
Стрессоустойчивость коухена: Тест самооценки стрессоустойчивости (с.Коухена и г.Виллиансона)26
Тест самооценки стрессоустойчивости (с.Коухена и г.Виллиансона)26
Инструкция.Прочитайте вопрос и выберите наиболее подходящий ответ.
Насколько часто неожиданные неприятности выводят вас из равновесия?
Никогда — 0. Почти никогда — 1. Иногда — 2. Довольно часто — 3. Очень часто — 4.
Насколько часто вам кажется, что самые важные вещи в вашей жизни выходят из под вашего контроля?
Никогда — 0. Почти никогда — 1. Иногда — 2. Довольно часто — 3. Очень часто — 4.
Как часто вы чувствуете себя «нервозным», подавленным?
Никогда — 0. Почти никогда — 1. Иногда — 2. Довольно часто — 3. Очень часто — 4.
Как часто вы чувствуете уверенность в своей способности справиться со своими личными проблемами?
Никогда
— 4. Почти никогда — 3. Иногда — 2. Довольно
часто — 1. Очень часто — 0.
Почти никогда — 3. Иногда — 2. Довольно
часто — 1. Очень часто — 0.
Насколько часто вам кажется, что все идет именно так, как вы хотите? Никогда — 4. Почти никогда — 3. Иногда — 2. Довольно часто — 1. Очень часто — 0.
Как часто вы в силах контролировать раздражение?
Никогда — 4. Почти никогда — 3. Иногда — 2. Довольно часто — 1. Очень часто — 0.
Насколько часто у вас возникает чувство, что вам не справиться с тем, что от вас требуют?
Никогда — 0. Почти никогда — 1. Иногда — 2. Довольно часто — 3. Очень часто — 4.
Часто ли вы чувствуете, что вам сопутствует успех?
Никогда — 4. Почти никогда — 3. Иногда — 2. Довольно часто — 1. Очень часто — 0.
Как часто вы злитесь по поводу вещей, которые вы не можете контролировать?
Никогда
— 0. Почти никогда — 1. Иногда — 2. Довольно
часто — 3. Очень часто — 4.
Почти никогда — 1. Иногда — 2. Довольно
часто — 3. Очень часто — 4.
Часто ли вы думаете, что накопилось столько трудностей, что их невозможно преодолеть?
Никогда — 0. Почти никогда — 1. Иногда — 2. Довольно часто — 3. Очень часто — 4.
Интерпретация результатов теста.Обработка результатов осуществляется путем подсчета суммы баллов набранных испытуемым по всем вопросам теста. Стрессоустойчивость определяется по
таблице 3.7. приведенной ниже на основании количества баллов набранных испытуемым и его возраста.
Таблица 3.7. Оценка стрессоустойчивости
|
Оценка стрессоустойчивости |
Воз |
раст | ||
|
18-29 |
30-44 |
45-54 |
55-64 | |
|
Отлично |
0,5 |
2,0 |
1,8 |
1,3 |
|
Хорошо |
6,8 |
6,0 |
5,7 |
5,2 |
|
Удовлетворительно |
14,2 |
13,0 |
12,6 |
11,9 |
|
Плохо |
24,2 |
2,3 | 22,6 |
21,9 |
|
Очень плохо |
34,2 |
33,0 |
23,3 |
31,8 |
Ниже
предлагается тест, который позволяет
получить оценку вашего уровня
стрессоустойчивости. Вы получите тем
более объективный результат, чем более
искренними будут ваши ответы. Обведите
кружком подходящий вариант по каждому
утверждению (при отсутствии бланков на
листочках рядом с номером вопроса
ставиться балл).
Вы получите тем
более объективный результат, чем более
искренними будут ваши ответы. Обведите
кружком подходящий вариант по каждому
утверждению (при отсутствии бланков на
листочках рядом с номером вопроса
ставиться балл).
Таблица 3.8.
Тест на определение стрессоустойчивости личности
|
Утверждения |
редко |
иногда |
часто |
|
1. Я думаю, что меня недооценивают в коллективе |
1 |
2 |
3 |
|
2. |
1 |
2 |
3 |
|
3. Я переживаю за качество своей работы |
1 |
2 |
3 |
|
4. Я бываю, настроен агрессивно |
1 |
2 |
3 |
| 5. Я не терплю критики в свой адрес |
1 |
2 |
3 |
|
6. |
1 |
2 |
3 |
|
7. Я стараюсь быть лидером там, где это возможно |
1 |
2 |
3 |
|
8. Меня считают человеком настойчивым и напористым |
1 |
2 |
3 |
|
9. Я страдаю бессонницей |
1 |
2 |
3 |
|
10. |
1 |
2 | 3 |
|
11. Я эмоционально и болезненно переживаю неприятность |
1 |
2 |
3 |
|
12. У меня не хватает времени на отдых |
1 |
2 |
3 |
|
13. У меня возникают конфликтные ситуации |
1 |
2 | 3 |

 Я бываю раздражителен
Я бываю раздражителен Своим недругам я могу дать отпор
Своим недругам я могу дать отпор
28
Психология личности» Сост. Н.В.Киршева,
Н.В.Рябчикова. — М., Геликон, 1995
Н.В.Киршева,
Н.В.Рябчикова. — М., Геликон, 1995
68
Окончание табл. 3.8.
|
Утверждения |
редко |
иногда |
часто |
|
14. Мне недостает власти, чтобы реализовать себя |
1 |
2 |
3 |
|
15. У меня не хватает времени, чтобы заняться любимым делом |
1 |
2 |
3 |
|
16. |
1 |
2 |
3 |
|
17. Я испытываю страх, что не поступлю в институт |
1 |
2 |
3 |
|
18. Я действую сгоряча, а затем переживаю за свои дела и поступки. |
1 |
2 |
3 |
 Я все делаю быстро
Я все делаю быстроДалее подсчитайте суммарное число баллов, которое было набрано, и определите, каков уровень вашей стрессоустойчивости, по таблице:
Таблица
3. 9.
9.
Тест на определение стрессоустойчивости личности
|
Суммарное число баллов |
Уровень вашей стрессоустойчивост и |
|
51 -54 |
1 — очень низкий |
|
53-50 |
2 — низкий |
|
49-46 |
3 — ниже среднего |
|
45-42 |
4 — чуть ниже среднего |
|
41 -38 |
5 — средний |
|
37-34 |
6 — чуть выше среднего |
|
33-30 |
7 — выше среднего |
|
29-26 |
8 — высокий |
|
18-22 |
9 — очень высокий |
Чем
меньше (суммарное число) баллов вы
набрали, тем выше ваша стрессоустойчивость,
и наоборот. Если у вас 1-й и даже 2-й уровень
стрессоустойчивости, то вам необходимо
кардинально менять свой образ жизни.
Если у вас 1-й и даже 2-й уровень
стрессоустойчивости, то вам необходимо
кардинально менять свой образ жизни.
Тест самооценки стрессоустойчивости С. Коухена и Г. Виллиансона[45]. Психология стресса и методы коррекции
Тест самооценки стрессоустойчивости С. Коухена и Г. Виллиансона[45]. Психология стресса и методы коррекцииВикиЧтение
Психология стресса и методы коррекции
Щербатых Юрий Викторович
Содержание
Тест самооценки стрессоустойчивости С. Коухена и Г. Виллиансона[45]
1. Насколько часто неожиданные неприятности выводят вас из равновесия?
2. Насколько часто вам кажется, что самые важные вещи в вашей жизни выходят из-под вашего контроля?
3. Как часто вы чувствуете себя «нервозным», подавленным?
4. Как часто вы чувствуете уверенность в своей способности справиться со своими личными проблемами?
5. Насколько часто вам кажется, что все идет именно так, как вы хотите?
Насколько часто вам кажется, что все идет именно так, как вы хотите?
6. Как часто вы в силах контролировать раздражение?
7. Насколько часто у вас возникает чувство, что вам не справиться с тем, что от вас требуют?
8. Часто ли вы чувствуете, что вам сопутствует успех?
9. Как часто вы злитесь по поводу вещей, которые вы не можете контролировать?
10. Часто ли вы думаете, что накопилось столько трудностей, что их невозможно преодолеть?
Интерпретация результатов теста – средние данные по возрастным группам (цит. по Я. Воробейчик, 2004).
Данный текст является ознакомительным фрагментом.
Причуды самооценки
Причуды самооценки
Еще в начале прошлого века выдающийся американский психолог Уильям Джемс высказал суждение, что близкий круг общения в немалой мере формирует личность человека. Недавние психологические эксперименты подтвердили наблюдение Джемса и даже позволили
Недавние психологические эксперименты подтвердили наблюдение Джемса и даже позволили
Обновление самооценки
Обновление самооценки Быть самим собой иногда нелегко. Многие достигают зрелости с сомнениями в собственной адекватности. Даже если мы миновали подростковый возраст, не получив серьезных травм и ушибов, став молодыми взрослыми мы сталкиваемся с сильным давлением,
Уровень самооценки
Уровень самооценки Многие исследователи отмечают, что большое влияние на характер реакций, которые демонстрирует человек в условиях эмоционального стресса, оказывает структура самооценки личности. Считается, что люди с заведомо заниженной самооценкой обычно в большей
Тест № 9 Тест СПИЛБЕРГЕРА-ХАНИНА. ОЦЕНКА ЭМОЦИОНАЛЬНОГО СОСТОЯНИЯ (УРОВЕНЬ РЕАКТИВНОЙ И ЛИЧНОЙ ТРЕВОЖНОСТИ)
Тест № 9
Тест СПИЛБЕРГЕРА-ХАНИНА. ОЦЕНКА ЭМОЦИОНАЛЬНОГО СОСТОЯНИЯ (УРОВЕНЬ РЕАКТИВНОЙ И ЛИЧНОЙ ТРЕВОЖНОСТИ)
С помощью данного теста определяется уровень тревожности на момент его выполнения (РТ), отражающий реакцию на кратковременную сиюминутную ситуацию и уровень
ОЦЕНКА ЭМОЦИОНАЛЬНОГО СОСТОЯНИЯ (УРОВЕНЬ РЕАКТИВНОЙ И ЛИЧНОЙ ТРЕВОЖНОСТИ)
С помощью данного теста определяется уровень тревожности на момент его выполнения (РТ), отражающий реакцию на кратковременную сиюминутную ситуацию и уровень
Тест № 36 ОПРОСНИК ДЛЯ ОПРЕДЕЛЕНИЯ УРОВНЯ САМООЦЕНКИ
Тест № 36 ОПРОСНИК ДЛЯ ОПРЕДЕЛЕНИЯ УРОВНЯ САМООЦЕНКИ Для того чтобы определить уровень самооценки, необходимо сложить баллы по всем 32 суждениям:»очень часто» – 4 балла;»часто» – 3 балла;»иногда» – 2 балла;»редко» – 1 балл;»никогда» – 0 баллов.Сумма от 0 до 25 баллов говорит о
Тест ВМ (ориентировочный тест школьной зрелости — вербальное мышление)
Тест ВМ
(ориентировочный тест школьной зрелости — вербальное мышление)
И. ИразекКакое животное больше — лошадь или собака? Лошадь = 0, неправильный ответ = -5.Утром мы завтракаем, а в полдень.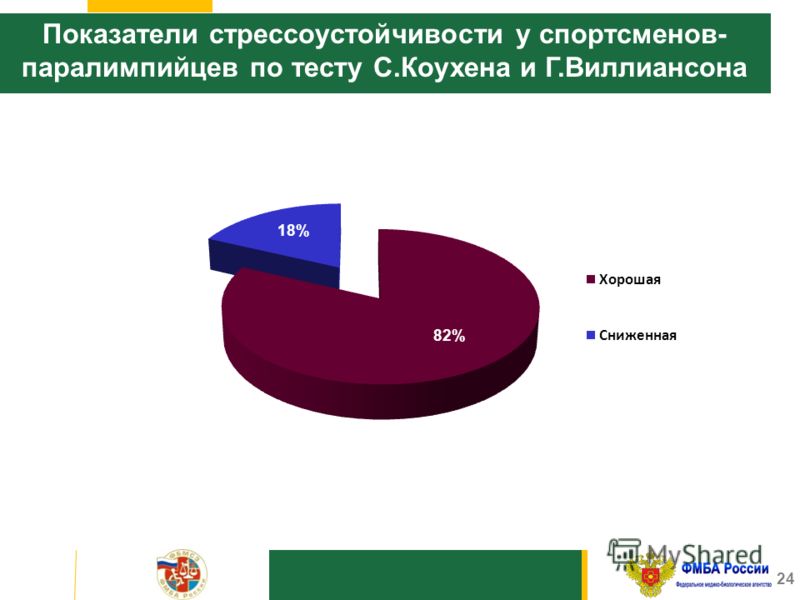 ..?Обедаем. Мы едим суп, макароны и мясо = 0.Полдничаем, ужинаем, спим и др. неправильные
..?Обедаем. Мы едим суп, макароны и мясо = 0.Полдничаем, ужинаем, спим и др. неправильные
Причуды самооценки
Причуды самооценки Еще в начале прошлого века выдающийся американский психолог Уильям Джемс высказал суждение, что близкий круг общения в немалой мере формирует личность человека. Недавние психологические эксперименты подтвердили наблюдение Джемса и даже позволили
Понятие самооценки
Понятие самооценки Иметь здоровую самооценку – то есть принимать самого себя – значит понимать свои чувства и реакции и действовать исходя из этого понимания (в своих интересах или в интересах других). Это относится ко всем сторонам жизни, а не только к ситуациям, когда
Структура самооценки
Структура самооценки
Самооценка имеет сложную структуру. Здоровая самооценка не означает, что надо все время быть сильным и чего-то добиваться. Она вмещает широкий эмоциональный спектр, самые разные реакции. Поэтому правильнее характеризовать самооценку через систему
Здоровая самооценка не означает, что надо все время быть сильным и чего-то добиваться. Она вмещает широкий эмоциональный спектр, самые разные реакции. Поэтому правильнее характеризовать самооценку через систему
Навыки стрессоустойчивости
Навыки стрессоустойчивости Управление собственными состояниями. Самовнушение. Медитация. Саморегуляция. Как использовать процедуры активизации терпимости к любым типам оппонентов с целью повышения внутренней уверенности и хладнокровия, – техники усмирения гордыни
Тест № 1 Уровень вашей самооценки
Тест № 1 Уровень вашей самооценки КлючI. У вас высокая самооценка. Вы явно не отягощены комплексом неполноценности, спокойно реагируете на критические замечания в свой адрес и редко сомневаетесь в своих действиях. Но не чрезмерно ли вы порой довольны собой? Быть
Тест №1.
 Уровень вашей самооценки
Уровень вашей самооценки
Тест №1. Уровень вашей самооценки
Система самооценки
Система самооценки Работа над характером напрямую связана с самооценкой. Вот мой способ. Я пишу шутку и тестирую ее со сцены на публике. Если публика громко смеется, я ставлю шутке оценку «1», смеется не очень сильно – ставлю «2», редко смеется – оценка «3».Когда я начал
Повышение самооценки
Повышение самооценки Прямо с настоящего момента начните хвалить себя за каждую мелочь и делайте это по возможности каждое утро.? Вовремя проснулись? Скажите себе, что вы молодец.? Приготовили завтрак? Похвалите себя за это, скажите, что вы все сделали великолепно.? Что-то
Теория привязанности: межличностные факторы стрессоустойчивости и защитные процессы
Теория привязанности: межличностные факторы стрессоустойчивости и защитные процессы Джон Боулби, английский психиатр и психоаналитик, сформулировал свою теорию привязанности в 1960-е годы (Bowlby, 1969, 1973) В отличие от традиционного психоаналитическго подхода,
Повышение устойчивости сельскохозяйственных культур к экологическим стрессам путем редактирования генома: опыт экстремофильных растений
Сохранить цитату в файл
Формат: Резюме (текст)PubMedPMIDAbstract (текст)CSV
Добавить в коллекции
- Создать новую коллекцию
- Добавить в существующую коллекцию
Назовите свою коллекцию:
Имя должно содержать менее 100 символов
Выберите коллекцию:
Не удалось загрузить вашу коллекцию из-за ошибки
Повторите попытку
Добавить в мою библиографию
- Моя библиография
Не удалось загрузить делегатов из-за ошибки
Повторите попытку
Ваш сохраненный поиск
Название сохраненного поиска:
Условия поиска:
Тестовые условия поиска
Электронная почта: (изменить)
Который день? Первое воскресеньеПервый понедельникПервый вторникПервая средаПервый четвергПервая пятницаПервая субботаПервый деньПервый рабочий день
Который день? ВоскресеньеПонедельникВторникСредаЧетвергПятницаСуббота
Формат отчета: SummarySummary (text)AbstractAbstract (text)PubMed
Отправить максимум:
1 шт. 5 шт. 10 шт. 20 шт. 50 шт. 100 шт. 200 шт.
5 шт. 10 шт. 20 шт. 50 шт. 100 шт. 200 шт.
Отправить, даже если нет новых результатов
Необязательный текст в электронном письме:
Создайте файл для внешнего программного обеспечения для управления цитированием
Полнотекстовые ссылки
Тейлор и Фрэнсис
Полнотекстовые ссылки
Обзор
. 2022 23 мая; 1-16.
дои: 10.1080/07388551.2022.2042481. Онлайн перед печатью.
Мохамед Куэн 1 2 , Педро Гарсия-Капаррос 3 , Ричард М. Твайман 4 , Чедли Абделли 1 , Хенда Махмуди 5 , Стефан Шилберг 6 , Ахмед Дебез 1
Принадлежности
- 1 Центр биотехнологии Borj-Cedria (CBBC), Лаборатория экстремофильных растений (LPE), Hammam-Lif, Тунис.
- 2 Факультет биологических наук и территории, Университет Молизе, Пеше, Италия.
- 3 Агрономический факультет высшей инженерной школы, Университет Альмерии, CIAIMBITAL, Альмерия, Испания.
- 4 Twyman Research Management Ltd, Скарборо, Великобритания.
- 5 Международный центр биозасоленного земледелия, Академический городок, рядом с Университетом Зайда, Дубай, Объединенные Арабские Эмираты.
- 6 Фраунгоферовский институт молекулярной биологии и прикладной экологии IME, Ахен, Германия.
- PMID: 35606905
-
DOI:
10.
 1080/07388551.2022.2042481
1080/07388551.2022.2042481
Обзор
Mohamed Kouhen et al. Критический обзор биотехнологий. .
. 2022 23 мая; 1-16.
дои: 10.1080/07388551.2022.2042481. Онлайн перед печатью.
Авторы
Мохамед Куэн 1 2 , Педро Гарсия-Капаррос 3 , Ричард М. Твайман 4 , Чедли Абделли 1 , Хенда Махмуди 5 , Стефан Шилберг 6 , Ахмед Дебез 1
Принадлежности
- 1 Центр биотехнологии Borj-Cedria (CBBC), Лаборатория экстремофильных растений (LPE), Hammam-Lif, Тунис.

- 2 Факультет биологических наук и территорий, Университет Молизе, Пеше, Италия.
- 3 Агрономический факультет высшей инженерной школы, Университет Альмерии, CIAIMBITAL, Альмерия, Испания.
- 4 Twyman Research Management Ltd, Скарборо, Великобритания.
- 5 Международный центр биозасоленного земледелия, Академический городок, рядом с Университетом Зайда, Дубай, Объединенные Арабские Эмираты.
- 6 Фраунгоферовский институт молекулярной биологии и прикладной экологии IME, Ахен, Германия.
- PMID: 35606905
-
DOI:
10.
 1080/07388551.2022.2042481
1080/07388551.2022.2042481
Абстрактный
В фундаментальных и прикладных науках редактирование генома стало незаменимым инструментом, особенно универсальная и адаптируемая система CRISPR/Cas9. Использование CRISPR/Cas9 в растениях позволило модифицировать многие ценные признаки, в том числе устойчивость к стрессам окружающей среды, что является важным аспектом, когда речь идет об обеспечении продовольственной безопасности в условиях изменения климата. Набор инструментов CRISPR обеспечивает более быструю и точную селекцию растений, облегчая: мультиплексное редактирование генов, построение генных пирамид и de novo одомашнивание. В этой статье мы обсуждаем самые последние достижения в области CRISPR/Cas9 и альтернативных систем на основе CRISPR, а также технические проблемы, которые еще предстоит решить. Пересмотр последних исследований, подтверждающих концепцию и функциональные характеристики, действительно позволил лучше понять количественные признаки, влияющие на урожайность и стрессоустойчивость. Кроме того, мы уделяем особое внимание применению технологии CRISPR/Cas9 в отношении экстремофильных растений в связи с их важностью на промышленном, экологическом и экономическом уровнях. Эти еще не изученные генетические ресурсы могут обеспечить средства для защиты наших культур от угрозы изменения климата, тем самым обеспечив продовольственную безопасность в следующем столетии.
Кроме того, мы уделяем особое внимание применению технологии CRISPR/Cas9 в отношении экстремофильных растений в связи с их важностью на промышленном, экологическом и экономическом уровнях. Эти еще не изученные генетические ресурсы могут обеспечить средства для защиты наших культур от угрозы изменения климата, тем самым обеспечив продовольственную безопасность в следующем столетии.
Ключевые слова: CRISPR/Cas9; Изменение климата; абиотический стресс; урожайность сельскохозяйственных культур; селекция растений.
Похожие статьи
-
Современные тенденции в редактировании генома растений: всеобъемлющий обзор набора инструментов CRISPR/Cas9.
Раззак А., Салим Ф., Канвал М., Мустафа Г., Юсаф С., Имран Аршад Х.М., Хамид М.К., Хан М.С., Джойя Ф.А. Раззак А. и др. Int J Mol Sci.
 201919 августа; 20 (16): 4045. дои: 10.3390/ijms20164045.
Int J Mol Sci. 2019.
PMID: 31430902
Бесплатная статья ЧВК.
Обзор.
201919 августа; 20 (16): 4045. дои: 10.3390/ijms20164045.
Int J Mol Sci. 2019.
PMID: 31430902
Бесплатная статья ЧВК.
Обзор. -
Технология CRISPR/Cas9 для улучшения агрономических характеристик и перспектив в сельском хозяйстве.
Рао М.Дж., Ван Л. Рао М.Дж. и соавт. Планта. 2021 8 сентября; 254(4):68. doi: 10.1007/s00425-021-03716-y. Планта. 2021. PMID: 34498163 Обзор.
-
Применение технологии редактирования генома CRISPR/Cas9 для улучшения сельскохозяйственных культур, выращиваемых в тропическом климате: последние достижения, перспективы и проблемы.
Хак Э., Танигути Х., Хассан М.М., Бховмик П., Карим М.Р., Смих М., Чжао К., Рахман М., Ислам Т. Хак Э. и др. Фронт завод науч. 2018 8 мая; 9:617.
 doi: 10.3389/fpls.2018.00617. Электронная коллекция 2018.
Фронт завод науч. 2018.
PMID: 29868073
Бесплатная статья ЧВК.
Обзор.
doi: 10.3389/fpls.2018.00617. Электронная коллекция 2018.
Фронт завод науч. 2018.
PMID: 29868073
Бесплатная статья ЧВК.
Обзор. -
Современное состояние технологии CRISPR и проектирования засухоустойчивых, засоленных и термоустойчивых сельскохозяйственных культур.
Ченнакесавулу К., Сингх Х., Триведи П.К., Джайн М., Ядав С.Р. Ченнакесавулу К. и др. Отчет о растительных клетках, март 2022 г.; 41 (3): 815-831. doi: 10.1007/s00299-021-02681-w. Epub 2021 19 марта. Представитель клеток растений 2022. PMID: 33742256 Обзор.
-
Критический обзор: последние достижения в использовании технологии CRISPR/Cas9 для повышения урожайности и смягчения последствий глобального продовольственного кризиса.
Рашид А.
 , Гилл Р.А., Хассан М.У., Махмуд А., Кари С., Заман К.У., Ильяс М., Аамер М., Батул М., Ли Х., Ву З.
Рашид А. и др.
Curr выпускает Mol Biol. 2021 11 ноября; 43 (3): 1950-1976. дои: 10.3390/cimb43030135.
Curr выпускает Mol Biol. 2021.
PMID: 34889892
Бесплатная статья ЧВК.
Обзор.
, Гилл Р.А., Хассан М.У., Махмуд А., Кари С., Заман К.У., Ильяс М., Аамер М., Батул М., Ли Х., Ву З.
Рашид А. и др.
Curr выпускает Mol Biol. 2021 11 ноября; 43 (3): 1950-1976. дои: 10.3390/cimb43030135.
Curr выпускает Mol Biol. 2021.
PMID: 34889892
Бесплатная статья ЧВК.
Обзор.
Посмотреть все похожие статьи
Цитируется
-
Обеспечение торговли продуктами с измененными генами в Азии и Австралазии: развивающийся нормативный ландшафт и перспективы на будущее.
Джонс МГК, Фосу-Ньярко Дж., Икбал С., Адил М., Ромеро-Альдемита Р., Аруджанан М., Касаи М., Вей Х., Прасетья Б., Нугрохо С., Мьюетт О., Мансур С., Аван М.Дж.А., Ордонио Р.Л., Рао С.Р. , Поддар А., Хандлби П., Ямсупасит Н., Ху К. Джонс МГК и др. Растения (Базель). 2022 сен 27;11(19)):2538. doi: 10.3390/plants11192538. Растения (Базель).
 2022.
PMID: 36235403
Бесплатная статья ЧВК.
Обзор.
2022.
PMID: 36235403
Бесплатная статья ЧВК.
Обзор.
Типы публикаций
Полнотекстовые ссылки
Тейлор и Фрэнсис
Укажите
Формат: ААД АПА МДА НЛМ
Отправить по номеру
Реакция растений на механический стресс
Механические раздражители вместе с соответствующими механизмами восприятия растений и тонко настроенной тигмоморфогенетической реакцией представляют научный и практический интерес с середины XVII века. В качестве новой области исследования механических напряжений возникает много проблем. Действительно, исследования различных видов растений (однолетних/многолетних) и органов растений (стебель/корень) с использованием различных подходов (полевой, лабораторный и in silico/вычислительный) не дали достаточных результатов, которые часто препятствуют практическому применению полученных знаний.
1. Введение
В ответ на различные механические стрессоры растения проявляют различные морфологические реакции, которым предшествует ряд биохимических изменений. Эти реакции зависят от типа механического стресса (MS), а также от стадии развития растения и биологических характеристик вида (например, см. [1] [2] ). Это обрисовывает общие тигмоморфогенетические характеристики стебля и корня; исследователи отмечают, что в меру современных знаний и, как показали другие авторы, например, [3] , универсальной реакции на РС у древесных и однолетних растений нет ( Рисунок 1 ).
Рисунок 1. Резюме реакции на механический стресс однолетних (арабидопсис; левая панель) и древесных растений (правая панель). ( A ) Молекулярные реакции, связанные с механическим стрессом, в стебле арабидопсиса. ( B ) Зоны дифференцированного роста в древесных стеблях растений, подвергающихся механическому воздействию (реакционная древесина, RW; нормальная древесина, СЗ; супротивная древесина, OW).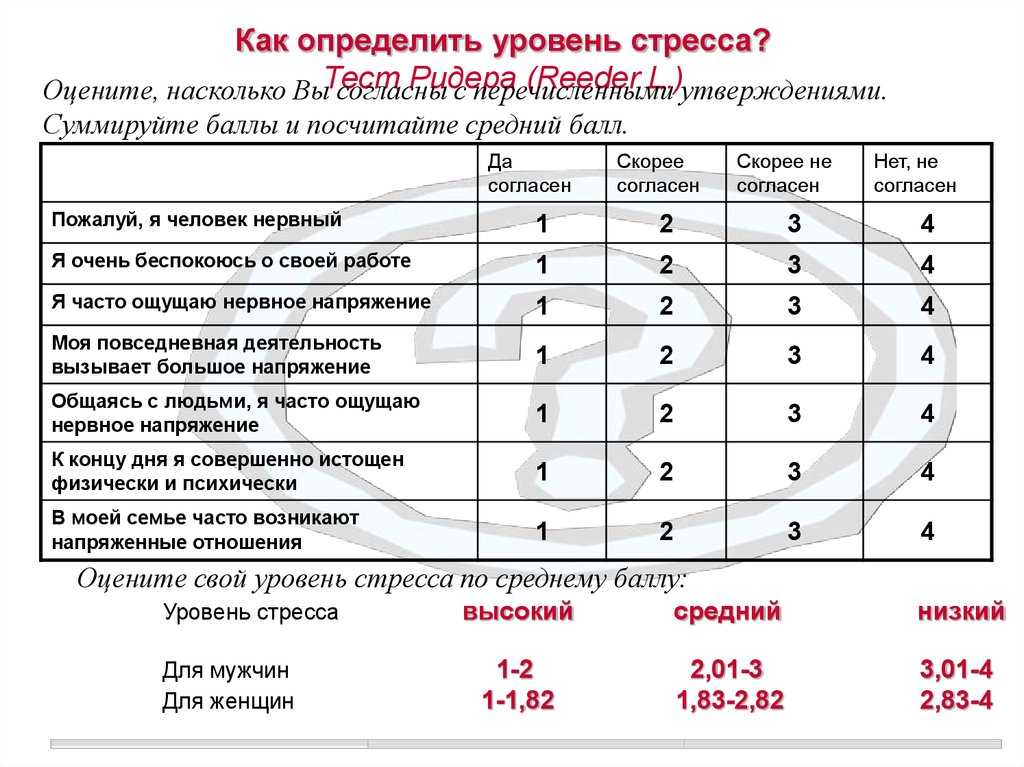 ( C ) Расположение и характеристики образования натяжной древесины (TW), т. е. RW в стебле вида покрытосеменных. ( D ) Расположение и характеристики образования прессованной древесины (CW), т.е. RW в стебле голосеменных видов. ( E ) Уменьшение роста в длину и увеличение радиальной толщины стебля древесных растений. ( F ) Увеличение зоны растяжения с радиально-симметричными изменениями расширения и удлинения клеток в корне арабидопсиса. ( G ) Специфическое для стимула быстрое и преходящее увеличение цитозольного кальция в корне арабидопсиса. ( H ) Апопластное подщелачивание, закисление цитоплазмы и производство апопластных активных форм кислорода (АФК). ( I ) Боковая инициация корня как реакция либо на гравитропное искривление, либо на ручное изгибание древесного корня. ( J ) Распределение боковых корней в форме «двустороннего веера» в условиях склона.
( C ) Расположение и характеристики образования натяжной древесины (TW), т. е. RW в стебле вида покрытосеменных. ( D ) Расположение и характеристики образования прессованной древесины (CW), т.е. RW в стебле голосеменных видов. ( E ) Уменьшение роста в длину и увеличение радиальной толщины стебля древесных растений. ( F ) Увеличение зоны растяжения с радиально-симметричными изменениями расширения и удлинения клеток в корне арабидопсиса. ( G ) Специфическое для стимула быстрое и преходящее увеличение цитозольного кальция в корне арабидопсиса. ( H ) Апопластное подщелачивание, закисление цитоплазмы и производство апопластных активных форм кислорода (АФК). ( I ) Боковая инициация корня как реакция либо на гравитропное искривление, либо на ручное изгибание древесного корня. ( J ) Распределение боковых корней в форме «двустороннего веера» в условиях склона. ( K ) Асимметричный отклик трех участков изгиба корней на вогнутой и выпуклой стороне изогнутых древесных корней.
( K ) Асимметричный отклик трех участков изгиба корней на вогнутой и выпуклой стороне изогнутых древесных корней.
2. Тигмоморфогенез в стебле
Согласно обзору Braam [4] , тигмоморфогенез по-разному воздействует на виды растений, и сенсорные стимулы могут вызывать различные реакции в надземных органах растений, т. е. в листьях плотоядных растений, видоизмененных листьях/ стебли у вьющихся растений, цветы у некоторых видов, у которых возможно самоопыление, и т. д. Наиболее характерной иллюстрацией этого является тигмоморфогенетическая реакция Mimosa pudica, также называемая «не прикасайся ко мне». Опосредуя моторный орган, названный pulvinus, листочки растения быстро складываются в ответ на экзогенный MS, используя быстрый электрический сигнал на большие расстояния [5] и кальциевые флюсы [6] .
2.1. Реакция однолетних растений на механический стресс: модель Arabidopsis
В ответ на механические раздражители, такие как ветер или прикосновение, стебли претерпевают физиологические изменения и изменения в развитии, которые повышают устойчивость к последующему MS ( Рисунок 1 A). Как правило, растения, растущие в ветреную погоду, короче, коренастее и часто имеют измененную гибкость [7] [8] .
Как правило, растения, растущие в ветреную погоду, короче, коренастее и часто имеют измененную гибкость [7] [8] .
У арабидопсиса, как розеточного растения, онтогенетическая функция стебля отличается от многолетнего аналога, в котором он способствует долговременной стабильности, структурной и механической приспособленности, и где вышеупомянутые функции не являются приоритетными [9] [10] [11] . Однако биологические особенности арабидопсиса, т. е. возможность индуцирования стебля во вторичную структуру путем декапитации, уменьшения освещенности («условия короткого дня») или увеличения весовой нагрузки стебля [4] [12] , наряду с наличием знаний/данных о широком спектре физиомолекулярных процессов у видов, были использованы с точки зрения изучения механизмов стволовой реакции на РС. Было показано, что стимуляция ветром пропорционально влияет на степень ветвления и базальную продуктивность растений арабидопсиса 9.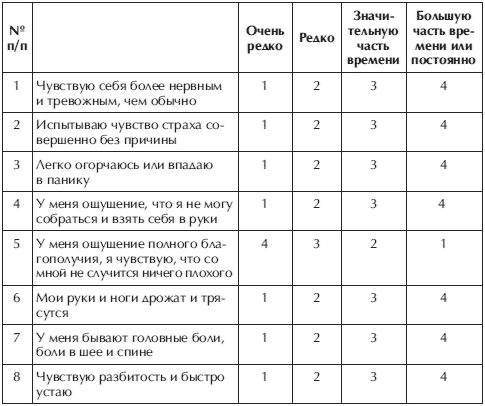 0069 [13] .
0069 [13] .
Случайное открытие набора генов Arabidopsis touch (TCH) вызвало интерес к тигмоморфогенетическим молекулярным механизмам [3] [14] . Роль, которую играет семейство генов TCH, не ограничивается рассеянным склерозом, поскольку они были связаны с активацией посредством экзогенного ауксина и/или брассиностероида и колебаниями свободного цитозольного иона кальция (Ca 2+ ) в качестве вторичного мессенджера в различных сигналах. пути трансдукции [14] , открытие новых исследовательских каналов. Обычно наблюдаемое увеличение толщины стебля в ответ на MS не всегда происходит у арабидопсиса, но за счет приложения веса к стеблю тип силы сжатия может вызвать образование камбиоподобных тканей [3] . Было обнаружено, что ауксин поддерживает образование вторичной ксилемы, и предполагается, что три гена фактора ответа на ауксин (ARF) (ARF2, ARF4 и ARF12) играют особенно важную роль во время образования древесины [15] . Из ранее упомянутого семейства генов R2R3-MYB четыре фактора транскрипции MYB рассматриваются как кандидаты в регуляторные гены для формирования древесины, и три из них (AtMYB77, AtMYB73, AtMYB44), по-видимому, имеют сходные функции в развитии стебля [15] . Некоторые из генов TCH были дополнительно связаны с передачей сигналов jasmonate [16] . Механостимуляция включает сигнальные пути жасмоновой кислоты (ЖАК) как часть регуляции камбия, которая индуцирует продукцию ЖАК и экспрессию генов биосинтеза ЖАК [17] и необходим для регуляции роста раны [18] . Было обнаружено, что катанин-зависимая динамика микротрубочек повышает способность клеток реагировать на РС за счет повышения способности клеток адаптироваться к их росту в соответствии с соседями [19] . Подтверждена также роль этилена, ауксина, цитокининов и гиббереллинов в развитии сосудов арабидопсиса [20] .
Из ранее упомянутого семейства генов R2R3-MYB четыре фактора транскрипции MYB рассматриваются как кандидаты в регуляторные гены для формирования древесины, и три из них (AtMYB77, AtMYB73, AtMYB44), по-видимому, имеют сходные функции в развитии стебля [15] . Некоторые из генов TCH были дополнительно связаны с передачей сигналов jasmonate [16] . Механостимуляция включает сигнальные пути жасмоновой кислоты (ЖАК) как часть регуляции камбия, которая индуцирует продукцию ЖАК и экспрессию генов биосинтеза ЖАК [17] и необходим для регуляции роста раны [18] . Было обнаружено, что катанин-зависимая динамика микротрубочек повышает способность клеток реагировать на РС за счет повышения способности клеток адаптироваться к их росту в соответствии с соседями [19] . Подтверждена также роль этилена, ауксина, цитокининов и гиббереллинов в развитии сосудов арабидопсиса [20] .
2.2. Реакция стебля древесных растений — роль реактивной древесины
Многолетнее растение связано с широким спектром морфологических и физиологических признаков, которые, вероятно, обусловлены более широким диапазоном экологических и сезонных сигналов, с которыми сталкиваются эти растения по сравнению с их однолетними аналогами [21] . В стебле многолетних древесных растений механическое напряжение вызывало уменьшение прироста удлинением при увеличении радиальной толщины, т.е. уменьшение высоты и увеличение диаметра соответственно [4] [22] , оказывая видимое и непосредственное влияние на урожайность/производство биомассы ( Рисунок 1 B). Целью тигмоморфогенетической реакции древесных растений является переориентация невертикальной оси, которая достигается за счет образования RW в точках, где сила (сжатие или растяжение) может вытолкнуть стебель в исходное положение [23] . Эта реакция возникает из-за неоднородности активности камбиальной области и в основном затрагивает древесину, также называемую вторичной ксилемой, которая обеспечивает механическую поддержку и дальнюю проводимость воды и питательных веществ [24] [25] [26] . Древесина естественным образом состоит из микрофибрилл целлюлозы, расположенных в матрице гемицеллюлозы и лигнина, которые под нагрузкой и с течением времени проявляют анатомические и химические деформации [27] .
В стебле многолетних древесных растений механическое напряжение вызывало уменьшение прироста удлинением при увеличении радиальной толщины, т.е. уменьшение высоты и увеличение диаметра соответственно [4] [22] , оказывая видимое и непосредственное влияние на урожайность/производство биомассы ( Рисунок 1 B). Целью тигмоморфогенетической реакции древесных растений является переориентация невертикальной оси, которая достигается за счет образования RW в точках, где сила (сжатие или растяжение) может вытолкнуть стебель в исходное положение [23] . Эта реакция возникает из-за неоднородности активности камбиальной области и в основном затрагивает древесину, также называемую вторичной ксилемой, которая обеспечивает механическую поддержку и дальнюю проводимость воды и питательных веществ [24] [25] [26] . Древесина естественным образом состоит из микрофибрилл целлюлозы, расположенных в матрице гемицеллюлозы и лигнина, которые под нагрузкой и с течением времени проявляют анатомические и химические деформации [27] . Эти изменения могут снизить ценность древесины с экономической точки зрения, что побудило значительный объем литературы рассмотреть реакцию ствола на MS [28] . MS также считается необходимым для дифференцировки клеток ксилемы, но точные механизмы его воздействия не ясны [26] [29] . Как только ствол сгибается, асимметричная реакция проявляется в виде образования RW и противоположной древесины (OW) (вставка 1; 90-193, рис. 1, 90-117 C). Расположение и характеристики RW различаются у голосеменных и покрытосеменных, соответственно, и сжатой древесины (CW) и растянутой древесины (TW) (вставка 1), и они дополнительно влияют на гидравлические и механические свойства древесины за счет изменений свойств древесины [ 26] [30] . Различия как в CW, так и в TW обусловлены видовыми характеристиками и возрастом, условиями окружающей среды, типом стресса и степенью сжатия [23] [24] [25] [26] [27] [28] .
Эти изменения могут снизить ценность древесины с экономической точки зрения, что побудило значительный объем литературы рассмотреть реакцию ствола на MS [28] . MS также считается необходимым для дифференцировки клеток ксилемы, но точные механизмы его воздействия не ясны [26] [29] . Как только ствол сгибается, асимметричная реакция проявляется в виде образования RW и противоположной древесины (OW) (вставка 1; 90-193, рис. 1, 90-117 C). Расположение и характеристики RW различаются у голосеменных и покрытосеменных, соответственно, и сжатой древесины (CW) и растянутой древесины (TW) (вставка 1), и они дополнительно влияют на гидравлические и механические свойства древесины за счет изменений свойств древесины [ 26] [30] . Различия как в CW, так и в TW обусловлены видовыми характеристиками и возрастом, условиями окружающей среды, типом стресса и степенью сжатия [23] [24] [25] [26] [27] [28] . Тем не менее, оба используют сходные базовые механизмы для восприятия стрессового стимула и тигмоморфогенетической реакции, которые различаются структурным и механическим контекстом [23] и будут кратко изложены в следующих параграфах ( Рисунок 1 D, E).
Тем не менее, оба используют сходные базовые механизмы для восприятия стрессового стимула и тигмоморфогенетической реакции, которые различаются структурным и механическим контекстом [23] и будут кратко изложены в следующих параграфах ( Рисунок 1 D, E).
КС формируется на нижней (вогнутой) сжатой стороне изогнутого стебля/ветви у голосеменных многолетников ( Рисунок 1 D), и его основная функция состоит в том, чтобы вернуть отклоняющийся шток в вертикальное положение за счет напряжения сжатия [31] . Изменения, связанные с CW, являются одной из основных причин снижения качества древесины и изделий из волокна [32] . Сжатие вызывает продольную усадку по сравнению с обычной древесиной (NW), что связано с большим углом микрофибрилл и повышенным одревеснением, которые использовались для исследования механического поведения древесины и способствуют снижению жесткости CW 9.0069 [27] [28] [33] . Другой анатомической особенностью КС являются укороченные трахеиды по сравнению с СЗ и ОВ того же дерева, с изменением формы и деформацией кончиков [25] [28] . В дополнение к более высокому содержанию лигнина КС также содержит более высокие количества (1-4)-β-галактана и более низкие количества целлюлозы, маннана и ксилана [28] .
Другой анатомической особенностью КС являются укороченные трахеиды по сравнению с СЗ и ОВ того же дерева, с изменением формы и деформацией кончиков [25] [28] . В дополнение к более высокому содержанию лигнина КС также содержит более высокие количества (1-4)-β-галактана и более низкие количества целлюлозы, маннана и ксилана [28] .
Вставка 1. Глоссарий терминов, относящихся к реакции древесной структуры на механическое воздействие.
Реакция древесины (RW) — естественная реакция древесных растений на механическое воздействие посредством асимметричного образования вторичной ксилемной ткани, направленная на усиление структуры и перенаправление роста [30] [34] ;
Нормальная древесина (СЗ) — древесина, образовавшаяся в отсутствие раздражителя [30] ;
Изгибаемая древесина (FW) — специфическая древесина, образованная сосудистым камбием деревьев, растущих в ветреную погоду, характеризующаяся повышенным образованием вторичной ксилемы и пониженным модулем упругости по сравнению с нормальной древесиной [30] ;
Сжатая древесина (CW) — RW у голосеменных растений, образующаяся на нижней стороне наклоненных стеблей или ветвей, характеризующаяся высоким содержанием лигнина и низким содержанием целлюлозы из-за создания сжимающей силы, толкающей ствол вверх [30] [34] . Ткань с такими же характеристиками отмечена на нижней (вогнутой) стороне корня тополя изогнутого [30] [35] ;
Ткань с такими же характеристиками отмечена на нижней (вогнутой) стороне корня тополя изогнутого [30] [35] ;
Натяжная древесина (TW) — RW у двудольных покрытосеменных, образующаяся на верхней стороне наклоняющихся стеблей или ветвей в процессе переориентации, характеризующаяся низким содержанием лигнина и высоким содержанием целлюлозы [30] [34] ;
Противоположная древесина (OW) — часть асимметричного отклика на механическое воздействие, расположенная напротив RW, характеризующаяся промежуточными свойствами между NW и RW [30] [35] ;
TW образуется на верхней (выпуклой) растянутой стороне изогнутого стебля/ветви у покрытосеменных многолетников ( Рисунок 1 E). Подобно CW, у разных видов TW демонстрирует широкий спектр организационных вариаций (см. подробный обзор в 9).0069 [36] ), но его основной анатомической характеристикой являются G-волокна, представляющие собой волокна ксилемы с меньшим радиальным диаметром, имеющие дополнительный толстый слой на внутренней стороне вторичной стенки и образующие дополнительный слой с полупрозрачной желеобразный внешний вид, который не очень связан с остальными слоями клеточной стенки, названный G-слоем [24] [25] . G-слой в основном полностью состоит из целлюлозы, но подтверждено присутствие лигнина, ксилоглюканов и белков, синтезирующих ксилоглюканы, пектинов, белков рамногалактуронана I, арабиногалактана и арабиногалактана [23] [25] . Однако, поскольку обнаруженное содержание лигнина было минимальным или отсутствовало, обычно считается, что TW имеет повышенное содержание целлюлозы и пониженное содержание лигнина [23] [24] как другую сторону ранее упомянутой отрицательной корреляции. Слой G также характеризуется более высокой пористостью, допускающей более высокое содержание воды, что, как предполагается, является причиной желеобразного вида и способности слоя G к поперечному набуханию/усадке [23] . Образование TW является результатом повышенной скорости деления клеток, т.е. камбиальной активности [25] .
G-слой в основном полностью состоит из целлюлозы, но подтверждено присутствие лигнина, ксилоглюканов и белков, синтезирующих ксилоглюканы, пектинов, белков рамногалактуронана I, арабиногалактана и арабиногалактана [23] [25] . Однако, поскольку обнаруженное содержание лигнина было минимальным или отсутствовало, обычно считается, что TW имеет повышенное содержание целлюлозы и пониженное содержание лигнина [23] [24] как другую сторону ранее упомянутой отрицательной корреляции. Слой G также характеризуется более высокой пористостью, допускающей более высокое содержание воды, что, как предполагается, является причиной желеобразного вида и способности слоя G к поперечному набуханию/усадке [23] . Образование TW является результатом повышенной скорости деления клеток, т.е. камбиальной активности [25] .
Роль фитогормонов в тигмоморфогенетической реакции и их участие в формировании КС и ТВ были признаны и изучены, но результаты и выводы не согласованы и их трудно сравнивать (подробный обзор см. [28] для CW и [23] для TW). Это связано не только с общим участием фитогормонов во многих аспектах развития растений, но и с комбинированным действием гормонов друг с другом и с другими звеньями механизма регуляции стресса. Подводя краткий итог, можно сказать, что в формировании КС в основном участвуют ауксин и этилен, а также редуцированные эндогенные цитокинины и абсцизовая кислота 9.0069 [28] [37] . В формировании TW, хотя роль этилена постоянно подтверждается, роль ауксина еще четко не определена, но, по-видимому, играет решающую роль в формировании RW [23] [24] . Совсем недавно цитокинины и брассиностероиды также были связаны с образованием TW посредством [30] и [34] соответственно. Роль гиббереллина в формировании CW была отвергнута, но были некоторые доказательства его роли в формировании TW, где было показано, что он способен индуцировать камбиальный рост и дифференцировку G-волокон [23] [34] [38] .
[28] для CW и [23] для TW). Это связано не только с общим участием фитогормонов во многих аспектах развития растений, но и с комбинированным действием гормонов друг с другом и с другими звеньями механизма регуляции стресса. Подводя краткий итог, можно сказать, что в формировании КС в основном участвуют ауксин и этилен, а также редуцированные эндогенные цитокинины и абсцизовая кислота 9.0069 [28] [37] . В формировании TW, хотя роль этилена постоянно подтверждается, роль ауксина еще четко не определена, но, по-видимому, играет решающую роль в формировании RW [23] [24] . Совсем недавно цитокинины и брассиностероиды также были связаны с образованием TW посредством [30] и [34] соответственно. Роль гиббереллина в формировании CW была отвергнута, но были некоторые доказательства его роли в формировании TW, где было показано, что он способен индуцировать камбиальный рост и дифференцировку G-волокон [23] [34] [38] .
Предыдущие исследования в основном были сосредоточены на анатомических и морфологических характеристиках RW. Такого же внимания не уделялось молекулярным механизмам вторичного роста [12] , и текущий интерес действительно сосредоточен на молекулярных и сигнальных аспектах образования RW. Однако единого мнения о том, является ли сравнение модельных видов арабидопсисом и многолетними растениями, а также различными многолетними видами, не достигнуто. В то время как ключевые регуляторы наблюдались, когда дело доходит до вторичного развития как травянистых, так и древесных растений [20] , проблема остается в том, что образование RW не происходит естественным образом у травянистых модельных видов, таких как Arabidopsis [39] . Использование тополя в качестве модельного вида действительно помогает преодолеть некоторые трудности, но дефицит молекулярных исследований многолетних видов и тем более голосеменных остается [39] [40] . В CW анализ экспрессии генов выявил повышенную регуляцию генов, участвующих в гравитропной реакции стебля, то есть биосинтезе лигнина, этиленобразующих ферментах и белках клеточной стенки (биосинтетические ферменты, углеводный обмен и регуляторные белки, биосинтез монолигнола, арабиногалактана и пролина). богатые белками) [28] [32] . Особый интерес представляет семейство R2R3-MYB, которое регулирует метаболизм лигнина и фенилпропаноида во время образования древесины и участие которого было подтверждено у хвойных [40] . Пилат и др. [39] представил всесторонний обзор геномных исследований TW, указывающий на потенциал TW для лучшего понимания молекулярных механизмов образования древесины и их свойств. Совсем недавно исследования, посвященные ранним изменениям в транскриптоме тополя, способствовали лучшему пониманию тигмоморфогенеза. Помьес и др. [41] исследовали ответ на однократное или многократное сгибание и пришли к выводу, что, хотя основные изменения экспрессии генов происходят в первые два часа после сгибания, существует несколько механистических путей, участвующих в ответе, начиная с генов, участвующих в общий ответ на абиотический стресс (АФК, Ca 2+ и передача сигналов жасмоновой кислоты) на более специфические гены, участвующие в развитии клеточной стенки и древесины.
В CW анализ экспрессии генов выявил повышенную регуляцию генов, участвующих в гравитропной реакции стебля, то есть биосинтезе лигнина, этиленобразующих ферментах и белках клеточной стенки (биосинтетические ферменты, углеводный обмен и регуляторные белки, биосинтез монолигнола, арабиногалактана и пролина). богатые белками) [28] [32] . Особый интерес представляет семейство R2R3-MYB, которое регулирует метаболизм лигнина и фенилпропаноида во время образования древесины и участие которого было подтверждено у хвойных [40] . Пилат и др. [39] представил всесторонний обзор геномных исследований TW, указывающий на потенциал TW для лучшего понимания молекулярных механизмов образования древесины и их свойств. Совсем недавно исследования, посвященные ранним изменениям в транскриптоме тополя, способствовали лучшему пониманию тигмоморфогенеза. Помьес и др. [41] исследовали ответ на однократное или многократное сгибание и пришли к выводу, что, хотя основные изменения экспрессии генов происходят в первые два часа после сгибания, существует несколько механистических путей, участвующих в ответе, начиная с генов, участвующих в общий ответ на абиотический стресс (АФК, Ca 2+ и передача сигналов жасмоновой кислоты) на более специфические гены, участвующие в развитии клеточной стенки и древесины.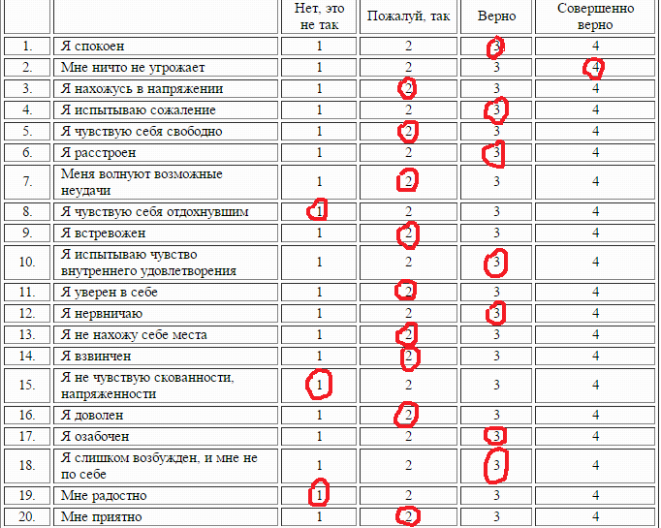 Используя инновационное изотропное устройство, Lopez et al. [34] удалось выделить ранний (30 минут) молекулярный ответ на гравистимуляцию, снова подтвердив активность клеточной стенки и образования древесины и отметив около 200 генов, регулируемых ксилемой, которые еще не были функционально охарактеризованы.
Используя инновационное изотропное устройство, Lopez et al. [34] удалось выделить ранний (30 минут) молекулярный ответ на гравистимуляцию, снова подтвердив активность клеточной стенки и образования древесины и отметив около 200 генов, регулируемых ксилемой, которые еще не были функционально охарактеризованы.
3. Тигмоморфогенез в корнях
Препятствия, с которыми сталкиваются корни при проникновении в почву, неизменно вызывают тигмоморфогенез [16] . Исследования корневой экологии сталкиваются с многочисленными проблемами, начиная от биологических характеристик органа и заканчивая дизайном соответствующих экспериментальных исследований [42] . Эти трудности еще более усугубляются в случае тигмоморфогенеза корней из-за ранее упомянутых факторов вариабельности MS в плане эксперимента и трудностей с отбором проб.
3.1. Реакция молодых корней на механический стресс
Воздействие механических раздражителей на первичные корни вызывает изменения направления роста, которые могут изменить расположение боковых корней (БК), при этом БК исходят с выпуклой стороны возникающих кривых, а не находятся в предустановленное распределение [43] [44] [45] [46] [47] . Каскад событий, ведущих к образованию LR из клеток перицикла ксилемы, хорошо изучен у Arabidopsis, что, как показано, строго связано с преходящим пространственно-временным накоплением ауксинов вдоль оси родительского корня [48] [49 ] . В частности, Ditengou et al. [43] наблюдали делокализацию переносчика ауксина PIN1 в одиночной протоксилемной клетке с последующим накоплением ауксина в месте индукции боковых корешков. Другое исследование подтвердило излучение LR на выпуклой стороне изгиба в момент времени всего 20 с переходного изгиба 9.0069 [50] . Эмиссия LR также была связана с передачей сигналов кальция, которая транслирует механические силы в реакцию развития в корнях [50] . Поскольку растения обнаруживают механические раздражители для выявления соседних барьеров и изменяют модели своего роста, чтобы приспособиться к окружающей среде, другим широко используемым подходом в исследованиях рассеянного склероза является воздействие на корневой барьер.
Каскад событий, ведущих к образованию LR из клеток перицикла ксилемы, хорошо изучен у Arabidopsis, что, как показано, строго связано с преходящим пространственно-временным накоплением ауксинов вдоль оси родительского корня [48] [49 ] . В частности, Ditengou et al. [43] наблюдали делокализацию переносчика ауксина PIN1 в одиночной протоксилемной клетке с последующим накоплением ауксина в месте индукции боковых корешков. Другое исследование подтвердило излучение LR на выпуклой стороне изгиба в момент времени всего 20 с переходного изгиба 9.0069 [50] . Эмиссия LR также была связана с передачей сигналов кальция, которая транслирует механические силы в реакцию развития в корнях [50] . Поскольку растения обнаруживают механические раздражители для выявления соседних барьеров и изменяют модели своего роста, чтобы приспособиться к окружающей среде, другим широко используемым подходом в исследованиях рассеянного склероза является воздействие на корневой барьер. Внешние и эндогенно генерируемые механические силы постоянно вызывают стимул-специфическое, быстрое и преходящее увеличение цитозольного Ca 2+ [51] . Было показано, что барьерное воздействие вызывает апопластное подщелачивание, подкисление цитоплазмы и выработку апопластных активных форм кислорода (АФК) [52] . Якобсен и др. [53] использовали анализ барьерной системы in vitro для изучения реакции корня арабидопсиса на кратковременный механический импеданс (до 30 часов) посредством глобального профилирования транскрипции. Результаты выявили радиально-асимметричные изменения в расширении и удлинении клеток и уменьшении длины корня в дополнение к более короткому расстоянию от кончика корня до выхода корневого волоска (9).0193 Рисунок 1 F). ROS, сигнальные гены, связанные с дифференциальными градиентами этилена и ауксина, и активация транскрипции ROS были частью ранней реакции корней арабидопсиса [53] . Недавно была установлена роль ауксина в тигмотропизме во время взаимодействий растение-препятствие, где сообщалось, что PIN-FORMED (PIN)-опосредованный транспорт полярного ауксина делает возможным изгибание корня перед избеганием препятствия [54] .
Внешние и эндогенно генерируемые механические силы постоянно вызывают стимул-специфическое, быстрое и преходящее увеличение цитозольного Ca 2+ [51] . Было показано, что барьерное воздействие вызывает апопластное подщелачивание, подкисление цитоплазмы и выработку апопластных активных форм кислорода (АФК) [52] . Якобсен и др. [53] использовали анализ барьерной системы in vitro для изучения реакции корня арабидопсиса на кратковременный механический импеданс (до 30 часов) посредством глобального профилирования транскрипции. Результаты выявили радиально-асимметричные изменения в расширении и удлинении клеток и уменьшении длины корня в дополнение к более короткому расстоянию от кончика корня до выхода корневого волоска (9).0193 Рисунок 1 F). ROS, сигнальные гены, связанные с дифференциальными градиентами этилена и ауксина, и активация транскрипции ROS были частью ранней реакции корней арабидопсиса [53] . Недавно была установлена роль ауксина в тигмотропизме во время взаимодействий растение-препятствие, где сообщалось, что PIN-FORMED (PIN)-опосредованный транспорт полярного ауксина делает возможным изгибание корня перед избеганием препятствия [54] . Некоторые авторы предполагают, что у арабидопсиса сила натяжения, действующая на выпуклой стороне изогнутого корня, вызывает увеличение содержания Са 2+ уровней в специфических клетках перицикла, становясь «клеткой-основателем» нового бокового корня [50] [55] ( Рисунок 1 G). Это увеличение Ca 2+ приводит к: (а) изменению АФК и закислению цитозоля, которые, как известно, вызывают сигнальные события; и (b) подщелачивание клеточной стенки, которое, как известно, укрепляет матрикс клеточной стенки (, рис. 1, E). Diaz-Sala [56] предположил, что механочувствительные ионные каналы, присутствующие на плазматических мембранах, могут генерировать электрические потенциалы действия, которые распространяются на короткие расстояния от клетки к клетке по сети плазматической мембраны и через плазмодесмы (или, альтернативно, через клетки флоэмы). на более длинном расстоянии), вызывая модификации клеточных стенок, создавая специфические взаимодействия между клеточной стенкой и цитоскелетом и изменяя динамику микротрубочек.
Некоторые авторы предполагают, что у арабидопсиса сила натяжения, действующая на выпуклой стороне изогнутого корня, вызывает увеличение содержания Са 2+ уровней в специфических клетках перицикла, становясь «клеткой-основателем» нового бокового корня [50] [55] ( Рисунок 1 G). Это увеличение Ca 2+ приводит к: (а) изменению АФК и закислению цитозоля, которые, как известно, вызывают сигнальные события; и (b) подщелачивание клеточной стенки, которое, как известно, укрепляет матрикс клеточной стенки (, рис. 1, E). Diaz-Sala [56] предположил, что механочувствительные ионные каналы, присутствующие на плазматических мембранах, могут генерировать электрические потенциалы действия, которые распространяются на короткие расстояния от клетки к клетке по сети плазматической мембраны и через плазмодесмы (или, альтернативно, через клетки флоэмы). на более длинном расстоянии), вызывая модификации клеточных стенок, создавая специфические взаимодействия между клеточной стенкой и цитоскелетом и изменяя динамику микротрубочек.
3.2. Реакция древесных корней
На древесных растениях имеется очень мало исследований, касающихся анатомических, морфологических, биохимических и молекулярных аспектов реакции древесных корней на наклон или напряжение изгиба [30] [35] [57 ] [58] ( Рисунок 1 I). древесный рост корней в условиях естественного склона привел к асимметричной корневой системе, обозначенной как «двусторонне-веерная форма», в которой боковые корни развивались как вниз, так и вверх по склону [59] [60] [61] ( Рисунок 1 J). Однако механические ограничения не вызывают одинаковой реакции стебля и корня. Кроме того, корни, подвергающиеся одинаковым механическим ограничениям, которые накладываются на стебель, могут развивать совершенно разные RW. Действительно, у растений тополя при изгибании на «верхней» выпуклой растянутой стороне ствола или ветки образуется ДВ, а на «нижней», вогнутой, сжатой стороне после изгиба формируется КС, как у голосеменных стеблей. Хеллгрен и др. [62] установили, что образование ДВ у тополя не опосредовано изменением уровня индол-3-уксусной кислоты (ИУК) в тканях камбия. Напротив, более высокое количество эндогенной ИУК было обнаружено на стороне камбиальной области, формирующей КС, которая могла действовать как пространственный регулятор камбиальной активности, увеличивая скорость клеточного деления и сообщая ключевую позиционную информацию клеткам камбиальной области. окружающие зону ткани для дифференцировки/инициации RW [35] [63] [64] .
Хеллгрен и др. [62] установили, что образование ДВ у тополя не опосредовано изменением уровня индол-3-уксусной кислоты (ИУК) в тканях камбия. Напротив, более высокое количество эндогенной ИУК было обнаружено на стороне камбиальной области, формирующей КС, которая могла действовать как пространственный регулятор камбиальной активности, увеличивая скорость клеточного деления и сообщая ключевую позиционную информацию клеткам камбиальной области. окружающие зону ткани для дифференцировки/инициации RW [35] [63] [64] .
Было показано, что реакция MS древесных корней временно и пространственно модулируется сложным взаимодействием различных путей передачи сигнала, включая активные формы кислорода (АФК), гормоны (индолуксусная кислота, гиббереллины, АБК и этилен) и специфические молекулярные факторы, регулирующие отложение лигнина, целостность клеточных стенок и формирование боковых корней Трупиано и др. [58] постулирует модель распределения механической силы, в которой выпуклая и вогнутая стороны каждого изогнутого корневого сектора подвергаются различному распределению механической силы, при этом силы растяжения приложены к выпуклой стороне, а силы сжатия сосредоточены на вогнутой стороне. Боковые и секторальные стратегии использовались изогнутыми корнями для поддержания поглощения и транспортировки воды в деформирующем состоянии, вызванном силами растяжения и сжатия; это привело к увеличению толщины ксилемы на сжатой стороне и усилению образования латеральных корней в месте натяжения (9).0193 Рисунок 1 К).
Боковые и секторальные стратегии использовались изогнутыми корнями для поддержания поглощения и транспортировки воды в деформирующем состоянии, вызванном силами растяжения и сжатия; это привело к увеличению толщины ксилемы на сжатой стороне и усилению образования латеральных корней в месте натяжения (9).0193 Рисунок 1 К).
После 6-месячного стресс-теста на изгиб корней деревянистые корни Populus nigra продемонстрировали образование реакционной древесины (RW) из-за сил сжатия на вогнутой стороне [35] , демонстрируя характерные для корней характеристики по сравнению с полученными в изогнутом стволе. RW одревесневших корней характеризуется низкой плотностью сосудов и высоким содержанием лигнина, в основном запускается ауксином и связан с индукцией активности клеток камбия [35] . Исследование также обеспечивает некоторое начальное понимание механизмов, контролирующих эту индуцированную сжатием древесину на вогнутой стороне, характеризующуюся активацией специфических белков, которые управляют деформацией клеточной стенки, одревеснением и дифференцировкой ксилемы [35] . Аналогичное исследование с использованием более короткого (2 месяца) стресс-теста на сгибание корней наблюдало специфичное для сектора изгиба распределение фитогормонов (ауксина, цитокинина и абсцизовой кислоты), которое отражало адаптацию к силам, специфичным для сжатия или растяжения [69] . Эти изменения позже подтвердились в механике появления всходов арабидопсиса. В недавнем исследовании была предложена модель для объяснения механики изгиба гипокотиля [70] , в которой сообщается, что максимумы ауксина генерируются на внутренней стороне изгиба за счет полярного транспорта ауксина через компоненты механизма транспорта ауксина PIN3, PIN4, PIN7 и AUX1, продвижение пектина с высокой степенью метилэтерификации и, следовательно, придание жесткости стенке и снижение скорости удлинения клеток; это противоположно внешней стороне, которая имела низкий уровень ауксина, что способствовало деметилэстерификации пектина, ослаблению клеточной стенки и более быстрому удлинению клеток [70] .
Аналогичное исследование с использованием более короткого (2 месяца) стресс-теста на сгибание корней наблюдало специфичное для сектора изгиба распределение фитогормонов (ауксина, цитокинина и абсцизовой кислоты), которое отражало адаптацию к силам, специфичным для сжатия или растяжения [69] . Эти изменения позже подтвердились в механике появления всходов арабидопсиса. В недавнем исследовании была предложена модель для объяснения механики изгиба гипокотиля [70] , в которой сообщается, что максимумы ауксина генерируются на внутренней стороне изгиба за счет полярного транспорта ауксина через компоненты механизма транспорта ауксина PIN3, PIN4, PIN7 и AUX1, продвижение пектина с высокой степенью метилэтерификации и, следовательно, придание жесткости стенке и снижение скорости удлинения клеток; это противоположно внешней стороне, которая имела низкий уровень ауксина, что способствовало деметилэстерификации пектина, ослаблению клеточной стенки и более быстрому удлинению клеток [70] . В другом исследовании изогнутых древесных корней растений тополя De Zio et al. [30] наблюдал асимметричный отраслевой ответ. Эти различия выражаются в измеренных параметрах, с более высокой концентрацией лигнина в вышеуказанном секторе изгиба (АБС) и меньшим количеством углеводов на вогнутой стороне АБС, а также уменьшенным количеством индолуксусной кислоты (ИУК) в выпуклом секторе. стороны как изгиба (BS), так и ниже изгиба (BBS), а также формирование RW за счет повышенной активности камбиальных клеток на вогнутой стороне BS и BBS [30] . Было обнаружено, что эти изменения строго коррелируют со способностью клеток сосудистого камбия воспринимать специфические сигналы и, в свою очередь, управлять специфическими генами, ведущими к образованию RW (в направлении вогнутой стороны) или боковых корней (в направлении выпуклой стороны). Недавно Димитрова и соавт. [71] предоставили новую информацию о координации ответа, связи и потенциальных сигнальных путях, которые были асимметрично активированы вдоль основной оси корня, которые в основном делегировались Ca 2+ (для образования новых боковых корней) и АФК (для гравитропной реакции и накопления лигнина).
В другом исследовании изогнутых древесных корней растений тополя De Zio et al. [30] наблюдал асимметричный отраслевой ответ. Эти различия выражаются в измеренных параметрах, с более высокой концентрацией лигнина в вышеуказанном секторе изгиба (АБС) и меньшим количеством углеводов на вогнутой стороне АБС, а также уменьшенным количеством индолуксусной кислоты (ИУК) в выпуклом секторе. стороны как изгиба (BS), так и ниже изгиба (BBS), а также формирование RW за счет повышенной активности камбиальных клеток на вогнутой стороне BS и BBS [30] . Было обнаружено, что эти изменения строго коррелируют со способностью клеток сосудистого камбия воспринимать специфические сигналы и, в свою очередь, управлять специфическими генами, ведущими к образованию RW (в направлении вогнутой стороны) или боковых корней (в направлении выпуклой стороны). Недавно Димитрова и соавт. [71] предоставили новую информацию о координации ответа, связи и потенциальных сигнальных путях, которые были асимметрично активированы вдоль основной оси корня, которые в основном делегировались Ca 2+ (для образования новых боковых корней) и АФК (для гравитропной реакции и накопления лигнина). Кроме того, некоторые данные указывают на то, что вогнутая сторона изогнутого сектора, где механические силы наиболее интенсивны, сообщается с другими (соседними и дальними) секторами, индуцируя пространственно связанные стратегии, обеспечивающие поглощение воды и сопровождающие клеточные модификации . 71] . Предполагается, что связь между этими частями осуществляется с помощью сигналов на короткие расстояния, таких как химические и электрические сигналы, гидравлические импульсы плазматической мембраны или плазмодесмы и меристематические коннектомы [72] , чтобы преодолевать большие расстояния и приспосабливать корневое тело к окружающей среде.
Кроме того, некоторые данные указывают на то, что вогнутая сторона изогнутого сектора, где механические силы наиболее интенсивны, сообщается с другими (соседними и дальними) секторами, индуцируя пространственно связанные стратегии, обеспечивающие поглощение воды и сопровождающие клеточные модификации . 71] . Предполагается, что связь между этими частями осуществляется с помощью сигналов на короткие расстояния, таких как химические и электрические сигналы, гидравлические импульсы плазматической мембраны или плазмодесмы и меристематические коннектомы [72] , чтобы преодолевать большие расстояния и приспосабливать корневое тело к окружающей среде.
В течение последних нескольких десятилетий исследовательские усилия позволили получить частичное представление о реакции корней растений на рассеянный склероз. Однако остаются большие пробелы, особенно в отношении специфических физиологических, молекулярных и генетических процессов, связанных с механосенсорикой и механотрансдукцией.