
Автор эпигенетической теории: рассказ о возникновении эпигенетической теории эволюции
рассказ о возникновении эпигенетической теории эволюции
Ирина Лагунина: Продолжая рассказ в научной рубрике нашей программы об альтернативных эволюционных учениях, сегодня мы поговорим о теории, которая возникла в связи с развитием эпигенетики. Ее открытие показало, что наш организм может унаследовать даже те изменения, которые вызваны простыми внешними воздействиями среды.О том, как возникла эпигенетическая теории эволюции, рассказывает доктор биологических наук Александр Марков. С ним беседует Ольга Орлова
Ольга Орлова: Помимо эволюционных теорий, которые возникали в течение истории развития науки, открытия научные 20 века обогатили нас тоже возникновением новых эволюционных теорий. Одна из них так называемая эпигенетическая. Александр, расскажите, в чем ее суть и может ли она называться полноценной эволюционной теорией?
Александр Марков: Что касается эпигенетической теории, ее развивает сотрудник нашего Палеонтологического института Михаил Александрович Шишкин.
Ольга Орлова: Вы в этом смысле первооткрыватель, получаетесь.
Александр Марков: То, что я сейчас собираюсь пытаться более-менее понятным языком изложить суть этой теории может, быть я ошибаюсь, но насколько мне известно, это будет первая попытка. Во-первых, необходимо иметь в виду, что связь между генотипом и фенотипом не является однозначной. Уже давно ушли в прошлое времена, когда считалось, что каждому признаку организма соответствует какой-то ген.
Ольга Орлова: Что за такой признак отвечает такой-то ген, и мы можем установить прямую связь. На самом деле связь не прямая и может ее не быть.
Александр Марков: В действительности генетики, когда говорят о генах какого-то признака, то это своеобразный жаргон, который имеет на самом деле вполне нормальный правильный смысл, только он обычно не формулируется. Когда говорят, что есть ген серых глаз и ген карих глаз — это значит на самом деле, не то, что есть какой-то ген, сидит в хромосоме, и он вызывает развитие карих глаз у человека, а другой вариант этого гена вызывает развитие зеленых глаз. Ничего подобного. Для развития глаза нужно согласованное действие огромного количества генов, вообще говоря, всего генома. Если все эти гены не будут работать, то не будет не только глаза того или иного цвета, не будет организма жизнеспособного. Только на фоне всего генетического многообразия всей этой системы генетической может проявиться тот или иной признак. Так вот, когда говорят «ген карих глаз» имеется в виду, во-первых, всегда в сравнении между двумя состояниями признака, есть карие глаза, а есть зеленые глаза, и есть генетические различия. Например, в каком-то гене какой-то нуклеатид разный. Если он аденин, то получаются карие глаза, если гуанин, то получаются зеленые глаза, но только при условии наличия всех остальных генов на своих местах и так далее.
Когда говорят, что есть ген серых глаз и ген карих глаз — это значит на самом деле, не то, что есть какой-то ген, сидит в хромосоме, и он вызывает развитие карих глаз у человека, а другой вариант этого гена вызывает развитие зеленых глаз. Ничего подобного. Для развития глаза нужно согласованное действие огромного количества генов, вообще говоря, всего генома. Если все эти гены не будут работать, то не будет не только глаза того или иного цвета, не будет организма жизнеспособного. Только на фоне всего генетического многообразия всей этой системы генетической может проявиться тот или иной признак. Так вот, когда говорят «ген карих глаз» имеется в виду, во-первых, всегда в сравнении между двумя состояниями признака, есть карие глаза, а есть зеленые глаза, и есть генетические различия. Например, в каком-то гене какой-то нуклеатид разный. Если он аденин, то получаются карие глаза, если гуанин, то получаются зеленые глаза, но только при условии наличия всех остальных генов на своих местах и так далее.
Так же говорят о генах, определяющих те или иные особенности поведения. То есть можно выявить генетические различия, которые вызывают какие-то поведенческие различия. Например, сказать, что носители такого генетического варианта на 10% чаще проявляют агрессию, дерутся в школе по сравнению с носителями другого варианта. И говорят: вот это ген агрессии мы нашли. Но понятно, что на самом деле это жаргон такой. Никто на самом деле не думает, что этот ген берет и создает агрессию, речь идет всегда о различиях между генами, которым соответствуют различия между признаками. И что еще важно, даже если это учитывать, нужно помнить, что определение генами фенотипа, то есть признаков организма, всегда вероятностно, то есть не строгая вещь.
 Если у нас такой-то генетический вариант, значит будет точно такой же признак. Обычно речь идет о вероятностях.
Если у нас такой-то генетический вариант, значит будет точно такой же признак. Обычно речь идет о вероятностях.Ольга Орлова: Означает ли это, что, скажем, при наличии определенного генетического варианта у нас в пяти случаях будет этот признак, а в шестом уже не будет?
Александр Марков: Именно так обычно дело и обстоит.
Ольга Орлова: То есть мы должны просто ожидать вероятность появления этого признака в большей или меньшей степени.
Александр Марков: Совершенно верно. Дело в том, что геном как таковой – это молекулярный аппарат, который существует и действует на уровне одной клетки. Он внутри каждой клетки сидит в ядре и там действует. Это не некая общеорганизменная вещь — это в каждой клетке сидит. И работает, проявляется тоже на уровне клетки. И максимум, что может реально непосредственно определять – это строение, поведение клетки одной, да и то с большими оговорками. А как же из одной клетки, из оплодотворенного яйца развивается многоклеточный организм? Совершенно неправильно думать что в геноме записан какой-то чертеж строения будущего организма, что можно найти соответствие между каким-то участком генома и какой-то частью организма. Например, форма носа записана тут в геноме, а форма уха записана здесь. Вот таких соответствий нет в принципе. То есть геном – это не чертеж организма, а это в лучшем случае наиболее подходящая аналогия – это рецепт.
А как же из одной клетки, из оплодотворенного яйца развивается многоклеточный организм? Совершенно неправильно думать что в геноме записан какой-то чертеж строения будущего организма, что можно найти соответствие между каким-то участком генома и какой-то частью организма. Например, форма носа записана тут в геноме, а форма уха записана здесь. Вот таких соответствий нет в принципе. То есть геном – это не чертеж организма, а это в лучшем случае наиболее подходящая аналогия – это рецепт.
Представим себе рецепт приготовления какого-нибудь снадобья целебного: смешайте, подогрейте, помешайте, охладите, добавьте капельку крови лягушки, опять вскипятите.

Ольга Орлова: Если это уместно в этой аналогии спросить, мы все знаем, что у каждой хозяйки, несмотря на то, что рецепт один, а отвары, которые она будет варить, будут чуть-чуть отличаться в большей или меньшей степени. Справедливо ли это по отношению к аналогии с рецептом генома?
Александр Марков: Вы очень глубокое замечание сделали. Аналогии можно продолжать — это будет очень интересно. Хозяйка вносит что-то свое в данный рецепт. Когда она передает этот рецепт другой хозяйке, у другой хозяйки получается немножко по-другому. Это называется ненаследование приобретенных признаков.
Ольга Орлова: Эту изюминку вкуса, которая есть только в ее отваре, в ее блюде.
Александр Марков: Вот то, что не передается при передаче рецепта — это есть как раз аналог, почему у на эволюция идет по Дарвину, а не по Ламарку, почему приобретенные признаки не наследуются, потому что передаются рецепты голые. Эпигенетическая теория как раз делает акцент на том, что между генотипом и фенотипом нет строгого соответствия, что генотип как бы не предопределяет строго фенотип, что между генотипом и фенотипом лежит такая вещь, как очень сложный процесс индивидуального развития. Как из одной клетки образуется целый организм, реализация этого рецепта. Это очень сложный процесс, который только отчасти предопределен строением генотипа.
Очень важная идея, которая положена в основу эпигенетической теории, состоит в том, что индивидуальное развитие организма представляет сложную целостную систему, которая в результате естественного отбора очень устойчива и способна компенсировать разные нарушения и возмущения. Если, например, мы вмешаемся в развитие зародыша, то во многих случаях он все равно разовьется в нормальный взрослый организм, он скомпенсирует. Если мы разделим даже зародыш пополам на ранних стадиях развития, из каждой половинки может развиться нормальный взрослый организм. Но конечно, очень сильное воздействие может привести к уродству, к разным отклонениям. Но штука в том, что подчеркивает Михаил Александрович Шишкин в своей теории, что совершенно неважно, какое это воздействие — мутация или внешнее воздействие.
На само деле очень много экспериментальных данных свидетельствуют о том, что часто одни и те же изменения можно вызвать и путем мутаций, и путем внешних воздействий.
 Например, у мухи-дрозофилы известен целый ряд, огромное количество мутаций. Так вот, оказывается, что почти все проявления этих мутаций можно получить, просто издеваясь над эмбрионом, над личинкой так или иначе. Например, подвергая личинку развивающуюся тепловому шоку, мы можем получить таких же уродцев, которые получаются без нагревания в случае генетической мутации, те же самые отклонения, нарушения развития.
Например, у мухи-дрозофилы известен целый ряд, огромное количество мутаций. Так вот, оказывается, что почти все проявления этих мутаций можно получить, просто издеваясь над эмбрионом, над личинкой так или иначе. Например, подвергая личинку развивающуюся тепловому шоку, мы можем получить таких же уродцев, которые получаются без нагревания в случае генетической мутации, те же самые отклонения, нарушения развития.Ольга Орлова: А вот вопрос: допустим, экспериментально можно показать, но означает ли это, если мы получим один и тот же результат при разном воздействии мутационном или внешнем, означает ли это, что это на самом деле неважно, что это не сказывается на развитии организма и потомства и вида, популяции, каким способом получен этот дефект?
Александр Марков: Естественно, разница принципиальная. Если у мухи произошла мутация, она стала четырехкрылой получаться в нормальных условиях — это будет унаследовано, и ее потомство будет иметь четыре крыла, хотя этот признак, как и все мутации, имеет не стопроцентное проявление, может и не проявиться, но с большой вероятностью он будет проявляться. А если путем нагревания, если мы все время будем нагревать в каждом поколении, если, допустим, условия среды изменились, все время жарко там, где развивается личинка, то у нас получится то же самое, все мухи будут четырехкрылые из поколения в поколение. И с точки зрения естественного отбора, который смотрит на фенотип,а не на генотип, все равно, по какой причине у мухи четыре крыла.
А если путем нагревания, если мы все время будем нагревать в каждом поколении, если, допустим, условия среды изменились, все время жарко там, где развивается личинка, то у нас получится то же самое, все мухи будут четырехкрылые из поколения в поколение. И с точки зрения естественного отбора, который смотрит на фенотип,а не на генотип, все равно, по какой причине у мухи четыре крыла.
Ольга Орлова: И на этом основании автор эпигенетической теории Шишкин решил, что это неважно?
Александр Марков: И он решил, что эволюционные изменения начинаются не с мутаций, а с этих самых изменений нормального развития, которое происходит при резком изменении условий. Он считает, не только он, многие его предшественники, что индивидуальное развитие — это некая система, в которой есть один магистральный путь, по которому все особи идут к своему нормальному взрослому виду, и вот эта система, она так устроена, что у нее есть дополнительные ответвления, которые реализуются с малой вероятностью при различных нарушениях. И вот на эти боковые пути развития может свернуть либо в результате мутации, либо в результате какого-нибудь внешнего сильного воздействия, либо какого-то их сочетания. Причем разные воздействия могут приводить к одному и тому же эффекту.
И вот на эти боковые пути развития может свернуть либо в результате мутации, либо в результате какого-нибудь внешнего сильного воздействия, либо какого-то их сочетания. Причем разные воздействия могут приводить к одному и тому же эффекту.
Эпигенетическая теория личности Эриксона
Теория Эрика Эриксона возникла из практики психоанализа. Он трактует структуру личности так же, как и 3. Фрейд (как состоящую из «Оно», «Я», «Сверх-Я»), стадии развития личности, открытые Фрейдом, не отвергаются Эриксоном, а усложняются и как бы заново осмысливаются с позиции нового исторического времени. Психосоциальная концепция развития личности, разработанная Эриксоном, показывает тесную связь психики человека и характера общества, в котором он живет. Сравнение воспитания детей в индейских племенах с воспитанием белых американских детей позволило ему сделать вывод, что в каждой культуре имеется особый стиль воспитания детей — он всегда принимается матерью как единственно правильный. Этот стиль определяется тем, что ожидает от ребенка общество, в котором он живет. Каждой стадии развития человека соответствуют свои, присущие данному обществу, ожидания, которые человек может оправдать или не оправдать. Все детство человека — от рождения до юности — рассматривается Эриксоном как длительный период формирования зрелой психосоциальной идентичности, в результате которого человек приобретает объективное чувство принадлежности к своей социальной группе, понимание неповторимости своего индивидуального бытия. Ядром, вокруг которого строятся личность, является приобретение эго-идентичности.
Каждой стадии развития человека соответствуют свои, присущие данному обществу, ожидания, которые человек может оправдать или не оправдать. Все детство человека — от рождения до юности — рассматривается Эриксоном как длительный период формирования зрелой психосоциальной идентичности, в результате которого человек приобретает объективное чувство принадлежности к своей социальной группе, понимание неповторимости своего индивидуального бытия. Ядром, вокруг которого строятся личность, является приобретение эго-идентичности.
Эриксон ввел понятие «групповой идентичности«, которая формируется с первых дней жизни. Ребенок ориентирован на включение в определенную социальную группу, начинает понимать мир так, как эта группа. Но постепенно у ребенка формируется и «эгоидентичность«, чувство устойчивости и непрерывности своего «Я», несмотря на то, что идут многие процессы изменения. Формирование эго-идентичности — длительный процесс, включает ряд стадий развития личности. Каждая стадия характеризуется задачами этого возраста, а задачи выдвигаются обществом. Но решение задач определяется уже достигнутым уровнем психомоторного развития человека и духовной атмосферой общества, в котором человек живет.
Но решение задач определяется уже достигнутым уровнем психомоторного развития человека и духовной атмосферой общества, в котором человек живет.
- На стадии младенчества (1-я стадия) главную роль в жизни ребенка играет мать, она кормит, ухаживает, дает ласку, заботу, в результате чего у ребенка формируется базовое доверие к миру. Базовое доверие проявляется в легкости кормления, хорошем сне ребенка, нормальной работе кишечника, умении ребенка спокойно ждать мать (не кричит, не зовет, ребенок уверен, что мать придет и сделает то, что нужно). Динамика развития доверия зависит от матери. Здесь важно не количество пищи, а качество ухода за ребенком, важна уверенность матери в своих действиях. Если мать тревожна, невротична, если обстановка в семье напряженная, если ребенку уделяют мало внимания (например, ребенок в доме сирот), формируется базовое недоверие к миру, устойчивый пессимизм. Сильно выраженный дефицит эмоционального общения с младенцем приводит к резкому замедлению психического развития ребенка.
- 2-я стадия раннего детства связана с формированием автономии и независимости, ребенок начинает ходить, обучается контролировать себя при выполнении актов дефекации; общество и родители приучают ребенка к аккуратности, опрятности, начинают стыдить за «мокрые штанишки». Социальное неодобрение открывает глаза ребенка внутрь, он чувствует возможность наказания, формируется чувство стыда. В конце стадии должно быть равновесие «автономии» и «стыда». Это соотношение будет положительно благоприятным для развития ребенка, если родители не будут подавлять желания ребенка, не будут бить за провинности.
- В возрасте 3-6 лет, на 3-й стадии, ребенок уже убежден, что он личность, так как он бегает, умеет говорить, расширяет область овладения миром, у ребенка формируется чувство предприимчивости, инициативы, которое закладывается в игре ребенка. Игра очень важна для развития ребенка, т. е. формирует инициативу, творчество, ребенок осваивает отношения между людьми посредством игры, развивает свои психические возможности: волю, память, мышление и пр.
 Но если родители сильно подавляют ребенка, не уделяют внимания его играм, это отрицательно влияет на развитие ребенка, способствует закреплению пассивности, неуверенности, чувства вины.
Но если родители сильно подавляют ребенка, не уделяют внимания его играм, это отрицательно влияет на развитие ребенка, способствует закреплению пассивности, неуверенности, чувства вины. - В младшем школьном возрасте (4-я стадия) ребенок уже исчерпал возможности развития в рамках семьи, и теперь школа приобщает ребенка к знаниям о будущей деятельности, передает технологический опыт культуры. Если ребенок успешно овладевает знаниями, новыми навыками, он верит в свои силы, уверен, спокоен, но неудачи в школе приводят к появлению, а порой и к закреплению, чувства своей неполноценности, неверия в свои силы, отчаяния, потери интереса к учебе. При неполноценности ребенок как бы снова возвращается в семью, она для него убежище, если родители с пониманием стараются помочь ребенку преодолеть трудности в учебе. В случае, если родители лишь ругают и наказывают за плохие оценки, чувство неполноценности у ребенка закрепляется порой на всю его жизнь.
- В подростковом возрасте (5-я стадия) формируется центральная форма эгоидентичности.
 Бурный физиологический рост, половое созревание, озабоченность тем, как он выглядит перед другими, необходимость найти свое профессиональное призвание, способности, умения — вот вопросы, которые встают перед подростком, и это уже требования общества к подростку о самоопределении. На этой стадии заново встают все критические прошедшие моменты. Если на ранних стадиях у ребенка сформировалась автономия, инициатива, доверие к миру, уверенность в своей полноценности, значимости, то подросток успешно создает целостную форму эго-идентичности, находит свое «Я», признание себя со стороны окружающих. В противном случае происходит диффузия идентичности, подросток не может найти свое «Я», не осознает своих целей и желаний, происходит возврат, регрессия к инфантильным, детским, иждивенческим реакциям, появляется смутное, но устойчивое чувство тревоги, чувство одиночества, опустошенности, появляется постоянное ожидание чего-то такого, что может изменить жизнь, но сам человек активно ничего не предпринимает, появляется страх перед личным общением и неспособность эмоционально воздействовать на лиц противоположного пола, враждебность, презрение к окружающему обществу, чувство «непризнания себя» со стороны окружающих людей.
Бурный физиологический рост, половое созревание, озабоченность тем, как он выглядит перед другими, необходимость найти свое профессиональное призвание, способности, умения — вот вопросы, которые встают перед подростком, и это уже требования общества к подростку о самоопределении. На этой стадии заново встают все критические прошедшие моменты. Если на ранних стадиях у ребенка сформировалась автономия, инициатива, доверие к миру, уверенность в своей полноценности, значимости, то подросток успешно создает целостную форму эго-идентичности, находит свое «Я», признание себя со стороны окружающих. В противном случае происходит диффузия идентичности, подросток не может найти свое «Я», не осознает своих целей и желаний, происходит возврат, регрессия к инфантильным, детским, иждивенческим реакциям, появляется смутное, но устойчивое чувство тревоги, чувство одиночества, опустошенности, появляется постоянное ожидание чего-то такого, что может изменить жизнь, но сам человек активно ничего не предпринимает, появляется страх перед личным общением и неспособность эмоционально воздействовать на лиц противоположного пола, враждебность, презрение к окружающему обществу, чувство «непризнания себя» со стороны окружающих людей. Если человек нашел себя, то идентификация облегчается.
Если человек нашел себя, то идентификация облегчается. - На 6-й стадии (молодость) для человека актуальным становится поиск спутника жизни, тесное сотрудничество с людьми, укрепление связей со своей социальной группой, человек не боится обезличивания, он смешивает свою идентичность с другими людьми, появляется чувство близости, единства, сотрудничества, интимности с определенными людьми.
- Однако, если диффузия идентичности переходит и на этот возраст, человек замыкается, закрепляется изоляция, одиночество. 7-я — центральная стадия — взрослый этап развития личности. Развитие идентичности идет всю жизнь, идет воздействие со стороны других людей, особенно детей, они подтверждают, что ты им нужен. Положительные симптомы этой стадии: личность вкладывает себя в хороший, любимый труд и заботу о детях, удовлетворена собой и жизнью. Если не на кого излить свое «Я» (нет любимой работы, семьи, детей), то человек опустошается, намечается застой, косность, психологический и физиологический регресс.
 Как правило, такие отрицательные симптомы сильно выражены, если личность была подготовлена к этому всем ходом своего развития, если всегда были отрицательные выборы на этапах развития.
Как правило, такие отрицательные симптомы сильно выражены, если личность была подготовлена к этому всем ходом своего развития, если всегда были отрицательные выборы на этапах развития. - После 50 лет (8-я стадия) происходит создание завершенной формы эго-идентичности на основе всего пути развития личности, человек переосмысливает всю свою жизнь, осознает свое «Я» в духовных раздумьях о прожитых годах. Человек должен понять, что его жизнь — это неповторимая судьба, которую не надо переделывать, человек «принимает» себя и свою жизнь, осознает необходимость в логическом завершении жизни, проявляет мудрость, отстраненный интерес к жизни перед лицом смерти. Если «принятия себя и жизни» не произошло, человек чувствует разочарование, теряет вкус к жизни, чувствует, что жизнь прошла неверно, зря.
По мнению Эриксона, человек на протяжении жизни переживает восемь психосоциальных кризисов, специфических для каждого возраста, благоприятный или неблагоприятный исход которых определяет возможность последующего расцвета личности.
- Первый кризис человек переживает на первом году жизни. Он связан с тем, удовлетворяются или нет основные физиологические потребности ребенка ухаживающим за ним человеком. В первом случае у ребенка развивается чувство глубокого доверия к окружающему его миру, а во втором — недоверие к нему.
- Второй кризис связан с первым опытом обучения, особенно с приучением ребенка к чистоплотности. Если родители понимают ребенка и помогают ему контролировать естественные отправления, ребенок получает опыт автономии. Напротив, слишком строгий или слишком непоследовательный внешний контроль приводит к развитию у ребенка стыда или сомнений, связанных главным образом со страхом потерять контроль над собственным организмом.
- Третий кризис соответствует второму детству. В этом возрасте происходит самоутверждение ребенка. Планы, которые он постоянно строит и которые ему позволяют осуществить, способствуют развитию у него чувства инициативы. Наоборот, переживание повторных неудач и безответственности могут привести его к покорности и чувству вины.
- Четвертый кризис происходит в школьном возрасте. В школе ребенок учится работать, готовясь к выполнению будущих задач. В зависимости от царящей в школе атмосферы и принятых методов воспитания у ребенка развивается вкус к работе или же, напротив, чувство неполноценности как в плане использования средств и возможностей, так и в плане собственного статуса среди товарищей.
- Пятый кризис переживают подростки обоего пола в поисках идентификации (усвоения образцов поведения значимых для подростка других людей). Этот процесс предполагает объединение прошлого опыта подростка, его потенциальных возможностей и выборов, которые он должен сделать. Неспособность подростка к идентификации или связанные с ней трудности могут привести к ее «распылению» или же к путанице ролей, которые подросток играет или будет играть в аффективной, социальной и профессиональной сферах.
- Шестой кризис свойственен молодым взрослым людям. Он связан с поиском близости с любимым человеком, вместе с которым ему предстоит совершать цикл «работа-рождение детей- отдых», чтобы обеспечить своим детям надлежащее развитие.
 Отсутствие подобного опыта приводит к изоляции человека и его замыканию на самом себе.
Отсутствие подобного опыта приводит к изоляции человека и его замыканию на самом себе. - Седьмой кризис переживается человеком в сорокалетнем возрасте. Он характеризуется развитием чувства сохранения рода (генеративности), выражающегося, главным образом, в «интересе к следующему поколению и его воспитанию». Этот период жизни отличается высокой продуктивностью и созидательностью в самых разных областях. Если, напротив, эволюция супружеской жизни идет иным путем, она может застыть в состоянии псевдоблизости (стагнация), что обрекает супругов на существование лишь для самих себя с риском оскудения межличностных отношений.
- Восьмой кризис переживается во время старения. Он знаменует собой завершение предшествующего жизненного пути, а разрешение зависит от того, как этот путь был пройден. Достижение человеком цельности основывается на подведении им итогов своей прошлой жизни и осознании ее как единого целого, в котором уже ничего нельзя изменить. Если человек не может свести свои прошлые поступки в единое целое, он завершает свою жизнь в страхе перед смертью и в отчаянии от невозможности начать жизнь заново.

(PDF) Теории эволюции и их история в перспективе философии науки
299
невозможно, и нам остаётся довольствоваться косвенными
свидетельствами (например, находками ископаемых). Но при этом
эволюция организмов является не гипотетическим предположением, а, как
констатировал немецкий философ Хуго Динглер, «историческим фактом,
который является достоверным в той же мере, что и любые другие факты
истории, как, скажем, поход Александра, появление Альп, возникновение
христианства и т.п., которые методически могут быть установлены только
на точно таких же принципах» [9, p. 232]. То, что в случае эволюции речь
идёт об исторических фактах, не может оспариваться никаким
образованным и непредвзятым человеком [10, p. 15], и в этом единодушны
все теоретики эволюции и биологи (например, [11, p. 17—77; 12, p. 139; 13,
p. 162]). Но при этом о механизмах и деталях исторического развития
живого имеется множество гипотез и теорий. Реконструкция прошлого
Реконструкция прошлого
часто остаётся временной и спекулятивной из-за неполных и часто
противоречивых данных палеонтологии, сравнительной анатомии,
биогеографии, систематики и генетики [2; 14, 15; 16]. В целом,
эволюционную теорию удобно разделять на две составляющие: эволюция
как исторический факт (сам факт эволюционного развития, филогенез) и
факторы эволюции, т.е. механизмы приводящие в движение
эволюционную машину.
Сочетание различных факторов эволюции (например, естественный
отбор, изотропические мутации, рекомбинация, изоляции, «волны жизни»
и т.д.) может являться предметом обсуждения в науке. То, что
естественный отбор, мутации и т. п. существуют и в состоянии изменить
биологический вид, то есть привести к эволюции, является в настоящее
время установленным фактом. Спорным всегда остаётся то, какое значение
имеют отдельные факторы, как они взаимодействуют и существуют ли ещё
какие-нибудь помимо них. Когда Дарвин опубликовал свою знаменитую
Когда Дарвин опубликовал свою знаменитую
Эпигенетическая теория личности Эрика Эриксона
Истоки теории личности Эриксона
На становление теории личности американского психоаналитика Э. Эриксона большое влияние оказали труды по психоанализу. Ученый принимал структуру личности, разработанную 3. Фрейдом, создав психоаналитическую концепцию отношений «Я» и общества. Особенное внимание он обращал на значение «Я», которое может являться ведущим в развитии личности. Эриксон полагал, что основы человеческого «Я» заложены в социальных особенностях организации общества.
К этому выводу он пришел путем наблюдений за особыми типами личностных изменений, которые происходили с людьми в Америке после войны. Люди становились в то время более тревожными, жесткими, апатичными, смятенными. После принятия идеи неосознанной мотивации, ученый большое внимание в своих исследованиях стал уделять процессу социализации.
Особенности эпигенетической теории Эриксона
Работы Эриксона ознаменовали начало нового метода исследования психики — психоисторического. Он представлял собой использование психоанализа к исследованию развития личности при учете исторического периода, в котором живет человек. Посредством данного метода Эриксон анализирует биографии М. Лютера, М. Ганди, Б. Шоу, Т. Джефферсона и др. Также он подверг исследованию истории жизни современников, включая детей и взрослых. Психоисторический метод потребовал одинакового внимания как к психологии индивида, так и к характеристикам общества, в котором живет человек.
Основную задачу Эриксон ставил в разработку новой психоисторической теории развития личности, которая учитывает конкретную культурную среду.
Ученый провел полевые этнографические исследования, касающиеся воспитания детей в 2 индейских племенах. Он подверг сравнению их с воспитанием детей в городах США, обнаружив, что для каждой культуры характерен особенный стиль материнства. Такой стиль каждая мать воспринимает в качестве единственно правильного, но стиль в любом случае определялся тем, что именно ожидала от ребенка в будущем социальная группа, к которой он принадлежит, включая племя, класс или касту.
Такой стиль каждая мать воспринимает в качестве единственно правильного, но стиль в любом случае определялся тем, что именно ожидала от ребенка в будущем социальная группа, к которой он принадлежит, включая племя, класс или касту.
Для каждого полового этапа развития характерны собственные, присущие именно этому обществу ожидания. Их индивиды могут оправдать или не оправдать, либо включаясь в общество, либо отвергаясь им. Подобные соображения Э. Эриксона становятся основной двух важнейших понятий его концепции, которая представляла собой групповую идентичность и эго-идентичность.
Групповая идентичность в основании имеет положение, что мама с первого дня жизни ориентирует воспитание ребенка на включение его в определенную социальную группу, включая формирование характерного для этой группы мироощущения.
Формирование эго-идентичности происходит параллельно с групповой, создавая у субъекта чувство устойчивости и непрерывности собственного «Я», вне зависимости от изменений, происходящих с людьми в процессе его роста и развития.
Стадии жизненного пути личности
Э. Эриксон определил стадии жизненного пути личности. Для каждой из них характерна специфическая задача, выдвигаемая обществом, которое также формирует содержание развития на различных этапах жизненного цикла. Тем не менее, решение задач, в соответствии с воззрениями Эриксона, зависит и от уже достигнутого уровня психомоторного развития человека, включая общую духовную атмосферу общества, в котором он живет.
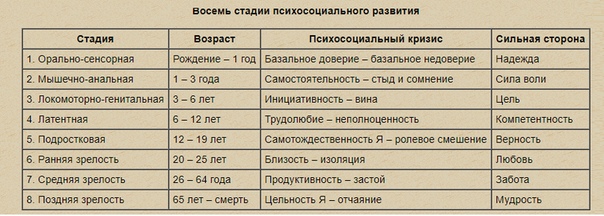
В табл. 1 рассмотрены этапы жизненного пути личности, разработанные Э. Эриксоном.
Нужна помощь преподавателя?
Опиши задание — и наши эксперты тебе помогут!
Описать заданиеТаблица 1. Этапы жизненного пути личности Э. Эриксона
|
Старость |
Интеграция |
|
|
разочарование в жизни |
|
Зрелость |
Творчество |
|
|
застой |
|
Молодость |
Интимность |
|
|
изоляция |
|
Подростковый возраст |
Идентичность |
|
(латентный этап) |
диффузия |
|
|
идентичности |
|
Школьники |
Достижение |
|
(латентный этап) |
неполноценность |
|
Игровой возраст |
Инициативность |
|
(фаллический этап) |
вина |
|
Ранний возраст |
Автономия |
|
(анальный этап) |
стыд, сомнение |
|
Младенчество |
Доверие |
|
(оральный этап) |
отсутствие доверия |
Что касается кризиса развития, то для него характерны все форм идентичности. Э. Эриксон считал, что основной кризис идентичности происходит в юношеском возрасте. Если процессы развития происходят удачно, то обретается «взрослая идентичность», а при возникновении трудностей развития отмечается задержка формирования идентичности.
Э. Эриксон считал, что основной кризис идентичности происходит в юношеском возрасте. Если процессы развития происходят удачно, то обретается «взрослая идентичность», а при возникновении трудностей развития отмечается задержка формирования идентичности.
Эриксон назвал интервал между юностью и взрослым состоянием «психосоциальным мораторием». Его можно считать временем, когда молодые люди через пробы и ошибки стремятся определить собственное место в жизни. Бурность протекания этого кризиса зависит от того, насколько удачно разрешились предыдущие кризисы (доверие, независимость, активность и др.) и от духовной атмосферы в обществе. В случае, если кризис на ранних фазах не был успешно преодолен, отмечается задержка формирования идентичности.
Э. Эриксоном было введено понятие ритуализации, которая в поведении означает построенное на соглашении взаимодействие нескольких людей. Оно может возобновляться через определенные промежутки времени в повторяющихся обстоятельствах (ритуалы взаимного узнавания, приветствие, критика и др. ). Ритуал при однократном возникновении способен последовательно включаться в систему, которая проявляется на более высоких уровнях и становится частью последующих стадий.
). Ритуал при однократном возникновении способен последовательно включаться в систему, которая проявляется на более высоких уровнях и становится частью последующих стадий.
Эпигенетическая теория Эриксона называется эпигенетической концепцией жизненного пути личности. В соответствии с ней для всего, что растет, характерен общий план. Исходя из него происходит развитие отдельных элементов, при этом каждый из них имеет максимально благоприятный период для развития.
Это происходит до тех пор, пока все элементы в своем развитии не создадут функциональное целое.
Эриксон полагал, что последовательность этапов является результатом биологического созревания, при этом содержание развития можно определить тем, что ожидает общество от индивида. Ученый признавал, что его периодизация не должна рассматриваться в качестве теории личности. Ее можно считать лишь ключом к построению подобной теории.
Эпигенетическая теория эволюции: ivanov_p — LiveJournal
- ivanov_p (ivanov_p) wrote,
ivanov_p
ivanov_p
 Это была не одна теория, их рядом развивалось несколько вариантов. Это была замечательная альтернатива мейнстриму. И поэтому, конечно, было глухо: усилия одиночек вязли в глухоте, как это всегда и бывает. (История эта заканчивается обычно без всякого оптимизма: забвением). С тех восьмидесятых многое изменилось, и окружающий фон соперничающих концепций стал совсем иной. Вокруг, конечно, молбиология, кладизм, плюрализм и чрезвычайное снижение интереса ко всяким теоретическим компонентам, и к теории адаптаций, и к системным аспектам теории эволюции. Это статья М.Шишкина, создателя эпигенетической теории эволюции — 2018 года. Собственно, можно видеть, как это излагается сейчас на английском, а не в 80-х на русском, и можно наблюдать, что сорок прошедших лет ничего не изменили: автор не нуждается в прошедшем развитии, цитаты те же, что можно было дать и в 82-м году. Интересно также и то, что тут воспроизводится теория, уровень которой много выше современного. Но, кажется, она не только выше, она уже — среди того, что прошло.
Это была не одна теория, их рядом развивалось несколько вариантов. Это была замечательная альтернатива мейнстриму. И поэтому, конечно, было глухо: усилия одиночек вязли в глухоте, как это всегда и бывает. (История эта заканчивается обычно без всякого оптимизма: забвением). С тех восьмидесятых многое изменилось, и окружающий фон соперничающих концепций стал совсем иной. Вокруг, конечно, молбиология, кладизм, плюрализм и чрезвычайное снижение интереса ко всяким теоретическим компонентам, и к теории адаптаций, и к системным аспектам теории эволюции. Это статья М.Шишкина, создателя эпигенетической теории эволюции — 2018 года. Собственно, можно видеть, как это излагается сейчас на английском, а не в 80-х на русском, и можно наблюдать, что сорок прошедших лет ничего не изменили: автор не нуждается в прошедшем развитии, цитаты те же, что можно было дать и в 82-м году. Интересно также и то, что тут воспроизводится теория, уровень которой много выше современного. Но, кажется, она не только выше, она уже — среди того, что прошло. Автора цитируют в основном российские исследователи, внимания его публикации не привлекают. Это ответы, которые не нужны — прошло время, когда еще были живы вопросы, на которые тут пытаются ответить.
Автора цитируют в основном российские исследователи, внимания его публикации не привлекают. Это ответы, которые не нужны — прошло время, когда еще были живы вопросы, на которые тут пытаются ответить.-
Новое Средневековье и Новый Рим
Не первой молодости книга о «впадении в средневековье», на примере США. Примерно то же сказано уже стотыщ раз. Критика современного общества понятна.…
-
Классификация энтомологов-программистов
«Программисты часто сталкиваются с багами. «Баги» это — жуки. Они же — насекомые. Ученые, занимающиеся насекомыми — энтомологи. То есть программисты…
-
Анатомия человеческих сообществ — книга о сознании
В книге нет ни интересных фактов, ни доказательных рассуждений — нет ничего. Весь объем — произвольные допущения и невнятные рассуждения в пользу…
Photo
Hint http://pics.livejournal.com/igrick/pic/000r1edq
Теории старения.
 Часть вторая
Подробный материал опубликован в четвертом номере журнала Les Nouvelles Esthétiques за 2017 год
Часть вторая
Подробный материал опубликован в четвертом номере журнала Les Nouvelles Esthétiques за 2017 год Мутационная теория старения и теория нестабильности генома
Постулат данных теория — накопление мутационных изменений в геноме приводит к старению организма на всех уровнях организации. Одним из первых автор этой теории был американский физик Лео Силард. Будучи ядерным физиком, в 159 году он предположил, что ионизирующее излучение приводит к ускорению старения. Позднее, в 1961 году, Линдоп и Ротблат показали, что радиация сокращает жизнь крыс и мышей. И только позднее было показано, что в данном случае основной причиной сокращения жизни является рост канцерогенеза, но не физиологическое ускорение старения.
Мутация ДНК противостоят механизмы распознавания повреждений/репарации ДНК. В настоящее время механизмы репарации ДНК хорошо изучены, в том числе и на примере кожных фибробластов. Харт и Сетлоу в 1974 году показали, что активность механизмов репарации ДНК в кожных фибробластах, облученных УФ-радиацией, зависит от максимальной продолжительности жизни данного вида животных. Пропорциональной зависимости между продолжительностью жизни и активностью механизмов репарации ДНК не установлено, но ясно, что эти механизмы играют существенную роль в увеличении продолжительности жизни.
Пропорциональной зависимости между продолжительностью жизни и активностью механизмов репарации ДНК не установлено, но ясно, что эти механизмы играют существенную роль в увеличении продолжительности жизни.
Специально разработанные методики in vivo изучения интенсивности мутаций показали рост таковых по мере старения организма. Тем не менее на тех же мышах показано, что для одних клеток/тканей рост количества мутаций с возрастом достоверный, а для других — нет. Сегодня ясно, что по мере взросления рост мутаций увеличивается, но остается неизменным в старости. И общий уровень накопленных мутаций даже в старческом возрасте незначителен для того, чтобы оказывать влияние на продолжительность жизни.
Основываясь на выраженности функциональных возрастных нарушений, свойственных ряду органов (например, мозгу и сердцу), и на несоответствии незначительности накопленных мутационных изменений в ДНК клеток этих органов, в настоящий момент исследователи не рассматривают данные теории как основополагающие.
Теория репликативного старения и теломерная теория
В 1961 году профессор анатомии Калифорнийского университета Леонард Хейфлик при изучении деления человеческих фибробластов в монослое обнаружил предел данного деления — около 50 раз. В 1971 году советский молекулярный биолог Алексей Оловников для объяснения экспериментальных данных Хейфлика предположил, что причина лимитированного количества делений клетки заключается в укорочении хромосомы при каждом делении. А уже в 1985 году была открыта обратная транскриптаза, которой является теломераза (спустя 24 года коллектив американских ученых получил Нобелевскую премию за открытие механизма старения, предсказанного Оловниковым).
Разнообразные исследования Хейфлика показали, что клетки запоминают количество предшествующих делений. При увеличении возраста пробанда на 1 год количество делений в культуре фибробластов снижается на 0,2. Это значит, что репликативный потенциал фибробластов человека за сто лет снижается всего на 20 делений, а весь этот потенциал достаточен для жизни в 250 лет.
Важно, что в эксперименте Хейфлика in vitro клетки отделяли друг от друга при помощи трипсина и инкубировали в присутствии сыворотки. Как оказалось позднее, это не так уж и безобидно: нарушается стабильность плазмолеммы и ядерной мембраны, что приводит к деградации мембранных белков, полисом и негистоновых ядерных белков. Скорее всего, не внутренние часы, а именно 50-кратная обработка типсином вызывала старение и гибель культуры фибробластов. Таким образом, в 1978-1979 гг. исследователи из нескольких стран провели ревизию теории Хейфлика. Было обнаружено, что ряд клеток подвергается делению около 170 раз, а культура сохраняет стабильность гораздо более длительное время (рис. 1).
РИС. 1 Результат исследования Хейфлика; ревизия результатов его исследования
Исследования теломер и толемераз позволили раскрыть многие особенности жизнедеятельности клетки, но не добавили яности в вопросе старения.
Теломера — это концевая несимметричная часть молекулы ДНК, состоящая их гексануклеотидных (TTAGGG) повторов. Теломера необходима для корректного разведения обеих целей ДНК, инициирования репликации, профилактики связывания двух цепей друг с другом и потери генетического укорачивается на 50-100 нуклеотидных пар, что за 50 делений составит 2-3 тыс. нуклеотидных пар, и это около 20% всей длины теломеры. Критическое укорочение теломеры и/или ее повреждение являются причинами запрета деления клетки (рис. 2).
Теломера необходима для корректного разведения обеих целей ДНК, инициирования репликации, профилактики связывания двух цепей друг с другом и потери генетического укорачивается на 50-100 нуклеотидных пар, что за 50 делений составит 2-3 тыс. нуклеотидных пар, и это около 20% всей длины теломеры. Критическое укорочение теломеры и/или ее повреждение являются причинами запрета деления клетки (рис. 2).
В исследованиях показано, что фибробласты 90-летних доноров делились всего на 20 раз меньше, чем фибробласты, полученные от плода человек а. То есть старыми фибробласты стали не потому, что много делились. Математический подсчет количества делений гемопоэтических клеток указывает, что оно может составлять 400-450 раз у 60-летнего пробанда. Таким образом, неизвестно, существует ли предел делений клеток in vivo.
РИС. 2 Укорочение и повреждение теломеры как причины запрета деления клетки
Попытки обосновать существование генетически запрограммированного предела продолжительности жизни ссылками на предел клеточных делений нельзя признать убедительными. Это тем более важно на фоне того, что выполнение высокоспециализированной функции оказывается несовместимым с клеточным делением (например, нервные и мышечные клетки), а снижение митотической активности является нормальным физиологическим процессом, связанным с дифференцировкой клеток.
Это тем более важно на фоне того, что выполнение высокоспециализированной функции оказывается несовместимым с клеточным делением (например, нервные и мышечные клетки), а снижение митотической активности является нормальным физиологическим процессом, связанным с дифференцировкой клеток.
Вместе с тем очевидно, что переход части клеток в постмитотическое состояние создает предпосылки к снижению регенераторных возможностей организма и в конечном счете — к уменьшению числа функционирующих клеток, что действительно наблюдается с возрастом. Поэтому исследование кинетики клеточных популяций на разных этапах жизни организма может иметь большое значение для выяснения механизмов, определяющих продолжительность жизни.
Теория дисдифференцировки и эпигенетическая теория
Ричард Катлер в 1991 году предположил, что зрелая (дифференцированная) клетка в процессе своей жизни (старения) может подвергаться изменениям в реакциях транскрипции. Иными словами, происходит ее дисдифференцировка — строгий паттерн экспрессируемых генов со временем «размывается» (ослабевает), приводя к изнашиванию морфологии/функции клеток и тканей.

Эта теория основана в том числе и на том, что экспрессия генов в дифференцированных клетках (например, мозга) более гетерогенная и широкая у особой с более выраженным старением.
Схожего мнения придерживался Владимир Фролькис; его адаптационно-регуляторная теория старения гласит, что при нарушении активности работы регуляторных генов страдает функция структурных генов. Это может привести к выключению традиционных генов и включению ранее не работавших генов (и появлению в клетке не свойственных ей белков).
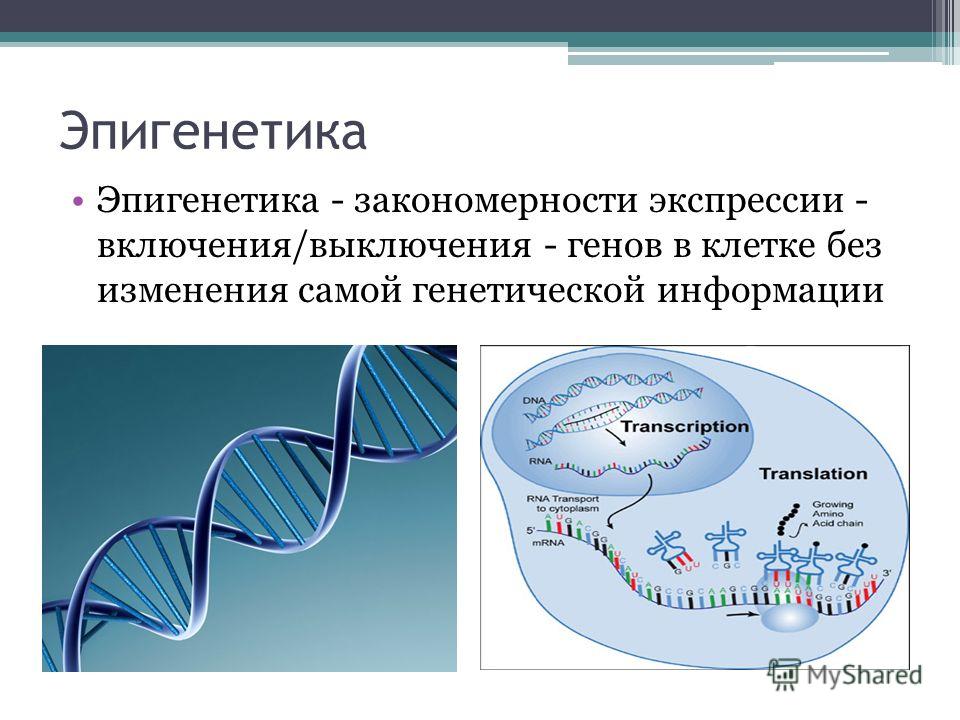
Вышеуказанные теории получили второе дыхание в связи с развитием эпигенетической теории старения. Эпигенетика изучает процессы наследования, не затрагивающие последовательности нуклеотидов в ДНК, а также то, каким образом из поколения в поколение экспрессия одного и того же паттерна генов наследуется без изменений. Или каким образом обеспечивается продолжительный контроль за единообразием экспрессии определенного паттерна генов в долгоживущей дифференцированной клетке, несмотря на воздействующие на нее факторы. Или каким образом в митотических клетках наследуются изменениям в функции генов, которые не могут быть объяснены изменениями в последовательности ДНК.
Эпигенетические механизмы контроля за стабильностью экспрессии генов включают:
- метилирование ДНК;
- ацетилирование гистонных белков;
- экспрессию микроРНК.
Эти механизмы стимулируются как внешними, так и внутренними факторами (энергетический метаболизм, питание, физические нагрузки) и влияют на процессы старения и развитие возраст-ассоциированных заболеваний.
В исследованиях было обнаружено, что для более старых клеток характерны иные «узоры» метилирования ДНК и ацетилирования гистонных белков по сравнению с молодыми. Например, эпигенетические различия у однояйцевых близнецов в старшем возрасте были значительно более выраженными, чем в молодом. Это подтверждает тот факт, что один и тот же геном в процессе жизни может подвергаться различным эпигенетическим модификациям с исходом в разную траекторию фенотипа (например, более или менее старый).
Старческая немощь (потеря веса, мышечная слабость, саркопения) ассоциирована с глобальным снижением метилирования ДНК. Старение в принципе ассоциировано со снижением метилирования ДНК в определенных локусах. Отмеченное с возрастом снижением ацетилирования гистонных белков приводит к более плотной упаковке хроматина, что затрудняет экспрессию генов.
Эпигенетическая теория старения выглядит многообещающей, но еще предстоит провести широкий спектр биологических исследований.
Эпигенетическая Теория Эволюции Как Концептуальная Модель Развития Системы Управления Образованием
Author
Abstract
Развитие системы управления образованием все более усложняется за счет повышения сложности самой системы образования, а также появления новых вызовов и угроз для ее развития. Данные обстоятельства вызвали к жизни возникновение целого спектра новых моделей развития системы управления образованием. Одной из таких моделей является эпигенетическая теория эволюции. Автор приводит аргументы в пользу возможности этой теории как концептуальной модели развития системы управления образованием.
Suggested Citation
Download full text from publisher
Corrections
All material on this site has been provided by the respective publishers and authors. You can help correct errors and omissions. When requesting a correction, please mention this item’s handle: RePEc:scn:017885:15898845. See general information about how to correct material in RePEc.
For technical questions regarding this item, or to correct its authors, title, abstract, bibliographic or download information, contact: (CyberLeninka). General contact details of provider: http://cyberleninka.ru/ .
If you have authored this item and are not yet registered with RePEc, we encourage you to do it here. This allows to link your profile to this item. It also allows you to accept potential citations to this item that we are uncertain about.
We have no references for this item. You can help adding them by using this form .
If you know of missing items citing this one, you can help us creating those links by adding the relevant references in the same way as above, for each refering item. If you are a registered author of this item, you may also want to check the «citations» tab in your RePEc Author Service profile, as there may be some citations waiting for confirmation.
Please note that corrections may take a couple of weeks to filter through the various RePEc services.
Эволюционные последствия эпигенетического наследования
Изучение эпигенетического наследования перешло от документирования его существования к изучению его эволюционных последствий. Из множества открытых областей исследований есть три ключевых вопроса, на которые мы особенно хотели бы обратить внимание.
Является ли эпигенетическое наследование обычно адаптивным?
Модели предполагают, что эпигенетическое наследование родительского фенотипа может быть адаптивным в медленно меняющихся и коррелированных средах, поскольку родитель и потомство чаще всего находятся в одних и тех же условиях окружающей среды (Jablonka and Raz 2009; Uller et al.2015). Однако прямых проверок этого прогноза пока нет. С другой стороны, если родители и потомство живут в отрицательно коррелированной среде, родительский фенотип не должен передаваться по наследству, но родители все еще могут предвидеть среду, в которой находится потомство. Таким образом, развитие негенетического родительского эффекта может быть адаптивным, и это предсказание недавно получило экспериментальную поддержку (Dey et al., 2016). Это соответствует широкой концепции родительских эффектов (Uller et al. 2015) и предполагает, что опережающие эффекты (такие как эпигенетическое наследование) могут развиваться, если окружающая среда предсказуема для разных поколений.Кроме того, ожидается, что адаптивная обусловленная окружающей средой эпигенетическая наследственность будет более распространена у организмов с коротким жизненным циклом, чем, например, у долгоживущих млекопитающих (Houri-Zeevi and Rechavi 2017). Чтобы исследовать эти вопросы и получить более глубокое понимание эволюционной динамики эпигенетического наследования, важно сосредоточиться на разнообразном наборе систем исследования с известной степенью экологической изменчивости.
Влияет ли эпигенетическая наследственность на генетическую адаптацию?
Эпигенетическое наследование может способствовать адаптации двумя способами.Во-первых, адаптивное эпигенетическое наследование, вызванное окружающей средой, может приблизить популяцию к пику приспособленности, чем если бы они полагались только на генетические изменения (Kronholm and Collins 2016). Если новая среда стабильна, в конечном итоге могут последовать генетические изменения в соответствии с теми же принципами, что и генетическая фиксация первоначально индуцированных фенотипов (т. Е. Генетическая ассимиляция). Во-вторых, из-за высокой скорости эпимутаций (Graaf et al. 2015) отбор может воздействовать на случайно индуцированные эпигенетические варианты, которые, если они унаследованы, могут способствовать адаптации в аналогичном сценарии, описанном выше.Однако разделить генетические и эпигенетические вариации сложно, но работа с клональными организмами, которые вторгаются в новые среды, может быть подходящей отправной точкой.
Насколько важна отцовская эпигенетическая наследственность?
Хотя мы все больше осознаем, что отцы могут иметь большое влияние на наследование между поколениями, мы далеки от понимания того, столь же важны такие отцовские эффекты, как и материнские. Кроме того, мы должны также изучить и определить важность эякулята в целом в отцовской передаче между поколениями.Помимо спермы, самцы также передают факторы самкам через семенную жидкость, и такие факторы могут иметь долгосрочные последствия как для самок, так и для потомства (Crean et al. 2014; Bromfield et al. 2014). Отцовские трансгенерационные эффекты опосредуются женскими реакциями, которые также открывают потенциальный сексуальный конфликт из-за экспрессии генов в потомстве (Crean and Bonduriansky 2014). Эксперименты, направленные на выяснение этих эффектов, позволят нам исследовать относительную важность и механизмы отцовского трансгенерационного эпигенетического наследования.
Заключительные замечания
Как видно из этого набора статей, эпигенетика и эпигенетическая наследственность могут иметь огромное влияние на эволюцию. Эпигенетическая наследственность между поколениями не только должна приниматься во внимание при оценке количественных генетических параметров, но также может реагировать на отбор и влиять на адаптацию к новым условиям. Таким образом, возникающая область эпигенетического наследования имеет много общего с хорошо развитой в настоящее время областью фенотипической пластичности, которая перестала считаться неприятной и стала основной областью исследований, имеющей большое значение для адаптации.Пластичность и эпигенетическое наследование могут иметь общие механизмы, которые регулируют экспрессию генов. В обоих случаях степень неоднородности окружающей среды и ее стабильность на протяжении поколений становятся определяющим фактором, влияющим на их эволюцию. Материалы этого специального выпуска дают важную картину состояния поля адаптивного эпигенетического наследования, подчеркивают его эволюционные последствия и указывают на важные направления дальнейшего развития.
Идея этого специального выпуска была частично основана на симпозиуме под названием «Эволюционные последствия мобильных элементов, эпигенетика и негенетическая наследственность», состоявшемся на конференции Европейского общества эволюционной биологии в Гронингене, Нидерланды, в августе 2017 г. .
Социальная конструкция социального эпигенома и более широкого биологического контекста | Эпигенетика и хроматин
Невозможно отделить эпигенетику от генетики
В мае 2016 года в New Yorker появилась статья исследователя рака и писателя Сиддхартха Мукерджи о том, как факторы окружающей среды могут изменять активность генов без изменения последовательности ДНК. [37], вызвали сильную критическую реакцию со стороны генетиков, эпигенетиков и других биологов [38].Они критиковали Мукерджи, подчеркивая модификацию гистонов и метилирование ДНК, игнорируя основную роль факторов транскрипции и РНК в процессе транскрипции. Марк Пташне и Джон Грилли указали на важность специфичности: «Развитие требует высокоспецифичного последовательного включения и выключения наборов генов. Факторы транскрипции и РНК обеспечивают эту специфичность, но ферменты, которые модифицируют гистоны, не могут: каждая нуклеосома (и, следовательно, каждый ген) кажется ферменту одинаковым.«Не только специфичность клеточной идентичности, но и реакция на стресс, как известно на протяжении десятилетий, обусловлено действием специфических ДНК-связывающих белков (и, реже, молекул РНК), которые регулируют транскрипцию генов» [39]. В исключительном и весьма похвальном ответе Мукерджи поблагодарил своих критиков за их «чрезвычайно подробные комментарии», признав, что совершил ошибку, «упустив из виду ключевые области науки» [38].
Исключение механизмов, основанных на последовательности ДНК как основной причине изменений экспрессии генов, также является серьезным недостатком в аргументах социальных эпигенетиков.Подчеркивая предполагаемую роль метилирования ДНК и ацетилирования гистонов в контроле регуляции генов, они не упоминают факторы транскрипции. Как прокомментировали Стивен Хеникофф и Джон Грили, факторы транскрипции «на самом деле обладают многими необходимыми свойствами регулятора клеточной памяти или медиатора влияний окружающей среды» [4].
Модификаторы гистонов или ферменты, которые переносят метильные группы на ДНК (метилтрансферазы), не имеют специфических ДНК-связывающих доменов и не направлены конкретно на определенные гены.Таким образом, факторы транскрипции необходимы для нацеливания событий регуляции транскрипции на специфические последовательности ДНК, иногда связываясь с длинными некодирующими РНК в этих событиях [40]. Они также опосредуют влияние окружающей среды на активность генов и поддерживают клеточную память в зависимости от последовательности. Кроме того, факторы транскрипции участвуют в начальных стадиях инактивации Х-хромосомы и импринтинга, который затем поддерживается метилированием ДНК [12]. Адриан Бёрд считает, что это тесное взаимодействие между эпигенетическими метками и генетикой растворяет самобытность эпигенетики.«И я считаю, что это хорошо» [41].
Функция метилирования ДНК спорна.
Роль метилирования ДНК и модификации гистонов в биохимических событиях, которые регулируют гены, все еще четко не установлена и является спорной. Существуют группы организмов, такие как нематоды и некоторые насекомые, такие как дрозофила, которые не метилируют свои геномы. Бесспорно, что метилирование ДНК не подавляет активные промоторы генов, но влияет на гены, которые уже молчат [42, 43].Также общепризнано, что одной из основных функций метилирования цитозина ДНК является его решающее участие в таких процессах, как молчание транспозонов, импринтинг и инактивация Х-хромосомы [44,45,46,47]. Здесь также факторы, специфичные для последовательности ДНК, такие как факторы транскрипции или РНК, нацелены на метилтрансферазы в соответствующие части генома.
Некоторые авторы предполагают, что метилирование цитозином повторяющихся последовательностей ДНК и транспозонов представляет собой систему защиты генома [44, 46].Тимоти Бестор и его соавторы пояснили, что, несмотря на множество корреляций между активацией транскрипции и деметилированием, причинно-следственная связь не была продемонстрирована, а имеющиеся данные не подтверждают «существование биохимической системы, которая регулирует эмбриогенез с помощью запрограммированного метилирования и деметилирования регуляторных органов». последовательности. » Они также считают, что «на сегодняшний день нет разумных доказательств существования сложной биохимической системы, которая активирует и репрессирует гены посредством обратимого метилирования ДНК» [46].Авторы критикуют отсутствие надежных критериев в исследованиях, утверждающих, что гены регулируются «динамическим запрограммированным метилированием и деметилированием ДНК во время развития».
Bestor et al. предполагают, что «паттерны метилирования генома млекопитающих представляют собой эволюционную адаптацию системы защиты генома, которая наделяет геномы способностью самовоспроизводиться инактивировать определенные области генома, что по существу необратимо на протяжении жизни организма». Они согласны с Ptashne и Greally [39], что активация и репрессия генов во время развития контролируются хорошо установленными и законсервированными механизмами, основанными на белках и РНК.Таким образом, метилирование ДНК, которое наиболее сильно подчеркивается социальными эпигенетиками как механизм регуляции генов, по-видимому, не играет роли в включении и выключении генов.
Окружающая среда не оказывает длительного воздействия на изменение эпигенетических меток
По словам Адриана Берда, достоверных данных о влиянии окружающей среды на «эпигеном» человека нет [41]. Ответ на сигналы окружающей среды обычно опосредуется специфическими белками, такими как факторы транскрипции или, по терминологии Марка Пташне, рекрутерами [48].
Паттерны метилирования ДНК, однажды установленные в клетке факторами транскрипции, могут реплицироваться и передаваться в дочерние клетки ферментом DNMT1 независимо от факторов транскрипции; то же самое может быть верно и для некоторых гистоновых меток. Это открывает возможность того, что метилом может напрямую зависеть от окружающей среды, например, из-за острой нехватки ферментных кофакторов, таких как доноры метила, необходимых для метилтрансфераз, или из-за присутствия ферментных ингибиторов, таких как 2-гидроксиглутарат, который ингибирует деметилазы.Социальные эпигенетики могли использовать этот факт для своих рассуждений, но они проигнорировали все другие факты и контексты, в частности то, что почти во всех случаях окружающая среда действует на фенотип посредством регуляции транскрипции и клеточной дифференциации. Большая часть стабильности и клеточной памяти основана на сетях регуляции генов, включающих петли обратной связи [я благодарен анонимному рецензенту за эту информацию].
Согласно Эдит Херд и Роберту Мартиенсену, эпигенетическая изменчивость может реагировать на окружающую среду, но это не означает, что она каким-либо образом влияет на адаптивную приспособленность.Таким образом, у Drosophila вызванная тепловым шоком или осмотическим стрессом репрессия гена white может передаваться по материнской и отцовской линии в течение нескольких поколений, но затем возвращается в нормальное состояние. У мышей Agouti матери могут изменять цвет шерсти своего потомства с помощью специфической диеты, состоящей из доноров метильных групп, но этот эффект теряется к третьему поколению, указывая на то, что влияние диеты не является стабильным или действительно трансгенерационным [49].
Организмы реагируют на окружающую среду посредством взаимодействия многих факторов, в первую очередь специфических ДНК-связывающих белков.На дрожжах было показано, что стресс окружающей среды, такой как тепло, окисление, кислотность или голод, влияет на различные гены по-разному, то есть ответ зависит от последовательности ДНК, с факторами транскрипции и комплексом, модифицирующим мультипротеиновый хроматин, активируя чувствительные к стрессу гены. в ответ на стрессоры [50]. Этот мультибелковый комплекс эволюционно консервативен; в дрожжах он ацетилирует и деубиквитинирует гистоны [51].
Некоторые исследования указывают на изменение решений клеточной судьбы в ответ на дефицит питательных микроэлементов или на эндокринные разрушители у мышей через факторы транскрипции.Химические вещества, нарушающие работу эндокринной системы, изменяют функцию нормальной эндокринной системы и представляют собой одну из основных областей эпигенетических исследований [12]. В хорошо изученном случае мыши, подвергшиеся внутриутробному воздействию определенных химических веществ (из семейства оловоорганических, члены которого используются в качестве пестицидов), накапливали жир от рождения до взрослого возраста. Эти фенотипические эффекты, по-видимому, опосредуются рецепторами, которые заставляют мезенхимальные стволовые клетки дифференцироваться преимущественно в клон адипоцитов (жировых клеток).Это означает, что они не требуют перепрограммирования определенного типа клеток [12].
Проверяя гипотезу о том, что виктимизация молодых людей в детстве и подростковом возрасте связана с метилированием ДНК, Marzi et al. показали, что такие анализы страдали серьезными методологическими недостатками (они были затруднены курением табака и / или не выдержали контрольных тестов со-близнецов) [52]. Анализ шести генов-кандидатов в ответ на стресс (NR3C1, FKBP5, BDNF, AVP, CRHR1, SLC6A4) не выявил предполагаемых ассоциаций с метилированием ДНК.Сделав вывод о том, что их эпидемиологический анализ эпигенетических эффектов стресса в раннем возрасте не подтвердил гипотезу о сильных изменениях метилирования ДНК у пострадавших молодых людей, авторы рекомендовали, чтобы «нам нужно было смириться с возможностью того, что эпигенетическая эпидемиология еще не улучшилась. сопоставлены с экспериментальными, нечеловеческими моделями в раскрытии биологической оболочки стресса »[52]. Если вы прислушаетесь к этому совету, это значительно уменьшит заблуждение относительно эпигенетических меток.
Недавнее исследование показало, что, действительно, изменения в поведении курения были связаны с изменениями в метилировании ДНК, которые зависели от стимула в геноме человека, но не зависели от генетических и экологических факторов риска, поскольку данные о близнецах, противоречащие курению, не учитывались. совпадение [53].Основываясь на этих выводах и указывая на методические проблемы социальных эпигеномных исследований в целом, такие как их низкая статистическая мощность, некоторые социологи рекомендуют вместо этого полагаться на геномную методологию: «С появлением и растущей надежностью геномных методологий социологи оказались в завидном положении. принять эти инструменты и интегрировать их в свои исследования »[54].
Нет доказательств трансгенерационной эпигенетической наследственности у людей
Эпигенетическое наследование у растений и нематод
Большинство ученых, рассмотренных в этой статье, согласны с существованием трансгенерационного наследования приобретенных признаков через РНК у нематод и через метилирование у растений.Но доказательства того, что наследование между поколениями имеет эпигенетическую основу у млекопитающих, редки [49].
Трансгенерационное эпигенетическое наследование неясной функции является обычным явлением у растений. На сегодняшний день нет доказательств того, что наследуемые признаки адаптивны. Эпигенетическое наследование у растений обычно связано с мобильными элементами, вирусами или трансгенами и может быть, как предполагалось для млекопитающих, побочным продуктом стратегий защиты зародышевой линии [49]. В последние годы новое политическое движение, которое сопровождается растущей симпатией к Сталину, обратилось к эпигенетике, чтобы реабилитировать ошибочные эксперименты по яровизации, проведенные агрономом Трофимом Лысенко, ставленником Сталина [55].Яровизация, влияние температуры и сезона на время цветения растений, была открыта немецким ботаником Густавом Гасснером в 1918 году и затем широко применена Лысенко, который утверждал, что эффекты яровизации передаются по наследству [56]. Отсутствие научной строгости в его работе анализировалось в другом месте, как и разрушительные политические и экономические последствия практики Лысенко (см., Например, [55, 57].
Было показано, что яровизация, которая происходит после продолжительных периодов холода). , приводит к эпигенетическому подавлению репрессора цветков в сложном процессе, который включает два белковых комплекса и метилирование.Но в отличие от заявлений нового движения сторонников Лысенко, память о яровизации не сохраняется в следующем поколении, потому что она надежно сбрасывается в зародышевой линии и раннем эмбрионе [49].
Трансгенерационное эпигенетическое наследование было наиболее надежно продемонстрировано многими исследователями на нематоде C. elegans , где малые РНК могут проникать в зародышевую линию и опосредовать наследственное молчание транскрипции в последующих поколениях (нематоды не метилируют свои геномы).Примером может служить наследование между поколениями на протяжении многих поколений малых интерферирующих РНК, нацеленных на гены, имеющие отношение к хемотаксису, питанию или подавлению вирусного генома червя [58,59,60]. В этих исследованиях Одед Рехави и его соавторы предполагают — но пока не могут показать, — что обнаруженные ими механизмы могут обеспечить адаптивные преимущества для червя. Механизмы основаны на генах и, следовательно, подвержены естественному отбору. Эти гены, которые «необходимы для этого многопоколенческого эффекта» передачи РНК, нацелены на другие гены, играющие роль в питании [59].Малые РНК транскрибируются и, в отличие от метильных групп, содержат генетическую информацию. По этой причине, а также из-за отсутствия доказанной до настоящего времени адаптивности утверждение Рехави и др. О том, что «наши результаты, таким образом, подтверждают ламарковскую концепцию наследования приобретенного признака» [60], неуместно. Наследование приобретенных черт от поколения к поколению не обязательно должно быть ламаркистским, т. Е. Иметь адаптивное и эволюционное значение. Результаты по нематодам нелегко применить к людям.У нематод очень короткое время генерации и, в отличие от высших животных, они обладают РНК-зависимыми РНК-полимеразами, которые могут копировать небольшие молекулы РНК для многих поколений. Кроме того, в отличие от C. elegans , большинство предполагаемых трансгенеративных унаследованных у людей признаков, таких как последствия голодания, являются пагубными.
Отсутствие доказательств трансгенерационного эпигенетического наследования у людей и его редкость у других млекопитающих
Многие из возможных примеров эпигенетического наследования, которые были предложены для людей, касаются скорее межпоколенческих, чем трансгенерационных эффектов и редко исключают изменения последовательности ДНК как основную причину наследственности [49, 61].Родительские или межпоколенческие эффекты возникают, когда матка подвергается воздействию токсинов, вирусов (например, краснухи), вредной пищевой или гормональной среды, которая напрямую влияет на развивающийся эмбрион и его зародышевую линию. Это воздействие обычно влияет на первое поколение, но иногда и на внуков. Напротив, трансгенерационные эффекты относятся к поколениям, которые не подвергались воздействию исходной среды, то есть к правнукам и не только.
Межпоколенческие эффекты наблюдаются у людей и других млекопитающих, но есть два раунда эффективного перепрограммирования и стирания метилирования ДНК в развитии тотипотентных клеток в раннем эмбрионе, а также во время дифференцировки половых клеток.Широко распространено мнение, что это перепрограммирование предотвращает наследование большинства эпигенетических меток, хотя некоторые локусы генов избегают его. Некоторые исследователи приписывают ему эволюционное значение: «Похоже, что эволюция пошла на многое, чтобы гарантировать эффективное удаление любых потенциально вредных закладок, которые могли быть наложены жизненным опытом родителей», и они приходят к выводу, что «хотя на это было обращено много внимания потенциальные последствия наследования между поколениями для здоровья человека пока мало поддерживаются »[49].
Совсем недавно John Edwards et al. продемонстрировали, что динамика деметилирования и реметилирования во время раннего развития более сложна, чем предполагалось ранее [47]. Они показали, что только последовательности, которые, по-видимому, имеют мало доказательств биологической функции, такие как старые и неактивные остатки транспозонов, сателлитная и другая повторяющаяся ДНК, подвергаются двойной волне деметилирования и реметилирования. Напротив, другие последовательности, такие как подавляющее большинство промоторов CpG-островков, не подвержены этим волнам метилирования и деметилирования, потому что они неметилированы на всех стадиях.Специфичное для пола метилирование в импринтирующих контрольных областях деметилируется только в первом раунде; тогда как небольшая популяция молодых, богатых CpG транспозонов в значительной степени избегает обоих раундов деметилирования.
Авт. Показали, более того, что паттерны геномного метилирования регуляторных последовательностей по существу статичны во время развития и что деметилирование промоторов при активации транскрипции скорее всего является следствием, а не причиной активации. Ссылаясь на доказательства того, что только около 10% генома млекопитающих является функциональным и что среди основных биологических функций метилирования ДНК — наследственная репрессия транскрипции ретротранспозонов и инактивация Х-хромосомы в женских клетках, авторы считают, что «большая часть метилирования ДНК также, вероятно, будет иметь место. не иметь значительной биологической функции »[47].
По словам Бернхарда Хорстхемке, большинство исследований, в которых утверждается, что продемонстрировали трансгенерационную эпигенетическую наследственность через метилирование ДНК или РНК сперматозоидов, — исследования, которые показали реакцию на факторы метаболизма окружающей среды (диета с высоким содержанием жиров, ожирение, диабет, недоедание и травмы) в мыши и крысы — все еще ждут независимого подтверждения [61]. По словам Хорстхемке, очень сложно предоставить убедительные доказательства трансгенерационной эпигенетической наследственности у млекопитающих, особенно у людей, потому что ее изучение осложняется генетической наследственностью, а также влиянием экологии и культуры.Некоторые исследования, такие как исследования трансгенеративного воздействия эндокринных разрушителей и диеты с высоким содержанием жиров на метилом ДНК, были оспорены другими.
В ключевом исследовании якобы долгосрочных эффектов эндокринных разрушителей сообщается, что воздействие на беременных самок крыс эндокринного разрушителя винклозолина влияло на мужскую фертильность в последующих поколениях и что это было связано с эпигенетическими изменениями в зародышевой линии [62]. Эмма Уайтлоу обратила внимание на исследования, опровергающие подобные утверждения [63].Между тем широко цитируемое исследование Iqbal et al. убедительно показали, что эти эпигенетические изменения корректируются событиями репрограммирования зародышевой линии в следующем поколении [64]. Согласно Уайтлоу, доказательства эпигенетических эффектов, сохраняющихся более чем на одно поколение, как предполагалось в исследованиях трансгенерационных эффектов голландской голодной зимы и посттравматического стрессового расстройства после нападений на всемирный торговый центр [65, 66], неубедительны. Она добавляет тревожное наблюдение о том, что исследования, опровергающие эту идею, в основном отсутствуют в литературе: «Очень сложно публиковать отрицательные результаты, какими бы важными они ни были.В результате положительные исследования «кажутся неоспоримыми для тех, кто не работает в этой области» [63].
Согласно Хорстхемке, рост случаев сердечно-сосудистых и метаболических заболеваний у взрослых потомков беременных женщин, пострадавших от тяжелого недоедания во время голландского «Хонгервинтера», был вызван не передачей эпигенетической информации по материнской зародышевой линии, а прямым следствие воздействия в матке [61]. Он цитирует исследования, показывающие, что паттерны патологического метилирования ДНК могут быть результатом мутации в соседнем гене, которая влияет на патологическое метилирование промотора в этом гене.Поскольку это зависит от последовательности ДНК, передача этого паттерна метилирования следующему поколению не является примером трансгенерационного эпигенетического наследования.
Маюми Ивасакия и Ежи Пашковски утверждают, что перспектива факторов окружающей среды, включая стресс и материнскую заботу, унаследованных через эпигенетические изменения и влияющих на последующие поколения, «столь же интригующая, сколь и тревожная, поскольку можно представить себе, что накопление стресса воспоминания нескольких поколений могли затруднить принятие жизненных решений »[67].Исследуя высвобождение вредных эпигенетически подавленных транспозонов посредством абиотического стресса, они обнаружили механизм, который делает эту активацию только временной, быстро сбрасывая вызванные стрессом эпигенетические состояния, тем самым стирая «эпигенетическую стрессовую память» и тем самым предотвращая их митотическое распространение и наследование между поколениями. Они показали, что этот механизм сохраняется у растений и млекопитающих.
Методологические проблемы исследований ассоциаций в масштабе эпигенома
Исследования ассоциаций в масштабе всего эпигенома (EWAS), i.д., исследования изменений метилирования ДНК в отдельных геномах или геномах популяций широко используются для изучения того, могут ли изменения метилирования ДНК быть связаны с корреляцией фенотипов болезни с воздействием окружающей среды, в частности теми, которые происходят задолго до фенотипа. появился. Статистические проблемы и проблема невоспроизводимости таких исследований здесь не рассматриваются. Вместо этого в этом разделе освещаются до сих пор нерешенные проблемы относительно интерпретации EWAS, как это было определено Туули Лаппалайнен и Джоном Греалли [12].Сосредоточившись на интерпретируемости даже четко продемонстрированных изменений метилирования ДНК, авторы обнаруживают множество проблем, в том числе следующие:
-
«Часто расплывчатые определения и термины», которые используются при обсуждении эпигенетики;
-
Тот факт, что метилирование ДНК может изменяться «в ответ на разнообразные воздействия», в том числе наличие систематических различий в пропорциях клеточных подтипов между тестируемыми группами.
-
Тот факт, что большая часть различий в метилировании ДНК между людьми может быть связана с последовательностью ДНК. Исследование Gertz et al. семьи из трех поколений и неродственных особей показали, что последовательность ДНК составляет до 80% вариабельности метилирования ДНК [68]. По словам авторов, «большая часть вариаций в метилировании ДНК может быть объяснена генотипом», влияние которого на паттерны метилирования ДНК «значительно превосходит влияние импринтинга на паттерны метилирования в масштабе всего генома.Они пришли к выводу, что генотип необходимо будет принимать во внимание при оценке метилирования ДНК в контексте заболевания.
-
Обратная причинная связь, т.е. изменение метилирования ДНК как следствие транскрипции, отражающее, а не вызывающее различия в экспрессии генов. Это наблюдалось в ряде случаев (например, [69]; и в целом генетические и эпигенетические факторы тесно взаимосвязаны (см. Раздел «Невозможно отделить эпигенетику от генетики»).Lappalainen и Greally заключают, что тот факт, что «многие EWAS не измеряют и не учитывают генетические эффекты метилирования ДНК», является одной из причин нынешних проблем интерпретируемости исследований EWAS [12].
Статистические недостатки исследований, утверждающих, что продемонстрировали трансгенерационную эпигенетическую наследственность у людей, указаны Кевином Митчеллом, который указывает, например, на то, что шум интерпретируется как свидетельство, или на обоснование широких общих утверждений о трансгенерационных эпигенетических эффектах крошечными статистическими различиями [70].Статистические задачи в данной статье не рассматриваются.
Эпигенетика в психологии | ДНК Noba
расшифровывается как дезоксирибонуклеиновая кислота, и хотя ДНК каждого человека уникальна для этого человека, она на 99,9% похожа на ДНК любого другого человека на планете. [Изображение: CC0 Public Domain, https://goo.gl/m25gce]Раннее детство — это не только период физического роста; это также время умственного развития, связанного с изменениями в анатомии, физиологии и химии нервной системы, которые влияют на психическое здоровье на протяжении всей жизни.Когнитивные способности, связанные с обучением и памятью, рассуждением, решением проблем и развитием отношений, продолжают проявляться в детстве. В этот критический или чувствительный период развитие мозга происходит быстрее, чем в любой другой: каждую секунду создается более 700 нейронных связей. Здесь сложные взаимодействия ген-среда (или взаимодействия генотип-среда, G × E) служат для увеличения числа возможных контактов между нейронами, поскольку они оттачивают свои синаптические свойства и возбудимость у взрослых.Многие слабые связи образуются с разными нейронными целями; впоследствии они претерпевают реконструкцию, при которой исчезает большинство связей и остается несколько стабильных связей. Эти структурные изменения (или пластичность) могут иметь решающее значение для развития зрелых нейронных сетей, поддерживающих эмоциональное, когнитивное и социальное поведение. Генерация различных морфологических, физиологических и поведенческих результатов из одного генома в ответ на изменения в окружающей среде формирует основу для «фенотипической пластичности», которая имеет фундаментальное значение для того, как организмы справляются с изменчивостью окружающей среды, ориентируются в современном мире и решают будущие проблемы.
Задача психологии состояла в том, чтобы интегрировать результаты генетических и экологических (социальных, биологических, химических) факторов, включая качество привязанности младенца к матери, в изучение личности и наше понимание возникновения психических заболеваний. Эти исследования продемонстрировали, что общие вариации последовательности ДНК и редкие мутации составляют лишь небольшую часть (1–2%) от общего риска наследования черт личности и психических расстройств (Dick, Riley, & Kendler, 2010; Gershon, Alliey -Rodriguez, & Liu, 2011).Кроме того, исследования, которые пытались изучить механизмы и условия, при которых вариации последовательности ДНК влияют на развитие и функции мозга, были затруднены сложными причинно-следственными отношениями (Petronis, 2010). Большая неучтенная наследуемость личностных черт и психического здоровья предполагает участие дополнительных молекулярных и клеточных механизмов.
Эпигенетика может дать ответы на эти важные вопросы и относится к передаче фенотипа с точки зрения экспрессии генов в отсутствие изменений в последовательности ДНК — отсюда и название эпи- (греч. Επί- выше) генетика ( Waddington, 1942; Wolffe & Matzke, 1999).Появление высокопроизводительных методов, таких как подходы на основе секвенирования для изучения распределения регуляторов экспрессии генов по всему геному, привело к коллективному описанию «эпигенома». В отличие от последовательности генома, которая статична и одинакова почти во всех клетках, эпигеном очень динамичен, различаясь между типами клеток, тканями и областями мозга (Gregg et al., 2010). Недавние исследования предоставили понимание эпигенетической регуляции путей развития в ответ на ряд факторов внешней среды (Dolinoy, Weidman, & Jirtle, 2007).Эти факторы окружающей среды в раннем детстве и подростковом возрасте могут вызывать изменения в экспрессии генов, создающие риск психического здоровья и хронических физических состояний. Таким образом, изучение взаимодействий генетика-эпигенетика-среда с точки зрения развития может определить природу неправильной регуляции генов при психологических расстройствах.
В этом модуле будет представлен обзор основных компонентов эпигенома и рассмотрены темы недавних эпигенетических исследований, которые имеют отношение к психологии, чтобы сформировать биологическую основу для взаимодействия между сигналами окружающей среды и геномом в регулировании индивидуальных различий в физиологии. , эмоции, познание и поведение.
Однояйцевые близнецы — прекрасный пример эпигенетики. Хотя у них одна и та же ДНК, их уникальный жизненный опыт заставит одни гены (а не другие) проявить себя. Вот почему со временем однояйцевые близнецы начинают выглядеть и вести себя по-другому. [Изображение: M., https://goo.gl/VU5iJv, CC BY-NC-SA 2.0, https://goo.gl/Toc0ZF]Почти все клетки нашего тела генетически идентичны, но наше тело генерирует много разных типов клеток, организованных в разные ткани и органы и экспрессирующих разные белки.В каждом типе клетки млекопитающего около 2 метров геномной ДНК разделено на ядерные хромосомы. Однако ядро клетки человека, содержащее хромосомы, имеет диаметр всего около 2 мкм. Чтобы достичь этого 1000000-кратного уплотнения, ДНК оборачивается вокруг группы из 8 белков, называемых гистонами. Эта комбинация ДНК и гистоновых белков образует особую структуру, называемую «нуклеосомой», основной единицей хроматина, которая представляет собой структурное решение для поддержания и доступа к сильно уплотненному геному.Эти факторы изменяют вероятность того, что ген будет экспрессироваться или заглушаться. Таким образом, на клеточные функции, такие как экспрессия генов, репликация ДНК и образование определенных типов клеток, влияют различные паттерны структуры хроматина, включая ковалентную модификацию как гистонов (Kadonaga, 1998), так и ДНК (Razin, 1998).
Важно отметить, что эпигенетические вариации также проявляются на протяжении всей жизни. Например, хотя однояйцевые близнецы имеют общий генотип и являются генетически идентичными и эпигенетически подобными в молодом возрасте, с возрастом они становятся все более непохожими по своим эпигенетическим паттернам и часто проявляют поведенческие, личностные или даже физические различия и имеют разные уровни риска. при тяжелом заболевании.Таким образом, понимание структуры нуклеосомы является ключом к пониманию точного и стабильного контроля экспрессии и регуляции генов, обеспечивая молекулярный интерфейс между генами и изменениями клеточной активности, вызванными окружающей средой.
Метилирование ДНК — наиболее изученная эпигенетическая модификация, влияющая на экспрессию генов. ДНК состоит из четырех типов встречающихся в природе азотистых оснований: аденина (A), тимина (T), гуанина (G) и цитозина (C). В геномах млекопитающих метилирование ДНК происходит в основном по остаткам цитозина в контексте цитозинов, за которыми следуют гуанины (динуклеотиды CpG), с образованием 5-метилцитозина по клеточному образцу (Goll & Bestor, 2005; Law & Jacobsen, 2010; Suzuki & Bird, 2008 г.).Ферменты, осуществляющие метилирование ДНК, называются ДНК-метилтрансферазами (DNMT), которые катализируют перенос метильной группы в цитозин (Adams, McKay, Craig, & Burdon, 1979). Все эти ферменты экспрессируются в центральной нервной системе и динамически регулируются во время развития (Feng, Chang, Li, & Fan, 2005; Goto et al., 1994). Влияние метилирования ДНК на функцию гена варьируется в зависимости от периода развития, в течение которого происходит метилирование, и местоположения метилированного цитозина.Метилирование ДНК в регуляторных областях генов (промоторные и энхансерные области) обычно приводит к подавлению генов и снижению экспрессии генов (Ooi, O’Donnell, & Bestor, 2009; Suzuki & Bird, 2008; Sutter and Doerfler, 1980; Vardimon et al. , 1982). Это мощный регуляторный механизм, обеспечивающий экспрессию генов только при необходимости. Таким образом, метилирование ДНК может широко влиять на развитие мозга человека, а возрастная неправильная регуляция метилирования ДНК связана с молекулярным патогенезом нарушений развития нервной системы.
Жизненный опыт, как и стрессовое событие в детстве, может вызвать модификацию гистоновых белков (на фото), чтобы помочь адаптироваться к окружающей среде. Например, в ответ на стрессовое событие может произойти гистоновая модификация ДНК человека, чтобы стимулировать более осторожную личность — чтобы избежать будущих стрессовых столкновений. [Изображение: Zephyris, https://goo.gl/gGrSQd, CC BY-SA 3.0, https://goo.gl/kB1Ogc]Модификация гистоновых белков включает важную эпигенетическую метку, связанную с экспрессией генов.Одной из наиболее тщательно изученных модификаций является ацетилирование гистонов, которое связано с активацией генов и повышенной экспрессией генов (Wade, Pruss, & Wolffe, 1997). Ацетилирование гистоновых хвостов опосредуется противоположной ферментативной активностью гистоновых ацетилтрансфераз (HAT) и гистоновых деацетилаз (HDAC) (Kuo & Allis, 1998). Например, ацетилирование гистона в регуляторных областях генов ферментами HAT обычно связано с деметилированием ДНК, активацией генов и повышенной экспрессией генов (Hong, Schroth, Matthews, Yau, & Bradbury, 1993; Sealy & Chalkley, 1978).С другой стороны, удаление ацетильной группы (деацетилирование) ферментами HDAC обычно связано с метилированием ДНК, подавлением генов и снижением экспрессии генов (Davie & Chadee, 1998). Взаимосвязь между паттернами модификаций гистонов и активностью генов свидетельствует о существовании «гистонового кода» для определения программ экспрессии генов, специфичных для клеток (Jenuwein & Allis, 2001). Интересно, что недавние исследования с использованием моделей на животных продемонстрировали, что модификации гистонов и метилирование ДНК определенных генов опосредуют долгосрочные поведенческие эффекты уровня заботы, испытываемого в младенчестве.
Развитие личности — это активный процесс адаптации, происходящий в социально-экономическом контексте. Например, близость или степень позитивной привязанности между родителем (обычно матерью) и младенцем и родительские вложения (включая снабжение питательными веществами, обеспечиваемыми родителем), которые определяют опыт раннего детства, также программируют развитие индивидуальных различий в реакции на стресс в мозгу , которые затем влияют на память, внимание и эмоции. С точки зрения эволюции, этот процесс дает потомству возможность физиологически корректировать профили экспрессии генов, способствуя организации и функционированию нервных цепей и молекулярных путей, которые поддерживают (1) биологические защитные системы для выживания (например,g., стрессоустойчивость), (2) репродуктивный успех, способствующий установлению и устойчивости в нынешней среде, и (3) адекватное воспитание в следующем поколении (Bradshaw, 1965).
Наиболее полным на сегодняшний день исследованием вариаций родительского вклада и эпигенетической наследственности у млекопитающих является исследование материнской реакции на стресс у крыс. У крысят материнское воспитание (вылизывание и уход) в течение первой недели жизни связано с долгосрочным программированием индивидуальных различий в реакции на стресс, эмоциональности, когнитивных способностях и репродуктивном поведении (Caldji et al., 1998; Фрэнсис, Диорио, Лю и Мини, 1999; Лю и др., 1997; Майерс, Брунелли, Шаир, Сквайр и Хофер, 1989; Стерн, 1997). Во взрослом возрасте потомство матерей, которые демонстрируют повышенный уровень вылизывания и ухода за щенками в течение первой недели жизни, демонстрирует повышенную экспрессию рецептора глюкокортикоидов в гиппокампе (структура мозга, связанная с реакцией на стресс, а также обучением и памятью) и более низкую гормональная реакция на стресс по сравнению со взрослыми животными, выращиваемыми плохо вылизывающими и ухоженными матерями (Francis et al., 1999; Лю и др., 1997). Более того, детеныши крыс, которые получали низкие уровни материнского вылизывания и ухода в течение первой недели жизни, показали снижение ацетилирования гистонов и повышенное метилирование ДНК нейрон-специфического промотора гена глюкокортикоидного рецептора (Weaver et al., 2004). Затем экспрессия этого гена снижается, количество рецепторов глюкокортикоидов в головном мозге уменьшается, и животные демонстрируют более высокую гормональную реакцию на стресс на протяжении всей своей жизни. Влияние материнской заботы на реакцию гормона стресса и поведение потомства можно устранить в зрелом возрасте с помощью фармакологического лечения (ингибитор HDAC трихостатин А, TSA) или диетических аминокислотных добавок (метилдонор L-метионин), лечения, влияющего на ацетилирование гистонов, ДНК метилирование и экспрессия гена рецептора глюкокортикоидов (Weaver et al., 2004; Weaver et al., 2005). Эта серия экспериментов показывает, что ацетилирование гистонов и метилирование ДНК промотора гена глюкокортикоидного рецептора является необходимым звеном в процессе, ведущем к долгосрочным физиологическим и поведенческим последствиям плохого ухода за матерью. Это указывает на возможную молекулярную мишень для лечения, которое может обратить вспять или уменьшить следы жестокого обращения в детстве.
Родительская забота в детстве имеет важные и последующие последствия для развития человека, которые сохраняются даже во взрослой жизни.[Изображение: The White Ribbon Alliance, https://goo.gl/KgY6N5, CC BY-NC-SA 2.0, https://goo.gl/Toc0ZF]В нескольких исследованиях была предпринята попытка определить, в какой степени результаты модели животные могут быть переданы человеку. Изучение посмертной ткани мозга здоровых людей показало, что человеческий эквивалент промотора гена глюкокортикоидного рецептора (промотор 1F экзона NR3C1) также уникален для человека (Turner, Pelascini, Macedo, & Muller, 2008). Аналогичное исследование новорожденных показало, что метилирование промотора гена глюкокортикоидных рецепторов может быть ранним эпигенетическим маркером материнского настроения и риска повышенных гормональных реакций на стресс у младенцев в возрасте 3 месяцев (Oberlander et al., 2008). Хотя необходимы дальнейшие исследования для изучения функциональных последствий этого метилирования ДНК, эти результаты согласуются с нашими исследованиями на новорожденных и взрослых потомках матерей с низким уровнем облизывания и ухода, которые показывают повышенное метилирование ДНК промотора гена глюкокортикоидного рецептора, пониженное содержание глюкокортикоидов. экспрессия гена рецептора и усиление гормональной реакции на стресс (Weaver et al., 2004). Исследование мозговой ткани жертв суицида показало, что промотор гена рецептора глюкокортикоидов человека также более метилирован в мозге людей, переживших жестокое обращение в детстве (McGowan et al., 2009). Эти данные свидетельствуют о том, что метилирование ДНК опосредует эффекты ранней среды как у грызунов, так и у людей, и указывает на возможность новых терапевтических подходов, вытекающих из трансляционных эпигенетических исследований. Действительно, сходные процессы в сопоставимых эпигенетических лабильных регионах могут объяснить, почему взрослые потомки матерей с высоким и низким уровнем облизывания / ухода демонстрируют широко распространенные различия в экспрессии генов гиппокампа и когнитивных функциях (Weaver, Meaney, & Szyf, 2006).
Однако этот вид исследований ограничен недоступностью образцов человеческого мозга.Трансляционный потенциал этого открытия будет значительно увеличен, если соответствующая эпигенетическая модификация может быть измерена в доступной ткани. Изучение образцов крови взрослых пациентов с биполярным расстройством, которые также ретроспективно сообщали о своем опыте жестокого обращения и пренебрежения в детстве, показало, что степень метилирования ДНК промотора гена глюкокортикоидного рецептора человека имеет прямую положительную связь с описанным десятилетиями жестокого обращения с детьми. ранее.Для взаимосвязи между молекулярной мерой и зарегистрированным историческим воздействием величина эффекта чрезвычайно велика. Это открывает ряд новых возможностей: учитывая большой размер эффекта и постоянство этой ассоциации, измерение метилирования промотора GR может эффективно превратиться в анализ крови, измеряющий физиологические следы, оставленные в геноме ранним опытом. Хотя этот анализ крови не может заменить современные методы диагностики, эта уникальная и дополнительная информация дополняет наши знания о том, как болезнь может возникать и проявляться на протяжении всей жизни.В ближайшем будущем будет изучаться вопрос о том, добавляет ли эта мера ценность по сравнению с простым сообщением о ранних невзгодах, когда дело доходит до прогнозирования важных результатов, таких как реакция на лечение или самоубийство.
Независимо от того, знали ли ваши родители науку, говорящую вам есть овощи в детстве, вы действительно становитесь здоровее и сильнее — по крайней мере, ваша ДНК. [Изображение: Министерство сельского хозяйства США, https://goo.gl/tpyYzA, CC BY 2.0, https://goo.gl/BRvSA7]Старая пословица «ты то, что ты ешь» может быть верной не только в физический уровень: еда, которую вы выбираете (и даже то, что выбрали ваши родители, бабушки и дедушки), отражается на вашем личном развитии и риске заболевания во взрослой жизни (Wells, 2003).Питательные вещества могут обращать вспять или изменять метилирование ДНК и модификации гистонов, тем самым изменяя экспрессию критических генов, связанных с физиологическими и патологическими процессами, включая эмбриональное развитие, старение и канцерогенез. Похоже, что питательные вещества могут влиять на эпигеном либо путем прямого ингибирования ферментов, которые катализируют метилирование ДНК или модификации гистонов, либо путем изменения доступности субстратов, необходимых для этих ферментативных реакций. Например, матери крыс, которых кормили диетой с низким содержанием доноров метильных групп во время беременности, производили потомство со сниженной экспрессией DNMT-1, сниженным метилированием ДНК и повышенным ацетилированием гистонов в промоторных областях определенных генов, включая рецептор глюкокортикоидов, и повышенной экспрессии генов в печень молодого потомства (Lillycrop, Phillips, Jackson, Hanson, & Burdge, 2005) и взрослого потомства (Lillycrop et al., 2007). Эти данные предполагают, что питание в раннем возрасте может влиять на эпигенетическое программирование в мозге не только на раннем этапе развития, но и во взрослой жизни, тем самым влияя на здоровье на протяжении всей жизни. В этом отношении эпигенетика питания рассматривается как привлекательный инструмент для предотвращения педиатрических болезней развития и рака, а также для замедления процессов, связанных со старением.
Наилучшие свидетельства, касающиеся воздействия неблагоприятных условий окружающей среды на развитие и здоровье, получены из исследований детей беременных женщин во время двух голодовок среди гражданского населения во время Второй мировой войны: блокады Ленинграда (1941–44) (Bateson, 2001). и голландская голодная зима (1944–1945) (Stanner et al., 1997). Во время голода в Нидерландах женщины, которые ранее хорошо питались, подвергались низкому потреблению калорий и связанным с этим стрессовым факторам окружающей среды. Женщины, перенесшие голод на поздних сроках беременности, рожали младенцев меньшего размера (Lumey & Stein, 1997), и у этих детей был повышенный риск инсулинорезистентности в более позднем возрасте (Painter, Roseboom, & Bleker, 2005). Кроме того, у потомства, которого недоедали до рождения, позже в зрелом возрасте наблюдалось нарушение толерантности к глюкозе, даже когда пищи было больше (Stanner et al., 1997). Голод на различных стадиях беременности был связан с широким спектром рисков, таких как увеличение ожирения, более высокие показатели ишемической болезни сердца и более низкая масса тела при рождении (Lumey & Stein, 1997). Интересно, что при обследовании 60 лет спустя у людей, подвергшихся пренатальному голоданию, было обнаружено снижение метилирования ДНК по сравнению с их не подвергавшимися воздействию однополых братьев и сестер (Heijmans et al., 2008).
Нейропластичность — это изменение нервных путей и синапсов, которое позволяет нам узнавать новые вещи и запоминать их.[Изображение: Джерри Шоу, https://goo.gl/JBqlY7, CC BY-SA 3.0, https://goo.gl/eLCn2O]Воспоминания — это воспоминания о реальных событиях, хранящиеся в нашем мозгу. Но как наш мозг может формировать и хранить эти воспоминания? Эпигенетические механизмы влияют на активность генома в головном мозге, вызывая долгосрочные изменения в передаче сигналов, организации и морфологии синапсов, которые, в свою очередь, поддерживают обучение и память (Day & Sweatt, 2011).
Активность нейронов в гиппокампе мышей связана с изменениями метилирования ДНК (Guo et al., 2011), а нарушение работы генов, кодирующих механизм метилирования ДНК, вызывает нарушения обучения и памяти (Feng et al., 2010). Метилирование ДНК также участвует в поддержании долговременной памяти, например, в фармакологическом подавлении метилирования ДНК и нарушении памяти (Day & Sweatt, 2011; Miller et al., 2010). Эти открытия указывают на важность метилирования ДНК в обеспечении синаптической пластичности и когнитивных функций, которые нарушаются при психологическом заболевании.
Изменения модификаций гистонов могут также влиять на формирование долговременной памяти, изменяя доступность хроматина и экспрессию генов, имеющих отношение к обучению и памяти.Формирование памяти и связанное с этим усиление синаптической передачи сопровождается увеличением ацетилирования гистонов (Guan et al., 2002) и изменениями метилирования гистонов (Schaefer et al., 2009), которые способствуют экспрессии генов. Напротив, повышение нейрональной активности гистондеацетилазы, которая способствует молчанию генов, приводит к снижению синаптической пластичности и ухудшению памяти (Guan et al., 2009). Фармакологическое ингибирование гистондеацетилаз усиливает формирование памяти (Guan et al., 2009; Levenson et al., 2004), далее предполагая, что (де) ацетилирование гистонов регулирует этот процесс.
У людей генетические дефекты генов, кодирующих метилирование ДНК и хроматиновый аппарат, оказывают глубокое влияние на когнитивные функции и психическое здоровье (Jiang, Bressler, & Beaudet, 2004). Двумя наиболее описанными примерами являются синдром Ретта (Amir et al., 1999) и синдром Рубинштейна-Тайби (RTS) (Alarcon et al., 2004), которые представляют собой серьезные нарушения умственной отсталости. И MECP2, и CBP высоко экспрессируются в нейронах и участвуют в регуляции экспрессии нервных генов (Chen et al., 2003; Мартинович и др., 2003).
Пациенты с синдромом Ретта имеют мутацию в последовательности ДНК в гене MECP2. MECP2 играет много важных ролей в клетке: одна из этих ролей — считывать последовательность ДНК, проверять метилирование ДНК и связываться с участками, содержащими метилирование, тем самым предотвращая присутствие неправильных белков. Другие роли MECP2 включают содействие присутствию определенных необходимых белков, обеспечение правильной упаковки ДНК в клетке и помощь в производстве белков.Функция MECP2 также влияет на экспрессию генов, которые поддерживают развитие дендритов и синапсов и гиппокамп-зависимую память (Li, Zhong, Chau, Williams, & Chang, 2011; Skene et al., 2010). Мыши с измененной экспрессией MECP2 обнаруживают повышение ацетилирования гистонов во всем геноме, гибель нейронных клеток, повышенную тревогу, когнитивные дефициты и социальную изоляцию (Shahbazian et al., 2002). Эти находки подтверждают модель, в которой метилирование ДНК и MECP2 составляют клеточно-специфический эпигенетический механизм для регуляции модификации гистонов и экспрессии генов, которые могут быть нарушены при синдроме Ретта.
Пациенты с RTS имеют мутацию в последовательности ДНК в гене, называемом CBP. Одна из этих ролей CBP — связываться со специфическими гистонами и способствовать ацетилированию гистонов, тем самым способствуя экспрессии генов. В соответствии с этой функцией, пациенты с RTS обнаруживают снижение ацетилирования гистонов и когнитивную дисфункцию по всему геному во взрослом возрасте (Kalkhoven et al., 2003). Дефицит обучаемости и памяти объясняется нарушением нейронной пластичности (Korzus, Rosenfeld, & Mayford, 2004). Подобно RTS у людей, мыши с мутацией CBP плохо справляются с когнитивными задачами и обнаруживают пониженное ацетилирование гистонов по всему геному (для обзора см. Josselyn, 2005).Было обнаружено, что в мозге мышей CBP действует как эпигенетический переключатель, способствующий рождению новых нейронов в головном мозге. Интересно, что этот эпигенетический механизм нарушается в мозге плодов мышей с мутацией CBP, которые, будучи детенышами, обнаруживают ранние поведенческие дефициты после удаления и отделения от своей матери (Wang et al., 2010). Эти находки обеспечивают новый механизм, посредством которого экологические сигналы, действуя через ферменты, модифицирующие гистоны, могут регулировать эпигенетический статус и тем самым напрямую способствовать нейрогенезу, который регулирует нейроповеденческое развитие.
Вместе эти исследования демонстрируют, что неправильная регуляция эпигенетических модификаций и их регуляторных ферментов способна управлять заметными недостатками нейрональной пластичности и когнитивной функции. Знания, полученные в результате этих исследований, могут помочь лучше понять другие психические расстройства, такие как депрессия и суицидальное поведение.
На рисунке выше изображен хроматин, спиралевидная макромолекула, участвующая в депрессии. [Изображение: Zephyris, https://goo.gl/6DBQ1g, CC BY-SA 3.0, https: // goo.gl / eLCn2O]Исследования в масштабе всего эпигенома идентифицировали несколько десятков сайтов с изменениями метилирования ДНК в генах, участвующих в развитии мозга и путях нейротрансмиттеров, которые ранее были связаны с психическими заболеваниями (Mill et al., 2008). Эти расстройства являются сложными и обычно начинаются в молодом возрасте и вызывают пожизненную инвалидность. Часто ограниченная польза от лечения делает эти заболевания одними из самых обременительных для отдельных людей, семей и общества. Стало очевидным, что усилия по выявлению основных причин сложных психических расстройств могут значительно выиграть от исследований, связывающих воздействие окружающей среды с изменениями, наблюдаемыми в отдельных клетках.
Эпигенетические события, которые изменяют структуру хроматина для регулирования программ экспрессии генов, были связаны с поведением, связанным с депрессией, и действием антидепрессантов, и все больше доказательств сходных механизмов, происходящих в посмертном мозге депрессивных людей. У мышей социальное избегание привело к снижению экспрессии генов гиппокампа, важных в опосредовании депрессивных реакций (Tsankova et al., 2006). Аналогичным образом было обнаружено, что стресс хронического социального поражения снижает экспрессию генов, участвующих в нормальной обработке эмоций (Lutter et al., 2008). В соответствии с этими находками, уровни гистоновых маркеров повышенной экспрессии генов были подавлены в посмертных образцах мозга человека от людей с историей клинической депрессии (Covington et al., 2009).
Введение антидепрессантов увеличивало гистоновые маркеры повышенной экспрессии генов и обращало вспять репрессию генов, вызванную стрессом поражения (Lee, Wynder, Schmidt, McCafferty, & Shiekhattar, 2006; Tsankova et al., 2006; Wilkinson et al., 2009). Эти результаты подтверждают использование ингибиторов HDAC против депрессии.Соответственно, было обнаружено, что несколько ингибиторов HDAC проявляют антидепрессивный эффект, модифицируя отдельные клеточные мишени (Cassel et al., 2006; Schroeder, Lin, Crusio, & Akbarian, 2007).
Также появляется все больше доказательств того, что аберрантная экспрессия генов, возникающая в результате изменения эпигенетической регуляции, связана с патофизиологией суицида (McGowan et al., 2008; Poulter et al., 2008). Таким образом, возникает соблазн предположить, что существует эпигенетически обусловленная сниженная способность к экспрессии генов, которая необходима для обучения и памяти, в мозгу жертв суицида.
Хотя есть некоторые свидетельства того, что дисфункциональное воспитание может увеличить вероятность шизофрении (эпигенетически наследуемого заболевания), у некоторых людей, у которых есть как предрасположенность, так и стрессовая среда, никогда не развивается психическое заболевание. [Изображение: Стив Уайт, CC0 Public Domain, https://goo.gl/m25gce]Хотя клеточные и молекулярные механизмы, влияющие на физическое и психическое здоровье, долгое время находились в центре внимания нейробиологии, только в последние годы внимание переключилось на на эпигенетические механизмы, лежащие в основе динамических изменений экспрессии генов, ответственных за нормальную когнитивную функцию и повышенный риск психических заболеваний.Связи между ранним окружением и эпигенетическими модификациями предполагают механизм, лежащий в основе взаимодействий ген-окружение. Само по себе неблагоприятное воздействие окружающей среды в раннем возрасте не является достаточной причиной психического заболевания, поскольку многие люди, перенесшие тяжелое жестокое обращение или травмы в детстве, остаются здоровыми. Становится все более очевидным, что унаследованные различия в сегментах определенных генов могут смягчать последствия неблагоприятных факторов и определять, кто чувствителен, а кто устойчив, посредством взаимодействия генов и окружающей среды.Гены, такие как рецептор глюкокортикоидов, по-видимому, смягчают влияние невзгод в детстве на психические заболевания. Примечательно, что были выявлены эпигенетические модификации ДНК, которые могут лежать в основе длительного воздействия окружающей среды на биологические функции. Это новое эпигенетическое исследование указывает на новую стратегию понимания взаимодействия генов и окружающей среды.
Следующее десятилетие исследований покажет, можно ли использовать этот потенциал для разработки новых терапевтических возможностей, которые могут изменить следы, которые ранняя среда оставляет на геноме.Однако, как обсуждается в этом модуле, эпигеном не статичен и может формироваться сигналами развития, нарушениями окружающей среды и болезненными состояниями, что представляет собой экспериментальную проблему при поиске эпигенетических факторов риска психологических расстройств (Rakyan, Down, Balding, И Бек, 2011). Размер образца и требуемый эпигеномный анализ зависят от количества пораженных тканей, а также от типа и распределения эпигенетических модификаций. Сочетание исследований карт генетических ассоциаций с исследованиями развития на уровне всего эпигенома может помочь выявить новые молекулярные механизмы для объяснения особенностей наследования черт личности и трансформировать наше понимание биологических основ психологии.Важно отметить, что эти эпигенетические исследования могут привести к выявлению новых терапевтических целей и позволить разработать улучшенные стратегии ранней диагностики, профилактики и лучшего лечения психологических и поведенческих расстройств.
роль эпигенетики в эволюции человека | Горизонты биологических наук: Международный журнал исследований студентов
Аннотация
Этот обзор призван выделить ключевые области, в которых изменения эпигенома сыграли важную роль в эволюции и развитии нашего вида.Во-первых, будет краткое введение в тему эпигенетики, чтобы обрисовать текущее понимание предмета и проинформировать читателя об основных механизмах и функциях эпигенома. Это приведет к более подробному рассмотрению роли, которую играют эпигенетические изменения в быстрой эволюции нашего вида и появлении от наших предков, а также о человеческих ускоренных регионах, которые сыграли свою роль в этом. Обсуждение подчеркивает, как эпигенетика помогла и препятствовала развитию нашего вида посредством изменений в эпигеноме в более современное время, обсуждая примеры задокументированных случаев, когда показано, что эпигенетика сыграла роль в эволюции человечества.
Введение
Эволюция человека часто рассматривается как нечто абстрактное и далекое, что-то, что сыграло роль в развитии наших давно забытых предков, но не влияет на современное состояние нашего вида. Даже когда мы рассматриваем генетические условия, которые когда-то обеспечивали эволюционное преимущество популяциям ранних людей, но теперь стали опасными, часто думают, что современная медицина компенсирует и, в значительной степени, сводит на нет их симптомы.К ним относятся такие состояния, как серповидноклеточная анемия и сахарный диабет 1 типа; Многие из людей, страдающих этими заболеваниями, живут полноценной и здоровой жизнью благодаря современной медицине. Поэтому мы рассматриваем эти условия через призму болезни, а не как пережитки нашей эволюционной истории. Одно время серповидно-клеточная анемия обеспечивала важную защиту от малярии (Wiesenfeld, 1967), и есть свидетельства того, что сахарный диабет 1 типа развился у первых европейцев в результате адаптации к более холодному климату (Moalem et al., 2005). По мере того, как технологии развиваются и компенсируют эволюционные движущие силы этих черт, они больше не так выгодны для людей, которые их носят, как когда-то. Однако эволюция — это непрерывный процесс, от которого не исключены и современные люди, несмотря на то, что развитие науки снизило роль отбора. Люди не боятся последствий эволюции, и недавние исследования в области эпигенетики служат напоминанием об этом факте.
Эпигенетика — это механизм контроля генов, который может способствовать или подавлять экспрессию генов без изменения генетического кодирования организма (Feinberg, 2008).Другими словами, эпигенетика представляет собой систему, с помощью которой можно изменять экспрессию генов человека без изменения последовательности их генома. Наше текущее понимание выявило некоторые из контролирующих эпигенетических процессов, регулирующих экспрессию генов, которые называются эпигенетическими «метками». Например, метилирование ДНК, изменение молекул гистонов, которые удерживают вместе суперструктуры ДНК посредством метилирования или ацетилирования, и различные процессы, зависящие от РНК и Dicer-белка, которые ингибируют экспрессию генов.В совокупности сумма всех этих эпигенетических меток у человека называется эпигеномом. Этот обзор будет в основном сосредоточен на исследованиях, связанных с метилированием ДНК, поскольку это наиболее широко изученный эпигенетический механизм, но будут затронуты и другие аспекты эпигенома. Уже известно, что метилирование ДНК очень важно; например, крайние случаи деметилирования, представляющие собой потерю контроля над экспрессией генов, связаны с онкогенезом (Feinberg and Tycko, 2004).Этот обзор, однако, не будет сосредоточен на влиянии, которое изменения в эпигеноме могут иметь на здоровье человека, вместо этого он будет обсуждать роль, которую этот механизм контроля экспрессии генов сыграл в эволюции нашего вида, как появление человеческого виды и последствия, которые он оказал в последнее время.
Прежде чем мы сможем полностью осознать влияние, которое эпигенетика оказала на наш вид, необходимо более твердое понимание того, что такое эпигенетика и каким образом она может изменять экспрессию генов.Контроль над генами — увлекательная область генетики, и это особенно интересно тем, кто увлекается эволюцией человека. Эпигенетическое влияние на экспрессию генов, возможно, возникло как защита от транспозонов, паразитарной ДНК, которая прыгает в геноме и может нарушать работу генов, вставляя в их середину (Slotkin and Martienssen, 2007). Возможный механизм защиты может быть достигнут посредством метилирования ДНК, как показано на рис. 1. Выключение этих мобильных элементов и предотвращение или ограничение повреждения генома организма дает важное преимущество тем первым видам, которые разработали этот механизм.В конце концов этот процесс превратился в метод стимулирования и репрессии генов хозяина (Feinberg, 2008), которые можно было не только приобретать на протяжении всей жизни человека, но и передавать его потомству (Jones, 2012). Этот механизм сайленсинга генов, возможно, также позволил развитию многоклеточных организмов, позволяя одному геному адаптировать свои экспрессируемые гены в каждой отдельной клетке внутри более крупного организма (Badyaev, 2014).
Рисунок 1.
Метилирование как защита от транспозонов. На рисунке показано, как метилирование может помочь организму защитить себя от транспозонов. Со временем ДНК, кодирующая жизненно важные белки для транспозона, станет метилированной и перестанет экспрессироваться, таким образом, захватив транспозон в его текущем положении в геноме хозяина. Это предотвращает «прыжок» транспозона и возможное нарушение экспрессии жизненно важных генов в другом месте генома хозяина.
Рисунок 1.
Метилирование как защита от транспозонов.На рисунке показано, как метилирование может помочь организму защитить себя от транспозонов. Со временем ДНК, кодирующая жизненно важные белки для транспозона, станет метилированной и перестанет экспрессироваться, таким образом, захватив транспозон в его текущем положении в геноме хозяина. Это предотвращает «прыжок» транспозона и возможное нарушение экспрессии жизненно важных генов в другом месте генома хозяина.
Хотя эпигенетика представляет собой относительно новое понимание систем, участвующих в контроле и экспрессии генов, она также представляет собой нечто очень важное — фундаментальную переоценку теории эволюции.Приобретенные признаки, хотя и не являются изменениями генома, могут передаваться по наследству (Jones, 2012). В этом обзоре будут рассмотрены последствия этого для концепции эволюции человека и выделены интересные примеры и тематические исследования, в которых эти эффекты заметны.
Способ наследования
Изучение эпигенетики выявило интересный аспект этого метода контроля экспрессии генов. Метилирование ДНК и другие эпигенетические метки не изменяют гены, на которые они влияют на уровне последовательности, но, тем не менее, изменяют экспрессию этих генов.Кроме того, эти знаки могут быть приобретены на протяжении всей жизни человека, и, если они хранятся в их гаметах, эти знаки передаются по наследству. В этом разделе основное внимание будет уделено способам наследования этих меток.
Полуконсервативный характер митоза приводит к появлению двух наборов дочерних хроматид, по одному в каждом наборе, несущем эпигенетические метки из исходной хромосомы (Feinberg, 2008), как показано на рис. 2. Это позволяет переносить эпигенетические метки из материнские клетки дочерним клеткам в соматической ткани.Это объясняет, как эти отметки могут сохраняться у человека, но не как они передаются следующему поколению потомства.
Рисунок 2.
Сохранение эпигенетических меток во время митоза.
На рисунке показано, как эпигенетические метки могут поддерживаться в эпигеноме отдельной клетки во время митоза. Как видно из ( A ), исходная хромосома содержит эпигенетические метки на обеих хроматидах, а в ( B ) обе дочерние хромосомы содержат некоторые из эпигенетических меток материнской хромосомы из-за полуконсервативной природы митоза.
Рисунок 2.
Сохранение эпигенетических меток во время митоза.
На рисунке показано, как эпигенетические метки могут поддерживаться в эпигеноме отдельной клетки во время митоза. Как видно из ( A ), исходная хромосома содержит эпигенетические метки на обеих хроматидах, а в ( B ) обе дочерние хромосомы содержат некоторые из эпигенетических меток материнской хромосомы из-за полуконсервативной природы митоза.
Эти метки могут также сохраняться в дочерних хроматидах во время мейоза, в результате чего все гаметы несут эпигенетические метки индивидуума происхождения.Однако многие из этих следов удаляются в процессе образования гамет. Теперь понятно, что некоторые гены защищены от этого процесса деметилирования, в результате чего метки сохраняются в эпигеноме их гамет (Giuliani et al., 2015). Общая механика процесса оставляет некоторые вопросы без ответа, но известно, что модификация гистонов является ключом к идентификации участков генома потомства для метилирования после оплодотворения гамет (Samson et al., 2014).
Метки метилирования могут быть унаследованы от матери (Giuliani et al., 2015) или отцовской гаметы (Soubry, 2015). Через отца потомство может унаследовать широкий спектр меток метилирования, причем большинство этих меток каким-то образом влияет на пищеварительную систему ребенка (Soubry, 2015). Таким образом, собственная диета отца может влиять на развитие и адаптацию его ребенка, чтобы он лучше подходил к условиям питания, в которых он жил. Эпигеном матери-родителя оказывает гораздо большее влияние на условия питания. Мать имеет вдвое большее влияние на эпигеном своего потомства, во-первых, за счет собственных эпигенетических адаптаций, приобретенных ею в периконцепционный период своей жизни (Giuliani et al., 2015), а во-вторых, во время самой беременности (Heijmans et al., 2008). Сравнительно повышенное влияние эпигенома матери еще раз подчеркивается в случаях голода. Если затронутым родителем является мать, последствия вредных эпигенетических меток более сильно выражены в фенотипе потомства, особенно если голод наступает на ранних сроках беременности (Tobi et al., 2009). В качестве примера этой пагубной природы гипометилирования можно взглянуть на голландскую зиму голода, хорошо задокументированный пример голода в современном мире, который произошел с 1944 по 1945 год из-за блокировки, препятствовавшей перемещению топлива и продуктов питания в Нидерландах. .Это голодание привело к гипометилированию гена IGF-2 , гена, ответственного за образование инсулиноподобного фактора роста 2. Этот белок необходим для роста и развития плода, поэтому метилирование генов и последующее молчание привело к увеличению метаболических заболеваний у младенцев (Heijmans et al., 2008). Способность родительского недоедания оказывать явно негативное и вредное воздействие на эпигеном потомства будет более подробно изучена позже в обзоре.
Не все наследуемые эпигенетические метки унаследовали пищеварительную систему потомства, при этом многие из них влияют на иммунологические возможности потомства. Это было очень полезно и выгодно для кочевых народов, и пример этого можно увидеть при сравнении народов оромо и народов амхара в Эфиопии (Alkorta-Aranburu et al., 2012). В этом случае оказывается, что эпигенетические метки на самом деле способствуют иммунологической изменчивости среди вновь прибывшей популяции. Это будет рассмотрено более подробно в разделе «Примеры из практики» далее в обзоре.Другой случай унаследованных эпигенетических меток, влияющих на адаптацию человека к окружающей среде, — это сильное влияние, которое эпигеном человека оказывает на гомеостатическую систему (Gluckman, Hanson, and Spencer, 2005). На адаптацию человека к окружающей среде сильно влияют собственные приобретенные черты родителей, и по мере того, как сменяются поколения, каждое поколение будет накапливать все больше и больше эпигенетических меток, которые влияют на гомеостатические системы их поколений. В конечном итоге это приведет к тому, что популяция будет генетически похожа на первоначальных поселенцев, но будет более адаптирована к окружающей среде.Этот процесс приведет к возникновению гомеостатического фенотипа, который лучше компенсирует их естественный климат за более короткий промежуток времени, чем если бы фенотип популяции изменился под воздействием только комбинации генетической мутации и эволюционного давления.
Влияние на развитие человека как вида
Чтобы понять влияние эпигенетики на наше развитие в современных людях, мы должны сравнить области метилирования генов, наблюдаемые у нашего вида и наших ближайших ныне живущих родственников.Многие из областей в геноме человека, которые метилированы, не являются генами, уникальными для человека, с наибольшими различиями в метилировании, происходящими в областях ДНК, связанных с факторами транскрипции (или TF) и контролем генов (Hernando-Herraez et al., 2015). Это связано с тем, что TF имеют широкое влияние на выраженный фенотип индивидуума из-за того, что эти факторы функционируют как форма регуляции экспрессии генов, тем самым способствуя или подавляя другие гены в геноме. Даже небольшие различия в эпигеноме, окружающем ТФ, могут приводить к широкому различию фенотипов между особями одного и того же вида из-за их широкого влияния (Heyn et al., 2013). Таким образом, можно только предположить, насколько важны эти фенотипические изменения в вариации, которая отделяет нас от наших предков.
Следует отметить эпигенетические изменения, которые, как известно, произошли в регионах с ускоренным развитием человека (или HAR) (Hubisz and Pollard, 2014). HAR — это участки ДНК, которые претерпели быстрые изменения с момента появления человеческого вида, намного превышающие нормальную скорость мутаций. Эти регионы выделяются из-за чрезвычайно ускоренного темпа мутаций, которым они подверглись, и, как многие понимают, ответственны за быстрое отклонение людей от других видов (Hubisz and Pollard, 2014).Точная природа роли, которую играют эпигенетические изменения в HARs, неясна, но важность их роли несомненна, поскольку эпигенетические изменения, возможно, предшествуют последовательным изменениям в ДНК (Бадяев, 2014). Предлагаемая теория состоит в том, что эти отметки на самом деле способствовали возникновению мутаций в генах, ответственных за существование нашего вида.
Изучение эпигеномов наших родственных видов проливает свет на относительно большое расхождение, которое произошло с момента нашего появления от наших дальних родственников, расхождение такого роста, что его нельзя объяснить исключительно нуклеотидными изменениями (Hernando-Herraez et al., 2015). Высказывались даже предположения, что эпигенетические изменения могут иметь большее влияние на дарвиновскую эволюцию вида, чем геномные мутации (Бадяев, 2014), и эта область исследований только добавляет веса этим утверждениям.
Влияние на эволюцию современного человека
Современные люди выживали и процветали в самых разных средах на протяжении тысячелетий, от арктической тундры до пустынь Сахары. Ключом к этому успеху всегда была уникальная человеческая способность быстро адаптироваться, и эпигенетика сыграла роль в этой способности адаптироваться.В то время как культурная адаптация к окружающей среде, такая как смена одежды или ритуального поведения, являются наиболее наглядными признаками этой приспособляемости, не менее важны более тонкие генетические и эпигенетические изменения, которым подвергается популяция, когда она живет на определенной территории в течение нескольких поколений. Например, популяция, которая жила в засушливой среде, будет нести множество генетических мутаций, которые сделают ее более подходящей для засушливого климата. Если происходит катастрофический сдвиг климата и их исконные земли внезапно становятся холодными и сырыми, они могут адаптироваться к ношению более толстой одежды (Cavalli-Sforza and Feldman, 1983) для защиты от холода и могут даже принять новые обычаи и ритуалы, связанные с гигиеническим поведением для защиты. против новых болезней, которые прижились в регионе (Wiesenfeld, 1967).Однако эта популяция все еще будет нести многие генетические мутации, которые сделали их подходящими для их старой среды, пока давление отбора не позволит новым мутациям компенсировать эти генетические реликвии.
Важной идеей является способ рассмотрения каждого отдельного типа адаптации в сравнении друг с другом. Культурная адаптация — это универсальный термин, охватывающий все искусственные адаптации, которые человек может использовать, чтобы чувствовать себя более комфортно в окружающей среде (Cavalli-Sforza and Feldman, 1983).Одежда и развитие новых обычаев — все это полезные инструменты перед лицом экологических проблем, но они представляют собой краткосрочные адаптации, которые не влияют на выраженные фенотипы вида. Для сравнения, генетические изменения, такие как распространенность серповидной анемии в регионах, подверженных вспышкам малярии, представляют собой гораздо более долгосрочную адаптацию. Чтобы добиться этих изменений, нужно больше времени, и от них не так легко избавиться после того, как их полезность исчерпала себя, например, когда человек просто меняет свою одежду в соответствии с погодой (Laland, Odling-Smee, and Myles, 2010).
Что же тогда представляют собой эпигенетические изменения в этой модели? Во-первых, они иллюстрируют среднесрочные адаптации, находящиеся между культурными изменениями и генетической эволюцией во времени, которое требуется человеку для их приобретения (Giuliani et al., 2015). В этой модели понимания адаптации человека эпигенетические изменения также служат механизмом хронометража, помогая смягчить негативные эффекты генетических реликвий, приобретенных популяциями предков под различным эволюционным давлением (Бадяев, 2014).Заглушая старые гены, которые когда-то служили жизненно важной цели, эпигенетика также помогает предотвратить накопление сложности в организме, заглушая старые, менее часто транскрибируемые гены (Бадяев, 2014), почти так же, как метилирование ДНК борется с нанесенным ущербом. транспозонами (Slotkin, Martienssen, 2007).
Хороший способ изучить эту модель адаптации — рассмотреть, каким образом каждое из этих изменений повлияет на гипотетическую популяцию, которая внезапно стала подвергаться суровому, холодному климату.Эта популяция очень быстро адаптируется, во-первых, за счет усиления защиты от непогоды за счет более толстой одежды. Хотя это эффективный метод сохранения тепла, их тела еще не адаптировались к холоду, и поэтому их гены, контролирующие гомеостаз, будут по-прежнему функционировать так же, как и в более теплом климате, что может быть значительно расточительным и, возможно, вредным. . Если когда-то их пот помогал удерживать тепло от повреждения их тел, теперь он расходует воду.На этом этапе, по прошествии значительного числа поколений, эпигенетические изменения начнут проявляться под давлением отбора. Метилирование ДНК и модификации гистонов будут накапливаться, точно настраивая их гомеостатическую экспрессию генов на более холодную среду. Это приводит к подавлению генов, которые лучше подходят для более жаркого климата, и способствует экспрессии других генов, которые дают преимущество в более холодном климате. Наконец, через еще большее количество поколений новые аллели закрепятся в популяциях, представляющих новые гены.Эти новые гены будут кодировать новые белки, которые тем или иным образом обеспечат селективное преимущество, которое почти постоянно проявляется в экспрессии, если не дает преимущества.
В рассмотренной выше модели краткосрочной и долгосрочной адаптации, чем больше период времени требуется для приобретения и адаптации, тем более значительными будут изменения в физиологии человека и выраженном фенотипе. Еще один момент, подчеркнутый этой моделью, заключается в том, что чем дольше адаптация приобретается, тем меньше вероятность ее утраты.Ведь намного легче снять куртку, чем спонтанно потерять ген, отвечающий за повышение метаболической активности. Здесь снова появляется эпигенетика, поскольку она не только заставляет замолчать старые гены, которые больше не нужны, под влиянием давления отбора, но и вносит большую пластичность в экспрессию генов (Giuliani et al., 2015), позволяя людям которые несут одинаковый или невероятно похожий геном с измененной экспрессией генов. Благодаря этому механизму эпигенетика допускает вариабельность фенотипов, необходимых для адаптации и отбора (Tobi et al., 2009).
Примеры из практики
Интересным исследованием эпигенетики, которое представляет собой пример ее роли в эволюции человека, является исследование различных эпигенетических маркеров между народами оромо и амхара в высокогорье Эфиопии, которое выявило нечто удивительное: исследователи ожидали обнаружить, что представители народов оромо , которые являются мигрантами в высокогорье, будут иметь увеличение эпигенетических меток вокруг генов, связанных с потреблением кислорода или производством красных кровяных телец.Это приспособления, которые уже были у народов амхара, позволяющие им успешно жить на своей возвышенной родине. Вместо этого многие эпигенетические маркеры, обнаруженные исследователями в популяции оромо, связаны с генами, связанными с иммунной системой (Alkorta-Aranburu et al., 2012). Более интересным было то, что эти отметки не были однородными среди населения, а вместо этого широко варьировались от человека к человеку (Alkorta-Aranburu et al., 2012). Это, по-видимому, демонстрирует эпигенетические метки, действующие как катализатор внесения вариаций в экспрессию генов, что приводит к широкому спектру фенотипов и ответов для борьбы с новыми микробиологическими угрозами, которым мигрирующая популяция подвергалась по прибытии в регион.
Здесь эти эпигенетические метки компенсируют отсутствие иммунологической адаптации народов оромо к этому новому климату по сравнению с коренным народом Амхара, смягчая ущерб, нанесенный популяциям оромо в промежуточный период до того, как может произойти генетическая мутация, которая обеспечивает более сильную защита.
Как обсуждалось ранее, эпигенетика играет ключевую роль в пищевой адаптации индивидов, производя индивидов, несущих эпигенетические метки, которые делают их более приспособленными к диете их родителей.Толерантность к лактозе — один из способов проявления этой эпигенетической пищеварительной адаптации (Ingram et al., 2009). Фенотипы «неоднородной» толерантности к лактозе наблюдались в популяциях, лишенных мутации толерантности к лактозе. С увеличением доступности молочных продуктов во всем мире можно ожидать, что эпигенетическая модификация, вызывающая более слабую толерантность к лактозе, будет только увеличиваться, по крайней мере, до тех пор, пока мутация толерантности к лактозе не распространится в глобальном генофонде.
Различные генетические состояния влияют на эритроциты и их способность поглощать кислород.К ним относятся серповидноклеточная анемия и талассемия, которые возникают только после того, как у человека полностью переключится на гемоглобин взрослых (Sripichai et al., 2009). В случае серповидной анемии известно, что это состояние придает устойчивость к малярии. По мере того, как противомалярийные методы лечения становятся все более и более эффективными, а уничтожение комаров начинает сдерживать рост инфекций, это стало условием, которое в настоящее время в основном служит бременем для молодых служб здравоохранения во всем мире.Поскольку давление отбора на эти популяции изменилось, эпигеном этих популяций также отреагировал. В двух различных исследованиях были обнаружены эпигенетические маркеры, которые могут вызывать сохранение гемоглобина плода (или POFH) (Sripichai et al., 2009; Sankaran, Xu, and Orkin, 2010). POFH — это состояние, при котором человек никогда не переключается на гемоглобин взрослого и, таким образом, избегает экспрессии мутаций серповидной анемии и талассемии. Хотя они все еще несут эти мутации, эти люди не проявляют вредных фенотипов из-за эпигенетических маркеров, которые ингибируют связанные гены.Это показано на рис. 3.
Рисунок 3.
Эпигенетическая профилактика серповидной анемии.
На рисунке показано, как эпигенетические метки могут предотвратить экспрессию мутации серповидных клеток. ( A ) показывает, что происходит у человека, который несет мутации, но не имеет никаких эпигенетических меток, подавляющих ген переключения взрослого гемоглобина. У этих людей в конечном итоге разовьется серповидноклеточная анемия. В ( B ) ген переключения гемоглобина взрослого человека подавляется эпигенетическими метками, и поэтому ген серповидной анемии никогда не экспрессируется, поскольку индивидуум поддерживает выработку гемоглобина плода.
Рисунок 3.
Эпигенетическая профилактика серповидной анемии.
На рисунке показано, как эпигенетические метки могут предотвратить экспрессию мутации серповидных клеток. ( A ) показывает, что происходит у человека, который несет мутации, но не имеет никаких эпигенетических меток, подавляющих ген переключения взрослого гемоглобина. У этих людей в конечном итоге разовьется серповидноклеточная анемия. В ( B ) ген переключения гемоглобина взрослого человека подавляется эпигенетическими метками, и поэтому ген серповидной анемии никогда не экспрессируется, поскольку индивидуум поддерживает выработку гемоглобина плода.
Уязвимости эпигенетики
В этом обзоре изменения эпигенома человека обсуждались в чрезмерно позитивном свете. Такие изменения могут быть механизмом, обеспечивающим более быструю форму адаптации, чем генетические мутации (Giuliani et al., 2015). Он также может действовать как механизм отсчета времени в более старых, а теперь и вредных генах (Бадяев, 2014). Наконец, это может быть влиятельная реакция на дарвиновское давление на человека или популяцию (Alkorta-Aranburu et al., 2012). Как и в любом процессе адаптации, существует отрицательная сторона, и, как и генетические мутации, эпигенетические метки часто наносят вред здоровью человека. Например, существует известная связь между гипердеметилированием и онкогенезом (Feinberg and Tycko, 2004), но в этом обзоре будут рассмотрены негативные аспекты изменений эпигенома с точки зрения их влияния на эволюцию человека.
Эпигенетика может действовать как механизм отсчета времени, подавляя гены, которые отжили свое предназначение.Однако стоит отметить, что эта функция эпигенетических меток имеет ограничения, поскольку во многих случаях рассматриваемый ген не заглушается полностью (Бадяев, 2014), а вместо этого экспрессируется с более низкой скоростью. С дарвиновской точки зрения это нежелательное последствие, поскольку индивидуум выживет, но его потомство теперь будет обладать чертой, снижающей его способность к адаптации. Таким образом могут накапливаться многие черты, снижающие общую приспособленность вида.
Роль, которую эпигенетика играет в пищеварительной адаптации, также является палкой о двух концах.Это было продемонстрировано исследованиями многочисленных случаев голода, включая современные примеры, такие как голландская зима голода (Tobi et al., 2009; Giuliani et al., 2015; Soubry, 2015). Исследования показывают, что подверженность родителей голоду приводит к накоплению отрицательных черт у потомства из-за нарушения регуляции меток метилирования, включая склонность к диабету и ожирению, а также увеличение частоты сердечно-сосудистых заболеваний (Heijmans et al., 2008).
Наконец, и, возможно, самым пугающим аспектом эпигенома является способность людей, переживших травмирующие переживания, которые подвергли их разум и тело экстремальному стрессу, приобретать, а затем передавать полученные эпигенетические метки и черты этого времени в их жизни.Наиболее ярким примером этого является накопление эпигенетических меток у потомков переживших Холокост, что приводит к заметному увеличению посттравматического стрессового расстройства, депрессии и ожирения, что является результатом дифференциального метилирования гена FKBP5 (Yehuda et al., 2015) . Хотя эти эффекты менее серьезны, чем генетические мутации, эффекты изменений эпигенома, в силу того, что они менее сильны по своему влиянию на фенотип, позволяют накапливать признаки, которые, если бы они были выражены на уровне последовательности, не были бы сохранены. новое поколение.
Заключение
В этом обзоре обсуждались различные эффекты, которые эпигеном оказывает на эволюцию нашего вида. Его положительная способность действовать как ответ на давление отбора и как способ смягчения вредных мутаций может быть выгодной. Однако его значительный контроль над экспрессией генов также может привести к вредным последствиям, например как путь к онкогенезу или как механизм накопления черт, снижающих приспособленность и приспособляемость.Самое важное, что нужно понимать об эпигеноме, — это то, что он не всегда связан с положительным или отрицательным влиянием на развитие нашего вида, а скорее с тем, что он придает пластичность нашему геному (Giuliani et al., 2015). Поощряя вариабельность и адаптивность нашего вида, эпигенетические механизмы контроля экспрессии генов гарантируют, что человечество может выживать и процветать в любом количестве сред. Эпигенетика является важной частью причины, по которой наш вид стал настолько адаптируемым, и эта черта, как часто думают, отличает нас от тех, кого мы часто считаем менее развитыми и развитыми животными, с которыми мы обитаем на этой Земле.В самом деле, можно утверждать, что эпигенетика отвечает за и предоставляет нашему виду инструменты, которые действительно сделали нас уникальными в нашей способности завоевывать любую среду обитания и адаптироваться практически к любому климату. Изучение эпигенетики также сделало эволюцию нашего вида менее абстрактной и отдаленной; Теперь мы можем лучше понять влияние различных черт на уровне поколений и лучше наблюдать движущие факторы, стоящие за изменениями нашего вида. Что еще более важно, это свидетельствует о том, что человечество не находится выше и не остается равнодушным к последствиям избирательного давления.Наконец, более глубокое понимание эпигенетики изменило то, как мы думаем об эволюции, составив фундаментальное переосмысление темы и того, как этот механизм позволяет нам приобретать черты в течение всей жизни и передавать эти черты нашему потомству. Конечно, выживание наиболее приспособленных остается золотым правилом эволюции, но, углубляясь в эпигеном, мы понимаем, что черты, управляющие приспособленностью, и, следовательно, приспособленностью индивидуума, более изменчивы и податливы, чем если бы они рассматривались исключительно через призму объективности. генетическая мутация и унаследованные черты.
Самый важный урок, извлеченный из изучения эпигенома нашего вида, заключается в том, что оно дало понимание факторов, отделяющих нас от наших ближайших живых родственников в животном царстве, которые нельзя объяснить только генетическими мутациями.
Outlook
Центральной теоремой, на основе которой был сделан этот обзор, является понимание эпигенетики с точки зрения ее роли в обеспечении среднесрочной адаптации. Чтобы развить эту гипотезу и подтвердить доказательства, представленные в этом обзоре, необходимо провести более глубокое исследование появления эпигенетических меток у популяций, сталкивающихся с изменяющимся давлением отбора.Работа, проделанная по изучению и сравнению эпигенома аборигенных и недавно мигрировавших популяций в высокогорных районах Эфиопии, является убедительным примером того, как можно проводить это исследование (Alkorta-Aranburu et al., 2012). Более того, идеальной областью, которая могла бы пролить еще больше информации о роли эпигенетики в эволюции человека, является сравнение популяций Верхнего и Нижнего Нила. Эти районы исторически сталкивались с аналогичными угрозами в виде малярии, но развитие вдоль Нижнего Нила, подкрепленное инвестициями в профилактику со стороны египетского правительства, в последние десятилетия резко снизило темпы перехода малярии комарами.Кроме того, в обеих этих областях отмечены значительные уровни серповидной анемии (El-Hazmi, Al-Hazmi and Warsy, 2011). Сочетание обоих этих факторов делает этот регион идеальным для исследования возникающих эпигеномных изменений в условиях изменения давления отбора. Можно предсказать, что эпигенетические метки, заглушающие эффекты серповидноклеточной анемии (Sripichai et al., 2009), как обсуждалось ранее, станут более распространенными в этих областях, но это не может быть известно наверняка, как это было в случае с предсказанным. изменения эпигенома в Эфиопском нагорье (Alkorta-Aranburu et al., 2012).
Биография автора
Когда я закончил факультет генетики Университета Глазго, мои интересы начали сосредоточиваться на области эпигенетики довольно поздно в моей академической карьере. Однако после того, как я погрузился в эпигенетику и огромное количество вопросов и теорий, поднятых этой темой, которые перевернули генетику с ног на голову, эпигенетика очаровала меня. Я надеюсь, что когда-нибудь либо проведу исследование влияния эпигенетики на эволюцию человека, либо стану журналистом, специализирующимся на этой теме.
Благодарности
Я хотел бы поблагодарить моего консультанта доктора Криса Финли за его поддержку на протяжении всего моего исследования и указание на конечную цель моего исследования эпигенетики. Он предоставил начальные документы, которые дали мне более глубокое представление о работе по исследованию роли эпигенома в развитии человеческого вида.
Список литературы
Алькорта-Аранбуру
,г.
,Билл
,К.М.
, Витонский,
Д. Б.
и др. . (2012
)Генетическая архитектура адаптации к большой высоте в Эфиопии
,PLoS Genetics
,8
(12
),e1003110
.
Бадяев
,AV
(2014
)Эпигенетическое разрешение «проклятия сложности» в адаптивной эволюции сложных признаков
,The Journal of Physiology
,592
,2251
—2260
2260 .
Cavalli-Sforza
,LL
иFeldman
,MW
(1983
)Культурная адаптация против генетической
,Труды Национальной академии наук
,80
(80
)4993
—4996
.
Эль-Хазми
,MAF
,Аль-Хазми
,AM
иWarsy
,AS
(2011
)Серповидноклеточная болезнь в арабских странах Ближнего Востока
,Индийский медицинский журнал Исследования
,134
(5
),597
—610
.
Feinberg
,A. P.
(2008
)Эпигенетика в эпицентре современной медицины
,Журнал Американской медицинской ассоциации
,299
(11
),1345
— 13502.
Feinberg
,A. P.
иTycko
,B.
(2004
)История рака Эпигенетика
,Nature Reviews. Рак
,4
(2
),143
—153
.
Giuliani
,C.
,Bacalini
,M. G.
,Sazzini
,M.
et al. . (2015
)Эпигенетическая сторона адаптации человека: гипотезы, доказательства и теории
,Анналы биологии человека
,42
(1
),1
—9
.
Gluckman
,P. D.
,Hanson
,M. A.
иSpencer
,H.G.
(2005
)Прогнозные адаптивные реакции и эволюция человека
,Тенденции в экологии и эволюции
,20
(10
),527
—533
.
Heijmans
,B. T.
,Tobi
,E. W.
,Stein
,A. D.
et al. . (2008
)Стойкие эпигенетические различия, связанные с пренатальным воздействием голода у людей
,Proceedings of the National Academy of Sciences
,105
(44
),17046
—17049
.
Hernando-Herraez
,I.
,Heyn
,H.
,Fernandez-Callejo
,M.
et al. . (2015
)Взаимодействие между метилированием ДНК и расхождением последовательностей в недавней эволюции человека
,Nuclear Acids Research
,43
(17
),8204
—8214
.
Heyn
,H.
,Moran
,S.
,Hernando-Herraez
,I.
et al. . (2013
)Метилирование ДНК способствует естественной изменчивости человека
,Genomic Research
,23
,1363
—1372
.
Hubisz
,MJ
иPollard
,KS
(2014
)Изучение происхождения и функций регионов с ускоренным развитием человека проливает свет на их роль в эволюции человека
,Current Opinion
,29
,15
—21
.
Ingram
,C. J. E.
,Mulcare
,C. A.
,Itan
,Y.
et al. . (2009
)Переваривание лактозы и эволюционная генетика устойчивости лактазы
,Human Genetics
,124
(6
),579
—591
.
Jones
,PA
(2012
)Функции метилирования ДНК: островки, стартовые сайты, тела генов и за их пределами
,Nature Reviews Genetics
,13
(7
),484
—492
.
Laland
,KN
,Odling-Smee
,J.
иMyles
,S.
(2010
)Как культура сформировала геном человека: объединение генетики и гуманитарных наук
,Обзоры природы Генетика
,11
(2
),137
—148
.
Моалем
,С.
,Этаж
,К. Б.
,Перси
,М.E.
et al. . (2005
)Самое приятное в диабете 1 типа: криопротекторная эволюционная адаптация
,Медицинские гипотезы
,65
(1
),8
—16
.
Samson
,M.
,Jow
,M. M.
,Wong
,C. C.
et al. . (2014
)Спецификация и глобальное перепрограммирование гистона Эпигенетические метки во время формирования гамет и раннего развития эмбриона у C.elegans
,PLoS Genetics
,10
(10
),e1004588
.
Sankaran
,VG
,Xu
,J.
иOrkin
,SH
(2010
)Достижения в понимании переключения гемоглобина
,Британский журнал гематологии
(
2
),181
—194
.
Слоткин
,р.K.
иMartienssen
,R.
(2007
)Мобильные элементы и эпигенетическая регуляция генома
,Nature Reviews Genetics
,8
(4
),272
—285
.
Субри
,A.
(2015
)Эпигенетическое наследование и эволюция: взгляд отца на влияние питания
,Прогресс в биофизике и молекулярной биологии
,118
(1–2
), 79 —85
.
Sripichai
,O.
,Kiefer
,C. M.
,Bhanu
,N. V.
et al. . (2009
)Цитокин-опосредованное увеличение гемоглобина плода связано с модификацией гистона глобина и перепрограммированием фактора транскрипции
,Кровь
,114
(11
),2299
—2306
.
Tobi
,E. W.
,Lumey
,L.H.
,Таленс Рудольф
,P.
et al. . (2009
)Различия в метилировании ДНК после воздействия пренатального голода являются обычными и зависят от времени и пола
,Human Molecular Genetics
,18
(21
),4046
—4053
.
Wiesenfeld
,S. L.
(1967
)Серповидноклеточный признак в биологической и культурной эволюции человека
,Science (New York, N.Ю.)
,157
(3793
),1134
—1140
.
Yehuda
,R.
,Daskalakis
,N. P.
,Bierer
,L. M.
et al. . (2015
)Воздействие холокоста вызвало межпоколенческие эффекты метилирования FKBP5
,Биологическая психиатрия
,80
,652
—656
.Заметки автора
Руководитель: Кристофер Финлей, Университет Глазго, Глазго G12 8QQ, Великобритания
Как выпускник факультета генетики Университета Глазго, мои интересы начали сосредотачиваться на области эпигенетики довольно поздно в моей академической карьере.Однако после того, как я погрузился в эпигенетику и огромное количество вопросов и теорий, поднятых этой темой, которые перевернули генетику с ног на голову, эпигенетика очаровала меня. Я надеюсь, что когда-нибудь либо проведу исследование влияния эпигенетики на эволюцию человека, либо стану журналистом, специализирующимся на этой теме.
© Автор, 2017. Опубликовано Oxford University Press.
Это статья в открытом доступе, распространяемая в соответствии с условиями некоммерческой лицензии Creative Commons Attribution (http: // creativecommons.org / licenses / by-nc / 4.0 /), который разрешает некоммерческое повторное использование, распространение и воспроизведение на любом носителе при условии правильного цитирования оригинальной работы. По вопросам коммерческого повторного использования обращайтесь по адресу [email protected].(PDF) Эпигенетические механизмы кембрийского взрыва
Nelson R. Cabej
CABEJ
ЭПИГЕНЕТИЧЕСКИЕ
МЕХАНИЗМЫ
КАМБРИЙСКИЙ ВЗРЫВ
ВЗРЫВ
ЭПИГЕНЕТИЧЕСКИЙ МЕХАНИЗМ
ЭПИГЕНЕТИЧЕСКИЙ МЕХАНИЗМ
ВЗРЫВ
9780128 143117
ISBN 978-0-12-814311-7
Уникально опирается на эпигенетическую теорию эволюции для объяснения быстрой диверсификации метазоан
того периода
Кембрийский взрыв стал беспрецедентно быстрой диверсификацией животных
542 миллиона лет назад.В эволюционный «миг» примерно 10 миллионов лет, из которых
представляют лишь 0,3% времени существования жизни на Земле, появились все 30 существующих планов тела
и основные группы животных.
За последние полвека был выдвинут ряд гипотез, объясняющих кембрийский взрыв
, но эти теории не имеют подтверждающих доказательств или не объясняют все изменения, произошедшие во время кембрийского взрыва, из
.В книге «Эпигенетические механизмы кембрийского взрыва
» автор использует свою эпигенетическую теорию эволюции для объяснения причинной основы кембрийского взрыва
. Он представляет как эмпирические доказательства, так и теоретическую аргументацию
в поддержку этой эпигенетической теории.
Эпигенетические механизмы кембрийского взрыва предоставляет читателям базовые знания о
эпигенетическом объяснении этой биологической загадки и помогает ответить на оставшиеся вопросы
кембрийского взрыва для эволюционных биологов, эпигенетиков и других научных
исследователей.
Основные характеристики
● Объясняет кембрийский взрыв с полностью эпигенетической точки зрения
● Использует причинный, а не описательный подход к явлению
● Обеспечивает широкий круг читателей, в том числе тех, кто обладает только базовыми биологическими знаниями, в то время как
соблюдение научной строгости
Об авторе
Д-р Нельсон Р. Кабей является автором книг «Эпигенетические принципы эволюции» (Elsevier, 2011) и
«Построение самой сложной структуры на Земле» (Elsevier, 2013), в которых он исследует роль
эпигенетических механизмов в эволюции организма.Он опубликовал более 40 научных
статей и 10 книг на английском и его родном албанском языке по эволюционной биологии,
эпигенетике и биологии развития. Доктор Кабедж получил докторскую степень по биологии в университете
в Тиране, Албания, и в настоящее время работает научным сотрудником на кафедре биологии в университете
.
Эпигенетические изменения: Ламарк, просыпайся, тебя разыскивают в конференц-зале!
Если мы хотим услышать, что говорят нам окаменелости, мы должны выяснить, как они стали такими, какими они были (и как их потомки стали такими, какие они есть сегодня).В течение нескольких поколений ученые обращались к дарвиновской эволюции (естественный отбор, действующий на случайные мутации) почти исключительно для объяснения всего этого. Вопросы считались решенными, если можно было предоставить дарвиновский счет.
Это произошло не потому, что любой закон природы показывает, что дарвинизм должен быть правильным объяснением. Скорее потому, что, как выразился Ричард Докинз, «дарвинизм — единственная известная теория, которая в принципе способна объяснить определенные аспекты жизни» (стр. 287, Blind Watchmaker, 1986).
Но когда мы посмотрели на горизонтальный перенос генов (ГПГ), мы обнаружили, что утверждение Докинза неверно.
Напомним, что дарвинизм предполагает вертикальную передачу генов от общего предка потомкам. Горизонтальный перенос генов означает передачу генов от одного организма к другому при контакте, , независимо от происхождения любой формы жизни. Да, ХПГ — это форма эволюции. Но это резко ослабляет статус дарвинизма как «единственно известной теории». Любое дарвиновское утверждение об эволюции должно сначала исключить ГПГ как возможное объяснение.И, как мы вскоре увидим, это также должно исключать эпигенетику.
Почему этому историческому сдвигу в бремени доказывания уделяется сравнительно мало внимания? Вероятно, это связано с всеобщим принятием дарвинизма как культурной метафоры и философии жизни. Например, можно вспомнить, как Amazon цитирует «целенаправленный дарвинизм» и максимально использует теорию Дарвина в качестве защиты от недавнего разоблачения условий труда в фирме. Концепции, которые продвигает Amazon, бессмысленны с научной точки зрения, но имеют культурное значение.А культура заглушает науку.
Таким образом, при разговоре с окаменелостями (или нынешними живыми формами) наша задача состоит в том, чтобы прислушиваться к тому, что они говорят, а не к тому, что говорят дарвиновские интерпретаторы окаменелостей (и почти всего остального).
Это подводит нас к эпигенетике. Жан-Батист Ламарк (1744-1829) был одним из первых эволюционистов, который предположил, что формы жизни могут получать информацию из окружающей среды и передавать ее своим генам. Дарвинисты в течение многих десятилетий увольняли его, хотя и не высмеивали его (хотя, как это бывает, не Дарвин).Но основная направленность его идеи недавно нашла отражение в эпигенетике.
В том, как произошло возрождение, есть ирония. Ключевым научным достижением 1990-х годов стало картирование генома человека. Это были дни, когда лауреат премии Вальтер Гилберт, превознося проект «Геном человека», держал компакт-диск с данными и сообщал своей аудитории: «Вот человек; это я!»
Но что, если это не так? Что, если это просто компакт-диск, содержащий важную информацию о Гилберте, но ни в коем случае не всю историю? Что, если остальная часть истории создается непрерывным потоком жизни? Ученые только сейчас начинают узнавать о нераспознанном до сих пор «втором генетическом коде», эпигеноме, который значительно увеличивает как сложность, так и уникальность информации в живых системах.
Эпигенетика — это изучение систем и процессов, с помощью которых экспрессия генов может быть изменена, не случайным образом, как в дарвинизме, а посредством конкретных, предсказуемых, повторяемых и исследуемых событий — а затем унаследованных в измененном состоянии. Исследования показали, что метилирование является ключевым механизмом. Описывается так:
Не все гены активны постоянно. Метилирование ДНК — один из нескольких эпигенетических механизмов, которые клетки используют для контроля экспрессии генов…. распространенный инструмент эпигенетической передачи сигналов, который клетки используют для блокировки генов в положении «выключено».В последние десятилетия исследователи многое узнали о метилировании ДНК, в том числе о том, как это происходит и где это происходит, а также обнаружили, что метилирование является важным компонентом многих клеточных процессов, включая эмбриональное развитие, геномный импринтинг, инактивацию Х-хромосомы. , и сохранение стабильности хромосом.
Эпигенетические изменения, такие как метилирование, не изменяют ДНК клетки, но они меняют то, как она функционирует. Например, в исследованиях на грызунах стресс может запускать серию химических реакций, которые определяют, насколько активными будут некоторые гены в передаче воздействия родительского стресса.
Такое наследование приобретенных черт постепенно получает признание в академической науке, но мы все еще находимся на ранней стадии понимания механизмов. Или, как выразился один исследователь, «… мы все чертовски увлечены выяснением того, как они работают». И еще один: «Хотя многие ломают голову над дырами в предлагаемом механизме, немногие предполагают, что лежащий в основе феномен — это сказка».
Оказывается, большинство генов не столько эгоистичны, сколько готовы тусоваться с не той компанией и усваивать дурные привычки.Вы так думаете, если прочитаете это в специальном выпуске Science :
.Молекулярное наследие выходит за рамки переноса генов и включает митохондрии и эпигенетику (Lane et al., P. 756). Наконец, процессы вынашивания и родов также формируют потомство, и преждевременные роды в настоящее время находятся в центре внимания исследований (Romero et al., P. 760).
Вот еще несколько результатов, которые дают некоторое представление о масштабах изменений, которые эпигенетика привносит в наше понимание истории жизни:
— Бактерии эпигенетически переписывают свою ДНК. Карл Циммер отмечает в Quanta :
Для студентов, изучающих историю биологии, этот вид наследственности перекликается с в значительной степени дискредитированной теорией, выдвинутой натуралистом Жаном-Батистом Ламарком в начале 19 века. Ламарк выступал за наследование приобретенных черт. … Появление генетики, казалось, разрушило эту идею. Похоже, что у опыта не было никакого способа изменить гены, которые организмы передавали своему потомству. Но CRISPR показал, что микробы переписывают свою ДНК информацией о своих врагах — информация, которую показал Баррангу, может иметь значение между жизнью и смертью их потомков.
Новые методы анализа помогут определить роль, которую бактериальная эпигенетика играет в создании устойчивости к антибиотикам.
— Лабораторные исследования показывают, как это работает на животных. Дело не только в окружающей среде матки, потому что вклад родителей-мужчин через изменения в сперме очень велик. Из Природа :
Самцы крыс, получавшие пищу с высоким содержанием жиров, например, рожали дочерей с аномальным метилированием ДНК в поджелудочной железе. Самцы мышей, получавших диету с низким содержанием белка, имеют потомство с измененной экспрессией генов холестерина в печени.А самцы мышей с предиабетом имеют аномальное метилирование сперматозоидов и передают повышенный риск диабета следующим двум поколениям.
Эффекты эпигенетического стресса, обнаруженные у крыс, могут сохраняться в течение нескольких поколений. Точно так же в недавнем исследовании на мышах, описанном в Nature , РНК сперматозоидов показала наследственные признаки травмы еще для двух поколений. А от голодающих мышей родились щенки, потомство которых грозило диабетом. В другом исследовании на мышах было обнаружено, что «вообще без каких-либо изменений в ДНК, метильные группы могут быть добавлены или вычтены, и эти изменения унаследованы так же, как мутация в гене.”
— Эпигенетика может влиять на поведение животных и в последующих поколениях. «Эпигенетическая наследственность между поколениями не является решаемой областью — она очень постоянно меняется», — признает один исследователь, но похоже, что эпигенетическая память пережитого стресса может передаваться из поколения в поколение и переходить от клетки к клетке на раннем этапе развития: «Это не меняет последовательность генов, а скорее то, как упакована ДНК и как гены экспрессируются ». Недавние исследования на грызунах показывают, что «воздействие наркотиков, алкоголя и стресса на родителей-животных может повлиять на развитие мозга и поведение их потомства.”
Например, из одного исследования о том, как мыши могут унаследовать страх перед запахом от своего родителя-самца, мы узнали:
Каким-то образом информация об устрашающем запахе проникает в семенники мыши и приводит к более низкому метилированию гена Olfr151 в ДНК сперматозоидов. Исследователи даже провели эксперименты с использованием экстракорпорального оплодотворения, чтобы убедиться, что отец каким-то образом не передал страх перед ацетофеноном через взаимодействие с матерью. Эпигенетическая подстройка сперматозоидов сохраняется в ДНК потомства, что приводит к усилению экспрессии рецептора в носу животных и, в конечном итоге, к повышенной чувствительности к запаху.
Конечно, предстоит еще много работы, и это открытие было поставлено под сомнение, как «слишком хорошее, чтобы быть правдой». Но авторы отметили, что они не тщательно отобрали данные; они включили в дополнительные материалы эксперименты, которые не дали статистически значимых результатов.
Если The Scientist не вешает авторов исследования эпигенетики на сушку, то, скорее всего, потому, что редакторы думают, что есть что сказать в пользу их модели. Это может помочь решить давние вопросы, например, о том, как животные, кажется, «знают» то, что они не узнали на собственном опыте.Сказать, что их знания — это «инстинкт», просто означает, что мы не знаем, как они это знают. Эпигенетические исследования, напротив, могут выявить конкретный механизм передачи информации.
— Эпигенетика также может влиять на здоровье человека. Унаследовать «хорошие гены» — это нормально (мы много слышали об этом от евгеников более века!). Но что, если пакет «хороших генов» будет поврежден при транспортировке и попадет туда? Гены могут передаваться по наследству в бодрствующем состоянии и могут влиять на метаболизм, поведение и предрасположенность к заболеваниям, включая болезни сердца и диабет.От одного исследователя:
По словам Исии, основная мысль заключается в следующем: «Я надеюсь, что люди понимают, что различные стрессы могут изменять экспрессию генов без изменения последовательности ДНК». Он говорит, что самые молодые из нас — развивающиеся эмбрионы и младенцы — могут быть особенно чувствительны к такого рода эпигенетическим изменениям, вызванным стрессом, и «нам следует более осторожно относиться к ним».
В другом исследовании травмированные беременные женщины, которым было около 9-11, не только имели значительно более низкие уровни кортизола в слюне, но и их дети, измеренные после рождения.Исследователи отметили, что эффект был наиболее очевиден, когда подвергшиеся воздействию дети находились в третьем триместре. Последующие исследования на крысах, подвергшихся стрессу, выявили различия в метилировании ДНК, способе химической модификации ДНК.
Аналогичным образом, проект «Ледяной шторм» обнаружил «отличительную« сигнатуру »в ДНК детей, рожденных после массивного ледяного шторма в Квебеке» (1998):
Тринадцать лет спустя исследователи обнаружили, что ДНК в Т-клетках — типе клеток иммунной системы — 36 детей демонстрирует отличительные паттерны метилирования ДНК.Исследователи впервые пришли к выводу, что материнские невзгоды предсказывают степень метилирования ДНК в Т-клетках. «Эпигенетическая» подпись играет роль в способе самовыражения генов. Это исследование также является первым, показывающим, что именно объективное воздействие стресса (например, дни без электричества), а не степень эмоционального стресса у беременных женщин, вызывает долгосрочные изменения в эпигеноме их младенцев.
Пока еще рано, но есть также некоторые свидетельства того, что бедность родителей может способствовать детскому ожирению.Приватизация может перепрограммировать ДНК в утробе матери, чтобы повышать или понижать определенные гены, связанные с аппетитом. Может быть, «всякий раз, когда вы находите еду, ешьте все, что можно»? Это может продолжаться и во взрослой жизни, когда дефицита действительно нет. Как объясняет один исследователь: «Если генетика — это алфавит, то эпигенетика — это написание, которое определяет активность наших клеток».
Итак, роль эпигенетики в ожирении в целом, некоторых иммунитетах, хронических заболеваниях и даже раке сейчас активно изучается. Что касается рака, некоторые исследователи обнаружили, что эпигенетическая регуляция необходима для обеспечения правильного количества хромосом в дочерних клетках после деления, и «опухолевые клетки часто имеют либо слишком мало, либо слишком много хромосом, что приводит к неправильной экспрессии ряда хромосом. гены.”
Многие из этих результатов могут нуждаться в модификации или замене, но мы больше не можем исключать возможность того, что изменения в диете прадедов и дедушек или в окружающей среде влияют на здоровье или продолжительность жизни последующих поколений. Эти изменения не являются дарвиновской генетической наследственностью; у родителя не было случайно мутировавшего «гена», скажем, ожирения, а скорее опыт (возможно, хронический голод), который изменил способ экспрессии генов, контролирующих аппетит. Изменения передаются, по крайней мере, через поколение или около того.Таким образом, эпигенетику не следует путать с генетическим или каким-либо другим детерминизмом; это не пророчество, а просто инструмент для оценки источников риска.
— Может ли эпигенетика влиять на поведение человека, а также на его здоровье? Возможно; например, жестокое обращение в раннем возрасте может изменить экспрессию генов у детей:
Исследователи обнаружили связь между родителями детей и конкретным геном (называемым геном рецептора глюкокортикоидов), который отвечает за важные аспекты социального функционирования и здоровья.
Если такие выводы подтвердятся, они могут пролить свет на «прерванные усыновления», когда спасенные дети продолжают вести себя так, как если бы они жили в среде с низким уровнем доверия / высокой угрозой. В любом случае, «рожденный плохим» просто больше не подходит для объяснения. Если эпигенетика дает достоверные сведения, мы, по крайней мере, можем надеяться на более реалистичные стратегии реабилитации.
По мере того как эпигенетика постепенно становится общепринятой в научной культуре, медицинский геномик Стефан Бек считает, что эпигеномная эпидемиология «находится на той же стадии, на которой геномная эпидемиология была восемь лет назад, когда большинство исследований были небольшими и редко выявляли одни и те же генетические варианты для какого-либо одного заболевания.Материалы семинара для конференции в Швеции в марте 2015 года «Эпигенетика как точка встречи между природой и воспитанием» предлагают проблему: «Существуют препятствия с точки зрения подозрения некоторых ученых в отношении биологических объяснений и упрощения социальных и философских вопросов. проблемы среди некоторых ученых. Две культуры должны быть соединены мостом, и мы предполагаем, что мост — это эпигенетика ».
Да, действительно. Рассмотрим спорный пересмотр дарвиновской теории рас в недавней книге научного писателя Николаса Уэйда « Беспокойное наследование». Эпигенетика делает такие теории неуместными, а также спорными. Вопрос не в том, проявляются ли определенные характеристики чаще в одной группе, чем в другой, а в том, каков механизм? Как сказал бы Майкл Бихи?
Эпигенетика должна привести к пересмотру учебников, говорят некоторые европейские исследователи, возможно, не подозревая о последователях Дарвина, которые разбили лагерь возле офисов органов образования, защищая свою финансируемую за счет налогов область. Тем не менее, освещение эпигенетики в самом последнем издании популярного университетского текста дарвиновского эволюциониста Кена Миллера увеличилось.В книге теперь есть определение эпигенетики со ссылкой на абзац по этой теме.
Некоторые, тем не менее, противостоят эпигенетике и кричат СТОП !! В конце концов, он очень новый и не приживается с легкостью. Цитата из «ключевой концепции» из The Scientist гласит: «Концепция клеточной памяти, передаваемая потомству, может быть не такой безумной, как кажется». В эпигенетике нет ничего сумасшедшего, если только человек не привержен генетическому фатализму, включающему «ген жира», «ген гея», «ген неверности», «ген насилия» и т. Д., которые якобы «заставляют» людей что-то делать.
«Прекратить шумиху вокруг эпигенетики и ламаркианской эволюции», Real Clear Science требует, отмечая, что «млекопитающие — совершенно разные звери» от червей и растений, где было продемонстрировано эпигенетическое наследование:
Поэтому очень скептически относитесь к исследованиям, в которых утверждается, что обнаружены эффекты для здоровья из-за эпигенетической наследственности. Шумиха может скоро утихнуть, и концепция ламаркистской эволюции может снова вернуться в могилу.
В основе этого всплеска может лежать философия, если мы обратимся к статье в Cell со ссылкой на RCS : «Поскольку геном человека был секвенирован, термин« эпигенетика »все чаще ассоциируется с надеждой на то, что мы больше, чем просто сумма наших генов ».
Это не «надежда», это факт. И обвинение в «шумихе» звучит странно, учитывая широко распространенную дарвиновскую шумиху вокруг всего, что связано с генами, приписывая «генам» все явления, от плохого вождения до удивления Вселенной.
Real Clear Science продолжает беспокоиться о том, что эпигеном угнетает женщин:
Эпигенетика — это следующая большая область, которую СМИ, подстрекатели и политические взломщики попытаются использовать. … Авторы обеспокоены, возможно, справедливо, что ажиотаж в СМИ вокруг эпигенетики снова обратится к матерям. Будет ли правительство снова регулировать то, что беременные женщины могут есть, пить и делать? И если да, то почему бы не регулировать и поведение мужчин? В конце концов, эпигенетика может повлиять на качество спермы.
Авторы, о которых идет речь, опубликовали статью в журнале Nature , в которой говорится: «Общество: не обвиняйте матерей — небрежное обсуждение эпигенетических исследований того, как ранняя жизнь влияет на здоровье поколений, может нанести вред женщинам». Это странное беспокойство, поскольку большинство матерей с радостью избежали бы передачи хронического заболевания, , если бы у них была правильная информация. И эпигенетика в основном предназначена для установления правильной информации.
Между тем, бывший главный лоббист Дарвина в американских школах сказал:
Было почти облегчением, когда один антиэволюционист заявил, что книги должны быть отвергнуты, потому что они не включают эпигенетику.По крайней мере, аргумент об эпигенетике появился сравнительно недавно (возможно, ему всего 5-8 лет). Согласно креационному мышлению, включение эпигенетики в учебники биологии ослабит эволюцию, потому что эпигенетика является свидетельством против эволюции.
Обратите внимание на любопытный термин «аргумент эпигенетики», как будто все согласны с тем, что единственный потенциал эпигенетики — это политическая стратегия.
Но жизнь продолжается … Некоторые спорят, действительно ли эпигенетика «ламаркистская». Помимо придирок, наш мир — не мир компакт-диска «This Is You». 1 Некоторые признанные эксперты даже говорят: «Эпигенетика может влиять на генетику» и что она «является одним из основных факторов межпоколенческих изменений, а не просто пассивным компонентом». Тем временем эпигеномика дебютирует благодаря десятилетию разработки программ анализа данных.
Итак, эпигенетика не собиралась уходить в могилу, а инвестировала в более крупное оборудование.
Многие результаты эпигенетики, как и многие находки в области горизонтального переноса генов, могут быть пересмотрены или заменены, но направление вряд ли изменится.Так как же эпигенетика участвует в эволюции? На этот счет есть два мнения. Говорят, что эпигеном развивается быстрее, чем геном, но большинство его изменений длится не более нескольких поколений. Возможно, это потому, что они перекрываются дальнейшими изменениями из-за новых условий: в своем новаторском исследовании исследователи пришли к выводу, что «эпигенетические изменения происходят на много порядков чаще, чем обычные мутации ДНК, но также часто недолговечны».
Однако эпигеномное разнообразие, согласно другим исследованиям, позволяет растениям приспосабливаться к различным условиям окружающей среды по всему миру.Изменения в среде обитания растений могут серьезно повлиять на эволюцию животных. Таким образом, это может сводиться к вопросу о том, каких изменений мы ищем. Что считается изменением?
В той мере, в какой эпигенетика ставит под сомнение упрощенные дарвиновские или другие схемы эволюции, ее главная роль может заключаться в «спойлере». Например, недавно шведские исследователи обнаружили, что эпигенетика, а не дарвиновская эволюция, является причиной широкого разнообразия типов домашних кур. Они говорят, что их результаты явно не согласуются с «традиционным дарвиновским объяснением», что интересно с учетом того факта, что собственное разведение голубей Дарвина часто используется как способ представления его теории общественности.
Итак, что мы можем теперь сказать об эволюции, истории жизни ?: Если мы не ищем чудесных преобразований, мы действительно наблюдаем некоторую эволюцию. Мы видели, что формы жизни могут продвигаться к одной и той же цели без общего предка (конвергентная эволюция). Совместное использование генов (горизонтальный перенос генов) может привести к появлению новых свойств у формы жизни. Эпигенетика может привести к тому, что потомство унаследует от своих родителей черты, которые были созданы при жизни их родителей, а не унаследованы от предыдущего общего предка.
Но возникает и другая возможность: может ли форма жизни эволюционировать, когда теряет вместо того, чтобы приобретать черты (деволюция)? Мы увидим.
Примечания:
(1) Цитируется у Дороти Нелкин, «Менее эгоистичен, чем священный? Гены и религиозный импульс в эволюционной психологии », в: Хилари Роуз и Стивен Роуз, ред., Увы, Бедный Дарвин: аргументы против эволюционной психологии (Лондон: Random House, Vintage, 2001), с. 18.
См. Остальную часть серии на сегодняшний день на «Поговорите с окаменелостями: давайте посмотрим, что они говорят в ответ.”
Изображение: Жан-Батист де Моне Шевалье де Ламарк, Шарль Тевенин [общественное достояние], через Wikimedia Commons.
Можем ли мы действительно унаследовать травму?
Одна рабочая теория основана на исследованиях на животных. В серии недавних исследований ученые из Медицинской школы Университета Мэриленда под руководством Трейси Бейл вырастили мышей-самцов в сложных условиях, периодически наклоняя клетки или оставляя свет включенным на ночь. Такое воспитание, по сути, травмирующее детство, меняет последующее поведение генов этих мышей таким образом, что меняет то, как они справляются с выбросами гормонов стресса.
И это изменение, в свою очередь, тесно связано с изменениями в том, как их потомство справляется со стрессом: а именно, молодые мыши оцепенели или менее реагируют на гормоны по сравнению с контрольными животными, — сказал доктор Бейл, директор университета. Центр эпигенетических исследований детского здоровья и развития мозга. «Это ясные и последовательные результаты», — сказала она. «Эта область значительно продвинулась вперед всего за последние пять лет».
Возможно, лучшее объяснение того, как такие следы травм могут быть прикреплены к сперматозоидам отца, дает Оливер Рандо из Массачусетского университета.Его исследования, в том числе на мышах, сосредоточились на придатке яичка, трубке возле яичек, где сперматозоиды загружаются перед эякуляцией. «Там они учатся плавать в течение нескольких дней, и их гены могут быть помечены», — сказал доктор Рандо.
Молекулы, которые влияют на изменения, по-видимому, являются «малыми РНК», фрагментами генетического материала, о котором ученые все еще изучают, сказал доктор Рандо.
«Эта трубка производит небольшие РНК и доставляет их к сперматозоидам по мере их развития, что позволяет предположить, что существует место, которое чувствует условия окружающей среды отца и может изменять упаковку РНК, доставляемых через сперму ребенку», — сказал доктор.- сказал Рандо. Он не делает никаких широких заявлений, кроме этого.