, которые показывают значительные различия в кинетике связывания между клетками (кинетика связывания для большего количества клеток показана в таблице S2). Этот метод также позволяет нам исследовать кинетику связывания в разных местах одной и той же клетки. представляет собой графики кинетических кривых связывания ацетилхолина с nAChR в различных местах клетки, которые показывают большие вариации связывания.показывает максимальную деформацию мембраны по краю ячейки, где цвет представляет деформацию в нанометрах. Гетерогенное распределение рецепторов мембранных белков в клетках играет важную роль в клеточных функциях, включая взаимодействие между различными клетками ( 8 , 9 ).
Неоднородность низкомолекулярных взаимодействий с рецепторами клеточных мембран.( A и C ) Фазово-контрастные изображения фиксированных трансфицированных hα4β2 клеток SH-EP1, где числами отмечены анализируемые клетки.Масштабные линейки, 20 мкм. ( B ) Кинетика связывания клеток 1 и 2 (100 нМ ацетилхолина). ( D ) Кинетика связывания клеток 3 и 4 (100 нМ ацетилхолина). ( E ) Кинетика связывания в различных местах клетки в (F) (100 нМ ацетилхолина). ( F ) Фазово-контрастное изображение фиксированной клетки SH-EP1, трансфицированной hα4β2. Цвет показывает степень деформации края клеточной мембраны. Масштабная линейка 20 мкм.
Таблица 1
Константы скорости ассоциации ( k на ), константы скорости диссоциации ( k от ) и константы равновесия ( K D ) для четырех ячеек, как показано на.| k на (M −1 s −1 ) | k off (s −1 ) 61 D (нМ) | Ячейка 1 | 7,3 ± 0,2 × 10 5 | 1,61 ± 0,02 × 10 -2 | 22,0 ± 0,6 | Ячейка 2 9 ± 0,3 × 10 5 | 2.17 ± 0,03 × 10 −2 | 26,1 ± 0,7 | Ячейка 3 | 3,4 ± 0,2 × 10 5 | 2,98 ± 0,03 × 10 −2 | 88,0 ± 4,0 | | 5,8 ± 0,9 × 10 5 | 1,72 ± 0,01 × 10 −2 | 29,9 ± 0,5 | Среднее значение | 6,2 × 10 5 | 2,12 × 10 — 2 | 41,5 | SD | 2.1 × 10 5 | 0,62 × 10 −2 | 31,2 | |
ОБСУЖДЕНИЕ
Важность изучения молекулярных взаимодействий с мембранными белками побудила многие усилия по разработке безметочных технологий обнаружения в реальном времени анализ кинетики взаимодействия. Примеры включают микровесы на кристаллах кварца ( 33 ) и методы SPR ( 10 ). Однако сигналы этих методов пропорциональны массе молекулы, что затрудняет кинетическое измерение взаимодействий малых молекул с мембранными белками.Локализованный SPR в металлических наночастицах увеличивает предел обнаружения ( 34 ), а эффект экстраординарного оптического пропускания (EOT) от нанопор в металлической пленке предлагает уникальные плазмонные характеристики ( 35 ), которые показывают многообещающие возможности для чувствительного обнаружения молекулярных взаимодействий. Однако эти подходы не применялись для изучения молекулярных взаимодействий с мембранными белками в клетках, а их чувствительность зависит от молекулярной массы, что затрудняет изучение молекулярных взаимодействий для малых молекул.Насколько нам известно, настоящая работа является первым прямым измерением в реальном времени кинетики связывания малых молекул с мембранными белками в интактных клетках без мечения.
Метод основан на законе термодинамики, который является общим и применимым для анализа взаимодействий как больших, так и малых молекул с мембранными белками. Однако степень деформации мембраны зависит от механических свойств мембраны, которые не учитываются в формуле. 1. Механические вклады в свободную энергию клеточной мембраны включают энергию изгиба и натяжения Хельфриха ( 36 ).В присутствии мембранных белков с концентрацией ϕ свободная энергия также включает энтропийный вклад мембранных белков [ f (ϕ)] и взаимодействия между мембранными белками (Λ H ϕ) и мембраной, учитывая по ( 37 )
F = ∫ [κ2 (2H − C0) 2 + σ − ΛHφ + f (φ)] dA
(3)
где κ — модуль изгиба, H — среднее кривизна мембраны, C 0 — спонтанная кривизна, а Λ — коэффициент связи.Присутствие мембранных белков (последние члены в уравнении 3) влияет на модуль упругости мембраны, приводя к эффективному модулю изгиба
, что указывает на изменение модуля изгиба мембраны в присутствии белков. Эта деформация мембраны, вызванная связыванием белков, наблюдалась в различных мембранных системах ( 38 — 40 ).
Деформация мембраны, индуцированная связыванием лиганда, также ожидается из рассмотрения на молекулярном уровне. В общем, связывание лиганда с белковым рецептором в клеточной мембране приводит к конформационному изменению рецептора, которое влияет на взаимодействие рецептора с окружающими липидными молекулами и, таким образом, вызывает деформацию мембраны.Например, в случае связывания ацетилхолина с nAChR конформационные изменения nAChR включают перемещение гидрофобных остатков от поры, вращение гидрофильных остатков по направлению к поре и увеличение размера пор с ~ 3 до ~ 8 Å ( 30 ). Ожидается, что перераспределение гидрофобных / гидрофильных остатков и увеличение размера рецептора приведет к искажению клеточной мембраны через член связывания рецептор-липид (Λ H ϕ) в уравнениях. 3 и 4. Исходя из этих соображений, сигнал связывания, обнаруживаемый с помощью настоящего подхода, зависит от конкретных взаимодействий лиганд-рецептор и рецептор-липид и общих механических свойств мембраны, которые не обязательно зависят от размера лиганда.Для исследований кинетики связывания нужно только предположить, что деформация мембраны пропорциональна количеству лиганда, связывающегося с мембранным рецептором, что, как ожидается, будет верным при низкой плотности лиганда. Наблюдаемое согласие между настоящим исследованием и литературой подтверждает предположение для изучаемых здесь систем.
В дополнение к механической амплификации, субнанометровое разрешение, предлагаемое алгоритмом дифференциального обнаружения, обеспечивает чувствительное обнаружение молекулярного связывания в отдельных клетках, что также важно для успеха настоящего метода.Предел обнаружения в субнанометровом масштабе может быть дополнительно улучшен за счет удаления шума от различных источников. На рисунке S5 показан спектр мощности шума движения края соты. На низких частотах (<10 Гц) спектр мощности можно описать линейной функцией с наклоном, близким к -2, что указывает на броуновское движение как основной источник шума. Однако на более высоких частотах он значительно отличается от поведения броуновского шума, что показывает важность шума от других источников, таких как источник света и камера.Измеряемая здесь механическая деформация зависит от прочности связывания и покрытия поверхности мембранным белком, а также от механических свойств клеточной мембраны. В настоящем исследовании пределы обнаружения WGA и ацетилхолина оцениваются в 1,5 нМ для клеток CP-D и 1,4 нМ для клеток SH-EP1-hα4β2 на основе критерия 3 SD.
Пространственное разрешение на уровне отдельной клетки позволяет нам количественно оценить не только межклеточную вариацию кинетики связывания (от A до D), но также вариацию от области к области в пределах одной клетки.Например, показывает относительно большие сигналы связывания ячейки в областях, где ячейка взаимодействует с соседней ячейкой (красный цвет внутри). Точно так же соответствующие области соседней клетки также показывают более сильные сигналы связывания, чем другие области (рис. S6A). Это явление также наблюдалось в других клетках (рис. S6B). Мы связываем наблюдаемые вариации от региона к региону с локальными вариациями покрытия мембранным белком и механических свойств клетки.
Настоящий метод демонстрирует возможность изучения кинетики связывания малых молекул с мембранными белками на фиксированных и живых клетках.Фиксация параформальдегидом (PFA) является обычной практикой для фиксации клеток и широко используется в иммунофлуоресценции и иммуноферментных анализах. Литература также показала, что фиксация PFA не влияет на связывание nAChR (α4β2) ( 41 ). Мы обнаружили сходную кинетику связывания для фиксированных и живых клеток ( 41 ) и что кинетика связывания или константы равновесия, полученные с помощью настоящего метода, согласуются с литературными сообщениями ( 10 , 31 , 32 ).Эти наблюдения показывают, что фиксация не влияет на кинетику связывания для изучаемых здесь систем. Однако фиксация в целом может влиять на активность белков и должна учитываться при интерпретации данных. Сходство кинетики связывания фиксированных и живых клеток также предполагает, что последующий эффект в исследуемых здесь клетках не оказал значительного влияния на кинетику связывания. Несмотря на успех, индуцированные молекулярным связыванием последующие эффекты, такие как перестройки цитоскелета ( 42 ), необходимо учитывать при применении настоящего метода для изучения молекулярного связывания в живых клетках.
Наконец, метод основан на оптической микроскопии, которая проста и совместима с различными методами оптической визуализации, включая флуоресцентную визуализацию. Эти возможности будут полезны для скрининга лекарств и анализа различных клеточных процессов, в которых участвуют мембранные белки. Последнее особенно важно, потому что по мере экспоненциального увеличения числа трехмерных структур мембранных белков изучение кинетики взаимодействия молекул с мембранными белками становится все более важным.
МАТЕРИАЛЫ И МЕТОДЫ
Материалы
WGA и хлорид ацетилхолина были приобретены у Sigma-Aldrich. Моноклональные антитела против EGFR были приобретены у EMD Millipore (номер по каталогу 05-101). Первичные моноклональные антитела крысы к субъединице α4 nAChR (клон 299) и козий иммуноглобулин G крысы (IgG) Alexa Fluor 488 были приобретены у Abcam Inc. Для всех экспериментов по связыванию в качестве буфера использовали 1 × PBS (pH 7,4). Все образцы были приготовлены в буфере 1 × PBS.
Клеточные культуры
Клетки SH-EP1-hα4β2 культивировали во влажном инкубаторе при 37 ° C, 5% CO 2 и относительной влажности 70%.Клетки культивировали в 25-сантиметровой колбе 2 , содержащей модифицированную по Дульбекко среду Игла (DMEM; Lonza) с 10% фетальной бычьей сывороткой (Life Technologies) и пенициллин-стрептомицин (BioWhittaker). Когда было достигнуто примерно 80% слияния, клетки пассировали с 0,05% трипсин-ЭДТА (Life Technologies).
КлеткиCP-D культивировали в инкубаторе при 37 ° C, 5% CO 2 и относительной влажности 70%. Клетки культивировали в колбе 2 объемом 25 см, содержащей 1 × Keratinocyte-SFM (Life Technologies) и пенициллин-стрептомицин (BioWhittaker).Когда было достигнуто примерно 80% слияния, клетки пассировали с 0,05% трипсин-ЭДТА (Life Technologies).
Клетки A431 культивировали в инкубаторе при 37 ° C, 5% CO 2 и относительной влажности 70%. Клетки культивировали в 25-см колбе 2 , содержащей DMEM (Lonza) с пенициллин-стрептомицином (BioWhittaker). Когда было достигнуто примерно 80% слияния, клетки пассировали с 0,05% трипсин-ЭДТА (Life Technologies).
Для экспериментов клетки культивировали в течение ночи на предметных стеклах без покрытия (микропокровное стекло 22 × 60 мм, VWR), помещенных поверх силиконовой лунки (FlexiPERM, Greiner Bio-One), чтобы позволить клеткам прикрепиться к поверхности.Клетки на предметных стеклах также культивировали в инкубаторе при 37 ° C с 5% CO 2 и относительной влажностью 70%. Клетки инкубировали в 4% PFA в течение 10 мин при комнатной температуре для фиксации и дальнейшего анализа. Перед измерением маленькую силиконовую лунку заменили на самодельную полидиметилсилоксановую лунку (ПДМС) длиной 2 см, шириной 1 см и высотой 1 см.
Для измерения кинетики связывания на живых клетках клетки также культивировали в течение ночи на предметных стеклах без покрытия в силиконовой лунке и помещали в инкубатор при 37 ° C с 5% CO 2 и относительной влажностью 70%.Перед экспериментом маленькую силиконовую лунку заменили на самодельную PDMS-лунку большего размера. Измерение проводили сразу после замены культуральной среды на раствор для визуализации живых клеток (Life Technologies) для поддержания жизнеспособности клеток.
Установка оптической визуализации
Использовался инвертированный микроскоп (Olympus X81), оборудованный конденсатором фазы 2 и объективом фазы 2 40x, при освещении сверху ячеек с образцом. Для эксперимента с ацетилхолином дополнительный 0.5-кратный зум-объектив использовался с камерой CCD.
Иммунофлуоресцентная микроскопия
Для иммунофлуоресценции nAChR клетки фиксировали 4% PFA в PBS в течение 10 мин при комнатной температуре, а затем трижды промывали 1 × PBS. Фиксированные клетки инкубировали в 1% бычьем сывороточном альбумине в PBS при 37 ° C в течение 1 часа для блокирования клеточной поверхности. Добавляли первичное антитело, и клетки инкубировали в течение 30 мин при 37 ° C. Клетки промывали трижды буфером PBS, затем добавляли вторичное антитело, меченное Alexa Fluor 488, и клетки инкубировали еще 30 мин.Флуоресцентное изображение получали на той же установке для фазово-контрастного изображения после трехкратной промывки лунки PBS. Для иммунофлуоресцентного анализа использовали набор оптических фильтров (возбуждение, 420-480 нм; эмиссия, 515 нм).
Калибровка дифференциального оптического обнаружения движения края ячейки
Связь между движением края ячейки и измеренным изменением интенсивности дифференциального изображения была определена и откалибрована с использованием следующей процедуры.Чтобы уменьшить цифровой шум, связанный с конечным размером пикселей, плотность пикселей каждого изображения была увеличена в пять раз за счет добавления дополнительных пикселей с использованием подхода билинейной интерполяции. Расстояние между двумя пикселями в интерполированном изображении составляло 37 нм (Pike F032B CCD, Allied Vision Technologies). Край одной ячейки был выбран вручную, и был определен центр тяжести (O) ячейки (рис. S7A). Была установлена полярная система координат, в которой центроид служил полюсом. Перемещение края ячейки рассчитывали через каждые 1 мкм, начиная с 0 ° (рис.S7, B и C). Область интереса (1,85 × 3,70 мкм) в определенной точке края ячейки (точка A) была сдвинута на разное количество пикселей снаружи внутрь ячейки (рис. S7D) (перпендикулярно касательной линии в точке A). ), и были определены соответствующие изменения в дифференциальной интенсивности изображения (рис. S7E). Было обнаружено, что связь между разностной интенсивностью изображения и движением края ячейки (пикселей) является линейной в пределах определенного диапазона (рис. S7E, между двумя красными пунктирными линиями), который служил калибровочной кривой для определения движения края ячейки (механическое деформация) от дифференциальной интенсивности изображения (рис.S7F). Обратите внимание, что каждый пиксель соответствует 0,037 мкм, что позволило нам преобразовать движение в пикселях в микрометры.
Общая процедура метода дифференциального обнаружения для отслеживания движения края ячейки показана на рис. S8. Сначала вручную определялся край ячейки. Затем калибровочная кривая в каждом месте вдоль края ячейки была получена с использованием процедуры, описанной ранее. Используя эти калибровочные кривые, мы определили движения краев в разных местах краев ячеек.Хотя мы вручную выбрали границу ячейки в текущем исследовании, результат нечувствителен к выбору, пока граница находится в линейной области на рис. S7E, что составляет около 20 пикселей (0,74 мкм). Обычно анализ одной ячейки, включая выбор границы ячейки, калибровку, отслеживание границ и кинетические данные, с помощью портативного компьютера занимает около 5 минут. В будущем возможен автоматический выбор границы ячейки и более эффективный расчет.
Анализ данных
Кинетика связывания k на , k off и K D были получены путем аппроксимации экспериментальных кинетических кривых с помощью кинетической модели первого порядка (аппроксимация методом наименьших квадратов) , а ошибки — это ошибки подгонки.Среднее значение и стандартное отклонение кинетики связывания между различными клетками также были рассчитаны и показаны в таблицах S1-S3. Вариабельность кинетики связывания между разными клетками намного больше, чем ошибки подбора.
Благодарности
Мы благодарим J. Wu (Неврологический институт Барроу) за предоставленные клетки, а также H. Xu и Q. Chen (Amgen Inc.) за обсуждение. Финансирование: Мы благодарим Фонд Гордона и Бетти Мур и NSFC (грант № 21327008) за финансовую поддержку. Вклад авторов: Y.G., X.S. и N.T. разработал исследование; Ю.Г. провели исследование и проанализировали данные; X.S. и С. помогли с приборами; F.Z. подготовленные образцы; N.T. задумал проект; N.T. и H.-Y.C. руководил экспериментами; и Ю.Г. и Н. написал газету. Конкурирующие интересы: Авторы заявляют, что у них нет конкурирующих интересов. Доступность данных и материалов: Данные представленных анализов доступны по запросу у соответствующего автора.
ДОПОЛНИТЕЛЬНЫЕ МАТЕРИАЛЫ
Дополнительные материалы к этой статье доступны по адресу http://advances.sciencemag.org/cgi/content/full/1/9/e1500633/DC1
Взаимодействия WGA и гликопротеинов в живой клетке
Рис. . S1. Взаимодействие АЗП и гликопротеинов в живой клетке.
Взаимодействие антител против EGFR с EGFR в фиксированных клетках A431
Рис. S2. Взаимодействие антител против EGFR с EGFR в фиксированных клетках A431.
Отрицательный контроль взаимодействия нАХР и ацетилхолина
Рис.S3. Отрицательный контроль взаимодействия нАХР и ацетилхолина.
Иммунофлуоресценция нАХР
Рис. S4. Фазово-контрастные и иммунофлуоресцентные изображения nAChR-положительных и nAChR-отрицательных клеток.
Спектр мощности шума движения края соты фиксированной соты CP-D
Рис. S5. Спектр мощности шума движения края ячейки фиксированной ячейки CP-D.
Деформация мембраны, вызванная молекулярным связыванием, вдоль краев клеток
Рис. S6. Деформация мембран по краям клеток, вызванная молекулярным связыванием.
Калибровка интенсивности дифференциального изображения и движения края ячейки
Рис. S7. Калибровка дифференциального обнаружения движения края ячейки.
Дифференциальный метод обнаружения
Рис. S8. Схема, иллюстрирующая процедуру метода дифференциального обнаружения.
Статистический анализ взаимодействий WGA и гликопротеинов на фиксированных клетках CP-D
Таблица S1. Кинетика связывания между WGA и гликопротеином на разных фиксированных клетках CP-D.
Статистический анализ взаимодействий WGA и гликопротеинов на живых клетках CP-D
Таблица S2. Кинетика связывания между WGA и гликопротеином на разных живых клетках CP-D.
Статистический анализ взаимодействия ацетилхолина и nAChR на фиксированных клетках SH-EP1-hα4β2.
Таблица S3. Кинетика связывания ацетилхолина и nAChR на разных фиксированных клетках SH-EP1-hα4β2.
Ссылка ( 43 )
ССЫЛКИ И ПРИМЕЧАНИЯ
1. White S. H., Обзорная статья о биофизическом разрезе мембранных белков.Природа 459, г. 344–346 (2009). [PubMed] [Google Scholar] 2. Эрлих П., Обращение в патологию по химиотерапии: научные принципы, методы и результаты. Ланцет 2, 445–451 (1913). [Google Scholar] 3. Хопкинс А. Л., Грум К. Р., Геном, подверженный наркотикам. Nat. Rev. Drug Discov. 1, 727–730 (2002). [PubMed] [Google Scholar] 4. Коупленд Р. А., Помплиано Д. Л., Мик Т. Д., Время пребывания лекарства и мишени и его значение для оптимизации отведений. Nat. Rev. Drug Discov. 5, 730–739 (2006). [PubMed] [Google Scholar] 5.Суинни Д. К., Роль кинетики связывания в терапевтически полезном действии лекарственного средства. Curr. Opin. Drug Discov. Devel. 12, 31–39 (2009). [PubMed] [Google Scholar] 6. Р. Э. Эрнст, К. Н. Хай, Т. Р. Гласс, К. Чжао, в книге «Терапевтические моноклональные антитела: от лаборатории к клинике», , З. Ан, изд. (Уайли, Хобокен, Нью-Джерси, 2009 г.). [Google Scholar] 7. Changeux J.-P., 50 лет аллостерических взаимодействий: перипетии моделей. Nat. Rev. Mol. Cell Biol. 14, 819–829 (2013). [PubMed] [Google Scholar] 8.Альмендро В., Марусык А., Поляк К., Клеточная гетерогенность и молекулярная эволюция рака. Анну. Преподобный Патол. 8, 277–302 (2013). [PubMed] [Google Scholar] 10. Ван В., Ян Ю., Ван С., Нагарадж В. Дж., Лю К., Ву Дж., Тао Н., Измерение и картирование кинетики связывания мембранных белков в отдельных живых клетках без использования меток. Nat. Chem. 4, 846–853 (2012). [Бесплатная статья PMC] [PubMed] [Google Scholar] 11. Им Х., Виттенберг Н. Дж., Лесуффлер А., Линдквист Н. С., О С.-Х., Биочувствительность мембранных белков с помощью плазмонных массивов нанопор и липидных мембран, охватывающих поры.Chem. Sci. 1, 688–696 (2010). [Бесплатная статья PMC] [PubMed] [Google Scholar] 12. Мэйнард Дж. А., Линдквист Н. К., Сазерленд Дж. Н., Лесуффлер А., Уоррингтон А. Э., Родригес М., О С.-Х., Поверхностный плазмонный резонанс для высокопроизводительного лигандного скрининга мембраносвязанных белков. Biotechnol. Дж. 4, 1542–1558 (2009). [Бесплатная статья PMC] [PubMed] [Google Scholar] 14. Бамбардекар К., Клеман Р., Блан О., Шардес К., Ленн П.-Ф., Прямая лазерная манипуляция раскрывает механизм клеточных контактов in vivo.Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 112, 1416–1421 (2015). [Бесплатная статья PMC] [PubMed] [Google Scholar] 15. Парк Ю., Бест К. А., Бадизадеган К., Дасари Р. Р., Фельд М. С., Куриабова Т., Хенле М. Л., Левин А. Дж., Попеску Г., Измерение механики эритроцитов при морфологических изменениях. Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 107, 6731–6736 (2010). [Бесплатная статья PMC] [PubMed] [Google Scholar] 16. Парк Й., Бест К. А., Авт. Т., губернатор Н. С., Сафран С. А., Попеску Г., Суреш С., Фельд М. С., Метаболическое ремоделирование мембраны эритроцитов человека.Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 107, 1289–1294 (2010). [Бесплатная статья PMC] [PubMed] [Google Scholar] 17. Ву Г. Х., Цзи Х. Ф., Хансен К., Тундат Т., Датар Р., Кот Р., Хаган М. Ф., Чакраборти А. К., Маджумдар А., Происхождение наномеханического движения кантилевера, вызванного биомолекулярными взаимодействиями. Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 98, 1560–1564 (2001). [Бесплатная статья PMC] [PubMed] [Google Scholar] 18. Кросс С. Э., Цзинь Ю.-С., Рао Дж., Гимзевски Дж. К., Наномеханический анализ клеток онкологических больных.Nat. Nanotechnol. 2, 780–783 (2007). [PubMed] [Google Scholar] 19. Лулевич В., Циммер К. К., Хун Х.-с., Цзинь Л.-в., Лю Г.-у., Механика отдельных клеток обеспечивает чувствительные и количественные средства для исследования взаимодействий амилоид-β-пептида и нейронных клеток. Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 107, 13872–13877 (2010). [Бесплатная статья PMC] [PubMed] [Google Scholar] 20. Гуань Ю., Шань Х., Ван С., Чжан П., Тао Н., Обнаружение связывания молекул с помощью механического отклика оптических волокон, вызванного зарядом .Chem. Sci. 5, 4375–4381 (2014). [Бесплатная статья PMC] [PubMed] [Google Scholar] 21. Тао Н. Дж., Буссад С., Хуанг В. Л., Аречабалета Р. А., Д’Аньез Дж., Спектроскопия поверхностного плазмонного резонанса высокого разрешения. Rev. Sci. Instrum. 70, 4656–4660 (1999). [Google Scholar] 22. Шан Х., Патель У., Ван С., Иглесиас Р., Тао Н., Визуализация локального электрохимического тока с помощью поверхностного плазмонного резонанса. Наука 327, 1363–1366 (2010). [PubMed] [Google Scholar] 23. Делл А., Моррис Х. Р., Определение структуры гликопротеинов масс-спектрометрией.Наука 291, 2351–2356 (2001). [PubMed] [Google Scholar] 24. Смит Л., Хохмут Р. М., Влияние агглютинина зародышей пшеницы на вязкоупругие свойства мембраны эритроцитов. J. Cell Biol. 94, 7–11 (1982). [Бесплатная статья PMC] [PubMed] [Google Scholar] 25. Эванс Э., Люнг А., Адгезивность и жесткость мембраны эритроцитов по отношению к связыванию агглютининов зародышей пшеницы. J. Cell Biol. 98, 1201–1208 (1984). [Бесплатная статья PMC] [PubMed] [Google Scholar] 26. Лу Дж., Ван В., Ван С., Шань Х., Ли Дж., Тао Н., Электрохимическая импедансная спектроскопия на основе плазмонов: приложение к молекулярному связыванию. Анальный. Chem. 84, 327–333 (2012). [Бесплатная статья PMC] [PubMed] [Google Scholar] 27. Шань X., Фанг Ю., Ван С., Гуань Ю., Чен Х.-Й., Тао Н., Обнаружение зарядов и молекул с помощью самоорганизующихся нано-осцилляторов. Nano Lett. 14, 4151–4157 (2014). [PubMed] [Google Scholar] 28. Шуллер Х. М., Спровоцирован ли рак измененной передачей сигналов никотиновых ацетилхолиновых рецепторов? Nat. Преподобный Рак 9, 195–205 (2009). [PubMed] [Google Scholar] 29.Тали А., Корринджер П.-Дж., Гедин Д., Лестаж П., Ченжакс Ж.-П., Никотиновые рецепторы: аллостерические переходы и терапевтические мишени в нервной системе. Nat. Rev. Drug Discov. 8, 733–750 (2009). [PubMed] [Google Scholar] 30. Альбукерке Э. Х., Перейра Э. Ф. Р., Алкондон М., Роджерс С. В., Никотиновые ацетилхолиновые рецепторы млекопитающих: от структуры к функции. Physiol. Ред. 89, 73–120 (2009). [Бесплатная статья PMC] [PubMed] [Google Scholar] 31. Итон Дж. Б., Пэн Дж .-Х., Шредер К. М., Джордж А.А., Фрайер Дж. Д., Кришнан К., Булман Л., Куо Ю.-П., Стейнлейн О., Лукас Р. Дж., Характеристика человеческих α4β2-никотиновых ацетилхолиновых рецепторов, стабильно и гетерологично экспрессируемых в нативных никотиновых рецепторах SH-EP1 человеческих эпителиальных клетках. Мол. Pharmacol. 64, 1283–1294 (2003). [PubMed] [Google Scholar] 32. Йенсен А. А., Миккельсен И., Фрёлунд Б., Бройнер-Осборн Х., Фальч Э., Крогсгаард-Ларсен П., Гомологи карбамоилхолина: новые и сильные агонисты нейрональных никотиновых рецепторов ацетилхолина.Мол. Pharmacol. 64, 865–875 (2003). [PubMed] [Google Scholar] 33. Пей З., Сен-Гирон Дж., Кек К., Ингемарссон Б., Ааструп Т., Анализ углеводов на поверхности клеток в реальном времени с использованием биосенсора QCM: подход на основе лектинов. Биосенс. Биоэлектрон. 35, 200–205 (2012). [PubMed] [Google Scholar] 34. Анкер Дж. Н., Холл В. П., Ляндрес О., Шах Н. К., Чжао Дж., Ван Дайн Р. П., Биосенсор с плазмонными наносенсорами. Nat. Матер. 7, 442–453 (2008). [PubMed] [Google Scholar] 35. Эббесен Т. В., Лезец Х.Дж., Гэми Х. Ф., Тио Т., Вольф П. А., Необычайная оптическая передача через решетку отверстий субволновой длины. Природа 391, 667–669 (1998). [Google Scholar] 36. Хельфрих В., Упругие свойства липидных бислоев: теория и возможные эксперименты. Z. Naturforsch. C 28, 693–703 (1973). [PubMed] [Google Scholar] 37. Лейблер С., Неустойчивость кривизны мембран. J. Phys. 47, 507–516 (1986). [Google Scholar] 38. Циммерберг Я., Козлов М. М., Как белки вызывают искривление клеточной мембраны. Nat. Ред.Мол. Cell Biol. 7, 9–19 (2006). [PubMed] [Google Scholar] 40. Каллан-Джонс А., Бассеро П., Распределение липидов и белков мембран, обусловленное кривизной. Curr. Opin. Solid State Mater. Sci. 17, 143–150 (2013). [Google Scholar] 41. Вальехо Ю. Ф., Бюиссон Б., Бертран Д., Грин В. Н., Хроническое воздействие никотина активирует никотиновые рецепторы по новому механизму. J. Neurosci. 25, 5563–5572 (2005). [Бесплатная статья PMC] [PubMed] [Google Scholar] 42. Шабо В., Куэрье К. М., Эшер Э., Эймес В., Гранбуа М., Шарет П. Г., Биосенсинг на основе поверхностного плазмонного резонанса и живых клеток. Биосенс. Биоэлектрон. 24, 1667–1673 (2009). [PubMed] [Google Scholar] 43. Чжоу Ю., Гоэнага А.-Л., Хармс Б. Д., Цзоу Х., Лу Дж., Конрад Ф., Адамс Г. П., Шоберл Б., Нильсен Ю. Б., Маркс Дж. Д., Влияние внутренней аффинности на функциональное связывание и биологическую активность антител к EGFR. Мол. Рак Тер. 11, 1467–1476 (2012). [Бесплатная статья PMC] [PubMed] [Google Scholar]Произошла ошибка при настройке вашего пользовательского файла cookie
Этот сайт использует файлы cookie для повышения производительности.Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка вашего браузера для приема файлов cookie
Существует множество причин, по которым cookie не может быть установлен правильно. Ниже приведены наиболее частые причины:
- В вашем браузере отключены файлы cookie. Вам необходимо сбросить настройки своего браузера, чтобы он принимал файлы cookie, или чтобы спросить вас, хотите ли вы принимать файлы cookie.
- Ваш браузер спрашивает вас, хотите ли вы принимать файлы cookie, и вы отказались.Чтобы принять файлы cookie с этого сайта, нажмите кнопку «Назад» и примите файлы cookie.
- Ваш браузер не поддерживает файлы cookie. Если вы подозреваете это, попробуйте другой браузер.
- Дата на вашем компьютере в прошлом. Если часы вашего компьютера показывают дату до 1 января 1970 г., браузер автоматически забудет файл cookie. Чтобы исправить это, установите правильное время и дату на своем компьютере.
- Вы установили приложение, которое отслеживает или блокирует установку файлов cookie.Вы должны отключить приложение при входе в систему или проконсультироваться с системным администратором.
Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу. Чтобы предоставить доступ без файлов cookie потребует, чтобы сайт создавал новый сеанс для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файле cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в cookie; никакая другая информация не фиксируется.
Как правило, в файлах cookie может храниться только информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта. Например, сайт не может определить ваше имя электронной почты, пока вы не введете его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступа к остальной части вашего компьютера, и только сайт, который создал файл cookie, может его прочитать.
Руководство по открытию запроса на поддержкудля Cisco Kinetic для городов
Введение
В этом документе описывается, как обратиться за помощью в Центр технической поддержки (TAC) по проблемам или вопросам, связанным с Cisco Kinetic for Cities (CKC).
Как подать запрос в службу поддержки (SR) для получения помощи по вопросам или вопросам, связанным с CKC?
Чтобы иметь возможность поднять SR с помощью Cisco TAC, вам необходимо выполнить следующее:
- Имя пользователя и пароль Cisco.com (CCO).
Приведенные здесь шаги объяснят вам, как создать его, если у вас его нет. - Номер вашего контракта на поддержку.
Этот 9-значный номер должен был быть предоставлен вашим торговым представителем или партнером. Если вы не знаете номер, лучше всего связаться с ними.
Шаг 1. Создайте учетную запись CCO для доступа к сервисам Cisco
Если у вас уже есть учетная запись CCO для любого другого продукта или услуги Cisco, вы можете пропустить этот шаг и войти в систему с учетной записью, которая уже существует на этом шаге. 2.
Чтобы создать новую учетную запись, перейдите по адресу https://identity.cisco.com/ui/tenants/global/v1.0/enrollment-ui и заполните форму своей личной информацией, как показано на изображении. Лучше всего использовать ваш рабочий адрес электронной почты и служебный адрес, чтобы ускорить проверку контракта на более позднем этапе.
После того, как вы заполнили форму, нажмите Зарегистрироваться и дождитесь, пока на ваш почтовый ящик придет подтверждение по электронной почте.
Как только вы получите 6-значный код, введите его в форму, как показано на изображении.
Если все в порядке, вы успешно создали учетную запись CCO для доступа к сервисам Cisco.
Шаг 2. Свяжите свой номер контракта на поддержку со своим CCO ID
Этот шаг необходим для предоставления права на ваш идентификатор CCO для открытия SR с Cisco.
Перейдите к Support Case Manager (SCM) с помощью: https://mycase.cloudapps.cisco.com/case
Войдите в систему, используя свой существующий или только что созданный идентификатор CCO из шага 1., как показано на изображении.
Если вы получили это сообщение на следующем экране, это означает, что еще нет контрактов, связанных с вашим идентификатором CCO.
Если вы не видите сообщение или хотите добавить дополнительный контракт к своему идентификатору CCO, вы можете использовать следующую ссылку: https: // identity.cisco.com/ui/tenants/global/v1.0/profile-ui/personal
Нажмите Да , чтобы продолжить.
На следующей странице в разделе «Личные» укажите свой адрес и название компании, как показано на изображении.
После того, как вы правильно ввели информацию, перейдите в раздел Access Management , как показано на изображении.
В Access Management щелкните Add Access , чтобы связать ваш контракт на поддержку с вашим идентификатором CCO ID, как показано на изображении.
Выберите Создание дела TAC и RMA, Загрузка программного обеспечения, инструменты поддержки и соответствующий контент на Cisco.com , как показано на изображении.
На следующей странице выберите определение контракта и введите номер контракта, который вы получили от торгового представителя или который был указан в вашем заказе, как показано на изображении.
Как упоминалось ранее, если у вас нет номера, обратитесь к торговому представителю или партнеру.
После того, как вы выполнили эти шаги, нажмите Отправить , чтобы отправить форму.
Может пройти некоторое время, прежде чем контракт будет добавлен, и это зависит от того, насколько легко связать информацию идентификатора CCO ID с информацией контракта.
Если все пойдет хорошо, вы получите электронное письмо, в котором будет указано, что добавление контракта к вашему идентификатору CCO было одобрено, и вы можете перейти к шагу 3.
Шаг 3. Откройте запрос на поддержку для CKC
Последний шаг в процессе — открыть фактический SR для вашей проблемы / вопроса.
В случае, если вы захотите открыть еще один в будущем, вы, очевидно, можете пропустить первые два шага и сразу начать с этого.
Имейте в виду, что дела с высокой степенью серьезности (S1 / S2) можно открывать только по телефону.
По этим номерам можно связаться с Cisco Interactive Network (CIN). https://www.cisco.com/c/en/us/support/web/tsd-cisco-worldwide-contacts.html.
Убедитесь, что ваш идентификатор CCO, номер контракта и эта технология / вспомогательная технология готовы, чтобы ускорить процесс.
Перейдите к SCM, используя этот URL-адрес: https://mycase.cloudapps.cisco.com/case.
Войдите в систему, используя свой идентификатор CCO из шага 1., как показано на рисунке.
В SCM вы можете видеть существующие обращения для вашего CCO ID. Чтобы открыть новое дело, нажмите кнопку Открыть новое дело , как показано на изображении.
На следующем экране, если все идет хорошо с шагом 2, вы сможете ввести / найти номер вашего контракта для CKC.
В списке выберите любой действительный PID продукта CKC (который начинается с SC-CIM), как показано на изображении.
Затем выберите серьезность для вашего SR. Как уже упоминалось, проблемы S1 и S2 могут быть подняты только по телефону в CIN (см. Начало шага 3).
Добавьте заголовок и краткое описание проблемы / вопроса, имеющего отношение к CKC, как показано на изображении.
После завершения щелкните Вручную выберите технологию и выберите, как показано на изображении.
Это гарантирует, что SR немедленно получит правильную команду, которая хорошо осведомлена о CKC.
Затем выберите проблемную область, которая лучше всего соответствует типу запроса на обслуживание, который вы поднимаете, и заполните оставшуюся часть формы.
После завершения нажмите кнопку «Отправить», чтобы подать запрос на обслуживание.
Если все пойдет хорошо, необходимо создать запрос и направить его опытному инженеру, который свяжется с вами для работы над проблемой.
На протяжении всего оставшегося срока службы SR вы можете управлять всем через SCM.
В основном общение происходит по электронной почте. Убедитесь, что вы всегда включаете [email protected] в поле Кому: и номер SR в строке темы.
Таким образом, все коммуникации будут связаны с правильным регистром и будут видны в SCM, как показано на изображении.
Дополнительную информацию о SCM и способах его использования можно найти здесь: https://www.cisco.com/c/en/us/support/web/tools/scm/help/en-US/index.html
Фотодеградация левофлоксацина в водных и органических растворителях: кинетическое исследование
1. Д. Э. Кинг, Р. Мэлоун и С. Х. Лилли, Новая классификация и обновленная информация о хинолоновых антибиотиках, Am. Fam. Physician 61 (2000) 2741-2748. Искать в Google Scholar
2. К. Куммерер, Антибиотики в водной среде — обзор, часть 1, Chemosphere 75 (2009) 417-434; DOI: 10.1016 / j.bbr.2011.03.031. Поиск в Google Scholar
3. Дж. Бурхенн, М. Людвиг и М.Spiteller, Полярные продукты фотодеградации хинолонов, определенные с помощью ВЭЖХ / МС / МС, Chemosphere 38 (1999) 1279-1286; DOI: 10.1016 / j.bbr.2011.03.031.Поиск в Google Scholar
4. М. Будаи, П. Гроф, А. Циммер, К. Папай, И. Хлебович и К. Лудани, Фотодеградация липосом, индуцированная УФ-светом. инкапсулированные фторхинолоны: исследование MS, J. Photochem. Photobiol. A: Chem. 198 (2008) 268-273; DOI: 10.1016 / j.jphotochem.2008.04.004. Поиск в Google Scholar
5. Э. М. Тифенбахер, Э.Haen, B. Przybilla и H. Kurz, Фотодеградация некоторых хинолонов, используемых в качестве противомикробных терапевтических средств, J. Pharm. Sci. 83 (1994) 463-467; DOI: 10.1002 / jps.2600830403. Поиск в Google Scholar
6. К. Торниайнен, С. Таммилехто и В. Ульви, Влияние pH, типа буфера и концентрации лекарственного средства на фотодеградацию ципрофлоксацина, Int. J. Pharm. 132 (1996) 53-61; DOI: 10.1016 / j.bbr.2011.03.031.Поиск в Google Scholar
7. Х. Р. Н. Сальгадо, П. Р. Х. Морено, А. Л. Брага, Э.E. S. Schapoval, Фотодеградация спарфлоксацина и выделение продуктов его распада с помощью препаративной ВЭЖХ, Rev. Cienc. Ферма. Basica Apl. 26 (2005) 47-54. Поиск в Google Scholar
8. Х. Хубицка, Я. Кшек, Б. Зуромска, М. Вальчак, М. Зилевски и Д. Павловски, Определение фотостабильности и продуктов фотодеградации моксифлоксацина в наличие ионов металлов в растворах и твердой фазе. Кинетика и идентификация фотопродуктов, Photochem. Photobiol.11 (2012) 351-357; DOI: 10.1039 / c1pp05259d.Search in Google Scholar
9. Дж. Бурхенн, М. Людвиг, П. Николудис и М. Спителлер, Фотолитическое разложение фторхинолонкарбоновых кислот в водном растворе. Часть I: Первичные фотопродукты и период полураспада, Environ. Sci. Загрязнение. Res. 4 (1997) 10-15; DOI: 10.1007 / BF02986278. Поиск в Google Scholar
10. Дж. Бурхенн, М. Людвиг и М. Спителлер, Фотолитическое разложение фторхинолонкарбоновых кислот в водном растворе. Часть II: Выделение и структурное выяснение полярных фотометаболизма, Environ.Sci. Загрязнение. Res. 4 (1997) 61-67; DOI: 10.1007 / BF02986278. Поиск в Google Scholar
11. Г. Виола, Л. Фаччоло, М. Кантон, Д. Ведальди, Ф. Далл Аква, Г. Г. Алоизи, М. Амелия, А. Барбафина, Ф. Элисей и Л. Латтерини, Фотофизические и фототоксические свойства антибактериальных фторхинолонов левофлоксацина и моксифлоксацина, Chem. Биодайверы. 1 (2004) 782-801; DOI: 10.1002 / cbdv.2004. Поиск в Google Scholar
12. Й. Йошида, Э. Сато и Р. Морой, Продукты фотодеградации левофлоксацина в водном растворе, Arzneim.Форш. 43 (1993) 601-606.Search in Google Scholar
13. MRM Santoro, NM Kassab, AK Singh и ERM Kedor-Hackmam, Количественное определение гатифлоксацина, левофлоксацина, ломефлоксацина и пефлоксацина фторхинолоновых антибиотиков в жидких фармацевтических препаратах с помощью высокоэффективных фторхинолоновых антибиотиков. хроматография, J. Pharm. Биомед. Анальный. 40 (2006) 179–184; DOI: 10.1016 / j.jpba.2005.06.018. Поиск в Google Scholar
14. К. Г. Хэтчард, К. А. Паркер, Новый чувствительный химический актинометр.II. Ферриоксалат калия как стандартный химический актинометр, Proc. Royal Soc. (Лондон) А 235 (1956) 518-536; DOI: 10.1098 / rspa.1956.0102. Поиск в Google Scholar
15. Б. К. Сингх, Д. В. Парват и С. К. Шукла, Система быстрой идентификации по цветному тесту для скрининга поддельных фторхинолонов, E-J. Chem. 6 (2009) 377-384; DOI: 10.1155 / 2009/870286. Поиск в Google Scholar
16. М. Т. Монтеро, Д. Саиз, Р. Ситжес, Дж. Л. Васкес и Дж. Х. Боррелл, Влияние физико-химических свойств фторхинолонов на эффективность инкапсуляции в липосомы, Int.J. Pharm. 138 (1996) 113-120; DOI: 10.1016 / 0378-5173 (96) 04543-7. Поиск в Google Scholar
17. JAO Gonzalez, MC Mochon и FJB de la Rosa, Спектрофлуориметрическое определение левофлоксацина в таблетках, человеческой моче и сыворотке, Talanta 52 (2000) 1149-1156; DOI: 10.1016 / S0039- -9140 (00) 00484-7. Поиск в Google Scholar
18. Б. Де Витте, Х. В. Лангенхове, К. Хемельсоэт, К. Демеестер, П. Де Виспелер, В. В. Спейбрук и Дж. Девульф , Озонирование левофлоксацина в воде: параметры процесса, определяющие скорость, и выяснение путей реакции, Chemosphere 76 (2009) 683-689; DOI: 10.