
Механизмы противостояния манипуляции: Методы противостояния манипуляциям | Статья на Psysovet.ru
Методы противостояния манипуляциям | Статья на Psysovet.ru
На тему манипуляций написано немало. Для современного человека проблема противостояния манипуляциям является чрезвычайно актуальной, ведь сталкиваться с этим явлением приходится буквально каждый день. Как ни печально осознавать, но нами все время кто-то пытается манипулировать. А если говорить начистоту, мы тоже это делаем с окружающими, не всегда даже понимая, чем занимаемся. Попробуем разобраться с понятием «манипуляция», «манипулятивное воздействие» и определить подход к этому явлению в нашей повседневной жизни.
Манипуляция как способ взаимодействия
Манипуляция неотделима от взаимодействия людей, от общения и от отношений. Абсолютно в любом типе коммуникаций есть возможность оказаться объектом манипулирования со стороны партнера. Это связано с тем, что между людьми часто возникает конфликт интересов. А стратегий взаимодействия в конфликте, согласно исследователям К. Томасу и Р. Киллмену, всего пять. Вот как они выглядят.
Вот как они выглядят.
- Уступка. Когда человек жертвует своими интересами в пользу интересов другого, например, чтобы не испортить отношения с ним.
- Уход от конфликта. Когда оба партнера уклоняются от взаимодействия по спорному вопросу, например, если предмет конфликта им несущественен.
- Компромисс. Когда оба партнера частично соблюдают свой интерес, а частично – интерес своего партнера. При этом отношения между ними не портятся и даже развиваются.
- Сотрудничество. Когда оба партнера ценят отношения и стараются удовлетворить интересы каждого из них максимально. Возможна, только если конфликт не задевает жизненно важных интересов партнеров. Если же эти интересы задеты, то остается единственная стратегия:
- Соперничество. При этой стратегии один из партнеров преследует свои интересы в максимальной степени, не учитывая интересов другой стороны. Отношения при реализации такого способа взаимодействия, безусловно, страдают.
Мы вспомнили о поведении людей в конфликте и о стратегии соперничества не случайно. Именно она лежит в основе понимания манипуляции как способа воздействия. Дело в том, что манипуляция является одной из разновидностей стратегии соперничества. Соперничество, как в природе, так и в военной науке, может осуществляться как открытым способом, так и скрытым. Так, например, во время войны командир, перед которым стоит задача захвата новой территории противника, перед выбором военной стратегии всегда взвешивает, обладает ли он достаточным ресурсов для прямой атаки. И если приходит к выводу, что ресурсы (людей, боеприпасы) лучше экономить, то может принять в качестве основной стратегии не атаку, а осаду. При таком подходе он имеет шансы, не потеряв своих солдат, получить тот же самый результат, проявив терпение и дождавшись, пока сам противник сдастся без боя, не выдержав тягот осажденного положения. Похожим образом ведут себя и некоторые животные, выслеживая свою добычу из укрытия, засады, и экономя свои силы, предпринимая решающее действие лишь в тот момент, когда их жертва ни о чем не подозревает.
Именно она лежит в основе понимания манипуляции как способа воздействия. Дело в том, что манипуляция является одной из разновидностей стратегии соперничества. Соперничество, как в природе, так и в военной науке, может осуществляться как открытым способом, так и скрытым. Так, например, во время войны командир, перед которым стоит задача захвата новой территории противника, перед выбором военной стратегии всегда взвешивает, обладает ли он достаточным ресурсов для прямой атаки. И если приходит к выводу, что ресурсы (людей, боеприпасы) лучше экономить, то может принять в качестве основной стратегии не атаку, а осаду. При таком подходе он имеет шансы, не потеряв своих солдат, получить тот же самый результат, проявив терпение и дождавшись, пока сам противник сдастся без боя, не выдержав тягот осажденного положения. Похожим образом ведут себя и некоторые животные, выслеживая свою добычу из укрытия, засады, и экономя свои силы, предпринимая решающее действие лишь в тот момент, когда их жертва ни о чем не подозревает. Они не бегут за ней по лесу или степи, не тратят энергию в борьбе, а лишь выжидают удобный момент. Преимущество скрытой атаки очевидно. Ровно так же ведут себя манипуляторы. Будучи убеждены, что их партнер по коммуникации обладает чем-то ценным для манипулятора, но не желая просить это у него напрямую, манипуляторы предпринимают действие, направленное на то, чтобы ввести жертву в заблуждение, состояние сконфуженности и, воспользовавшись этим, получить свой выигрыш. Очень важно понимать, что манипуляторы, так же как и прямые агрессоры, преследуют исключительно собственные интересы и нисколько не заботятся об интересах своего партнера. Ценность отношений для манипуляторов в этот момент не существенна. При этом если бы у манипулятора было достаточно сил, он бы просто отнял то, чем ему хочется завладеть. В связи с этим важно знать, что если вы окажете сопротивление манипулятору, это может привести к открытой форме конфликта.
Они не бегут за ней по лесу или степи, не тратят энергию в борьбе, а лишь выжидают удобный момент. Преимущество скрытой атаки очевидно. Ровно так же ведут себя манипуляторы. Будучи убеждены, что их партнер по коммуникации обладает чем-то ценным для манипулятора, но не желая просить это у него напрямую, манипуляторы предпринимают действие, направленное на то, чтобы ввести жертву в заблуждение, состояние сконфуженности и, воспользовавшись этим, получить свой выигрыш. Очень важно понимать, что манипуляторы, так же как и прямые агрессоры, преследуют исключительно собственные интересы и нисколько не заботятся об интересах своего партнера. Ценность отношений для манипуляторов в этот момент не существенна. При этом если бы у манипулятора было достаточно сил, он бы просто отнял то, чем ему хочется завладеть. В связи с этим важно знать, что если вы окажете сопротивление манипулятору, это может привести к открытой форме конфликта.
Подводя итог вышесказанному, определим манипуляцию как стратегию достижения собственных интересов, скрытую от осознания второго партнера с помощью изменения его сознания и без учета его интересов (а иногда – с нанесением ему ущерба).
Этапы манипулятивного воздействия
Манипуляцию можно представить как последовательность определенных этапов, своего рода сценарий или танец с заранее известными движениями. Известны они, конечно, лишь манипулятору, да и то, лишь в том случае, если он сам их осознает. Кстати, осознанная манипулятором манипуляция – не что иное, как мошенничество. Однако наша жизнь далеко не исчерпывается мошенничеством, манипуляций и их видов намного больше, и как мы писали выше, далеко не всякий манипулирующий отдает себе отчет в своих действиях. Это, впрочем, совсем не мешает выделить во всех манипуляциях одни и те же этапы или шаги.
- Сначала манипулятор сообщает своей жертве некую информацию, в которой правда смешана с ложью в определенной пропорции. Пропорция эта такова, что «жертва» не способна ее сразу различить. Именно это создает у «жертвы» ощущение некоторой растерянности, недоумения, ощущения «что-то не так» или “SOS” Но в этот момент «жертва» еще не способна понять, что именно и продолжает коммуникацию.

- Манипулятор продолжает осуществлять атаку на психику жертвы дополнительной информацией, а «жертва», не в силах сопротивляться дискомфорту, вызванному противоречием, улавливаемым на предыдущем шаге, вытесняет сигнал «SOS» в область бессознательного. С этого момента манипулятор может делать с «жертвой» что угодно.
Замечательной иллюстрацией этого является известный сюжет о Буратино, Лисе Алисе и Коте Базилио. В песне Алисы и Базилио отражены ключевые мишени манипуляторов, стремящихся путем обмана достичь своего интереса.
«На хвастуна не нужен нож Ему немного подпоешь, И делай с ним что хошь» «На жадину не нужен нож Ему покажешь медный грош И делай с ним что хошь» «На дурака не нужен нож Ему с три короба наврешь И делай с ним что хошь»
3. Манипулятор предлагает «жертве» выбор из двух зол, а «жертва» выбирает «наименьшее зло», тем самым приводя манипулятора к выигрышу, а себя – к проигрышу и ощущению использованности.
Основной критерий распознавания манипуляций – наличие эмоционального шлейфа после общения с манипулятором. Если вы чувствуете, что после какого-то взаимодействия осталось ощущение проигрыша, использованности, обмана, можете не сомневаться, что у вас произошел контакт с манипулятором. Такое может случиться и в личных отношениях с близким человеком, и в результате первой в жизни встречи. Эмоциональный осадок безошибочно подскажет вам, к какому типу взаимодействия можно отнести произошедшее.
Самый эффективный способ сопротивляться манипулятивному воздействию – как можно раньше обнаружить, что вы подвергаетесь манипуляции. Без осознанности на этом этапе все остальное бесполезно. Как только что вы видели в схеме осуществления манипуляции, сознание «жертвы» манипулятора настолько меняется, что пропадает способность критически оценивать происходящее. Именно это и дает шанс манипулятору добиться желаемого. Поэтому первое, что важно сделать – понять, что вы в опасности. Это не такая уж простая задача, как может показаться на первый взгляд. Наша психика крайне противится любому дискомфорту и негативным переживаниям. Людям свойственно отгонять неприятные мысли подальше. Если бы мы были устроены иначе, манипуляторы всех мастей не имели бы такого успеха. Реальность же такова, что человек не желает слушать подсказки своей интуиции и упрямо мчится вперед, к тому самому проигрышу и эмоциональному осадку.
Наша психика крайне противится любому дискомфорту и негативным переживаниям. Людям свойственно отгонять неприятные мысли подальше. Если бы мы были устроены иначе, манипуляторы всех мастей не имели бы такого успеха. Реальность же такова, что человек не желает слушать подсказки своей интуиции и упрямо мчится вперед, к тому самому проигрышу и эмоциональному осадку.
Схема противостояния манипуляции
Итак, самое важное – улавливать сигнал SOS, посылаемый вам вашим мозгом. Это может ощущаться как смутное беспокойство, какая-то неясность, ощущение противоречия, несоответствия в том, что вы слышите от партнера по общению. Никогда не пропускайте этот сигнал. Научитесь его удерживать, как бы это ни было неприятно.
Следующее важное решение – это дать себе возможность оказаться на безопасном расстоянии от источника смущающей вас информации. Вам необходимо физически прекратить доступ к своему сознанию от того, кто ввел вас в смущение. Воспользуйтесь этим временем, чтобы проанализировать ситуацию. Ваше сознание мгновенно прояснится, к вам вернется способность объективно оценивать информацию.
Ваше сознание мгновенно прояснится, к вам вернется способность объективно оценивать информацию.
На следующем шаге вам необходимо разобраться, сколько лжи вы только что получили вперемешку с правдой. Различать правду и ложь не всегда так уж просто. Есть ряд признаков, по которым можно достоверно судить о том, что вы подвергаетесь манипуляциям. Остановимся на этих ловушках манипулятора чуть подробнее.
Признаки манипуляций
- Использование двусмысленных слов и выражений позволяет манипулятору избегать ответственности за сказанное. Интонационно или с помощью подтекста манипулятор придает слову выгодный ему смысл. Пример двусмысленного вопроса из известного детского произведения: «Вы перестали пить коньяк по утрам? » показывает, что иногда двусмысленность может привести в тупик, не оставляя шанса на достойный ответ.
- Использование ложной эквивалентности, когда ваш собеседник соединяет в одной фразе истинное и ложное, проводя между ними параллели
- Использование ложной причинно-следственной связи, когда из первой части высказывания как будто вытекает вторая, хотя на самом деле это не так.

Цитата из произведения «Собачье сердце»:
Заведующая — профессору Преображенскому: «Предлагаю вам купить несколько журналов в пользу детей Германии. По полтиннику штука». Профессор Преображенский: «Нет, не возьму». Заведующая: «Вы не сочувствуете детям Германии? » Преображенский: «Сочувствую». Заведующая: «А, полтинника жалко? »
В этом примере наглядно видно, что отказ Преображенского от покупки журналов интерпретирован заведующей как эквивалент отсутствия сочувствия к детям, а так же сделан ложный вывод о наличии причинно-следственной связи между отказом от покупки и жадностью в отношении полтинника.
Использование ложного вывода, когда меняют местами причину и следствие.
Использование ложной генерализации часто применяется манипуляторами, когда они вносят в речь такие слова, как «всегда», «никогда», «все», «ничто», «везде», «нигде».
Использование искусственного преуменьшения или преувеличения, например, «ты мог бы хотя б когда-нибудь прийти на встречу вовремя? ».
Узурпация права на оценку – очень частый прием манипуляторов. «Ты сегодня как будто места себе не находишь, все время дергаешься, нервничаешь» — типичный пример того, как манипулятор сообщает партнеру по общению непрошенную оценку.
Придание негативной эмоциональной окраски обычным словам также способно смутить и выбить из колеи. «С твоей добротой ты нас по миру пустишь» — наглядный пример подобного высказывания.
Как себя вести после распознавания манипуляции
Как только вы однозначно определили, в чем вам солгал манипулятор, неизбежно встает вопрос о том, что с этим делать. Выхода существует всего три.
- Разрыв отношений. Этот вариант подходит, если мы столкнулись с беззастенчивой манипуляцией при первом же контакте и не видим необходимости в перевоспитании партнера, проще выйти из отношений. Так же может быть подходящим выводом, когда отношения исчерпали себя, и шансов на конструктивное взаимодействие нет.
- Добровольный отказ от борьбы, сдача позиций.
 Этот вариант является наименее приятным, но иногда вынужденным. Бывают ситуации, когда мы осознанно решаем отдать что-то манипулятору, понимая, что в противном случае потеряем еще больше. Несмотря на то, что принятие такого решения не уберегает нас от реального ущерба, осознанность собственного выбора позволяет избежать эмоционального шлейфа и чувства проигрыша. Выбор в пользу сокращения издержек является вполне рациональной стратегией.
Этот вариант является наименее приятным, но иногда вынужденным. Бывают ситуации, когда мы осознанно решаем отдать что-то манипулятору, понимая, что в противном случае потеряем еще больше. Несмотря на то, что принятие такого решения не уберегает нас от реального ущерба, осознанность собственного выбора позволяет избежать эмоционального шлейфа и чувства проигрыша. Выбор в пользу сокращения издержек является вполне рациональной стратегией. - Применить техники противостояния манипуляциям. Этот вариант подойдет всем, кто считает необходимым тренировать навыки отстаивания собственных границ в отношениях.
Приемы и методики противостояния манипуляциям
Основной принцип всех техник: выбирать собственную интерпретацию манипулятивного сообщения. Этот принцип обращает оружие манипулятора против него самого и соответствует подходу психологического айкидо.
Существует немало техник противостояния манипуляциям, и каждая из них имеет свою область применения и свои ограничения. Для эффективного использования приемов необходимо свободно владеть навыками распознавания манипуляций и свободно чувствовать себя в коммуникационном пространстве. Обучаться таким техникам можно на специализированных тренингах или в личной работе с коучем или психологом. Лучше всего, если вы выберете из множества приемов те, которые наилушим образом подойдут к вашим ситуациям и вашему складу характера. В этом материале мы ограничимся перечислением многообразия приемов, способных оказать эффективное сопротивление манипуляторам.
Для эффективного использования приемов необходимо свободно владеть навыками распознавания манипуляций и свободно чувствовать себя в коммуникационном пространстве. Обучаться таким техникам можно на специализированных тренингах или в личной работе с коучем или психологом. Лучше всего, если вы выберете из множества приемов те, которые наилушим образом подойдут к вашим ситуациям и вашему складу характера. В этом материале мы ограничимся перечислением многообразия приемов, способных оказать эффективное сопротивление манипуляторам.
Итак, к наиболее эффективным можно отнести методы
«Наведения тумана или заезженной пластинки». Общий смысл техники – соглашаться с какой-то частью из высказывания манипулятора, лишая его агрессивной энергии и снижая накал коммуникации. Технику можно повторять много раз, что и дало ей второе название. Действует изматывающее на манипулятора, у него полностью пропадает желание доставать свою жертву.
«Удушения в объятиях или приписывания позитивных намерений». Из названия следует, что содержанием техники является обезоруживание манипулятора путем приписывания ему положительных намерений, которые он мог и не иметь в виду. Хорошо действует на лиц, облеченных властью и нуждающихся в подтверждении своей значимости.
Из названия следует, что содержанием техники является обезоруживание манипулятора путем приписывания ему положительных намерений, которые он мог и не иметь в виду. Хорошо действует на лиц, облеченных властью и нуждающихся в подтверждении своей значимости.
«Конкретизирующие вопросы» позволяет в самом начале захода на манипуляцию продемонстрировать, что вы намерены прояснять все двусмысленности и неоднозначности в сообщении, которое направляет в ваш адрес манипулятор. Действует эффективно на тех, кто на самом деле надеялся сконфузить вас и вызвать чувство вины или стыда. Ее применение актуально в ситуациях, когда вам необходимо добиться какой-то цели от манипулятора. Не рекомендуется использовать для того, чтобы «взять манипулятора измором», вопросы должны носить содержательный характер и создавать впечатление человека, желающего разобраться, а не переиграть манипулятора. Приветствуются дополнительные разъяснения о причинах задавания вопросов: «Я лишь хочу как можно лучше вас понять. .. ».
.. ».
«Конструктивная конфронтация» показывает манипулятору, что ему не удастся провести вас, и что вы хорошо понимаете, где правда, а где ложь. Уместна в ситуации равных отношений и готовности перейти к конструктивному сотрудничеству у обеих сторон. Дает шанс отношениям, которые в принципе являются сбалансированными, но претерпевают временный сбой.
«Вопросы о цели коммуникации» призвана избежать компрометирующих ситуаций и испорченной репутации.
Пример:
Коллега- новому сотруднику «Кстати, наш руководитель рассказал кое-что нелицеприятное о вашем прежнем опыте»
Новый сотрудник: «Почему вы решили проинформировать меня об этом именно сейчас, когда наш руководитель отсутствует на своем рабочем месте? »
«Ассертивный сценарий» имеет смысл использовать для того, чтобы четко обозначить свои границы в отношениях и предупредить о возможных неизбежных последствиях, если партнер по коммуникации продолжит манипулирование. Основные элементы этого подхода – использование Я-сообщений, формирование пожеланий к изменению ситуации, обозначение неизбежных последствий.
Метод «Серого камня» является крайней мерой при взаимодействии с патологическими личностями, с которыми по ряду причин вы не можете прервать отношения. Основное содержание метода состоит в усилиях, направленных на создание максимально скучного и неинтересного образа для манипулятора, избегание любых причин для провокаций и зависти манипулятора, отвлечение внимания манипулятора на несущественные аспекты вашей жизни с тем, чтобы самые значимые и ценные для вас области оставались в безопасности.
Все эти методики и приемы вы сможете освоить на специальном тренинге, посвященном противостоянию манипуляциям, либо научиться им самостоятельно, в реальных жизненных ситуациях. Самое важное при противостоянии манипуляциям – это помнить, что у вас есть право свободного человека отказываться от того, что не соответствует вашим интересам и защищать свои границы. И, конечно, прислушиваться к голосу собственной интуиции, которая никогда не подводит вас и всегда старается предупредить о надвигающейся опасности. Ваша главная задача – развивать контакт с собственными чувствами и повышать уровень осознанности. И тогда никакой манипулятор не будет вам страшен!
Ваша главная задача – развивать контакт с собственными чувствами и повышать уровень осознанности. И тогда никакой манипулятор не будет вам страшен!
При написании статьи использовались материалы трехгодичного курса лекций и практических занятий Елены Лопухиной «Обучение психодраме», а так же материалы блогера tanja-tank.
3 универсальных приема защиты от манипуляций — Alexandra Klimenko на vc.ru
Меня зовут Александра Клименко, я продакт-менеджер (теперь уже скорее предприниматель), автор телеграм-канала «Нормально делай, нормально будет» о digital продуктах и основатель Simulator.skillslab.center — онлайн-тренажера для развития навыков soft skills.
1866 просмотров
Последние 7 лет помимо работы продакт-менеджером я вместе с мужем изучаю переговоры, конфликты и манипуляции в коммуникациях. Именно про манипуляции сегодня и хочется поговорить.
Думаю, хоть раз в своей жизни вы сталкивались с манипулятивным поведением коллег, друзей или родных. Не всегда получается четко объяснить, чем так неприятна манипуляция, а главное — почему так сложно ей противостоять. Чаще всего мы понимаем, что нами манипулировали, уже после того, как отреагировали на ситуацию не лучшим образом.
Не всегда получается четко объяснить, чем так неприятна манипуляция, а главное — почему так сложно ей противостоять. Чаще всего мы понимаем, что нами манипулировали, уже после того, как отреагировали на ситуацию не лучшим образом.
Я постараюсь рассказать, как мы боремся с манипуляциями сами и учим бороться наших учеников. Для этого сначала давайте посмотрим поближе на механизм манипуляций.
Манипуляция — это скрытое воздействие через вызов у человека определенных эмоций: вины, страха и т.д.
Во-первых, манипуляции могут быть нацелены на получение какой-то выгоды. Вот вам пример ситуации:
На встрече друзей один из них попросил у более богатого друга деньги в долг. В ответ на отказ (у второго принцип — деньги в долг не давать) говорит: «У тебя ведь денег много, а другу помочь жалко!»
Видите здесь манипуляцию? Манипулятор пытается вызвать у друга стыд за его богатство и сыграть на чувстве вины, который требует другу помочь. Чуть дальше мы обязательно поговорим о том, что делать в такой ситуации.
Чуть дальше мы обязательно поговорим о том, что делать в такой ситуации.
Далее, «долгосрочного эффекта» достигают манипуляции, которые изменяют социальные роли манипулятора и манипулируемого.
Стоп, мы же говорили об эмоциях — причем здесь социальные роли?
Дело в том, что все мы состоим в обществе и играем в нем ту или иную роль. В разных ситуациях Вы можете быть руководителем или подчиненным, родителем, ребенком, другом или коллегой. Но еще у каждой роли есть оттенки, которые делают их сложнее. Так, если авторитету опытного руководителя доверяют без сомнений, то слова слабого или неопытного руководителя могут оспариваться, а задачи игнорироваться без особых последствий. Давайте посмотрим на примере:
Руководитель не первый раз просит опытного сотрудника пояснения в каком-то вопросе. Сотрудник громко (слышно соседям по офису) говорит: «Опять тебе непонятно, что мне, придется еще раз объяснять?»
Видно, что манипулятор пытается вызывать чувство стыда из-за незнания чего-то руководителем.
Теперь давайте представим, что таких эпизодов накопилось уже немало, а руководитель на них никак не отвечает. Команде начнет казаться, что у них действительно туго соображающий, не очень опытный руководитель. Кто-то перестанет доверять такому руководителю, кто-то начнет работать менее эффективно, кто-то вообще начнет искать новую команду с другим руководителем, у которого будет чему поучиться.
Бывают еще «манипуляции ради манипуляций». Это когда манипулятор не преследует никакой рациональной цели, не хочет менять социальные роли, а просто старается достичь комфорта за счет других. Неуверенность в себе, страх за несоответствие ожиданиям общества легче игнорировать, если тонко намекнуть на несостоятельность собеседников из-за их пола, возраста или внешнего вида. Вот вам еще пример:
При устройстве на работу сотрудника попросили от руки заполнить документы для HR департамента.Забирая, анкету, HR сказала «Вам бы почерк исправить! Пишите как курица лапой!»
Разобрались с тем, в каких ситуациях применяются манипуляции. Теперь к самому важному — что с ними делать?
Как бороться с манипуляциями
Прием 1. Вскрытие цели
Один из приемов, который мы тренируем — это вскрытие цели манипулятора.
Манипуляция строится вокруг скрытого воздействия. Когда вы показываете цель манипулятора, вы лишаете его рычагов давления.
Ответ следует строить по следующей формуле:
Ты хочешь, чтобы <я испытал(а) эмоцию> и сделал(а) <желаемая цель манипулятора>?
Например, давайте представим, что богатый друг из первого примера сдался и все-таки дал манипулятору денег в долг. Прошло время возвращать, но на вопрос о долге он слышит такое:
Для ответа вскрываем манипуляцию. Эмоция — стыд, желаемая цель — чтобы долг не требовали обратно.
Тогда хорошим ответом будет, например, такой: «Ты хочешь, чтоб мне стало стыдно за то, что я хочу вернуть свои деньги и я простил тебе долг?»
Прием 2. Показать манипуляцию
Не всегда удается верно и быстро определить эмоцию и цель, хотя саму манипуляцию вы и почувствовали в том, что сказал собеседник. Можно воспользоваться вторым способом — показать манипуляцию.
Суть в том, чтобы назвать вещи своими именами и показать собеседнику, что он вами манипулирует. В этом приёме самое главное — не бояться манипуляцию назвать.
Например, в ответ на «Когда для тебя деньги стали дороже дружбы?»
можно ответить так: «Зачем ты сейчас мною манипулируешь?»Необходимо, однако, быть готовым к тому, что манипулятор сразу же начнет отрицать, говорить, что пошутил(а), что вы все придумали. Здесь важно проявить последовательность и вне зависимости от ответа собеседника спокойно обозначить, что с вами так нельзя общаться.
Прием 3. Игнорирование
Игнорирование
Этот приём мы любим меньше всего, но он может быть полезен, когда вы думаете, что манипуляция одноразовая и больше не повторится.
В этом случае главное — не идти на поводу у ложной эмоции и сделать вид, что не замечаете стремления манипулятора подтолкнуть вас к определенным действиям.
Например, на «Когда для тебя деньги стали дороже дружбы?» можно ответить «Ты наличными отдашь или на карту переведешь?»
Этот прием не лучший. Помните, если манипуляция повторилась более одного раза — она может привести к изменению социальных ролей. Зато в споре с малоприятными персонажами, которых вы больше никогда не встретите, игнорирование поможет сохранить вам силы 🙂
Здесь мы рассмотрели три универсальных приема борьбы с манипуляциями. На самом деле их намного больше. Если же вам актуально научиться противостоять манипуляциям, решать конфликты и управлять переговорами, попробуйте Симулятор жизненных ситуаций. Первая глава доступна бесплатно, чтобы оценить, подходит ли вам такой формат обучения.
В заключение я предлагаю вам в комментариях потренироваться и оставить ответы на несколько следующих манипуляций, а мы их прокомментируем:
Молодая пара студентов (22 года и 20 лет) решила снять квартиру в Москве и съехать от родителей. Владелец квартиры при знакомстве говорит: «Молодые вы! Проблем с вами будет много»
Руководитель вызвал сотрудника, который допускает ошибку в важных документах в очередной раз из-за невнимательности. Сотрудник, выслушав претензию руководства говорит: «Ну так уволь меня, если тебе не нравится как я работаю!»
Мать одной из подруг невесты решила помочь с пошивом платьев для подружек на свадьбу. Невеста обрадовалась и приняла помощь. Спустя несколько дней после свадьбы мать подруги спрашивает невесту: «А за платья ты когда заплатишь? Ясное дело, что не за «спасибо» я ВСЕ ЭТО делала!»
5 видов манипуляций и как им противостоять
Отличить манипуляцию не всегда просто. Некоторые маскируются под комплименты и безобидные просьбы. Мы попросили психолога рассказать, как понять, что вами хотят воспользоваться, и противостоять этому.
Некоторые маскируются под комплименты и безобидные просьбы. Мы попросили психолога рассказать, как понять, что вами хотят воспользоваться, и противостоять этому.
Оксана Стрижкова, практикующий психолог. Окончила ЮУрГУ по профилю «Психодиагностика и консультирование» и МГУ по программе «Социальная психология». Стаж — три с половиной года.
Манипуляции — осознанное или неосознанное давление, когда один человек использует психологически уязвимые места другого. Обычно он это делает, чтобы удовлетворить собственные желания. Манипулятор не учитывает эмоциональные потребности другого. Из-за манипуляций испаряется равенство в отношениях, что ведёт к их разрушению. Некоторые манипуляции выглядят вполне безобидно. Мы можем их использовать, даже не осознавая, что вредим отношениям.
Манипуляция на чувстве любви: «Если ты не сделаешь чего-то, ты меня не любишь!»
Она строится на логической ошибке — гиперобобщении. Манипулятор преувеличивает значение отказа, считает, что любящий человек обязан делать всё, что тот захочет. Это нереалистичное, искажённое представление об отношениях. Однако некоторые верят, что должны делать всё, чтобы доказать свою любовь. Тогда манипуляция срабатывает.
Манипулятор преувеличивает значение отказа, считает, что любящий человек обязан делать всё, что тот захочет. Это нереалистичное, искажённое представление об отношениях. Однако некоторые верят, что должны делать всё, чтобы доказать свою любовь. Тогда манипуляция срабатывает.
Как отвечать: «Я не могу сделать того, о чём ты просишь, потому что… Но это не значит, что я тебя не люблю».
Манипуляции на чувстве вины и жалости: «Что ты за человек такой, если не хочешь мне помочь?»
Мы хотим быть хорошими и одобряемыми. Это один из механизмов социальной адаптации. И наше уязвимое место. Если оно не защищено или у человека развит комплекс вины, манипуляция попадает в самое яблочко. Она тоже строится на гиперобобщении. Манипулятор строит из себя жертву или оценивает другого человека целиком по одному действию. «Если сделаешь, как я хочу, значит, ты хороший. В противном случае ты плохой». Человек бросается доказывать свою «хорошесть», спасая себя от нежелательной оценки.
В противном случае ты плохой». Человек бросается доказывать свою «хорошесть», спасая себя от нежелательной оценки.
Как отвечать: «Я — хороший человек. Мой отказ не означает обратного. Конкретно сейчас я не могу (или не хочу) помочь, потому что…».
Манипуляции на чувстве долга: «Я в тебя столько вложила, ты обязана делать, что я скажу»
Этот приём бьёт в ещё одно уязвимое место — чувство благодарности. При частом употреблении подобных фраз оно превращается в тяжёлое чувство долга. Человек понимает, что за всё полученное он будет платить свободой. Изначальной договорённости о таком обмене не было. Но в итоге человек оказывается на крючке, потому что цену за «всё хорошее, что было для тебя сделано» назначает манипулятор.
Как отвечать: «Я не просила столько делать для меня. Это был твой выбор. Мы не договаривались, что я тебе буду за это должна».
Это был твой выбор. Мы не договаривались, что я тебе буду за это должна».
Манипуляции на неуверенности: «У меня больше опыта, я лучше знаю»
Неуверенность — плодородная почва для манипуляций. Когда человек не верит в себя или свою позицию, он становится лёгкой мишенью. Манипулятор пытается подавить своим авторитетом. В ход могут идти аргументы низкого качества. Например, «Я старше, значит, я прав». Он всеми средствами добивается, чтобы позиция оппонента померкла на его фоне. Однако такие аргументы редко означают компетентность.
Как отвечать: «Опыт и возраст не играют роли в этой ситуации. У меня есть чёткие аргументы в защиту своей позиции».
Манипуляции на чувстве гордости: «Сделай, пожалуйста, за меня отчёт, ты же такая умная и отзывчивая»
Фраза кажется безобидной и даже приятной. Человека хвалят. Однако, чтобы присвоить и не потерять эти комплименты, ему приходится доказывать, что он их достоин. А для этого нужно сделать отчёт. И всё, что попросит манипулятор. На самом деле он говорит следующее: «Ты умная и отзывчивая, пока делаешь, что я хочу. Если ты откажешь, получается, ты не такая». Всем хочется быть на хорошем счету. Но если одобрение других слишком важно, человек будет делать даже то, чего не хочет, лишь бы заслужить его.
Человека хвалят. Однако, чтобы присвоить и не потерять эти комплименты, ему приходится доказывать, что он их достоин. А для этого нужно сделать отчёт. И всё, что попросит манипулятор. На самом деле он говорит следующее: «Ты умная и отзывчивая, пока делаешь, что я хочу. Если ты откажешь, получается, ты не такая». Всем хочется быть на хорошем счету. Но если одобрение других слишком важно, человек будет делать даже то, чего не хочет, лишь бы заслужить его.
Как отвечать: «Да, я умная и отзывчивая. Но делать чужой отчёт — не моя обязанность, поэтому я откажусь».
Как противостоять манипуляциям
Манипуляции по-разному влияют на человека. Например, он уверенно чувствует себя в отношениях, но не в работе. Тогда на нём не сработает манипуляция на чувстве любви, но сработает на чувстве гордости, если этим воспользуется коллега. Однако есть общие рекомендации. Манипуляции срабатывают, если человек не уверен в себе, излишне пытается понравиться другим и не может защитить свои границы.
Манипуляции срабатывают, если человек не уверен в себе, излишне пытается понравиться другим и не может защитить свои границы.
Чаще говорите нет
Если между вами и другим человеком не было договорённости, что вы должны что-то делать, вы можете отказать. Начинайте с маленьких вещей. Например, скажите подруге, что не можете посидеть с её котом, если у вас уже другие планы. Постепенно переходите к чётким «нет» в семье или на работе. Так вы прокачиваете личные границы. Через них манипуляциям пробиться сложнее.
Прислушивайтесь к своим желаниям
В следующий раз, когда откликнетесь на чью-то просьбу, спросите себя: «Зачем я это делаю?» Постарайтесь ответить честно. Возможно, это не искренняя помощь. Вы просто хотите доказать свою любовь, работоспособность, талант. Или избежать чувства вины за то, что не помогли, ведь человек столько для вас сделал.
Работайте с самооценкой
Гораздо сложнее воспользоваться человеком, который уверен, что ему не надо никому ничего доказывать. Ведь он сам знает о своих способностях, таланте и уме. Когда манипулятор попытается доказать, что вы — плохой и недостойный человек, здоровая самооценка поможет этому противостоять. Учитесь замечать свои достоинства и вспоминайте о них почаще. Принимайте комплименты, а неудачи воспринимайте как полезный опыт. Ставьте реалистичные цели и отстаивайте свои границы, чтобы не подкармливать неуверенность в том, что вы чего-то не можете.
- Теги
- Отношения
- Психология
- Vkontakte
- Viber
Манипуляция партнером в супружеских отношениях
Автор: Гейфман Александра Альбертовна
Рубрика: Психология
Опубликовано
в
Молодой учёный
№18 (256) май 2019 г.
Дата публикации: 05.05.2019 2019-05-05
Статья просмотрена: 2773 раза
Скачать электронную версию
Скачать Часть 1 (pdf)
Библиографическое описание:Гейфман, А. А. Манипуляция партнером в супружеских отношениях / А. А. Гейфман. — Текст : непосредственный // Молодой ученый. — 2019. — № 18 (256). — С. 5-7. — URL: https://moluch.ru/archive/256/58631/ (дата обращения: 01.10.2022).
В данной статье рассматриваются манипуляции между супругами, манипулирование в подобных отношениях для получения собственной выгоды. Манипуляция в супружеских парах являются достаточно распространенной проблемой в отношениях, поэтому эта тема является актуальной на сегодняшний день.
Ключевые слова: манипуляция, отношение, манипулятор, механизм влияния, ожидаемая реакция.
На протяжении всей истории социум был основой выживания человека. Чтобы общество могло существовать как целостное образование, должны были возникнуть механизмы влияния на отдельных его участников.
Поэтому практически каждый человек наделен таким механизмом влияния, который делает его уязвимым для положительного или отрицательного состояния другого человека и общества.
Стремясь избавиться от негативного воздействия, некоторые люди привыкают действовать реактивно, не задумываясь поддаваясь на манипуляции. Таким образом, человек лишает себя возможности пересмотреть ситуацию и действовать по-другому. Люди, которые находятся рядом с манипулятором, также привыкают к тому, что они могут быть вынуждены действовать через определенное чувство, и начинают манипулировать сами.
Не секрет, что любое общение является, прежде всего, управлением. Взаимодействие с другими людьми основано на том, что мы всегда хотим чего-то друг от друга и управляем друг другом для достижения цели. То же самое происходит и в супружеских отношениях, потому что не всегда желания близких людей одинаковы, и если не удается договориться или спросить, они начинают влиять друг на друга посредством манипуляций.
Взаимодействие с другими людьми основано на том, что мы всегда хотим чего-то друг от друга и управляем друг другом для достижения цели. То же самое происходит и в супружеских отношениях, потому что не всегда желания близких людей одинаковы, и если не удается договориться или спросить, они начинают влиять друг на друга посредством манипуляций.
Под манипуляцией понимается психологическое воздействие, исполнение которого ведёт к скрытому возбуждению у другого человека намерений, не совпадающих с его актуально существующими планами и желаниями. [1]
Манипуляция основана на легком когнитивном диссонансе, который возникает из-за смешения правды и лжи. Достаточно самого маленького искажения фактов, чтобы возникло дискомфортное ощущение от противоречивости данных.
Отношения в супружеских парах часто строятся на манипулятивных действиях, осуществляемых сознательно или неосознанно. Манипуляции в супружеских отношениях направлены на то, что бы удовлетворить личные потребности за счет другого человека, при этом обходя прямое и открытое взаимодействие с ним.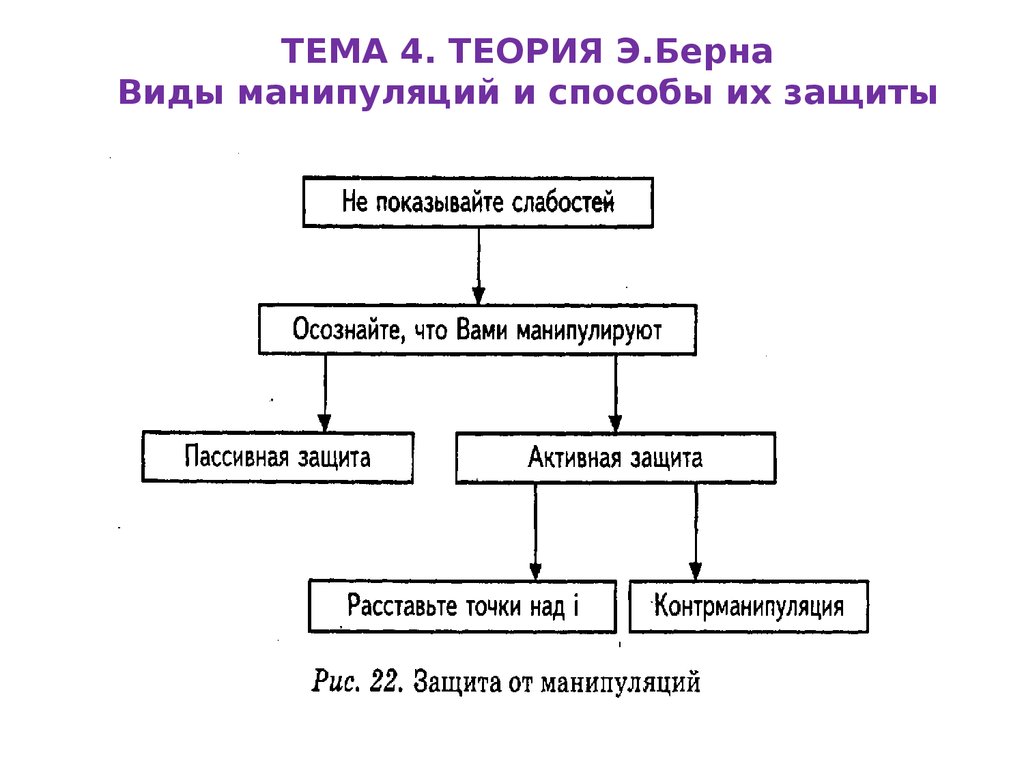 Другими словами, в таких отношениях один из партнеров стремиться к удовлетворению своих потребностей искаженным путем и может даже не задумываться о нуждах другого.
Другими словами, в таких отношениях один из партнеров стремиться к удовлетворению своих потребностей искаженным путем и может даже не задумываться о нуждах другого.
Манипулирующий супруг проявляет осознанную жестокость, во что бы то ни стало, заставляя другого человека полностью подчиниться своей воле, Как правило, манипулирующий находит слабое место у своего партнера и воздействует на эти слабости.
Манипулятор характеризуется эгоистичным характером, стремлением к власти в личных отношениях и неспособностью по-настоящему любить и сострадать. Подчиняющийся, напротив, располагает мягким характером, чувствительностью, эмоциональной зависимостью.
Манипуляция в супружеских отношениях может быть трех видов:
– Негативной – против интересов партнера.
– Позитивной – в рамках обоюдных интересов. Может быть и развлечением, и игрой, и заботой.
– Нейтральной – с точки зрения интересов партнера.
В руках умелого манипулятора супруг незаметно для себя самого принимает несвойственные решения, соглашается на нежелательные действия. Со временем такие отношения становятся нормой семьи, а достижение личных целей манипулятора оборачивается в корыстную игру в одни ворота. [2, с. 109]
Со временем такие отношения становятся нормой семьи, а достижение личных целей манипулятора оборачивается в корыстную игру в одни ворота. [2, с. 109]
Манипулирование в супружеских отношениях имеет свои особенности у супругов разного пола и с различным стажем брака. Данный факт конкретизируется в следующих предположениях:
- Супруги, состоящие в браке более 10 лет, более склонны к манипуляции, чем супруги, брачный стаж которых менее 5 лет.
- Склонность к манипуляции в супружеских отношениях у мужчин выше, чем у женщин.
- Имеются отличия в приемах манипулирования, которые используют в супружеском общении мужчины и женщины.
В супружеских отношениях существует несколько основных методов манипуляций, которыми пользуется манипулятор для достижения своих личных целей:
-
Манипуляция любовью. Очень часто один из супругов хоть раз, но говорил своему мужу или жене – «если любишь – сделаешь». Это самая слабая сторона эмоционально-зависимого.
 Ведь страх быть отвергнутым или потерять любимого – один из самых сильных. Поэтому манипулятор может добиться от супруга исполнения своих целей посредством давления на чувства.
Ведь страх быть отвергнутым или потерять любимого – один из самых сильных. Поэтому манипулятор может добиться от супруга исполнения своих целей посредством давления на чувства.
- Манипуляция гневом. Манипулятор, когда видит, что его просьбы не выполнены, может сорваться на супруга, будет кричать и топать ногами, принудит сдаться и подчиниться манипуляции. Если один раз пойти у него на поводу – значит, крики и скандалы станут неотъемлемой частью супружеской жизни.
- Манипуляция чувством вины. Эмоционально-зависимый манипулятор будет пытаться манипулировать своим партнером словами «это из-за тебя я стал таким», потому что он сознательно делает из себя жертву.
-
Манипуляция шантажом. Обещание манипулятора бросить воздействовать на волю супруга скрывает под собой желание вознаграждения. Если этот метод не действует, в ход идет шантаж со стороны манипулятора. Если он угрожает самоубийством – нельзя верить ему. Такие люди слишком себя любят, чтобы пойти на такое.
 [2, с. 111]
[2, с. 111]
На самом деле, манипуляции в супружеских отношениях – явление вполне привычное. Более того, любые взаимоотношения по своей природе манипулятивны. У каждого есть цель, и каждый хочет ее достичь. Новсе должно быть в меру. Если супружеские отношения полностью построены на различного рода манипуляциях, когда в них нет места простому легкому общению и радости пребывания вдвоем – тогда необходимо задуматься о том, как не поддаваться на манипуляции.
Находясь внутри супружеских отношений, будучи супругом, сложно даже признаться себе, что супружеский партнер манипулирует. Но естественно и обязательно одно – манипуляции надо пресекать, неважно – сознательно вами управляют или манипулятор сам подвержен психологическим проблемам. Жить с манипулятором, стараться его понять, да еще и защитить себя от его манипуляций очень сложная задача.
Следует понять, что борьба против манипуляций только заставляет напрягаться, усиливает тревогу, а главное – разрушает основу отношений: отсутствует возможность быть открытым, свободное решение быть рядом, быть собой.
Некоторые люди привыкли находиться в образе жертвы, и они могут уверенно держаться за любую сторону оппозиции – сами манипулировать или поддаваться манипуляции.
Манипуляция во время конфликта в семье потенциально опасна, даже если используется во благо. Поэтому важно знать, как вести себя с манипулятором в супружеских отношениях. В зависимости от ситуации, необходимо выбрать наиболее подходящую модель поведения: [3]
- Завершение контакта.
Если манипулятор не желает отступать, нужно отойти от него, не доказывать ничего и не оправдываться. Следует предложить завершить разговор для успокоения эмоций.
- Обратите внимание, имеет ли манипулятор моральное право говорить то, что говорит.
- Отсутствие ожидаемой реакции.
Такая техника защиты от манипулятора в супружеских отношениях крайне полезна, когда манипулятор заранее знает, как супруг себя поведет: заплачет, уйдет, начнет оправдываться и т. п. Либо, когда он пытается вывести на конфликт. Суть техники отсутствия ожидаемой реакции в ответе на манипуляцию согласием.
п. Либо, когда он пытается вывести на конфликт. Суть техники отсутствия ожидаемой реакции в ответе на манипуляцию согласием.
- Выражение благодарности за заботу.
Техника «задабривания» работает, когда партнер начинает манипулировать, чтобы привлечь к себе внимание. Однако противостоять можно, дав положительные эмоции вместо вызываемых (чувство вины, обиды и т.п.).
- Игнорирование подтекста, отвечая манипулятору только на сказанное.
- Конструктивный ответ.
Разбор манипуляции на 2 части: реальный факт и эмоциональная нагрузка. Отвечать следует только на факт с приведением доказательств. Максимально конструктивно.
- Задавание конкретных уточняющих вопросов.
Они помогут остановить напор манипулятора и ослабить пыл. Их суть – перевести разговор от эмоционального в конструктивное русло.
-
Задать прямой вопрос о цели манипуляции.

Смысл этого метода – дать понять манипулятору, что вы понимаете его мотивы и не поддадитесь им. Вопрос должен быть максимально открытым, произнесенным без претензии и эмоций.
Следовательно, чтобы противостоять манипуляции в супружеских отношениях и не дать развиться конфликту на этой почве, важно не вступать в начатую игру. При первых же намеках на нее – агрессивную, скрытую, дружелюбную – нужно дать понять манипулятору, что вы осознаете его намерения. [3]
Таким образом, манипуляции в супружеских отношениях говорят о не состоятельности и отсутствии готовности взять ответственность за свою жизнь полностью на себя. Человек, который манипулирует другим, делает второго зависимым от себя и сам является зависимым, в первую очередь от подобного способа выстраивать отношения. В таких семьях отсутствует способность создавать открытые и чистые взаимодействия ради общего счастья и страдать будут оба супруга.
Литература:
-
Доценко Е.
 Л. Психология манипуляции: феномены, механизмы и защита. М.: ЧеРо, 1997. 344 с.
Л. Психология манипуляции: феномены, механизмы и защита. М.: ЧеРо, 1997. 344 с.
- Сотникова Т. Ю. Манипуляции в созависимых отношениях //Стратегические направления развития науки, образования, технологий. – 2017. – С. 108-112.
- Танасов Г.Г. Манипулятивное поведение в отношениях женщин и мужчин с партнером противоположного пола / Г.Г. Танасов, О.А. Сысоева. Г. Краснодар: Научный журнал КубГАУ, №69(05), 2011.
Основные термины (генерируются автоматически): манипуляция, отношение, манипулятор, механизм влияния, ожидаемая реакция, супруг.
Ключевые слова
манипуляция, манипулятор, отношение, механизм влияния, ожидаемая реакцияманипуляция, отношение, манипулятор, механизм влияния, ожидаемая реакция
Похожие статьи
Основные технологии
манипулирования в общенииМанипуляция – один из наиболее интересных видов человеческого поведения, это неотъемлемая часть социальной культуры.
Степень успешности манипуляции зависит от того, насколько широк арсенал этих средств и насколько манипулятор гибок в их использовании.
Гендерный аспект в вопросе склонности к
манипулированиюМанипуляция — это вид психологического воздействия, искусное исполнение которого ведет к скрытому возбуждению у другого человека намерений
Эрих Фромм выдвигает вторую причину манипулирования: желая получить любовь, манипулятор стремиться получить полную…
Технологии психологических
манипуляций и управленческих…То есть манипуляция есть искусство психологического воздействия, такой его вид, при котором манипулятор способен вызвать у объекта намерения, не совпадающие с реальными
То есть освещать совершенно лживую, но ожидаемую и желаемую массами информацию.
Политическое
манипулирование: суть и механизмы защитыВ данной статье рассматривается процесс политического манипулирования, определяются характерные признаки политического манипулирования, анализируются вербальные и невербальные составляющие политического манипулирования…
Характеристика личности
манипулятора | Статья в сборнике…Манипуляция — это вид психологического воздействия, искусное исполнение которого ведет к скрытому возбуждению у другого человека намерений. Эрих Фромм выдвигает вторую причину манипулирования: желая получить любовь, манипулятор стремиться получить полную…
Манипулятивные технологии в современном информационном…
Одной из главных проблем современного информационного общества, для которого характерна, с одной стороны, глобализация информационной системы, а, с другой стороны, фрагментация сознания людей. ..
..
Современные манипулятивные приемы | Статья в журнале…
Именно воздействие на массовое сознание для изменения поведения общественности в корыстных целях стало основой деятельности
Так, немецкий социолог Г.Франке под манипуляцией понимает «своего рода психическое воздействие, которое проводится тайно…
Проблема
манипуляции массовым сознанием как фактор…— скрытое воздействие, факт которого не должен быть замечен объектом манипуляции (успех манипуляции
— отношение к манипулируемым людям не как к личностям, а как к объектам, особого рода вещам
Данный метод также называется «проблема-реакция-решение».
Манипуляция сознанием в массовой культуре | Статья в журнале…
В данной статье рассматривается проблема манипуляции сознанием, анализируются способы манипуляции, а также ее последствия. Оценивается роль массовой культуры, как фактора формирования данного явления.
Оценивается роль массовой культуры, как фактора формирования данного явления.
Похожие статьи
Основные технологии
манипулирования в общенииМанипуляция – один из наиболее интересных видов человеческого поведения, это неотъемлемая часть социальной культуры.
Степень успешности манипуляции зависит от того, насколько широк арсенал этих средств и насколько манипулятор гибок в их использовании.
Гендерный аспект в вопросе склонности к
манипулированиюМанипуляция — это вид психологического воздействия, искусное исполнение которого ведет к скрытому возбуждению у другого человека намерений
Эрих Фромм выдвигает вторую причину манипулирования: желая получить любовь, манипулятор стремиться получить полную…
Технологии психологических
манипуляций и управленческих. ..
..
То есть манипуляция есть искусство психологического воздействия, такой его вид, при котором манипулятор способен вызвать у объекта намерения, не совпадающие с реальными
То есть освещать совершенно лживую, но ожидаемую и желаемую массами информацию.
Политическое
манипулирование: суть и механизмы защитыВ данной статье рассматривается процесс политического манипулирования, определяются характерные признаки политического манипулирования, анализируются вербальные и невербальные составляющие политического манипулирования…
Характеристика личности
манипулятора | Статья в сборнике…Манипуляция — это вид психологического воздействия, искусное исполнение которого ведет к скрытому возбуждению у другого человека намерений. Эрих Фромм выдвигает вторую причину манипулирования: желая получить любовь, манипулятор стремиться получить полную…
Эрих Фромм выдвигает вторую причину манипулирования: желая получить любовь, манипулятор стремиться получить полную…
Манипулятивные технологии в современном информационном…
Одной из главных проблем современного информационного общества, для которого характерна, с одной стороны, глобализация информационной системы, а, с другой стороны, фрагментация сознания людей…
Современные манипулятивные приемы | Статья в журнале…
Именно воздействие на массовое сознание для изменения поведения общественности в корыстных целях стало основой деятельности
Так, немецкий социолог Г.Франке под манипуляцией понимает «своего рода психическое воздействие, которое проводится тайно…
Проблема
манипуляции массовым сознанием как фактор…— скрытое воздействие, факт которого не должен быть замечен объектом манипуляции (успех манипуляции
— отношение к манипулируемым людям не как к личностям, а как к объектам, особого рода вещам
Данный метод также называется «проблема-реакция-решение».
Манипуляция сознанием в массовой культуре | Статья в журнале…
В данной статье рассматривается проблема манипуляции сознанием, анализируются способы манипуляции, а также ее последствия. Оценивается роль массовой культуры, как фактора формирования данного явления.
|
||||||||||
|
||||||||||
|
||||||||||
|
||||||||||
|
© 2006-2022 Издательство ГРАМОТА разработка и создание сайта, поисковая оптимизация: krav.  ru ru
|
||||||||||
 Вопросы теории и практики (входит в перечень ВАК). Тамбов: Грамота, 2017. № 1. Ч. 1. С. 156-160.
Вопросы теории и практики (входит в перечень ВАК). Тамбов: Грамота, 2017. № 1. Ч. 1. С. 156-160. Отмечается, что термин «обнажение приема», встречающийся в работах литературоведческой направленности, может также применяться в лингвопрагматических исследованиях контрманипуляции для наименования речевой тактики, связанной с обнаружением и разоблачением адресатом используемого адресантом приема манипулятивного речевого воздействия. Выявленная тактика рассматривается на различном массмедийном материале: телевизионной передаче, радиопередаче и газетном тексте.
Отмечается, что термин «обнажение приема», встречающийся в работах литературоведческой направленности, может также применяться в лингвопрагматических исследованиях контрманипуляции для наименования речевой тактики, связанной с обнаружением и разоблачением адресатом используемого адресантом приема манипулятивного речевого воздействия. Выявленная тактика рассматривается на различном массмедийном материале: телевизионной передаче, радиопередаче и газетном тексте. Бесплатный просмотрщик PDF-файлов можно скачать здесь.
Бесплатный просмотрщик PDF-файлов можно скачать здесь.  … к. полит. н. М., 2011. 235 с.
… к. полит. н. М., 2011. 235 с. Изд-е 6-е, перераб. и доп. Ростов-на-Дону: Феникс, 2006. 224 с.
Изд-е 6-е, перераб. и доп. Ростов-на-Дону: Феникс, 2006. 224 с. Ю. Моделирование манипуляций сознанием масс в политическом процессе с помощью коммуникационного поля // Вестник Нижегородского университета им. Н. И. Лобачевского. 2011. № 6: в 3-х ч. Ч. 1. С. 326-331.
Ю. Моделирование манипуляций сознанием масс в политическом процессе с помощью коммуникационного поля // Вестник Нижегородского университета им. Н. И. Лобачевского. 2011. № 6: в 3-х ч. Ч. 1. С. 326-331. Ф. Контрсуггестия и история // История и психология: сб. М.: Мысль, 1972. С. 7-35.
Ф. Контрсуггестия и история // История и психология: сб. М.: Мысль, 1972. С. 7-35.
 М.: АСТ, 2016. 192 с.
М.: АСТ, 2016. 192 с.Как противостоять манипуляциям: способы защиты от попыток управлять вами
Как противостоять манипуляциям, если вами пытаются управлять?
Манипуляция – слово достаточно неприятное. Особенно когда употребляется оно по отношению к вашей персоне. Но, так или иначе, с манипулированием сталкивается каждый из нас ежедневно.
Сюда можно отнести как попытки кого-либо из окружения влиять на наши личные поступки и решения, так и привычное воздействие на нас рекламы с билбордов или дисплеев гаджетов.
Весьма логично, что вам сразу же захочется защититься от попыток кого бы то ни было склонять вас к определенным действиям и поступкам. Тогда необходимо четко понимать, какие методы защиты от манипуляции вообще существуют. Об этом и побеседуем.
Как противостоять манипуляциям в реальной жизни?
На самом деле, защититься от манипуляций не трудно. Ведь на все есть свой рецепт и решение. Но прежде чем мы затронем тему механизма защиты от манипуляций, стоит усвоить три основных и достаточно важных момента. Их понимание поможет в использовании тех методик, которые будут описаны в дальнейшем.
Но прежде чем мы затронем тему механизма защиты от манипуляций, стоит усвоить три основных и достаточно важных момента. Их понимание поможет в использовании тех методик, которые будут описаны в дальнейшем.
Итак, три условия, которые необходимо соблюдать:
- ваша речь и формулировки должны быть четкими и ясными;
- ваша интонация должна соответствовать ситуации и быть внятной;
- каждый ваш ответ на реплику манипулятора (человека, пытающегося навязать вам свою волю и свои решения) обязан быть основательным и уверенным.
Усвоили? Отлично, тогда идем дальше. Ведь на этих трех аспектах зиждется все, что будет перечислено ниже. А ниже следуют основные способы защиты от манипуляции, которые однажды были кропотливо собраны известным американским социальным психологом Филиппом Зимбардо.
Способы защиты от манипуляции
Чувствуете, что коллега, родственник или любой другой человек пытается навязать вам свое мнение, заставив вас поступать так, как нужно ему?
Такие действия могут быть как в форме активного эмоционального убеждения собеседника, так и в попытке “надавить на жалость”, либо даже в чем-то обвинить. Чтобы стать хозяином своих собственных решений, откроем 12 способов, которые обязательно помогут противостоять манипуляциям подобного характера.
Чтобы стать хозяином своих собственных решений, откроем 12 способов, которые обязательно помогут противостоять манипуляциям подобного характера.
Читайте по теме Мощные современные приемы манипуляции людьми
Распознавайте и отражайте манипуляцию
-
“Выход из шаблона”.
Это метод, который заключается в следующем – научитесь вести себя в диалоге, выходя из свойственной вам формы и манеры общения. Попробуйте на себе роль человека с противоположным характером. К примеру, отреагируйте так, как того вовсе не ожидает манипулятор. Главное – даже не думайте при этом оправдываться или пререкаться.
-
Примите свои ошибки и недостатки.
Ясно и четко заявите, что вы действительно допустили ошибку. Согласившись в том, в чем вас обвиняют – вы обезоружите своего собеседника и сбросите с себя груз ненужных обязательств, построенных на угрызениях совести.
-
Не ведитесь на навязывание манипулятором его картины мира.
Предложите альтернативу того, как лично вы видите обсуждаемую ситуацию.
 Находитесь в готовности вовремя сделать шаг назад и заявить о неприятии того видения, которое навязывается вам другим человеком.
Находитесь в готовности вовремя сделать шаг назад и заявить о неприятии того видения, которое навязывается вам другим человеком. -
Не пугайтесь кратковременного отказа от благ.
Которое может последовать за прекращением контактов с манипулятором (тех же денег или прочих материальных ресурсов). Если человек открыто навязывает вам выгодные ему действия – порвите ваши отношения и рассматривайте полученные от этого знания, как полезный в будущем опыт.
-
Будьте готовы отказаться от общения с манипулятором.
Этот пункт сопряжен с предыдущим, но кое в чем все же отличается. Здесь речь идет о нематериальных вещах. Не бойтесь сказать манипулирующему вами человеку в лицо слово “хватит”. Примите то, что вы самодостаточны и способны жить без расположения со стороны кого-либо. Это может быть как отказ от товарищеского общения, так и от проявлений симпатии противоположным полом.
-
Не позволяйте вас торопить.
Часто манипулятор умело торопит вас в принятии решений.
 И делать он это может искусно и довольно эмоционально. Не ведитесь на подобные уловки, в любой ситуации требуйте время на раздумия.
И делать он это может искусно и довольно эмоционально. Не ведитесь на подобные уловки, в любой ситуации требуйте время на раздумия. -
Настаивайте на внятных и конкретизированных объяснениях той или иной ситуации.
Также не реагируйте на провокации, нацеленные на то, чтобы выставить вас глупым. Если какие-либо объяснения были двусмысленны – требуйте конкретики!
-
Не слишком заигрывайтесь в модель отношений “хозяин – гость”.
Находясь в условиях, которые приписывают вам поведение гостя, не слишком увлекайтесь данной ролью. Ведь часто такие правила ощутимо ограничивают вашу персональную свободу действий, прикрываясь при этом общепринятой моделью поведения.
-
Не ждите от человека безосновательной симпатии и даже любви.
Это один из наиболее важных принципов, на котором базируются механизмы защиты от манипуляции. Нет, вовсе не значит, что нужно везде ожидать подвох и подставу. Это означает лишь то, что на отношения (особенно с незнакомыми вам людьми) стоит смотреть все-таки чуточку прагматичнее.

-
Попробуйте себя в роли беспристрастной и холодной личности.
Как ни странно, при этом можно спокойно оставаться добрым, отзывчивым и хорошим человеком. Однако же критическое мышление и оценивание действий других людей без эмоциональной составляющей значительно увеличат ваши шансы на спокойное и уверенное достижение собственных целей.
-
Искорените из себя чувство вины, диктуемое вам кем-то извне.
Рассматривайте ошибки как опыт и стремитесь при этом их не совершать, однако никогда не “переваривайте” в голове все то, что уже давно прошло. Именно регулярное переживание ситуаций из прошлого, основанных на ощущении собственной вины, способны выбить вас из колеи и сделать подвластным человеку-манипулятору.
-
Действуйте не исходя из устоявшихся привычек и законов, а исходя из собственных суждений.
Примите идею того, что вы можете изменяться – ведь вы живой человек. Даже если от вас привыкли ожидать одного, а вы решили поступить иначе – вы имеете полное на это право.

Делать так, как выгодно – не значит быть плохим
Не стоит забывать, что почти каждый в мире старается извлечь свою выгоду. Зачастую люди разнятся лишь тем, что одни принимают такую идею, а другие считают ее чем-то совершенно неприемлемым и сами же из-за этого страдают. Если хотите преуспеть – отнесите себя к первому типу. Это абсолютно не значит, что вы станете эгоистом. Наоборот вы сможете взглянуть на многие вещи с холодной головой и освободиться от оков надуманных обязательств и правил, которыми умело пользуются в своих целях манипуляторы.
Устойчивость к паклитакселу: молекулярные механизмы и фармакологические манипуляции
Обзор
. 2003 г., февраль; 3 (1): 1–19.
дои: 10.2174/156800
33754. Р З Юсуф
1
, З. Дуан, Д. Е. Ламендола, Р. Т. Пенсон, М. В. Зайден
Дуан, Д. Е. Ламендола, Р. Т. Пенсон, М. В. Зайден
принадлежность
- 1 Отделение гематологии/онкологии Массачусетской больницы общего профиля, Бостон, Массачусетс 02114, США.
- PMID: 12570657
- DOI: 10.2174/156800 33754
Обзор
R Z Yusuf et al. Цели лекарств против рака Curr. 2003 Февраль
. 2003 г., февраль; 3 (1): 1–19.
дои: 10.2174/156800
33754.Авторы
Р З Юсуф
1
, З. Дуан, Д. Э. Ламендола, Р. Т. Пенсон, М. В. Зайден
Дуан, Д. Э. Ламендола, Р. Т. Пенсон, М. В. Зайден
принадлежность
- 1 Отделение гематологии/онкологии Массачусетской больницы общего профиля, Бостон, Массачусетс 02114, США.
- PMID: 12570657
- DOI: 10.2174/156800 33754
Абстрактный
Прошло примерно десять лет с тех пор, как Управление по санитарному надзору за качеством пищевых продуктов и медикаментов (FDA) одобрило паклитаксел для лечения резистентной к платине эпителиальной карциномы яичников. С момента одобрения препарат нашел терапевтическое применение в различных режимах и при широком спектре злокачественных новообразований эпителия. Его новый механизм действия дал надежду на то, что он продемонстрирует противоопухолевую активность в опухолевых клетках с множественной лекарственной устойчивостью. К сожалению, как и в случае с другими химиотерапевтическими препаратами, обычно наблюдается резистентность. Лабораторные исследования определили широкий спектр механизмов резистентности, включая сверхэкспрессию гена множественной лекарственной устойчивости (MDR-1), молекулярные изменения в молекуле-мишени (бетатубулин), изменения в белках, регулирующих апоптоз, и контрольных точках митоза, а в последнее время — изменения в составе липидов и, возможно, гиперэкспрессия интерлейкина 6 (ИЛ-6). В этом обзоре описываются молекулярные данные in vitro, которые определяют и поддерживают различные механизмы резистентности, и критически оцениваются доказательства участия этих механизмов в клинически значимой резистентности к паклитакселу. В этом обзоре также исследуются фармакологические попытки модулировать резистентность к паклитакселу, главным образом путем ингибирования помпы оттока лекарственного средства MDR-1.
Его новый механизм действия дал надежду на то, что он продемонстрирует противоопухолевую активность в опухолевых клетках с множественной лекарственной устойчивостью. К сожалению, как и в случае с другими химиотерапевтическими препаратами, обычно наблюдается резистентность. Лабораторные исследования определили широкий спектр механизмов резистентности, включая сверхэкспрессию гена множественной лекарственной устойчивости (MDR-1), молекулярные изменения в молекуле-мишени (бетатубулин), изменения в белках, регулирующих апоптоз, и контрольных точках митоза, а в последнее время — изменения в составе липидов и, возможно, гиперэкспрессия интерлейкина 6 (ИЛ-6). В этом обзоре описываются молекулярные данные in vitro, которые определяют и поддерживают различные механизмы резистентности, и критически оцениваются доказательства участия этих механизмов в клинически значимой резистентности к паклитакселу. В этом обзоре также исследуются фармакологические попытки модулировать резистентность к паклитакселу, главным образом путем ингибирования помпы оттока лекарственного средства MDR-1. Обсуждаются будущие направления исследований лекарственной устойчивости и ее фармакологических манипуляций в клинике.
Обсуждаются будущие направления исследований лекарственной устойчивости и ее фармакологических манипуляций в клинике.
Похожие статьи
-
Модуляция лекарственной устойчивости альфа-тубулином в устойчивых к паклитакселу клеточных линиях рака легких человека.
Хан Э.К., Герке Л., Тахир С.К., Кредо Р.Б., Чериан С.П., Шам Х., Розенберг С.Х., Нг С. Хан Э.К. и др. Евр Джей Рак. 2000 авг; 36 (12): 1565-71. doi: 10.1016/s0959-8049(00)00145-3. Евр Джей Рак. 2000. PMID: 10
5 -
Лапатиниб (Tykerb, GW572016) обращает множественную лекарственную устойчивость в раковых клетках путем ингибирования активности члена 1 подсемейства B и члена 2 G АТФ-связывающей кассеты.
Дай С.Л., Тивари А.К., Ву С.П., Су XD, Ван С.
 Р., Лю Д.Г., Эшби С.Р. младший, Хуан И, Роби Р.В., Лян И.Дж., Чен Л.М., Ши Си Джей, Амбудкар С.В., Чен З.С., Фу Л.В.
Дай С.Л. и соавт.
Рак Рез. 2008 1 октября; 68 (19): 7905-14. doi: 10.1158/0008-5472.CAN-08-0499.
Рак Рез. 2008.
PMID: 18829547
Бесплатная статья ЧВК.
Р., Лю Д.Г., Эшби С.Р. младший, Хуан И, Роби Р.В., Лян И.Дж., Чен Л.М., Ши Си Джей, Амбудкар С.В., Чен З.С., Фу Л.В.
Дай С.Л. и соавт.
Рак Рез. 2008 1 октября; 68 (19): 7905-14. doi: 10.1158/0008-5472.CAN-08-0499.
Рак Рез. 2008.
PMID: 18829547
Бесплатная статья ЧВК. -
Опухолевые эндотелиальные клетки приобретают лекарственную устойчивость за счет активации MDR1 посредством передачи сигналов VEGF в микроокружении опухоли.
Акияма К., Ога Н., Хида Ю., Кавамото Т., Садамото Ю., Исикава С., Майши Н., Акино Т., Кондо М., Мацуда А., Иноуэ Н., Синдо М., Хида К. Акияма К. и др. Ам Джей Патол. 2012 март; 180(3):1283-1293. doi: 10.1016/j.ajpath.2011.11.029. Epub 2012 13 января. Ам Джей Патол. 2012. PMID: 22245726
-
Не только P-гликопротеин: амплификация ABCB1-содержащего участка хромосомы 7q21 придает множественную лекарственную устойчивость раковым клеткам за счет скоординированной сверхэкспрессии набора белков, связанных с устойчивостью.

Дженовезе И., Илари А., Ассараф Ю.Г., Фази Ф., Колотти Г. Дженовезе I и др. Обновление устойчивости к наркотикам. 2017 Май; 32:23-46. doi: 10.1016/j.drup.2017.10.003. Epub 2017 16 октября. Обновление устойчивости к наркотикам. 2017. PMID: 2
76 Обзор. -
Множественная лекарственная устойчивость: ретроспектива и перспективы противораковой лекарственной терапии.
Перес-Томас Р. Перес-Томас Р. Курр Мед Хим. 2006;13(16):1859-76. дои: 10.2174/0
706777585077. Курр Мед Хим. 2006. PMID: 16842198 Обзор.
Посмотреть все похожие статьи
Цитируется
-
Соображения по разработке наночастиц для совместной доставки нуклеиновых кислот и противораковых препаратов для устранения химиорезистентности.

Элджек С., Дэвид С., Фаггад А., Чурпа И., Аллард-Ваннье Э. Элджек С. и др. Int J Pharm X. 6 сентября 2022 г .; 4: 100126. doi: 10.1016/j.ijpx.2022.100126. электронная коллекция 2022 дек. Международный J Pharm X. 2022. PMID: 36147518 Бесплатная статья ЧВК.
-
Роль оси CircRNA/miRNA/mRNA в лекарственной устойчивости рака молочной железы.
Газиморади М.Х., Бабашах С. Газиморади М.Х. и соавт. Фронт Онкол. 2022 5 сент.; 12:966083. doi: 10.3389/fonc.2022.966083. Электронная коллекция 2022. Фронт Онкол. 2022. PMID: 36132137 Бесплатная статья ЧВК. Обзор.
-
Флупиртин в лечении вызванной таксанами боли у онкологических больных.
Пол А., Разак М.А., Биной А., Деви П.У., Павитран К.
 Пол А и др.
Индийский J Palliat Care. 2022 июль-сен;28(3):262-265. дои: 10.25259/IJPC_41_2021. Epub 2021 21 октября.
Индийский J Palliat Care. 2022.
PMID: 36072241
Бесплатная статья ЧВК.
Пол А и др.
Индийский J Palliat Care. 2022 июль-сен;28(3):262-265. дои: 10.25259/IJPC_41_2021. Epub 2021 21 октября.
Индийский J Palliat Care. 2022.
PMID: 36072241
Бесплатная статья ЧВК. -
Гинзенозид Rg5 сенсибилизирует устойчивые к паклитакселу клетки цервикальной аденокарциномы человека к паклитакселу и усиливает противораковое действие паклитаксела.
Рамеш Дж., Тилакан Р.С., Гопалакришнан Р.М., Виджаяпупати С., Доршель А., Венугопал Б. Рамеш Дж. и др. Гены (Базель). 2022 24 июня; 13 (7): 1142. дои: 10.3390/гены13071142. Гены (Базель). 2022. PMID: 35885925 Бесплатная статья ЧВК.
-
Участие ДНК-метилтрансферазы 1 (DNMT1) и белков, связанных с множественной лекарственной устойчивостью, в цитотоксичности, вызванной 2-метоксиэстрадиолом, в клетках EC109/Taxol.

Ян Ц., Го С., Сюй Ю., Дуань С., Ван Х., Фэн Ц., Чжан Н. Ян Кью и др. Транс Рак Res. 2021 янв;10(1):10-21. doi: 10.21037/tcr-20-2678. Транс Рак Res. 2021. PMID: 35116235 Бесплатная статья ЧВК.
Просмотреть все статьи «Цитируется по»
Типы публикаций
термины MeSH
вещества
Биология упражнений с отягощениями: манипулирование переменными программы упражнений с отягощениями определяет реакцию клеточных и молекулярных сигнальных путей
. 2008;38(7):527-40.
doi: 10.2165/00007256-200838070-00001.
Барри А. Спиринг
1
, Уильям Дж. Кремер, Джеффри М. Андерсон, Лоуренс Э. Армстронг, Брэдли С. Ниндл, Джефф С. Волек, Карл М. Мареш
Спиринг
1
, Уильям Дж. Кремер, Джеффри М. Андерсон, Лоуренс Э. Армстронг, Брэдли С. Ниндл, Джефф С. Волек, Карл М. Мареш
Принадлежности
принадлежность
- 1 Лаборатория функциональных возможностей человека, кафедра кинезиологии, Коннектикутский университет, Сторрс, Коннектикут 06269, США.
- PMID: 18557656
- DOI: 10.2165/00007256-200838070-00001
Барри А. Спиринг и др. Спорт Мед. 2008.
. 2008;38(7):527-40.
doi: 10. 2165/00007256-200838070-00001.
2165/00007256-200838070-00001.
Авторы
Барри А. Спиринг 1 , Уильям Дж. Кремер, Джеффри М. Андерсон, Лоуренс Э. Армстронг, Брэдли С. Ниндл, Джефф С. Волек, Карл М. Мареш
принадлежность
- 1 Лаборатория функциональных возможностей человека, кафедра кинезиологии, Коннектикутский университет, Сторрс, Коннектикут 06269, США.
- PMID: 18557656
- DOI: 10.2165/00007256-200838070-00001
Абстрактный
Последние достижения в области молекулярной биологии прояснили некоторые механизмы, регулирующие рост скелетных мышц. Логично предположить, что мышечные физиологи применили эти инновации к изучению упражнений с отягощениями (RE), поскольку RE представляет собой наиболее мощный естественный стимул для роста скелетных мышц взрослого человека. Однако по мере того, как это молекулярное направление исследований переходит к исследованиям на людях, ученые должны понимать фундаментальные принципы RE для эффективного планирования таких экспериментов. Поэтому мы представляем здесь обновленную парадигму биологии RE, которая объединяет фундаментальные принципы RE с современными знаниями о мышечных клеточных и молекулярных сигналах. RE вызывает последовательный каскад, состоящий из: (i) активации мышц; (ii) сигнальные события, возникающие в результате механической деформации мышечных волокон, гормонов и иммунных/воспалительных реакций; (iii) синтез белка за счет усиления транскрипции и трансляции; и (iv) гипертрофия мышечных волокон. В этой парадигме RE считается «восходящим» сигналом, который определяет конкретные события нисходящего потока.
Логично предположить, что мышечные физиологи применили эти инновации к изучению упражнений с отягощениями (RE), поскольку RE представляет собой наиболее мощный естественный стимул для роста скелетных мышц взрослого человека. Однако по мере того, как это молекулярное направление исследований переходит к исследованиям на людях, ученые должны понимать фундаментальные принципы RE для эффективного планирования таких экспериментов. Поэтому мы представляем здесь обновленную парадигму биологии RE, которая объединяет фундаментальные принципы RE с современными знаниями о мышечных клеточных и молекулярных сигналах. RE вызывает последовательный каскад, состоящий из: (i) активации мышц; (ii) сигнальные события, возникающие в результате механической деформации мышечных волокон, гормонов и иммунных/воспалительных реакций; (iii) синтез белка за счет усиления транскрипции и трансляции; и (iv) гипертрофия мышечных волокон. В этой парадигме RE считается «восходящим» сигналом, который определяет конкретные события нисходящего потока. Следовательно, манипулирование переменными программы острой RE (т. е. выбор упражнений, нагрузка, объем, продолжительность периода отдыха и порядок упражнений) изменяет уникальный «отпечаток пальца» стимула RE и впоследствии изменяет последующие клеточные и молекулярные реакции.
Следовательно, манипулирование переменными программы острой RE (т. е. выбор упражнений, нагрузка, объем, продолжительность периода отдыха и порядок упражнений) изменяет уникальный «отпечаток пальца» стимула RE и впоследствии изменяет последующие клеточные и молекулярные реакции.
Похожие статьи
-
Избранный вклад: острые клеточные и молекулярные реакции на упражнения с отягощениями.
Хаддад Ф., Адамс Г.Р. Хаддад Ф. и др. J Appl Physiol (1985). 2002 г., июль; 93 (1): 394–403. doi: 10.1152/japplphysiol.01153.2001. J Appl Physiol (1985). 2002. PMID: 12070230
-
Активация мышечных волокон не зависит от нагрузки и продолжительности повторений, когда упражнения с отягощениями выполняются до отказа.
Мортон Р.
Клиническое испытание. В., Сонне М.В., Фариас Зунига А., Мохаммад И.З., Джонс А., МакГлори С., Кейр П.Дж., Потвин Дж.Р., Филлипс С.М.
Мортон Р.В. и соавт.
Дж. Физиол. 2019 сен; 597 (17): 4601-4613. дои: 10.1113/JP278056. Epub 2019 27 июля.
Дж. Физиол. 2019.
PMID: 312
В., Сонне М.В., Фариас Зунига А., Мохаммад И.З., Джонс А., МакГлори С., Кейр П.Дж., Потвин Дж.Р., Филлипс С.М.
Мортон Р.В. и соавт.
Дж. Физиол. 2019 сен; 597 (17): 4601-4613. дои: 10.1113/JP278056. Epub 2019 27 июля.
Дж. Физиол. 2019.
PMID: 312 -
Роль интенсивности упражнений с отягощениями на адаптацию мышечных волокон.
Фрай переменного тока. Фрай АС. Спорт Мед. 2004;34(10):663-79. doi: 10.2165/00007256-200434100-00004. Спорт Мед. 2004. PMID: 15335243 Обзор.
-
Индуцированные сокращением изменения TNF-альфа и Akt-опосредованной передачи сигналов связаны с увеличением миофибриллярного белка в скелетных мышцах крыс.
Карагунис Л.Г., Яспелкис Б.Б. 3-й, Ридер Д.
 В., Ланкастер Г.И., Хоули Дж.А., Коффи В.Г.
Карагунис Л.Г. и соавт.
Eur J Appl Physiol. 2010 июль; 109 (5): 839-48. doi: 10.1007/s00421-010-1427-5. Epub 2010 13 марта.
Eur J Appl Physiol. 2010.
PMID: 20229019
В., Ланкастер Г.И., Хоули Дж.А., Коффи В.Г.
Карагунис Л.Г. и соавт.
Eur J Appl Physiol. 2010 июль; 109 (5): 839-48. doi: 10.1007/s00421-010-1427-5. Epub 2010 13 марта.
Eur J Appl Physiol. 2010.
PMID: 20229019 -
Индуцированные физической нагрузкой фосфопротеины в скелетных мышцах.
Дешмукх А.С., Хоули Дж.А., Зират Дж.Р. Дешмух А.С. и др. Int J Obes (Лондон). 2008 Сентябрь; 32 Приложение 4: S18-23. doi: 10.1038/ijo.2008.118. Int J Obes (Лондон). 2008. PMID: 18719593 Обзор.
Посмотреть все похожие статьи
Цитируется
-
На пути к новой парадигме тренировок с отягощениями с помощью мониторинга скорости: критический и сложный рассказ.
Гонсалес-Бадильо Х.
 Дж., Санчес-Медина Л., Рибас-Серна Х., Родригес-Роселл Д.
Гонсалес-Бадильо Дж. Дж. и др.
Открытый мед. спорт. 2022 16 сентября; 8 (1): 118. doi: 10.1186/s40798-022-00513-z.
Открытый мед. спорт. 2022.
PMID: 36114395
Бесплатная статья ЧВК.
Обзор.
Дж., Санчес-Медина Л., Рибас-Серна Х., Родригес-Роселл Д.
Гонсалес-Бадильо Дж. Дж. и др.
Открытый мед. спорт. 2022 16 сентября; 8 (1): 118. doi: 10.1186/s40798-022-00513-z.
Открытый мед. спорт. 2022.
PMID: 36114395
Бесплатная статья ЧВК.
Обзор. -
Воспроизводимость и применимость рекомендаций по традиционным силовым тренировкам.
Эредиа-Эльвар Х.Р., Эрнандес-Лугедо Х., Майкас-Перес Л., Нотарио-Алонсо Р., Гарначо-Кастаньо М.В., Гарсия-Фернандес П., Мате-Муньос Х.Л. Эредиа-Элвар Дж. Р. и соавт. Биология (Базель). 2022 2 июня; 11 (6): 851. doi: 10.3390/biology11060851. Биология (Базель). 2022. PMID: 35741372 Бесплатная статья ЧВК.
-
Биокомпартментализация инсулиноподобного фактора роста-I в крови, интерстициальной жидкости и мышцах до и после 3 месяцев хронических упражнений с отягощениями.

Стерчала А.Дж., Пирс Дж.Р., Барнс Б.Р., Урсо М.Л., Матени Р.В., Скофилд Д.Э., Фланаган С.Д., Мареш К.М., Замбраски Э.Дж., Кремер В.Дж., Ниндл Б.К. Стерчала А.Дж. и соавт. J Appl Physiol (1985). 2022 1 июля; 133 (1): 170-182. doi: 10.1152/japplphysiol.00592.2021. Epub 2022 9 июня. J Appl Physiol (1985). 2022. PMID: 35678743
-
Влияние прекондиционирования гипоксии-гипероксии на показатели мышечного повреждения после острой силовой нагрузки у спортсменов-мужчин.
Chen PW, Hsu CC, Lai LF, Chi CP, Yu SH. Чен П.В. и соавт. Фронт Физиол. 2022 19 апр;13:824210. doi: 10.3389/fphys.2022.824210. Электронная коллекция 2022. Фронт Физиол. 2022. PMID: 35514339Бесплатная статья ЧВК.
-
Миокины и тренировка с отягощениями: обзор повествования.

Zunner BEM, Wachsmuth NB, Eckstein ML, Scherl L, Schierbauer JR, Haupt S, Stumpf C, Reusch L, Moser O. Зуннер БЭМ и др. Int J Mol Sci. 2022 23 марта; 23 (7): 3501. дои: 10.3390/ijms23073501. Int J Mol Sci. 2022. PMID: 35408868 Бесплатная статья ЧВК. Обзор.
Просмотреть все статьи «Цитируется по»
использованная литература
-
- Медицинские спортивные упражнения. 2002 фев; 34 (2): 364-80 — пабмед
-
- наук СТКЭ. 2004 27 июля; 2004 (244): re11 — пабмед
-
-
J Нейрофизиол.
 1965 май; 28:560-80
—
пабмед
1965 май; 28:560-80
—
пабмед
-
J Нейрофизиол.
-
- J Appl Physiol (1985). 2001 июль; 91 (1): 57-64 — пабмед
-
- Бр Дж Спорт Мед. 2007 г., июнь; 41 (6): 346-8; обсуждение 348 — пабмед
Типы публикаций
термины MeSH
вещества
Программируемое по времени дозирование лекарств позволяет манипулировать, подавлять и обращать устойчивость к антибиотикам in vitro
Введение
Появление бактерий с множественной лекарственной устойчивостью, или «супербактерий», представляет непосредственную угрозу для нашего общества и ускоряется рядом факторов, таких как чрезмерное и неправильное использование существующих антибиотиков, а также сокращение поставок антибиотиков 1,2 . Было предложено использовать комбинацию препаратов в качестве многокомпонентной стратегии, например, комбинированную терапию и циклическое применение антибиотиков, чтобы справиться с текущей ситуацией. На сегодняшний день был проведен широкий спектр исследований для выяснения влияния одного, а также нескольких препаратов на эволюцию бактерий. Фармакокинетические/фармакодинамические исследования in vitro выявили оптимальные режимы дозирования с использованием нескольких препаратов, которые могут эффективно подавлять рост бактерий и предотвращать появление лекарственно-устойчивых мутантов 3,4,5,6 . Лабораторные исследования эволюции с использованием секвенирования всего генома 7,8,9,10,11,12 и анализа транскриптома 13,14 были применены для изучения долгосрочной динамики адаптации бактерий к стрессовым лекарственным средам, а также для получения информации о сложные взаимосвязи между лекарственной устойчивостью и генетическими изменениями и изменениями экспрессии генов.
Было предложено использовать комбинацию препаратов в качестве многокомпонентной стратегии, например, комбинированную терапию и циклическое применение антибиотиков, чтобы справиться с текущей ситуацией. На сегодняшний день был проведен широкий спектр исследований для выяснения влияния одного, а также нескольких препаратов на эволюцию бактерий. Фармакокинетические/фармакодинамические исследования in vitro выявили оптимальные режимы дозирования с использованием нескольких препаратов, которые могут эффективно подавлять рост бактерий и предотвращать появление лекарственно-устойчивых мутантов 3,4,5,6 . Лабораторные исследования эволюции с использованием секвенирования всего генома 7,8,9,10,11,12 и анализа транскриптома 13,14 были применены для изучения долгосрочной динамики адаптации бактерий к стрессовым лекарственным средам, а также для получения информации о сложные взаимосвязи между лекарственной устойчивостью и генетическими изменениями и изменениями экспрессии генов. Теоретическое моделирование использовалось для понимания эволюции бактерий с системной точки зрения. Различные модели, включая фамакодинамику 15,16 , популяционная генетика 17,18 и динамика популяций 19,20 были разработаны для изучения результатов, полученных в результате экспериментальных исследований. Цикличность антибиотиков также изучалась в клинических условиях более 30 лет, в частности широко изучалась цикличность аминогликозидов из-за повышения лекарственной устойчивости, опосредованной плазмидами, несущими ферменты аминогликозидов 21,22 . Велосипед в отделении интенсивной терапии также активно исследовался, потому что инфекции, вызванные лекарственно-устойчивыми бактериями, могут быть смертельными, если лечение не удается 23 .
Теоретическое моделирование использовалось для понимания эволюции бактерий с системной точки зрения. Различные модели, включая фамакодинамику 15,16 , популяционная генетика 17,18 и динамика популяций 19,20 были разработаны для изучения результатов, полученных в результате экспериментальных исследований. Цикличность антибиотиков также изучалась в клинических условиях более 30 лет, в частности широко изучалась цикличность аминогликозидов из-за повышения лекарственной устойчивости, опосредованной плазмидами, несущими ферменты аминогликозидов 21,22 . Велосипед в отделении интенсивной терапии также активно исследовался, потому что инфекции, вызванные лекарственно-устойчивыми бактериями, могут быть смертельными, если лечение не удается 23 .
Несмотря на ряд клинических, лабораторных и теоретических исследований с различными комбинациями антибиотиков 20,24,25,26,27,28 , они показали неоднозначные результаты отчасти из-за отсутствия стандартных процедур для проведения экспериментов. Однако недавние исследования показывают, что использование побочной чувствительности, при которой бактериальные штаммы, устойчивые к антибиотику, проявляют повышенную чувствительность к другим антибиотикам 12,13,29 , может быть ключом к подавлению или обращению эволюции бактериальной лекарственной устойчивости 30 . Таким образом, использование различных антибиотиков с временной модуляцией является многообещающим методом-кандидатом для эффективного дозирования нескольких лекарств, поскольку смена антибиотиков может избирательно гибнуть устойчивые штаммы из-за затрат на приспособление, связанных с лекарственной устойчивостью, при этом позволяя чувствительным штаммам разрастаться. К настоящему времени несколько лабораторных эволюционных исследований, основанных на побочной чувствительности, продемонстрировали эффективность чередования антибиотиков с определенными комбинациями препаратов и режимами дозирования 9,10,31,32 . Тем не менее, всестороннее экспериментальное исследование эффективных комбинаций антибиотиков для подавления возникновения лекарственной устойчивости бактерий еще предстоит провести.
Однако недавние исследования показывают, что использование побочной чувствительности, при которой бактериальные штаммы, устойчивые к антибиотику, проявляют повышенную чувствительность к другим антибиотикам 12,13,29 , может быть ключом к подавлению или обращению эволюции бактериальной лекарственной устойчивости 30 . Таким образом, использование различных антибиотиков с временной модуляцией является многообещающим методом-кандидатом для эффективного дозирования нескольких лекарств, поскольку смена антибиотиков может избирательно гибнуть устойчивые штаммы из-за затрат на приспособление, связанных с лекарственной устойчивостью, при этом позволяя чувствительным штаммам разрастаться. К настоящему времени несколько лабораторных эволюционных исследований, основанных на побочной чувствительности, продемонстрировали эффективность чередования антибиотиков с определенными комбинациями препаратов и режимами дозирования 9,10,31,32 . Тем не менее, всестороннее экспериментальное исследование эффективных комбинаций антибиотиков для подавления возникновения лекарственной устойчивости бактерий еще предстоит провести.
В этой работе мы исследуем эффективные комбинации антибиотиков, которые подавляют развитие лекарственной устойчивости в условиях циклического стресса, и то, как эволюция множества лекарств может быть связана с эволюцией в условиях одного антибиотика. Мы обнаруживаем уникальные эволюционные закономерности (то есть временное развитие бактериальной лекарственной устойчивости), при которых возникновение лекарственной устойчивости к одному из антибиотиков, включенных в цикл, полностью подавляется. На основе полученных данных построена простая математическая модель, которая позволяет нам предсказать судьбу эволюции многих лекарств на основе эволюционных паттернов в условиях одного лекарства. Затем, объединив экспериментальные данные и прогнозы модели, мы демонстрируем, что обращение вспять развившейся множественной лекарственной устойчивости возможно путем временной модуляции стресса от антибиотиков.
Результаты
План эксперимента и платформа
Это исследование направлено на получение информации об эволюции бактерий в условиях колеблющегося стресса антибиотиков путем разработки серии долгосрочных (24 дня) лабораторных экспериментов по эволюции. Эксперименты были разработаны для воздействия на бактериальные культуры одиночного стресса антибиотиками и ряда конфигурируемых чередующихся стрессов антибиотиков. Тип препарата и концентрация применяемого препарата либо переключались (включались/выключались), либо колебались в диапазоне переменных периодов времени (рис. 1а). Для достижения этого типа программируемого состояния наркотиков в окружающей среде мы разработали и построили автоматизированную платформу морбидостата 9.0009 7 , применение которого позволило оценить развитие устойчивости к антибиотикам (измеряемой как изменения чувствительности) в зависимости от состава нескольких лекарств, относительных концентраций и времени (дополнительная рис. 1). В морбидостате концентрация антибиотика автоматически контролировалась по пользовательскому алгоритму 7 ; при достижении бактериальной культурой определенной плотности на 12 мл бактериальной культуры вводили 1 мл питательной среды, содержащей антибиотик, а при низкой плотности клеток добавляли среду, не содержащую антибиотик (Методы).
Эксперименты были разработаны для воздействия на бактериальные культуры одиночного стресса антибиотиками и ряда конфигурируемых чередующихся стрессов антибиотиков. Тип препарата и концентрация применяемого препарата либо переключались (включались/выключались), либо колебались в диапазоне переменных периодов времени (рис. 1а). Для достижения этого типа программируемого состояния наркотиков в окружающей среде мы разработали и построили автоматизированную платформу морбидостата 9.0009 7 , применение которого позволило оценить развитие устойчивости к антибиотикам (измеряемой как изменения чувствительности) в зависимости от состава нескольких лекарств, относительных концентраций и времени (дополнительная рис. 1). В морбидостате концентрация антибиотика автоматически контролировалась по пользовательскому алгоритму 7 ; при достижении бактериальной культурой определенной плотности на 12 мл бактериальной культуры вводили 1 мл питательной среды, содержащей антибиотик, а при низкой плотности клеток добавляли среду, не содержащую антибиотик (Методы). Концентрация антибиотика во вводимой питательной среде как минимум в 10 раз превышала минимальную ингибирующую концентрацию (МПК) штамма дикого типа. Как правило, антибиотики добавляли по крайней мере несколько раз после того, как плотность клеток превышала пороговый уровень, и, таким образом, концентрацию антибиотика повышали до диапазона концентраций лекарственного средства, обеспечивающего отбор мутантов, устойчивых к лекарственным препаратам (так называемое «окно селекции мутантов» 9).0009 33 ). Использование автоматизированной системы культивирования клеток было важно не только для обеспечения программируемой дозировки, но и система морбидостата позволяла поддерживать бактериальные популяции в фазе экспоненциального роста. Таким образом, ожидалось, что платформа обеспечит хорошую фенотипическую воспроизводимость между параллельными экспериментами 7 , в то же время позволяя генетически разнообразным популяциям сосуществовать 34,35 . Обратите внимание, что здесь мы определили штамм Escherichia coli MG1655 дикого типа как восприимчивый штамм, в отличие от клинического определения, согласно которому штамм можно лечить антибиотиком в рекомендуемой дозировке.
Концентрация антибиотика во вводимой питательной среде как минимум в 10 раз превышала минимальную ингибирующую концентрацию (МПК) штамма дикого типа. Как правило, антибиотики добавляли по крайней мере несколько раз после того, как плотность клеток превышала пороговый уровень, и, таким образом, концентрацию антибиотика повышали до диапазона концентраций лекарственного средства, обеспечивающего отбор мутантов, устойчивых к лекарственным препаратам (так называемое «окно селекции мутантов» 9).0009 33 ). Использование автоматизированной системы культивирования клеток было важно не только для обеспечения программируемой дозировки, но и система морбидостата позволяла поддерживать бактериальные популяции в фазе экспоненциального роста. Таким образом, ожидалось, что платформа обеспечит хорошую фенотипическую воспроизводимость между параллельными экспериментами 7 , в то же время позволяя генетически разнообразным популяциям сосуществовать 34,35 . Обратите внимание, что здесь мы определили штамм Escherichia coli MG1655 дикого типа как восприимчивый штамм, в отличие от клинического определения, согласно которому штамм можно лечить антибиотиком в рекомендуемой дозировке. Устойчивые штаммы определяли как любые (эволюционировавшие) бактериальные образцы, МПК которых превышали МИК штамма дикого типа.
Устойчивые штаммы определяли как любые (эволюционировавшие) бактериальные образцы, МПК которых превышали МИК штамма дикого типа.
( a ) Схема эксперимента по эволюции бактерий при циклическом воздействии двух антибиотиков. E. coli дикого типа культивировали в системе морбидостата с динамическими концентрациями антибиотика. Каждые 90 353 t 90 354 дня (где 90 353 t 90 354 = 1, 3 и 6) препараты меняли попеременно, а образцы клеточных культур ежедневно отбирали и хранили при -80°C. Всего через 12 или 24 дня определяли МИК собранных образцов. ( b ) Эволюционные траектории устойчивости к KAN и POL (зеленая и желтая кривые соответственно) в условиях применения одного антибиотика. Траектории параллельных экспериментов были показаны бледными линиями, а среднее значение параллельных экспериментов было сплошной линией с пустыми кружками.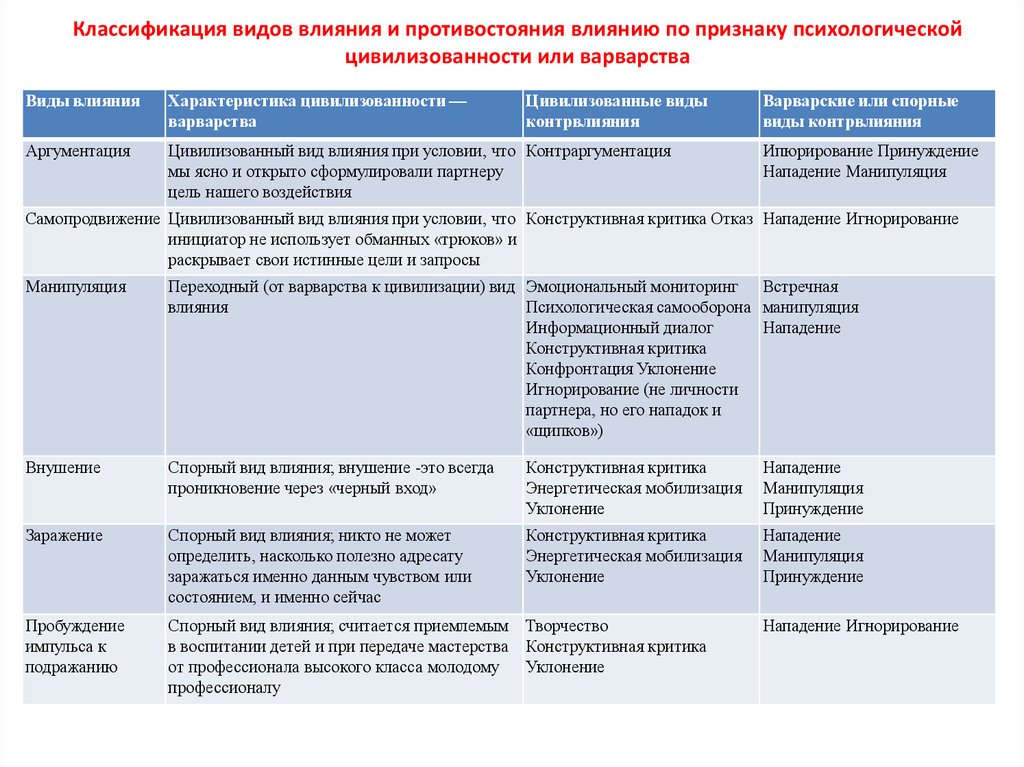 Уровни устойчивости измеряли как кратное изменение МИК по сравнению с диким типом, рассчитанное по log 2 (МИК и /МИК WT ), где МИК i и МИК WT представляют собой МИК образцов и -го дня и E. coli дикого типа соответственно. ( c ) Траектории сопротивления KAN и POL при циклировании с 3-дневным интервалом. Бледные линии указывают траектории параллельных экспериментов, а среднее значение показано сплошной линией с закрашенными кружками. Бледный фон указывает на используемые антибиотики. ( d , e ) эволюционные траектории устойчивости к CHL и POL (синий и желтый) в условиях одного антибиотика и при циклировании с 3-дневным интервалом соответственно. Аналогично, для ( f , g ) устойчивость к CHL и NIT (синий и фиолетовый) и ( h ) и ( i ) устойчивость к KAN и NAL (зеленый и оранжевый). Размер выборки составляет n= 3 для состояния с одним антибиотиком и n= 2 для циклического применения антибиотиков (биологические дубликаты).
Уровни устойчивости измеряли как кратное изменение МИК по сравнению с диким типом, рассчитанное по log 2 (МИК и /МИК WT ), где МИК i и МИК WT представляют собой МИК образцов и -го дня и E. coli дикого типа соответственно. ( c ) Траектории сопротивления KAN и POL при циклировании с 3-дневным интервалом. Бледные линии указывают траектории параллельных экспериментов, а среднее значение показано сплошной линией с закрашенными кружками. Бледный фон указывает на используемые антибиотики. ( d , e ) эволюционные траектории устойчивости к CHL и POL (синий и желтый) в условиях одного антибиотика и при циклировании с 3-дневным интервалом соответственно. Аналогично, для ( f , g ) устойчивость к CHL и NIT (синий и фиолетовый) и ( h ) и ( i ) устойчивость к KAN и NAL (зеленый и оранжевый). Размер выборки составляет n= 3 для состояния с одним антибиотиком и n= 2 для циклического применения антибиотиков (биологические дубликаты).
Увеличить
Бактериальная эволюция при чередующихся стрессах антибиотиков
Начиная с изогенного штамма E. coli , была проведена серия 24-дневных экспериментов в системе морбидостата с двумя чередующимися стрессами антибиотиков (рис. 1а), а также 12-дневные опыты в условиях однократного антибиотикостресса. Антибиотики были выбраны из разных классов, чтобы они имели разные мишени (таблица 1), что сводит к минимуму риск развития потенциальной перекрестной резистентности между антибиотиками 12,13,29 . Чтобы оценить воспроизводимость эволюционных траекторий, эксперименты были продублированы для каждой пары циклов антибиотиков и трижды для каждого отдельного антибиотика. Эксперименты с обратным порядком циклирования антибиотиков также проводились в двух экземплярах, чтобы оценить влияние порядка циклирования. Эти эксперименты показали, что бактериальные популяции демонстрировали две эволюционные закономерности в развитии лекарственной устойчивости в циклических условиях, как показано на рис. 1. Первая группа развила устойчивость только к одному из циклированных антибиотиков, тогда как вторая группа стала устойчивой к обоим. антибиотики. Мы называем эти устойчивые к антибиотикам состояния бактериальных популяций «состоянием устойчивости к одному лекарству» и «множественной лекарственной устойчивостью» соответственно. Во второй категории наблюдались два различных паттерна: Колебание 30 с постепенным увеличением базовой линии (рис. 1g) и снижением максимального сопротивления (рис. 1i). В первом случае скорость адаптации к хлорамфениколу (ХЛ) и нитрофурантоину (НИТ) была задержана (крутизна подобранных логистических кривых k =0,4±0,03 и 0,19±0,01 день -1 , среднее ± стандартная ошибка среднего, соответственно) по сравнению со случаем однократного антибиотического стресса (рис. 1е, к =0,79±0,13 и 0,55±0,05 сут -1 соответственно). В последнем случае лекарственная устойчивость развивалась быстро и с той же скоростью, что и в случае с одним антибиотиком 9.
1. Первая группа развила устойчивость только к одному из циклированных антибиотиков, тогда как вторая группа стала устойчивой к обоим. антибиотики. Мы называем эти устойчивые к антибиотикам состояния бактериальных популяций «состоянием устойчивости к одному лекарству» и «множественной лекарственной устойчивостью» соответственно. Во второй категории наблюдались два различных паттерна: Колебание 30 с постепенным увеличением базовой линии (рис. 1g) и снижением максимального сопротивления (рис. 1i). В первом случае скорость адаптации к хлорамфениколу (ХЛ) и нитрофурантоину (НИТ) была задержана (крутизна подобранных логистических кривых k =0,4±0,03 и 0,19±0,01 день -1 , среднее ± стандартная ошибка среднего, соответственно) по сравнению со случаем однократного антибиотического стресса (рис. 1е, к =0,79±0,13 и 0,55±0,05 сут -1 соответственно). В последнем случае лекарственная устойчивость развивалась быстро и с той же скоростью, что и в случае с одним антибиотиком 9.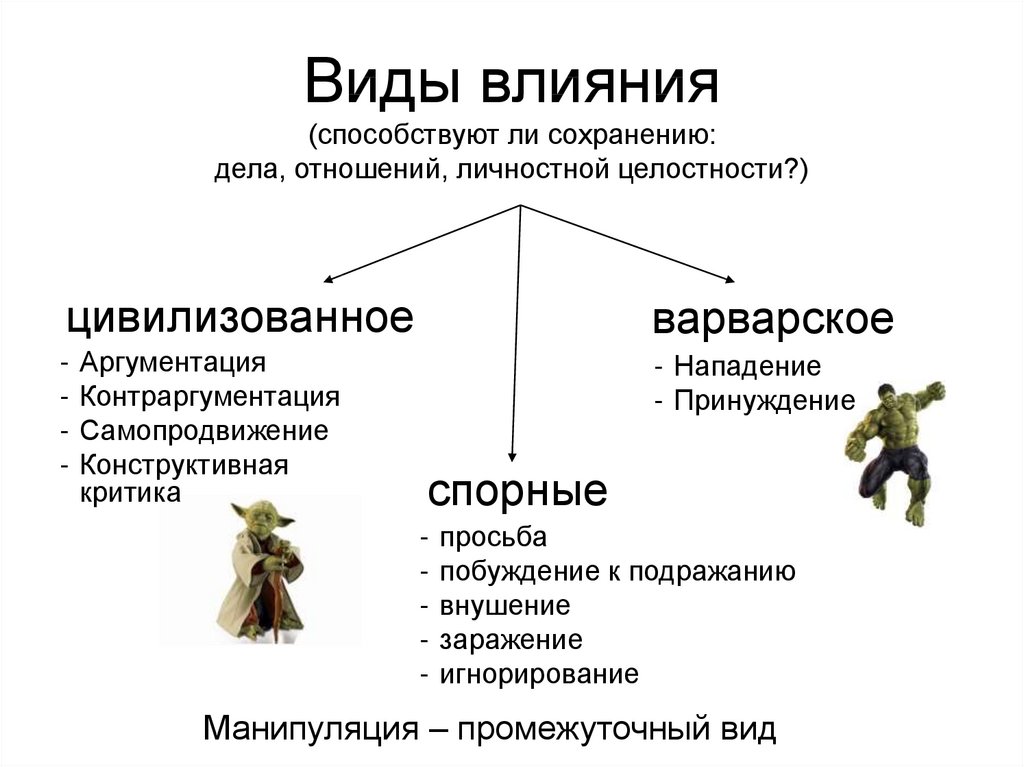 0353 k =1,7±0,12 и 31,9±3,12 дня -1 для канамицина (КАН) и налидиксовой кислоты (НАЛ), соответственно, для циклического случая и k =7,11±6,01 и 5,06±1,65 дня -1 на один случай антибиотика. Однако максимальный уровень устойчивости к NAL был значительно снижен ( P <0,001, два образца Уэлча t — тест. Дополнительный рисунок 4a), в то время как уровень устойчивости к KAN был сопоставим со случаем с одним антибиотиком. Эволюционные траектории воспроизводились между параллельными экспериментами, и эксперименты по эволюции с обратным порядком циклирования антибиотиков показали аналогичные эволюционные закономерности (дополнительная рис. 2a).
0353 k =1,7±0,12 и 31,9±3,12 дня -1 для канамицина (КАН) и налидиксовой кислоты (НАЛ), соответственно, для циклического случая и k =7,11±6,01 и 5,06±1,65 дня -1 на один случай антибиотика. Однако максимальный уровень устойчивости к NAL был значительно снижен ( P <0,001, два образца Уэлча t — тест. Дополнительный рисунок 4a), в то время как уровень устойчивости к KAN был сопоставим со случаем с одним антибиотиком. Эволюционные траектории воспроизводились между параллельными экспериментами, и эксперименты по эволюции с обратным порядком циклирования антибиотиков показали аналогичные эволюционные закономерности (дополнительная рис. 2a).
Полноразмерная таблица
Хотя развитие множественной лекарственной устойчивости ожидалось, результаты циклирования с полимиксином B (POL), показанные на рис. 1c,e, оказались неожиданными.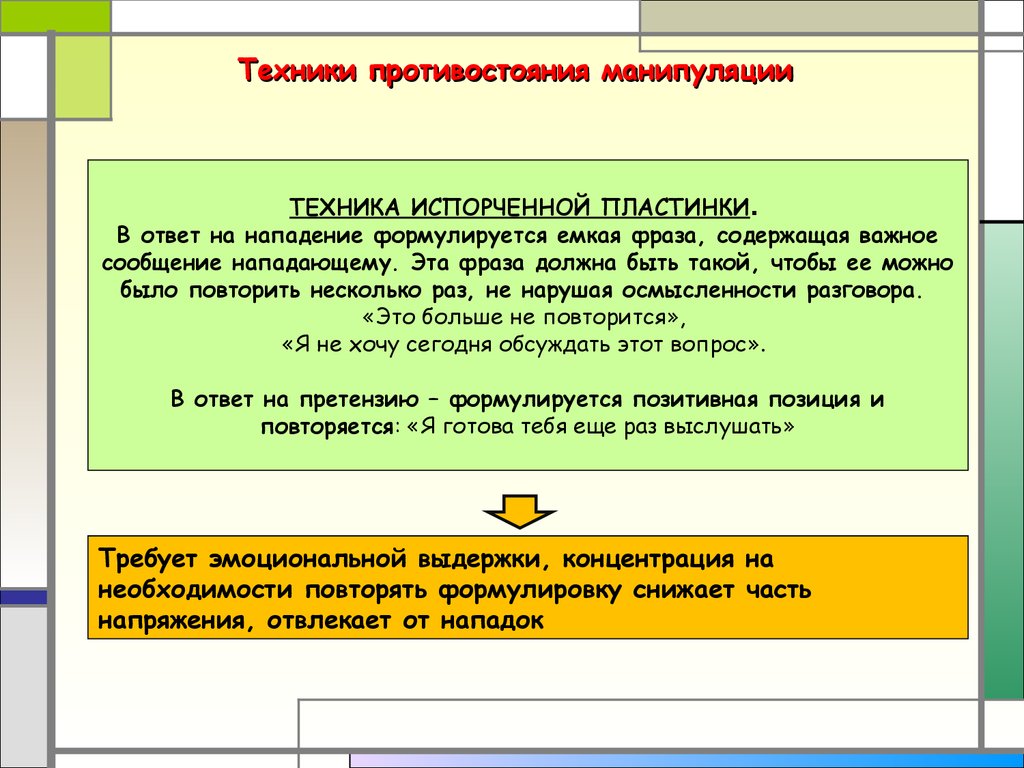 Это связано с тем, что в зависимости от комбинации антибиотиков подавлялось либо развитие резистентности к ПОЛ, либо противодействующий антибиотик. В результате бактериальная популяция была устойчива только к одному антибиотику. Напротив, резистентность развивалась во всех случаях при культивировании в условиях стресса с одним антибиотиком (рис. 1б, г, е, з). Чтобы лучше понять это поведение, бактериальная популяция подвергалась воздействию различных комбинаций антибиотиков (дополнительная рис. 3). Здесь антибиотики чередовались с интервалом в 1 день, потому что более короткие интервалы имели тенденцию задерживать развитие устойчивости к антибиотикам (дополнительная рис. 4b – i). Результаты показали, что циклическое использование POL часто приводило бактериальную популяцию к состоянию устойчивости к одному лекарству или удерживало популяцию в этом состоянии в течение длительного периода (дополнительная рис. 3a). Напротив, чередование с другими антибиотиками во всех случаях приводило к множественной лекарственной устойчивости (дополнительная рис.
Это связано с тем, что в зависимости от комбинации антибиотиков подавлялось либо развитие резистентности к ПОЛ, либо противодействующий антибиотик. В результате бактериальная популяция была устойчива только к одному антибиотику. Напротив, резистентность развивалась во всех случаях при культивировании в условиях стресса с одним антибиотиком (рис. 1б, г, е, з). Чтобы лучше понять это поведение, бактериальная популяция подвергалась воздействию различных комбинаций антибиотиков (дополнительная рис. 3). Здесь антибиотики чередовались с интервалом в 1 день, потому что более короткие интервалы имели тенденцию задерживать развитие устойчивости к антибиотикам (дополнительная рис. 4b – i). Результаты показали, что циклическое использование POL часто приводило бактериальную популяцию к состоянию устойчивости к одному лекарству или удерживало популяцию в этом состоянии в течение длительного периода (дополнительная рис. 3a). Напротив, чередование с другими антибиотиками во всех случаях приводило к множественной лекарственной устойчивости (дополнительная рис. 3b).
3b).
Бактериальная эволюция в условиях стресса с одним антибиотиком
Мы предположили, что состояние одиночной лекарственной устойчивости при ПОЛ может быть связано с эволюционными паттернами резистентности в условиях одного антибиотика. Действительно, развитие резистентности к ПОЛ в условиях стресса от одного антибиотика демонстрирует уникальный эволюционный паттерн с относительно длительной «тихой фазой», когда у популяции не развивается лекарственная устойчивость (до 6 дней), за которой следует резкое увеличение, как показано на рис. Рис. 5. Таким образом, закономерности эволюции других профилей устойчивости к антибиотикам можно разделить на два типа: (1) быстрое развитие устойчивости к NAL, KAN и рифампицину, повышающееся до максимального уровня с небольшой фазой молчания или без нее, и ( 2) постепенное повышение резистентности к ХЛ, НИТ, ампициллину и тетрациклину с течением времени 7,8 . Циклическое применение ПОЛ с антибиотиками из первой категории приводит к подавлению резистентности к ПОЛ, в то время как у популяций резистентность к ПОЛ развивалась на относительно ранней стадии при циклическом применении антибиотиков из второй категории.
Профиль побочной чувствительности
Мы также исследовали профили перекрестной резистентности и побочной чувствительности 12,13,29 , используя образцы последнего дня из эволюционных экспериментов в условиях одного антибиотика (рис. 2). Это показало, что штаммы, устойчивые к ПОЛ, проявляли побочную чувствительность ко многим антибиотикам, указывая на то, что наличие устойчивости к ПОЛ делает бактериальные клетки более восприимчивыми к другим антибиотикам, чем штаммы дикого типа. С другой стороны, штаммы, которые уже развили устойчивость к другим антибиотикам, часто развивали перекрестную устойчивость к другим антибиотикам или оставались нейтральными. Хотя эти результаты в целом согласовывались с предыдущими исследованиями чувствительности к побочным эффектам (дополнительная рис. 12) 29,36 , некоторые побочные профили чувствительности (например, KAN) были менее выражены по сравнению с предыдущими случаями. Это может быть связано с различной силой отбора во время эволюции между предыдущими и нашими случаями, поскольку известно, что профиль побочной чувствительности/перекрестной устойчивости зависит от силы отбора 36 .
MIC образцов последнего дня из эволюционных экспериментов в условиях одного антибиотика были протестированы со всеми восемью антибиотиками. Синие и красные сетки указывают на то, что пара антибиотиков чувствительна к побочным эффектам и перекрестно резистентна соответственно. Числа в сетках log 2 — трансформируемый МИК кратность смены. Значения, указанные в матрице, были округлены. Исходные данные см. в дополнительной таблице 2.
Изображение полного размера
Возможные механизмы подавления лекарственной устойчивости
Эксперименты по эволюции нескольких и отдельных лекарств продемонстрировали различные закономерности эволюции в зависимости от типов и комбинаций антибиотиков. В то время как сообщалось о стратегиях цикла антибиотиков, которые уничтожали бактерии сублетальными дозами антибиотиков 10 , в нашем случае сохранялся экспоненциальный рост, но подавлялось появление устойчивости к антибиотикам (рис. 1c, e). Эти различные паттерны эволюционных траекторий резко контрастируют с предыдущими лабораторными исследованиями эволюции цикла антибиотиков, где постоянно наблюдалось монотонное увеличение лекарственной устойчивости 9 . Чтобы дать возможные объяснения разнообразным эволюционным моделям, мы сначала рассмотрим условия роста в предыдущих и наших исследованиях.
1c, e). Эти различные паттерны эволюционных траекторий резко контрастируют с предыдущими лабораторными исследованиями эволюции цикла антибиотиков, где постоянно наблюдалось монотонное увеличение лекарственной устойчивости 9 . Чтобы дать возможные объяснения разнообразным эволюционным моделям, мы сначала рассмотрим условия роста в предыдущих и наших исследованиях.
Метод последовательного переноса, использованный в предыдущем исследовании 9 , бактериальная популяция испытывает экспоненциальную и стационарную фазы во время культивирования в течение ночи. В то время как скорость роста является основным фактором эволюции устойчивости в экспоненциальной фазе, другие факторы, такие как межклеточное взаимодействие и конкуренция за ресурсы, будут играть роль в стационарной фазе. Таким образом, эволюция с использованием метода серийного переноса была бы медленнее, чем в случае сохранения экспоненциальной фазы (то есть морбидостата), поскольку бактериям необходимо адаптироваться как к условиям экспоненциальной, так и к стационарной фазам. Циклическое использование антибиотиков еще больше замедлит процесс адаптации из-за побочной чувствительности, дополнительных затрат на приспособление из-за устойчивости ко второму антибиотику и других факторов 9 .
Циклическое использование антибиотиков еще больше замедлит процесс адаптации из-за побочной чувствительности, дополнительных затрат на приспособление из-за устойчивости ко второму антибиотику и других факторов 9 .
Напротив, скорость эволюции резистентности в морбидостате в первую очередь определяется скоростью роста, в частности скоростью роста нового мутанта по сравнению с популяцией 8 . Это могло бы объяснить закономерности эволюции при стрессе с одним антибиотиком, поскольку скорость роста (то есть приспособленность) устойчивого к антибиотикам мутанта может варьироваться в зависимости от используемых антибиотиков. Например, если требуется только одна единственная мутация, чтобы стать сильной устойчивостью к антибиотику, такой устойчивый мутант быстро распространится по популяции в присутствии антибиотика, и резистентность на уровне популяции резко возрастет. С другой стороны, если улучшение приспособленности за счет мутации относительно невелико, эволюция будет медленно увеличиваться. Эти механизмы могут объяснить быстрое или постепенное усиление эволюционных паттернов (например, KAN или CHL, соответственно, дополнительная рис. 5).
Эти механизмы могут объяснить быстрое или постепенное усиление эволюционных паттернов (например, KAN или CHL, соответственно, дополнительная рис. 5).
Эволюция при однократном стрессе POL показала уникальную картину с относительно продолжительной молчаливой фазой, которая не наблюдалась при использовании других протестированных антибиотиков. Простым объяснением этой закономерности может быть предположение, что для устойчивости к ПОЛ необходимы множественные мутации. Однако это кажется неправдоподобным, поскольку воспроизводимые эволюционные траектории устойчивости к ПОЛ (например, рис. 1д) не могут быть объяснены с учетом присущей генетической мутации стохастичности. Кроме того, как описано ниже, данные последовательности всего генома не подтверждают объяснение (дополнительная таблица 1). Вместо этого мы предполагаем двухэтапный механизм адаптации устойчивости к ПОЛ, поскольку известно, что существуют негенетические механизмы устойчивости к ПОЛ, такие как модификация 9 липида А. 0009 37 и гетерорезистентность 38 . Недавние исследования показали, что гетерорезистентность к ПОЛ достигается за счет усиления синтеза путресцина и белка YceI без каких-либо генетических мутаций. Кроме того, высвобождение молекул из высокорезистентных субпопуляций гетерорезистентных бактерий защищает менее резистентные бактерии от ПОЛ и других стрессовых воздействий антибиотиков 39,40 . Эти данные свидетельствуют о том, что популяции E.coli в этом исследовании могли использовать негенетические механизмы для преодоления стресса ПОЛ на начальном этапе эволюции, когда концентрация препарата была низкой. Затем это могло переключиться на генетические механизмы, то есть полезные мутации, придающие устойчивость к ПОЛ по мере увеличения концентрации препарата, что может объяснить ступенчатый эволюционный характер устойчивости к ПОЛ.
0009 37 и гетерорезистентность 38 . Недавние исследования показали, что гетерорезистентность к ПОЛ достигается за счет усиления синтеза путресцина и белка YceI без каких-либо генетических мутаций. Кроме того, высвобождение молекул из высокорезистентных субпопуляций гетерорезистентных бактерий защищает менее резистентные бактерии от ПОЛ и других стрессовых воздействий антибиотиков 39,40 . Эти данные свидетельствуют о том, что популяции E.coli в этом исследовании могли использовать негенетические механизмы для преодоления стресса ПОЛ на начальном этапе эволюции, когда концентрация препарата была низкой. Затем это могло переключиться на генетические механизмы, то есть полезные мутации, придающие устойчивость к ПОЛ по мере увеличения концентрации препарата, что может объяснить ступенчатый эволюционный характер устойчивости к ПОЛ.
Основываясь на закономерностях эволюции отдельных лекарств, мы затем рассмотрим эволюционные закономерности цикличности антибиотиков, в частности цикличности KAN-POL (рис.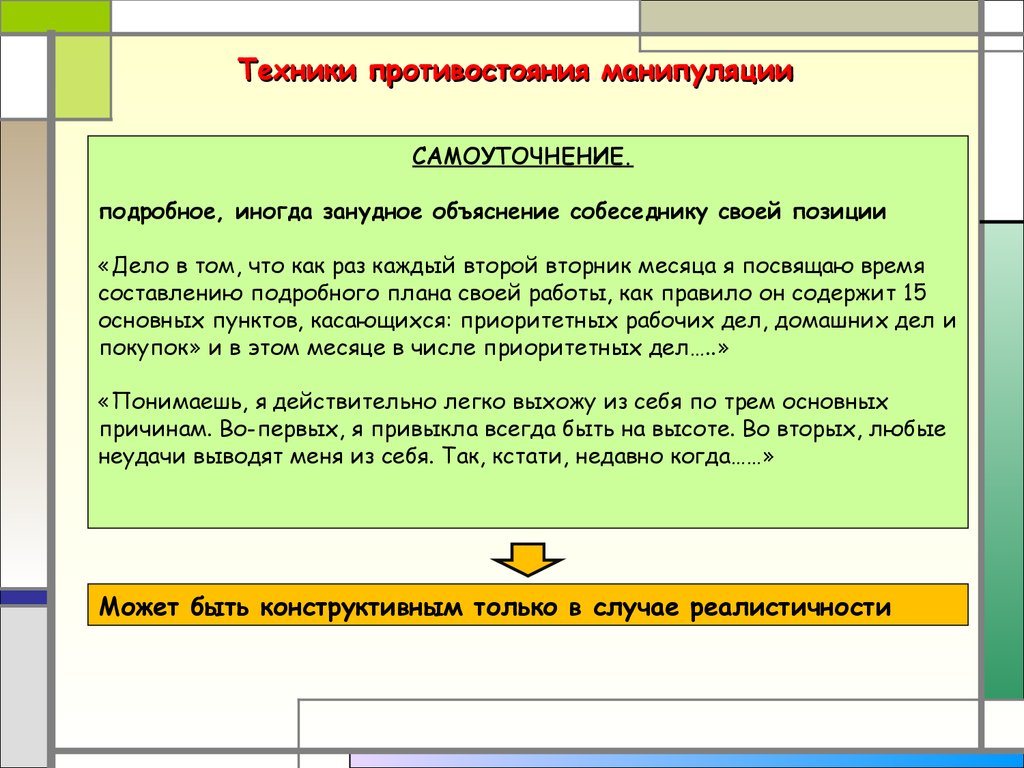 1c) и POL-CHL (рис. 1e). В первом случае бактериальная популяция может сначала приобрести устойчивость к KAN, поскольку это было показано быстро на начальном этапе эволюционных экспериментов (рис. 1б). Учитывая, что резистентность к KAN, как правило, влечет за собой затраты на приспособление, это может значительно задержать эволюцию, чтобы приобрести дополнительную устойчивость к POL за счет продления фазы молчания, особенно потому, что устойчивость к POL сопряжена с большими затратами на приспособление 37 . Действительно, в аналогичном случае с циклами NAL-POL (дополнительная рис. 3a) появление устойчивости к POL было в значительной степени отсрочено, что позволяет предположить, что устойчивость к POL появилась после того, как стоимость устойчивости к NAL была компенсирована вторичными адаптациями 41,42 . Это также предполагает, что, если циклирование KAN-POL продолжалось в течение более длительного периода, устойчивость к POL может проявиться на более позднем этапе эксперимента.
1c) и POL-CHL (рис. 1e). В первом случае бактериальная популяция может сначала приобрести устойчивость к KAN, поскольку это было показано быстро на начальном этапе эволюционных экспериментов (рис. 1б). Учитывая, что резистентность к KAN, как правило, влечет за собой затраты на приспособление, это может значительно задержать эволюцию, чтобы приобрести дополнительную устойчивость к POL за счет продления фазы молчания, особенно потому, что устойчивость к POL сопряжена с большими затратами на приспособление 37 . Действительно, в аналогичном случае с циклами NAL-POL (дополнительная рис. 3a) появление устойчивости к POL было в значительной степени отсрочено, что позволяет предположить, что устойчивость к POL появилась после того, как стоимость устойчивости к NAL была компенсирована вторичными адаптациями 41,42 . Это также предполагает, что, если циклирование KAN-POL продолжалось в течение более длительного периода, устойчивость к POL может проявиться на более позднем этапе эксперимента. Последний случай циклирования CHL-POL, при котором устойчивость к CHL была полностью подавлена, можно объяснить обратимым характером устойчивости к CHL (что также наблюдалось при устойчивости к POL, как описано ниже). На дополнительном рисунке 2 было замечено, что развившаяся резистентность к CHL возвращалась обратно к уровню восприимчивости, когда стресс от антибиотиков был переключен на POL. Это указывает на то, что резистентные к ХЛ мутанты, появившиеся в период стресса ХЛ, погибали при замене антибиотика на ПОЛ. Если бы скорость циклирования была более частой, уровень устойчивости к CHL оставался бы низким, потому что мутанты, устойчивые к CHL, были бы удалены до того, как они стали бы большинством в популяции.
Последний случай циклирования CHL-POL, при котором устойчивость к CHL была полностью подавлена, можно объяснить обратимым характером устойчивости к CHL (что также наблюдалось при устойчивости к POL, как описано ниже). На дополнительном рисунке 2 было замечено, что развившаяся резистентность к CHL возвращалась обратно к уровню восприимчивости, когда стресс от антибиотиков был переключен на POL. Это указывает на то, что резистентные к ХЛ мутанты, появившиеся в период стресса ХЛ, погибали при замене антибиотика на ПОЛ. Если бы скорость циклирования была более частой, уровень устойчивости к CHL оставался бы низким, потому что мутанты, устойчивые к CHL, были бы удалены до того, как они стали бы большинством в популяции.
Математическая модель бактериальной мультилекарственной эволюции
Исходя из приведенных выше результатов, мы предположили, что три фактора были вовлечены в эволюционные паттерны при циклическом стрессе антибиотиков, а именно: (1) продолжительность молчаливой фазы, (2) скорость адаптация и (3) профили перекрестной резистентности и побочной чувствительности. Чтобы изучить полученные результаты с более широкой точки зрения, мы разработали простую модель, включающую три фактора для описания эволюционных паттернов (дополнительные методы). Следует отметить, что мы использовали феноменологическую модель, а не общепринятые модели популяционной генетики 8,42,43 . Это связано с тем, что предыдущие исследования показали, что, несмотря на различные лежащие в основе генетические изменения, бактериальная эволюция устойчивости к стрессу может демонстрировать удивительно сходные фенотипические траектории 7,8,44,45,46 . Кроме того, с точки зрения прогнозирования было бы полезно, если бы модель основывалась на экспериментальных параметрах, которые можно относительно легко измерить. Таким образом, здесь мы разработали теоретическую модель, основанную только на относительных уровнях МПК.
Чтобы изучить полученные результаты с более широкой точки зрения, мы разработали простую модель, включающую три фактора для описания эволюционных паттернов (дополнительные методы). Следует отметить, что мы использовали феноменологическую модель, а не общепринятые модели популяционной генетики 8,42,43 . Это связано с тем, что предыдущие исследования показали, что, несмотря на различные лежащие в основе генетические изменения, бактериальная эволюция устойчивости к стрессу может демонстрировать удивительно сходные фенотипические траектории 7,8,44,45,46 . Кроме того, с точки зрения прогнозирования было бы полезно, если бы модель основывалась на экспериментальных параметрах, которые можно относительно легко измерить. Таким образом, здесь мы разработали теоретическую модель, основанную только на относительных уровнях МПК.
Модель была основана на двух предположениях: (1) Развитие устойчивости бактерий к антибиотику A , именуемый R A , постоянно активируется при воздействии антибиотика A . Однако уровень сопротивления в конце концов насыщается из-за затрат на фитнес 47 , как мы видели выше. (2) Увеличение R A отрицательно влияет на устойчивость к другому антибиотику B или R B в значительной степени, если антибиотики представляют собой побочную чувствительную пару
Однако уровень сопротивления в конце концов насыщается из-за затрат на фитнес 47 , как мы видели выше. (2) Увеличение R A отрицательно влияет на устойчивость к другому антибиотику B или R B в значительной степени, если антибиотики представляют собой побочную чувствительную пару
9, 29, 12,12
. В противном случае это имеет небольшой эффект (нейтральная или кросс-резистентная пара). Концептуально модель можно описать как систему с положительной саморегуляцией и двойными петлями отрицательной обратной связи (рис. 3а). Модель имеет три ключевых параметра: продолжительность фазы молчания θ , скорость адаптации α и коэффициент побочной чувствительности/перекрестного сопротивления β. Эти параметры модели были определены на основе подгонки параметров экспериментальных результатов при однократном стрессе антибиотика. Рисунок 3: Теоретические модели развития множественной лекарственной устойчивости. ( a ) Схематическая диаграмма модели, состоящей из положительной саморегуляции и двойной отрицательной обратной связи. ( b ) Фазовое пространство эволюционной динамики нескольких лекарств с POL и KAN. Два сплошных круга обозначают стабильные устойчивые состояния, соответствующие состояниям с одиночной и множественной лекарственной устойчивостью соответственно. Разорванный круг — это нестабильное устойчивое состояние. МИК были нормализованы к единице по максимальной МИК эволюции одного антибиотика. ( c ) Фазовое пространство эволюционной динамики множественных лекарств с НИТ и ХЛ. В этом случае существует только одно стабильное устойчивое состояние, состояние множественной лекарственной устойчивости. ( d – f ) Смоделированные эволюционные траектории устойчивости бактерий с POL и KAN, POL и NIT и CHL и NIT соответственно. Антибиотики циклировали с интервалом в 1 день. ( г – i ) Экспериментальные данные, полученные из системы морбидостата.
Изображение в полный размер
Геометрическая структура модели, показанной на рис. 3b,c, объясняет происхождение уникальных закономерностей эволюции устойчивости к ПОЛ. Нулевые линии сопротивления POL (желтая кривая на рис. 3b) и сопротивления KAN (зеленая) пересекаются в трех точках, две из которых являются устойчивыми стационарными состояниями (обозначены сплошными кружками). Таким образом, теоретически эволюция бактериальной резистентности к POL и KAN может привести либо к полирезистентному, либо к одиночному лекарственно-устойчивому состоянию. Однако в случае циклирования с KAN у бактериальной популяции сначала развилась резистентность к KAN из-за быстрой скорости адаптации (определяемой по α ), а резистентность к ПОЛ вообще не развивалась, несмотря на циклическое применение антибиотиков (рис. 3d,g). Напротив, когда POL циклировали с антибиотиком с медленной скоростью адаптации, таким как NIT, эволюция заканчивается мультирезистентным состоянием (рис. 3e, h). Это связано с тем, что резистентность к ПОЛ быстро увеличивалась до того, как система достигла состояния одного антибиотика из-за медленной скорости адаптации НИТ. Геометрическая структура уникальна для случая с POL, который имеет большую тихую фазу (определяемую θ ). Напротив, нульклины имеют только одно пересечение при циклическом применении антибиотиков, отличных от ПОЛ (рис. 3с). Это означает, что эволюция при циклировании антибиотиков всегда приводит к состоянию множественной лекарственной устойчивости в этой системе (рис. 3f, i). Действительно, циклические эксперименты с использованием антибиотиков, отличных от POL, всегда приводили к состоянию множественной лекарственной устойчивости, насколько мы тестировали (дополнительная рис. 3b).
Эволюционные траектории, смоделированные моделью, хорошо согласовывались с экспериментальными результатами с точки зрения конечных состояний лекарственной устойчивости (рис. 4), а также закономерностей эволюции (дополнительная рис. 6). На рис. 4а, б нормированный уровень лекарственной устойчивости для одного из антибиотиков, включенных в цикл, был нанесен по сравнению с другим антибиотиком. Например, цикличность KAN-POL, где KAN и POL представлены как препараты № 1 и № 2 соответственно, была нанесена на карту на x , потому что была развита только устойчивость к KAN, в то время как устойчивость к POL была полностью подавлена. В целом результаты можно разделить на две группы, то есть состояния с одиночной и множественной лекарственной устойчивостью, как отмечалось выше. Первые случаи (например, КАН-ПОЛ) откладывались по осям, а вторые (например, НАЛ-КАН) — в верхней правой части графика. Чтобы проиллюстрировать предсказуемость теоретической модели, нормализованные предсказанные значения MIC были нанесены на график относительно нормализованных наблюдаемых значений MIC (рис. 4c, d). В целом предсказания модели показали хорошую корреляцию с экспериментальными результатами ( r 2 =0,44 и 0,64 соответственно). Мы также построили нулевую модель математической модели в качестве сравнения. В нулевой модели все коэффициенты побочной чувствительности/перекрестного сопротивления β были установлены равными нулю. Сравнение прогнозов нулевой модели и экспериментальных результатов показало плохую корреляцию ( r 2 = 0,03 и 0,20. Дополнительный рис. 9). Поскольку в нулевой модели нет взаимодействия между устойчивостью к антибиотикам из-за β =0, оба уровня лекарственной устойчивости приближались к максимуму со скоростью, определяемой α . Этот результат показывает, что взаимодействие между α и β имеет решающее значение для различных эволюционных моделей и, следовательно, демонстрирует предсказуемость математической модели.
( a ) Диаграмма рассеяния наблюдаемых МИК первого препарата по сравнению со вторым препаратом из выборок последних дней циклических экспериментов с интервалом в 1 день. Обратите внимание, что метка для каждой точки указывает порядок цикла антибиотиков. Например, KAN-POL означает, что KAN использовался в первый день (обозначен как «Препарат № 1» на этикетке оси), а POL — во второй день («Препарат № 2»), а затем циклически повторялся с заказом. Из-за этого обозначения КАН-ПОЛ и ПОЛ-КАН нанесены на x и y осей соответственно, хотя в обоих случаях развивалась только устойчивость к KAN (дополнительный рис. 3a). ( b ) Диаграмма рассеяния предсказанных MIC от конечной временной точки смоделированных результатов. Обратите внимание, что в результаты был включен эффект снижения затрат на фитнес для циклов NAL и POL. ( c , d ) Диаграммы рассеяния предсказанных МИК против наблюдаемых МИК для первого и второго лекарственного средства соответственно. Обратите внимание, что те же данные, что и для 9Здесь используются 0405 a и b .
Изображение в натуральную величину
Следует также отметить, что в некоторых случаях модель и эксперименты не совпадали. Например, цикл CHL-POL привел к устойчивости к одному лекарству (только устойчивость к POL), в то время как модель предсказывала, что бактериальная популяция приведет к состояниям с множественной лекарственной устойчивостью (дополнительные рисунки 3a и 7a). Как обсуждалось в предыдущем разделе, дополнительный эффект бактериальной резистентности, такой как обратимая резистентность к CHL, может быть связан с экспериментальной моделью цикла антибиотиков и должен быть включен в модель для точного воспроизведения поведения. В случае циклирования с использованием НАЛ и ПОЛ экспериментальные результаты показали, что резистентность к ПОЛ появилась на более позднем этапе эксперимента. Однако теоретическая модель предсказывала одно состояние лекарственной устойчивости (только резистентность к NAL). Это говорит о том, что имел место какой-то дополнительный адаптационный процесс, снижающий затраты на приспособление к резистентности к NAL, возникшей во время эксперимента с циклами (например, компенсаторные мутации 9). 0009 41,42 ). Действительно, модель воспроизводила экспериментальные результаты, когда такой эффект учитывался путем уменьшения коэффициента побочной чувствительности / перекрестного сопротивления β до нуля в определенный момент во время эксперимента (дополнительный рис. 6b и дополнительные методы).
Получено приближенное аналитическое решение модели, которое описывает, что нормированная максимальная устойчивость к антибиотику A определяется как в простейшей форме (Дополнительные методы). Это означает, что максимальный уровень устойчивости определяется соотношением профиля перекрестной устойчивости/побочной чувствительности и скорости адаптации. Такой эффект α и β действительно наблюдались в экспериментальных результатах (дополнительная рис. 3): циклическое применение антибиотиков с побочно-чувствительной парой (то есть большой β ) снижало максимальный уровень устойчивости по сравнению с уровнем с перекрестно-резистентной парой (маленький β ), например, пары CHL и KAN и CHL и NAL соответственно. Уравнение также показывает, что если антибиотик A с большим α циклически повторяется с антибиотиком B с малым α 903:54 эволюция заканчивается в последний день, что подтверждается экспериментальными результатами.
Обращение вспять лекарственной устойчивости путем модуляции стресса антибиотиков
Одним из важных следствий этой модели является то, что эволюцию бактериальной лекарственной устойчивости можно рассматривать как динамическую систему, поскольку предложенная здесь модель представляет собой простую детерминированную систему. Такой динамический системный взгляд на биологические системы был ранее предложен в контексте дифференцировки стволовых клеток и эволюции лекарственной устойчивости раковых клеток 48,49 . В современном контексте эта точка зрения предполагает, что лекарственно-устойчивые состояния бактериальной популяции могут быть направлены из одного состояния в другое, если стрессы антибиотиков модулируются извне. На самом деле теоретически было показано, что циклическое применение антибиотиков с парой синергетических препаратов может отбирать чувствительные бактерии, уничтожая лекарственно-устойчивые из популяции 20 . Чтобы проверить это экспериментально, мы исследовали обратимость развившейся устойчивости к антибиотикам, поскольку ее можно рассматривать как переход между состояниями устойчивости. В частности, здесь мы в основном сосредоточились на резистентности к ПОЛ, потому что циклическое применение этого антибиотика имеет два состояния резистентности (рис. 3b), а также потому, что он является одним из «последних» антибиотиков 37 . Модель предсказывала, что развившуюся резистентность можно устранить из бактериальной популяции, если цикличность антибиотиков переключится на одиночный антибиотический стресс (дополнительная рис. 7a). Чтобы подтвердить это экспериментально, мы заразили бактериальные популяции из циклических экспериментов последовательным воздействием противодействующих антибиотиков. Результаты показали, что резистентность действительно можно обратить вспять даже с последнего дня цикла антибиотикотерапии за счет повышения резистентности к противодействующему антибиотику (рис. 5а). Следует также отметить, что не только одиночная, но и множественная лекарственная устойчивость в популяции бактерий может быть устранена путем воздействия противодействующего антибиотика (случай POL-NIT на дополнительной рис. 7b). Обратимость не зависела от развившейся резистентности или противодействия применяемым антибиотикам (дополнительные рисунки 2b и 7b). Однако были также случаи, когда устойчивость к антибиотикам не обращалась, особенно в случаях, когда популяция развивалась в условиях одного стресса от антибиотика (дополнительная рис. 7c).
Мы изучили механизм развития резистентности и обратимости и определили условия, которые способствовали этому явлению: Генетическая и фенотипическая гетерогенность в бактериальной популяции. Во-первых, мы секвенировали полный геном устойчивых к ПОЛ мутантов, развившихся в условиях одиночного или циклического стресса ПОЛ. Хотя в эволюционировавших штаммах был обнаружен ряд мутаций (дополнительная таблица 1), мы идентифицировали мутации в генах basS и secD как ключевые мутации, которые придают устойчивость к POL (рис. 5b и дополнительная рис. 8). Двухкомпонентная система BasSR (также называемая PmrAB) регулирует пути модификации липополисахарида и, как известно, придает устойчивость к ПОЛ 37 . SecD, часть белково-транслоказного комплекса Sec, принадлежит к семейству экспортеров множественных лекарственных препаратов 50 , относящемуся к семейству резистентных клубеньков и делений клеток, и известно, что он играет вспомогательную роль в устойчивости к противомикробным пептидам 51 . Результаты секвенирования показали, что изменение устойчивости к антибиотикам возможно при наличии генетических вариаций в гене basS или secD в популяции (рис. 5b и дополнительная рис. 8). Напротив, в генетически однородной популяции по аллель basS или secD (дополнительная рис. 8). Этот результат свидетельствует о том, что циклическое применение антибиотиков предотвратило селективную смену и, таким образом, сохранило генетическую гетерогенность в популяции даже после 24-дневного воздействия антибиотиков. Этот результат подтверждается тем фактом, что популяции, развившиеся в условиях однократного стресса антибиотиками, не демонстрировали обратного развития лекарственной устойчивости, поскольку популяция была генетически однородной. Это генетическое разнообразие отразилось и на уровне фенотипа. Наблюдаемые нами МИК отдельных изолятов были разнообразны и отличались от МИК всей популяции (черные кружки и крестик на рис. 5в) 34,35 . Эти генетические и фенотипические различия могут частично объяснить изменение развившейся резистентности, потому что воздействие противодействующего антибиотика отбирает менее устойчивые к POL и более устойчивые к CHL клетки из-за побочной чувствительности.
( a ) Эволюционные траектории устойчивости к ПОЛ и КХЛ (желтый и синий соответственно). Цвета фона указывают на используемые антибиотики. ( b ) Частоты аллелей гена basS . Цифры на карте указывают частоту мутантов в популяциях, оцененную на основе анализа последовательности генов. ( c ) Диаграмма рассеяния МИК отдельных изолятов на 24-й и 30-й день (черные и красные кружки соответственно). Размер выборки составляет 90 353 n = 90 354 70 для обоих случаев. Более темный цвет соответствует более высокой частоте. MIC всей популяции показан черным или красным крестом соответственно.
Полноразмерное изображение
Обсуждение
Хотя циклическое применение антибиотиков ранее считалось менее эффективным, чем комбинированное лечение антибиотиками 26,27 , недавние теоретические и экспериментальные исследования показывают, что, напротив, использование побочной чувствительности различных антибиотиков является ключом к разработке эффективных методы лечения, которые могут подавить или обратить вспять лекарственную устойчивость 9,10,20 . Недавние исследования показывают, что можно вернуть устойчивость бактерий к одному антибиотику, используя антибиотики во временном разделении 10,29,52 . Хотя побочная чувствительность была известна более 60 лет 53 , важность этого явления для подавления развития лекарственной устойчивости не была хорошо осознана до недавнего времени 54 , что может объяснить неудачные клинические исследования или исследования на животных по цикличности антибиотиков. В этом исследовании мы провели серию лабораторных экспериментов по эволюции для экспериментального исследования эффекта чередования антибиотиков. Наши результаты показывают, что несколько комбинаций лекарств могут ингибировать эволюционные пути к множественной лекарственной устойчивости, полностью подавляя развитие бактериальной устойчивости к одному из препаратов, включенных в цикл. Кроме того, теоретическая модель, которую мы разработали, показала, что эволюция бактериальных популяций против множественной лекарственной устойчивости может быть не только предсказана, но и обращена вспять путем модулирования стресса отбора окружающей среды, и это было успешно продемонстрировано экспериментально. Представленная здесь модель является общей моделью эволюции многих лекарств, поэтому она не ограничивается конкретными комбинациями лекарств. Поскольку модельные предсказания множественной лекарственной устойчивости основаны на закономерностях эволюции при стрессе от одного лекарственного средства, модель потенциально может быть использована для предсказания эволюции других множественных лекарств, если доступны данные об эволюции одного лекарства. Таким образом, поскольку эксперименты по эволюции отдельных лекарств требуют меньше испытаний, чем комбинированные испытания, применение модели может предложить способ поиска эффективных комбинаций антибиотиков для циклирования на основе небольшого числа экспериментов.
Наши результаты показывают, что сниженная эффективность антибиотика может быть восстановлена путем модуляции стресса, вызванного антибиотиками. Однако следует соблюдать осторожность при переводе этих результатов в клинические условия, учитывая, что в наших экспериментах здесь учитывались только хромосомные мутации de novo . Дальнейшие эксперименты, такие как эволюционные эксперименты с устойчивостью к антибиотикам, опосредованной плазмидами, потребуются для выяснения возможности подавления резистентности циклическим циклированием антибиотиков в более практических ситуациях. Эволюционные эксперименты с различными режимами дозирования с чередованием антибиотиков могут стать еще одним возможным путем для лучшего понимания устойчивости бактерий к антибиотикам. В отличие от системы морбидостата, где основное внимание уделяется отбору мутантов, Динамическая модель in vitro имитирует фармакокинетические профили антибиотиков 55 , что может дать полезную информацию об эффекте циклического применения антибиотиков, в частности пар, чувствительных к побочным эффектам, и о том, как может развиваться лекарственная устойчивость in vivo . Кроме того, эксперименты по циклированию антибиотиков с использованием побочной чувствительности также могут быть распространены на модели животных. Модель тканевой клетки 56 , например, позволит проводить постоянный отбор проб бактериальных популяций, а также концентрацию лекарственного средства во время чередующегося лечения антибиотиками. Параллельно с этими возможными дальнейшими экспериментами мы предвидим, что более глубокое понимание динамики популяции в условиях приема нескольких лекарств и улучшенное теоретическое моделирование потенциально устранят трудоемкие комбинаторные эксперименты с несколькими лекарствами. Взятые вместе, они могут привести к новым методам лечения, которые устранят множественную лекарственную устойчивость бактерий.
Методы
Бактериальные штаммы, условия культивирования и антибиотики
Штамм Escherichia coli MG1655 был приобретен в DSMZ (Германия) и высеян штрихами до отдельных колоний. Отобрали одну единственную колонию и использовали ее на протяжении всего исследования. Бактериальные клетки выращивали в 30-мл плоскодонных стеклянных флаконах (VWR, 548-0155) в инкубаторе-шейкере (Grant Instruments, ES-20) в течение 18 ч при 30°C перед экспериментами в морбидостате. Лизогенный бульон Миллера (фунт. 10 г л -1 Триптон, 5 г л -1 Дрожжевой экстракт, 10 г л -1 NaCl) использовали в качестве культуральной среды для всех экспериментов. Все антибиотики и питательные среды, используемые в этом исследовании, были приобретены у Sigma-Aldrich, если не указано иное. Растворы антибиотиков готовили из порошков. Их растворяют в растворителях (таблица 1), стерилизуют фильтрованием и выдерживают при 4°С в центрифужных пробирках. Их периодически тестировали путем измерения MIC E. coli (MG1655) 57 . Все лекарственные растворы обновлялись перед разложением.
Строительство системы морбидостата
Система морбидостата была построена на основе инструкций (дополнительный рис. 1a) 58 . Флакон с морбидостатом был сконструирован с использованием 30-мл плоского дна с полипропиленовой завинчивающейся крышкой (дополнительный рис. 1e). Завинчивающаяся крышка была снабжена четырьмя отверстиями, три из которых были оснащены изготовленными на заказ переходниками для трубок, состоящими из иглы, шприца и уплотнительной ленты из политетрафторэтилена (ПТФЭ), для добавления или удаления жидкостей. Два адаптера для пробирок с короткими иглами использовались для добавления свежей среды LB (называемой «свежей средой») и свежей среды LB, содержащей высокую концентрацию антибиотика («лекарственная среда») соответственно. Адаптер трубки с длинной иглой использовали для удаления лишней жидкости из флакона с морбидостатом, чтобы поддерживать постоянный объем бактериальной культуры. Последнее четвертое отверстие на крышке используется для забора фильтрованного воздуха. Весь узел пробирки для культивирования был изготовлен из автоклавируемых материалов.
Свежую и лекарственную среду перекачивали во флакон с помощью перистальтических насосов 12 В постоянного тока (Watson Marlow, 400FD/A1) со скоростью потока 1,2 мл мин -1 . 16-канальный параллельный перистальтический насос (Watson Marlow, 205U) использовали в качестве дренажного насоса для удаления избыточной жидкости из флаконов с морбидостатом. Флаконы и насосы соединены силиконовой трубкой и трубкой из ПТФЭ. Все насосы управляются компьютером с помощью платы Arduino MEGA (Arduino) с изготовленной на заказ монтажной печатной платой (дополнительный рис. 1g). На специальной печатной плате размещены четыре микросхемы из восьми транзисторов Дарлингтона (Texas Instruments, ULN2803A), которые управляют до 32 перистальтических насосов.
Оптическую плотность (ОП) бактериальных клеток измеряли с помощью согласованной пары инфракрасного светодиода (OSRAM, SFh5550) и фототранзистора (Vishay Semiconductor, BPW96B). Узлы держателей трубок, в которых размещаются источник инфракрасного света и фотодиод, были изготовлены на трехмерном принтере (Stratasys, Connex 500) с использованием материала VeroBlack (дополнительный рисунок 1f). Источник инфракрасного света и детектор располагались под углом 135°, чтобы максимизировать чувствительность детектора. Индуцированные светом изменения напряжения на детекторе измеряются с аналоговых входов другой платы Arduino MEGA. Схемы освещения и обнаружения показаны на дополнительном рис. 1h. Значения напряжения позже преобразуются в OD на основе калибровочной кривой (дополнительный рис. 1i).
Окончательно собранный аппарат вмещает 16 флаконов с морбидостатом в инкубаторе с регулируемой температурой (Lucky Reptile, HN-2UK, дополнительные рис. 1c, d). Культуры клеток во флаконах с морбидостатом постоянно перемешиваются небольшими мешалками во время работы. Магнитные мешалки, устанавливаемые непосредственно под флаконы с морбидостатом, выполнены с компьютерными вентиляторами (SUNON, KD0504PFS2.11.GN), трехмерными печатными магнитодержателями и неодимовыми магнитами (COMUS, M1219-4).
Работа системы морбидостата
Стеклянные флаконы, трубки и бутылки, используемые для экспериментов, перед использованием автоклавировали. После автоклавирования силиконовые и ПТФЭ трубки для морбидостата стерилизовали Virkon (Fisher Scientific) и этанолом по отдельности, затем дважды промывали стерильной водой. Бактериальные клетки, хранившиеся при -80°С, оттаивали и переносили в автоклавированные флаконы с морбидостатом с 12 мл среды LB, в которой бактериальные клетки были разведены в 60 раз. Затем собранные флаконы, содержащие клеточные культуры, помещали в держатели флаконов с морбидостатом.
Специальное программное обеспечение, которое отслеживает рост бактерий во флаконах и управляет насосами через платы Arduino, было написано на Python. Это программное обеспечение реализует алгоритм морбидостата, как указано (дополнительный рис. 1b) 50 , который работал следующим образом: мутность каждой клеточной культуры непрерывно измерялась каждые 2 с, затем усреднялась по 30 образцам (то есть усреднялась по 1 мин). ). Каждые 12 мин культуру бактерий во флаконе разбавляли 1 мл свежей или лекарственной среды. Среду не вносили, если ОП культуры клеток ниже 0,15, чтобы избежать вымывания бактериальных клеток из культуры. Во время разбавления проверялись два условия: (1) Если текущая усредненная ОП выше пороговой ОП (ОП THR ∼0,4) и (2) если текущая ОП больше, чем ОП во время предыдущего разбавления. При соблюдении обоих условий к культуре клеток добавляют 1 мл лекарственной среды. В противном случае добавляется свежая среда. Обратите внимание, что мы использовали более высокий порог OD, чем в предыдущем исследовании (OD THR = 0,15) 7 . Это связано с тем, что некоторые антибиотики демонстрируют резкие изменения кривой доза-эффект (например, ампициллин и KAN, дополнительная рис. 13), что затрудняет контроль ингибирования роста у морбидостата, если OD THR низкий.
Концентрацию антибиотика в лекарственной среде начинали с 10-кратной МИК в начале длительного эксперимента. Если бактериальная популяция приобретает устойчивость к антибиотикам, а концентрация лекарства внутри флакона с морбидостатом составляет более 60% от концентрации лекарственной среды, то концентрация антибиотика в лекарственной среде увеличивается в два раза по сравнению с предыдущей концентрацией (дополнительная рис. 14). ).
Один эксперимент с морбидостатом проводился в течение 22 часов. Явного образования биопленок на внутренней стенке флакона с морбидостатом в конце эксперимента не наблюдалось. Культуры клеток отбирали из всех 16 флаконов в морбидостате и хранили в глицериновом растворе при -80 °C.
Анализ данных Морбидостата
Данные журнала операций Морбидостата были проанализированы с помощью пользовательских сценариев Matlab. Скорость роста мк (единица измерения: ч -1 ) рассчитывали по формуле й разбавление. Δ t составляет 600 с. Концентрацию антибиотика А при n -м разведении [A] n рассчитывали по следующему уравнению.
Где V – объем клеточной культуры во флаконе с морбидостатом (то есть 12 мл), Δ V объем свежей или лекарственной среды, добавляемой при каждом разведении (1 мл), [A] запас — концентрация антибиотика А в исходной лекарственной среде (варьируется). Статистические тесты, то есть s.e.m. и P -значение по тесту Уэлча для двух образцов t , были выполнены с использованием R. концентрации антибиотика (-кратное серийное разведение). Развитые штаммы из экспериментов с морбидостатом, хранившиеся при -80°C, оттаивали и переносили в 384-луночные микропланшеты (Fisher) с 65 мкл среды LB. Степень разбавления здесь была 1:50. Микропланшеты инкубировали при 30°С в течение 18 ч при встряхивании со скоростью 1000 об/мин. перед анализом на чашках с агаром MIC. Чашки с агаром LB готовили в омни-чашках (Fisher). Агаровую среду LB автоклавировали и после охлаждения агара добавляли антибиотики. Клетки, выращенные в 384-луночных планшетах, переносили в планшеты с агаром с помощью штифтового репликатора (V&P Scientific) 59 . Чашки с агаром инкубировали при 30°С в течение 18 ч. После инкубации рост клеток оценивали по наличию или отсутствию видимых колоний. Минимальную концентрацию антибиотика без видимой колонии определяли как МИК. Для каждого измерения проводили четыре повторных эксперимента и рассчитывали средние значения. Устойчивость эволюционировавшего мутанта к антибиотикам рассчитывали как log 2 -трансформированных кратных изменений МИК по сравнению со штаммами дикого типа, то есть где МИК[А] WT и re MIC[A] EVO указывают МИК дикого типа (предкового) и эволюционировавшего мутанта против антибиотика А соответственно.
При сравнении экспериментальных результатов с теоретической моделью моделирования оба данных были нормализованы к единице. Для экспериментальных данных значения относительной устойчивости к антибиотикам нормализовали путем деления значения МИК на максимальное значение МИК усредненных данных по эволюции стресса от одного антибиотика.
Логистическая подгонка экспериментальных данных
Эволюционные траектории устойчивости бактерий к антибиотику были снабжены логистической функцией, где L указывает на максимальный относительный уровень МИК, k на крутизну кривой (наклон холма), а m на среднюю точку кривой . Подгонка выполнялась с помощью написанного на заказ скрипта Python с использованием пакета SciPy. Длительность фазы молчания θ рассчитывали как момент времени, когда f (x)≥0,1, то есть .
Профилирование одного клона
Эволюционировавшие штаммы из экспериментов с морбидостатом, хранившиеся при температуре -80 °C, высевали штрихами на чашки с агаром LB для отбора отдельных колоний для анализа МИК. После 15 часов инкубации при 30 °C не менее 25 колоний из каждого образца отбирали случайным образом и растворяли в 180 мкл LB в 96-луночном планшете по отдельности. После инкубации 96-луночного планшета в течение ночи при 30 °С мы удаляем 65 мкл этих клеточных образцов в пустой 384-луночный планшет для анализа МИК, как описано в 2.5.
Извлечение генома и полногеномное ресеквенирование
С помощью настольного секвенатора Illumina MiSeq было секвенировано всего 40 образцов: предок, образцы на 12-й день из всех 19 популяций в условиях одного антибиотика и образцы на 24-й день из 20 популяций в условиях циклического применения антибиотиков и контроль. Каждый эволюционировавший штамм разводили 1:100 в 5 мл LB без лекарственного средства и выращивали в течение ночи (18 ч) при 30 °C перед экстракцией ДНК. Мы подтвердили, что ночной рост без антибиотиков не меняет уровень устойчивости к антибиотикам двумя способами. Во-первых, устойчивые образцы, развившиеся в условиях одного антибиотика (дополнительная рис. 5, образцы последнего дня), выращивали в течение ночи с антибиотиками и без них. Значения MIC были одинаковыми независимо от стресса от антибиотиков (дополнительная рис. 10). Во-вторых, измеряли колониеобразующую единицу устойчивого штамма, чтобы увидеть, изменяется ли уровень устойчивости во время инкубации в течение ночи без антибиотиков. Использовали последний дневной образец циклов POL и CHL (рис. 5а, день 24), и сравнивали частоту выживших между замороженным запасом и ночной культурой запаса, выращенного без антибиотиков. Однако существенной разницы между этими двумя условиями не наблюдалось (дополнительная рис. 11). Геномную ДНК выделяли из 1 мл культур с использованием набора Wizard Genomic Purification Kit (Promega #9).FB022). Этап расщепления РНКазой был добавлен после процесса выделения генома. Библиотеки для секвенирования готовили с использованием набора для подготовки ДНК-библиотек Nextera XT.
Биоинформатический анализ секвенирования генома
Полученные последовательности приведены в соответствие с эталонными данными MG1655 (NC_000913.2). Выравнивание выполнялось с помощью Bowtie2. SNP и вставки были идентифицированы с помощью SAMtools. Идентификацию мутаций анализировали с использованием breseq (ссылка 60). Для каждого штамма было достигнуто минимальное 100-кратное покрытие. Выбранные предполагаемые варианты (SNP и инделы), обнаруженные с помощью ресеквенирования всего генома, были проверены с помощью ПЦР с последующим секвенированием по Сэнгеру. Праймеры, используемые для секвенирования по Сэнгеру, были разработаны с помощью Primer3web.
Определение частоты аллелей
Частоты аллелей определяли по интенсивностям флуоресцентных сигналов данных секвенирования по Сэнгеру, то есть по отношению интенсивности сигнала мутантного SNP к общему сигналу. Последовательности выравнивали с помощью DNA Baser (www.dnabaser.com/). Хотя это не точный способ определения частот аллелей, анализ использовался для определения того, является ли эволюционировавшая популяция генетически гомогенной или гетерогенной.
Математическая модель
Численные решения для теоретической модели были рассчитаны в программах Matlab (Mathworks, США) и Julia (http://julialang. org/) с использованием пакета Sundials. Более подробную информацию о модели можно найти в дополнительных методах.
Доступность данных
Данные полногеномного секвенирования были депонированы в Национальном центре биотехнологической информации (NCBI) Sequence Read Archive (SRA) со следующими образцами кодов доступа: SRS1749080 (CHL), SRS1749079 (POL), SRS1749078 (POL) -КХЛ), SRS1749077 (CHL-POL_D6), SRS1749084 (CHL-POL_D10), SRS1749083 (CHL-POL_D18), SRS1749082 (CHL-POL_D24), SRS1749081 (MG1655 дикого типа) и SRS1749076 (контроль). Специальное программное обеспечение, используемое в этом исследовании, доступно по адресу http://www.chem.gla.ac.uk/cronin/e_media/. Авторы заявляют, что все другие соответствующие данные, подтверждающие результаты исследования, включены в эту опубликованную статью и ее дополнительную информацию или могут быть получены от соответствующих авторов по запросу.
Дополнительная информация
Как цитировать эту статью: Yoshida, M. et al . Программируемое по времени дозирование лекарств позволяет манипулировать, подавлять и устранять резистентность к антибиотикам in vitro . Нац. коммун. 8, 15589 doi: 10.1038/ncomms15589 (2017).
Примечание издателя: Springer Nature остается нейтральной в отношении юрисдикционных претензий в опубликованных картах и институциональной принадлежности.
Каталожные номера
-
Лаксминараян, Р. и др. Устойчивость к антибиотикам — необходимость глобальных решений. Заражение ланцетом. Дис. 13 , 1057–1098 (2013).
Google ученый
-
Сильвер, Л. Л. Проблемы открытия антибактериальных средств. клин. микробиол. 24 , 71–109 (2011).
КАС Google ученый
-
Смирнова М.
 В. и др. Антистафилококковая фармакодинамика линезолида отдельно и в комбинации с доксициклином в in vitro динамическая модель. Дж. Чемотер. 23 , 140–144 (2011).
В. и др. Антистафилококковая фармакодинамика линезолида отдельно и в комбинации с доксициклином в in vitro динамическая модель. Дж. Чемотер. 23 , 140–144 (2011). КАС Google ученый
-
Фирсов А.А. и др. Основанное на фармакокинетике прогнозирование эффектов комбинаций антибиотиков на резистентные мутанты Staphylococcus aureus : модельные исследования in vitro с линезолидом и рифампицином. Дж. Чемотер http://dx.doi.org/10.1080/1120009X.2016.1245174 (2017).
-
Фирсов А. А., Востров С. Н., Лубенко И. Ю., Портной Ю. А. и Зиннер С. Х. Предотвращение селекции резистентного штамма Staphylococcus aureus с помощью моксифлоксацина плюс доксициклин в модели in vitro : аддитивный эффект динамической модели комбинация. Междунар. Дж. Антимикроб. Агенты 23 , 451–456 (2004).
КАС Google ученый
-
Gloede, J.
, Scheerans, C., Derendorf, H. & Kloft, C. In vitro фармакодинамические модели для определения действия антибактериальных препаратов. J. Антимикроб. Чемотер. 65 , 186–201 (2010). КАС Google ученый
-
Топрак Э. и др. Эволюционные пути к антибиотикорезистентности в условиях динамично поддерживаемого лекарственного отбора. Нац. Жене. 44 , 101–105 (2011).
Google ученый
-
Шеверо, Г. и др. Количественная оценка детерминант эволюционной динамики, ведущей к лекарственной устойчивости. PLoS Биол. 13 , e1002299 (2015).
Google ученый
-
Ким, С., Либерман, Т. Д. и Кишони, Р. Чередование антибиотиков сдерживает эволюционные пути к множественной лекарственной устойчивости. Проц. Натл акад. науч.
США 111 , 14494–14499 (2014). КАС Google ученый
-
Fuentes-Hernandez, A. et al. Использование последовательного режима для элиминации бактерий при сублетальных дозах антибиотиков. PLoS Биол. 13 , e1002104 (2015).
Google ученый
-
Pena-Miller, R. et al. Когда самая мощная комбинация антибиотиков вызывает наибольшую бактериальную нагрузку: переход от улыбки к хмурому взгляду. PLoS Биол. 11 , e1001540 (2013).
КАС Google ученый
-
Лазар, В. и др. Бактериальная эволюция гиперчувствительности к антибиотикам. Мол. Сист. биол. 9 , 700 (2013).
Google ученый
-
Suzuki, S., Horinouchi, T. & Furusawa, C.
Прогнозирование устойчивости к антибиотикам по профилям экспрессии генов. Нац. коммун. 5 , 5792 (2014). КАС Google ученый
-
Suzuki, S., Horinouchi, T. & Furusawa, C. Фенотипические изменения, связанные со стоимостью приспособленности штаммов кишечной палочки, устойчивых к антибиотикам. Мол. Биосист. 12 , 414–420 (2016).
КАС Google ученый
-
Друсано, Г. Л., Луи, А., Макгоуэн, А. и Хоуп, В. Подавление возникновения резистентности у патогенных бактерий: сохранение нашего порошка сухим. Антимикроб. Агенты Чемотер. 60 , 1183–1193 (2016).
КАС Google ученый
-
Друсано, Г. Л., Хоуп, В., Макгоуэн, А. и Луи, А. Подавление возникновения резистентности у патогенных бактерий: сохранение нашего порошка сухим.
Антимикроб. Агенты Чемотер. 60 , 1194–1201 (2016). КАС Google ученый
-
Mira, P.M. et al. Рациональный дизайн планов лечения антибиотиками: стратегия лечения для управления эволюцией и обращения вспять резистентности. PLoS ONE 10 , e0122283 (2015).
Google ученый
-
Никол, Д. и др. Направление эволюции с последовательной терапией для предотвращения появления устойчивости бактерий к антибиотикам. Вычисление PLoS. биол. 11 , e1004493 (2015).
Google ученый
-
Пенья-Миллер, Р., Фуэнтес-Эрнандес, А., Рединг, К., Гудель, И. и Бердмор, Р. Проверка свойств оптимальности двойного лечения антибиотиками в двухлокусной, двухаллельной модель. JR Soc. Интерфейс 11 , 20131035 (2014).
Google ученый
-
Пена-Миллер Р.
, Ланеманн Д., Шуленбург Х., Акерманн М. и Бердмор Р. Оптимальное применение синергетических антибиотиков: теоретико-контрольный подход. J. R. Soc. Интерфейс 9 , 2488–2502 (2012 г.). КАС Google ученый
-
Gerding, D. N. et al. Резистентность к аминогликозидам и использование аминогликозидов: десятилетний опыт работы в одной больнице. Антимикроб. Агенты Чемотер. 35 , 1284–1290 (1991).
КАС Google ученый
-
Гердинг, Д. Н. Циклическое применение противомикробных препаратов: уроки, извлеченные из опыта применения аминогликозидов. Заразить. Хосп. Эпидемиол. 21 , С12–С17 (2000).
КАС Google ученый
-
Коллеф, М. Х. Является ли циклическое применение антибиотиков ответом на предотвращение возникновения бактериальной резистентности в отделении интенсивной терапии? клин.
Заразить. Дис. 43 , S82–S88 (2006). Google ученый
-
Lee, C.R., Cho, I.H., Jeong, B.C. & Lee, S.H. Стратегии минимизации устойчивости к антибиотикам. Междунар. Дж. Окружающая среда. Рез. Общественное здравоохранение 10 , 4274–4305 (2013 г.).
КАС Google ученый
-
Бал, А. М., Кумар, А. и Гулд, И. М. Неоднородность антибиотиков: от концепции к практике. Энн. Н. Я. акад. науч. 1213 , 81–91 (2010).
КАС Google ученый
-
Bergstrom, C. T., Lo, M. & Lipsitch, M. Экологическая теория предполагает, что цикличность противомикробных препаратов не снижает резистентность к противомикробным препаратам в больницах. Проц. Натл акад. науч. США 101 , 13285–13290 (2004 г.).
КАС Google ученый
-
Bonhoeffer, S.
, Lipsitch, M. & Levin, B. R. Оценка протоколов лечения для предотвращения устойчивости к антибиотикам. Проц. Натл акад. науч. США 94 , 12106–12111 (1997). КАС Google ученый
-
Abel zur Wiesch, P., Kouyos, R., Abel, S., Viechtbauer, W. & Bonhoeffer, S. Циклическая эмпирическая антибактериальная терапия в больницах: метаанализ и модели. PLoS Патог. 10 , e1004225 (2014).
Google ученый
-
Имамович, Л. и Соммер, М. О. А. Использование сетей побочной чувствительности для разработки протоколов чередования лекарств, позволяющих избежать развития резистентности. Науч. Перевод Мед. 5 , 204ra132–204ra132 (2013).
Google ученый
-
Бейм, М., Стоун, Л. К. и Кишони, Р. Эволюционные стратегии применения нескольких лекарств для устранения устойчивости к антибиотикам.
Science 351 , aad3292–aad3292 (2016). Google ученый
-
Ромхильд, Р., Барбоза, К., Бердмор, Р. Э., Янсен, Г. и Шуленбург, Х. Временные вариации в среде с антибиотиками замедляют развитие резистентности у патогенной синегнойной палочки. Эволюция. заявл. 8 , 945–955 (2015).
КАС Google ученый
-
Schenk, M. F. et al. Роль плейотропии при адаптации ТЕМ-1-??-лактамазы к двум новым антибиотикам. Эволюция. заявл. 8 , 248–260 (2015).
КАС Google ученый
-
Drlica, K. & Zhao, X. Гипотеза окна выбора мутантов обновлена. клин. Заразить. Дис. 44 , 681–688 (2007).
Google ученый
Lee, H.H., Molla, M.N., Cantor, C.
R. & Collins, J.J. Бактериальная благотворительность приводит к сопротивлению всего населения. Природа 467 , 82–85 (2010). КАС Google ученый
-
Конрад, Т. М., Льюис, Н. Э. и Палссон, Б. Ø. Эволюция микробной лаборатории в эпоху науки о геноме. Мол. Сист. биол. 7 , 509 (2011).
Google ученый
-
Оз, Т. и др. Сила давления отбора является важным параметром, определяющим сложность эволюции устойчивости к антибиотикам. Мол. биол. Эвол. 31 , 2387–2401 (2014).
КАС Google ученый
-
Олайтан А.О., Моран С. и Ролен Ж.-М. Механизмы резистентности к полимиксинам: приобретенная и врожденная резистентность бактерий. Перед. микробиол. 5 , 643 (2014).
Google ученый
-
Эль-Халфави, О.
М. и Вальвано, М. А. Гетерорезистентность к противомикробным препаратам: новая область, требующая ясности. клин. микробиол. 28 , 191–207 (2015). КАС Google ученый
-
Эль-Халфави, О. М. и Вальвано, М. А. Химическая связь устойчивости к антибиотикам высокоустойчивой субпопуляции бактериальных клеток. PLoS ONE 8 , e68874 (2013).
КАС Google ученый
-
Эль-Халфави, О. М. и Вальвано, М. А. Путресцин снижает вызванный антибиотиками окислительный стресс как механизм модуляции устойчивости к антибиотикам у Burkholderia cenocepacia. Антимикроб. Агенты Чемотер. 58 , 4162–4171 (2014).
Google ученый
-
Хьюз, Д. и Андерссон, Д. И. Эволюционные последствия лекарственной устойчивости: общие принципы для различных целей и организмов.
Нац. Преподобный Жене. 16 , 459–471 (2015). КАС Google ученый
-
Левин Б.Р., Перро В. и Уокер Н. Компенсаторные мутации, устойчивость к антибиотикам и популяционная генетика адаптивной эволюции бактерий. Генетика 154 , 985–997 (2000).
КАС Google ученый
-
Хегренесс М., Шореш Н., Хартл Д. и Кишони Р. Принцип эквивалентности включения благоприятных мутаций в бесполые популяции. Наука 311 , 1615–1617 (2006).
КАС Google ученый
-
Horinouchi, T. et al. Транскриптомный анализ параллельно эволюционировавших штаммов Escherichia coli в условиях этанольного стресса. BMC Genomics 11 , 579 (2010).
Google ученый
-
Horinouchi, T.
et al. Фенотипическая конвергенция в адаптивной эволюции бактерий к этанольному стрессу. BMC Evol. биол. 15 , 180 (2015). Google ученый
-
Фурусава, К. и Канеко, К. Глобальные отношения в колебаниях и реакции в адаптивной эволюции. J. R. Soc. Интерфейс 12 , 20150482 (2015).
Google ученый
-
Lenski, R. E. Бактериальная эволюция и стоимость устойчивости к антибиотикам. Междунар. микробиол. 1 , 265–270 (1998).
КАС Google ученый
-
Хуанг С. и Кауфман С. Как избежать аттрактора рака: обоснование и ограничения многоцелевых препаратов. Семин. Рак биол. 23 , 270–278 (2013).
КАС Google ученый
-
Фурусава, К.
и Канеко, К. Взгляд динамических систем на биологию стволовых клеток. Наука 338 , 215–217 (2012). КАС Google ученый
-
Вёрёш, А. и др. SecDF в составе sec-транслоказы способствует эффективной секреции токсинов Bacillus cereus и белков, ассоциированных с клеточной стенкой. PLoS ONE 9 , e103326 (2014).
Google ученый
-
Quiblier, C., Zinkernagel, A.S., Schuepbach, R.A., Berger-Bächi, B. & Senn, M.M. Вклад SecDF в устойчивость Staphylococcus aureus и экспрессию факторов вирулентности. ВМС микробиол. 11 , 72 (2011).
КАС Google ученый
-
Stone, L.K. et al. Соединения, которые выбирают против эффлюксной помпы, устойчивой к тетрациклину. Нац. хим. биол. 12 , 902–904 (2016).
КАС Google ученый
-
Шибальский В. и Брайсон В. Генетические исследования перекрестной устойчивости микробов к токсичным агентам. I. Перекрестная устойчивость кишечной палочки к пятнадцати антибиотикам. J. Бактериол. 64 , 489–499 (1952).
КАС Google ученый
-
Холл, М. Д., Хэндли, М. Д. и Готтесман, М. М. Сопротивление бесполезно? Множественная лекарственная устойчивость и побочная чувствительность. Trends Pharmacol. науч. 30 , 546–556 (2009).
КАС Google ученый
-
Фирсов А. А. и Зиннер С. Х. Использование методов моделирования для помощи в выборе антибиотиков. Курс. Заразить. Дис. 3 , 35–43 (2001).
Google ученый
-
Cui, X.
et al. Сублетальные дозы летального токсина Bacillus anthracis ингибируют воспаление с помощью липополисахарида и заражения Escherichia coli, но оказывают противоположное влияние на выживаемость. Дж. Заразить. Дис. 193 , 829–840 (2006). КАС Google ученый
-
Chait, R., Craney, A. & Kishony, R. Взаимодействие антибиотиков, направленное против резистентности. Природа 446 , 668–671 (2007).
КАС Google ученый
-
Топрак Э. и др. Создание морбидостата: автоматизированное устройство непрерывного культивирования для изучения устойчивости бактерий к лекарственным средствам при динамическом устойчивом ингибировании лекарственными препаратами. Нац. протокол 8 , 555–567 (2013).
Google ученый
-
Голдсмит М.
и Тауфик Д. С. Потенциальная роль фенотипических мутаций в эволюции экспрессии и стабильности белков. Проц. Натл акад. науч. США 106 , 6197–6202 (2009 г.). КАС Google ученый
-
Deatherage, D. E. & Barrick, J. E. Идентификация мутаций в микробах, эволюционировавших в лаборатории, на основе данных секвенирования следующего поколения с использованием breseq. Методы Мол. биол. 1151 , 165–188 (2014).
КАС Google ученый
Скачать ссылки
Механизмы резистентности при кастрационно-резистентном раке предстательной железы (КРРПЖ) — Чандрасекар
Обзорная статья
Thenappan Chandrasekar, Joy C. Yang, Allen C. Gao, Christopher P. Evans
Кафедра урологии, Калифорнийский университет, Дэвис, Калифорния, США
Адрес для переписки: Christopher P. Evans, MD. 4860 Y Street, Suite 3500, Сакраменто, Калифорния
, США. Электронная почта: [email protected].
Резюме: Несмотря на успехи в диагностике и лечении рака предстательной железы, заболеваемость раком предстательной железы остается высокой. Приблизительно у 20% мужчин заболевание развивается на поздних стадиях или с метастазами, в то время как 29 000 мужчин продолжают умирать от рака простаты каждый год. Терапия депривации андрогенов (АДТ) была стандартом лечения начального лечения распространенного или метастатического рака простаты с тех пор, как Хаггинс и Ходжес впервые представили концепцию андрогенной зависимости в 1972 году, но прогрессирование до кастрационно-резистентного рака простаты (КРРПЖ) происходит в течение 2 лет. -3 года от начала ADT. КРРПЖ, ранее определявшийся как гормонорезистентный рак предстательной железы, в настоящее время считается все еще зависимым от андрогенов. Множественные механизмы резистентности способствуют прогрессированию болезни, резистентной к кастрации, и андрогенный рецептор (AR) остается важным фактором в этом прогрессировании. Эти механизмы включают амплификацию AR и гиперчувствительность, мутации AR, приводящие к беспорядочным половым связям, мутации коактиваторов/корепрессоров, андроген-независимую активацию AR, а также внутриопухолевую и альтернативную продукцию андрогенов. Совсем недавно идентификация вариантов АР (АРВ) была установлена как еще один механизм прогрессирования КРРПЖ. Химиотерапия доцетакселом исторически была терапией первой линии при КРРПЖ, но в последние годы были введены новые препараты, нацеленные на некоторые из этих механизмов резистентности, тем самым обеспечивая дополнительное преимущество в выживаемости. К ним относятся ингибиторы передачи сигналов AR, такие как энзалутамид (Xtandi, ENZA, MDV-3100), и ингибиторы CYP17A1, такие как абиратерона ацетат (Zytiga). В конечном счете, эти агенты также не смогут подавить CRPC. Хотя некоторые механизмы, из-за которых эти агенты не работают, уникальны, многие из них имеют сходство с механизмами, способствующими прогрессированию КРРПЖ. Понимание этих механизмов резистентности к ADT и утвержденным в настоящее время методам лечения КРРПЖ поможет в будущих исследованиях таргетных методов лечения.
Ключевые слова: Устойчивый к кастрации; прогрессирование заболевания; устойчивость к лекарству; новообразования предстательной железы
Поступила 21.12.2014. Принята к публикации 28.02.2015.
Рак предстательной железы является наиболее часто диагностируемым некожным злокачественным новообразованием у мужчин и остается второй по значимости причиной смерти от рака у мужчин (1,2). Несмотря на успехи в скрининге и раннем выявлении рака предстательной железы, у значительной части мужчин по-прежнему наблюдается прогрессирующее или метастатическое заболевание — примерно 20% мужчин в недавних отчетах (3). Действительно, заболеваемость этим заболеванием остается высокой, более 29000 смертей, связанных с раком простаты, только в 2013 году (1).
Андроген-депривационная терапия (АДТ), стандарт лечения пациентов с биохимическим рецидивом после радикальной первичной терапии, местно-распространенным заболеванием или метастатическим заболеванием, продемонстрировала первоначальную пользу, но у большинства пациентов прогрессирует до кастрационно-резистентного заболевания. в течение 2-3 лет (4).
Кастрационно-резистентный рак предстательной железы (КРРПЖ), ранее называвшийся гормонорезистентным раком предстательной железы, в настоящее время рассматривается как прогрессирование заболевания, несмотря на медикаментозную или хирургическую кастрацию. Сдвиг парадигмы связан с пониманием того, что КРРПЖ не является гормонорезистентным — фактически ось андрогенов продолжает играть важную роль в функционировании и росте КРРПЖ. Действительно, в то время как другие пути могут способствовать резистентности к кастрации, рецептор андрогена (АР) остается наиболее важным фактором в континууме КРРПЖ.
Понимание механизмов резистентности, которые вызывают прогрессирование гормонально-наивного рака предстательной железы в резистентность к кастрации, является ключом к разработке будущей терапии. В этом обзоре мы рассмотрим современные знания о механизмах, ведущих к резистентности к кастрации, агентах, доступных в настоящее время для лечения КРРПЖ, и механизмах резистентности к этим агентам.
Фон
Понимание оси андрогенов является ключевым компонентом для понимания механизмов, посредством которых развивается устойчивость к кастрации.
Рецептор андрогена (AR)
Ген AR на Xq11-12 кодирует ядерный рецептор массой 110 кДа с четырьмя различными функциональными мотивами — амино-концевым доменом (NTD), ДНК-связывающим доменом, шарнирной областью и лиганд-связывающим доменом (LBD) (5 -7). Цитоплазматический рецептор связан белками теплового шока (в частности, шапероновым комплексом HSP90) в неактивном состоянии (8). Связывание андрогенов, особенно дигидротестостерона (ДГТ) или тестостерона, с LBD вызывает конформационные изменения, которые приводят к диссоциации HSP9.0, гомодимеризация рецептора, транслокация в ядро и связывание с андроген-отвечающими элементами (ARE) в промоторной области генов, регулируемых андрогенами (6,9). Это взаимодействие с промоторной областью находится под влиянием многих корегуляторов транскрипции. Было идентифицировано более 150 белков (10), и многие из них являются ферментами (гистоновые ацетилтрансферазы, метилтрансферазы, киназы), которые действуют, открывая структуру хроматина, чтобы способствовать транскрипции.
Андрогены
Рост рака предстательной железы и выживаемость зависят от андрогенов, основных лигандов АР. Тестостерон является основным циркулирующим андрогеном, примерно 90% которого продуцируется клетками Лейдига в яичках и 10% — корой надпочечников. Только небольшая часть (3%) циркулирующего тестостерона не связана и функционально активна, а остальная часть связана и секвестрирована глобулином, связывающим половые гормоны, и альбумином. Однако тестостерон не является основным функционально активным андрогеном в микроокружении предстательной железы. После диффузии в цитоплазму тестостерон под действием фермента 5α-редуктазы превращается в ДГТ, сродство которого к LBD AR в пять раз выше (11–13).
Физиологический уровень андрогенов необходим для стимуляции роста и предотвращения апоптотической гибели. Следовательно, пути воздействия АР разнообразны, но сосредоточены на функциях эпителиальных клеток просвета, включая продукцию белков семенной жидкости, таких как простатспецифический антиген (ПСА), и множественные гены в метаболическом пути, ведущие к увеличению синтеза белков и липидов. (14-16).
Стероидогенез, который приводит к выработке андрогенов, является важным путем для понимания, поскольку он может фундаментально изменяться при КРРПЖ. Тестостерон вырабатывается яичками и надпочечниками, а затем преобразуется в цитоплазме в ДГТ под действием 5α-редуктазы (17). Тем не менее, в присутствии АДТ исследования продемонстрировали устойчивые уровни внутриопухолевого ДГТ (18-21), предполагая активацию измененных путей стероидогенеза (20). Надпочечниковые источники тестостерона, не затронутые АДТ, и внутриопухолевые 9Синтез андрогенов 1543 de novo может быть источником стойкой лиганд-зависимой активности AR при КРРПЖ (22).
Андрогенная депривационная терапия (АДТ)
С тех пор, как Huggins и Hodges (23) впервые продемонстрировали зависимость рака предстательной железы от передачи сигналов андрогенов, ГТ с помощью медикаментозной или хирургической кастрации стала стандартом лечения метастатического и местно-распространенного заболевания. Хирургическая кастрация или двусторонняя орхиэктомия удаляет андрогены яичек из кровотока путем удаления источника. Медицинская кастрация достигается за счет использования различных классов средств. Агонисты и антагонисты ЛГРГ истощают выработку гипофизом лютеинизирующего гормона (ЛГ) за счет отрицательной обратной связи или конкурентного ингибирования, соответственно, что, в свою очередь, предотвращает выработку тестикулярного тестостерона (24). Антиандрогены действуют как конкурентные ингибиторы LBD AR, тем самым предотвращая андрогенную стимуляцию AR. Эти агенты в сочетании обеспечивают полную блокаду андрогенов (25, 26).
Устойчивость к кастрации
Несмотря на первоначальный ответ на блокаду андрогенов, у всех пациентов в конечном итоге развивается резистентность к кастрации. Резистентность к кастрации — это прогрессирование заболевания, либо клиническое (развитие метастазов, прогрессирование ранее существовавшего заболевания), либо биохимическое (три последовательных подъема уровня ПСА выше минимума) при кастрационном уровне циркулирующего тестостерона (<50 нг/дл). (27,28).
Действительно, биохимический рецидив ПСА, гена, регулируемого АР, измеряемый по уровням в сыворотке, свидетельствует о том, что КРРПЖ не является гормононечувствительным. При добавлении антиандрогенов первого поколения, таких как флутамид или бикалутамид, к схеме лечения пациентов с прогрессирующим или метастатическим заболеванием часто первоначально отмечается снижение уровня ПСА в сыворотке, что указывает на реакцию на прямую блокаду АР (29).,30). Однако, несмотря на антиандрогенную терапию, уровни ПСА в сыворотке снова будут повышаться, что свидетельствует о том, что агент начал действовать как агонист АР; это подтверждается снижением уровня ПСА, отмеченным при отмене антиандрогенов (31,32).
Еще одно доказательство критической роли андрогенной системы в развитии КРРПЖ заключается в обнаружении того, что, несмотря на кастрированные уровни сывороточного тестостерона, при КРРПЖ остается более высокий уровень внутриопухолевых андрогенов по сравнению с раком предстательной железы без гормонов (18). -21). Недавние исследования показали, что внутриопухолевые уровни андрогенов при КРРПЖ сходны с таковыми у эугонадных мужчин, а в некоторых случаях даже повышены (22,33).
AR персистирует в клетках CRPC, и реактивация этой оси с помощью следующих механизмов, по-видимому, приводит к развитию резистентности к кастрации.
AR-зависимые механизмы резистентности, ведущие к КРРПЖ
Большинство идентифицированных механизмов, приводящих к резистентности к кастрации, опосредуются AR или осью андрогенов. Как видно из рисунка 1 , их можно разделить на пять основных подмножеств.
Рисунок 1 Андроген-рецептор-зависимые механизмы резистентности при гормонально-наивном раке предстательной железы, ведущие к резистентности к кастрации. wtAR, рецептор андрогена дикого типа; АРВ, вариант рецептора андрогена; mutAR, мутированный рецептор андрогена; Т, тестостерон; ДГТ, дигидротестостерон; ГСПГ, глобулин, связывающий половые гормоны.
Амплификация и мутации AR/путь гиперчувствительности/беспорядочный путь
Низкий уровень андрогенов сохраняется, несмотря на блокаду андрогенов с помощью АДТ. В этом микроокружении подгруппа клеток развивает чувствительность к этим низким уровням андрогенов либо за счет амплификации AR (путь гиперчувствительности) (34), либо за счет развития мутаций AR, которые приводят к активации молекулами, отличными от андрогенов (неразборчивый путь) (35, 36).
Амплификация AR была идентифицирована в значительной части клеточных линий CRPC, в диапазоне от 30 до 80% (37,38). Это открытие редко встречается при гормонально-наивном раке предстательной железы и может быть связано с избирательным ростом клеток КРРПЖ (36). Это усиление позволяет КРРПЖ быть гиперчувствительным к низкому уровню андрогенов, что способствует прогрессированию заболевания (35). Поскольку 20% метастазов КРРПЖ имеют признаки амплификации АР, которые отсутствуют при метастатическом заболевании без применения гормонов, это также может способствовать метастазированию. Кроме того, недавние исследования показали, что экзогенная сверхэкспрессия AR может приводить к КРРПЖ.
В соответствии с этой концепцией замена валина на лейцин в кодоне 89 приводит к повышению уровня 5α-редуктазы в подмножестве КРРПЖ. Это приводит к более высокому уровню ДГТ, несмотря на низкий уровень циркулирующего тестостерона. Эта мутация чаще наблюдается у афроамериканцев и связана с более агрессивным раком предстательной железы с ранним началом (39,40).
В самом гене AR были выявлены различные точечные мутации, которые приводят к повышению активности AR в присутствии низких уровней андрогенов, а также неандрогенных стероидов, таких как прогестерон, гидрокортизон, эстрадиол и некоторые антагонисты AR. Замена треонина на аланин в кодоне 877 в клетках LNCAP (41, 42) и замена гистидина на тирозин в клетках CWR22 (43, 44) хорошо описаны в литературе; другие примеры включают L701H, V715M, W741C (45-47). Хотя большинство мутаций преимущественно связано с LBD, также были идентифицированы мутации в NTD и ДНК-связывающем домене (48, 49). ).
Коактиваторы и корепрессоры
Более 150 различных молекул уже идентифицированы как коактиваторы и корепрессоры АР (10). AR обычно задействует ряд корегуляторных комплексов, которые могут либо усиливать (коактиваторы), либо подавлять (корепрессоры) транскрипционную активность. Многие из этих корегуляторов представляют собой ферменты, которые служат для модуляции других белков в комплексе посредством фосфорилирования, метилирования, ацетилирования или убиквитилирования, но они также были идентифицированы как молекулярные шапероны, рекрутеры транскрипционного аппарата и регуляторы сплайсинга РНК (50-52).
Было обнаружено, что один коактиватор, FKBP51, который также является геном-мишенью AR, активируется в рецидивирующих опухолях LAPC-4, выращенных у кастрированных мышей (53). Он способствует образованию комплекса супершаперона, регулируя привлечение p23, ко-шаперона, к Hsp90, связанному с АТФ, что, в свою очередь, удерживает AR в конформации с высоким сродством к связыванию лиганда. Это способствует стимулируемой андрогенами транскрипционной активности и росту.
Коактиваторы стероидных рецепторов (SRC) представляют собой класс коактиваторов AR, способных проявлять ацетилтрансферазную активность, которая, в свою очередь, усиливает AR-индуцированную транскрипцию, способствуя образованию комплексов между AR-ассоциированными энхансерами/промоторами и сайтом начала транскрипции генов-мишеней AR (54). ). Класс SRC включает SRC-1, SRC-2 (TIF-2, GRIP-1, NcoA2) и SRC-3 (AIB). Сюй и др. . продемонстрировали, что все три были связаны с прогрессированием рака предстательной железы (55). Уэда и др. . идентифицированный SRC-1 при фосфорилировании MAPK под влиянием IL-6 был способен как к лиганд-зависимой, так и к лиганд-независимой активации AR (56). В частности, SRC-3 имеет тенденцию к чрезмерной амплификации при раке человека. Чанг и др. . продемонстрировали, что SRC-3 не избыточно экспрессируется при андрогензависимом раке предстательной железы, но сверхэкспрессируется при низкодифференцированном и более прогрессирующем раке предстательной железы и напрямую связан с прогрессированием рака предстательной железы — мыши с нокаутом SRC-3 были эффективно задержаны на хорошо дифференцированной стадии и неспособен прогрессировать в более тяжелую патологию (57).
Другие важные пути включают p300/CBP, который способствует андроген-независимой IL-6-опосредованной активации AR в присутствии STAT3 (58), а также LSD1 и JMJD2c, лизиндеметилаз, которые деметилируют белки гистонов h4 и приводят к усилению транскрипции, индуцированной AR. 59). Многие из этих молекул продемонстрировали АР-зависимые и АР-независимые эффекты, поскольку их взаимодействие не ограничивается АР. С другой стороны, при КРРПЖ были обнаружены пониженные уровни белков-корепрессоров.
Аберрантная активация (посттрансляционная модификация)/незаконный путь
В то время как все предшествующие механизмы опосредуют повышенную активность AR в присутствии лиганда, независимая от лиганда активация AR также является важным механизмом развития резистентности к кастрации. Различные исследования in vitro показали, что многочисленные факторы роста, цитокины и киназные пути усиливают передачу сигналов AR, тем самым способствуя прогрессированию резистентности к кастрации лиганд-независимым образом (60). Идентификация и характеристика этих лиганд-независимых путей может привести к дополнительным таргетным методам лечения.
Было установлено, что семейство белков NF-kB является важным компонентом онкогенного пути при множественных злокачественных новообразованиях человека. Существует пять различных белков NF-κB, включая хорошо изученный гетеродимер p65/p50, который, как было показано, конститутивно активен при раке предстательной железы. Другой из путей NF-κB, путь p100/p52, вызвал недавний интерес. Процессинг р100 в р52 с помощью таких молекул, как лимфотоксин β, фактор активации В-клеток, лиганд CD40 и stat3 (61) при раке предстательной железы приводит к значительной гиперплазии и индуцированному резистентному к кастрации росту. Это было достигнуто за счет ограничения опосредованного ADT апоптоза и ингибирования клеточного цикла, но это было сделано в присутствии продолжающейся экспрессии и активации AR, что предполагает, что p52 может активировать AR во время прогрессирования КРРПЖ. p52 опосредовал свои эффекты AR-зависимым образом, взаимодействуя непосредственно с NTD AR. Понижение уровня р52 в клетках С4-2 приводило к потере конститутивной активации АР, что предполагало независимую от андрогенов активацию АР (62).
Путь PI3K является еще одним важным участником этого процесса. Потеря белка-супрессора опухоли PTEN, который является отрицательным ингибитором пути PI3K/AKT, идентифицируется почти при всех метастатических формах рака предстательной железы. Его активация была связана с развитием CPRC в различных доклинических моделях (63-65). PI3K, особенно изоформа p110β, тесно связана с ростом и прогрессированием рака предстательной железы за счет базовой активации AKT в моделях рака предстательной железы. Путь PI3K/AKT находится ниже ключевых рецепторных тирозинкиназ (RTK), таких как EGFR, IGFR, c-met, но некоторые исследования предполагают независимую активацию этого пути (66). Хотя он также находится выше некоторых важных сигнальных белков, таких как mTOR, также было обнаружено, что AKT напрямую фосфорилирует AR в двух местах, Ser-217 и Ser-79. 1, особенно в кастрированном состоянии, хотя клиническое значение еще не определено (67).
Src, ключевой член семейства не-RTK, называемого семейством киназ Src (SFK), была в центре внимания нашей лаборатории и наших сотрудников. Src, за 25 лет с момента его открытия в качестве первого идентифицированного протоонкогена, стал мишенью для лечения множества других злокачественных новообразований (68,69). Наше исследование роли Src в развитии рака предстательной железы определило Src как ключевую молекулу во многих путях, которые способствуют прогрессированию рака предстательной железы (70,71). Src экспрессируется в обычно используемых линиях клеток CaP CWR22Rv1, DU145, LAPC-4, LNCaP и PC-3. Поскольку Src не является конститутивно активным, трудно сопоставить уровни экспрессии белка Src с клеточной пролиферацией или агрессивностью in vitro . Тем не менее, киназа Src находится ниже по течению от многих важных влияний на рак простаты — поскольку она активируется факторами роста, цитокинами, хемокинами и пептидом, высвобождающим гастрин, она оказывает плейотропное действие на рак простаты (68,70,71). Наша лабораторная группа продемонстрировала, что более высокая относительная активация Src была связана с худшими фенотипами рака простаты, в частности, DU145 и PC3, а использование нового ингибитора SRC AZD0530 помогло выяснить некоторые пути, опосредованные Src в клеточных линиях рака простаты (70). Активация киназы Src была связана с андроген-независимым ростом клеток (72-74), ингибированием антиапоптотических путей (75-77), миграцией и адгезией клеток (73) и инвазией опухоли (78), среди других аспектов Биология клеток рака предстательной железы. На основании этих доклинических данных AZD0530 (саракатиниб) был допущен к клиническим испытаниям фазы II, но было продемонстрировано, что он имеет минимальную клиническую эффективность в качестве монотерапии (79).). Отсутствие клинической эффективности было также отмечено для дазатиниба; в фазе III клинических испытаний доцетаксела с дазатинибом или плацебо у пациентов с КРРПЖ, ранее не получавших химиотерапию, не было отмечено улучшения общей выживаемости (80). Другие нетирозинкиназы, такие как Btk и Etk в семействе Tek нетирозинкиназ, также являются мишенью; недавняя работа Guo и его коллег продемонстрировала, что CTN06, новый двойной ингибитор Btk и Etk, индуцирует апоптоз и аутофагию, а также ресенсибилизирует клеточные линии к доцетакселу (81).
Пути факторов роста, такие как IGF и KGF, связывают и активируют AR в кастрированном состоянии. Рецепторы факторов роста, такие как IGF-1R, IL-6R и EGFR, контролируют критические нижестоящие пути роста и выживания, такие как передача сигналов MAPK, PI3K/AKT и STAT. Различные RTK, такие как Her-2/neu, EGFR и IGR-1R, повышают стабильность и активность AR, а в некоторых случаях способствуют независимости от андрогенов. Например, было обнаружено, что Her2/neu способствует росту клеток ксенотрансплантата посредством Ack1-киназы, которая фосфорилирует AR по тирозину-267 и активирует его (82). Нацеливание на эти пути показало некоторые перспективы: кабозантиниб (XL-184) ингибирует тирозинкиназы c-Met и VEGF, а в фазе II клинических испытаний продемонстрировал значительное преимущество, особенно у пациентов с КРРПЖ с метастазами в кости; Тем не менее, он не достиг своей первичной конечной точки (облегчение боли в костях) в исследовании фазы III, при этом не было значимой разницы в облегчении боли в костях между группами лечения и контроля (митоксантрон/преднизолон).
Измененный стероидогенез
КРРПЖ развивается при кастрационном уровне циркулирующих андрогенов. Однако было установлено, что внутриопухолевые уровни андрогенов в моделях КРРПЖ такие же или даже выше, чем у эугонадных мужчин, что свидетельствует об альтернативной продукции андрогенов (18-22,33). Вероятно, это связано с выработкой надпочечниками, особенно предшественников андрогенов надпочечникового происхождения, таких как дегидроэпиандростерон (ДГЭА) и его сульфатированная форма (ДГЭА-С), которые могут быть преобразованы в высокоактивный ДГТ через «черный ход» (83,84). ).
ДГЭА и ДГЭА-С, вырабатываемые надпочечниками, не подвержены влиянию ГТ и все еще обнаруживаются в кровотоке. Молекулы превращаются в андростендион (AD) либо в предстательной железе, либо в надпочечниках с помощью 3βHSD, кодируемого HSD3B. Существует 2 изоформы: 3βHSD1 в предстательной железе и периферических тканях, 3βHSD2 в надпочечниках (85). Последующее преобразование АД в ДГТ в отсутствие АДТ обычно происходит через тестостерон в качестве посредника и требует 17βHSD3 и AKR1C3 (кодируется HSD17B3 и AKR1C3 соответственно) и стероид-5α-редуктазы (две изоформы, кодируемые SRD5A1 и SRD5A2). ). Однако в присутствии АДТ последовательность может быть обратной, что приводит к тому, что 5α-АД (5α-дион) служит посредником между АД и ДГТ, полностью минуя тестостерон. Было показано, что этот альтернативный путь, называемый «5α-дионовым», преобладает при КРРПЖ (86,87).
В дополнение к использованию слабых надпочечниковых андрогенов в пути 5α-диона, недавняя оценка клеток CRPC выявила повышенную экспрессию стероидогенных ферментов, таких как HSD3B1, HSD3B2, HSD17B3, AKR1C3 и SRD5A1 (20,87-89), которые могут способствовать к de novo производство стероидов и андрогенов. Повышающая регуляция SRD5A1 и одновременно понижающая регуляция SRD5A2 приводят к более высоким уровням 5α-редуктазы-1, для которой АД является лучшим субстратом, чем тестостерон (90–9).2). Что вызывает изменения в транскрипции этих стероидогенных ферментов? Многие однонуклеотидные полиморфизмы были идентифицированы в пределах вышеуказанных ферментов, особенно HSD3B1 и HSD3B2 (93), но их клиническое значение еще не ясно.
Варианты AR
Более поздней разработкой стала идентификация сплайс-вариантов AR (AR-V), которые являются конститутивно активными, как правило, из-за потери С-концевого LBD (94-97). Действительно, усиление AR, наблюдаемое при CRPC, может способствовать развитию вариантов сплайсинга. Большинство клеточных линий CRPC демонстрируют низкие уровни AR-V, но 22RV1 экспрессируют уровни, сходные с полноразмерными AR (44).
Функциональное значение этих вариантов еще полностью не изучено. Прямое измерение вариантов сплайсинга было ограничено отсутствием специфичных к варианту антител, оставляя только вторичную оценку по уровням РНК. ARV7 является единственным вариантом, который имеет подходящее антитело для окрашивания, а иммуногистохимическое окрашивание установило повышенную экспрессию при КРРПЖ (95,96). Однако уровни транскрибируемой РНК могут не полностью отражать уровни белка. Это предполагает некоторый посттрансляционный контроль, который еще полностью не выяснен.
Тем не менее, Hörnberg и др. . сообщили о высоких уровнях экспрессии вариантов сплайсинга в костных метастазах по сравнению с раком предстательной железы без гормонов, и что это приводило к КРРПЖ и худшему прогнозу (98). Это исследование также продемонстрировало несоответствие между уровнями РНК и уровнями белка, что усложняет определение значимости вариантов сплайсинга в развитии и прогрессировании КРРПЖ.
Преобладающими вариантами являются АРВ-1, АРВ-7 и АРВ-567. Из вариантов наиболее подробно изучен АРВ-7 (9).5,96). Как описано выше, у него отсутствует LBD, он расположен в ядре и конститутивно активен. Было показано, что он регулирует как AR-регулируемые гены, так и уникальный набор AR-независимых генов (96), что позволяет предположить, что он играет перекрывающуюся, но различную роль по сравнению с полноразмерным AR в клетках рака предстательной железы (94).
В недавно открытом варианте ARV8 фактически отсутствует ДНК-связывающий домен. Следовательно, он остается в плазматической мембране, и его конститутивная активность ограничивается активацией клеточных сигнальных путей (9). 9). Например, Ян и др. . продемонстрировали повышенное фосфорилирование AR посредством EGF-опосредованной активации SRC в присутствии этого варианта; его последующий нокдаун был связан с потерей этого фосфорилирования.
Механизмы резистентности к современным методам лечения КРРПЖ
Основываясь на этом понимании развития КРРПЖ, в настоящее время существуют одобренные лекарства для лечения пациентов, резистентных к кастрации. Однако, несмотря на эти новые агенты, у всех пациентов в конечном итоге будет прогрессировать заболевание. Понимание средств, с помощью которых рак предстательной железы преодолевает эти методы лечения, поможет определить новые варианты лечения.
Ниже мы рассмотрим основные агенты, доступные в настоящее время, сосредоточив внимание на механизме их действия и текущих знаниях о сопротивлении их функции. На рис. 2 представлен обзор существующих и экспериментальных препаратов, влияющих на андрогенную ось. Как и следовало ожидать, многие из этих механизмов пересекаются, и эти общие пути являются потенциально важными мишенями в будущем.
Рисунок 2 Текущие и экспериментальные препараты, нацеленные на андрогенную ось. wtAR, рецептор андрогена дикого типа; АРВ, вариант рецептора андрогена; LBD, лиганд-связывающий домен; NTD, n-концевой домен; Т, тестостерон; ДГТ, дигидротестостерон; ГСПГ, глобулин, связывающий половые гормоны.
Доцетаксел
Доцетаксел является текущим стандартом лечения пациентов, у которых прогрессировал кастрационно-резистентный рак предстательной железы. SWOG 9916 и TAX327 продемонстрировали преимущество доцетаксела в выживаемости в течение 3 месяцев по сравнению с митоксантроном у пациентов с КРРПЖ (100–102), и до недавнего времени это был единственный одобренный первичный метод лечения КРРПЖ. Однако недавнее исследование CHAARTED, возможно, продемонстрировало роль доцетаксела в качестве начального варианта лечения гормонально-наивного рака предстательной железы в сочетании с ГТ, поскольку было обнаружено, что эта комбинация обеспечивает 17-месячное преимущество в выживаемости (103).
Доцетаксел является хорошо известным и изученным химиотерапевтическим средством, используемым для лечения различных злокачественных новообразований. Это антимитотическое химиотерапевтическое средство, которое действует путем связывания β-субъединиц тубулина в микротрубочках, тем самым стабилизируя их и предотвращая деполимеризацию, необходимую для митоза (104-106), которая вызывает апоптоз. В частности, при КРРПЖ доцетаксел приводит к фосфорилированию bcl-2 (В-клеточная лимфома 2), что вызывает активацию каспаз и апоптоз in vivo и in vitro (107,108). Кроме того, экспрессия AR снижена в клетках CRPC, обработанных доцетакселом, и считается, что это связано с ядерной локализацией AR и ингибированием передачи сигналов (109).
Отток лекарств при КРРПЖ обеспечивает устойчивость к доцетакселу. Белки множественной лекарственной устойчивости (MDRP) хорошо описаны в литературе и включают P-гликопротеин (P-gp), белок множественной лекарственной устойчивости 1 (MRP1) и белок устойчивости к раку молочной железы (BRCP). Эти молекулы вызывают активный отток множества терапевтических агентов. Было обнаружено, что клеточные линии DU145 и 22RV1, устойчивые к доцетакселу, сверхэкспрессируют P-gp (110), в то время как линии CRPC, подвергшиеся воздействию доцетаксела, имеют генетические вариации MDR1, которые более устойчивы к доцетакселу (111). Линии КРРПЖ, устойчивые к доцетакселу, также активируют изоформу β-тубулина класса III, что позволяет меньше связывать таксаны. Ингибирование β-тубулина класса III восстанавливает чувствительность к доцетакселу в тех же клетках (112, 113). Кроме того, резистентные к доцетакселу клетки, полученные из LNCAP, продемонстрировали мутацию F270I в β-тубулине класса I, который на исходном уровне имел более сильное связывание с таксанами (114).
Хотя указанные выше механизмы специфичны для доцетаксела, были идентифицированы и другие механизмы резистентности. Резистентность к доцетакселу была связана с путями апоптоза, в частности, с активацией p53 и активацией PAR1. p53 является важным регулятором клеточного цикла, избыточная экспрессия которого часто обнаруживается при раке предстательной железы. Клетки LNCAP со сверхэкспрессией p53 дикого типа более устойчивы к активности доцетаксела, чем клеточные линии DU145 и PC3, у которых активность p53 снижена или отсутствует (115). Чжу и др. . продемонстрировали это на устойчивых к доцетакселу клетках C42B in vitro — клетки, обработанные доцетакселом, имели фосфорилирование и активацию p53, но устойчивые к таксану C42B фосфорилирования не продемонстрировали (116). Было показано, что PAR1 посредством активации NF-κB снижает апоптоз, индуцированный доцетакселом (117).
Помимо блокирования индуцированного доцетакселом апоптоза, антимитотическая активность доцетаксела сама по себе непосредственно инициирует пути выживания в клеточных линиях рака предстательной железы. Связывание с микротрубочками инициирует такие пути, как N-концевая киназа c-Jun (JNK), которая, в свою очередь, приводит к активации различных факторов транскрипции, таких как STAT-1, STAT-3 и NF-κB. Было показано, что модели с нокдауном этих факторов транскрипции более чувствительны к цитотоксичности доцетаксела (115, 118).
Повышенная экспрессия цитокинов и хемокинов, таких как IL-6, IL-8 и CCL-2, и молекул шаперонов, таких как HSP27 и HSP90, была связана с резистентностью к доцетакселу, но клинически значимые ингибиторы этих метаболических путей не проявлялись. все же идентифицированы. OGX-011, антисмысловой препарат второго поколения, который ингибирует секрецию кластерина, белка-шаперона, вводили в сочетании с доцетакселом в исследованиях фазы III, но не достигли своей первичной конечной точки. Его активность сосредоточена на CLU, ключевом белке, который существует в двух формах: ядерной CLU (nCLU) и секретируемой CLU (sCLU).,120). После начала химиотерапии, особенно доцетакселом, в клетках рака предстательной железы наблюдается сдвиг баланса в сторону sCLU, что, как считается, связано с активацией STAT-1 (110, 121). Однако ингибирование sCLU с помощью антисмыслового олигонуклеотида повышает чувствительность клеток к доцетакселу (121, 122).
Наша лабораторная группа идентифицировала более 1600 генов с измененной экспрессией в устойчивых к таксанам клетках C42B, причем примерно 52% были активизированы. Из этого подмножества мы недавно идентифицировали ABCB1, который принадлежит к семейству переносчиков АТФ-связывающей кассеты (ABC), среди генов с максимальной активацией в устойчивых к таксанам клетках. ABCB1 был высоко экспрессирован в устойчивых к таксанам клетках C42B, но практически не обнаруживался в чувствительных к таксанам клетках C42B. Ингибирование экспрессии ABCB1 повышало чувствительность клеток C42B к доцетакселу, что затем было подтверждено в клеточной линии DU-145 (116). Было показано, что апигенен, природная молекула из семейства флавонов, идентифицированная Шукла и Гупта (123), способствует повышению чувствительности клеток к терапии доцетакселом.
Абиратерон и ингибиторы синтеза андрогенов
Абиратерона ацетат (Zytiga) представляет собой молекулу, структурно сходную с прегненолоном, которая действует как необратимый ингибитор цитохрома p450, семейства 17, подсемейства A, полипептида 1 (CYP17A1). CYP17A1 является членом класса ферментов цитохрома p450, которые служат катализатором окисления различных молекул. Он имеет две последовательные ферментативные функции в пути стероидогенеза, которые способствуют превращению прегненолона в ДГТ. Потеря активности CYP17A1 вызывает значительную потерю продукции андрогенов в периферических органах, особенно андрогенов надпочечниками. Было обнаружено, что он в 10-30 раз более эффективен, чем кетоконазол, который является неспецифическим ингибитором ферментов p450 и ранее использовался для быстрой абляции андрогенов (106). Испытание фазы III COU-AA-301 продемонстрировало 3,9Преимущество абиратерона/преднизона в месячной выживаемости по сравнению с плацебо/преднизоном у пациентов, у которых на фоне терапии доцетакселом наблюдалось прогрессирование заболевания (124). Последующее исследование COU-AA-302 продемонстрировало преимущества до химиотерапии, с улучшением выживаемости без рентгенологического прогрессирования, времени до начала химиотерапии и тенденцией к улучшению общей выживаемости (125).
Измененный стероидогенез обсуждался как один из механизмов развития КРРПЖ. В то время как клеточные линии КРРПЖ, не получавшие абиратерон, используют путь «5α-диона» для выработки внутриопухолевого ДГТ в обход тестостерона, они все еще зависят от надпочечниковых андрогенов. Необратимо ингибируя этот критический вышестоящий фермент в пути стероидогенеза, абиратерон эффективно вызывает значительное снижение внутриопухолевых уровней андрогенов, предотвращая продукцию надпочечниковых андрогенов.
Однако, несмотря на его эффективность в ингибировании пути стероидогенеза (126), действие абиратерона является неполным. Аттард и др. . продемонстрировали, что в то время как большинство метаболитов андрогенов в моче и андрогенов в сыворотке были подавлены, ингибирование CYP17 приводило к более высоким уровням мочевого метаболита 3α5α-17HP, что коррелировало с экскрецией андростерона, который является основным метаболитом 5α-восстановленных андрогенов, таких как ДГТ. 127). Это говорит о том, что использование абиратерона может подтолкнуть 17-гидроксипрогестерон к пути «5α-диона».
Как и следовало ожидать, избыточная экспрессия или мутации CYP17A1 также могут способствовать резистентности к абиратерону (128). Чанг и др. . продемонстрировали, что мутация HSD3B1 (1245C), ранее упоминавшаяся как способствующая прогрессированию КРРПЖ, также была обнаружена в моделях ксенотрансплантатов, устойчивых к абиратерону, хотя клиническое значение этого еще предстоит выяснить (93). Мостагель и др. . продемонстрировали, что клеточные линии, обработанные абиратероном, реагировали повышенной экспрессией CYP17A1, а также повышенной экспрессией ферментов пути стероидогенеза, включая AKR1C3 и HSD17B3 (129).).
В настоящее время разрабатываются другие ингибиторы синтеза андрогенов, в том числе TAK-700 (Orteronel) и VT-464 (Viamet), оба из которых более селективны в отношении ингибирования 17, 20-лиазы (130). TAK-700 находится в дальнейшей разработке, в настоящее время накапливается для другого клинического испытания фазы III, на этот раз оценивающего эффективность у пациентов с КРРПЖ, ранее не получавших химиотерапию; начальное исследование III фазы у пациентов, получавших доцетаксел, продемонстрировало улучшение рентгенологической выживаемости без прогрессирования заболевания (ОР 0,755), но оно не соответствовало первичной конечной точке улучшения общей выживаемости (ОР 0,89). 4) (130).
Энзалутамид и ингибиторы рецепторов андрогенов (AR)
В ответ на обнаружение множества опосредованных AR механизмов резистентности, ведущих к развитию КРРПЖ, было разработано новое поколение ингибиторов передачи сигналов рецепторов андрогенов. Основным агентом этого класса является энзалутамид (MDV-3100, ENZA, Xtandi), который, как было продемонстрировано, обладает многосторонним подходом — предотвращением связывания тестостерона с AR, ядерной транслокацией AR, связыванием AR с ДНК и рекрутированием коактиватора. (106). В то время как исследование AFFIRM III продемонстрировало 4,8-месячное преимущество выживаемости по сравнению с плацебо у пациентов с КРРПЖ, у которых доцетаксел оказался неэффективным, а исследование PREVAIL продемонстрировало общую выживаемость и выживаемость без рентгенологического прогрессирования по сравнению с плацебо у пациентов с КРРПЖ, ранее не получавших химиотерапию (131,132), не все пациенты получили пользу от лечения — у подгруппы пациентов продолжалось прогрессирование, что указывает на наличие значительных механизмов резистентности, которые необходимо выявить и устранить.
Одним из механизмов развития устойчивости КРРПЖ к энзалутамиду и потенциально другим методам лечения является процесс аутофагии. Аутофагия представляет собой катаболический процесс, который, помимо конститутивной активности при низком базальном уровне, активируется в ответ на стрессоры, позволяя клеткам использовать лизосомно-опосредованную деградацию клеточных белков и органелл для регенерации энергии (133-135). Аутофагия может использоваться раковыми клетками для продления их выживания в суровых условиях метаболического стресса в микроокружении опухоли, вызванного различными методами лечения, но чрезмерная или нерегулируемая аутофагия может подтолкнуть клетки к аутофагической гибели клеток или запрограммированной гибели клеток типа II (136, 137). . Действительно, было показано, что андрогенная депривация вызывает аутофагию, и, хотя точный механизм неизвестен, подавление mTOR, по-видимому, играет критическую роль (135, 138). Предыдущие исследования, проведенные нашей группой и другими, установили, что введение ингибиторов аутофагии либо в виде монотерапии, либо в сочетании с общепризнанной терапией дает эффективный цитотоксический результат. Мы продемонстрировали, что использование кломипрамина и метформина, обоих клинических ингибиторов аутофагии, значительно увеличивало цитотоксичность, связанную с энзалутамидом 9.1543 in vitro и на мышах — комбинация энзалутамид/кломипрамин уменьшала размер опухоли на 91% по сравнению с уменьшением на 78% при применении энзалутамида/метформина (135). В настоящее время проводится множество клинических испытаний, оценивающих роль ингибиторов аутофагии в качестве сопутствующей терапии (139), в том числе исследование в нашем учреждении, которое недавно было одобрено для оценки комбинированной терапии метформином и энзалутамидом.
AR также являются важными механизмами устойчивости к энзалутамиду, как и при развитии КРРПЖ. Сообщалось, что мутация Phe876Leu в LBD AR превращает энзалутамид в агонист AR, хотя клиническая значимость этого изменения не была задокументирована (140, 141). Подобные эффекты были отмечены для антиандрогенов первого поколения бикалутамида и флутамида.
Другим предполагаемым механизмом является путь «захвата глюкокортикоидного рецептора». Глюкокортикоидные рецепторы представляют собой ядерные рецепторы, сходные по структуре с АР. Глюкокортикоиды изначально оказывают подавляющее действие на рак предстательной железы и действительно часто назначаются в сочетании с ранним лечением КРРПЖ, включая химиотерапию и абиратерон. Однако ДНК-связывающий домен глюкокортикоидного рецептора очень похож на DBD AR (142, 143), и было показано, что глюкокортикоидный рецептор связывается со многими генами, регулируемыми AR, что позволяет предположить, что его повышенная регуляция у пациентов, получающих химиотерапию или ADT, может способствовать к устойчивости к энзалутамиду (144).
Многие из этих механизмов могут аналогичным образом воздействовать на будущие ингибиторы рецепторов андрогенов. Например, было показано, что ARN-509, еще один новый антагонист AR, который в настоящее время находится в фазе накопления в многоцентровом клиническом исследовании фазы III, чувствителен к той же мутации AR F876L, которая превращает его в агонист (145). Другие агенты, разрабатываемые в настоящее время, включают ODM-201.
Как недавно было опубликовано в New England Journal of Medicine , Antonarakis и соавторы продемонстрировали, что 20-40% циркулирующих опухолевых клеток у пациентов с КРРПЖ, получавших абиратерон и энзалутамид, имеют конститутивно активную АРВ7 (146). Однако, что еще более важно, они продемонстрировали в этом проспективном исследовании, что подгруппа мужчин с АРВ7 в циркулирующих опухолевых клетках имела значительно более низкую частоту ответа ПСА, более короткую выживаемость без прогрессирования и общую выживаемость по сравнению с мужчинами без экспрессии АРВ7. Это исследование, наше собственное исследование (62) и исследования других групп (94,145-147) демонстрируют, что АРВ являются важным механизмом устойчивости к новым агентам КРРПЖ. Лю и др. . продемонстрировали, что AR-V7 присутствует в ряде клеточных линий рака предстательной железы и что он способен активировать промотор PSA в клетках LNCaP и PC3 в отсутствие андрогена (148). С потерей LBD на AR, как видно в ARV-7, клеточные линии CRPC преодолевают потерю циркулирующих и внутриопухолевых андрогенов, опосредованную абиратероном. Потеря LBD и параллельное лиганд-независимое связывание AR с ARE считается основным механизмом резистентности к энзалутамиду. Ли и др. . продемонстрировали, что нокдаун AR-V ограничивает андроген-независимую скорость роста клеток CWR22Rv1 и восстанавливает чувствительность к антиандрогенам (147).
С растущим объемом данных, указывающих на важную роль АРВ-препаратов в развитии резистентности, и одновременным обнаружением того, что многие из существующих механизмов прогрессирования в КРРПЖ включают изменения в пути АР, воздействие на АР, по-видимому, станет следующим важным шагом. в терапии рака предстательной железы.
Наша лаборатория ранее идентифицировала никлозамид, клинически используемый для лечения гельминтозов, в качестве ингибитора АРВ-7, способствуя его деградации; совместное лечение энзалутамидом продемонстрировало синергический эффект (148). Точно так же мы также установили, что miR-let-7c, микроРНК семейства let-7, противодействует экспрессии AR посредством деградации c-myc, что приводит к ингибированию пролиферации рака предстательной железы (149). Другие также начали сосредотачиваться на самом АР как на мишени для терапии — либо уменьшая его экспрессию, либо способствуя его деградации. Лай и др. . идентифицировали ASC-J9, новый усилитель деградации AR, который в настоящее время используется клинически для лечения других патологий (150). Садар и его коллеги сосредоточились на EPI-001, небольшой молекуле, которая ингибирует N-концевой домен (NTD), который присутствует как в AR дикого типа, так и в вариантах AR (151).
Возможно, нацелившись на АР и его разновидности, мы сможем преодолеть недостатки современных методов лечения КРРПЖ.
Выводы
Рак предстательной железы, особенно местнораспространенный и метастатический, продолжает оставаться тяжелым бременем для системы здравоохранения. В то время как прогноз хороший для мужчин с диагнозом локализованного заболевания, прогноз остается плохим для мужчин с более поздними стадиями заболевания. Все современные методы лечения, от ADT до химиотерапии, просто замедляют прогрессирование заболевания, но все пациенты неизбежно прогрессируют на терапии. Понимание механизмов, посредством которых у этих пациентов развивается резистентность к ADT, а затем к доцетакселу, абиратерону и энзалутамиду, важно для определения будущих целей терапии.
Благодарности
Финансирование: Эта работа частично поддерживается грантами DOD PC111467 и Medivation/Astellas для CPE, NIH RO1 CA 165263−13 для H-JK и организацией Stand Up To Cancer — Prostate Cancer Foundation — Prostate Dream Team Translation Research Cancer Research Предоставьте SU2C-AACR-PCF DT0812 Эрику Смоллу, Оуэну Витте и CPE. Этот исследовательский грант стал возможен благодаря щедрой поддержке фонда Movember. Stand Up To Cancer — это программа Фонда индустрии развлечений, администрируемая Американской ассоциацией исследований рака: расходы на публикацию этой статьи были покрыты.
Сноска
Конфликт интересов: У авторов нет конфликта интересов, о котором следует заявить.
Каталожные номера
- Сигел Р., Ма Дж., Цзоу З. и др. Статистика рака, 2014 г. CA Cancer J Clin 2014; 64:9-29. [ПубМед]
- Ferlay J, Stelarova-Foucher E, Lortet-Tieulent J, et al. Модели заболеваемости и смертности от рака в Европе: оценки для 40 стран в 2012 г. Eur J Cancer 2013;49: 1374-403. [ПубМед]
- Studer UE, Hauri D, Hanselmann S, et al. Немедленная и отсроченная гормональная терапия у пациентов с раком предстательной железы, которым противопоказана местная лечебная терапия: результаты рандомизированного исследования SAKK 08/88. Дж. Клин Онкол 2004; 22:4109-18. [ПубМед]
- Harris WP, Mostaghel EA, Nelson PS, et al. Терапия депривации андрогенов: прогресс в понимании механизмов резистентности и оптимизации истощения андрогенов. Нат Клин Практ Урол 2009;6:76-85. [ПубМед]
- Hughes IA, Davies JD, Bunch TI, et al. Синдром нечувствительности к андрогенам. Ланцет 2012; 380:1419-28. [ПубМед]
- Шафи А.А., Йен А.Е., Вейгель Н.Л. Рецепторы андрогенов при гормонозависимом и кастрационно-резистентном раке предстательной железы. Pharmacol Ther 2013;140:223-38. [ПубМед]
- Гельманн ЕР. Молекулярная биология рецептора андрогенов. Дж. Клин Онкол 2002; 20:3001-15. [ПубМед]
- Ким Ю.С., Аларкон С.В., Ли С. и др. Обновление Hsp90 ингибиторов в клинических испытаниях. Curr Top Med Chem 2009; 9:1479-92. [ПубМед]
- Koochekpour S. Передача сигналов рецептора андрогена и мутации при раке предстательной железы. Азиат Дж. Андрол 2010; 12: 639-57. [ПубМед]
- Химерс Х.В., Тиндалл Диджей. Корегуляторы рецепторов андрогенов (AR): разнообразие функций, сходящихся и регулирующих транскрипционный комплекс AR. Endocr Rev 2007; 28:778-808. [ПубМед]
- Фельдман Б.Дж., Фельдман Д. Развитие андроген-независимого рака предстательной железы. Nat Rev Рак 2001; 1:34-45. [ПубМед]
- Линдзи Дж. , Кумар М.В., Гроссман М. и др. Молекулярные механизмы действия андрогенов. Витам Хорм 1994;49:383-432. [ПубМед]
- Рой А.К., Лавровский Ю.С., Сонг С.С. и др. Регуляция действия андрогенов. Витам Хорм 1999;55:309-52. [ПубМед]
- Кунья Г.Р., Кук П.С., Курита Т. Роль стромально-эпителиальных взаимодействий в гормональных реакциях. Arch Histol Cytol 2004;67:417-34. [ПубМед]
- Ниу Ю., Алтувайри С., Йех С. и др. Ориентация на стромальные рецепторы андрогенов при первичных опухолях предстательной железы на более ранних стадиях. Proc Natl Acad Sci U S A 2008; 105:12188-93. [ПубМед]
- Ю Дж., Ю Дж., Мани Р.С. и др. Интегрированная сеть слияний генов рецепторов андрогенов, polycomb и TMPRSS2-ERG при прогрессировании рака предстательной железы. Раковая клетка 2010; 17:443-54. [ПубМед]
- Грин С.М., Мостагель Э.А., Нельсон П.С. Действие и метаболизм андрогенов при раке предстательной железы. Mol Cell Endocrinol 2012;360:3-13. [ПубМед]
- Геллер Дж. , Альберт Дж., Нахтсхайм Д. и др. Уровни стероидов при раке предстательной железы — маркеры опухолевой дифференцировки и адекватности антиандрогенной терапии. Прог Клин Биол Рез 1979;33:103-11. [ПубМед]
- Mohler JL, Gregory CW, Ford OH 3rd, et al. Андрогенная ось при рецидивирующем раке предстательной железы. Clin Cancer Res 2004; 10:440-8. [ПубМед]
- Монтгомери Р.Б., Мостагель Э.А., Весселла Р. и соавт. Поддержание внутриопухолевых андрогенов при метастатическом раке предстательной железы: механизм кастрационно-резистентного роста опухоли. Рак Res 2008;68:4447-54. [ПубМед]
- Massie CE, Lynch A, Ramos-Montoya A, et al. Рецептор андрогена питает рак предстательной железы, регулируя центральный метаболизм и биосинтез. Эмбо Дж. 2011; 30:2719-33. [ПубМед]
- Мостагель Э.А. Абиратерон в лечении метастатического резистентного к кастрации рака предстательной железы. Cancer Manag Res 2014; 6:39-51. [ПубМед]
- Хаггинс С., Ходжес CV. Исследования рака предстательной железы. I. Влияние кастрации, введения эстрогенов и андрогенов на фосфатазы сыворотки при метастатическом раке предстательной железы. CA Cancer J Clin 1972; 22:232-40. [ПубМед]
- Кроуфорд ЭД, Хоу АХ. Роль антагонистов LHRH в лечении рака предстательной железы. Онкология (Уиллистон Парк) 2009 г.;23:626-30. [ПубМед]
- Агарвал Н., Хуссейн М. Лечение гормоночувствительного метастатического рака предстательной железы. Hematol Oncol Clin North Am 2013;27:1221-41. VIII. [ПубМед]
- Гроссманн М., Чунг А.С., Заджак Д.Д. Андрогены и рак предстательной железы; патогенез и депривационная терапия. Best Pract Res Clin Endocrinol Metab 2013; 27: 603-16. [ПубМед]
- Саад Ф., Хотте С.Дж. Руководство по лечению резистентного к кастрации рака предстательной железы. Can Urol Assoc J 2010;4:380-4. [ПубМед]
- Cookson MS, Roth BJ, Dahm P, et al. Кастрационно-резистентный рак предстательной железы: Руководство AUA. Дж Урол 2013;190:429-38. [ПубМед]
- Scher HI, Liebertz C, Kelly WK, et al. Бикалутамид при распространенном раке предстательной железы: история естественного и леченного заболевания. Дж. Клин Онкол 1997; 15:2928-38. [ПубМед]
- Fosså SD, Slee PH, Brausi M, et al. Флутамид по сравнению с преднизоном у пациентов с симптоматически прогрессирующим раком предстательной железы после андрогенабляционной терапии: исследование III фазы Европейской организации по изучению и лечению рака мочеполовой группы. Дж. Клин Онкол 2001; 19: 62-71. [ПубМед]
- Келли В.К., Шер Х.И. Снижение специфического антигена простаты после отмены антиандрогенов: синдром отмены флутамида. Дж. Урол 1993; 149:607-9. [ПубМед]
- Culig Z, Hoffmann J, Erdel M, et al. Переход от антагониста к агонисту рецептора андрогенов бикалутамида связан с прогрессированием опухоли предстательной железы в новой модельной системе. Бр Дж Рак 1999;81:242-51. [ПубМед]
- Kumagai J, Hofland J, Erkens-Schulze S, et al. Внутриопухолевая конверсия предшественников андрогенов в надпочечниках стимулирует рост клеток, активируемый рецепторами андрогенов, при раке предстательной железы более сильно, чем стероидогенез de novo. Простата 2013;73:1636-50. [ПубМед]
- Visakorpi T, Hyytinen E, Koivisto P, et al. Амплификация in vivo гена рецептора андрогенов и прогрессирование рака предстательной железы человека. Нат Жене 1995;9:401-6. [ПубМед]
- Gregory CW, Johnson RT Jr, Mohler JL, et al. Стабилизация рецепторов андрогенов при рецидивирующем раке предстательной железы связана с гиперчувствительностью к низким андрогенам. Рак Res 2001; 61: 2892-8. [ПубМед]
- Чен Ю, Сойерс CL, Шер HI. Ориентация на путь рецептора андрогена при раке предстательной железы. Curr Opin Pharmacol 2008;8:440-8. [ПубМед]
- Лю В., Се К.С., Чжу Ю. и др. Гомозиготные делеции и повторяющиеся амплификации связаны с новыми генами, участвующими в развитии рака предстательной железы. Неоплазия 2008; 10:897-907. [ПубМед]
- Taylor BS, Schultz N, Hieronymus H, et al. Интегративное геномное профилирование рака простаты человека. Раковая клетка 2010; 18:11-22. [ПубМед]
- Макридакис Н.М., ди Салле Э. , Райхардт Дж.К. Биохимическое и фармакогенетическое вскрытие стероидной 5 альфа-редуктазы человека II типа. Фармакогенетика 2000;10:407-13. [ПубМед]
- Скариано Дж.К., Трет Э., Альба Ф. и др. Полиморфизм V89L SRD5A2 связан с тяжестью заболевания у мужчин с ранним началом рака предстательной железы. Простата 2008;68:1798-805. [ПубМед]
- Сак Дж.С., Киш К.Ф., Ван С. и др. Кристаллографические структуры лиганд-связывающих доменов рецептора андрогенов и его мутанта T877A в комплексе с природным агонистом дигидротестостероном. Proc Natl Acad Sci U S A 2001;98:4904-9. [ПубМед]
- Судзуки Х., Акакура К., Комия А. и др. Мутация кодона 877 в гене рецептора андрогенов при распространенном раке предстательной железы: связь с синдромом отмены антиандрогенов. простата 1996;29:153-8. [ПубМед]
- Steketee K, Timmerman L, Ziel-van der Made AC, et al. Расширенная чувствительность к лигандам мутантов рецептора андрогенов, полученных путем случайной аминокислотной замены H874 и горячей точки мутации T877 при раке предстательной железы. Int J Рак 2002; 100: 309-17. [ПубМед]
- Dehm SM, Schmidt LJ, Heemers HV, et al. Сплайсинг нового экзона рецептора андрогена генерирует конститутивно активный рецептор андрогена, который опосредует резистентность к терапии рака предстательной железы. Рак Res 2008; 68: 5469-77. [ПубМед]
- ван де Вейнгарт Д.Дж., Мольер М., Люшер С.Дж. и др. Систематический структурно-функциональный анализ мутантов рецептора андрогенов Leu701 объясняет свойства мутанта рака предстательной железы L701H. J Biol Chem 2010; 285:5097-105. [ПубМед]
- Thompson J, Saatcioglu F, Janne OA, et al. Нарушение амино- и карбоксиконцевых взаимодействий рецептора андрогенов связано с нечувствительностью к андрогенам. Мол Эндокринол 2001;15:923-35. [ПубМед]
- Хара Т., Коно Дж., Накамура К. и др. Возможная роль адаптивной мутации в резистентности к антиандрогенам клеток рака предстательной железы. Простата 2005;65:268-75. [ПубМед]
- Gottlieb B, Beitel LK, Wu JH, et al. База данных мутаций генов рецепторов андрогенов (ARDB): обновление 2004 г. Хум Мутат 2004; 23:527-33. [ПубМед]
- Gottlieb B, Beitel LK, Nadarajah A, et al. База данных мутаций генов рецепторов андрогенов: обновление 2012 г. Хум Мутат 2012; 33:887-94. [ПубМед]
- Wolf IM, Heitzer MD, Grubisha M, et al. Коактиваторы и трансактивация ядерных рецепторов. J Cell Biochem 2008;104:1580-6. [ПубМед]
- Хермансон О, Гласс К.К., Розенфельд М.Г. Корегуляторы ядерных рецепторов: множественные способы модификации. Тенденции Endocrinol Metab 2002;13:55-60. [ПубМед]
- Агульник И.Ю., Вейгель Н.Л. Коактиваторы рецепторов андрогенов и рак предстательной железы. Adv Exp Med Biol 2008;617:245-55. [ПубМед]
- Ni L, Yang CS, Gioeli D, et al. FKBP51 способствует сборке Hsp90 шапероновый комплекс и регулирует передачу сигналов рецептора андрогена в клетках рака предстательной железы. Мол Селл Биол 2010;30:1243-53. [ПубМед]
- Wang Q, Carroll JS, Brown M. Пространственное и временное рекрутирование рецептора андрогена и его коактиваторов включает хромосомное образование петель и отслеживание полимеразы. Мол Селл 2005;19:631-42. [ПубМед]
- Сюй Дж., Ву Р.К., О’Мэлли Б.В. Нормальные и связанные с раком функции семейства коактиваторов стероидных рецепторов p160 (SRC). Нат Рев Рак 2009;9:615-30. [ПубМед]
- Уэда Т., Мавджи Н.Р., Бруховский Н. и др. Лиганд-независимая активация рецептора андрогена интерлейкином-6 и роль коактиватора-1 стероидного рецептора в клетках рака предстательной железы. J Biol Chem 2002; 277:38087-94. [ПубМед]
- Чунг А.С., Чжоу С., Ляо Л. и др. Генетическая абляция амплифицированного рака молочной железы 1 ингибирует спонтанное прогрессирование рака простаты у мышей. Рак Res 2007; 67: 5965-75. [ПубМед]
- Debes JD, Schmidt LJ, Huang H, et al. p300 опосредует андроген-независимую трансактивацию рецептора андрогена интерлейкином 6. Cancer Res 2002;62:5632-6. [ПубМед]
- Wissmann M, Yin N, Muller JM, et al. Совместное деметилирование с помощью JMJD2C и LSD1 способствует экспрессии генов, зависимых от рецепторов андрогенов. Nat Cell Biol 2007;9:347-53. [ПубМед]
- Ван К., Ли В., Чжан Ю. и др. Рецептор андрогена регулирует отдельную программу транскрипции при андроген-независимом раке предстательной железы. Сотовый 2009; 138: 245-56. [ПубМед]
- Н. Надиминти, В. Лу, С.О. Ли и др. Активация Stat3 процессинга NF-{kappa}B p100 включает ацетилирование, опосредованное CBP/p300. Proc Natl Acad Sci U S A 2006; 103:7264-9. [ПубМед]
- Н. Надиминти, Р. Туммала, С. Лю и др. NF-kappaB2/p52 индуцирует устойчивость к энзалутамиду при раке предстательной железы: роль рецептора андрогена и его вариантов. Мол Рак Тер 2013; 12:1629-37. [ПубМед]
- Штивельман Э., Пиво ТМ, Эванс С.П. Молекулярные пути и мишени при раке предстательной железы. Онкотаргет 2014; 5:7217-59. [ПубМед]
- Торен П., Зубейди А. Ориентация на путь PI3K/Akt при раке предстательной железы: проблемы и возможности Int J Oncol 2014;45:1793-801. (Обзор). [ПубМед]
- Фруман Д.А., Роммель К. PI3K и рак: уроки, проблемы и возможности. Nat Rev Drug Discov 2014; 13:140-56. [ПубМед]
- Цзян X, Чен С, Асара Дж. М. и др. Активация пути фосфоинозитид-3-киназы в клетках рака предстательной железы с дефицитом гомолога фосфата и тензина (PTEN) не зависит от рецепторных тирозинкиназ и опосредована каталитическими субъединицами p110beta и p110delta. J Biol Chem 2010; 285:14980-9. [ПубМед]
- Xin L, Teitell MA, Lawson DA, et al. Прогрессирование рака предстательной железы за счет синергизма АКТ с генотропным и негенотропным действием рецептора андрогена. Proc Natl Acad Sci U S A 2006;103:7789-94. [ПубМед]
- Vlaeminck-Guillem V, Gillet G, Rimokh R. SRC: маркер или фактор агрессивности рака предстательной железы. Фронт Онкол 2014; 4:222. [ПубМед]
- Боггон Т.Дж., Экк М.Дж. Структура и регуляция киназ семейства Src. Онкоген 2004; 23:7918-27. [ПубМед]
- Чанг Ю.М., Бай Л., Лю С. и др. Онкогенный потенциал и пути киназ семейства Src при раке предстательной железы, как показано AZD0530. Онкоген 2008;27:6365-75. [ПубМед]
- Чанг Ю. М., Кунг Х.Дж., Эванс С.П. Нерецепторные тирозинкиназы при раке предстательной железы. Неоплазия 2007;9:90-100. [ПубМед]
- Lee LF, Guan J, Qiu Y и др. Нейропептид-индуцированная независимость от андрогенов в клетках рака предстательной железы: роль нерецепторных тирозинкиназ Etk/Bmx, Src и киназы фокальной адгезии. Мол Селл Биол 2001;21:8385-97. [ПубМед]
- Ли Л.Ф., Луи М.К., Десаи С.Дж. и др. Интерлейкин-8 обеспечивает андроген-независимый рост и миграцию LNCaP: дифференциальные эффекты тирозинкиназ Src и FAK. Онкоген 2004;23:2197-205. [ПубМед]
- Гонг Дж., Чжу Дж., Гудман О.Б. мл. и др. Активация активности гистон-ацетилтрансферазы p300 и ацетилирование рецептора андрогена бомбезином в клетках рака предстательной железы. Онкоген 2006;25:2011-21. [ПубМед]
- Фань С., Гао М., Мэн К. и др. Роль передачи сигналов NF-kappaB в защите клеток, опосредованной фактором роста/рассеяния гепатоцитов. Онкоген 2005;24:1749-66. [ПубМед]
- Сумитомо М., Шен Р. , Голдберг Дж. С. и др. Нейтральная эндопептидаза способствует индуцированному форболовым эфиром апоптозу в клетках рака предстательной железы путем ингибирования нейропептид-индуцированной деградации протеинкиназы С-дельта. Рак Res 2000;60:6590-6. [ПубМед]
- Унни Э., Сан С., Нан Б. и др. Изменения в негенотропной передаче сигналов андрогеновых рецепторов коррелируют с переходом клеток LNCaP к независимости от андрогенов. Рак Res 2004; 64: 7156-68. [ПубМед]
- Recchia I, Rucci N, Festuccia C и др. Пирролопиримидиновые ингибиторы c-Src уменьшают рост, адгезию, подвижность и инвазию клеток рака предстательной железы in vitro. Европейский Рак J 2003; 39: 1927-35. [ПубМед]
- Лара П.Н. мл., Лонгмейт Дж., Эванс С.П. и др. Испытание фазы II ингибитора Src-киназы AZD0530 у пациентов с прогрессирующим раком предстательной железы, резистентным к кастрации: исследование Калифорнийского онкологического консорциума. Противораковые препараты 2009; 20:179-84. [ПубМед]
- Araujo JC, Trudel GC, Saad F, et al. Доцетаксел и дазатиниб или плацебо у мужчин с метастатическим резистентным к кастрации раком предстательной железы (READY): рандомизированное двойное слепое исследование фазы 3. Ланцет Онкол 2013;14:1307-16. [ПубМед]
- Guo W, Liu R, Bhardwaj G, et al. Воздействие на клетки рака предстательной железы Btk/Etk с помощью нового двойного ингибитора. Cell Death Dis 2014;5:e1409. [ПубМед]
- Вэнь И., Ху М.С., Макино К. и др. HER-2/neu способствует независимому от андрогенов выживанию и росту клеток рака предстательной железы посредством пути Akt. Рак Res 2000;60:6841-5. [ПубМед]
- Chang KH, Ercole CE, Sharifi N. Метаболизм андрогенов при раке предстательной железы: от молекулярных механизмов до клинических последствий. Бр Дж Рак 2014;111:1249-54. [ПубМед]
- Шарифи Н. Миниобзор: Метаболизм андрогенов при резистентном к кастрации раке предстательной железы. Мол Эндокринол 2013;27:708-14. [ПубМед]
- Simard J, Ricketts ML, Gingras S, et al. Молекулярная биология семейства генов 3бета-гидроксистероиддегидрогеназы/дельта5-дельта4 изомеразы. Endocr Rev 2005; 26:525-82. [ПубМед]
- Чанг К.Х., Ли Р., Папари-Зари М. и др. Синтез дигидротестостерона обходит тестостерон, вызывая резистентный к кастрации рак предстательной железы. Proc Natl Acad Sci U S A 2011;108:13728-33. [ПубМед]
- Йепуру М., Ву З., Кулкарни А. и др. Стероидогенный фермент AKR1C3 представляет собой новый селективный коактиватор андрогенных рецепторов, который способствует росту рака предстательной железы. Clin Cancer Res 2013;19:5613-25. [ПубМед]
- Титус М.А., Шелл М.Дж., Лих Ф.Б. и др. Уровни тестостерона и дигидротестостерона в тканях при рецидивирующем раке предстательной железы. Clin Cancer Res 2005; 11:4653-7. [ПубМед]
- Stanbrough M, Bubley GJ, Ross K, et al. Повышенная экспрессия генов, превращающих андрогены надпочечников в тестостерон, при андроген-независимом раке предстательной железы. Рак Res 2006; 66: 2815-25. [ПубМед]
- Тигпен А.Е., Кала К.М., Рассел Д.В. Характеристика клеточных линий яичников китайского хомячка, экспрессирующих изоферменты стероида 5 альфа-редуктазы человека. J Biol Chem 1993; 268:17404-12. [ПубМед]
- Рассел Д.В., Уилсон Д.Д. Стероидная 5-альфа-редуктаза: два гена/два фермента. Annu Rev Biochem 1994;63:25-61. [ПубМед]
- Локк Дж.А., Ганс Э.С., Лубик А.А. и др. Уровни андрогенов увеличиваются за счет внутриопухолевого стероидогенеза de novo во время прогрессирования резистентного к кастрации рака предстательной железы. Рак Res 2008; 68: 6407-15. [ПубМед]
- Чанг К.Х., Ли Р., Кури Б. и др. Мутация с усилением функции в синтезе ДГТ при резистентном к кастрации раке предстательной железы. Сотовый 2013; 154: 1074-84. [ПубМед]
- Дехм С.М., Тиндалл Диджей. Альтернативно сплайсированные варианты рецептора андрогена. Endocr Relat Cancer 2011;18:R183-96. [ПубМед]
- Guo Z, Yang X, Sun F и др. Новый вариант сплайсинга рецептора андрогена активируется во время прогрессирования рака предстательной железы и способствует росту, устойчивому к истощению андрогенов. Рак Res 2009;69:2305-13. [ПубМед]
- Hu R, Dunn TA, Wei S, et al. Лиганд-независимые варианты рецепторов андрогенов, полученные в результате сплайсинга криптических экзонов, означают гормонорефрактерный рак предстательной железы. Рак Res 2009;69:16-22. [ПубМед]
- Sun S, Sprenger CC, Vessella RL, et al. Устойчивость к кастрации при раке предстательной железы человека обеспечивается часто встречающимся сплайс-вариантом рецептора андрогена. J Clin Invest 2010;120:2715-30. [ПубМед]
- Хёрнберг Э., Юлитало Э.Б., Крналик С. и др. Экспрессия сплайс-вариантов рецептора андрогена в костных метастазах рака предстательной железы связана с резистентностью к кастрации и короткой выживаемостью. PLoS One 2011; 6: e19059. [ПубМед]
- Yang X, Guo Z, Sun F и др. Новый сплайс-вариант ассоциированного с мембраной рецептора андрогенов потенцирует пролиферативный ответ и реакцию выживания в клетках рака предстательной железы. J Biol Chem 2011; 286:36152-60. [ПубМед]
- Tannock IF, de Wit R, Berry WR, et al. Доцетаксел плюс преднизолон или митоксантрон плюс преднизолон при распространенном раке предстательной железы. N Engl J Med 2004;351:1502-12. [ПубМед]
- Серпа Нето А., Тобиас-Мачадо М., Каликс Р. и др. Десять лет терапии аденокарциномы предстательной железы на основе доцетаксела: систематический обзор и метаанализ 2244 пациентов в 12 рандомизированных клинических исследованиях. Clin Genitourin Рак 2011; 9: 115-23. [ПубМед]
- Марех И., Вакка А., Раньери Г. и др. Новые стратегии лечения резистентного к кастрации рака предстательной железы Int J Oncol 2012;40:1313-20. (Обзор). [ПубМед]
- Sweeney C, Chen YH, Carducci MA, et al. Влияние химиогормональной терапии на общую выживаемость (ОВ) по сравнению с гормональной терапией при гормоночувствительном новом метастатическом раке предстательной железы (mPrCa): рандомизированное исследование фазы III под руководством ECOG. J Clin Oncol 2014;32:абстр LBA2.
- Шеланский М.Л., Гаскин Ф., Кантор Ч.Р. Сборка микротрубочек без добавления нуклеотидов. Proc Natl Acad Sci U S A 1973;70:765-8. [ПубМед]
- McGrogan BT, Gilmartin B, Carney DN, et al. Таксаны, микротрубочки и химиорезистентный рак молочной железы. Биохим Биофиз Акта 2008;1785:96-132.
- Штернберг К.Н., Петрилак Д.П., Мадан Р.А. и др. Прогресс в лечении распространенного рака предстательной железы. Am Soc Clin Oncol Educ Book 2014.117-31. [ПубМед]
- Фаббри Ф., Амадори Д., Карлони С. и др. Митотическая катастрофа и апоптоз, индуцированные доцетакселом в гормонорезистентных клетках рака предстательной железы. J Cell Physiol 2008; 217:494-501. [ПубМед]
- Крамер Г., Шварц С., Хагг М. и др. Доцетаксел индуцирует апоптоз в гормонорезистентных карциномах предстательной железы во время нескольких циклов лечения. Бр Дж Рак 2006;94:1592-8. [ПубМед]
- Курода К., Лю Х., Ким С. и др. Доцетаксел подавляет экспрессию рецептора андрогена и простат-специфического антигена, но не простат-специфического мембранного антигена в клеточных линиях рака предстательной железы: последствия для суррогатного материнства ПСА. Простата 2009;69:1579-85. [ПубМед]
- O’Neill AJ, Prencipe M, Dowling C, et al. Характеристика и манипулирование клеточными линиями рака предстательной железы, устойчивыми к доцетакселу. Мол Рак 2011; 10:126. [ПубМед]
- Sissung TM, Baum CE, Deeken J, et al. Генетическая изменчивость ABCB1 влияет на токсичность и клинический исход у пациентов с андроген-независимым раком предстательной железы, получающих доцетаксел. Clin Cancer Res 2008; 14:4543-9. [ПубМед]
- Терри С., Плоссар Г., Аллори Ю. и др. Повышенная экспрессия бета-тубулина класса III при кастрационно-резистентном раке предстательной железы человека. Бр Дж Рак 2009;101:951-6. [ПубМед]
- Ploussard G, Terry S, Maille P, et al. Экспрессия бета-тубулина класса III предсказывает агрессивность опухоли предстательной железы и ответ пациента на химиотерапию на основе доцетаксела. Рак Res 2010;70:9253-64. [ПубМед]
- Хара Т., Ушио К., Нишиваки М. и др. Мутация в бета-тубулине и устойчивая зависимость от передачи сигналов рецептора андрогена в недавно созданной устойчивой к доцетакселу клеточной линии рака предстательной железы. Cell Biol Int 2010;34:177-84. [ПубМед]
- Ган Л., Ван Дж., Сюй Х. и др. Устойчивость к доцетаксел-индуцированному апоптозу в клетках рака предстательной железы посредством передачи сигналов p38/p53/p21. Простата 2011;71:1158-66. [ПубМед]
- Zhu Y, Liu C, Nadiminty N, et al. Ингибирование экспрессии ABCB1 преодолевает приобретенную резистентность к доцетакселу при раке предстательной железы. Мол Рак Тер 2013; 12:1829-36. [ПубМед]
- Тантивейкул К., Лоберг Р.Д., Мавоча С.К. и др. Опосредованная PAR1 активация NFkappaB способствует выживанию клеток рака предстательной железы посредством Bcl-xL-зависимого механизма. J Cell Biochem 2005; 96:641-52. [ПубМед]
- Доминго-Доменек Дж., Олива С., Ровира А. и др. Интерлейкин 6, мишень ядерного фактора каппаВ, предсказывает резистентность к доцетакселу при гормононезависимом раке предстательной железы, а ингибирование ядерного фактора каппаВ PS-1145 усиливает противоопухолевую активность доцетаксела. Clin Cancer Res 2006; 12:5578-86. [ПубМед]
- Чжун Б., Салман Д.А., Гилвари Д.Л. и др. Индукция кластерина с помощью AKT — роль в цитопротекции против доцетаксела в клетках опухоли предстательной железы. Мол Рак Тер 2010;9: 1831-41. [ПубМед]
- Zhang H, Kim JK, Edwards CA, et al. Кластерин ингибирует апоптоз, взаимодействуя с активированным Bax. Nat Cell Biol 2005;7:909-15. [ПубМед]
- Совери Р.Д., Хадащик Б.А., Со А.И. и др. Нокдаун кластерина с использованием антисмыслового олигонуклеотида OGX-011 повторно сенсибилизирует клетки рака предстательной железы PC-3, устойчивые к доцетакселу, к химиотерапии. BJU Int 2008; 102:389-97. [ПубМед]
- Magadoux L, Isambert N, Plenchette S, et al. Новые цели для мониторинга и преодоления резистентности к доцетакселу при кастрационно-резистентном раке предстательной железы Int J Oncol 2014;45:919-28. (обзор). [ПубМед]
- Шукла С., Гупта С. Подавление конститутивной и альфа-индуцированной фактора некроза опухоли активации ядерного фактора (NF)-kappaB и индукции апоптоза апигенином в клетках карциномы предстательной железы человека PC-3: корреляция с подавлением NF-kappaB- отзывчивые гены. Clin Cancer Res 2004; 10:3169-78. [ПубМед]
- де Боно Дж. С., Логотетис С. Дж., Молина А. и др. Абиратерон и увеличение выживаемости при метастатическом раке предстательной железы. N Engl J Med 2011;364:1995-2005. [ПубМед]
- Ryan CJ, Smith MR, de Bono JS, et al. Абиратерон при метастатическом раке предстательной железы без предшествующей химиотерапии. N Engl J Med 2013; 368: 138-48. [ПубМед]
- Аттард Г., Рейд А.Х., А’Херн Р. и др. Селективное ингибирование CYP17 с помощью абиратерона ацетата высокоэффективно при лечении резистентного к кастрации рака предстательной железы. Дж. Клин Онкол 2009; 27:3742-8. [ПубМед]
- Attard G, Reid AH, Auchus RJ, et al. Клинические и биохимические последствия ингибирования CYP17A1 абиратероном, вводимым с экзогенными глюкокортикоидами и без них, у кастрированных мужчин с распространенным раком предстательной железы. Дж. Клин Эндокринол Метаб 2012; 97:507-16. [ПубМед]
- Cai C, Chen S, Ng P, et al. Внутриопухолевый синтез стероидов de novo активирует рецепторы андрогенов при кастрационно-резистентном раке предстательной железы и усиливается при лечении ингибиторами CYP17A1. Рак Res 2011; 71: 6503-13. [ПубМед]
- Мостагель Э.А., Марк Б.Т., Плимат С.Р. и др. Резистентность к ингибированию CYP17A1 абиратероном при кастрационно-резистентном раке предстательной железы: индукция стероидогенеза и вариантов сплайсинга рецепторов андрогенов. Clin Cancer Res 2011;17:5913-25. [ПубМед]
- Агарвал Н., Ди Лоренцо Г., Сонпавде Г. и др. Новые агенты для лечения рака предстательной железы. Энн Онкол 2014; 25:1700-9. [ПубМед]
- Sternberg CN, de Bono JS, Chi KN, et al. Улучшение исходов у пожилых пациентов с метастатическим резистентным к кастрации раком предстательной железы, получавших ингибитор рецепторов андрогенов энзалутамид: результаты исследования III фазы AFFIRM. Энн Онкол 2014; 25:429-34. [ПубМед]
- Beer TM, Armstrong AJ, Rathkopf DE, et al. Энзалутамид при метастатическом раке предстательной железы до химиотерапии. N Engl J Med 2014; 371: 424-33. [ПубМед]
- Mizushima N, Levine B, Cuervo AM, et al. Аутофагия борется с болезнями посредством клеточного самопереваривания. Природа 2008;451:1069-75. [ПубМед]
- Се Z, Клионский DJ. Формирование аутофагосом: основной механизм и приспособления. Nat Cell Biol 2007;9:1102-9. [ПубМед]
- Нгуен Х.Г., Ян Дж.К., Кунг Х.Дж. и др. Воздействие на аутофагию преодолевает резистентность к энзалутамиду в устойчивых к кастрации клетках рака предстательной железы и улучшает терапевтический ответ в модели ксенотрансплантата. Онкоген 2014;33:4521-30. [ПубМед]
- Леоне Р.Д., Амаравади Р.К. Аутофагия: целевой стержень метаболизма раковых клеток. Тенденции Endocrinol Metab 2013;24:209-17. [ПубМед]
- Чен Н., Каранца В. Аутофагия как терапевтическая мишень при раке. Cancer Biol Ther 2011;11:157-68. [ПубМед]
- Bennett HL, Stockley J, Fleming JT, et al. Оказывает ли аутофагия, связанная с терапией абляции андрогенов (ААТ), эффект выживания в клетках рака предстательной железы LNCaP человека? BJU Int 2013;111:672-82. [ПубМед]
- Фэрроу Дж.М., Ян Дж.К., Эванс С.П. Аутофагия как модулятор и мишень при раке предстательной железы. Нат Рев Урол 2014;11:508-16. [ПубМед]
- Eisermann K, Wang D, Jing Y, et al. Мутация гена рецептора андрогенов, реаранжировка, полиморфизм. Перевод Андрол Урол 2013;2:137-47. [ПубМед]
- Корпал М., Корн Дж. М., Гао X и др. Мутация F876L в рецепторе андрогена придает генетическую и фенотипическую устойчивость к MDV3100 (энзалутамиду). Рак Дисков 2013;3:1030-43. [ПубМед]
- Шарифи Н. Стероидные рецепторы в большом количестве при раке предстательной железы. N Engl J Med 2014;370:970-1. [ПубМед]
- Denayer S, Helsen C, Thorrez L, et al. Правила узнавания ДНК рецептором андрогенов. Мол Эндокринол 2010;24:898-913. [ПубМед]
- Claessens F, Helsen C, Prekovic S, et al. Новые механизмы устойчивости к энзалутамиду при раке предстательной железы. Нат Рев Урол 2014;11:712-6. [ПубМед]
- Джозеф Дж.Д., Лу Н., Цянь Дж. и др. Клинически значимая мутация андрогенового рецептора придает устойчивость к антиандрогенам второго поколения энзалутамиду и ARN-509. . Рак Дисков 2013;3:1020-9. [ПубМед]
- Антонаракис Э.С., Лу С., Ван Х. и др. AR-V7 и устойчивость к энзалутамиду и абиратерону при раке предстательной железы. N Engl J Med 2014;371:1028-38. [ПубМед]
- Li Y, Chan SC, Brand LJ и др. Варианты сплайсинга рецептора андрогена опосредуют устойчивость к энзалутамиду в устойчивых к кастрации клеточных линиях рака предстательной железы. Рак Res 2013; 73: 483-9. [ПубМед]
- Лю С., Лу В., Чжу Ю. и др. Никлозамид ингибирует экспрессию вариантов рецепторов андрогенов и преодолевает резистентность к энзалутамиду при кастрационно-резистентном раке предстательной железы. Clin Cancer Res 2014;20:3198-210. [ПубМед]
- Надиминти Н., Туммала Р., Лу В. и др. МикроРНК let-7c подавляет экспрессию и активность рецепторов андрогенов посредством регуляции экспрессии Myc в клетках рака предстательной железы. J Biol Chem 2012; 287:1527-37. [ПубМед]
- Лай К.П., Хуан К.К., Чанг Ю.Дж. и др. Новый терапевтический подход к подавлению резистентного к кастрации рака предстательной железы с использованием ASC-J9 посредством нацеливания на рецептор андрогена в селективных клетках предстательной железы. Ам Дж. Патол 2013; 182:460-73. [ПубМед]
- Садар MD. Низкомолекулярные ингибиторы, нацеленные на «ахиллесову пяту» активности рецепторов андрогенов. Рак Res 2011; 71: 1208-13. [PubMed]
Цитируйте эту статью как: Чандрасекар Т., Ян Дж.С., Гао А.С., Эванс С.П. Механизмы резистентности при кастрационно-резистентном раке предстательной железы (КРРПЖ). Перевод Андрол Урол 2015;4(3):365-380. doi: 10.3978/j.issn.2223-4683.2015.05.02
Механизмы и манипулирование устойчивостью корней картофеля к мучнистой парше — НИЦ
Заражение корней мучнистой паршой нарушает функцию корней – это означает, что для компенсации плохого развития корней требуется больше орошения, удобрений и фунгицидов – и приводит к снижению урожайности. В рамках этого проекта исследователи изучают устойчивость корней к заражению мучнистой паршой. Это включает в себя изучение того, как патоген заражает корни и вызывает заболевание, что способствует устойчивости у некоторых сортов картофеля и могут ли механизмы устойчивости быть усилены или переданы.
Дата проекта
20 декабря 2018 г. – 1 июля 2022 г.
Посетить сайт
Запрос об этом проекте
Заполните информацию, чтобы отправить запрос об этом исследовательском проекте.
Имя
Фамилия
Адрес электронной почты
Контактный телефон
Организация Организация
Тип организации
Пожалуйста, выберите Акселератор или ИнкубаторКорпорацияГосударственное агентствоОтраслевая организацияОтраслевой консультантИнвесторЧастная исследовательская организацияГосударственная исследовательская организацияФинансирующая организацияИсследовательская организацияПоставщики исследовательских услугСтартап или масштабирование или малый и средний бизнесДругое
Промышленность
Пожалуйста, выберитеАльтернативный белокАквакультураМясной скотНапиткиЗлаковые культурыХлопокКросс-промышленностьМолочные продуктыЯйцаЭфирные маслаЛесное хозяйствоФруктыДичьМедоносные пчелыПриродные активыОрехиМасличныесеменаКомплексные продуктыПастбища, корма и кормаСвининаПтицаБобовые зернаОвца и баранинаСахарОвощиДикий промыселВиноШерсть3
9000 Страна
Please selectAfghanistanAland IslandsAlbaniaAlgeriaAmerican SamoaAndorraAngolaAnguillaAntarcticaAntigua And BarbudaArgentinaArmeniaArubaAustraliaAustriaAzerbaijanBahamasBahrainBangladeshBarbadosBelarusBelgiumBelizeBeninBermudaBhutanBoliviaBosnia And HerzegovinaBotswanaBouvet IslandBrazilBritish Indian Ocean TerritoryBrunei DarussalamBulgariaBurkina FasoBurundiCambodiaCameroonCanadaCape VerdeCayman IslandsCentral African RepublicChadChileChinaChristmas IslandCocos (Keeling) IslandsColombiaComorosCongoCongo, Democratic RepublicCook IslandsCosta RicaCote D’IvoireCroatiaCubaCyprusCzech RepublicDenmarkDjiboutiDominicaDominican RepublicEcuadorEgyptEl SalvadorEquatorial GuineaEritreaEstoniaEthiopiaFalkland Islands (Malvinas)Faroe IslandsFijiFinlandFranceFrench GuianaFrench PolynesiaFrench Southern TerritoriesGabonGambiaGeorgiaGermanyGhanaGibraltarGreeceGreenlandGrenadaGuadeloupeGuamGuatemalaGuernseyGuineaGuinea-BissauGuyanaHaitiHeard Island и острова МакдональдсСвятой Престол (Ватикан y State)HondurasHong KongHungaryIcelandIndiaIndonesiaIran, Islamic Republic OfIraqIrelandIsle Of ManIsraelItalyJamaicaJapanJerseyJordanKazakhstanKenyaKiribatiKoreaKuwaitKyrgyzstanLao People’s Democratic RepublicLatviaLebanonLesothoLiberiaLibyan Arab JamahiriyaLiechtensteinLithuaniaLuxembourgMacaoMacedoniaMadagascarMalawiMalaysiaMaldivesMaliMaltaMarshall IslandsMartiniqueMauritaniaMauritiusMayotteMexicoMicronesia, Federated States OfMoldovaMonacoMongoliaMontenegroMontserratMoroccoMozambiqueMyanmarNamibiaNauruNepalNetherlandsNetherlands AntillesNew CaledoniaNew ZealandNicaraguaNigerNigeriaNiueNorfolk IslandNorthern Mariana IslandsNorwayOmanPakistanPalauPalestinian Territory, OccupiedPanamaPapua New GuineaParaguayPeruPhilippinesPitcairnPolandPortugalPuerto RicoQatarReunionRomaniaRussian FederationRwandaSaint BarthelemySaint HelenaSaint Kitts And NevisSaint LuciaSaint MartinSaint Pierre And MiquelonSaint Vincent And GrenadinesSamoaSan MarinoSao Tome And ПринципыСаудовская АравияСенегалСербияСей chellesSierra LeoneSingaporeSlovakiaSloveniaSolomon IslandsSomaliaSouth AfricaSouth Georgia And Sandwich Isl. SpainSri LankaSudanSurinameSvalbard And Jan MayenSwazilandSwedenSwitzerlandSyrian Arab RepublicTaiwanTajikistanTanzaniaThailandTimor-LesteTogoTokelauTongaTrinidad And TobagoTunisiaTurkeyTurkmenistanTurks And Caicos IslandsTuvaluUgandaUkraineUnited Arab EmiratesUnited KingdomUnited StatesUnited States Outlying IslandsUruguayUzbekistanVanuatuVenezuelaViet NamVirgin Islands, BritishVirgin Islands, U.S.Wallis And FutunaWestern SaharaYemenZambiaZimbabwe
Пожалуйста, скажите нам, кто из следующих вы
Пожалуйста, выберите Предприниматель или поставщик решенийКорпоративный инвесторИсследователь или исследовательская организацияПроизводитель или агроном
Пожалуйста, расскажите нам немного больше о вашем запросе
Главный следователь
Вино Раджандран
Исследовательские организации
Университет Тасмании (UTAS)
Проект финансируется
Несколько отраслей
Альтернативный белок Кросс-индустрия Фрукты Орехи Другие сельские отрасли Пастбище, фураж и корма Овощи
Несколько отраслей
Альтернативный белок Кросс-индустрия Фрукты Орехи Другие сельские отрасли Пастбище, фураж и корма Овощи
Связанные исследовательские проекты
Искать во всех исследовательских проектахФрукты
CT20009 — Комплексная борьба с черной пятнистостью цитрусовых и коричневой пятнистостью «Император»
Австралийская цитрусовая промышленность сильно ориентирована на экспорт и должна тщательно регулировать использование пестицидов, чтобы обеспечить соответствие максимальным остаточным уровням (MRL), которые различаются на каждом зарубежном рынке. Этот проект направлен на определение дополнительных инструментов комплексного управления болезнями (IDM), чтобы помочь производителям бороться с черной пятнистостью цитрусовых, «императорской» коричневой пятнистостью и черной сердцевинной гнилью цитрусовых таким образом, чтобы обеспечить соответствие MRL.
Прочие сельские отрасли
Разработка региональных моделей риска грибковых заболеваний пиретрума
Пиретрум – это разновидность хризантемы, которая производит один из наиболее часто используемых несинтетических инсектицидов. Болезни, такие как лучевая гниль и желтовато-коричневая пятнистость, представляют большой риск для урожайности пиретрума в Австралии, и в рамках этого проекта предполагается разработать основу для лечения этих болезней в конкретных регионах.
Несколько отраслей
Инновации в действии
В рамках этого проекта изучаются инновационные способы проведения НИОКР в садоводческой отрасли Австралии посредством сотрудничества с консультантами по агробизнесу из частного сектора, которые часто являются надежными союзниками своих клиентов-производителей. Был разработан ряд мероприятий, направленных на повышение осведомленности консультантов и производителей о соответствующих исследованиях и разработках и передовых методах управления с целью достижения изменений в практике на фермах.
Есть вопросы?
Узнайте, как мы можем вам помочь.
Найдите ответы на наиболее часто задаваемые вопросы об исследовательских проектах, коммерческих возможностях, организациях и многом другом.
Остались вопросы или есть отзывы о сайте? Пожалуйста, свяжитесь с нами, заполнив форму запроса.
Часто задаваемые вопросы Узнать сейчас
Сравнение механизмов манипулирования хозяином в таксонах хозяина и паразита | Журнал экспериментальной биологии
Skip Nav Destination
ИЗМЕНЕНИЕ ПОВЕДЕНИЯ ХОЗЯИНА| 01 января 2013 г.
Кевин Д. Лафферти,
Дженни С. Шоу
Информация об авторе и статье
* Автор для переписки ([email protected])
Полученный: 12 апреля 2012 г.
Принято: 05 июля 2012 г.
Online Issn: 1477-9145
Print Issn: 0022-0949
© 2013. Опубликовано The Company of Biologist Ltd.
https://doi.org/10.1242/jeb.073668
История статьи
Получено:
12 апреля 2012 г.
Принято:
05 июля 2012 г.
Похожие материалы
Соответствующая статья была опубликована: КАК ВЕДОМЫЕ ПАРАЗИТЫ ПРЕВРАЩАЮТ ЖЕРТВ В ЗОМБИ
- Разделенный экран
-
Просмотры
- Содержание артикула
- Рисунки и таблицы
- Видео
- Аудио
- Дополнительные данные
- Экспертная оценка
-
Делиться
- MailTo
- Твиттер
-
Инструменты
-
Получить разрешения
-
Иконка Цитировать Цитировать
-
- Поиск по сайту
Citation
Кевин Д. Лафферти, Дженни С. Шоу; Сравнение механизмов манипулирования хозяином в таксонах хозяина и паразита. J Exp Biol 1 января 2013 г.; 216 (1): 56–66. doi: https://doi.org/10.1242/jeb.073668
Скачать файл цитаты:
- Ris (Zotero)
- Менеджер ссылок
- EasyBib
- Подставки для книг
- Менделей
- Бумаги
- КонецПримечание
- РефВоркс
- Бибтекс
Расширенный поиск
Паразиты влияют на поведение хозяина несколькими способами. Они могут изменять активность, микросреду обитания или и то, и другое. Для паразитов, передающихся трофически (в центре нашего исследования), снижение активности может ухудшить способность хозяев реагировать на хищников-окончательных хозяев, а повышенная активность и изменение выбора микросреды обитания могут увеличить частоту контактов между хозяевами и хищниками-окончательными хозяевами. При анализе паразитов, передающихся трофическим путем, больше групп паразитов изменяют активность, чем изменяют выбор микросреды обитания. Паразиты, заражающие позвоночных, с большей вероятностью ухудшали реакцию хозяина на хищников, тогда как паразиты, заражавшие беспозвоночных, с большей вероятностью усиливали контакт хозяина с хищниками. Место заражения может повлиять на то, как паразиты манипулируют своими хозяевами. Например, паразиты в центральной нервной системе кажутся особенно подходящими для манипулирования поведением хозяина. Манипулятивные паразиты обычно заселяют полость тела, мышцы и центральную нервную систему своих хозяев. Скребни в наборе данных отличались от других таксонов тем, что встречались исключительно в полости тела беспозвоночных. Кроме того, они с большей вероятностью изменяли выбор микросреды обитания, чем активность. Паразиты в полости тела (по разным типам паразитов) с большей вероятностью были связаны с повышенным контактом хозяина с хищниками. Паразиты могут манипулировать хозяином посредством выкачивания энергии, но большинство паразитов используют более изощренные средства. Например, паразиты нацелены на четыре физиологические системы, формирующие поведение как беспозвоночных, так и позвоночных: нервную, эндокринную, нейромодулирующую и иммуномодулирующую. Взаимосвязи между этими системами затрудняют выделение конкретных механизмов манипулирования поведением хозяина.
Ключевые слова:
модификация поведения, трофическая передача, микросреда обитания, нейромодуляция, иммуномодуляция, место заражения, Activity
Являются ли паразиты изощренными кукловодами поведения своего хозяина? Многие исследования показали, что различия в поведении инфицированных и неинфицированных хозяев полезны для паразитов. В других случаях логично ожидать адаптации паразита. В любом случае доказательства неуловимы, а различия в поведении инфицированных и неинфицированных носителей сами по себе не подразумевают манипуляций со стороны паразита. Например, патогенные паразиты могут влиять на поведение хозяина таким образом, что это не приносит пользы паразиту. В нашем обзоре мы предполагаем, что примеры, которые мы приводим, являются адаптациями паразитов, но читатели должны понимать, что поддержка этого предположения варьируется.
Паразиты атакуют четыре физиологические системы, формирующие поведение как беспозвоночных, так и позвоночных: нервную, эндокринную, нейромодулирующую и иммуномодулирующую (Adamo, 2002; Adamo, 2013; Beckage, 1993; Escobedo et al., 2009; Helluy, 2013; Moore, 2002). ; Thomas et al., 2005; Thompson and Kavaliers, 1994). Эти системы связаны и взаимодействуют через нейротрансмиттеров, гормонов, а также нейромодулирующих и иммуномодулирующих химических веществ. В результате определение точных механизмов, лежащих в основе модификации поведения хозяина, оказалось сложной задачей. Более того, химически опосредованные модификации могут быть результатом действия секретируемых паразитами и хозяином веществ в ответ друг на друга, что еще больше скрывает механистическую основу изменений (Adamo, 2013; Thomas et al. , 2005). Менее сложные паразиты могут манипулировать поведением хозяина, истощая энергию или повреждая ключевые системы, такие как центральная нервная система (ЦНС) (хотя воздействие на ЦНС, возможно, является сложным достижением).
В этом обзоре манипулятивных паразитов мы обсуждаем, как различные типы поведенческих манипуляций распределяются между таксонами хозяев и паразитов. Основная часть нашего обзора касается способов, которыми паразиты манипулируют своими хозяевами, и того, как такие факторы, как место заражения, различаются в зависимости от таксонов хозяина и паразита.
Влияет ли тип хозяина или паразита на эволюцию поведенческих манипуляций? Пулен (Poulin, 1994) провел метаанализ, чтобы выяснить, как таксоны хозяев и паразитов влияют на величину изменений в активности хозяина или выборе микросреды обитания. Он обнаружил, что нематоды, передающиеся нетрофическим путем, оказывают наибольшее влияние на поведение хозяина. Скребни влияли только на выбор микросреды обитания хозяина, а цестоды оказывали большое влияние на активность хозяина. Кроме того, манипулирование выбором микросреды обитания было сильнее у позвоночных хозяев, чем у беспозвоночных хозяев. Поскольку за последние два десятилетия было проведено значительное исследование манипулятивных паразитов, мы хотели изучить влияние таксонов хозяев и паразитов с помощью новых данных и другого подхода.
Чтобы понять, как манипуляции распределяются между хозяевами и паразитами, мы обобщили доступную литературу по модификации поведения паразитов, передающихся трофически, начиная с всестороннего обзора Мура (Moore, 2002), который мы обновили с использованием последних публикаций (таблица 1). Несколько видов были изучены на наличие некоторых родов паразитов (например, Polymorphus ), часто с одинаковыми результатами, полученными для разных видов. Это предполагало, что виды внутри рода не являются независимыми, и поэтому мы решили не использовать виды в качестве нашей единицы репликации. Нас интересовала группа паразитов (например, цестоды против скребней) как фактор, поэтому мы также не учитывали филогению. Наше решение состояло в том, чтобы рассматривать в качестве единицы репликации роды паразитов, а не виды. Чтобы сфокусировать наш вопрос, мы рассмотрели только примеры, для которых авторы постулировали изменение поведения, которое может сделать промежуточных хозяев более восприимчивыми к хищничеству со стороны конечных хозяев. У нас была информация о манипулировании хозяином из 55 родов паразитов, распределенных среди следующих таксонов паразитов: три простейших, 10 скребней, 17 дигенеев, 15 цестод и 10 нематод. Для двух родов мы не смогли установить детали поведенческих манипуляций. Мы разделили зарегистрированное поведение инфицированных хозяев на три типа: изменение выбора микросреды обитания и изменение активности (увеличение или уменьшение). Поскольку паразиты могут манипулировать более чем одним поведением одновременно (Poulin, 2013), мы отслеживали роды паразитов, для которых имелись данные о более чем одном типе манипуляции [хотя мы отмечаем, что расширение исследований, вероятно, обнаружит больше типов измененных поведения (Perrot-Minnot and Cézilly, 2013)]. Тридцать родов имели пониженную активность, а 12 родов — повышенную активность. Относительная частота повышенной или пониженной активности не зависела от того, наблюдались ли в роде эффекты микросреды обитания.
Таблица 1.
Краткая информация об манипуляциях с паразитами по родам
Вид Большой
Мы провели два типа анализов. Для первого анализа мы последовали за Пулином (Poulin, 1994) в противопоставлении измененной паразитом активности хозяина с выбором хозяином микросреды обитания, измененной паразитом. Типы поведения различались между измененной активностью (26), измененной микросредой (11) и измененной активностью и микросредой (16). Другими словами, манипулирование с большей вероятностью изменит активность, чем выбор микросреды обитания. Чтобы определить, была ли эта тенденция одинаковой для таксонов хозяев и паразитов, мы записали таксон паразита и сгруппировали хозяев по позвоночным и беспозвоночным (более точная группировка хозяев по типам не повлияла на результаты). Для статистического сравнения таксонов паразитов мы исключили простейших из-за небольшого размера выборки для этой категории. Мы искали модели логистической регрессии, которые позволяли бы конкурировать независимым переменным, выбирая окончательные модели на основе информационного критерия Акаике (Burnham and Anderson, 2004). В тех случаях, когда при сравнении были очевидны особенности, мы ограничивали анализ таксонами паразитов с представителями в нескольких категориях.
Поведение хозяина было в значительной степени связано с одномерным анализом таксона паразита (χ 2 = 15,2, d.f. = 6, P = 0,012; рис. 1), потому что больше скребней (все из которых были зарегистрированы от беспозвоночных хозяев) были чаще связаны с изменением выбора микросреды обитания, чем с изменением активности хозяев (согласно Poulin, 1994), тогда как другие группы больше связаны с активностью хозяев. Эффект скребней указал на проблему сингулярности при анализе таксона хозяина, поскольку все зарегистрированные скребни паразитировали в полости тела беспозвоночных хозяев, что делало невозможным определение того, была ли разница между хозяевами беспозвоночных и позвоночных обусловлена таксоном хозяина, таксоном паразита или местонахождением. инфекции. Многофакторный анализ, исключающий скребней и простейших, обнаружил значительное влияние таксона хозяина (χ 2 =7,6, d.f.=2, P =0,02), но взаимодействие между таксоном-хозяином и местом заражения было значительным (χ 2 =14,6, d.f.=4, P =0,006). На рис. 2 показано, что активность была более распространенным эффектом манипуляций у позвоночных хозяев, чем выбор микросреды обитания, но тип поведения у беспозвоночных хозяев не отличался.
Пулен (Poulin, 1994) пришел к несколько иным выводам: измененная активность и выбор микросреды обитания одинаково часто встречались в его анализе акатоцефалов, нематод, дигеней и цестод. Кроме того, инфицированные позвоночные хозяева имели более сильно измененный выбор микросреды обитания, чем беспозвоночные хозяева. Однако подход, переменные ответа и данные были разными для двух исследований. В частности, результаты Пулена были обусловлены нематодами, передающимися нетрофически, которые мы исключили из нашего анализа. После исключения нематод из таблицы Пулена 1 (Poulin, 1994), мы подсчитали, что три из его видов скребней изменили активность, тогда как шесть изменили выбор микросреды обитания. Что касается других его таксонов паразитов, два изменили активность и три изменили выбор микросреды обитания, результат согласуется с нашими выводами о том, что скребни отличаются от других таксонов паразитов своей склонностью манипулировать выбором хозяина.
Рис. 1.
УвеличитьСкачать слайд
Тип измененного поведения хозяев (активность, выбор микросреды обитания) по таксонам паразитов. Данные представляют 55 родов трофически передающихся паразитов. Для каждого рода может быть несколько видов и несколько исследований для каждого вида. Предполагалось, что измененное поведение является адаптивным и классифицируется в зависимости от того, нарушают ли они способность хозяина реагировать на хищника или заставляют хозяина искать среду обитания или проявлять поведение, которое подвергает его повышенному риску нападения хищников. Во многих случаях у представителей рода паразитов наблюдались оба типа поведенческих манипуляций.
Рис. 1.
УвеличитьСкачать слайд
Тип измененного поведения хозяина (активность, выбор микросреды обитания) по таксонам паразитов. Данные представляют 55 родов трофически передающихся паразитов. Для каждого рода может быть несколько видов и несколько исследований для каждого вида. Предполагалось, что измененное поведение является адаптивным и классифицируется в зависимости от того, нарушают ли они способность хозяина реагировать на хищника или заставляют хозяина искать среду обитания или вести себя так, что подвергают его повышенному риску нападения хищников. Во многих случаях у представителей рода паразитов наблюдались оба типа поведенческих манипуляций.
Мы также проанализировали альтернативную группу поведения. Основное отличие от нашего предыдущего анализа заключалось в том, что мы не объединяли повышенную и пониженную активность в одну категорию. Если паразитированный хозяин описывался как вялый, мы предполагали, что у него будет пониженная реакция на хищников-окончательных хозяев, тогда как, если он описывался как более активный, мы предполагали, что у него будет повышенный контакт с хищниками. Типы поведения распределялись поровну между усилением контакта с хищниками (19), снижение реакции на хищников (20), а также усиление контакта и снижение реакции (15). Исключая простейших и скребней, чтобы сосредоточиться на влиянии таксона хозяина (как указано выше), мы обнаружили, что хозяева позвоночных в большей степени ассоциировались со сниженной реакцией, тогда как хозяева беспозвоночных в большей степени ассоциировались с повышенным контактом (χ 2 =16,2, d.f.= 2, P =0,0003 (рис. 3). Однако остаточное влияние таксона паразита или места заражения на эту классификацию поведения хозяина не было очевидным.
Таким образом, два наших анализа расширяют предыдущий анализ Пулена (Poulin, 1994) дополнительными данными и другим акцентом. Скребни с большей вероятностью изменяли выбор микросреды обитания хозяина, в то время как другие паразиты с большей вероятностью изменяли активность хозяина. Паразиты позвоночных с большей вероятностью снижали реакцию хозяина на хищников, в то время как паразиты беспозвоночных с большей вероятностью усиливали контакт хозяина с хищниками.
Механизмы, лежащие в основе модификации поведения паразитами, загадочны, и хотя некоторые компоненты были идентифицированы для частей изменения поведения, описание всех потенциальных механизмов остается в центре внимания совместных исследований паразитологов, физиологов и нейробиологов. Здесь мы обсуждаем утечку энергии, место заражения, нейровоспаление и моноаминовые нейротрансмиттеры как возможные общие механизмы модификации поведения паразитами нескольких таксонов. Однако оказывается, что механизмы были изучены только в 19родов паразитов, поэтому по-прежнему трудно делать обобщения из современной литературы.
Рис. 2.
Посмотреть в большом размереСкачать слайд
Тип измененного поведения хозяина (активность, выбор микросреды обитания) по таксону хозяина после исключения скребней и простейших. Другие детали как на рис. 1.
Рис. 2.
Посмотреть в большом размереСкачать слайд
Тип измененного поведения хозяина (активность, выбор микросреды обитания) по таксону хозяина после исключения скребней и простейших. Остальные детали как на рис. 1.
Истощение энергии
Паразиты извлекают энергию в виде пищи из своих хозяев. Если хозяевам не хватает питательных веществ, их поведение может измениться. Если утечка энергии ухудшает их физиологию, они могут стать более вялыми или демонстрировать более низкую физическую работоспособность. В качестве альтернативы, если у хозяина истощается энергия, он может стать более активным и увеличить скорость поиска пищи. Любое изменение в поведении может принести пользу паразиту, ответственному за утечку энергии.
Истощение энергии может привести к недомоганию, которое мешает защите. Например, самая опасная часть жизни комара — это пить кровь хозяина, который не хочет быть укушенным. Люди приспособлены слышать и чувствовать комаров и способны прихлопывать и убивать их. Эта реакция помогает хозяевам избежать кровопотери и уменьшить воздействие болезней, переносимых комарами, таких как малярия. Малярия — тяжелое заболевание, вызывающее периодическую лихорадку и требующее от хозяина пополнения клеток крови, потребляемых паразитом. Люди в разгар малярийной лихорадки часто становятся недееспособными, и это подвергает их воздействию комаров, которые приземляются, чтобы непрерывно питаться кровью. Например, эксперименты показали, что мыши, зараженные малярией, больше не пытаются избегать комаров в лаборатории (Day and Edman, 19).83). Точно так же у инфицированных хозяев может быть меньше запасов энергии или способности напрягать мышцы, необходимые для спасения от хищников. Личинка ленточного червя Echinococcus granulosus , по-видимому, выводит из строя лося-промежуточного хозяина, оседая в легких, гипотетически уменьшая емкость легких и облегчая волкам нападение на лося (Joly and Messier, 2004).
Утечка энергии также может повлиять на выбор среды обитания. Риск, связанный с кормлением, может поставить хозяина в ситуации, когда он с большей вероятностью будет съеден хищником, который действует как окончательный хозяин для паразита. У рыб, зараженных личинками ленточных червей, потребность в кислороде выше, поэтому они проводят больше времени на поверхности (Джайлс, 19 лет).87). Энергия, потребляемая паразитами, может повлиять на решения о поиске пищи. Голодный хозяин может с большей вероятностью избегать убежища, чтобы увеличить скорость добычи пищи (Milinski, 1984).
Рис. 3.
Посмотреть в большом размереСкачать слайд
Тип измененного поведения хозяина (реакция хозяина, контакт с хозяином) по таксону хозяина после исключения скребней и простейших. Другие детали, как на рис. 1.
Рис. 3.
Просмотреть в большом размереСкачать слайд
Тип измененного поведения хозяина (реакция хозяина, контакт с хозяином) по таксону хозяина после исключения скребней и простейших. Остальные детали как на рис. 1.
Преимущество оттока энергии как поведенческой манипуляции заключается в том, что отбирать энергию у хозяина — это то, для чего паразиты уже выбраны. Это отличный пример того, как поведенческие манипуляции не должны быть изощренными. Однако утечка энергии приводит к общим изменениям в поведении, которые могут не быть нацелены на конкретного хищника или переносчика хозяина, что приводит к неэффективной передаче или подвергает хозяина риску из-за других источников опасности, которые не приносят пользы паразиту.
Место заражения
Паразиты-манипуляторы занимают различные места в своих хозяевах. Они обнаруживаются в полости тела, мышцах, ЦНС и других частях хозяина. Здесь мы начнем с изучения того, существуют ли какие-либо закономерности в использовании сайтов в таксономии хозяев или паразитов. Затем мы сосредоточимся на паразитах, которые заражают ЦНС хозяина.
Чтобы лучше понять распределение паразитов среди участков внутри хозяина, мы расширили наш анализ данных (выше), чтобы рассмотреть места заражения манипулятивными паразитами, заражающими позвоночных и беспозвоночных. Мы определили место инфекции как полость тела, мышцу, ЦНС или другое (сердце и кожу). Для упрощения паразиты, обнаруженные в ЦНС и других тканях, были отнесены к категории ЦНС.
Мы обнаружили закономерности, связанные с локализацией инфекции у паразитов, передающихся трофическим путем. Большинство (27) родов встречались в полости тела, 12 были связаны, по крайней мере частично, с ЦНС, 12 были в мышцах и четыре встречались в других тканях. Таксоны паразитов достоверно различались по наиболее частому месту заражения (χ 2 =30,3, d.f.=12, P =0,0026; рис. 4). Два простейших, использованных в анализе, встречались в ЦНС, а скребни встречались только в полости тела. Цестоды часто находились в полости тела, но занимали и другие участки. Нематоды имели тенденцию встречаться в равной степени в мышцах и полости тела. Трематоды использовались в самых разных местах, несколько примеров — в полости тела, мышцах, ЦНС и других местах, таких как кожа. Место заражения варьировалось в зависимости от того, был ли хозяин позвоночным или беспозвоночным (χ 2 = 22,0, d. f. = 3, P = 0,0001), но, как и прежде, это могло быть смешано однородным использованием хозяев скребней и простейших. После исключения этих групп по-прежнему сохранялась тенденция к относительно меньшему количеству паразитов позвоночных в полости тела и относительно меньшему количеству паразитов беспозвоночных в нервной системе (χ 2 = 16,9, d.f. = 12, P = 0,0007; рис. 5), без остаточного действия таксона паразита. Однако систематическая ошибка может быть связана с таксоном хозяина из-за отсутствия подробных сведений о месте заражения у беспозвоночных хозяев по сравнению с хозяевами позвоночных (например, местом заражения беспозвоночных по умолчанию часто является «гемоцель»).
Рис. 4.
Посмотреть в большом размереСкачать слайд
Очаг заражения таксоном паразита. Иногда авторы не определяли место заражения, поэтому использовалась другая литература. Обратите внимание, что полость тела является обычным значением по умолчанию для паразитов беспозвоночных. Все паразиты, которые в той или иной степени были вовлечены в центральную нервную систему (ЦНС), были отнесены к категории обитающих в ЦНС.
Рис. 4.
Посмотреть в большом размереСкачать слайд
Очаг заражения таксоном паразита. Иногда авторы не определяли место заражения, поэтому использовалась другая литература. Обратите внимание, что полость тела является обычным значением по умолчанию для паразитов беспозвоночных. Все паразиты, которые в той или иной степени были вовлечены в центральную нервную систему (ЦНС), были отнесены к категории обитающих в ЦНС.
Паразиты могут изменять поведение хозяина, занимая и повреждая ключевые системы органов. Например, церкарий Diplostomum spatheceum проникают через кожу пресноводной рыбы и мигрируют в глаз, где инцистируются в хрусталике и развиваются в метацеркарии. Тяжелые инфекции приводят к паразитарной катаракте. Зараженные рыбы проводят больше времени, кормясь на поверхности (Crowden and Broom, 1980), предположительно потому, что катаракта ухудшает их визуальную способность охотиться на водных ракообразных, что может сделать их более заметными для птиц-хищников. Зараженные хозяева менее склонны к побегу и более восприимчивы к имитации хищничества в лабораторных условиях (Seppälä et al., 2004; Seppälä et al., 2005). В другом примере корнеголовый ракушка Sacculina carcini сначала разрушает андрогенные железы своего краба-хозяина, что приводит к феминизации самцов-хозяев (Høeg, 1995). Трансгендерные крабы не производят потомства, но ведут себя как самки и заботятся о яйцах ракообразных паразитов, как о своих собственных. Повреждение органов может быть эффективным способом манипулирования хозяином, а в случае корнеголовых крабов это может быть психиатрическим эквивалентом лоботомии.
Ключевой системой органов для координации поведения хозяина является ЦНС. Паразиты, занимающие ЦНС, по-видимому, хорошо приспособлены для манипулирования поведением посредством повреждений или более тонких манипуляций. На самом деле в ЦНС своих хозяев вторгается широкий спектр таксонов паразитов, в том числе простейшие, дигенеи и паразитоидные насекомые (Adamo, 2002; Adamo, 2013; Escobedo et al. , 2009).; Кляйн, 2003; Либерсат и др., 2009 г.; Мур, 2002). Когда-то считалось, что ключевое преимущество паразитирования в ЦНС заключается в том, чтобы ускользнуть от иммунной системы (Szidat, 1969). Теперь мы знаем, что мозг содержит нативные иммунореактивные компоненты, где микроглия, астроциты и даже нейроны имеют на своей клеточной поверхности рецепторы, обнаруживающие патогены (Falsig et al., 2008). Фактически, мозг может повышать свою иммунную реактивность при заражении паразитами или патогенами, что может быть усилено за счет транспорта с помощью специфических белков-переносчиков гематоэнцефалического барьера (Erickson et al., 2012). Следовательно, ключевым преимуществом паразитирования ЦНС, по-видимому, является манипулирование поведением.
Рис. 5.
Просмотреть в большом размереСкачать слайд
Место заражения таксоном-хозяином после исключения скребней и простейших. Другие детали как на рис. 4.
Рис. 5.
Посмотреть в большом размереСкачать слайд
Место заражения таксоном-хозяином после исключения скребней и простейших. Прочие сведения см. на рис. 4.
Toxoplasma gondii — простейший паразит, инфицирующий теплокровных позвоночных, таких как грызуны, в качестве промежуточных хозяев и кошек в качестве окончательных хозяев (см. Flegr, 2013; McConkey et al., 2013; Вьяс, 2013; Вебстер, 2013). В промежуточном хозяине T. gondii размножается бесполым путем (острый токсоплазмоз) до образования тканевых кист в органах, мышцах и, в частности, ЦНС (хронический или латентный токсоплазмоз). Зараженные грызуны усиливают свое исследовательское поведение, и их даже привлекает запах кошачьей мочи, что должно повышать их восприимчивость к хищничеству со стороны кошек и передачу T. gondii его окончательному хозяину (Berdoy et al., 2000; Vyas et al. ., 2007; Вебстер и МакКонки, 2010). Любопытно, что T. gondii продемонстрировал сродство к заражению миндалевидного тела грызунов, области, занимающей центральное место в обработке страха (Berenreiterová et al., 2011). Реактивация хронической инфекции, обычно у лиц с ослабленным иммунитетом, может вызвать воспаление головного мозга (токсоплазматический энцефалит), после чего повышается иммунная реактивность и рекрутируются дендритные клетки в очаг инфекции (John et al. , 2011).
Одним из классических примеров модификации поведения хозяина является печеночная двуустка Dicrocoelium dendriticum во втором промежуточном муравьином хозяине. Взрослые гельминты размножаются в пищеварительном тракте жвачных животных (овец и крупного рогатого скота), выделяя свои яйца с фекалиями хозяина. Яйца проглатываются легочными улитками первого промежуточного хозяина, которые отхаркивают шарики слизи, содержащие инфекционных церкарий. Муравьи заражаются после проглатывания шарика слизи, который является предпочтительной пищей для муравья. Попав в пищеварительный тракт муравья, церкарии мигрируют по телу и инцистируются в гемоцеле муравья, за исключением одной, которая инцистируется в подпищеводном ганглии, контролирующем нижние челюсти. Зараженные муравьи проявляют любопытное поведение на закате, когда температура падает, когда они поднимаются на кончик травинки, зажимают ее челюстями и остаются неподвижными в течение ночи и следующего рассвета. Как только температура повышается, они спускаются вниз и возобновляют нормальную деятельность до следующего вечера. Такое измененное поведение ставит муравьев в выгодное положение для поедания пасущимся домашним скотом, который, как правило, пасется по вечерам и ранним утром, тем самым решая дилемму паразита о переходе от муравья к травоядному и увеличивая его шансы завершить свой жизненный цикл.
Дигенеи семейства Diplostomidae часто паразитируют в головном мозге своих вторых промежуточных хозяев-рыб и развиваются во взрослых особей в пищеварительном тракте окончательных хозяев птиц. Метацеркарии диплостомид крупны по сравнению с мозгом рыб и инцистируются прямо под поверхностью оптических долей, областью, задействованной в визуальной обработке (Muzzall and Kilroy, 2007; Shirakashi and Goater, 2005). Было показано, что метацеркарии Ornithodiplostomum ptychocheilus нарушают визуальную обработку и передвижение у своих хозяев (Shirakashi and Goater, 2005), нарушение, которое может повлиять на способность рыб обнаруживать птицу-хищника и убегать от нее. Однако это происходит до того, как метацеркарии становятся инфекционными для окончательного хозяина, поэтому нарушение оптомоторной активности может быть патологическим побочным эффектом инфекции (Shirakashi and Goater, 2005).
Euhaplorchis californiensis (Heterophyidae), еще один инцистирующий мозг дигеней, изменяет поведение второй промежуточной рыбы-хозяина, калифорнийского киллифиша ( Fundulus parvipinnis ). Зараженные киллифиши проявляют заметное плавательное поведение, что повышает вероятность того, что их съедят окончательные хозяева птицы (Lafferty and Morris, 1996). Эта повышенная восприимчивость к хищничеству со стороны птиц может быть связана с индуцированным паразитами подавлением реакции рыб на стресс. Интересно, E. californiensis метацеркарий скапливаются на стволе мозга при низкоинтенсивных инфекциях (Shaw et al., 2009). Церкарии, инцистирующие мозг, перемещаются в мозг по спинномозговым нервам и, следовательно, сначала проникают в ствол мозга, способствуя инцистированию ствола мозга (Hendrickson, 1979). Возможно, E. californiensis нашел способ извлечь выгоду из своего первоначального местоположения по умолчанию. Интересно, что у гаммарид метацеркарии, инцистирующие мозг, также проявляют предпочтение местонахождения (см. Helluy, 2013). Microphallus papillorobustus инцистируется в протоцеребрум своего гаммаридного хозяина ( Gammarus lacustris ) при единичных инфекциях (Helluy and Thomas, 2003). Хотя размер выборки был слишком мал, чтобы указать на значительную тенденцию, это говорит о том, что паразиты могут отдавать предпочтение определенным областям мозга.
Таким образом, паразиты встречаются в самых разных местах своих хозяев. Основным местом инфекции у беспозвоночных хозяев является полость тела, тогда как у позвоночных хозяев используются более разнообразные участки. Личинки скребней в наших выборках заражали только полость тела, которая также была обычным местом заражения цестод. Дигенейские трематоды использовали самые разнообразные участки. Многие роды используют ЦНС хозяина, по крайней мере, частично. Вопреки ожиданиям, иммунные реакции действительно происходят в ЦНС. Хотя это означает, что ЦНС не является безопасным убежищем для паразитов, она предоставляет им идеальное положение для манипулирования поведением, и, как обсуждается ниже, они могут использовать иммунную защиту хозяина для достижения этой цели.
Иммунная система
Вполне возможно, что паразиты, которым приходится договариваться с иммунной системой хозяина, разовьют способы извлечь выгоду из врожденных защитных механизмов хозяина (Adamo, 1997; Adamo, 2013; Helluy and Thomas, 2003; Thomas et al. , 2005). Кроме того, следует выбирать наследуемые манипулятивные стратегии, повышающие приспособленность паразита [см. Poulin (Poulin, 2010) и ссылки в нем]. Многие исследования выявляют возможные связи между иммунными реакциями хозяина и возникающими в результате физиологическими и поведенческими изменениями, которые паразиты могут использовать в качестве стратегий для усиления передачи (см. Adamo, 2013; McCusker and Kelley, 2013). Исследования, связанные с нейровоспалением, обсуждаются ниже.
Нейровоспаление — это распространенный иммунный ответ головного мозга на повреждение или проникновение патогенов. Несколько исследований показывают, что паразиты могут включать нейровоспалительные реакции хозяина в свою стратегию модификации поведения, такие как оксид азота (NO), стержневые клетки и изменение нейромодуляторов. Хеллуи и Томас (Helluy and Thomas, 2010) обнаружили повышенную активность NO вокруг стенки кисты церебральных метацеркарий у гаммарид-хозяев. Одна из основных эффекторных молекул иммунной системы, NO нацелена на опухоли и патогены как у беспозвоночных, так и у позвоночных (Богдан, 2001; Хеллуй и Томас, 2010), поэтому неудивительно, что повышенная активность NO вблизи вторгшихся паразитов не должна вызывать удивления. Однако NO также действует как нейротрансмиттер (хотя он не хранится в синаптических везикулах и синтезируется по мере необходимости), который может влиять на моноаминергическую активность мозга. Было показано, что NO влияет на высвобождение серотонина (5-HT) и дофамина (DA) у грызунов (Dunn, 2006; Frisch et al., 2000; Helluy and Thomas, 2010), поэтому повышенная активность NO, вероятно, играет роль в снижение серотонинергической активности наблюдается в мозге инфицированных гаммарид (Helluy, Thomas, 2003; Helluy, Thomas, 2010).
Костистые рыбы обладают палочковидными клетками, названными так из-за палочковидных структур, присутствующих внутри каждой клетки, основной функцией которых, по-видимому, является реакция на повреждение тканей, особенно в случаях паразитарной инфекции (Dezfuli et al. , 2007; Matisz et al. и др., 2010; Шмахтенберг, 2007). Метацеркарии, проникающие в мозг, вызывают сильную реакцию палочек у пресноводных гольянов, где палочки окружают червя-нарушителя и их плотность увеличивается по мере того, как развивающиеся метацеркарии наносят больший ущерб (Matisz et al., 2010). Родлет-клетки не обнаруживаются в головном мозге неинфицированных сородичей (Dezfuli et al., 2007). Палочки, как полагают, являются частью более крупной воспалительной реакции, которая включает усиление васкуляризации и пролиферацию фибробластов в месте повреждения (Matisz et al., 2010). Последнее интригует, если учесть, что инцистирование мозга E. californiensis metacercariae экспрессируют три типа факторов роста фибробластов на поверхности своих кист (Shaw et al., 2009). Факторы роста фибробластов стимулируют пролиферацию фибробластов, клеток, которые повсеместно распространены во всех типах соединительной ткани и продуцируют белки коллагена и другие компоненты внеклеточного матрикса. Они также активируются в ответ на повреждение тканей, способствуя заживлению и образованию рубцов. В ЦНС фибробласты играют нейропротекторную роль и могут способствовать нейрогенезу (Erickson et al., 2012; Guillemot and Zimmer, 2011). Остается неясным, почему метацеркарии способствуют активности фибробластов вблизи стенок их кист, но ингибирование факторов роста фибробластов, по-видимому, нарушает способность метацеркарий к агрегации in vitro (J. LaClair and K.D.L., неопубликованные данные).
Паразитарная инфекция вызывает иммунный ответ хозяина, направленный на преодоление вторгшегося паразита. Мало того, что паразиты должны преодолевать иммунную защиту хозяина, чтобы установить инфекцию, устойчивый характер инфекции означает, что они должны постоянно уклоняться или обманывать постоянную бдительность иммунной системы. Кроме того, многие паразиты демонстрируют способность использовать механизмы иммунной защиты хозяина в своих интересах. Тем не менее, растущий объем исследований указывает на обширную перекрестную связь между иммунной системой и другими нейромодулирующими системами (Demas et al. , 2011; Lowry et al., 2007; Webster Marketon and Glaser, 2008; Thompson and Kavaliers, 19).94), так что воздействие паразита на иммунную систему может, в свою очередь, воздействовать на нейромодуляторный путь. Эти системы взаимодействуют через избыточные сигнальные молекулы (например, цитокины, моноаминовые нейротрансмиттеры и гормоны стресса), которые облегчают взаимные взаимодействия обратной связи. Более того, недавние исследования показывают, что паразиты могут изменять нейромодуляторную активность хозяина, как обсуждается ниже.
Нейромодуляция
Нейромодуляторы — это молекулы, которые влияют на долговременные физиологические изменения, в отличие от быстродействующих химических сообщений в синапсе. Гормоны — хороший пример нейромодуляторов, потому что они вызывают изменения в организме, которые могут длиться недели, месяцы и даже годы. Однако традиционное различие между нейротрансмиттерами и нейромодуляторами стало размытым, поскольку было обнаружено, что многие нейротрансмиттеры вызывают длительные физиологические изменения в дополнение к их временным ролям в синаптической передаче. Кроме того, химическое вещество может действовать как нейротрансмиттер или нейромодулятор в зависимости от места его действия, например. у позвоночных норадреналин (норэпинефрин, NE) и DA высвобождаются из надпочечников в виде гормонов, но оба они также являются нейротрансмиттерами по всему телу и ЦНС (Nelson, 19).95). Некоторые из наиболее известных примеров модификации поведения хозяина включают индуцированные паразитами изменения активности нейромодуляторов. Многие паразито-хозяинные системы и задействованные механизмы были подробно рассмотрены (Adamo, 2013; Adamo, 2002; Kavaliers et al., 1999; Klein, 2003; Lefèvre et al., 2009; Libersat et al., 2009; Moore, 2002; Poulin, 2010; Thomas et al., 2005; Thompson and Kavaliers, 1994) и будут кратко изложены здесь.
Моноаминовые нейротрансмиттеры являются мощными нейромодуляторами. К ним относятся катехоламины DA, адреналин (адреналин, EP) и NE, индоламин 5-HT и октопамин (OA), обнаруженные у беспозвоночных. Моноамины влияют на многие типы поведения позвоночных и беспозвоночных, в том числе связанные с активностью, движением, стрессом, социальной активностью и размножением (Fabre-Nys, 19). 98; Либерсат и Пфлюгер, 2004 г.; Нельсон, 1995; Нельсон и Трейнор, 2007 г.; Оверли и др., 2007 г.; Вейгер, 1997; Винберг и Нильссон, 1993). Вызванные паразитами изменения OA, DA и 5-HT занимают видное место в системах хозяин-паразит различных таксонов, что указывает на значительный уровень эволюционной консервативности в структуре и функции этих химических веществ (Libersat and Pflueger, 2004; Pflüger and Stevenson, 2005; Weiger, 1997; Winberg and Nilsson, 1993; см. также Helluy, 2013; Libersat and Gal, 2013; McConkey et al., 2013; Vyas, 2013; Webster et al., 2013). Для сравнения, EP и NE еще не показали существенного влияния на измененное поведение хозяина (Helluy and Holmes, 19).90; Оверли и др., 2001; Шоу и др., 2009 г.; Стиббс, 1985). По этой причине мы сосредоточимся на OA, 5-HT и DA.
OA, по-видимому, аналогичен NE у беспозвоночных, у которых EP и NE имеют гораздо меньшее физиологическое значение, чем у позвоночных (Libersat and Pflueger, 2004; Roeder, 1999; Roeder et al., 2003). ОА является важным нейротрансмиттером и гормоном, который регулирует другие нейромодуляторы и влияет на многие виды поведения, включая реакции «бей или беги», стресс, агрессию, передвижение и кормление (Adamo, 2010; Beckage, 19).97; Либерсат и Пфлюгер, 2004 г.; Снеддон и др., 2003). Две хорошо известные системы «хозяин-паразит» демонстрируют значительную модификацию поведения хозяина в связи с измененной активностью ОА. Гусеницы табачной аскариды ( Manduca sexta ), зараженные личинками паразитоидной осы ( Cotesia congregata ), прекращают питаться и двигаться за 1 день до выхода личинок (Beckage, 1997). После вылупления осы поселяются на кутикуле своего хозяина и окукливаются, что занимает несколько дней. Измененное поведение хозяина, по-видимому, имеет решающее значение для выживания ос, потому что в противном случае гусеница может развернуться и поглотить прикрепленных личинок (Адамо, 2002). Такое поведение совпадает с повышением уровня ОА в гемолимфе хозяина, что является физиологическим изменением, которое, по-видимому, связано с присутствием личинок осы, а не с ядом осы (Adamo, 2002). Уменьшение питания, по-видимому, частично связано с усилением ОА, которое уменьшает сокращения передней кишки — механизм глотания гусеницы — за счет разрушения нейронов лобного ганглия (Адамо, 2002). Хотя личинки, по-видимому, не являются источником избытка ОА, точный механизм того, как личинки вызывают всплеск ОА, остается неизвестным (Адамо, 2002; Адамо, 2010).
ОА является нейротрансмиттером для двигательных путей у беспозвоночных, и, по-видимому, он нарушается скребнеобразным паразитом, который изменяет локомоцию своего хозяина, что в конечном итоге приносит пользу паразиту. Гаммариды ( G. lacustris ), инфицированные личинками скребней Polymorphus paradoxus , демонстрируют положительный фототаксис и необычное цепляющееся поведение, когда они всплывают на поверхность и реагируют на беспокойство, цепляясь за растительность или плавающий материал (Helluy and Holmes, 19).90). Это измененное поведение делает инфицированных гаммарид более восприимчивыми к поеданию пасущимися водоплавающими птицами, которые являются окончательными хозяевами P. parodoxus . Инъекция ОА инфицированным гаммаридам подавляет их реакцию цепляния (Helluy, Holmes, 1990).
Рис. 6.
Просмотреть в большом размереСкачать слайд
Типы механизмов, о которых сообщается для манипулятивных паразитов по типу хозяина. Все паразиты, которые в той или иной степени были вовлечены в центральную нервную систему (ЦНС), были отнесены к категории обитающих в ЦНС.
Рис. 6.
Просмотреть в большом размереСкачать слайд
Типы механизмов манипулятивных паразитов в зависимости от типа хозяина. Все паразиты, которые в той или иной степени были вовлечены в центральную нервную систему (ЦНС), были отнесены к категории обитающих в ЦНС.
Вызванные паразитами изменения в 5-НТ индоламина были зарегистрированы в нескольких системах хозяин-паразит с поразительной модификацией поведения. Гаммариды, инфицированные либо дигенейными метацеркариями ( M. papillorobustus ) или цистаканты скребня ( Polymorphus spp. и Pomphorhynchus spp.) являются фотофильными, что связано с повышенной активностью 5-НТ (Adamo, 2013; Helluy and Holmes, 1990; Lefèvre et al., 2009; Мур, 2002; Тейн и др., 2006). Притяжение к свету может быть вызвано у неинфицированных гаммарид повышением содержания 5-HT посредством прямой инъекции или фармацевтического препарата (Guler and Ford, 2010; Tain et al., 2006; Tain et al., 2007). 5-HT вводили неинфицированным G. lacustris также вызывает аберрантное цепляющееся поведение, проявляемое особями, инфицированными скребнем P. parodoxus (Helluy and Holmes, 1990). Исследования архитектуры нейронов в ЦНС выявляют повышенную серотонинергическую иммунореактивность (окрашивание) у гаммарид, инфицированных различными видами скребней, которые не находятся в прямом контакте с мозгом [таблица 3.1 в Lefèvre et al. (Lefèvre et al., 2009) (см. также Maynard et al., 1996; Tain et al., 2006)]. Напротив, гаммариды, инфицированные M. papillorobustus , единственный паразит, поражающий его мозг, проявляют более низкую серотонинергическую иммунореактивность в определенной оптической области (Helluy and Thomas, 2003). Снижение иммунного окрашивания у инфицированных M. papillorobustus гаммарид может быть частично связано с вызванными паразитами нарушениями морфологии серотонинергических нейронов, хотя дегенеративные нейроны также были обнаружены на противоположном от метацеркарий конце мозга, что позволяет предположить, что дегенерация могла быть результатом снижения серотонинергической иннервации (Helluy and Thomas, 2003). Иммуноокрашивание может выявить несоответствия в стационарном серотонинергическом содержании; однако информация не указывает направление изменений. Увеличение окрашивания 5-НТ может указывать либо на снижение высвобождения нейротрансмиттера, либо на увеличение синтеза для компенсации увеличения высвобождения (Adamo, 2013; Helluy and Thomas, 2003; Tain et al., 2006). Дополнительным типом дополнительного анализа может быть измерение как 5-HT, так и его метаболита, 5-гидроксииндолуксусной кислоты (5-HIAA), в мозге гаммарид с использованием ВЭЖХ; изменение относительных концентраций 5-HT и 5-HIAA указывает на направление оборота (Øverli et al. , 2001; Shaw et al., 2009).). Например, снижение 5-HT и увеличение 5-HIAA будет указывать на увеличение метаболизма 5-HT и, следовательно, метаболизма.
Изменение метаболизма 5-НТ также лежит в основе изменений в поведении хозяина у рыб, зараженных личинками гельминтов. Трехиглая колюшка ( Gasterosteus aculeatus ), зараженная личиночным цепнем Schistocephalus solidus , проявляет различные изменения в поведении, включая повышенное всплытие на поверхность и нарушение реакции побега, что должно сделать зараженную рыбу более восприимчивой к хищничеству со стороны птиц, окончательных хозяев для S. solidus (Barber et al., 2000; Øverli et al., 2001). Инфицированная рыба проявляет повышенную активность 5-НТ в стволе головного мозга, на что указывает увеличение отношения 5-НТ к его метаболиту, хотя это изменение характерно для рыб в условиях хронического стресса, и S. solidus действительно вредит своему хозяину ( Оверли и др., 2001). Напротив, киллифиши, инфицированные штаммом E. californiensis , демонстрируют индуцированное паразитами подавление серотонинергической активности ствола мозга, что является начальным компонентом врожденной реакции рыб на стресс. Нормальная реакция рыб на стресс состоит из начального повышения серотонинергической активности ствола мозга, что запускает каскад других физиологических процессов, включая всплеск циркулирующих гормонов стресса (Winberg and Nilsson, 19).93). Нарушение этой первоначальной физиологической реакции на стресс может в конечном итоге ухудшить реакцию побега киллифиша.
Поразительные изменения поведения позвоночных и беспозвоночных, вызванные 5-HT, демонстрируют повсеместное распространение 5-HT и его широкую функциональность у животных различной сложности. 5-HT является основным нервно-мышечным нейротрансмиттером, используемым у самых примитивных животных (например, книдарий), и его роль расширяется до нейромодулятора и нейрогормона у более сложных животных, где он контролирует дополнительные физиологические функции и поведение, включая реакции на стресс и иммунные проблемы (Weiger, 1997). Реакция на стресс состоит из активации гипоталамо-гипофизарно-надпочечниковой оси, где мозг сигнализирует о высвобождении гормонов стресса и моноаминов, которые, в свою очередь, активируют и регулируют лимфоциты и другие иммунные эффекторные белки, такие как цитокины (Webster Marketon and Glaser, 2008). . Вполне вероятно, что любое нейрохимическое вещество, играющее важную роль в физиологической реакции на стресс, будет влиять на иммунный ответ (Adamo, 2010; Wendelaar Bonga, 1997; de Jong-Brink et al., 2001; Demas et al., 2011; Escobedo et al. , 2009 г.; Webster Marketon and Glaser, 2008), и вызванные паразитами изменения в нейроактивных химических веществах хозяина могли возникнуть в результате взаимной коммуникации обеих систем (Adamo, 2013; Adamo and Baker, 2011; Escobedo et al., 2009; Thomas et al., 2005). ). Например, 5-НТ может активировать и регулировать активность гипоталамо-гипофизарно-надпочечниковой оси у млекопитающих (гипоталамо-гипофизарно-интерренекулярную у рыб), систему нейроэндокринных путей, которые координируют физиологическую реакцию на стресс и болезнь посредством общих эффекторных молекул, таких как нейромодуляторы. (Чаулофф, 2000; Клементс и др., 2003; Вебстер Маркетон и Глейзер, 2008; Винберг и др., 19).97) и, в свою очередь, регулируется иммунными белками (Dunn, 2006).
Было продемонстрировано, что измененная активность дофамина вызывает поведенческие изменения, наблюдаемые в поразительных примерах модификации поведения паразитами у ракообразных. DA стимулирует активность ракообразных и связан с усилением боевой позы и поведения (Sneddon et al., 2003). Береговые крабы ( Hemigrapsus crenulatus ), инфицированные цистакантами беспозвоночных Profilicollis antarticus , более активны, чем неинфицированные (Haye, Ojeda, 19).98). DA гемолимфы увеличивается у инфицированных крабов, что может способствовать повышенной активности (Rojas and Ojeda, 2005).
Паразитозависимые изменения в активности дофамина в головном мозге также очевидны у килллифов, инфицированных метацеркариями E. californiensis . Зараженные киллифиши имеют повышенную активность DA в стволе мозга (Shaw et al. , 2009). DA связан с повышенной локомоцией, агрессией, доминированием и репродуктивным поведением у рыб и млекопитающих (Winberg and Nilsson, 1993; Fabre-Nys, 19).98), а повышенная дофаминергическая активность у киллифи может способствовать усилению проявлений заметной плавательной активности (Shaw et al., 2009). Однако дофаминергические нейроны в стволе головного мозга также ингибируют серотонинергические нейроны, которые проецируются из ствола в средний мозг, и, таким образом, могут способствовать индуцированному паразитами подавлению активности 5-HT в стволе мозга, наблюдаемому у инфицированных киллифишей (Shaw et al., 2009).
В нескольких исследованиях изменения активности дофамина рассматриваются как возможный механизм, лежащий в основе измененного поведения, наблюдаемого у грызунов, инфицированных T. gondii [подробные обзоры см. в Webster and McConkey (Webster and McConkey, 2010) и Adamo (Adamo, 2013)]. Активность DA в головном мозге повышена у мышей, инфицированных T. gondii (Stibbs, 1985), а введение агониста рецептора DA вызывает поведение поиска новизны у неинфицированных мышей (Skallová et al., 2006). Эти результаты согласуются с общим представлением о том, что дофамин стимулирует передвижение, агрессию, доминирование и репродуктивное поведение у рыб и млекопитающих (Winberg and Nilsson, 199).3; Фабр-Нис, 1998).
Антипсихотические препараты так же эффективны, как и антипаразитарные препараты, в снижении вызванных Toxoplasma изменений поведения, и наиболее эффективным препаратом является антагонист дофаминовых рецепторов (Webster et al., 2006). Совсем недавно было обнаружено, что T. gondii экспрессирует гены тирозингидроксилазы, фермента, который синтезирует дофамин, а также увеличивает метаболизм дофамина в нейронах хозяина (Gaskell et al., 2009; Prandovszky et al., 2011). Эта повышенная сигнальная активность инфицированных нейронов может управлять изменениями поведения, связанными с DA, особенно в сильно дофаминергических миндалинах и базальных ганглиях, которые являются важными областями для формирования страха и двигательного поведения, соответственно (Prandovszky et al. , 2011).
Эти связанные с паразитами изменения активности моноаминов в различных системах паразит-хозяин, указанные выше, указывают на высокий уровень консервативной функции на протяжении всей эволюции беспозвоночных и позвоночных (Libersat and Pflueger, 2004; Pflüger and Stevenson, 2005; Weiger, 1997; Winberg и Нильсон, 1993). Однако детали специфических механизмов действия ОА, 5-НТ и ДА в формировании поведения зависят от конкретных типов рецепторов и их локализации в разных областях мозга и ЦНС (Adamo, 2013; Demas et al., 2011; Fabre- Нысь, 1998; Такахаши и др., 2011; Winberg and Nilsson, 1993), и это следует учитывать при интерпретации механистических исследований. Например, рецепторы 5-НТ могут снижать или усиливать агрессию в зависимости от их локализации в мозге позвоночных (Summers and Winberg, 2006; Takahashi et al., 2011).
Несмотря на обширные исследования, описанные в настоящем документе, остается неизвестным, изменяют ли паразиты нейромодуляторы хозяина, секретируя нейрохимические вещества, вызывающие изменения в нейрохимической активности хозяина, или секретируя химические вещества, которые вызывают реакцию хозяина, приводя к изменению нейрохимической активности (Adamo, 2013; de Jong -Бринк и др. , 2001; Томас и др., 2005). Экономным объяснением было бы последнее, учитывая, что паразиты часто намного меньше своих хозяев, и для них может быть слишком дорого производить точные химические вещества, необходимые для достижения уровней системных изменений, наблюдаемых у хозяев (Adamo, 2013; de Jong). -Бринк и др., 2001; Томас и др., 2005).
Из-за сложности и взаимосвязанности систем нейромодуляторов ученым сложно выделить конкретные механизмы, ответственные за дискретные изменения в поведении. Однако сложная природа этих систем может также позволить паразитам вызывать широкомасштабные изменения в поведении своих хозяев с минимальными усилиями. Например, моноамины регулируют друг друга в регуляторных петлях обратной связи, т.е. OA и DA стимулируют у насекомых поведение побега, которому противодействует 5-HT (Gavra and Libersat, 2011). Точно так же дофамин способствует передвижению, агрессии и другим видам доминантного поведения у позвоночных, в то время как 5-НТ ингибирует то же поведение (Winberg and Nilsson, 19). 93). Теоретически паразиту достаточно изменить активность одного из нейромодуляторов, участвующих в петле обратной связи, чтобы произвести регуляторные изменения в других компонентах.
Вызванные паразитами изменения в нейромодуляторах хозяина могли возникнуть в результате взаимного взаимодействия нейроэндокринной и иммунной систем (Adamo, 2013; Adamo and Baker, 2011; Escobedo et al., 2009; Thomas et al., 2005). Паразиты должны договариваться и преодолевать иммунную систему своих хозяев, и при этом могли извлечь выгоду из поведенческих изменений, которые произошли в результате изменений иммунного ответа в физиологии. Адамо (Adamo, 2013) отмечает, что для паразитов было бы всего лишь небольшим эволюционным шагом вызвать изменения в поведении хозяина из-за попыток иммуносупрессии хозяина. Действительно, паразиты используют различные нейрохимические методы для запуска изменений в физиологии хозяина, а не нацелены на какую-то одну систему или путь нейротрансмиттеров (Adamo, 2013; de Jong-Brink et al. , 2001; Lefèvre et al., 2009).; Томас и др., 2005). Высокая степень взаимосвязи и перекрестной коммуникации того, что называют нейроиммуноэндокринной системой (Escobedo et al., 2009), возможно, позволяет паразитам влиять на изменения во многих системах с минимальными энергетическими усилиями. Устойчивая активация эффекторных белков иммунной системы (цитокинов) необходима для поддержания устойчивости к острым и хроническим инфекциям T. gondii , и только это может изменить нейромодуляторную активность (Webster and McConkey, 2010). Еще одним доводом в пользу этой гипотезы является структура паразитов, неспособных изменить поведение неместных хозяев, что указывает на уровень локальной адаптации к иммунной системе нативного хозяина и, возможно, ключ к механизмам модификации поведения (Ballabeni and Ward, 1993; Корнет и др., 2010; Тейн и др., 2006).
Подводя итог разделу о механизмах, можно сказать, что у паразитов есть множество способов изменить поведение хозяина. Группа паразитов имела почти значительный эффект (χ 2 = 35,3, d. f. = 24, P = 0,064), при этом скребни были связаны с моноаминами, цестоды связаны с утечкой энергии и гормонами, а дигенеи не связаны с гормонами. Было обнаружено, что механизмы более или менее равномерно распределяются среди позвоночных и беспозвоночных хозяев (рис. 6). Эти результаты слишком преждевременны, чтобы делать выводы об общих закономерностях. Однако механизмы, используемые паразитами, распределены по группам, поэтому, по-видимому, существует множество независимых эволюций различных механизмов. Единственный механизм, доступный для всех паразитов, — это утечка энергии. Это несложный механизм, но в некоторых ситуациях он может способствовать передаче паразитов. Воздействие паразита может быть ограничено тканями хозяина, на которых он живет. По этой причине место заражения может дать ключ к разгадке механизмов поведенческих манипуляций. Из всех сайтов ЦНС больше всего связана со способностью манипулировать поведением хозяина и более представлена по сравнению с другими механизмами (рис. 5). Мы согласны с тем, что ЦНС не свободна от иммунной защиты. На самом деле паразиты могут использовать иммунную защиту в своих интересах, потому что воспаление связано с различными нейромодуляторами. Способность производить, подавлять или активировать нейромодуляторы является вершиной изощренности манипулятивных паразитов.
В этом обзоре мы предположили, что изменение поведения, связанное с паразитом, является адаптацией, которая приносит пользу паразиту. Существуют заслуживающие доверия примеры манипулятивных паразитов из самых разных таксонов. Скребни отличались от других паразитов тем, что они были зарегистрированы только в полости тела беспозвоночных и с большей вероятностью изменяли микросреду обитания хозяина, чем активность. Зараженные позвоночные хозяева с большей вероятностью проявляли нарушенную реакцию на хищников, а инфицированные хозяева-беспозвоночные чаще контактировали с хищниками. Паразиты в мышцах, ЦНС и других тканях чаще ассоциировались со сниженной реакцией на хищников (нарушение реакции), тогда как паразиты в полости тела чаще ассоциировались с повышенным контактом с хищниками. Немногие исследования показали три ключевых доказательства, необходимых для понимания того, как паразиты манипулируют своими хозяевами: изменение поведения, связанное с паразитизмом, положительная связь между измененным поведением и приспособленностью паразита, а также механизмы, с помощью которых паразиты изменяют поведение. Из них понимание механизмов является наиболее сложным. Несмотря на эти проблемы, у нас есть масса информации о том, как паразиты могут влиять на иммунную систему и нейромодулирующие химические вещества. Кажется разумным предположить, что эти сложные механизмы развились в ходе естественного отбора, чтобы изменить поведение хозяина в пользу паразита.
Финансирование
Это исследование не получило специального гранта от какого-либо финансирующего агентства в государственном, коммерческом или некоммерческом секторах.
Адамо
С. А.
(
1997
).
Как паразиты изменяют поведение своих насекомых-хозяев
. В
Parasites and Pathogens: Effects on Host Hormones and Behavior
(ed.
Beckage
N.E.
), стр.
231
–
245
.
Нью-Йорк
:
Чепмен энд Холл
.
Адамо
С. А.
(
2002
).
Модуляция модуляторов: паразиты, нейромодуляторы и изменение поведения хозяина
.
Поведение мозга. Эвол.
60
,
370
–
377
.
Адамо
С. А.
(
2010
).
Почему иммунный ответ должен активировать реакцию на стресс? Информация от насекомых (сверчок Gryllus texensis)
.
Поведение мозга. Иммун.
24
,
194
–
200
.
Адамо
С. А.
(
2013
).
Паразиты: нейробиологи эволюции
.
J. Расшир. биол.
216
,
3
–
10
.
Адамо
С. А.
,
Бейкер
Дж. Л.
(
2011
).
Сохраняющиеся признаки хронического стресса у различных типов: влияние длительного стресса на поведение и концентрацию нейрогормона октопамина у сверчка Gryllus texensis
.
Горм. Поведение
60
,
478
–
483
.
Баллабени
П.
,
Уорд
ИП
(
1993
).
Местная адаптация тремадота Diplostomum phoxini к европейскому гольяну Phoxinus phoxinus, его второму промежуточному хозяину
.
Функц. Экол.
7
,
84
–
90
.
Парикмахерская
I.
,
Хоар
Д.
,
Краузе
Дж.
(
2000
3).
Воздействие паразитов на поведение рыб: обзор и эволюционная перспектива
.
Rev. Fish Biol. Рыбы.
10
,
131
–
165
.
Бекедж
Н.Е.
(
1993
).
Эндокринные и нейроэндокринные отношения паразит-хозяин
.
Рецептор
3
,
233
–
245
.
Бекедж
Н.Е.
(
1997
).
Паразиты и патогены: влияние на гормоны хозяина и поведение
.
Нью-Йорк
:
Чепмен и Холл
.
Бердой
М.
,
Вебстер
Дж. П.
,
Макдональд
Д. В.
(
2000
).
Смертельное влечение у крыс, инфицированных Toxoplasma gondii
.
Проц. Р. Соц. Лонд. Б
267
,
1591
–
1594
.
Berenreiterová
M.
,
Flegr
J.
,
Kuběna
A. A.
,
NěMEC
стр.
(
2011
).
Распределение цист Toxoplasma gondii в головном мозге мыши с латентным токсоплазмозом: значение для гипотезы поведенческих манипуляций
.
PLoS ONE
6
,
e28925
.
Богдан
К.
(
2001
).
Оксид азота и иммунный ответ
.
Нац. Иммунол.
2
,
907
–
916
.
Бернем
К. П.
,
Андерсон
Д. Р.
(
2004
).
Мультимодельный вывод – понимание AIC и BIC при выборе модели
.
Соц. Методы Рез.
33
,
261
–
304
.
Шаулофф
Ф.
(
2000
).
Серотонин, стресс и кортикоиды
.
J. Psychopharmacol.
14
,
139
–
151
.
Клементс
S.
,
Moore
F. L.
,
Schreck
C. B.
(
2003
).
Доказательства того, что острая серотонинергическая активация потенцирует двигательно-стимулирующие эффекты кортикотропин-высвобождающего гормона у молоди чавычи (Oncorhynchus tshawytscha)
.
Горм. Поведение
43
,
214
–
221
.
Cornet
S.
,
Sorci
G.
,
Морет
Y.
(
2010
).
Биологическая инвазия и паразитизм: инвазионисты не страдают от физиологических изменений скребня Pomphorhynchus laevis
.
Паразитология
137
,
137
–
147
.
Крауден
А. Е.
,
Метла
Д. М.
(
1980
).
Влияние двуустки Diplostomum spathaceum на поведение ельца (Leuciscus leuciscus)
.
Аним. Поведение
28
,
287
–
294
.
День
Дж. Ф.
,
Эдман
Дж. Д.
(
1983
).
Малярия делает мышей восприимчивыми к питанию комарами, когда гаметоциты наиболее заразны
.
Ж. Паразитол.
69
,
163
–
170
.
де Йонг-Бринк
М.
,
Бергамин-Сассен
М.
,
Солис Сото
М.
(
2001
).
Множественные стратегии шистосом для удовлетворения своих потребностей в промежуточном хозяине-улитке
.
Паразитология
123
,
S129
–
S141
.
Demas
G. E.
,
Adamo
S. A.
,
French
S.
(
2011
).
Нейроэндокринно-иммунные перекрестные помехи у позвоночных и беспозвоночных: значение для защиты хозяина
.
Функц. Экол.
25
,
29
.
Dezfuli
B. S.
,
Giari
L.
,
Shinn
A. P.
(
2007
).
Роль палочек в воспалительной реакции в головном мозге Phoxinus phoxinus, инфицированном Diplostomum
.
Рыба Моллюски Immunol.
23
,
300
–
304
.
Данн
Эй Джей
(
2006
).
Влияние цитокинов и инфекций на нейрохимию головного мозга
.
Клин. Неврологи. Рез.
6
,
52
–
68
.
Эриксон
М. А.
,
Дохи
К.
,
Банки
В. А.
(
2012
).
Нейровоспаление: общий путь при заболеваниях ЦНС, опосредованный через гематоэнцефалический барьер
.
Нейроиммуномодуляция
19
,
121
–
130
.
Эскобедо
Г.
,
Лопес-Гриего
Л.
,
Моралес-Монтор
Дж.
(
2009
).
Нейроиммуноэндокринная модуляция хозяина паразитами-гельминтами: новая форма коэволюции хозяина и паразита?
Нейроиммуномодуляция
16
,
78
–
87
.
Фабр-Нис
К.
(
1998
).
Стероидный контроль моноаминов в отношении сексуального поведения
.
Ред. Репрод.
3
,
31
–
41
.
Falsig
J.
,
Van Beek
J.
,
Hermann
C.
,
Leist
M.
(
8 2008
).
Молекулярная основа для обнаружения проникающих патогенов в головной мозг
.
J. Neurosci. Рез.
86
,
1434
–
1447
.
Флегр
Дж.
(
2013
).
Влияние латентной токсоплазменной инфекции на личность, физиологию и морфологию человека: плюсы и минусы модели токсоплазма-человек при изучении гипотезы манипуляции
.
J. Расшир. биол.
216
,
127
–
133
.
Frisch
C.
,
Dere
E.
,
Silva
M. A. D.
,
Godecke
A.
,
Schrader
J.
,
Хьюстон
Дж. П.
(
2000
).
Превосходная эффективность прохождения водного лабиринта и усиление поведения, связанного со страхом, у мышей с дефицитом эндотелиальной синтазы оксида азота вместе с изменениями моноаминов в мозжечке и вентральном стриатуме
.
J. Neurosci.
20
,
6694
–
6700
.
Gaskell
E. A.
,
Smith
J. E.
,
Pinney
J. W.
,
Westhead
D. R.
,
McConkey
G. A.
(
2009
).
Уникальная аминокислотная гидроксилаза двойной активности у Toxoplasma gondii
.
PLoS ONE
4
,
e4801
.
Гавра
Т.
,
Либерсат
Ф.
(
2011
).
Вовлечение опиоидной системы в гипокинетическое состояние, индуцированное у тараканов паразитоидной осой
.
Дж. Комп. Физиол. А
197
,
279
–
291
.
Джайлз
Н.
(
1987
).
Риск хищничества и снижение активности рыб в поисках пищи: эксперименты с паразитированной и непаразитированной трехиглой колюшкой, Gasterosteus aculeatus L
.
J. Fish Biol.
31
,
37
–
44
.
Guillemot
F.
,
Zimmer
C.
(
2011
).
От колыбели до могилы: многочисленные роли факторов роста фибробластов в развитии нервной системы
.
Нейрон
71
,
574
–
588
.
Гулер
Ю.
,
Форд
А. Т.
(
2010
).
Антидепрессанты помогают амфиподам увидеть свет
.
Вода. Токсикол.
99
,
397
–
404
.
Хэй
П. А.
,
Охеда
Ф. П.
(
1998
).
Метаболические и поведенческие изменения у краба Hemigrapsus crenulatus (Milne-Edwards 1837), вызванные его скребневым паразитом Profilicollis antarcticus (Zdzitowiecki 1985)
.
J. Расшир. Мар биол. Экол.
228
,
73
.
Хеллуй
С.
(
2013
).
Паразито-индуцированные изменения сенсомоторных путей у гаммарид: побочное повреждение нейровоспаления?
J. Расшир. биол.
216
,
67
–
77
.
Хеллуй
С.
,
Холмс
Дж. К.
(
1990
).
Серотонин, октопамин и цепляние, вызванное паразитом Polymorphus paradoxus (Acanthocephala) у Gammarus lacustris (Crustacea)
.
Кан. Дж. Зул.
68
,
1214
–
1220
.
Хеллуй
С.
,
Томас
Ф.
(
2003
).
Влияние Microphallus papillorobustus (Platyhelminthes: Trematoda) на серотонинергическую иммунореактивность и архитектуру нейронов в головном мозге Gammarus insensibilis (Crustacea: Amphipoda)
.
Проц. Р. Соц. Б
270
,
563
–
568
.
Хеллуй
С.
,
Томас
Ф.
(
2010
).
Паразитарные манипуляции и нейровоспаление: данные из системы Microphallus papillorobustus (Trematoda) – Gammarus (Crustacea)
.
Паразит. Векторы
3
,
38
.
Хендриксон
Г. Л.
(
1979
).
Ornithodiplostomum ptychocheilus: миграция в мозг промежуточного хозяина рыбы, Pimephales promelas
.
Расшир. Паразитол.
48
,
245
–
258
.
Хёг
Дж. Т.
(
1995
).
Биология и жизненный цикл корнеголовых (Cirripedia)
.
J. Mar. Biol. доц. Великобритания
75
,
517
–
550
.
John
B.
,
Ricart
B.
,
Tait Wojno
E. D.
,
Harris
T. H.
,
Randall
L. M.
,
Кристиан
Д. А.
,
Грегг
B.
,
de Almeida
D. M.
,
Weninger
W.
,
Hammer
D. A.
, et al. . (
2011
).
Анализ поведения и транспорта дендритных клеток в головном мозге при токсоплазматическом энцефалите
.
PLoS Патог.
7
,
e1002246
.
Джоли
Д. О.
,
Мессье
Ф.
(
2004
).
Распространение Echinococcus granulosus у лосей: свидетельство индуцированной паразитами уязвимости перед хищничеством волков?
Экология
140
,
586
–
590
.
Кавальерс
М.
,
Колуэлл
Д. Д.
,
Холерис
Е.
(
1999
).
Паразиты и поведение: этофармакологический анализ и биомедицинские последствия
.
Неврологи. Биоповедение.
23
,
1037
–
1045
.
Кляйн
С. Л.
(
2003
).
Воздействие паразитов на непосредственные механизмы, опосредующие социальное поведение позвоночных
.
Физиол. Поведение
79
,
441
–
449
.
Лафферти
К. Д.
,
Моррис
А. К.
(
1996
). .
Экология
77
,
1390
–
1397
.
Lefèvre
T.
,
Adamo
S. A.
,
Biron
D. G.
,
Missé
D.
,
Hughes
D.
,
Томас
Ф.
(
2009
).
Вторжение похитителей тел: разнообразие и эволюция манипулятивных стратегий при взаимодействии хозяина и паразита
.
Доп. Паразитол.
68
,
45
–
83
.
Libersat
F.
,
Gal
R.
(
2013
).
Чему паразитоидные осы могут научить нас в отношении принятия решений у насекомых?
J. Расшир. биол.
216
,
47
–
55
.
Либерсат
F.
,
Pflueger
HJ
(
2004
).
Моноамины и управление поведением
.
Биология
54
,
17
–
25
.
Libersat
F.
,
Delago
A.
,
GAL
R.
(
2009
).
Манипулирование поведением хозяина с помощью паразитических насекомых и насекомых-паразитов
.
год. Преподобный Энтомол.
54
,
189
–
207
.
Lowry
C. A.
,
Hollis
J. H.
,
de Vries
A.
,
Pan
B.
,
Brunet
L. R.
,
Hunt
J. R. F.
,
Paton
J. F. R.
,
Van Kampen
E.
,
Knight
D. M. M.
,
Evans
D. M. M.
,
Evans
A.K.
,
. . (
2007
).
Идентификация иммунореактивной мезолимбокортальной серотонинергической системы: потенциальная роль в регуляции эмоционального поведения
.
Неврология
146
,
756
–
772
.
Matisz
C. E.
,
Goater
C. P.
,
Bray
D.
(
2010
).
Плотность и созревание стержневых клеток в мозговой ткани толстоголовых гольянов (Pimephales promelas), подвергшихся воздействию церкарий трематод
.
Междунар. Дж. Паразитол.
40
,
307
–
312
.
Maynard
B. J.
,
Demartini
L.
,
Wright
W. G.
(
1996
).
Gammarus lacustris, укрывающий Polymorphus paradoxus, проявляет измененные модели серотониноподобной иммунореактивности
.
Ж. Паразитол.
82
,
663
–
666
.
McConkey
G. A.
,
Martin
H. L.
,
Bristow
G. C.
,
Webster
J. P.
(
2013
).
Инфекция и поведение Toxoplasma gondii – место, место, место?
J. Расшир. биол.
216
,
113
–
119
.
Маккаскер
Р. Х.
,
Келли
К. В.
(
2013
).
Иммуно-нейральные связи: как реакция иммунной системы на инфекционные агенты влияет на поведение
.
J. Расшир. биол.
216
,
84
–
98
.
Милинский
М.
(
1984
).
Паразиты определяют оптимальную стратегию питания хищника
.
Поведение. Экол. Социобиол.
15
,
35
–
37
.
Мур
Дж.
(
2002
).
Паразиты и поведение животных
.
Нью-Йорк
:
Oxford University Press
.
Muzzall
P. M.
,
Kilroy
A. L.
(
2007
).
Tylodelphys scheuringi (Diplostomidae), заражающий мозг центрального гольяна Umbra limi, в Силвер-Крик, Мичиган, США
.
Комп. Паразитол.
74
,
164
.
Нельсон
Р. Дж.
(
1995
).
Введение в поведенческую эндокринологию
.
Сандерленд, Массачусетс
:
Sinauer Associates
.
Нельсон
Р. Дж.
,
Трейнор
Б. К.
(
2007
).
Нейронные механизмы агрессии
.
Нац. Преподобный Нейроски.
8
,
536
–
546
.
Верли
Ø.
,
Палл
М.
,
Борг
Б.
,
Джоблинг
М.
,
Винберг
С.
(
2001
).
Влияние инфекции Schistocephalus solidus на моноаминергическую активность головного мозга самок трехиглой колюшки Gasterosteus aculeatus
.
Проц. Р. Соц. Лонд. Б
268
,
1411
–
1415
.
Верли
Ø.
,
Соренсен
C.
,
Pulman
K. G. T.
,
Pottinger
T. G.
,
Korzan
W. J.
,
Summers
C. H.
,
Nilsson
G. E.
(
2007
).
Эволюционные предпосылки для стилей преодоления стресса: взаимосвязь между физиологическими, поведенческими и когнитивными чертами у немлекопитающих позвоночных
.
Неврологи. Биоповедение.
31
,
396
–
412
.
Перро-Минно
М.-Ж.
,
Сезилли
Ф.
(
2013
).
Исследование потенциальных нейромодуляторных систем, лежащих в основе паразитарных манипуляций: концепции, ограничения и перспективы
.
J. Расшир. биол.
216
,
134
–
141
.
Pflüger
HJ
,
Stevenson
P. A.
(
2005
).
Эволюционные аспекты октопаминергических систем с акцентом на членистоногих
.
Структура членистоногих. Дев.
34
,
379
.
Пулен
Р.
(
1994
).
Метаанализ поведенческих изменений, вызванных паразитами
.
Аним. Поведение
48
,
137
–
146
.
Пулен
Р.
(
2010
).
Манипулирование паразитами поведением хозяина: новости и часто задаваемые вопросы
.
Доп. Стад. Поведение
41
,
151
–
186
.
Пулен
Р.
(
2013
).
Манипуляции паразитами с личностью хозяина и поведенческие синдромы
.
J. Расшир. биол.
216
,
18
–
26
.
Прандовски
Э.
,
Гаскелл
Э.
,
Мартин
Х.
39003
0003J. P.
,
Webster
J. P.
,
McConkey
G. A.
22004 (310 90 0103 9004).
Нейротропный паразит Toxoplasma gondii увеличивает метаболизм дофамина
.
PLoS ONE
6
,
e23866
.
Редер
Т.
(
1999
).
Октопамин в беспозвоночных
.
Прог. Нейробиол.
59
,
533
–
561
.
Roeder
T.
,
SEIFERT
M.
,
Kähler
C.
,
Gewecke
M.
(
2003
4444444444444444444444.
.Тирамин и октопамин: антагонистические модуляторы поведения и метаболизма
.
Арх. Биохимия насекомых. Физиол.
54
,
1
–
13
.
Рохас
Дж. М.
,
Охеда
Ф. П.
(
2005
).
Измененные уровни дофамина, вызванные паразитом Profilicollis antarcticus у его промежуточного хозяина, краба Hemigrapsus crenulatus
.
Биол. Рез.
38
,
259
–
266
.
Шмахтенберг
О.
(
2007
).
Эпителиальные стражи или простейшие паразиты? Исследования на изолированных палочках к 100-летию загадки
.
Ред. Чил. История Нац.
80
,
55
.
Seppälä
O.
,
Karvonen
A.
,
Valtonen
E. T.
(
2004
).
Вызванное паразитами изменение поведения хозяина и восприимчивость к хищникам при взаимодействии глазной двуустки и рыбы
.
Аним. Поведение
68
,
257
–
263
.
Seppälä
O.
,
Karvonen
A.
,
Valtonen
E. T.
(
2005
).
Нарушение крипсии рыбы, зараженной паразитом, передающимся трофическим путем
.
Аним. Поведение
70
,
895
–
900
.
Shaw
J. C.
,
Korzan
W. J.
,
Carpenter
R. E.
,
Kuris
A. M.
,
Lafferty
K. D.
,
Лето
C. H.
,
Верли
Ø.
(
2009
).
Паразитическая манипуляция моноаминами головного мозга калифорнийской киллифии (Fundulus parvipinnis) с помощью трематоды Euhaplorchis californiensis
.
Проц. биол. науч.
276
,
1137
–
1146
.
Ширакаши
С.
,
Козел
КП
(
2005
).
Хронология вызванного паразитами изменения поведения рыб: влияние созревания паразита и опыта хозяина
.
Паразитология
130
,
177
–
183
.
Скаллова
А.
,
Кодым
П.
,
Фрынта
Д. 90
,
0003Дж.
(
2006
).
Роль дофамина в вызванных токсоплазмой поведенческих изменениях у мышей: этологическое и этофармакологическое исследование
.
Паразитология
133
,
525
–
535
.
Sneddon
L. U.
,
Braithwaite
V. A.
,
Gentle
M. J.
0004 (2003
).
Есть ли у рыб ноцицепторы? Доказательства эволюции сенсорной системы позвоночных
.
Проц. Р. Соц. Лонд. Б
270
,
1115
–
1121
.
Стиббс
Х. Х.
(
1985
).
Изменения концентрации катехоламинов и индоламинов в головном мозге мышей, инфицированных Toxoplasma gondii
.
Энн. Троп. Мед. Паразитол.
79
,
153
–
157
.
Саммерс
К. Х.
,
Винберг
С.
(
2006
).
Взаимодействие нервной регуляции стресса и агрессии
.
J. Расшир. биол.
209
,
4581
–
4589
.
Сидат
Л.
(
1969
).
Структурное развитие и поведение новых стригеатоидных метацеркарий из субтропических рыб Южной Америки
.
Дж. Рыба. Рез. Совет может.
26
,
753
.
Tain
L.
,
Perrot-Minnot
M.-J.
,
Сезилли
Ф.
(
2006
).
Изменение поведения хозяина и серотонинергическая активность мозга, вызванные скребнями: свидетельство специфичности
.
Проц. биол. науч.
273
,
3039
–
3045
.
Tain
L.
,
Perrot-Minnot
M.-J.
,
Сезилли
Ф.
(
2007
).
Дифференциальное влияние Pomphorhynchus laevis (Acanthocephala) на серотонинергическую активность головного мозга у двух родственных видов-хозяев
.
Биол. лат.
3
,
69
–
72
.
Takahashi
A.
,
Quadros
I. M.
,
De Almeida
R. M. M.
,
Miczek
K. A.
(
Miczek
K. A.
(
K. A.
(
K. A.
(
K. A.
(
.0003
2011
).
Мозговые рецепторы и транспортеры серотонина: инициация и прекращение эскалации агрессии
.
Психофармакология
213
,
183
–
212
.
Thomas
F.
,
Adamo
S.
,
Moore
J.
(
2005
).
Паразитическая манипуляция: где мы и куда нам идти?
Поведение. Процессы
68
,
185
–
199
.
Томпсон
С. Н.
,
Кавальерс
М.
(
1994
).
Физиологические основы вызванных паразитами изменений поведения хозяина
.
Паразитология
109
,
S119
–
S138
.
Вьяс
А.
(
2013
).
Паразито-усиленный выбор партнера и снижение врожденного страха у крыс, инфицированных Toxoplasma gondii
.
J. Расшир. биол.
216
,
120
–
126
.
Вьяс
А.
,
Ким
С.-К.
,
Джакомини
Н.
,
Бутройд
JC
,
Sapolsky
R. M.
(
2007
).
Поведенческие изменения, вызванные заражением грызунов токсоплазмой, очень специфичны для отвращения кошачьих запахов
.
Проц. Натл. акад. науч. США
104
,
6442
–
6447
.
Webster
J. P.
,
McConkey
G. A.
(
2010
).
Поведение хозяина, измененное Toxoplasma gondii: сведения о механизме действия
.
Фолиа Паразитол.
57
,
95
–
104
.
Webster
J. P.
,
Lamberton
P. H. L.
,
Donnelly
C. A.
,
Torrey
E. F.
(
0003
2006
).
Паразиты как возбудители аффективных расстройств человека? Влияние антипсихотических препаратов, стабилизаторов настроения и противопаразитарных препаратов на способность Toxoplasma gondii изменять поведение хозяина
.
Проц. Р. Соц. Лонд. Б
273
,
1023
–
1030
.
Webster
J. P.
,
Кошик
М.
,
Bristow
G.C.
,
McConkey
G.A.
(
2013
).
Инфекция Toxoplasma gondii, от хищничества до шизофрении: может ли поведение животных помочь нам понять поведение человека?
J. Расшир. биол.
216
,
99
–
112
.
Webster Marketon
J. I.
,
Glaser
R.
(
2008
).
Гормоны стресса и иммунная функция
.
Сотовый. Иммунол.
252
,
16
–
26
.
Weiger
W. A.
(
1997
).
Серотонинергическая модуляция поведения: филогенетический обзор
.
Биол. Преподобный Кэмб. Филос. соц.
72
,
61
–
95
.
Венделаар Бонга
Ю. Е.
(
1997
).
Реакция рыб на стресс
.
Физиол.
77
,
591
–
625
.