
Что такое таксис: Таксис (биология) — это… Что такое Таксис (биология)?
Таксисы — это… Что такое Таксисы?
таксисы — (от греч. taxis расположение по порядку) ориентирующие компоненты поведенческих актов, врожденные способы пространственной ориентации в сторону благоприятных (положительные Т.) или неблагоприятных (отрицательные Т.) условий среды. У растений… … Большая психологическая энциклопедия
Таксисы — (от греч. taxis расположение), двигательные реакции свободно передвигающихся организмов (животных, бактерий, некоторых грибов), а также некоторых клеток (сперматозоидов) свободно передвигающихся организмов в ответ на односторонне действующий… … Экологический словарь
ТАКСИСЫ — (от греч. taxis расположение) направленные движения организмов (простейшие животные и низшие растения), отдельных клеток или их органелл под влиянием односторонне действующего стимула света (фототаксис), температуры (термотаксис), химических… … Большой Энциклопедический словарь
ТАКСИСЫ
 taxis расположение), двигат. реакции в ответ на односторонне действующий стимул, свойственные свободно передвигающимся организмам (бактерии, нек рые грибы и водоросли, животные), нек рым клеткам и органоидам (споры и гаметы, обладающие… … Биологический энциклопедический словарь
taxis расположение), двигат. реакции в ответ на односторонне действующий стимул, свойственные свободно передвигающимся организмам (бактерии, нек рые грибы и водоросли, животные), нек рым клеткам и органоидам (споры и гаметы, обладающие… … Биологический энциклопедический словарьтаксисы — двигательные реакции подвижных микроорганизмов под влиянием одностороннего раздражения–светом (фототаксис), хим. веществами (хемотаксис), температурой (термотаксис) и др.; различают Т. положительные (движение к раздражителю) и Т. отрицательные… … Словарь микробиологии
таксисы — (от греч. táxis расположение), направленные движения организмов (простейшие животные и низшие растения), отдельных клеток или их органелл под влиянием односторонне действующего стимула света (фототаксис), температуры (термотаксис), химических… … Энциклопедический словарь
Таксисы — (от греч. taxis расположение) двигательные реакции свободно передвигающихся микроорганизмов и простейших растений, а также некоторых клеток многоклеточных организмов (зооспор, сперматозоидов, лейкоцитов) и отдельных частей клеток (ядер,… … Большая советская энциклопедия
ТАКСИСЫ — (от греч. taxis расположение), направленные движения организмов (простейшие ж ные и низшие р ния), отд. клеток или их органелл под влиянием односторонне действующего стимула света (фототаксис), темп ры (термотаксис), хим. в в (хемотаксис), в т.ч … Естествознание. Энциклопедический словарь
taxis расположение), направленные движения организмов (простейшие ж ные и низшие р ния), отд. клеток или их органелл под влиянием односторонне действующего стимула света (фототаксис), темп ры (термотаксис), хим. в в (хемотаксис), в т.ч … Естествознание. Энциклопедический словарь
таксисы — (гр. taxis расположение по порядку) биол. двигательные реакции свободно передвигающихся низших растений и животных, а также отдельных клеток (зооспоры, сперматозоиды, лейкоциты) и частей клеток (ядра, пластиды) под влиянием одностороннего… … Словарь иностранных слов русского языка
ТАКСИСЫ — (от греч. táxis расположение), направленное движение бактерий, слизистых грибов (миксомицетов), одноклеточных водорослей и животных, а также отдельных клеток (лейкоцитов, сперматозоидов) многоклеточных организмов и органелл (ядер, пластид) … Ветеринарный энциклопедический словарь
Таксис
Та́ксис
(от греч. τάξις — построение, порядок, расположение) — языковая категория, характеризующая временны́е
отношения между действиями (в широком смысле, включая любые
разновидности предикатов):
одновременность/неодновременность, прерывание, соотношение главного и
сопутствующего действия и т. п. Таксис включает аспектуальную
(видовую) характеристику комплекса соотносимых во времени действий
и может взаимодействовать с причинно-следственными,
уступительно-противительными и другими значениями.
τάξις — построение, порядок, расположение) — языковая категория, характеризующая временны́е
отношения между действиями (в широком смысле, включая любые
разновидности предикатов):
одновременность/неодновременность, прерывание, соотношение главного и
сопутствующего действия и т. п. Таксис включает аспектуальную
(видовую) характеристику комплекса соотносимых во времени действий
и может взаимодействовать с причинно-следственными,
уступительно-противительными и другими значениями.
Понятие таксиса в его отличиях от понятия времени (см. Время) было определено Р. О. Якобсоном: таксис
характеризует сообщаемый факт по отношению к другому сообщаемому
факту и безотносительно к факту сообщения (в отличие от времени,
характеризующего сообщаемый факт по отношению к факту сообщения). Если
время относится к числу категорий («шифтеров»), содержащих указания на
отношение сообщаемого факта и/или его участников к факту сообщения либо
к его участникам (ср. категории лица,
наклонения,
засвидетельствованности), то таксис представляет собой одну из
категорий («не-шифтеров»), не содержащих этих указаний (ср. категории
рода, числа, залога).
Якобсоном был введён и сам термин «таксис» как «греческий прообраз»
предложенного Л. Блумфилдом термина «порядок» (order). Вместе с тем широко известны другие термины:
«относительное время», «временна́я соотнесённость» (А. И. Смирницкий) и
др. Э. Кошмидер писал о «временном отношении данного факта к другому в
комплексе фактов, называемом ситуацией», и анализировал такие
«ситуационные типы», как «что-то происходило, и тем временем произошло
что-то другое», «когда что-то происходило, что-то произошло» и
т. п.
категории
рода, числа, залога).
Якобсоном был введён и сам термин «таксис» как «греческий прообраз»
предложенного Л. Блумфилдом термина «порядок» (order). Вместе с тем широко известны другие термины:
«относительное время», «временна́я соотнесённость» (А. И. Смирницкий) и
др. Э. Кошмидер писал о «временном отношении данного факта к другому в
комплексе фактов, называемом ситуацией», и анализировал такие
«ситуационные типы», как «что-то происходило, и тем временем произошло
что-то другое», «когда что-то происходило, что-то произошло» и
т. п.
Таксис может быть выражен различными (морфологическими, синтаксическими, морфолого-синтаксическими, лексическими) средствами, которые образуют функционально-семантическое поле,
пересекающееся с полями темпоральности и аспектуальности. Таксис
существует в каждом языке, однако как особая грамматическая категория, отличающаяся от вида и времени, — лишь в тех языках, где имеется
соответствующая специальная система грамматических форм. Якобсон, говоря о таксисе как
грамматической категории, приводит в качестве примера факты нивхского языка, а также (со ссылкой на
Б. Л. Уорфа) языка хопи; в числе «грамматических понятий, выражаемых
русскими глагольными формами», он рассматривает выражаемый деепричастием зависимый таксис, указывающий на
сообщаемый факт, сопутствующий другому, главному сообщаемому факту.
Как отмечает Ю. С. Маслов, во многих языках таксис не выступает в
качестве особой грамматической категории, а объединяется в рамках одной
комбинированной категории либо со временем, либо с видом.
Якобсон, говоря о таксисе как
грамматической категории, приводит в качестве примера факты нивхского языка, а также (со ссылкой на
Б. Л. Уорфа) языка хопи; в числе «грамматических понятий, выражаемых
русскими глагольными формами», он рассматривает выражаемый деепричастием зависимый таксис, указывающий на
сообщаемый факт, сопутствующий другому, главному сообщаемому факту.
Как отмечает Ю. С. Маслов, во многих языках таксис не выступает в
качестве особой грамматической категории, а объединяется в рамках одной
комбинированной категории либо со временем, либо с видом.
Некоторые разновидности таксиса в ряде языков (французском, английском, немецком, болгарском и др.)
находят выражение в формах относительных времён, в частности со
значением предшествования (в формах плюсквамперфекта, будущего
предварительного, в одной из функций форм перфекта). В подобных формах на передний план
могут выступать аспектуальные функции, исторически предшествовавшие
выражению хронологических отношений между действиями.
При исследовании таксиса как функционально-семантического поля важно определить соотношение и взаимосвязи его центральных и периферийных компонентов. Так, в русском языке таксис выступает как функционально-семантическое поле с полицентрической структурой; в сфере зависимого таксиса грамматическим центром являются конструкции с деепричастием совершенного и несовершенного вида, а в области независимого таксиса — сочетания видо-временных форм: а) в предложениях с придаточным времени, б) в предложениях с однородными сказуемыми и в сложносочинённых предложениях (при возможном взаимодействии с лексическими элементами типа «сначала», «а затем» и т. п.).
- Смирницкий А. И., Морфология английского языка, М., 1959;
- Кошмидер Э., Очерк науки о видах польского глагола. Опыт синтеза, [пер. с польск.], в кн.: Вопросы глагольного вида, М., 1962;
- Якобсон Р. О., Шифтеры, глагольные категории и русский
глагол, пер. с англ.
 , в кн.: Принципы типологического анализа языков
различного строя, М., 1972;
, в кн.: Принципы типологического анализа языков
различного строя, М., 1972; - Маслов Ю. С., К основаниям сопоставительной аспектологии, в кн.: Вопросы сопоставительной аспектологии, в. 1, Л., 1978;
- Бондарко А. В., Функциональная грамматика, Л., 1984;
- Слюсарева Н. А., Проблемы функциональной морфологии современного английского языка, М., 1986;
- Теория функциональной грамматики. Аспектуальность. Временная локализованность. Таксис, Л., 1987.
А. В. Бондарко.
Таксис
Таксисы иногда используют в борьбе с вредными насекомыми. Например, используется отрицательный хемотаксис платяной моли (Tineola biselliella Hümm.) для защиты мехов, шерстяных тканей, ковров путем пересыпания их нафталином или используется положи-тельный фототаксис путем лова различных вредных насекомых на свет лампы для определения наилучших сроков борьбы с ними и т. д.[ …]
Таксисы объясняют нам многие явления в жизни насекомых. Капустная моль (Р lutetia maculipennis Curt.), например, откладывает яйца исключительно на крестоцветные растения потому, что ею руководит в данном случае хемотаксис — раздражение воспринимается через органы обоняния. Хлопковая совка (Chlori-dea obsoleta F.) заселяет хлопковые посевы только со времени бутонизации, так как железистые волоски на листьях и стеблях хлопчатника, выделяющие щавелевую и муравьиную кислоты, до этого времени не функционируют, здесь решающее значение имеет также хемотаксис.[ …]
Капустная моль (Р lutetia maculipennis Curt.), например, откладывает яйца исключительно на крестоцветные растения потому, что ею руководит в данном случае хемотаксис — раздражение воспринимается через органы обоняния. Хлопковая совка (Chlori-dea obsoleta F.) заселяет хлопковые посевы только со времени бутонизации, так как железистые волоски на листьях и стеблях хлопчатника, выделяющие щавелевую и муравьиную кислоты, до этого времени не функционируют, здесь решающее значение имеет также хемотаксис.[ …]
Таксисы простые перемещения животных, связанные с избеганием неблагоприятных факторов и поиском оптимальных условий.[ …]
ТАКСИСЫ [от гр. taxis — расположение] — двигательные реакции свободно передвигающихся организмов (животные, бактерии), а также некоторых клеток (сперматозоидов) в ответ на односторонне действующий фактор. Источником раздражения может быть свет (фототаксис), влага (гидротаксис), температура (термотаксис), химические вещества (хемотаксис) и др.[ …]
Термином таксис в настоящее время обозначают движения в ответ на раздражения, которые так легко наблюдать у низших животных (фиг. 120). Френкель и Гунн (1940), давшие первый обзор этих основных форм поведения, различают: 1) кинезис — ненаправленные реакции, такие, как общее избегание неблагоприятных условий; 2) таксисы в узком смысле — направленные реакции, ориентированные прямо к раздражителю или в противоположную сторону (мотылек, летящий к свету), и 3) поперечную ориентацию,- или движение под некоторым углом к направлению стимула, такую, как хорошо известная ориентация по световому компасу и нахождение направления у пчел (см. фон Фриш, 1955). Нельзя провести четкой и твердой границы между таксисами и рефлексами, но рефлексы рассматриваются в общем как реакции специфических органов или частей тела в ответ на раздражение; и те и другие могут изменяться в результате приобретенного опыта. Хорошо известный условный рефлекс служит началом приобретенной в результате обучения поведенческой реакции. Когда-то считали, что ее можно выработать только у высших животных, но, по-видимому, ее можно выработать и у низших червей и даже у простейших.
120). Френкель и Гунн (1940), давшие первый обзор этих основных форм поведения, различают: 1) кинезис — ненаправленные реакции, такие, как общее избегание неблагоприятных условий; 2) таксисы в узком смысле — направленные реакции, ориентированные прямо к раздражителю или в противоположную сторону (мотылек, летящий к свету), и 3) поперечную ориентацию,- или движение под некоторым углом к направлению стимула, такую, как хорошо известная ориентация по световому компасу и нахождение направления у пчел (см. фон Фриш, 1955). Нельзя провести четкой и твердой границы между таксисами и рефлексами, но рефлексы рассматриваются в общем как реакции специфических органов или частей тела в ответ на раздражение; и те и другие могут изменяться в результате приобретенного опыта. Хорошо известный условный рефлекс служит началом приобретенной в результате обучения поведенческой реакции. Когда-то считали, что ее можно выработать только у высших животных, но, по-видимому, ее можно выработать и у низших червей и даже у простейших.
Значение таксисов для этих грибов, однако, не очень велико. Без помощи токов воды зооспоры могут пройти лишь очень ограниченное расстояние. В увлажненной почве, помещенной в чашки Петри, зооспоры Phytophthora cinnamomi достигали кончиков корней, удаленных от них не больше 7 см.[ …]
ХЕМОТАКСИС — см. в ст. Таксисы. ХИМИЧЕСКАЯ ДЕГРАДАЦИЯ ПОЧВЫ — см. в ст. Деградация почвы. ХИМИЧЕСКАЯ ЗАЩИТА РАСТЕНИЙ — система мероприятий по борьбе с вредителями, сорняками и болезнями растений или предотвращению их распространения путем обработки растений ядохимикатами. См. Пестициды.[ …]
ТОНОТАКСИС [от гр. tonos — напряжение, давление и таксис] — направленное движение организмов как ответная реакция на изменение давления среды.[ …]
Недавно Торнли предложил математическую модель фплло-таксиса, основные положения которой близки постулированным в Теории подавления. В основе модели лежит предположение о том, что каждый листовой примордий является местом образования «морфогена» М. Интенсивность действия морфо-гена зависит от возраста, размера и расстояния его источника от апекса. Предполагается, что морфоген должен диффузно распространяться от зачатков листьев и интенсивность его действия понижается со скоростью, пропорциональной его концентрации. Согласно этой модели, заложение нового листового примордия может происходить только в том месте, где концентрация М падает до определенного критического значения. Это положение идентично положению теории подавления. Математическая модель, основанная на этих исходных постулатах, предполагает, что при определенных условиях угол между соседними примордиями .будет приближаться к наблюдаемому значению 137,5°. Таким образом, данный подход также показывает, что расположение листовых примордиев на апексе побега может быть предсказано на основе довольно простых исходных допущений.[ …]
Интенсивность действия морфо-гена зависит от возраста, размера и расстояния его источника от апекса. Предполагается, что морфоген должен диффузно распространяться от зачатков листьев и интенсивность его действия понижается со скоростью, пропорциональной его концентрации. Согласно этой модели, заложение нового листового примордия может происходить только в том месте, где концентрация М падает до определенного критического значения. Это положение идентично положению теории подавления. Математическая модель, основанная на этих исходных постулатах, предполагает, что при определенных условиях угол между соседними примордиями .будет приближаться к наблюдаемому значению 137,5°. Таким образом, данный подход также показывает, что расположение листовых примордиев на апексе побега может быть предсказано на основе довольно простых исходных допущений.[ …]
У организмов, лишенных нервной системы (простейшие и растения), раздражимость проявляется в виде тропизмов, таксисов и настий. У организмов, имеющих нервную систему, раздражимость проявляется в виде рефлекторной деятельности. У животных восприятие внешнего мира осуществляется через первую сигнальную систему, тогда как у человека в процессе исторического развития сформировалась еще и вторая сигнальная система. Благодаря раздражимости организмы уравновешиваются со средой. Избирательно реагируя на факторы среды, организмы «уточняют» свои отношения со средой, в результате чего возникает единство среды и организма.[ …]
У животных восприятие внешнего мира осуществляется через первую сигнальную систему, тогда как у человека в процессе исторического развития сформировалась еще и вторая сигнальная система. Благодаря раздражимости организмы уравновешиваются со средой. Избирательно реагируя на факторы среды, организмы «уточняют» свои отношения со средой, в результате чего возникает единство среды и организма.[ …]
Зачатки водорослей в виде спор, гамет и зигот не совсем стихийно разносятся водой. Они обладают различного рода таксисами, определяющими направление их движения в зависимости от внешних раздражителей: света (ф о т о т а к с и с), температуры (термотаксис), химических веществ, содержащихся в воде (хемотаксис). Способностью двигаться обладают не только зооспоры, но и споры, лишенные жгутиков. У них наблюдается амебоидное движение, при котором сначала образуется выступ, а затем в него перемещается содержимое всей споры.[ …]
Поведение насекомых определяется внешними раздражениями и проявляется во врожденных, или безусловных, рефлексах, таксисах и условных рефлексах. [ …]
[ …]
Простые рефлексы представляют собой автоматическую реакцию на внешние раздражения. К более сложным рефлексам относятся таксисы — определенные движения насекомых по отношению раздражителя. Раздражителями являются свет, тепло, запах и другие факторы среды. Таксисы бывают положительными и отрицательными, в зависимости от движения насекомого к источнику раздражения или удаления от него.[ …]
АЭРОТАКСИС — движение аэробных микроорганизмов к источнику кислорода (положительный А.) или анаэробных — от него (отрицательный А.). См. также Таксисы. АЭРОТЕНК, аэротанк [от гр. аёг — воздух и англ. tank резервуар, бак] — искусственное сооружение для биологической очистки сточных вод путем окисления их микроорганизмами, представляющее собой несколько проточных резервуаров, продуваемых воздухом.[ …]
Человек применяет также различного рода механические и физико-механические мероприятия для борьбы с вредными насекомыми, причем учитывает и таксисы насекомых. Используются, например, светоловушкн для вылавливания вредных насекомых; пар, нагретая вода и сухой жар для борьбы с вредителями закрытого грунта (парников, оранжерей) и продуктов, хранимых на складах; для защиты посевов от нелетающих вредителей (жуков кравчиков — Lethrus sp. sp., не летающего при температуре ниже 18° С свекловичного долгоносика — Bothynoderes punctiventris Germ, и др.) вырывают заградительные канавы; производится вылавливание гусениц бабочек, прыгающих жуков, малоподвижных клопов путем применения различных механических уловителей и т. д. Некоторые вредители садов—гусеницы яблочной плодожорки (Carpo-capsa pomonella L.), чехликовые моли (Coleophora sp. sp.) и др. привлекаются для уничтожения накладываемыми на стволы деревьев ловчими поясами из мешковины или соломы, в которые они забираются для-окукливания. Разрабатываются методы использования: ультракоротких электроволн для очистки от амбарных и почвенных вредителей.[ …]
sp., не летающего при температуре ниже 18° С свекловичного долгоносика — Bothynoderes punctiventris Germ, и др.) вырывают заградительные канавы; производится вылавливание гусениц бабочек, прыгающих жуков, малоподвижных клопов путем применения различных механических уловителей и т. д. Некоторые вредители садов—гусеницы яблочной плодожорки (Carpo-capsa pomonella L.), чехликовые моли (Coleophora sp. sp.) и др. привлекаются для уничтожения накладываемыми на стволы деревьев ловчими поясами из мешковины или соломы, в которые они забираются для-окукливания. Разрабатываются методы использования: ультракоротких электроволн для очистки от амбарных и почвенных вредителей.[ …]
Атротаксис (А1Ьго1ах1з) — единственный род таксодиевых, распространенный в южном полушарии. Два ныне живущих вида растут в горных районах Западной Тасмании на высоте 900—1300 м. Ископаемые остатки атро-таксиса изредка находят в третичных отложениях Тасмании и Австралии.[ …]
Помимо инстинкта, имеются и более примитивные врожденные формы поведения, не имеющие характерной для инстинкта структуры, например, кинезис (изменение скорости хаотического движения в ответ на стимул), таксис (направленное движение по направлению к стимулу или от него), рефлексы типа коленного и др. Их роль в человеческом социальном поведении, вероятно, незначительна. Более того, противники биополитического подхода как раз и пытались сводить вклад биологического элемента в поведение человеческого индивида к коленному рефлексу или отдергиванию руки от раскаленного предмета. Как мы уже видели, фактически вклад биологии существенно более весом, и о нем будет идти речь в дальнейшем.[ …]
Их роль в человеческом социальном поведении, вероятно, незначительна. Более того, противники биополитического подхода как раз и пытались сводить вклад биологического элемента в поведение человеческого индивида к коленному рефлексу или отдергиванию руки от раскаленного предмета. Как мы уже видели, фактически вклад биологии существенно более весом, и о нем будет идти речь в дальнейшем.[ …]
Пусть теперь особи в популяции, кроме случайного блуждания, перемешаются по ареалу в определенном направлении со скоростью IV. Это может быть, например, направление увеличения концентрации пищи или направление уменьшения концентрации какого-либо вредного вешества. Такие явления в биологии называются таксисом. Возможна и другая интерпретация. Например, если это свободно плавающие водные организмы, то, кроме собственных случайных блужданий, они могут переноситься течениями. В этом случае М> — скорость течения.[ …]
Ориентация вытянутых в длину тел в потоке жидкости, как известно, обусловлена действием на них градиента скорости движущейся жидкости, возникающего в результате уменьшения скорости течения от центра к периферии потока.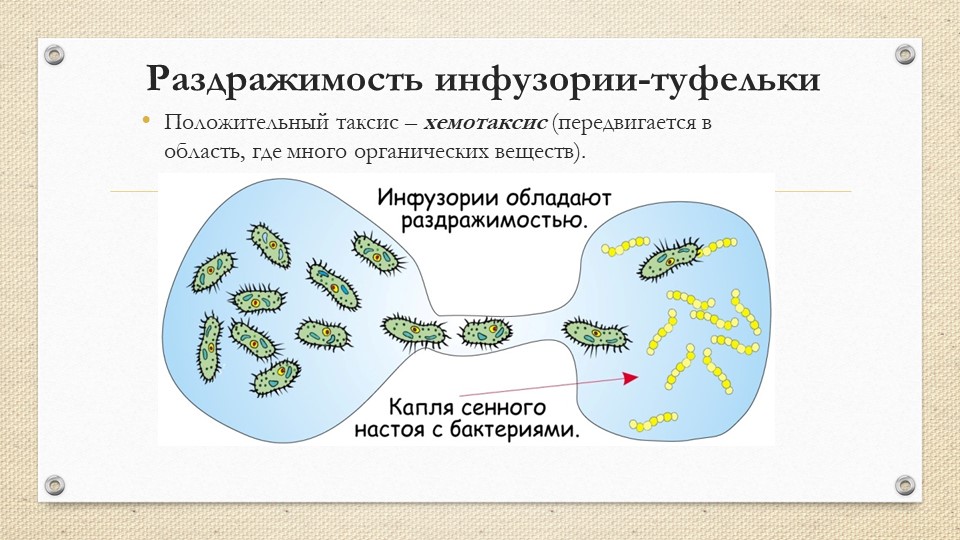 С этой позиции легко назвать причины, которые заставляют спермий ориентироваться в потоке жидкости своей продольной осью в направлении течения. Однако исследования таксисов у спермиев позволяют в ряде случаев допустить наличие механизма ориентации и движения в пространстве, которое нельзя объяснить лишь действием элементарных законов физики.[ …]
С этой позиции легко назвать причины, которые заставляют спермий ориентироваться в потоке жидкости своей продольной осью в направлении течения. Однако исследования таксисов у спермиев позволяют в ряде случаев допустить наличие механизма ориентации и движения в пространстве, которое нельзя объяснить лишь действием элементарных законов физики.[ …]
ТРОПИЗМЫ [от гр. tropos — поворот, направление] — направление ростовых движений (изгибы) органов растений, вызванные односторонним действием к.-л. фактора: света (фототропизм), земного притяжения (геотропизм), химических веществ (хемотропизм), температуры (термотропизм), воды (гидротропизм) и др. В основе Т. лежит неравномерный рост, вызываемый перераспределением в растении фитогормонов.[ …]
Поведение в самом широком смысле — это некие явные действия, предпринимаемые организмом для того, чтобы приспособиться к внешним условиям и обеспечить себе выживание. Кроме того, поведение — это важное средство, позволяющее особи войти в качестве составной части в организованные и регулируемые группы и сообщества. Поведение слагается из шести компонентов, относительное значение которых варьирует у разных организмов: 1) тропизмы; 2) таксисы; 3) рефлгксы; 4) инстинкты; 5) обучение; 6) интеллект. Вначале этологи1 старались четко различать «врожденное» (компоненты 1—4) и «приобретенное» поведение (компоненты б и 6), но теперь выяснилось, что так называемое «обучение» происходит на основе сложного комплекса рефлексов, инстинктов и других типов врожденного поведения, включая циркадные и другие врожденные ритмы, обсуждавшиеся в предыдущем разделе.[ …]
Поведение слагается из шести компонентов, относительное значение которых варьирует у разных организмов: 1) тропизмы; 2) таксисы; 3) рефлгксы; 4) инстинкты; 5) обучение; 6) интеллект. Вначале этологи1 старались четко различать «врожденное» (компоненты 1—4) и «приобретенное» поведение (компоненты б и 6), но теперь выяснилось, что так называемое «обучение» происходит на основе сложного комплекса рефлексов, инстинктов и других типов врожденного поведения, включая циркадные и другие врожденные ритмы, обсуждавшиеся в предыдущем разделе.[ …]
ТЕЛОМОРФОЗ [от гр. telos — конец, цель и morphe — форма, вид] — особое направление эволюции, означающее образование узкоспециализированных форм, приспособленных к специфическим условиям существования в узкой адаптивной зоне (напр., многие паразиты, глубоководные рыбы, обитатели подземных вод, ленивцы и др.). См. Стенобионты. ТЕЛОТАКСИС [от гр. telos — конец, цель и taxis расположение] — направленное перемещение животных к источнику стимуляции. См. Таксисы.[ …]
Таксисы.[ …]
Поведенческие (этологические) адаптации проявляются в самых разнообразных формах. Например, существуют формы приспособительного поведения животных, направленные на обеспечение оптимального теплообмена с окружающей средой. Приспособительное поведение может проявляться в создании убежищ, передвижениях в направлении более благоприятных, предпочитаемых температурных условий, выборе мест с оптимальной влажностью или освещенностью. Многим беспозвоночным свойственно избирательное отношение к свету, проявляющееся в приближениях или удалениях от источника (таксисах). Известны суточные и сезонные кочевки млекопитающих и птиц, включая миграции и перелеты, а также межконтинентальные перемещения рыб.[ …]
У насекомых особенно развиты термотаксис, фототаксис, гидротаксис, хемо- и трофотаксисы. Термотаксис вызывается действием температуры. Фототаксис вызывается светом. Положительный фототаксис проявляется у многих ночных бабочек и других насекомых, летящих на свет; у ряда других насекомых, например у яблонной плодожорки, проявляется отрицательный фототаксис, и они на свет не летят. Хемотаксис обусловлен действием химических веществ. Например, бабочки капустной белянки привлекаются запахом горчичных масел крестоцветных растений для откладки на них яиц. Изучение таксисов позволяет использовать их для борьбы с вредными насекомыми.[ …]
Хемотаксис обусловлен действием химических веществ. Например, бабочки капустной белянки привлекаются запахом горчичных масел крестоцветных растений для откладки на них яиц. Изучение таксисов позволяет использовать их для борьбы с вредными насекомыми.[ …]
Тропизмы и таксисы — это двигательные реакции организмов или их структур
Одним из главных свойств живого организма является движение или ответ на раздражающий фактор. У развитых организмов движением называется мышечный акт, реализация которого достигается за счет влияния нервного импульса на мышцу. Однако у элементарных организмов движение и ответ на раздражение приобретают несколько иную форму. В общем виде эти явления объединены в понятие «таксисы». Это двигательная реакция организма, его части или отдельной органеллы в направлении раздражителя или от него. У растений аналогичную трактовку имеет термин «тропизм». Таксисы и тропизмы могут быть положительными и отрицательными.
Источники раздражения
Источники раздражения, способные провоцировать таксисы — это факторы живой и неживой природы. Любые физические явления, биологические факторы или химические вещества способны вызывать движение организма, если от них зависит его жизнедеятельность. К примеру, хемотаксис — направленное движение к месту расположения химического вещества. Если клетка движется к той молекуле, которая имеет ценность как метаболический субстрат, то такой хемотаксис является положительным. Отрицательный хемотаксис — это намеренное увеличение расстояния между химическим веществом и клеткой. Примером положительного хемотаксиса является движение лейкоцита к месту воспаления.
Любые физические явления, биологические факторы или химические вещества способны вызывать движение организма, если от них зависит его жизнедеятельность. К примеру, хемотаксис — направленное движение к месту расположения химического вещества. Если клетка движется к той молекуле, которая имеет ценность как метаболический субстрат, то такой хемотаксис является положительным. Отрицательный хемотаксис — это намеренное увеличение расстояния между химическим веществом и клеткой. Примером положительного хемотаксиса является движение лейкоцита к месту воспаления.
Отрицательные химические таксисы — это активное бегство клеток или попытка отграничения от них, если вещества способны привести к их гибели. Также источником раздражения является электромагнитное излучение с различными длинами волн, жидкость, почва и другие факторы. В каждом случае таксис может быть положительным, то есть организм, его часть или отдельный его органоид, приближаются к раздражителю, или отрицательным. Отрицательные таксисы — это намеренное увеличение расстояния между организмом и раздражающим фактором.
Тропизм и таксис
Тропизм является частным примером таксиса у растений. Они имеют множество ориентиров, по отношению к которым двигаются в течение жизни или суточных циклов. К примеру, верхушки почти всех фотосинтезирующих растений обладают отрицательным геотропизмом и положительным гелиотропизмом. Это значит, что они стремятся достичь солнца с целью увеличения эффективности фотосинтеза. Растения также обладают положительным гидротропизмом, отрицательным термотропизмом.
Специфические тропизмы и таксисы
Разобравшись, что такое таксис в биологии, определение конкретных раздражителей для некоторых организмов позволяет понимать особенности их метаболизма. В частности, положительным термотропизмом обладают организмы, метаболизм которых должен протекать при высоких температурах. Также существует магнитотаксис, анемотаксис (движение в направлении воздуха), баротаксис, цитотаксис, реотаксис (в зависимости от течения в водоемах), гальванотаксис (по отношению к электрическому току). При этом таксис — это фундаментальный тип поведения одноклеточных или многоклеточных организмов. Только по отношению к ориентиру, в качестве которого выступает любой указанный выше фактор, организмы способны двигаться в живой природе.
При этом таксис — это фундаментальный тип поведения одноклеточных или многоклеточных организмов. Только по отношению к ориентиру, в качестве которого выступает любой указанный выше фактор, организмы способны двигаться в живой природе.
Таксисы и тропизмы / Словарь
Термины, нередко ошибочно трактуемые как синонимы, следует противопоставить друг другу как алгоритмы наведения на цель и способы ориентации вне зависимости от природы ориентирующего стимула (таксисы) и как собственные отношения к этим стимулам, связанные с внутренней мотивацией их выбора (тропизмы).
В соответствии с набором стимулов и отношений к ним насекомых различают положительные и отрицательные фото-, гео-, хемо-, гигро-, термотропизмы и тому подобное, подразумевая положительные и отрицательные реакции на гравитацию, источники света, химические стимулы, влажность и тепло. Механизм формирования тропизмов до сих пор остаётся неясным, но его биологический смысл часто вполне очевиден. Что касается таксисов, то наиболее явные из них — фобо-, клино-, тропо-, тело- и менотаксис.
Фоботаксис, иногда называемый методом проб и ошибок, представляет собой относительно примитивный, но наиболее общий алгоритм поведения в экстремальных условиях, угрожающих самому существованию организма. Он проявляется в замедлении или ускорении движения под влиянием стимула и в частоте изменения направления движения. Например, при повышенной температуре многие насекомые начинают быстро бегать, постоянно меняя направление движения; при этом вероятность их выхода за пределы зоны опасности возрастает. На основе фоботаксиса можно объяснить формирование скоплений насекомых в местах с наиболее благоприятными для них режимами влажности, освещённости и температуры. Клинотаксис определяет целенаправленность движения насекомого при последовательном выборе уровней возбуждения соответствующих рецепторов.
Например, личинки комнатной мухи, отличающиеся отрицательным фототропизмом, раскачивают переднюю часть тела с расположенными на ней фото-рецепторами, избирая направление движения, при котором рецепторы возбуждаются минимально. Тропотаксис представляет собой более совершенный алгоритм наведения на источник стимула, основанный на достижении равновесия в возбуждении симметричных рецепторов. Например, медоносная пчела, расставив в стороны антенны, легко воспринимает пахучую приманку, расположенную в одном из колен V-образной камеры.
При нарушении симметрии рецепторов, например закрашивая один глаз, мы вынуждаем насекомое к постоянным манежным (круговым) движениям даже в параллельных лучах света. Телотаксис предполагает наведение на стимул за счёт того, что его воспринимают рецепторы, направленные прямо вперёд. При этом даже ослеплённое на один глаз насекомое сохраняет верную ориентацию, не совершая манежных движений. Менотаксис, наводя на цель, предполагает фиксацию некоторого угла между направлением движения и ориентирующим стимулом. При этом избранные ориентиры могут быть несущественными для насекомого, но всегда в строго фиксированном положении по отношению к истинной цели. В частности, бабочки, привлечённые светом, демонстрируют менотаксис, который сравним со способами ориентации по азимуту.
См. Поведение.
Все термины на букву «Т»
Что такое Таксисы? Таксисы – это… Значение в биологическом словаре
Что значит «Таксисы»? Читайте ниже определение «Таксисы».
Что такое «Таксисы»? «Таксисы» – это:
(от греч. taxis — расположение), двигат. реакции в ответ на односторонне действующий стимул, свойственные свободно передвигающимся организмам (бактерии, нек-рые грибы и водоросли, животные), нек-рым клеткам и органоидам (споры и гаметы, обладающие жгутиками, лейкоциты, хлоропласты). Источниками раздражения могут быть свет (фототаксис), темп-ра (термотаксис), влага (гидротаксис), хим. в-ва (хемотаксис) и др. Движение может быть направлено к источнику стимуляции (положительный Т.) или от него (отрицательный Т.). В микробиологии и ботанике по характеру реагирования на раздражитель различают фоботаксисы — ненаправленные изменения курса движения (бактерии, зооспоры нек-рых фикомицетов) и топотаксисы — направленные перемещения по отношению к источнику раздражения (сперматозоиды мхов, папоротников). Жгутиковым водорослям свойственны оба типа Т. У животных Т. наз. Только направленные по отношению к стимулу перемещения (ненаправленные движения наз. кинезами), напр. реакция личинок мясной мухи на свет, бросок богомола на жертву, ориентация по солнцу у пчёл, муравьев, перелётных птиц, мигрирующих бабочек, рыб.
См. тропизмы.
Источник: Биологический энциклопедический словарь.
Другие значения
Определение термина в других источниках:
- Микробиология. Словарь терминов / Таксисы – это
Двигательные реакции подвижных микроорганизмов под влиянием одностороннего раздражения–светом (фототаксис), хим. веществами (хемотаксис), температурой (термотаксис) и др.; различают Т. положительные (движение к раздражителю) и Т. отрицательные (движение от раздражителя).
- Ботаника. Словарь терминов / Таксисы – это
ТАКСИСЫ — двигательные реакции самостоятельно передвигающихся микроорганизмов и простейших растений, а также некоторых клеток многоклеточных организмов (зооспор, сперматозоидов) и отдельных частей и органоидов клеток (ядер, пластид) в ответ на односторонне действующий фактор. В зависимости от природы раздражителя … далее…
См. также что такое
«Ситимобил» поменял схему работы с водителями такси
Сервис заказа такси «Ситимобил» 19 мая перешел на новую схему работы с водителями, следует из информации на странице таксопарка Gdel в Instagram. Представитель «Ситимобила» Георгий Лобушкин подтвердил, что агрегатор изменил схему работы с водителями.
Теперь в приложении сервиса для водителей есть только два режима. В режиме «не работаю» водители получают только предварительные заказы. А при выборе «на линии» «Ситимобил» сам назначает заказы с правом отказаться от них, в том числе если конечная точка маршрута находится за городом. Конечную точку водители видят только после поступления заказа и при условии достаточно большого количества баллов в приложении сервиса. Цена поездки теперь скрыта и отображается только после принятия заказа. До этого у «Ситимобила» был еще один режим – «предлагать». В нем водитель самостоятельно мог выбрать понравившийся ему заказ и сразу видел как точку назначения маршрута, так и цену поездки.
«Ситимобил» – второй после «Яндекс.Такси» по доле рынка агрегатор такси в Москве. Компания входит в совместное предприятие Сбербанка и Mail.ru Group (доля СП в сервисе – 95%, по данным ЕГРЮЛ). Миноритарные акционеры – «Мегафон» через АО «Мегалабс» (3%) и основатель «Ситимобила» Арам Аракелян (2%).
По словам Лобушкина, изменения были запланированы еще до пандемии. Он уверяет, что водители могут не опасаться, что им будут выпадать только дешевые заказы: они равномерно распределяются между исполнителями и компенсируются более выгодными предложениями. Новая модель сокращает время ожидания машины, это увеличит количество заказов для водителей и приведет к росту их заработка, объясняет он. Лобушкин добавляет, что в режиме «предлагать» водители могли находиться далеко от места посадки пассажиров, это увеличивало время поездки.
По похожей схеме уже давно работает «Яндекс.Такси». Система распределяет заказы между водителями автоматически с помощью алгоритмов, они зависят от рейтинга исполнителей и наличия на машине символики компании, говорит представитель «Яндекс.Такси» Ася Паноян. Она подтверждает, что такая модель помогает водителям увеличить доход, так как сводит к минимуму время в пути без пассажиров, а также уменьшает время ожидания машины для пользователей.
В отличие от «Яндекс.Такси» «Ситимобил» позволяет увидеть конечную точку маршрута и отказаться от поездки при определенных условиях, говорит Лобушкин. Водители «Яндекс.Такси» также видят конечную точку и могут отказываться от заказа при определенных условиях, утверждает представитель этой компании. Представитель еще одного агрегатора такси, Gett, не раскрыл подробностей модели своей работы с водителями.
От новой схемы работы с «Ситимобилом» пока только одним плюсы, уверяет представитель ГК «Бери такси». В ее автопарке около 2300 машин в 12 городах России: Москве, Санкт-Петербурге, Нижнем Новгороде, Ярославле, Перми, Ульяновске, Воронеже, Ростове, Самаре и др. Если раньше, по словам собеседника «Ведомостей», водители могли выбирать заказы по своему желанию и часто пропускали поездки, руководствуясь какими-то своими принципами, то теперь они работают как «в офисе»: выполняют предложенные сервисом поездки подряд. Насколько увеличилась выручка «Бери такси» после перехода на новую модель, по его словам, судить рано, для этого должно пройти хотя бы несколько недель.
Если сервис сам назначает заказы водителю, существует риск, что отказов от поездок со стороны пассажиров станет больше, а это может привести к дополнительным издержкам у водителей, которые тратят бензин, чтобы доехать до клиента, предупреждает Виталий Еремин, юрист таксомоторной компании «Авеню» (работает с «Яндекс.Такси»). Когда водитель выбирает заказ самостоятельно, по его словам, такое случается реже.
После перехода на новую схему работы «Ситимобил» потерял свое конкурентное преимущество перед «Яндекс.Такси», считает председатель общественного движения «Форум-такси» Олег Амосов. Если раньше исполнители могли видеть конечную точку маршрута, цену поездки и выбирали заказ среди понравившихся им вариантов, то теперь этого нет. Он считает, что это приведет к уходу профессиональных водителей из сервиса к более крупному игроку рынка, «Яндекс.Такси».
По оценке Амосова, в условиях пандемии число заказов такси по сравнению с началом года сократилось на 80% в Москве и на 60% в регионах. В столице количество поездок на такси, по данным департамента транспорта Москвы на утро 20 мая, упало на 46% год к году. Заказов на всех не хватает и «Ситимобил», по мнению Амосова, считает, что у водителей не будет другого варианта, кроме как использовать их сервис. В период пандемии водители хватаются за любые заказы у разных агрегаторов, чтобы хоть что-то заработать, подтверждает Еремин.
Такси определение и значение | Словарь английского языка Коллинза
Примеры «такси» в предложении
такси
Эти примеры были выбраны автоматически и могут содержать конфиденциальный контент. Подробнее… Что ж, другие страны платят рабочим лучше и позволяют им оставлять больше за счет более низких налогов.The Sun (2016)
Более низкие налоги и инвестиции в инфраструктуру могут способствовать росту.Times, Sunday Times (2017)
В своем отчете обвиняет дешевую выпивку и призывает к более высоким налогам на алкоголь и минимальной цене за единицу.The Sun (2016)
Восемь из десяти человек, опрошенных в ходе опроса, заявили, что высокие налоги удерживают их от продажи.Times, Sunday Times (2016)
Он знает, что более высокие налоги — худший из возможных вариантов, поскольку Британия сталкивается с проблемами Brexit.The Sun (2017)
Экономически нецелесообразно, не говоря уже о справедливости, ожидать, что молодые люди возьмут на себя это бремя более высоких налогов.Times, Sunday Times (2017)
Снижение налогов почти не упоминается.Times, Sunday Times (2017)
Эти компании позволяют руководителям советов избегать налогов на заработную плату.Times, Sunday Times (2015)
Все мы будем платить эти долги высокими налогами и нехваткой пайков на протяжении десятилетий.The Sun (2009)
Цена включает государственные налоги и плату за обслуживание.Солнце (2008)
Подробнее …
Нам нужны более жесткие экологические налоги, чтобы изменить поведение.Times, Sunday Times (2006)
У нас самые высокие налоги и самые низкие инвестиции.Кокетт, Ричард Размышляя о немыслимом (1994)
Сколько новых налогов ввело последнее правительство?Times, Sunday Times (2012)
Но убедитесь, что вы понимаете национальные и местные налоги и социальные сборы.Times, Sunday Times (2014)
Они опасались, что зеленые налоги или ограничения на полеты могут вызвать негативную реакцию избирателей.Times, Sunday Times (2006)
Причина — совпадение более высоких цен на нефть и низких налогов.Times, Sunday Times (2007)
Но собрать деньги с помощью новых налогов сложно.Times, Sunday Times (2010)
Повышение экологических налогов должно быть компенсировано снижением налогов в других местах.Times, Sunday Times (2008)
Глобальный спад побудил правительства рассмотреть вопрос о займах для снижения налогов.Times, Sunday Times (2008)
Топливный сбор составляет две трети экологических налогов.The Sun (2011)
Потому что более низкие налоги побуждают людей больше работать для себя и для людей, которых они любят.The Sun (2015)
Первое предполагает снижение налогов и дополнительные государственные расходы, в то время как второе потребует более высоких налогов и сокращения расходов.Times, Sunday Times (2008)
Возможно, наиболее вероятным будет введение новых налогов для оплаты снижения старых.Times, Sunday Times (2015)
Правильная сумма налогов на заработную плату указана в строке 13 этой формы.Христианство сегодня (2000)
Вы находитесь в стране с высокими налогами?Times, Sunday Times (2012)
На здание может быть заложена ипотека, а также налоги и общие сборы.Times, Sunday Times (2007)
Включает полный пансион, развлечения и все налоги и сборы.The Sun (2012)
Прибыль будет частично компенсирована налогами и влиянием хеджирования, введенного против падения цены акций китайского банка.Times, Sunday Times (2009)
определение такси в The Free Dictionary
налог · составляет
(tăk’sĭs) n. пл. tax · es (tăk′sēz)1. Биология Ответное движение свободно движущегося организма или клетки к или от внешнего стимула, такого как свет.
2. Медицина Перемещение части тела путем манипуляции в нормальное положение, например, после вывиха, перелома или грыжи.
[Греческий, расположение , от tassein, tag-, до аранжировки .]
Словарь английского языка American Heritage®, пятое издание. Авторские права © 2016 Издательская компания Houghton Mifflin Harcourt. Опубликовано Houghton Mifflin Harcourt Publishing Company. Все права защищены.
такси
(ˈtæksɪs) n1. (Биология) движение клетки или организма в определенном направлении в ответ на внешний стимул
2. (Хирургия) Операция Репозиционирование смещенного органа или части только ручными манипуляциями
[C18: через новую латынь с греческого: расположение, от tassein , чтобы разместить по порядку]
Collins English Dictionary — Complete and Unabridged , 12-е издание 2014 г. © HarperCollins Publishers 1991, 1994, 1998, 2000, 2003, 2006, 2007, 2009, 2011, 2014
налог • составляет
(ˈtæk sɪs)n., pl. налог • es (ˈtæk siz)
1. расположение или порядок, как в одной из физических наук.
2. ориентированное движение подвижного организма в ответ на внешний раздражитель, как на свет.
3. изменение положения смещенной части тела путем манипуляции без разрезания.
[1720–30; <Новая латынь <Греческий táxis, производная от tássein для организации]
-taxis
или-taxis, комбинированная форма, представляющая такси 1 : хемотаксис.
Random House Словарь колледжа Кернермана Вебстера © 2010 K Dictionaries Ltd. Авторские права 2005, 1997, 1991, Random House, Inc. Все права защищены.
taxis
Ответ простейшего или низшего животного на направленный стимул, обычно связанный с движением либо к стимулу, либо от него.
Словарь незнакомых слов от Diagram Group © 2008, Diagram Visual Information Limited
Поведение животных | Протокол
Drosophila как модельный организм
Обыкновенная плодовая муха, Drosophila melanogaster, — широко используемый модельный организм в биологии, хотя более широко ее можно признать любящим фрукты вредителем.Есть несколько причин, которые делают D. melanogaster отличным экспериментальным организмом: их примерно двухнедельное время генерации позволяет изучать несколько поколений, их легко содержать в очень маленьких пробирках, а их половой диморфизм позволяет исследователям легко различать самцов, у которых есть черная окраска кзади и самки с полосатым брюшком.
Томас Хант Морган и его ученики были одними из первых, кто использовал Drosophila в начале 20-го, 90-х, 90-х, -го века.В 1933 году Морган получил Нобелевскую премию за открытие роли хромосом в наследственности, которое было основано на его наблюдениях за мутантом Drosophila . Морган продолжил свои исследования, создав новых мутантов с помощью радиации. Присутствие множества мутантов с дискретным влиянием на внешний вид мухи позволило ему взглянуть на закономерности многофакторного наследования. Ключевым аспектом его ранних работ было обнаружение явных нарушений второго закона Менделя, который гласит, что гены разделяются независимо друг от друга.Наблюдения Моргана показали, что гены не будут расщепляться независимо, если они объединены в одной хромосоме. Кроме того, стало возможным определять расположение генов на хромосоме относительно друг друга посредством анализа того, как часто они рекомбинировали, что инициировало исследования картирования генов для определения местоположения гена и расстояний между генами. В результате сотрудники лаборатории Моргана получили две дополнительные Нобелевские премии за другие исследования генетики Drosophila .Успех Моргана и накопление информации о Drosophila вдохновили других принять их в качестве экспериментальных организмов, что расширило наши знания о них и постепенно повысило их ценность как модельных организмов. Другие ученые разработали множество мутаций. Более 27 000 уникальных линий Drosophila в настоящее время поддерживаются в качестве исходных культур в центрах хранения Drosophila и легко доступны для лабораторий, которые используют их в качестве инструментов для изучения различных аспектов биологии, включая поведение 1 .
Drosophila Этология
Поведение животных или этология — это изучение того, как животные взаимодействуют друг с другом и с окружающей их средой. Хотя это относительно молодая биологическая область, этология имеет важные последствия, поскольку выживание организма зависит от его поведения в отношении возможных партнеров, источников пищи и естественных препятствий. Изучение этих организмов помогает ученым понять контексты этого поведения и лежащие в основе механизмы того, как они развивались.Таким образом, цель изучения поведения животных — найти генетическую основу поведения, проявляемого отдельными лицами или группами лиц. Например, родственный отбор, впервые предложенный Уильямом Гамильтоном, предполагает, что люди будут вести себя альтруистично, принося жертву за члена семьи, а не за незнакомца 2 . Это связано с тем, что у родственных особей больше генов, чем у неродственных, и будущее размножение их родственниками способствует их собственной приспособленности или репродуктивному успеху.Следовательно, наблюдение за тем, как организмы, подобные Drosophila , взаимодействуют со своей средой, может позволить нам понять, как это поведение жестко запрограммировано. Кроме того, такое поведение дает представление о том, как эти организмы адаптировались, чтобы справляться с окружающей средой и выжить.
Направленное поведение обычно изучается, чтобы понять, как и почему организм движется в окружающей среде. Кинезис и такси — это две формы направленного поведения. Кинезис включает вызванные стимулом движения в случайных направлениях, такие как случайное движение мух, когда их прогоняют.С другой стороны, такси подразумевает движение, вызванное стимулами, в определенном направлении, например, мотыльки, летящие к лампочке. Поведение такси является положительным, если животное движется к стимулу, и отрицательным, если животное удаляется от стимула. Кроме того, поведение такси названо в зависимости от конкретного стимула, который его вызывает. Например, геотаксис — это реакция на гравитацию, где положительный геотаксис означает, что организм движется под действием силы тяжести. Фототаксис — это движение в ответ на свет, например, положительный фототаксис бабочек по направлению к лампочке.Точно так же хемотаксис — это притяжение или избегание переносимого по воздуху химического сигнала, такого как отрицательный хемотаксис многих хищников, включая людей, в ответ на запах брызг от скунса. Кроме того, существует множество других типов налогов, которые изучаются учеными, включая аэротаксис, баротаксис, гидротаксис и магнитотаксис, которые представляют собой движения в ответ на кислород, давление, воду и магнитные поля соответственно.
Изучение поведения дрозофилы
таксиРаспространенным и простым методом изучения поведения такси Drosophila является использование камеры выбора, которая позволяет наблюдать направленную реакцию мухи на конкретный стимул, такой как сила тяжести, свет или химическое вещество. Drosophila вставляются в центр камеры выбора, а затем позволяют свободно бродить по камере, которая устроена так, что альтернативные стимулы могут быть представлены с противоположных сторон. После периода блуждания количество мух считается по обе стороны от камеры выбора. Затем количество мух на каждой стороне камеры анализируется с помощью теста хи-квадрат, чтобы определить, есть ли предпочтение стимулу.
Сравнение направленного поведения мутантной и дрозофилы дикого типа позволяет исследователям определить роль мутировавшего гена в этом конкретном поведении.Более того, ученые также пытаются понять, как животные решают, что делать при наличии комбинации стимулов, включая противоречивые стимулы 3 . Вместе эти исследования могут помочь предсказать, как организмы будут реагировать на определенные раздражители или изменения факторов окружающей среды.
Список литературы
- Cook KR, Parks AL, Jacobus LM, Kaufman TC, Matthews K. Новые исследовательские ресурсы в Bloomington Drosophila Stock Center. Fly. 2010, Т.4, 1: 88-91.
- WD, Гамильтон. Альтруизм и связанные с ним явления, в основном у социальных насекомых. Ann Rev Ecol Systematics. 1972, Т. 3, 193-202.
- Гепнер Р., Сканата М.М., Бернат Н.М., Каплоу М., Гершоу М. Вычисления, лежащие в основе фототаксиса, запахового такси и мультисенсорной интеграции у дрозофилы. eLife. 2015, Т. 4, e06229.
Эволюция реакции такси у виртуальных бактерий: неадаптивная динамика
Abstract
Бактерии способны ощущать различные внешние раздражители и реагировать на них, причем реакции варьируются от стимула к стимулу и от вида к виду.Наиболее изученным является хемотаксис в модельном организме Escherichia coli , где динамика и структура основного пути хорошо охарактеризованы. Однако неясно, насколько хорошо это подробное знание применимо к механизмам, опосредующим ответы на другие стимулы или пути у других видов. Кроме того, появляется все больше экспериментальных доказательств того, что бактерии интегрируют ответы от различных стимулов, чтобы генерировать согласованный таксисный ответ. В настоящее время нам не хватает полного понимания различных структур и динамики путей и того, как достигается эта интеграция.Чтобы изучить различные структуры и динамику путей, которые могут лежать в основе реакции таксистов у бактерий, мы проводим компьютерное моделирование эволюции таксистов. Этот подход начинается с популяции виртуальных бактерий, которые перемещаются в виртуальной среде на основе динамики простых биохимических путей, по которым они находятся. Поскольку мутации приводят к изменениям в структуре и динамике путей, бактерии, способные лучше локализоваться в благоприятных условиях, получают избирательное преимущество. Мы обнаружили, что определенная динамика последовательно развивается при различных допущениях модели и различных условиях.Эта динамика, которую мы называем неадаптивной динамикой, напрямую связывает вероятность кувырка клетки с возрастающими стимулами. Динамика, которая является адаптивной в широком диапазоне условий, как видно из пути хемотаксиса E. coli , не эволюционирует в этих эволюционных моделированиях. Однако мы обнаружили, что нехватка стимулов и колебания во время эволюции приводят к сложной динамике путей, которая приводит как к адаптивной, так и к неадаптивной динамике в зависимости от уровней базальных стимулов. Дальнейший анализ эволюционирующих структур путей показывает, что эффективная динамика такси может быть опосредована всего двумя компонентами.Неадаптивная динамика, опосредующая таксисные реакции, объясняет экспериментальные наблюдения, сделанные на мутантных штаммах E. coli и Rhodobacter sphaeroides дикого типа, которые не могли быть объяснены с помощью стандартных моделей. Мы предполагаем, что такая динамика существует и у других бактерий и играет роль, связывая метаболическое состояние клетки и реакцию таксиса. Простота механизмов, опосредующих такую динамику, делает их кандидатом на роль предшественников более сложных реакций такси, включающих адаптацию.Это исследование предполагает тесную связь между условиями стимула во время эволюции и динамикой эволюционных путей. Когда эволюция моделировалась в условиях редких и колеблющихся стимулов, эволюционировавший путь содержал черты как адаптивной, так и неадаптивной динамики, предполагая, что эти два типа динамики могут иметь разные преимущества в различных условиях окружающей среды.
Сведения об авторе
Здесь мы изучаем, как могли развиться сигнальные сети, обеспечивающие хемотаксис.Мы смоделировали эволюцию виртуальных бактерий, которые могут исследовать окружающую среду, чередуя плавание и кувырок. Частота кувырков определяется выходом сигнальной сети, которая определяет уровни внеклеточных питательных веществ, в то время как репродуктивный успех бактерий определяется их способностью находить питательные вещества. Мы обнаружили, что в условиях обильного питания бактерии быстро развивают сигнальные сети, которые обеспечивают эффективный хемотаксис, когда повышение уровня питательных веществ увеличивает частоту переворачивания.Наши результаты обеспечивают объяснение сетевой динамики, лежащей в основе подобного поведения, наблюдаемого у некоторых мутантных штаммов Escherichia coli и у других видов бактерий. Напротив, E. coli дикого типа реагирует на повышение уровня питательных веществ снижением частоты их акробатических движений и адаптацией к постоянным уровням аттрактантов. Мы наблюдаем такую динамику адаптивной сети, когда повторяем эволюционное моделирование в условиях дефицита пищи. Эти данные предполагают, что (i) адаптация не является необходимой для эффективного хемотаксиса, (ii) древняя система минимального хемотаксиса могла использовать простую связь между сигнальной сетью и метаболическим состоянием, и (iii) условия окружающей среды являются одним из определяющих факторов. для эволюции адаптивных ответов.
Образец цитирования: Гольдштейн Р.А., Сойер О.С. (2008) Эволюция реакции такси у виртуальных бактерий: неадаптивная динамика. PLoS Comput Biol 4 (5): e1000084. https://doi.org/10.1371/journal.pcbi.1000084
Редактор: Сатору Мияно, Токийский университет, Япония
Поступила: 18 сентября 2007 г .; Принята к печати: 7 апреля 2008 г .; Опубликовано: 23 мая 2008 г.
Авторские права: © 2008 Goldstein, Soyer.Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Финансирование: RAG благодарит за поддержку Национальный институт медицинских исследований (Совет медицинских исследований, Великобритания). OSS выражает признательность Министерству университетов и исследований Италии за поддержку (проект MUR-FIRB RBPR0523C3).
Конкурирующие интересы: Авторы заявили, что никаких конкурирующих интересов не существует.
Введение
Бактериальные реакции на внешние раздражители и пути, с помощью которых они опосредованы, являются модельными системами для изучения молекулярных основ поведения. Большая часть исследований в этой области была сосредоточена на хемотаксисе, способности бактерий плавать по градиенту химического аттрактанта, как это было выполнено на модельном организме Escherichia coli . Спустя более 30 лет после первых исследований [1], [2] мы теперь обладаем обширными знаниями о лежащих в основе биохимических путях [3], [4].Вкратце, E. coli плавает в прямом направлении (претерпевая некоторую вращательную диффузию), когда обратимые моторные белки на его внешней мембране вращаются против часовой стрелки (CCW) и прикрепленные жгутики переплетаются, образуя эффективный пропеллер. Когда двигатели реверсируют и вращаются по часовой стрелке (CW), жгутики отделяются и вызывают падение бактерии, что приводит к изменению направления плавания. Частота переключения мотора связана с активностью рецептора набором белков, составляющих сигнальный путь.С увеличением уровней аттрактанта возбуждающая ветвь пути вызывает подавление вращения и акробатики CW, в то время как адаптационная ветвь заставляет клетку возвращаться к исходным уровням акробатика при постоянных концентрациях аттрактанта независимо от этого уровня концентрации. Первая ветвь включает связанную с рецептором киназу CheA и связанный с ней регулятор ответа CheY, который при фосфорилировании связывает мотор и увеличивает вероятность вращения CW. Адаптация достигается за счет контроля метилирования рецептора и, следовательно, чувствительности рецептора с помощью белков CheR и CheB.Комбинация этих двух ветвей приводит к тому, что частота акробатических движений приблизительно соответствует отрицательной производной от концентрации аттрактанта по времени [5]. Адаптация является отличительной чертой этого ответа, позволяя бактериям выполнять временные сравнения аттрактанта с высокой чувствительностью в широком динамическом диапазоне [5] — [9].
Хотя эта «парадигма хемотаксиса E. coli » хорошо известна, наши знания о таксисных реакциях на другие стимулы и у других видов [10] — [12] показывают, что производная реакция с адаптацией наблюдается у E.coli не универсален и не необходим для эффективного такси. Например, ответ на кислород в E. coli , как полагают, опосредуется рецептором Aer, у которого отсутствуют сайты метилирования, что указывает на отсутствие адаптации [13]. У Rhodobacter sphaeroides адаптация к постоянным раздражителям происходит намного медленнее или не происходит совсем [14]. У тех же видов рост в аэробных условиях приводит к «инвертированной» реакции хемотаксиса, когда увеличение концентрации аттрактанта вызывает увеличение частоты кувырков [15]; этот инвертированный ответ также наблюдается у некоторых галобактерий [16] и у некоторых мутантных штаммов E.coli , которые были «выпотрошены» из некоторых или большинства белков хемотаксиса [17], [18]. Интересно, что все эти природные и мутантные штаммы все еще проявляют способность к хемотаксису.
Такая разнообразная хемотаксическая динамика могла быть результатом нескольких путей, которые позволяют бактериям интегрировать информацию о внутреннем и внешнем состоянии для создания согласованного поведения таксиса [13]. Например, у R. sphaeroides генетические исследования показывают, что большое количество таксисных белков у этого вида организовано в несколько отдельных путей [19].Хемотаксический ответ на определенные молекулы у этого вида требует транспорта в клетку [20], демонстрируя связь между метаболизмом и хемотаксисом. Несмотря на большие усилия, нам все еще не хватает всестороннего понимания различных молекулярных механизмов, участвующих в реакциях бактериального таксиса, их основной динамики и того, как эти разные динамики интегрированы.
Здесь мы используем вычислительный подход для решения некоторых из этих вопросов путем моделирования эволюции реакции такси с использованием компьютерного моделирования движения бактерий и динамики их путей.Эти симуляции используют популяцию виртуальных бактерий, существующих в виртуальном мире, вместе с источником стимула, который, как предполагается, сигнализирует о наличии благоприятных условий. Бактерии начинаются с набора невзаимодействующих белков, а также с рецептора и обратимого двигателя. Взаимодействия между белками развиваются посредством случайных мутаций, при этом бактерии отбираются для размножения на основе их способности локализоваться в местах с благоприятными условиями. Эти эволюционные симуляции постоянно приводят к появлению бактерий с сильной способностью двигаться в направлении стимула.Интересно, что в условиях обильных стимулов эти бактерии развиваются так, что вероятность кувырка напрямую связана с уровнями стимулов без какой-либо адаптации. Мы обнаружили, что такая неадаптивная динамика может быть опосредована всего двумя сигнальными компонентами, что дает возможность метаболитов действовать как эффекторы. Условия моделирования, имитирующие среду с редкими или колеблющимися источниками стимулов, приводят к эволюции путей со сложной динамикой, которые имеют черты как неадаптивной, так и адаптивной динамики.В сочетании с экспериментальными наблюдениями эти результаты демонстрируют, что адаптивная динамика не является необходимой для эффективного хемотаксиса. Эта работа также предполагает, что неадаптивная динамика лежит в основе хемотаксиса, наблюдаемого у потрошенных штаммов E. coli и может играть роль в сложном таксисном поведении R. sphaeroides . Мы предполагаем, что механизмы, приводящие к такой динамике, существуют у современных бактерий и предоставляют способ точно настроить реакцию таксистов в различных условиях или связать ее с энергетическим состоянием клетки.
Результаты
Для изучения эволюции бактериальных такси мы используем виртуальные бактерии, которые перемещаются в компьютерной двухмерной среде, содержащей фиксированный источник стимула и периодические граничные условия. Таким образом, эта компьютерная среда имитирует естественную среду с обильными стимулами. Движение этих бактерий связано с динамикой сигнального пути, состоящего из нескольких белков, которые катализируют активацию и дезактивацию друг друга, что соответствует киназам и фосфатазам в реальных клетках (см. Методы).Эти белки включают рецептор, уровень активности которого напрямую связан с уровнем локального стимула, и эффектор, который при активации может связываться с обратимым двигателем, меняя его направление и заставляя бактерии падать. Эволюционное моделирование начинается с популяции бактерий, каждая из которых содержит определенное количество белков, которые изначально не взаимодействуют. Этим бактериям позволяют исследовать окружающую среду в течение «времени генерации», состоящего из определенного количества временных шагов.На каждом временном шаге бактерии либо могут продолжать плыть вперед, либо кувыркаться, ориентируясь в новом случайном направлении. Кроме того, обновляются концентрации активированных белков в путях каждой бактерии, и рассчитывается вероятность опрокидывания на следующем временном шаге на основе концентрации активированного эффектора. После этого времени генерации бактерии отбираются для репликации на основе интегрированного количества стимулов, с которыми они столкнулись. Во время репликации существует вероятность возникновения мутаций, которые изменяют структуру и параметры биохимического пути.Подводя итог, можно сказать, что эти эволюционные симуляции связывают мутационные события, происходящие на молекулярном уровне (то есть на уровне пути), с отбором на поведенческом уровне (то есть таксической реакцией). Обратите внимание, что это общая модель, в которой стимул представляет все, что способно активировать рецептор (например, химический аттрактант, свет, pH), и нет априорных предположений относительно структуры или динамики пути.
На рис. 1 показано среднее популяции приспособленности (встреченные стимулы) во время одного эволюционного моделирования для сигнального пути, состоящего из четырех белков.Как показано, значение приспособленности быстро улучшается в течение нескольких поколений и выходит на плато. Очевидно, что структура и динамика путей у виртуальных бактерий развиваются таким образом, чтобы опосредовать таксопарения. Такое поведение можно увидеть из среднего времени, проведенного населением в различных частях окружающей среды (см. Вставки на рис. 1). В то время как неэволюционировавшие популяции бактерий распределяются независимо от источника стимула, конечные популяции могут быстро совмещаться с ним. Это поведение опосредуется динамикой определенного биохимического пути; в установившемся состоянии, при отсутствии какого-либо сигнала, концентрация активированного эффектора находится на низком уровне, и бактерия в основном плавает, не переворачиваясь (см. рисунок 2 для типичной структуры и динамики пути, кинетические параметры показаны в наборе данных S1).Когда бактерия встречает более высокий уровень стимула, эффектор быстро активируется и остается активированным до тех пор, пока присутствует сигнал, что приводит к усилению движения бактерий. Мы обнаружили, что качественный характер этого типа динамики не зависит от уровня базальных стимулов (данные не показаны). Эта неадаптивная динамика позволяет бактериям проводить больше времени в областях с высоким стимулом и плавать прямо, когда уровень стимула уменьшается. В эволюционном моделировании, повторяемом пять раз для путей от 2 до 5 белков, этот механизм всегда развивался как доминирующий.
Рисунок 1. Эволюция реакции такси in silico.
Средняя приспособленность развивающейся популяции виртуальных бактерий. На вставке показано усредненное по времени распределение позиций населения в поколении 0, 200 (соответствует пригодности приблизительно 2,0) и 5000 (последнее поколение) в виде контурного графика. Области, обведенные более темными линиями, указывают на большее количество времени, проведенного в них. Обратите внимание, что в этих симуляциях вся совокупность начинается в сетке (30,30), в то время как источник стимула фиксируется в (50,50).
https://doi.org/10.1371/journal.pcbi.1000084.g001
Рис. 2. Динамика адаптивных и неадаптивных путей.
Временной ход концентрации фосфорилированного CheY (вверху), моделируемый моделью, представленной в [10], и временной ход концентрации активного эффектора для наиболее частого пути в эволюционном моделировании, описанном на рисунке 1 (внизу). На вставке изображен этот путь в карикатуре. В обоих моделированиях системе позволяют предварительно уравновесить 1000 временных шагов.Затем стимул, равный единице, добавляется в момент времени 1000 и удаляется в момент времени 2000. Кинетические параметры для показанного пути приведены в наборе данных S1.
https://doi.org/10.1371/journal.pcbi.1000084.g002
Структуры путей, полученные в результате этого моделирования, были разнообразными (см. набор данных S1), что указывает на то, что существует несколько возможных каскадов биохимических сигналов, которые могут опосредовать неадаптивную динамику. . В случае примера пути, показанного на рисунке 2, мы обнаружили, что рецептор действует как глобальный ингибитор, отключающий эффекторную активность в отсутствие стимулов.Входящие сигналы подавляют активность рецептора, позволяя наращивать эффектор, который участвует в петле обратной связи с одним из промежуточных белков, белком (см. Изображение на рисунке 2). Другой белок действует как киназа (то есть активатор) как на рецепторе, так и на белке, тем самым обеспечивая быстрое прекращение ответа при удалении стимула. Эта сложная структура пути и результирующая динамика обеспечивают эффективное поведение хемотаксиса, как описано выше. Однако подобная динамика может быть достигнута с гораздо более простыми схемами, содержащими только два белка (см. Обсуждение).
Используя простую аналитическую модель, мы можем зафиксировать движение бактерий, опосредованное неадаптивной динамикой (см. Методы). Эта модель показывает, что в простых средах представленная динамика должна приводить к накоплению бактерий примерно пропорционально локальному уровню стимула. Обратите внимание, что до тех пор, пока уровни стимула превышают определенный порог, этот механизм чувствителен только к соотношению относительных уровней, а не к их абсолютным величинам. Это говорит о том, что эффективная реакция такси может быть достигнута в широком динамическом диапазоне стимулов с динамикой пути, которая не отображает адаптацию к стимулу и приводит к увеличению вероятности кувырка с увеличением стимула.Обе эти динамические особенности резко контрастируют с хемотаксисным поведением E. coli , где этот путь обеспечивает снижение вероятности кувырка с увеличением стимула с последующей быстрой адаптацией [5] (см. Рисунок 2).
Существует ряд различных возможных объяснений того, почему таксисные пути, эволюционирующие в этих симуляциях, характеризуются «перевернутой» реакцией (т.е. реакция на усиление стимулов противоположна той, что наблюдается в E. coli ) и неадаптивной динамикой.Во-первых, моделируемые здесь эволюционные процессы могут сделать неадаптивные пути более доступными с эволюционной точки зрения. Во-вторых, может оказаться, что смоделированные экологические ситуации особенно хорошо подходят для такси, опосредованных такой динамикой. В частности, допущения о постоянстве и изобилии стимулов в окружающей среде могут снизить потребность в адаптации. В-третьих, могут быть другие факторы, такие как внутриклеточная коммуникация, рецепторы с несколькими состояниями (т.е. рецепторы с сайтами метилирования) и различные физические процессы [21], которые не включены в модель и могут быть важны для эволюции таксистов. ответы опосредованы другой динамикой.
Чтобы увидеть, является ли неадаптивная динамика результатом сложности развития адаптивной динамики, мы выполнили дополнительное моделирование. Они начались с исходной бактериальной популяции, содержащей биохимические пути с динамикой, подобной той, что обнаружена у E. coli [22]. В пяти отдельных симуляциях бактерии всегда развивались более подходящими путями с неадаптивной динамикой и инвертированной реакцией. Другими словами, в условиях этих симуляций (то есть при большом количестве стимулов) всегда существовал путь с неадаптивной динамикой, который мог бы опосредовать более эффективную реакцию такси, чем исходный адаптивный путь.Это указывает на то, что полученные нами результаты не связаны с отсутствием эволюционного пути к обычной динамике, наблюдаемой в E. coli . Однако это не указывает на то, что реакции такси, опосредованные неадаптивной динамикой, лучше, поскольку было невозможно воспроизвести все условия окружающей среды и другие, возможно, важные особенности, как упоминалось выше.
Чтобы изучить влияние условий окружающей среды на эволюцию хемотаксиса, мы выполнили два набора моделирования при (i) периодических граничных условиях и флуктуирующем источнике стимула и (ii) непериодических граничных условиях и фиксированном источнике стимула.Первый набор условий позволяет нам проверить гипотезу о том, что адаптивная динамика позволяет бактериям сохранять устойчивость реакции на колебания внешней среды или внутренних параметров [23]. Последние условия имитируют среду с ограниченными стимулами, где ожидается, что исследование будет более важным, чем эксплуатация. В пяти симуляциях, проведенных при каждом условии, мы не нашли путей с динамикой, которые адаптируются к широкому диапазону стимулов, как это было у E.coli . Однако в результате нескольких симуляций были получены пути, которые имели динамическое поведение, подобное поведению E. coli при некоторых условиях. Как показано на рисунке 3, эти пути дают «нормальный» ответ (то есть сниженную вероятность кувырка со стимулами) и имеют ограниченную адаптацию к непрерывным стимулам. Интересно, что большинство путей развития в непериодических граничных условиях демонстрируют динамику, зависящую от базальных уровней стимулов. Это в основном влияет на динамику адаптации, и мы наблюдаем один путь достижения идеальной адаптации при узком диапазоне уровней базальных стимулов (см. Рисунок 3).Большинство симуляций, выполняемых в условиях редких стимулов, приводили к примерно таким же уровням приспособленности, как показано на рисунке 1. Однако симуляции, выполняемые в этих условиях (ii), занимали гораздо больше времени (обычно более 2000 поколений), чтобы достичь этих уровней приспособленности. Взятые вместе, эти результаты показывают, что реалистичные и сложные условия окружающей среды приводят к эволюции сложной динамики путей, которые содержат особенности как адаптивной, так и неадаптивной динамики. Выявление роли каждого типа динамики в эффективности хемотаксиса требует дальнейшего детального анализа.
Рисунок 3. Разнообразные механизмы такси.
Динамика концентрации активных эффекторов для наиболее частых и уникальных путей, полученная в результате избранных эволюционных симуляций, проводимых в различных условиях окружающей среды (см. Основной текст). Каждая панель отображает динамику для определенной структуры пути, показанной на вставке. Системе позволяют уравновеситься на 1000 временных шагов на уровне фонового базального стимула. Стимул, равный единице, добавляется во время 1000 и удаляется во время 2000.Реакция пути на такие стимулы, подаваемые поверх выбранного базального уровня, показана линиями разных типов (базальный уровень равен 0, 1 и 2, соответственно, для сплошных, пунктирных и штриховых). Пути, показанные на верхних панелях, взяты из моделирования с периодическими граничными условиями и колеблющимся источником стимула, а пути, показанные на нижней панели, взяты из моделирования с непериодическими граничными условиями и фиксированным источником стимула. Кинетические параметры для этих путей приведены в наборе данных S1.
https: // doi.org / 10.1371 / journal.pcbi.1000084.g003
Обсуждение
Молекулярные системы, опосредующие таксисные реакции, наблюдаемые у бактерий, более сложны, чем предполагает доминирующая картина хемотаксиса E. coli . Бактерии могут улавливать и реагировать на различные факторы окружающей среды, возможно, интегрируя сигнал от различных биохимических путей. Недавние экспериментальные наблюдения на растущем числе видов бактерий и прошлые исследования на мутантных штаммах E.coli указывают на разнообразие молекулярных механизмов, участвующих в генерации этих ответов. Здесь мы приводим доказательства эффективности одной из возможных динамических схем. Основными особенностями этой динамики являются перевернутая реакция, приводящая к увеличению частоты кувырков с увеличением уровня стимула, а также отсутствие адаптации к постоянным стимулам. Мы показываем, что такая неадаптивная динамика легко развивается в различных условиях окружающей среды и допущениях модели и позволяет бактериям эффективно накапливаться в благоприятных условиях.
Эти данные дают возможное объяснение неадаптивной динамики и инвертированных ответов, наблюдаемых у дикого типа R. sphaeroides [14], [15], и инвертированных ответов, наблюдаемых у галобактерий [16] и потрошенных штаммов E. coli. [18]. В каждом случае наблюдались эффективные таксисные реакции, хотя точная природа лежащих в основе молекулярных механизмов не могла быть определена. Вероятно, что эти механизмы образуют системы, подобные представленным здесь путям.Анализ результатов моделирования с двумя белками показывает минимальные сигнальные системы для достижения таксисных ответов, опосредованных неадаптивной динамикой (см. Рисунок 4). Они включают связывание сигнала с эффектором через рецептор с саморегуляцией обоих белков (посредством аллостерических взаимодействий или процессов, таких как автофосфорилирование). Поразительная простота этих минимальных систем приводит к предположению, что неадаптивная динамика может быть достигнута даже без каких-либо сигнальных белков; небольшая молекула, которая является побочным продуктом метаболизма или попадает в клетку через транспортер, может напрямую регулировать вероятность переворачивания клетки.Мы предполагаем, что именно такой сценарий отвечает за хемотаксис, наблюдаемый в потрошеной кишечной палочке E. coli [18]. Связанный с аттрактантами метаболизм вызывает повышение уровня фумарата внутри клетки, который связывает мотор и увеличивает вероятность кувырка. Экспериментально было продемонстрировано, что фумарат может участвовать в хемотаксисе [24] и может управлять переключением моторики [25], хотя точная динамика того, как он может приводить к хемотаксису, была неизвестна.
Рисунок 4. Минимальные пути с неадаптивной динамикой.
Карикатура и динамика концентрации активного эффектора для наиболее частых и уникальных путей с 2 белками, полученными из 5 различных эволюционных симуляций. Пути слева и справа были найдены в 1 и 4 моделированиях соответственно. На нижних панелях показаны кривые соответствия для соответствующих имитаций и усредненное по времени распределение позиций окончательной популяции. Кинетические параметры для этих путей приведены в наборе данных S1.
https: // doi.org / 10.1371 / journal.pcbi.1000084.g004
Если неадаптивная динамика доступна и обеспечивает эффективную реакцию такси, почему мы наблюдаем адаптивную динамику у E. coli и других видов бактерий? Адаптивные механизмы могут быть более эффективными при изучении окружающей среды и достижении устойчивой реакции на меняющиеся стимулы. Эволюция сложной динамики в симуляциях, имитирующих редкие и колеблющиеся источники стимулов, поддерживает такие аргументы. Динамика этих путей содержала как адаптивные, так и неадаптивные особенности, что дополнительно указывает на возможную сложность поведения хемотаксиса.Как показали эксперименты с потрошеной кишечной палочкой E. coli и другими видами, неадаптивные пути, вероятно, функционируют в сочетании с каноническим механизмом и участвуют в тонкой настройке реакции такси в определенных условиях окружающей среды или в обеспечении связи между энергией. соответствующие реакции такси и хемотаксис. В качестве альтернативы, учитывая простоту требуемого молекулярного механизма, неадаптивная динамика может быть предшественником более сложных адаптивных механизмов.Обе гипотезы могут быть проверены с помощью конкретных экспериментальных установок и анализа последовательности белков, участвующих в реакции такси, соответственно.
Методы
Для изучения путей, лежащих в основе ответов такси, мы использовали ранее описанную модель пути [22]. Эта модель предполагает, что путь состоит из набора белков N p , каждый из которых может существовать в деактивированном или активированном состоянии (активация может соответствовать фосфорилированию, метилированию или любому другому типу химической или структурной модификации).Каждый белок способен в активированном состоянии вызывать активацию или дезактивацию любого из других белков. Первый белок в этом пути произвольно выбран в качестве рецептора, который может активироваться внешними стимулами, в то время как белок N p произвольно выбран в качестве эффектора. При активации он может связываться с моторным белком, вызывая реверсирование мотора, и бактерия падает. Биохимическая динамика для [ P i *], фракции белка i , который активируется, подчиняется (1), где k ij ( l ij ) — скорость, с которой активируется белок j активирует (деактивирует) белок i , k ii ( l ii ) — скорость самоактивации (деактивации) белка i , k 1A ( l 1A ) представляет собой скорость, с которой стимул активирует (деактивирует) рецепторный белок 1, [ A ] — это локальный уровень стимула, а δ — это дельта Кронекера.Предполагается, что общая концентрация каждого белка равна единице. Для простоты мы предполагаем, что любой конкретный белок может активировать или деактивировать другой, но не то и другое вместе. То есть k ij l ij = 0 для всех i и j . Мы формулируем это, рассматривая значение γ ij , где положительные (отрицательные) значения γ ij соответствуют положительным значениям k ij ( l ij ).Это может быть выражено как (2) где H (x) — это ступенчатая функция Хевисайда, равная единице, если x положительна, и нулю в противном случае. Вероятность p Tumble перекатывания белка на любом заданном временном шаге дается формулой (3), где γ m — сродство белка N p к двигателю.
Эволюционное моделирование проводится с популяцией N бактерий = 1000 бактерий, существующих в двумерном квадратном пространстве 100 × 100 с периодическими граничными условиями.Уровень стимула определяется как распределение Гаусса, расположенное в центре (50, 50) пространства с максимальным значением 10,0 и шириной 7,1. В каждом поколении каждая бактерия начинается в месте (30, 30), указанном в случайном направлении, со всеми концентрациями активированного белка, установленными на ноль. На каждом временном шаге биокинетические уравнения (уравнение 1) интегрируются на основе локального уровня стимула. Затем бактерия может либо упасть (выбрать новое случайное направление) с вероятностью p Tumble , либо плыть по прямой в текущем направлении на расстояние, определяемое скоростью плавания (0.2). Каждая бактерия поглощает количество стимула, пропорциональное уровню локального стимула, но не потребляет его. В конце шага N = 5000 временных шагов следующая популяция бактерий выбирается с помощью турнирного отбора: случайным образом выбирается набор из пяти бактерий, из которых назначается бактерия, которая накопила наибольшее количество стимулов. следующему поколению. Эта процедура повторяется (с заменой) N бактерий раз. Во время каждого процесса присваивания существует вероятность p мутации = 0.1 возникающей мутации. Эта мутация включает добавление или вычитание значения, распределенного по Гауссу (среднее значение 0, стандартное отклонение 0,1), к одному из случайно выбранных γ ij .
Таким образом, полученная модель пути отражает основную биохимию сигнальных путей и позволяет связывать внешние сигналы с вероятностью кувырка. Основные предположения модели заключаются в том, что белки могут находиться только в двух состояниях и что каждый белок может взаимодействовать с любым другим.Последнее предположение допускает обобщение модели без наложения ограничений. Каждая структура пути, которая может быть построена в представленной модели, может быть построена с использованием реальной биохимии (потенциально требующей большего количества белков). Тем не менее, мы проверили влияние наложения возможных ограничений на структуру пути на эволюцию реакции такси. Они включали требование, чтобы рецептор мог быть только киназой (т.е. он мог активировать только другие белки), чтобы каждый белок мог иметь только одно взаимодействие или что саморегуляция не допускалась в модели.Мы считаем, что такие ограничения не оказывают существенного влияния на результат эволюционного моделирования (данные не показаны).
Чтобы проверить влияние колеблющегося источника стимула в окружающей среде на эволюцию реакции такси, мы запускаем дополнительное моделирование, в котором ряд параметров выбирался независимо для каждого поколения. Они включали; (i) фоновое равномерное распределение стимулов, выбранное из равномерного распределения [0–5.0], (ii) высота пика гауссовского распределения стимулов, выбираемое из равномерного распределения [1.0–10,0], (iii) дисперсия (квадрат ширины) гауссовского распределения стимулов, выбранного из равномерного распределения [50,0–500,0], и (iv) начальное местоположение бактерий, выбранное случайным образом из всех точек в 100 × 100 пространство. Кроме того, для этих прогонов 20 процентов всех мутаций привели к удалению взаимодействия (установлено равным нулю), а не к модификации путем добавления случайно выбранного приращения. Результат этих симуляций был проанализирован, подвергнув путь определенному временному графику уровней стимула и отслеживая полученный ответ, как показано на рисунке 2.
Чтобы выполнить простую оценку эффективности реакции такси, опосредованной динамикой неадаптивного пути (см. Основной текст), мы рассматриваем ситуацию равновесия, когда существует постоянная концентрация бактерий и стимулов в любом месте. На каждом временном шаге бактерия может либо кувыркаться, либо плавать, поэтому мы можем выразить вероятность того, что бактерия, которая начинается в определенном месте, поплывет к нему, равной, где p 1 → 2 — это вероятность того, что плавающая бактерия локация 1 переместится в локацию 2.Если мы теперь обратимся к детальному балансу, общий поток бактерий, плывущих из, будет в точности равен потоку плывущих из, или. В общем, p 1 → 2 будет сложной функцией, которая включает информацию о предыдущем местоположении (и, следовательно, текущем направлении плавания) бактерии в, но если мы проигнорируем эти корреляции и предположим, что бактерии плавают в изотропным образом в простом пространстве, p 1 → 2 = p 2 → 1 .Далее мы можем предположить, что для описанной динамики неадаптивного пути концентрация эффектора пропорциональна локальному уровню стимула. В этих условиях, предполагая, что вероятность опрокидывания соответствует уравнению 3, несложно показать, что относительная концентрация бактерий в различных местах определяется выражением, где C — нормировочная константа. Другими словами, для достаточного усиления (γ m λ) и уровня стимула, стационарная концентрация бактерий пропорциональна уровню стимула, что приводит к эффективной реакции такси.
Дополнительная информация
Набор данных S1.
Наиболее частые и уникальные пути, являющиеся результатом всех этапов эволюции, описанных в основном тексте.
Структура пути представлена в виде матрицы, где в каждой строке указан коэффициент взаимодействия между данным белком и всеми остальными. Буквы A, R, P и E обозначают аттрактант, рецептор, белок и эффектор соответственно. Обратите внимание, что аттрактант может взаимодействовать только с рецептором. Сродство эффектора к двигателю (γm) показано рядом с матрицей взаимодействия.Коэффициенты для каждого пути используются для моделирования его динамики, как описано в разделе «Методы». Матрицы, для которых динамическое поведение показано в основном тексте, следующие: Рисунок 4: Все показанные матрицы для 2-белковых путей, фиксированные условия аттрактанта. Рисунок 2: Матрица из опыта 03 для путей с 4 белками, фиксированные условия аттрактанта. Рисунок 3, верхний ряд: матрицы из прогонов 02, 03 и 04 для путей с 4 белками, изменяющиеся условия аттрактанта. Рисунок 3, нижний ряд: матрицы из прогонов 02, 03 и 04 для путей с 4 белками, непериодические граничные условия.
https://doi.org/10.1371/journal.pcbi.1000084.s001
(0,46 МБ DOC)
Вклад авторов
Задумал и спроектировал эксперименты: RG OS. Проведены эксперименты: РГ ОС. Проанализированы данные: РГ ОС. Предоставленные реагенты / материалы / инструменты анализа: RG OS. Написал бумагу: РГ ОС.
Ссылки
- 1. Месибов Р., Адлер Дж. (1972) Хемотаксис по отношению к аминокислотам в Escherichia coli . J Bacteriol 112: 315–326.
- 2. Adler J, Hazelbauer GL, Dahl MM (1973) Хемотаксис сахаров в Escherichia coli . J Bacteriol 115: 824–847.
- 3. Falke JJ, Bass RB, Butler SL, Chervitz SA, Danielson MA (1997) Двухкомпонентный сигнальный путь бактериального хемотаксиса: молекулярный взгляд на передачу сигнала рецепторами, киназами и адаптационными ферментами. Annu Rev Cell Dev Biol 13: 457–512.
- 4. Брен А., Эйзенбах М. (2000) Как сигналы слышны во время бактериального хемотаксиса: белок-белковые взаимодействия при распространении сенсорного сигнала.J Bacteriol 182: 6865–6873.
- 5. Segall JE, Block SM, Berg HC (1986) Временные сравнения в бактериальном хемотаксисе. Proc Natl Acad Sci U S A 83: 8987–8991.
- 6. Berg HC, Brown DA (1972) Хемотаксис в Escherichia coli проанализирован с помощью трехмерного отслеживания. Природа 239: 500–504.
- 7. Macnab RM, Koshland DE Jr (1972) Механизм определения градиента в бактериальном хемотаксисе. Proc Natl Acad Sci U S A 69: 2509–2512.
- 8. Berg HC, Tedesco PM (1975) Переходный ответ на хемотаксические стимулы в Escherichia coli . Proc Natl Acad Sci U S A 72: 3235–3239.
- 9. Weis RM, Koshland DE Jr (1988) Обратимое метилирование рецептора необходимо для нормального хемотаксиса Escherichia coli в градиентах аспарагиновой кислоты. Proc Natl Acad Sci U S A 85: 83–87.
- 10. Rao CV, Kirby JR, Arkin AP (2004) Дизайн и разнообразие бактериального хемотаксиса: сравнительное исследование на Escherichia coli и Bacillus subtilis .PLoS Biol 2: E49.
- 11. Szurmant H, Ordal GW (2004) Разнообразие механизмов хемотаксиса среди бактерий и архей. Microbiol Mol Bio Rev 68: 301–319.
- 12. Wadhams GH, Armitage JP (2004) Осмысление всего этого: бактериальный хемотаксис. Nat Rev Mol Cell Bio 5: 1024–1037.
- 13. Armitage JP (1997) Поведенческие реакции бактерий на свет и кислород. Arch Microbiol 168: 249–261.
- 14. Пул PS, Armitage JP (1988) Подвижный ответ Rhodobacter sphaeroides на хемотаксическую стимуляцию.J Bacteriol 170: 5673–5679.
- 15. Packer HL, Armitage JP (2000) Инвертированные поведенческие реакции у Rhodobacter sphaeroides дикого типа на временные стимулы. FEMS Microbiol Lett 189: 299–304.
- 16. Hoff WD, Jung KH, Spudich JL (1997) Молекулярный механизм фотосигнализации архейных сенсорных родопсинов. Анну Рев Биоф Биом 26: 223–258.
- 17. Stock J, Kersulis G, Koshland DE Jr (1985) Ни метилирующие, ни деметилирующие ферменты не требуются для бактериального хемотаксиса.Ячейка 42: 683–690.
- 18. Барак Р., Эйзенбах М. (1999) Хемотаксический ответ клеток Escherichia coli , лишенных известного механизма хемотаксиса, но содержащих сверхэкспрессированный CheY. Mol Microbiol 31: 1125–1137.
- 19. Shah DS, Porter SL, Martin AC, Hamblin PA, Armitage JP (2000) Тонкая настройка бактериального хемотаксиса: анализ поведения Rhodobacter sphaeroides в аэробных и анаэробных условиях путем мутации основных оперонов хемотаксиса и генов cheY.EMBO J 19: 4601–4613.
- 20. Ingham CJ, Armitage JP (1987) Участие транспорта в хемотаксисе Rhodobacter sphaeroides . J Bacteriol 169: 5801–5807.
- 21. Диллон Р., Фаучи Л., Гавер Д., 3-й (1995) Микромасштабная модель бактериального плавания, хемотаксиса и транспорта субстрата. Дж. Теор Био 177: 325–340.
- 22. Soyer OS, Pfeiffer T, Bonhoeffer S (2006) Моделирование эволюции путей передачи сигналов. Дж. Тео Био 241: 223–232.
- 23. Баркай Н., Лейблер С. (1997) Устойчивость в простых биохимических сетях. Nature 387: 913–917.
- 24. Montrone M, Eisenbach M, Oesterhelt D, Marwan W (1998) Регулирование частоты переключения и смещения бактериального жгутикового двигателя с помощью CheY и фумарата. J Bacteriol 180: 3375–3380.
- 25. Prasad K, Caplan SR, Eisenbach M (1998) Фумарат модулирует вращение жгутиков бактерий, уменьшая разность свободной энергии между состояниями двигателя по часовой стрелке и против часовой стрелки.J Mol Bio 280: 821–828.
такси | такси | Альтернативные формы |
Такси — это альтернативная форма такси .Во множественном числе от | taxi | lang = en разница между такси и таксисостоит в том, что такси (нерегулярные), а такси .Как существительные, разница междутакси и такси состоит в том, что такси (нерегулярно), а такси (биология) — это направленное движение организма в ответ на стимул или такси может быть.Как глаголтакси — ( такси ).Другие сравнения: в чем разница?
|
В чем разница между совместным использованием поездок и такси?
Брайан О Саттер Личный вред Дорожные аварии Ответ на этот конкретный вопрос может во многом зависеть от того, кого вы спрашиваете.По словам компаний по совместному использованию поездок, таких как Uber и Lyft, совместное использование поездок — это услуга, в которой используются преимущества современных технологий, чтобы обеспечить более непринужденный подход к поездке. Водители — это просто парни (или девушки) со своими автомобилями, которые работают в свое свободное время и могут взять столько пассажиров, сколько захотят.Тем не менее, эти компании также быстро отмечают, что их водители придерживаются строгих стандартов профессионализма во время работы, и как водители, так и пассажиры могут оценивать и проверять друг друга для дальнейшего использования.
Однако представитель компании такси с большей вероятностью скажет вам, что совместное использование поездок — это служба такси, которая использует новый формат, чтобы срезать углы и избегать правил, которые обеспечивают безопасность водителей такси и пассажиров и возможность доставить минимум стандарт качества. Пятизвездочная рейтинговая система может не дать вам полного представления о том, что вас ждет, особенно для начинающих водителей и пассажиров, и в то время как Uber и Lyft могут требовать определенного стандарта качества от водителей и их транспортных средств, они не могут предоставить своим сотрудникам, работающим по контракту, такой же уровень надзора, как менеджер службы такси.
Страховой вопрос
Одним из препятствий между компаниями, занимающимися каршерингом, и компаниями такси является то, какой размер страховки подходит для каждой услуги. Такси, как правило, сильно застрахованы с момента включения водителем зажигания до момента парковки на стоянке компании, и страховые взносы за это покрытие являются основными расходами для таксомоторных компаний.
Однако это требование о покрытии не обязательно распространяется на совместное использование поездок, поэтому службы такси заинтересованы в том, чтобы законодатели распространяли это требование о страховании на такие компании, как Uber и Lyft.Он не только защищает пассажиров, водителей и несчастных пешеходов, которые могут попасть в аварию, но и дает таксомоторным компаниям шанс на борьбу.
Точно так же и сервисы совместного использования пассажиров знают, что у них будет конкурентное преимущество, если они смогут удерживать премии, которые они должны платить, на как можно более низком уровне. Сначала это означало согласование стандартов регулирования для каждого города, но штаты играют все более важную роль в установлении общих правил.
Флорида V. Типовой законопроект
Стандартный законопроект о страховании совместного использования пассажиров, который прошел через законодательные собрания 20 штатов, уже требует большей страховки, чем типичная служба такси, но применяется только тогда, когда приложение службы активно и пассажир в машине.Однако один сенатор от Флориды выдвинул встречное предложение: вместо этого применяется постоянно меньшая минимальная страховая ставка.
Это требование напоминает правила страхования, применяемые к такси, и поэтому реакция оказалась именно такой, как вы и ожидали: долгожданное изменение для таксомоторных компаний, но повод держаться подальше от Флориды для Uber. По их словам, это изменение нелепо в свете гибкого рабочего графика их водителей, а поскольку водители Uber — это просто люди в своих автомобилях, пока у них не появится пассажир, имеет смысл требовать только обычную страховку Флориды, пока водитель ищет и вождение по тарифу.
Каким бы способом ни закончился счет за совместное использование поездок, все еще могут быть определенные пробелы благодаря тому, что водители Uber и Lyft считаются подрядчиками, а не сотрудниками со всеми вытекающими правами и компенсациями работникам. Таким образом, в городах Флориды, которые принимают услуги совместного использования пассажиров, может наблюдаться рост числа связанных с ними исков о травмах, которые не покрываются никаким страховым полисом.
3 Такси, регулирование и рост транспортных сетевых компаний | Между общественной и частной мобильностью: изучение роста транспортных услуг на базе технологий
Хинц, Г.2015. Эмануэль приветствует универсальное приложение для такси. Crain’s Chicago Business , 4 мая. Http://www.chicagobusiness.com/article/20150504/BLOGS02/150509959/emanuel-hails-a-universal-taxi-app.
ITRE. 1998. Обзор изменений в правилах использования такси в Цинциннати, Индианаполисе и Сиэтле. Международный фонд такси и ливреи, Кенсингтон, Мэриленд.
La Croix, S., J. Mak, and W. Miklius. 1992. Оценка альтернативных механизмов предоставления услуг такси в аэропорту. Обзор логистики и транспорта , Vol. 28, № 2, 147–166.
LeighFischer. 2015. Отчет ACRP 108: Коммерческий наземный транспорт в аэропортах: передовой опыт. Совет по исследованиям транспорта Национальной академии, Вашингтон, округ Колумбия
MacMillan, D., and T. Demos. 2015. Uber оценивается более чем в 50 миллиардов долларов. The Wall Street Journal , 31 июля. Http://www.wsj.com/articles/uber-valued-at-more-than-50-billion-1438367457.
Май-Дык, К.2015. Лос-Анджелес потребует от всех такси использовать «Uber-like» мобильные приложения к лету. Los Angeles Times , 16 января. Http://www.latimes.com/local/lanow/la-me-ln-los-angeles-taxi-app-20150116-story.html.
Мец, C. 2014. Доходы Uber в 12 раз превышают доходы Lyft, говорится в новом исследовании. Wired.com , 11 сентября. Http://www.wired.com/2014/09/ubers-revenue12-times-bigger-lyfts-new-study-says.
Надь, Э. 2014. Увеличение доли: рост продаж и доходов Lyft в 2014 году в пять раз. Fast Company , 11 ноября. Http://www.fastcompany.com/3038350/most-innovative-companies/increased-shares-lyfts-rides-and-revenue-grew-five-fold-in-2014.
Нельсон, Л. Дж., И К. Шеперд. 2015. LAX становится крупнейшим аэропортом США, разрешающим пикапы Uber и Lyft. Los Angeles Times , 16 июля. Http://www.latimes.com/local/lanow/la-me-ln-uber-legal-lax-20150716-story.html#page=1.
Nelson \ Nygaard Consulting Associates. 2004. Исследование правил использования такси : Рабочий документ № 1. Город Сакраменто, Калифорния.
NYC TLC. 2014. Мобильность для всех жителей Нью-Йорка. 11 июня. Http://www.nyc.gov/html/mopd/downloads/pdf/Mobility_for_All_New_Yorkers_Disabled_Accessiblity_Plan_6_11_14.pdf.
ПрайсВотерхаус. 1993. Анализ дерегулирования и перерегулирования такси. International Taxicab Foundation, Кенсингтон, Мэриленд.
Шаллер Консалтинг. 2000. Исследование потребности в разрешениях на использование такси в городе Сан-Диего. Город Сан-Диего, Калифорния.
Шаллер Консалтинг. 2006. Исследование наземного транспорта Форт-Уэрта. Город Форт-Уэрт, штат Техас.
Шаллер, Б. 2007. Въездной контроль для регулирования такси. Транспортная политика , Vol. 14, № 6, с. 490–506.
Шоу, Л. К., Дж. Гилберт, К. Бишоп и Э. Прюитт. 1983. Правила использования такси в городах США. Министерство транспорта США, Вашингтон, округ Колумбия
.