
Суточные и годовые ритмы: Ритмы суточные
Ритмы суточные
Суточная, сезонная и многолетняя периодичность внешних условий и проявление внутренних (эндогенных) ритмов организмов, флуктуации популяций достаточно синхронно отражается в цикличности всего сообщества — биоценоза.[ …]
Суточные циклы наиболее резко выражены в условиях климата высокой континентальности, где значительная разница между дневными и ночными температурами. В пустынях Средней Азии многие животные ведут ночной образ жизни летом, а зимой переходят на дневной (змеи, пауки и др.). Однако суточные ритмы наблюдаются во всех географических зонах, и даже в тундре в полярный день растения закрывают и открывают свои цветки в соответствии с этими ритмами.[ …]
Суточные ритмы протекают на фоне годовых, годовые — на фоне многолетних. Существуют также вековые, многолетние ритмы, например изменения климата (похолодание — потепление, иссушение — увлажнение).[ …]
Ритмы поступления на поверхность почвы солнечной энергии и влаги (суточной, годовой, многолетней) и биологические ритмы развития растительности обусловили резко выраженную сезонную цикличность почвообразования, имеющую огромное значение для формирования почвенного плодородия.
Суточный ритм жизни в известных пределах специфичен для вида, его отдельных популяций, отдельных этапов развития и рыб, находящихся в разном биологическом состоянии. Суточная ритмика жизни вырабатывается, в первую очередь, как приспособление к добыванию пищи и защите от врагов. Рыба связана со своими кормовыми объектами и врагами сложной системой отношений, одним из отражений которых является суточный ритм жизни.[ …]
В суточном ритме меняются и функциональные параметры экосистемы — интенсивность фотосинтеза и переработки первичной биологической продукции во вторичную. Лишь в почве, заселенной армадой простейших и беспозвоночных животных, жизнь в ночные часы замедляется незначительно. Сезонные ритмы. Обитатели экосистемы хорошо адаптированы к смене времен года: растения на зиму сбрасывают листья, животные «утепляются», увеличивая прослойку жира и густоту шерстного покрова, впадают в спячку или мигрируют в более благоприятные и теплые условия (птицы), зайцы меняют «маскировочные халаты» и становятся белыми и т.
Изучение суточных ритмов активности стрекоз — одно из необходимых условий для оценки их роли в трофических сетях биоценозов, например, в регуляции численности гнуса или вредителей сельского и лесного хозяйств, а также представляет интерес с точки зрения выяснения адаптивных возможностей этих насекомых.[ …]
Борисов С.Н. Суточный ритм активности Anax parthenope Selys (Odo— nata, Libellulidae) в условиях аридной зоны // Докл.[ …]
Циркадные и суточные ритмы лежат в основе способности организма чувствовать время. Механизм, ответственный за такую периодическую активность — будь то питание или размножение, — получил название «биологических часов».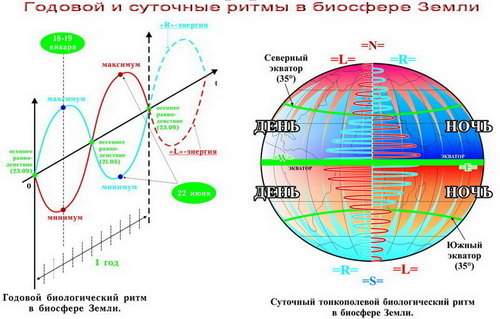 Поразительная точность работы биологических часов, управляющих жизнедеятельностью многих растений и животных, является объектом исследований ученых разных стран мира (рис. 7.13).[ …]
Поразительная точность работы биологических часов, управляющих жизнедеятельностью многих растений и животных, является объектом исследований ученых разных стран мира (рис. 7.13).[ …]
В случае человека суточные и циркадные ритмы затрагивают многие свойства и физиологические процессы (температура тела, артериальное давление, митотическая активность клеток, биоэлектрическая активность мозга, количество тромбоцитов в периферической крови и т. д.). Предполагают, что эти ритмы генетически контролируются, т. к. у нейроспоры и дрозофилы установлены так называемые «часовые» гены, которые детерминируют циркадные ритмы этих организмов. Установлено также, что циркадный ритм присущ синтезу мРНК на «часовых» генах и что существуют «часовые» белки, способные ингибировать экспрессию «часовых» генов.[ …]
В настоящее время суточные ритмы стрекоз наиболее полно изучены в аридных условиях на юго-западе Таджикистана. В зависимости от распределения максимумов здесь выделяются три основных типа активности: преимущественно дневная — для термоксерофильных видов на протяжении всего сезона и гигрофильных в холодное время года (ранней весной, поздней осенью и зимой), преимущественно утренневечерняя — для гигрофильных видов в теплое время года и преимущественно вечерняя (сумеречная) — для ряда видов сем.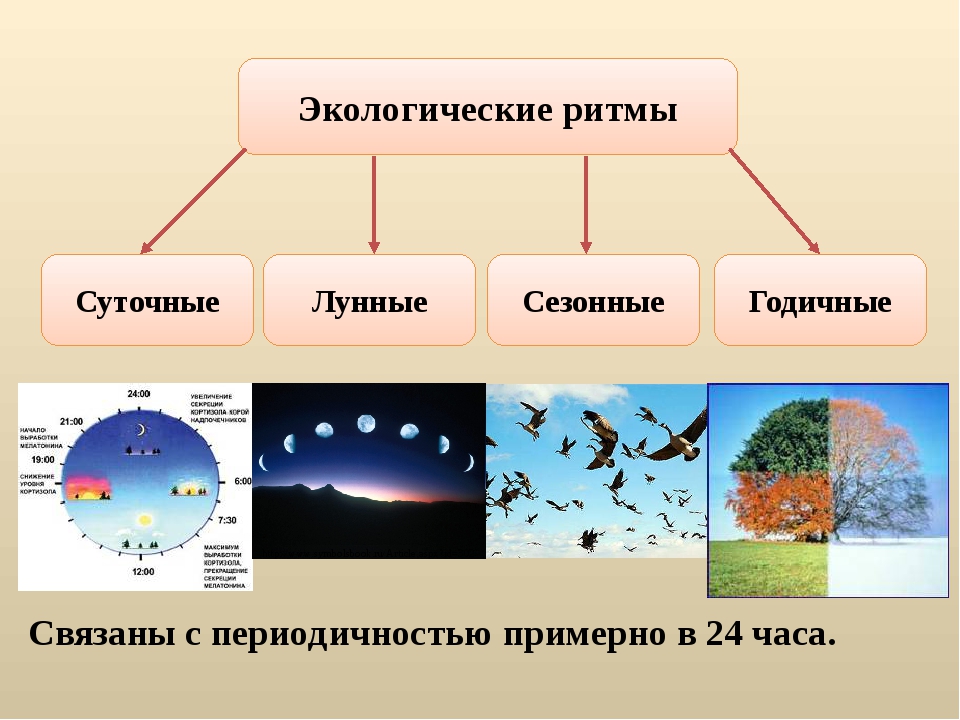
Еще лучше изучена суточная ритмика у животных: циклы деления, конъюгации, биолюминесценции у простейших, время спаривания, откладки яиц, вылупление из куколок у насекомых, ритмическая деятельность желез у позвоночных и многое другое. В ряде случаев зафиксированы точные параметры таких циклов и их составляющих. Так, суточные круговые движения вьющихся стеблей Phaseolus vulgaris складываются из трех параметров (скорость движения, его вертикальная и горизонтальная составляющие), обнаруживающих ультрадиан-ные ритмы с периодом 80—110 мин (В. Millet, W. Koukkari, 1990). Эксперименты с мадрепоровыми кораллами Acropora acuminata показали, что меченый углерод включается в ткани только в дневное время с максимумом в послеполуденные часы; интенсивность видимой кальцификации максимальна в полдень и минимальна в полночь (D. Barnes, G. Grassland, 1978). Опыты со слизнями LaevicauUs alta выявили четкую периодичность уровня фосфорилазной активности с максимумом в 0 ч и минимумом в 12 ч. Циклы такого же типа с максимумом в 20 ч и минимумом в 8 ч отмечены у скорпиона Hetemmetrus fulvipes, также ведущего ночной образ жизни.
В целом сведения о суточной активности стрекоз очень фрагментарны. Необходимо изучение ритмики отдельных видов с учетом всех видов частной активности и факторов, влияющих на них. Важно выяснить особенности ритмов стрекоз в экстремальных условиях среды, например, в высоких широтах, в высокогорье, зимой на юге, а у распространенна видов — в разных ландшафтно-географических районах.[ …]
Реакция организма на суточную смену режима освещения (день и ночь) называется фотопериодизмом. В связи с фотопериодизмом в организме изменяются процессы обмена веществ, роста и развития. Фотопериодичность — это один из главных факторов, влияющих на биологические часы организма, определяющие его физиологические ритмы в соответствии с изменениями в окружающей среде.[ …]
Однако чисто эндогенных ритмов, как и чисто экзогенных, по-видимому, не существует. Воздействие внешних факторов подчас существенно изменяет характер суточного ритма, определенный эндогенной программой. Это и понятно: активность животных отражает весь слоясный комплекс их взаимоотношений со средой и должна быть максимально синхронизирована с постоянно меняющимися условиями жизни. Абсолютное сохранение автономных, независимых от среды циклов биологически не выгодно так же как и абсолютное «подчинение» активности ежеминутно меняющимся условиям. Сочетание устойчивых эндогенных ритмов с корректирующим влиянием внешних факторов — оптимальное решение этой биологической задачи.[ …]
Воздействие внешних факторов подчас существенно изменяет характер суточного ритма, определенный эндогенной программой. Это и понятно: активность животных отражает весь слоясный комплекс их взаимоотношений со средой и должна быть максимально синхронизирована с постоянно меняющимися условиями жизни. Абсолютное сохранение автономных, независимых от среды циклов биологически не выгодно так же как и абсолютное «подчинение» активности ежеминутно меняющимся условиям. Сочетание устойчивых эндогенных ритмов с корректирующим влиянием внешних факторов — оптимальное решение этой биологической задачи.[ …]
Циклические изменения сообществ отражают суточную, сезонную и многолетнюю периодичность внешних условий и проявления эндогенных ритмов организмов. Суточная динамика экосистем связана главным образом с ритмикой природных явлений и носит строго периодический характер. Нами уже было рассмотрено, что в каждом биоценозе имеются группы организмов, активность жизни у которых приходится на разное время суток. Одни активны днем, другие — ночью. Отсюда в составе и в соотношении отдельных видов биоценоза той или иной экосистемы происходят периодические изменения, так как отдельные организмы на определенное время выключаются из него. Суточную динамику биоценоза обеспечивают как животные, так и растения. Как известно, у растений в течение суток изменяются интенсивность и характер физиологических процессов — ночью не происходит фотосинтез, нередко у растений цветки раскрываются только в ночные часы и опыляются ночными животными, другие приспособлены к опылению днем. Суточная динамика в биоценозах, как правило, выражена тем сильнее, чем значительнее разница температур, влажности и других факторов среды днем и ночью.[ …]
Суточную динамику биоценоза обеспечивают как животные, так и растения. Как известно, у растений в течение суток изменяются интенсивность и характер физиологических процессов — ночью не происходит фотосинтез, нередко у растений цветки раскрываются только в ночные часы и опыляются ночными животными, другие приспособлены к опылению днем. Суточная динамика в биоценозах, как правило, выражена тем сильнее, чем значительнее разница температур, влажности и других факторов среды днем и ночью.[ …]
Фотопериодизм нельзя отождествлять с обычными внешними суточными ритмами, которые у животных и человека переходят во врожденные свойства вида и становятся внутренними (эндогенными) ритмами. Но в отличие от изначально внутренних ритмов они могут отличаться от точной цифры — 24 часа — на 15—20 минут, и поэтому их называют циркадными (в переводе — близкие к суткам).[ …]
Изменения облика биоценозов, связанные с динамикой активности, сезонными ритмами в жизни растений и животных, сменами видового состава на протяжении годового цикла, отражают комплекс приспособлений экосистемы в целом к суточной и сезонной динамике условий существования. Характерной чертой такого типа динамики является сохранение принципиальных свойств данной экосистемы, поддержание ее целостности и функциональной устойчивости. Даже сезонные изменения видового состава входят в общую характеристику каждой данной экосистемы, поскольку закономерно повторяются год за годом.[ …]
Характерной чертой такого типа динамики является сохранение принципиальных свойств данной экосистемы, поддержание ее целостности и функциональной устойчивости. Даже сезонные изменения видового состава входят в общую характеристику каждой данной экосистемы, поскольку закономерно повторяются год за годом.[ …]
Напряжение популяции развивается циклично, что обусловлено солнечными, сезонными, суточными и другими природными ритмами. Детерминированные природными процессами, они далеко не всегда сбалансированы с ритмами производственно-технологической деятельности человека, его социальной активностью. Циклы напряжения популяции необходимо рассматривать с учетом миграционных потоков населения. Если популяция города замещается на 20—30% в год, воздействие локальных циклических природно-производ-ственных условий на приезжих будет иным, чем на прошедших период адаптации. Вместе с этим состояние напряжения данной популяции, куда вливаются 20—30% вновь приезжающих, меняется в зависимости от величины и состава уехавшей части. [ …]
[ …]
В опытах американских ученых (R. Knopoka, S, Benzer, 1971) обработка дрозофил мутагеном нарушала ритмы локомоторной активности и выкукливания. Более подробный анализ позволил идентифицировать ген («per»), нормальный аллель которого определяет околосуточный ритм, а мутации его влияют на биоритмы: у одних мутантов изменялась длительность эндогенного ритма, другие оказывались апериодичными (J. Takahashi, 1992). Наследуемый характер суточной ритмики выявлен и в опытах с мелкими грызунами; в некоторых случаях удавалось выявить гены, ответственные за наследование биоритмов (В. Mumm et al., 1988; J. Miller, 1989).[ …]
Суточная ритмика жизнедеятельности проявляется прежде всего в питании рыб. Можно сказать, что в период нагула ряд других биологических ритмов (суточные миграции, образование стай, рассеивание рыб и пр.) связан с ритмом питания у рыб. На этот вопрос можно ответить с достаточной определенностью и сказать, что суточная ритмика, например, питания рыб различна у взрослых рыб, молоди и сеголетков. Но вот вопрос о том, различна ли суточная ритмика питания рыб в пределах популяции, скажем, взрослых рыб одного и того же стада или расы или даже вида, не только не исследован, но даже и не ставился.[ …]
Но вот вопрос о том, различна ли суточная ритмика питания рыб в пределах популяции, скажем, взрослых рыб одного и того же стада или расы или даже вида, не только не исследован, но даже и не ставился.[ …]
Изменения в природной среде происходят непрерывно с определенной ритмичностью; выделяют суточные, связанные с вращением Земли вокруг своей оси; годовые—с движением Земли по орбите вокруг Солнца и многолетние, которые во многом обусловлены главным образом активностью Солнца и внутренними (эндогенными) процессами в недрах Земли. Суточные и годовые ритмы оказывают влияние на жизнедеятельность организмов, аналогичное влияние оказывают ритмы, связанные, например, с фазами Луны и т. п. В целом географическая оболочка развивается поступательно, от простого к сложному, от низшего к высшему.[ …]
В своих исследованиях мы придерживались терминологии, предложенной в монографии BJB. Чернышева /1984/ «Суточные ритмы активности насекомых», — первой сводке в мировой литературе. Прежде чем перейти к обсуждению методов изучения суточной активности, поясним, что она означает по отношению к стрекозам (рассматривается только для имаго). По определению BJ5. Чернышева, активность-это любая деятельность насекомого. Средством для ее осуществления служит подвижность. Б.Ф. Белышев /19676/ признаком активности стрекоз считает полёт, -гак как с ним целиком связано питание и размножение — важнейшие жизненные функции. Представления о лёте и активности отождествляют и другие авторы /Заика, Воронова, 1977; Заика, 1980; Олигер, 1980, 1985; Борисов, 1985в, 1987/, что в целом неоправданно сужает диапазон понятия активности стрекоз, фактически ограниченный периодом с момента начала движения после ночного покоя до полного прекращения двигательной активности перед ночевкой. Иначе, между началом или прекращением активности и непосредственно лётом существуют переходные периоды, когда насекомые проявляют двигательную активность (например, выползают из укрытий), но из-за действия каких-либо факторов еще или уже не способны к полету. Эти периоды соответствуют времени потенциальной готовности — ВПГ /Чернышев, 1973, 1981, 1984/. Знание ВПГ имеет непосредственно практическое значение при профилактике гельминтоз-ных заболеваний, переносчиками которых являются стрекозы.
По определению BJ5. Чернышева, активность-это любая деятельность насекомого. Средством для ее осуществления служит подвижность. Б.Ф. Белышев /19676/ признаком активности стрекоз считает полёт, -гак как с ним целиком связано питание и размножение — важнейшие жизненные функции. Представления о лёте и активности отождествляют и другие авторы /Заика, Воронова, 1977; Заика, 1980; Олигер, 1980, 1985; Борисов, 1985в, 1987/, что в целом неоправданно сужает диапазон понятия активности стрекоз, фактически ограниченный периодом с момента начала движения после ночного покоя до полного прекращения двигательной активности перед ночевкой. Иначе, между началом или прекращением активности и непосредственно лётом существуют переходные периоды, когда насекомые проявляют двигательную активность (например, выползают из укрытий), но из-за действия каких-либо факторов еще или уже не способны к полету. Эти периоды соответствуют времени потенциальной готовности — ВПГ /Чернышев, 1973, 1981, 1984/. Знание ВПГ имеет непосредственно практическое значение при профилактике гельминтоз-ных заболеваний, переносчиками которых являются стрекозы. Во время потенциальной готовности стрекозы наиболее уязвимы и могут в массе поедаться птицами.[ …]
Во время потенциальной готовности стрекозы наиболее уязвимы и могут в массе поедаться птицами.[ …]
Более всего естественная среда обитания изменяется в крупных городах. Этому способствуют специфический ритм жизни, психоэмоциональная обстановка труда и быта и пр. Как отмечает В.И.Торшин, интенсивность солнечной радиации в городах на 15-20% ниже, чем в прилегающей местности, тогда как среднегодовая температура примерно на 1,5°С выше, не столь значительны суточные и сезонные колебания температуры, чаще возникают туманы, больше осадков (в среднем на 10%), ниже атмосферное давление.[ …]
В ходе исторического развития организмы в ответ на смену дня и ночи, на смену времен года, т. е. в ответ на основные ритмы Земли, обусловленные ее вращением вокруг Солнца, выработали в процессе адаптогенеза способность к ритмической жизнедеятельности, что получило название биоритмов. Характерная особенность биоритмов заключается в том, что они осуществляются синхронно с процессами, протекающими в среде периодически. Различают суточные ритмы (24-часовые) и околосуточные, которые протекают во время от 20 до 28 часов и которые называют циркадными (от лат. cirka — вокруг, около, dies — день).[ …]
Различают суточные ритмы (24-часовые) и околосуточные, которые протекают во время от 20 до 28 часов и которые называют циркадными (от лат. cirka — вокруг, около, dies — день).[ …]
В этой связи можно считать, что структура человека представ-Ляет собой совокупность множества колебательных контуров. На своих частотах и ритмах функционируют сердце, легкие, желудок Др., то есть все внутренние органы. Сейчас в организме человека явлено около 300 суточных ритмов. И если бы в природе не су-ствовало такого многообразия ритмов, развитие организмов Эт Ло остановиться на уровне простейших биосистем. Исходя из сЯтс°’ Можно сказать> что колебательные процессы, а к ним отно-и0 я Магнитные и электромагнитные излучения, составляют ос-а У Нащего мироздания. Но какие из них «вредные» для человека, «полезные», не всегда ясно.[ …]
Кроме поведенческих, физиологических, морфогенетических реакций к ним относятся особенности морфологии организмов, способствующих экономии или рассеиванию энергии, диапауза, спячка, ритм суточной активности, сезонные миграции, запасание пищи. [ …]
[ …]
К гигрофильным видам относятся представители рода Ischnura (i. elegans, I. evansi, L fountainei ) и, видимо, еще некоторые равнокрылые стрекозы. Для видов этой группы в теплый период года характерен бимодальный ритм с максимумами активности в утренние и вечерние часы. Таким образом, они избегают воздействия высоких температур при низкой влажности воздуха (рис. 10, 16). К этой же группе относится своеобразный по типу ритм Anax parthe— поре и, вероятно, некоторые другие виды сем. Мощные вечерние лики охотничьей активности приурочены к сумеркам и совпадают с максимумами активности гнуса (рис. 10, II ). Отмечена также активность утром, в сумерках и ночью, чего не наблюдается у стрекоз в умеренных широтах. Аналогичное поведение у близкого вида A. imperator описывает Корбет/Corbet, 1957/ На юге Украины вечерний охотничий лёт наблюдается у Brachytron. hafniense ( = pratense), Ае. affinis, Ае. coluberculus, Cordulia aenea /Олигер, 1985/. Для видов рода Aeschna активность с одним вечерним максимумом отмечена в лесостепи Восточного Приуралья и Западной Сибири /Харитонов, 1975а, б/. [ …]
[ …]
Каждая почва обладает определенным комплексом свойств и процессов с конкретными показателями их величин. В природной обстановке наблюдается динамичность свойств и процессов почв в связи с ритмами поступления на поверхность почвы света, тепла, влаги и ритмами биологической активности почв. Совокупность суточных и сезонных количественных и качественных изменений почвенных показателей называется почвенным режимом. Различают режимы: питательный, физико-химический, температурный, влажности, газового состава почвенного воздуха, окислительновосстановительный, микробиологический, ферментативный и др. Наряду с ними выделяют режимы, отражающие совокупность явлений, поступления, передвижения (переноса) и расхода элементов питания растений, влаги, воздуха и тепла в почвах: пищевой, водный, воздушный и тепловой.[ …]
Экология рыб представляет собою раздел ихтиологии, посвященный изучению образа жизни ркб, а именно: характера динамики их популяций, внутривидовых (стаи, косяки, колонии) и межвидовых группировок, распределения, миграций, суточного и сезонного ритма . жизни, характера пищевых взаимоотношей, размножения, и т. д. Экология изучает суточный, сезонный и жизненный цикл рыбы, характер взаимосвязей ее с абиотическим и биотическим окружением (Г. Никольский, 1955).[ …]
жизни, характера пищевых взаимоотношей, размножения, и т. д. Экология изучает суточный, сезонный и жизненный цикл рыбы, характер взаимосвязей ее с абиотическим и биотическим окружением (Г. Никольский, 1955).[ …]
Сравнение количества эхомишеней, регистрируемых на одних и тех же участках полигона и на полигонах в целом в разные часы .суток показывает, что оно не остается постоянным. Известно (Поддубный и др., 1978), что это явление связано с суточной ритмикой активности рыб, спецификой образа жизни разных видов и проявления у них Двигательных оборонительных и пищевых реакций. Обычно рыбам свойственны одно-, двух- или трехпиковые ритмы суточной активности, связанные с суточным ходом освещенности и использованием для кормления и отдыха удобных ш экологической, обстановке смежных биотопов. Многие рыбы во время отдыха затаиваются на склонах углублений дна, в подводных зарослях, среди затонувших деревьев и т. п. убежищах и становятся недоступны облову и эхолоту. Ритмы суточной активности видоспецифичны и их сезонные и годовые изменения достаточно стереотипны. [ …]
[ …]
С особенностями освещения связано строение рыбыее органа зрения, наличие или отсутствие органов свечения, развитие других органов чувств, окраска и др. Свет оказывает определенное влияние и на ход обмена веществ рыбы, на созревание половых продуктов. Таким образом, для большинства рыб свет является необходимым элементом их среды.[ …]
Ритмичность явлений в географической оболочке. Географическая оболочка Земли постоянно изменяется, усложняются взаимосвязи между ее отдельными компонентами. Эти изменения происходят во времени и пространстве. В природе существуют ритмы разной продолжительности. Короткие, суточные и годовые, ритмы особенно важны для живых организмов. Их периоды покоя и активности согласуются с этими ритмами. Суточный ритм (смена дня и ночи) обусловлен вращением Земли вокруг своей оси; годовой (смена времен года) — обращением Земли вокруг Солнца. Годовая ритмика проявляется в существовании периодов покоя и вегетации у растений, в линьке и миграции животных, в некоторых случаях — в спячке, размножении. Годовая ритмика в географической оболочке зависит от широты мест: в экваториальных широтах она выражена слабее, чем в умеренных или полярных.[ …]
Годовая ритмика в географической оболочке зависит от широты мест: в экваториальных широтах она выражена слабее, чем в умеренных или полярных.[ …]
Поскольку все сообщества, кроме глубоководных, почвенных и пещерных, испытывают на себе в течение суток изменение интенсивности света и температуры, можно ожидать, что в активности основной части популяций в большинстве сообществ наблюдается периодичность, связанная прямо или косвенно с этими изменениями. Периодичность сообщества является результатом синхронной активности в течение дня и ночи целых групп организмов. Некоторые, например, активны только ночью (ночные), другие — днем (дневные), а третьи — только в сумерки (сумеречные) .[ …]
Для растений вода — и условие, и ресурс. Влияние относительной влажности воздуха на организмы опосредовано температурой и скоростью ветра и ее трудно отделить от доступности воды вообще. Относительная влажность воздуха, температура и географическое распространение ТеЬарЫБ.[ …]
Наилучшее соответствие между организмами и изменяющимися условиями неизбежно предполагает некий компромисс между приспособлением -к переменам и способностью к их переживанию. В условиях многократного воздействия циклических изменений на последовательные поколения организмов естественный отбор привел к возникновению ряда особенностей образа жизни, которые и сами по себе являются циклическими. К числу таких особенностей относятся диапауза насекомых, ежегодное сбрасывание листвы листопадными деревьями, суточные движения листьев, приливно-отливный ритм перемещений у литоральных крабов, годовой цикл функционирования репродуктивных систем и сезонный цикл изменения густоты меха у млекопитающих.[ …]
В условиях многократного воздействия циклических изменений на последовательные поколения организмов естественный отбор привел к возникновению ряда особенностей образа жизни, которые и сами по себе являются циклическими. К числу таких особенностей относятся диапауза насекомых, ежегодное сбрасывание листвы листопадными деревьями, суточные движения листьев, приливно-отливный ритм перемещений у литоральных крабов, годовой цикл функционирования репродуктивных систем и сезонный цикл изменения густоты меха у млекопитающих.[ …]
Косвенные трансбиотические взаимоотношения между растениями (через животных и микроорганизмы). Важная экологическая роль животных в жизни растений состоит в участии в процессах опыления, распространения семян и плодов. Опыление растений насекомыми, получившее название энтомофилии, способствовало выработке ряда приспособлений как у растений, так и насекомых. Назовем здесь такие интересные адаптации энтомофилъных цветков как узоры, образующие «путевые нити» к нектарникам и тычинкам, нередко видимые только в ультрафиолетовых лучах, доступных для зрения насекомых; различие окраски цветков до и после опыления; синхронизация суточных ритмов раскрывания венчика и тычинок, обеспечивающие безошибочное попадание пыльцы на тело насекомого, а с него — на рыльце другого цветка, и т. д. (рис. 6.16).[ …]
д. (рис. 6.16).[ …]
БИОЛОГИЧЕСКИЕ РИТМЫ • Большая российская энциклопедия
БИОЛОГИ́ЧЕСКИЕ РИ́ТМЫ (биоритмы), периодически повторяющиеся изменения интенсивности и характера биологич. процессов и явлений. Под Б. р. в широком смысле понимают все циклич. процессы, протекающие в живых системах разного уровня организации. Периоды этих циклов варьируют от миллисекундных колебаний нейрональной активности до многолетних вариаций динамики популяций. Среди них выделяют т. н. рабочие, или физиологические, ритмы, которые являются необходимым условием функционирования биологич. систем (напр., ритмы дыхания и сокращений сердца). Их частоты определяются как физико-химич. особенностями ритмически работающих органов, так и потребностями организма. Так, во время сна сердечный ритм человека обычно составляет от 40 до 90 ударов в минуту, а при интенсивной физич. работе может возрастать до 200 и более ударов. Кроме того, всем живым организмам свойственны колебания, периоды которых равны осн. геофизич. циклам, обусловленным вращением Земли вокруг собств. оси (суточный ритм) и вокруг Солнца (годовой, или сезонный, ритм). Мн. виды организмов реагируют на изменения ночной освещённости, связанной с фазами Луны, а активность обитателей морского побережья подчиняется также ритмам приливов и отливов. Такие Б. р. называют адаптивными, т. к. они направлены на координацию жизнедеятельности организмов при колебаниях абиотических и биотических параметров среды. Напр., у растений ритмы фотосинтеза непосредственно определяются чередованием светлого и тёмного времени суток (фотопериодом). Ночная активность, характерная для мн. видов грызунов, повышает вероятность их выживания в условиях пресса хищников.
работе может возрастать до 200 и более ударов. Кроме того, всем живым организмам свойственны колебания, периоды которых равны осн. геофизич. циклам, обусловленным вращением Земли вокруг собств. оси (суточный ритм) и вокруг Солнца (годовой, или сезонный, ритм). Мн. виды организмов реагируют на изменения ночной освещённости, связанной с фазами Луны, а активность обитателей морского побережья подчиняется также ритмам приливов и отливов. Такие Б. р. называют адаптивными, т. к. они направлены на координацию жизнедеятельности организмов при колебаниях абиотических и биотических параметров среды. Напр., у растений ритмы фотосинтеза непосредственно определяются чередованием светлого и тёмного времени суток (фотопериодом). Ночная активность, характерная для мн. видов грызунов, повышает вероятность их выживания в условиях пресса хищников. Благодаря годовым ритмам, особенно значимым в условиях резко континентального климата, растения и животные могут заблаговременно подготовиться к изменению погодных условий, напр. к зимнему похолоданию. При этом одни из них залегают в спячку (сурки, суслики), у других уплотняется меховой покров и усиливается способность к термогенезу (лисицы, белки, хомячки и др.), у третьих происходит накопление во внеклеточной жидкости веществ, предотвращающих её замерзание при минусовых темп-рах (лягушки, улитки, некоторые виды растений и др.).
Благодаря годовым ритмам, особенно значимым в условиях резко континентального климата, растения и животные могут заблаговременно подготовиться к изменению погодных условий, напр. к зимнему похолоданию. При этом одни из них залегают в спячку (сурки, суслики), у других уплотняется меховой покров и усиливается способность к термогенезу (лисицы, белки, хомячки и др.), у третьих происходит накопление во внеклеточной жидкости веществ, предотвращающих её замерзание при минусовых темп-рах (лягушки, улитки, некоторые виды растений и др.).
Б. р. не являются простым отражением колебаний условий во внешней среде. Растения, животные или микроорганизмы, изолированные от осн. геофизич. циклов («датчиков времени»), сохраняют в течение длительного времени присущую им ритмич. активность. Однако наблюдаемые в условиях изоляции периоды свободнотекущих ритмов близки, но не равны внешним циклам. Из-за этого несовпадения Б. р. получили назв. «циркаритмы» (от лат. circa – около). Соответственно колебания с периодом близким к 24 часам называются околосуточными или циркадными (от лат. dies – день) ритмами, а с периодом близким к 12 месяцам – окологодовыми или цирканнуальными (от лат. annual – год) ритмами. Устойчивое отклонение периодов свободнотекущих ритмов от внешних циклов служит гл. аргументом в пользу их эндогенной природы. С этих позиций суточные, годовые (сезонные), лунные и приливные колебания функций организма рассматриваются как результат синхронизации биологич. часов внешними циклич. процессами, среди которых ведущую роль играет фотопериод.
Из-за этого несовпадения Б. р. получили назв. «циркаритмы» (от лат. circa – около). Соответственно колебания с периодом близким к 24 часам называются околосуточными или циркадными (от лат. dies – день) ритмами, а с периодом близким к 12 месяцам – окологодовыми или цирканнуальными (от лат. annual – год) ритмами. Устойчивое отклонение периодов свободнотекущих ритмов от внешних циклов служит гл. аргументом в пользу их эндогенной природы. С этих позиций суточные, годовые (сезонные), лунные и приливные колебания функций организма рассматриваются как результат синхронизации биологич. часов внешними циклич. процессами, среди которых ведущую роль играет фотопериод.
Периоды свободнотекущих ритмов могут не совпадать у разных систем организма. Так, у людей, изолированных от внешних датчиков времени, периоды колебаний темп-ры тела, выведения калия и гормонов коры надпочечников с мочой в ср. равны 25 часам. В то же время периоды ритмов сна и бодрствования, выведения кальция и натрия с мочой у 1/3 людей могут либо превышать 40 часов, либо быть короче 20 часов. Это свидетельствует о способности отд. систем организма к самостоят. генерации циклов. Представления о множественной локализации биологич. часов с циркадным периодом подкреплены исследованиями изолированных органов (надпочечники, сердце, печень и др.), сохраняющих ритмич. активность в течение нескольких суток вне организма. Но даже у одноклеточной водоросли Gonialax poliedra периоды свободнотекущих ритмов хемолюминисценции и передвижения могут не совпадать друг с другом, т. е. отд. клетка также обладает несколькими часами.
равны 25 часам. В то же время периоды ритмов сна и бодрствования, выведения кальция и натрия с мочой у 1/3 людей могут либо превышать 40 часов, либо быть короче 20 часов. Это свидетельствует о способности отд. систем организма к самостоят. генерации циклов. Представления о множественной локализации биологич. часов с циркадным периодом подкреплены исследованиями изолированных органов (надпочечники, сердце, печень и др.), сохраняющих ритмич. активность в течение нескольких суток вне организма. Но даже у одноклеточной водоросли Gonialax poliedra периоды свободнотекущих ритмов хемолюминисценции и передвижения могут не совпадать друг с другом, т. е. отд. клетка также обладает несколькими часами.
Рис. 1. Циркадная организация млекопитающих и человека. Центральные биологические часы в супрахиазматических ядрах (СХЯ) гипоталамуса благодаря нервным (вегетативная нервная система, ВНС) и эндокринны. ..
..
Пока детально изучены лишь механизмы эндогенных околосуточных ритмов. Их молекулярно-генетич. основу составляют внутриклеточные системы с отрицательной обратной связью, в которых полный цикл – экспрессия специализир. генов биологич. часов (т. н. часовых генов), подавление их активности собственными белковыми продуктами и последующая деградация белков – занимает ок. 24 часов. Дополнительные контуры обратной связи, регулирующие образование факторов транскрипции, дающих начало следующему циклу, стабилизируют работу биологич. часов. Периоды свободнотекущих ритмов остаются неизменными в широком диапазоне температур даже у одноклеточных организмов. Так, цианобактерии сохраняют ритм обменных процессов близкий к 25 часам при вариациях темп-ры от 30 до 60 °C.
На основе внутриклеточных биологич. часов формируются циркадные ритмы отд. органов многоклеточного организма. Их ритмич. активность координируется центр. водителями ритма. У млекопитающих и человека роль центр. часов выполняют супрахиазматические ядра (СХЯ) гипоталамуса, которые связаны с вегетативными нервными ганглиями и эпифизом (рис. 1). Гормон эпифиза мелатонин синхронизирует экспрессию часовых генов в разл. отделах нейроэндокринной системы и совместно с нейромедиаторами и гормонами желёз внутренней секреции обеспечивает координацию ритмич. процессов, которая проявляется в упорядоченном чередовании подъёмов и спадов функциональной активности разл. систем организма. Так, перед пробуждением после сна происходит всплеск секреции гормонов коры надпочечников (кортизола и кортикостерона), играющих ключевую роль в энергетич. обеспечении физич.
часов формируются циркадные ритмы отд. органов многоклеточного организма. Их ритмич. активность координируется центр. водителями ритма. У млекопитающих и человека роль центр. часов выполняют супрахиазматические ядра (СХЯ) гипоталамуса, которые связаны с вегетативными нервными ганглиями и эпифизом (рис. 1). Гормон эпифиза мелатонин синхронизирует экспрессию часовых генов в разл. отделах нейроэндокринной системы и совместно с нейромедиаторами и гормонами желёз внутренней секреции обеспечивает координацию ритмич. процессов, которая проявляется в упорядоченном чередовании подъёмов и спадов функциональной активности разл. систем организма. Так, перед пробуждением после сна происходит всплеск секреции гормонов коры надпочечников (кортизола и кортикостерона), играющих ключевую роль в энергетич. обеспечении физич. активности во время бодрствования, а увеличение секреции гормона роста, наблюдаемое в первой половине сна, стимулирует синтез белков, что необходимо для восстановления организма во время отдыха.
активности во время бодрствования, а увеличение секреции гормона роста, наблюдаемое в первой половине сна, стимулирует синтез белков, что необходимо для восстановления организма во время отдыха.
Рис. 2. У полярников в Антарктиде суточные ритмы температуры тела (А) и сила сердечных сокращений (В) реагируют на изменения фотопериода, а ритмы концентрации кортизола в слюне (Б) и частота пульса (Г…
Ход биологич. часов настраивается внешними циклич. процессами и, прежде всего, фотопериодом. У млекопитающих, включая человека, в фотопериодич. синхронизации Б. р. участвуют зрительные рецепторы, реагирующие на освещённость. Сигналы от них передаются к СХЯ гипоталамуса и далее к эпифизу. Свет подавляет секрецию мелатонина, который выделяется эпифизом исключительно в ночные часы, независимо от того, к какому времени суток приурочены сон или бодрствование конкретного вида животных. Благодаря влиянию на экспрессию генов биологич. часов мелатонин с одинаковым успехом синхронизирует циркадные ритмы ночных животных (мыши, крысы), у которых вечерний подъём активности совпадает с повышением концентрации гормона, и ритмы дневных видов (люди, обезьяны), у которых активность повышается утром при снижении его уровня.
Благодаря влиянию на экспрессию генов биологич. часов мелатонин с одинаковым успехом синхронизирует циркадные ритмы ночных животных (мыши, крысы), у которых вечерний подъём активности совпадает с повышением концентрации гормона, и ритмы дневных видов (люди, обезьяны), у которых активность повышается утром при снижении его уровня.
Мелатонин используется позвоночными животными и для синхронизации цирканнуальных ритмов жизнедеятельности, поскольку из всего многообразия сезонных феноменов именно изменения фотопериода являются наиболее надёжным индикатором времени года. При этом не столько количество мелатонина, сколько продолжительность его ночной секреции определяют сезонные морфофизиологич. преобразования, характер которых, в свою очередь, определяется образом жизни, свойственным определённому виду. Напр., осеннее увеличение ночной активности эпифиза стимулирует генеративную функцию у копытных, но подавляет её у грызунов.
Напр., осеннее увеличение ночной активности эпифиза стимулирует генеративную функцию у копытных, но подавляет её у грызунов.
Наряду с фотопериодом в синхронизации суточных ритмов участвуют и др. периодич. процессы, напр. у млекопитающих нарушение ритмов активности, вызванное разрушением СХЯ, восстанавливается при периодич. приёме пищи. Биологич. часы, координирующие работу разных органов, отличаются по способности к восприятию синхронизирующего влияния внешних «датчиков времени» разл. природы. В результате этого циркадная организация физиологич. процессов существенно меняется при быстром перемещении в новый часовой пояс, при сменной работе, при сезонных изменениях фотопериода, что особенно важно для жителей полярных районов (рис. 2). Наблюдаемые при этом отклонения в последовательности подъёмов и спадов Б. р. разл. систем организма (десинхроноз) отрицательно сказываются на работоспособности и сопротивляемости организма негативному влиянию разл. патогенных факторов. Наука, изучающая Б. р., – биоритмология, или хронобиология.
р. разл. систем организма (десинхроноз) отрицательно сказываются на работоспособности и сопротивляемости организма негативному влиянию разл. патогенных факторов. Наука, изучающая Б. р., – биоритмология, или хронобиология.
Суточные биоритмы: что мы о них знаем?
Автор фото, Getty Images
Подпись к фото,Начало нового дня. Суточные биоритмы характерны для всего живого на Земле
Знаете ли вы, что суточные биоритмы — или циркадные ритмы — характерны для всего живого на Земле: от грибов до человека?
У всего живого — в том числе и у крошечной бактерии, которую можно рассмотреть только под мощным микроскопом — есть суточные биоритмы: биологический процесс, который занимает приблизительно 24 часа и определяет ритм нашей жизни в целом.
А что вы знаете о том, как суточные биоритмы влияют на нас?
1.
 Суточные биоритмы существуют почти с появления жизни на Земле
Суточные биоритмы существуют почти с появления жизни на Земле Автор фото, Getty Images
Подпись к фото,Суточные биоритмы начали формироваться с появлением первой живой клетки на Земле
Считается, что первая живая клетка на Земле в светлое время суток под действием ультрафиолета получила повреждения, а ночью восстановилась. Человеческий организм работает также — ночью, во время сна, запускаются процессы восстановления.
2. Внутренние биологические часы есть не только у человека
Автор фото, Getty Images
Подпись к фото,У мимозы свои биоритмы — цветы открываются и закрываются даже ночью
Считается, что суточные биоритмы есть у всего живого на Земле, что способно вырабатывать энергию под действием солнечного света.
Однако несмотря на связь с внешними стимулами, такими как солнечный свет, циркадные ритмы имеют внутреннее, эндогенное происхождение, представляя, таким образом, биологические часы организма.
Французский ботаник Огюстен Пирам Декандоль еще в 1834 году определил, что период, с которыми мимоза открывает и закрывает листья, короче длины суток и составляет примерно 22-23 часа. То есть листья мимозы открываются в темное время суток.
3. Биологические часы задают ритм нашей жизни
Автор фото, Getty Images
Подпись к фото,Даже грибы живут по своим внутренним биологическим часам
Суточные биоритмы позволяют живым существам предвосхищать наступление ночи и дня, зимы и лета и дают нам возможность подготовиться к этим событиям.
4. У вас есть внутренние часы с функцией синхронизации
Автор фото, Getty Images
Подпись к фото,Забудьте о времени по Гринвичу. Ваши часы, отражающие суточные биоритмы, точнее и всегда с вами
Ваши главные биологические часы, которые отвечают за процессы синхронизации, находятся в гипоталамусе. Эти часы-синхронизаторы, как дирижер, посылают вам определенные регулирующие сигналы в разное время в течение суток.
5. У вас есть также «периферические часы»
Автор фото, Getty Images
Подпись к фото,Внутренние и периферические часы всегда находятся в процессе синхронизации
Все ваши органы и ткани имеют дополнительные — периферические — часы, которые синхронизируются с главными часами в вашем гипоталамусе.
6. Часы есть в каждой клетке
Автор фото, Getty Images
Подпись к фото,Каждая клетка имеет свои суточные биоритмы и часы
Каждая клетка вашего организма живет по своим внутренним часам, которые отвечают за изменения, происходящие в клетке в течение каждых 24 часов.
7. Годичный ритм
Автор фото, Getty Images
Подпись к фото,Медведь начинает готовится к зимней спячке задолго до наступления зимы
Когда ночи становятся длиннее, мозг вырабатывает больше мелатонина — гормона, который регулирует состояние сна и бодрствования.
Многие животные — например, олени — реагируют на такие изменения — начиная готовиться к зимней спячке или периоду размножения.
Человеческий организм также реагирует на эти изменения и зимой вырабатывает больше антител для того, чтобы бороться с различными инфекциями, характерными для холодного времени года.
8. Дневной свет помогает вам сохранять режим
Автор фото, EPA
Подпись к фото,Для здоровья необходимо получать достаточно солнечного света
Если вам не хватает солнечного света, то ваши биологические часы сбиваются с 24-часового ритма.
Сенсоры, которые располагаются в ваших глазах реагируют на свет и темноту и посылают сигнал в мозг, которые отвечают за процессы синхронизации в вашем организме.
9. Пора спать?
Автор фото, Getty Images
Подпись к фото,Как мы понимаем, что нам пора спать?
С того момента как вы проснулись, организм начинает готовиться ко сну.
Но вы не засыпаете до того времени, пока ваши биологические часы не скажут вам, что пора спать.
10. Рассинхронизация, или джетлаг
Автор фото, Getty Images
Подпись к фото,Организму нужно время, чтобы перевести ваши суточные биологические часы
Джетлаг — рассогласование циркадного ритма человека с природным суточным ритмом.
Мы говорим о «джетлгае», когда ваши биологические часы работают в одном часовом поясе, а другие части тела — печень, кишечник, мозг и мышцы — живет в другом часовом поясе.
Для того, чтобы работа этих часов синхронизировалась нужно приблизительно по дню на каждый часовой пояс. То есть если вы прилетели в город, где разница во времени по сравнению с вашим часовым поясом, составляет три часа, то вам понадобится около трех дней, чтобы ваш организм адаптировался.
11. Социальный «джетлаг»
Автор фото, Getty Images
Подпись к фото,Будильник говорит вам «пора просыпаться», а ваш организм говорит «надо еще поспать»
Социальный джетлаг характерен для тех, кто вынужден работать по скользящему графику, или тех, у кого «социальные» и «биологические» часы сильно рассинхронизированы.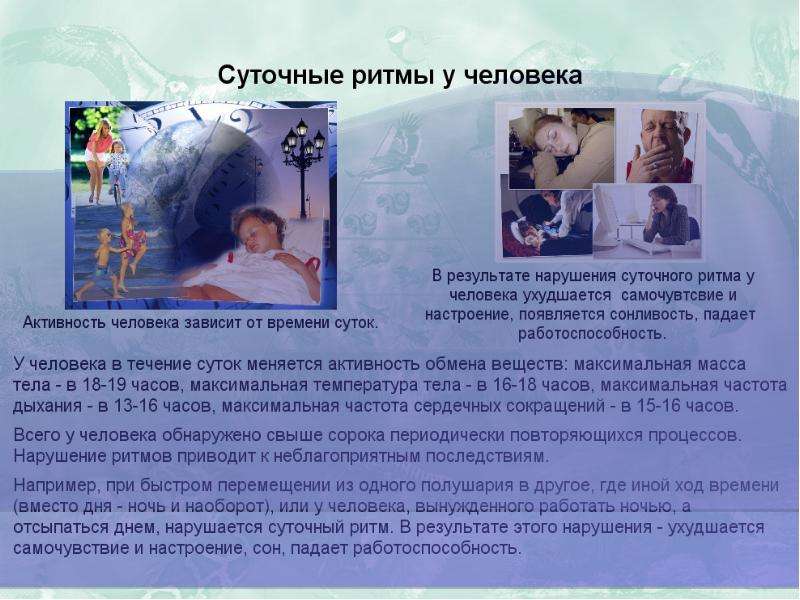
Если ваш будильник звонит тогда, когда биологические часы еще не дали сигнал к пробуждению, тогда вы живете в режиме «социального джетлага».
Как свидетельствуют многие исследования, рассогласование «социальных» и «биологических» часов повышает риск возникновения депрессии, заболеваний сердца, диабета, ожирения и даже рака.
12. Дайте подросткам поспать подольше
Автор фото, Getty Images
Подпись к фото,Для студента 7:00 — это то же самое что 5:00 для взрослого человека
Гормональные изменения в организме подростка предполагают, что сигнал к пробуждению их биологические часы будут давать как минимум на два часа позже, чем обычно.
Разбудить подростка в 7 часов утра — это то же самое, что разбудить 50-летнего человека в 5 утра.
По мере взросления подростка его биологические часы вернутся к прежнему ритму.
Приливные и годовые ритмы
Удары
волн о берег и смена сезонов могут
повлиять на эндогенные биологические
ритмы.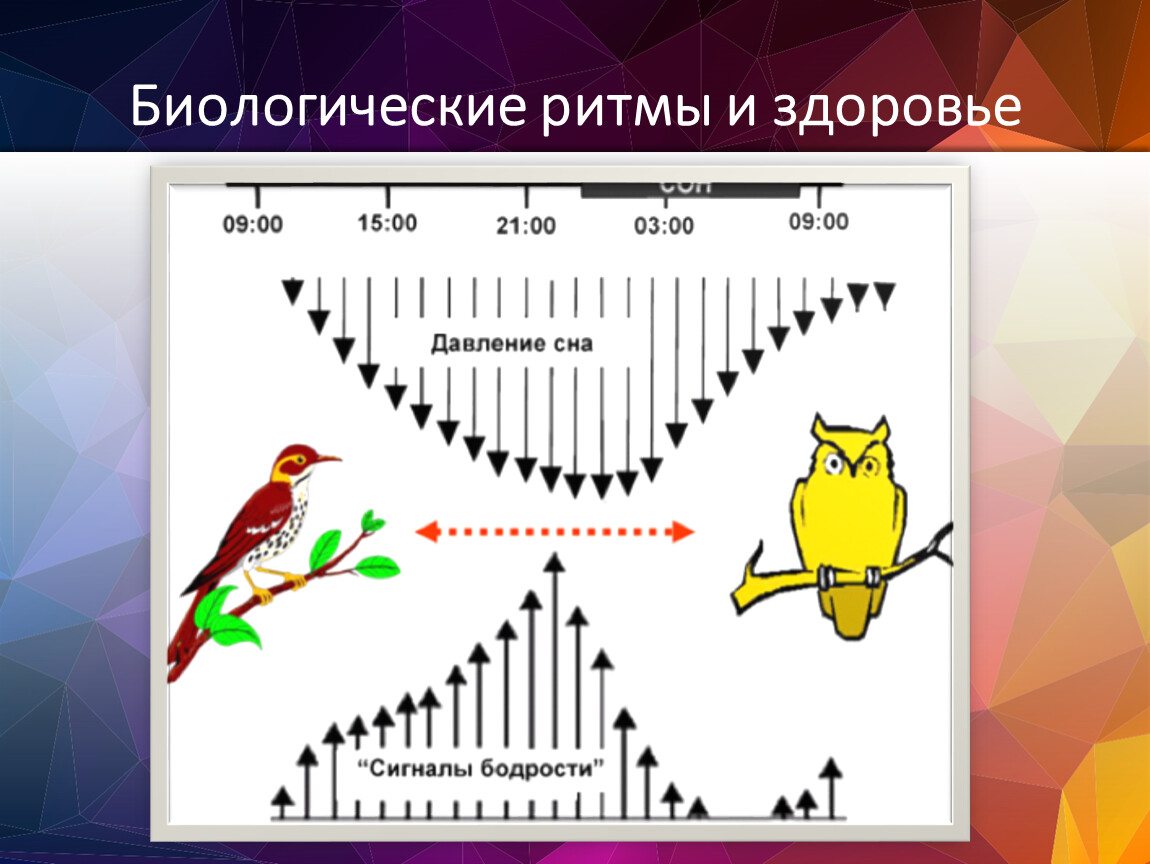 Эти приливные и годовые ритмы
являются самыми короткими и самыми
длинными соответственно.
Эти приливные и годовые ритмы
являются самыми короткими и самыми
длинными соответственно.
Неудивительно, что многие морские животные проявляют биологические ритмы, связанные с приливами и отливами и периодически повторяющиеся каждые 12,4 часа. Приспособление к этим ритмам позволяет животным выживать в условиях местообитаний с резкими циклическими изменениями, состоящими в том, что сначала территория покрывается водой, а затем за следующие 6 часов, открывается палящему солнцу и хищникам, сидящим в засаде не пляже.
Зелёный краб (Carcinas maenas) хорошо прячется среди камней или в расщелинах, когда прилив отступает. Однако он твёрдо знает, что можно выходить из своего убежища, когда вода снова начинает покрывать берег. Он один из тех животных, которые обитают в приливно- отливной зоне морских побережий и, появляясь и прячась, ярко демонстрирует ритмическое поведение.
Некоторые приливные
ритмы не являются эндогенными, они
запускаются только внешними воздействиями
– изменениями солёности, давления,
температуры, механическим воздействием,
причиняемым потоком воды.
Тем не менее существует ряд фиксированных эндогенных приливных ритмов, для которых внешние воздействия служат в основном для синхронизации с приливами, а не для их запуска. Даже после того, как североамериканские манящие крабы (род Uca) содержались в полной темноте, без приливов на протяжении 25 дней, они всё ещё демонстрировали обычный, связанный с приливами ритм появления и затаивания, подтверждая, что эти ритмы являются эндогенными. В местах обитания этих видов крабов, на берегу моря приливы опаздывают на 50 минут каждый день. Это запоздание было смоделировано в экспериментальном приливном ритме, созданным для этих крабов изучавшим их в начале 1970 года доктором Ж. Палмером. В отличие от зелёных, манящие крабы появляются при низком приливе для поисков партнёра и демонстрации территориального поведения и прячутся под водой во время высокого прилива.
Годовые ритмы
Годовые
ритмы с периодом в 365 дней служат причиной
разного рода поведенческих феноменов,
происходящих только один раз в году,
включая миграции животных в разных
уголках земного шара, зимнюю спячку и
даже смену оперения и окраски меха в
течении года.
Так как годовую ритмику составляют очень длительные периоды времени, их сложно смоделировать в экспериментах и соответственно выяснить, являются ли годовые ритмы, демонстрируемые животными, только запускаемыми внешними факторами или эндогенными.
Эксперименты, предполагают содержание животных в искусственных условиях постоянного света или темноты, или температуры в течении многих лет, для того чтобы пронаблюдать, сохраняется ли определённый ритм. Присутствие эндогенных ритмов было выявлено у нескольких различных видов. Например было показано, что годовой цикл линьки обоих видов – садовой славки (Sylvia borin) является эндогенным.
Птицы обоих видов содержались в постоянных условиях при 12-часовой освещённости и следующей за ней 12-часвой темноты на протяжении более чем двух лет. Этот режим исключал какие-либо сезонные изменения освещенности, и при нём годовой цикл линьки сохранился.
В лабораторных
экспериментах было доказано, что
изменения в семенниках обыкновенного
скворца (Sturnus
vulgaris)
управляются эндогенными ритмами. Таким
же образом при содержании многие годы
в постоянной темноте жука антренуса
(Anthrenus
verbasci)
он сохранял свой годовой ритм появления
взрослого насекомого из куколки. И
пятнистый олень (Cervus
nippon)
продолжал сбрасывать и отращивать рога
ежегодно, несмотря на световой режим,
экспериментально навязанный ему.
Таким
же образом при содержании многие годы
в постоянной темноте жука антренуса
(Anthrenus
verbasci)
он сохранял свой годовой ритм появления
взрослого насекомого из куколки. И
пятнистый олень (Cervus
nippon)
продолжал сбрасывать и отращивать рога
ежегодно, несмотря на световой режим,
экспериментально навязанный ему.
В другую очень важную
группу биологических ритмов, имеющих
огромное значение для высших и низших
организмов, входят сезонные (околосезонные),
годичные ритмы, обусловленные вращением
Земли вокруг Солнца. Сезонные изменения
растительного покрова Земли, миграция
птиц, зимняя спячка ряда видов животных
— это примеры ритмов с годичным периодом.
Сезонные колебания жизненных функций
характерны и для человека. Так, в регионах
с сезонными контрастами климата
интенсивность обмена веществ выше
зимой, чем летом. Холод является адекватным
стимулятором функции щитовидной железы.
Артериальное давление, количество
эритроцитов, гемоглобина обычно ниже
в жаркое время года.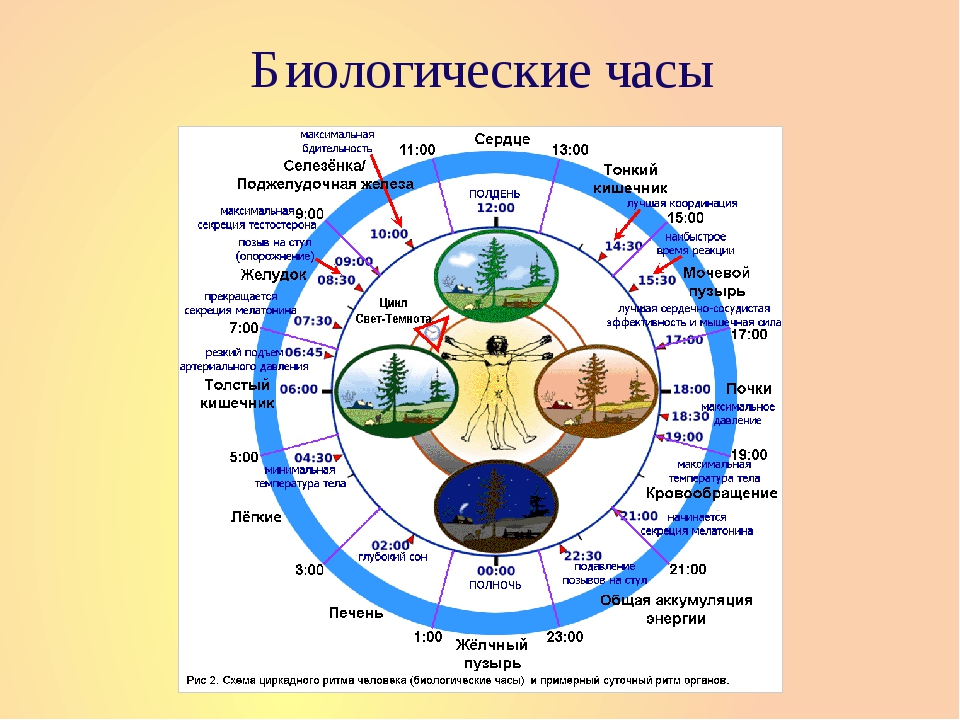 Весной и летом у
большинства людей работоспособность
выше, чем зимой. Пик выдающихся спортивных
достижений приходится на весенне-летний
и ранний осенний периоды. Хорошо известно
волнообразное течение многих заболеваний,
при котором периоды обострения сменяются
длительными ремиссиями: так, туберкулез
чаще обостряется весной, а язвенная
болезнь желудка и двенадцатиперстной
кишки — весной и осенью. В осенне-зимний
и весенний периоды выявляют наибольшее
число первичных больных инсулинозависимых
сахарным диабетом.
Весной и летом у
большинства людей работоспособность
выше, чем зимой. Пик выдающихся спортивных
достижений приходится на весенне-летний
и ранний осенний периоды. Хорошо известно
волнообразное течение многих заболеваний,
при котором периоды обострения сменяются
длительными ремиссиями: так, туберкулез
чаще обостряется весной, а язвенная
болезнь желудка и двенадцатиперстной
кишки — весной и осенью. В осенне-зимний
и весенний периоды выявляют наибольшее
число первичных больных инсулинозависимых
сахарным диабетом.
Сезонные колебания физиологических показателей у многих теплокровных в определенной мере повторяют суточные: в зимний период отмечается понижение обмена и двигательной активности, в весенне-летний — активизация физиологических процессов.
| 1
кафедра экологии г. Сыктывкар Министерство образования и науки Российской Федерации Федеральное агентство по образованию ГОУ ВПО «Сыктывкарский государственный университет» Химико-биологический факультет Кафедра экологии “Приспособительные ритмы жизни ” (10 класс) Выполнила: Студент 245 гр. Осташова Е.В. ___________________ Проверила: Уваровская О.В
___________________ Тема урока
“Приспособительные ритмы жизни ” Задачи урока: Образовательные – изучить определения понятий фотопериод и фотопериодизм; сформировать знания о значении суточных и годовых ритмах в жизни животных и растений. Развивающие – развитие
умения работать с карточками, систематизировать, сравнивать и обобщать полученные знания, выделять главное существенное, рефлексировать, слушать, кроме того, развитие навыков групповой самоорганизации, способности четко формулировать свои мысли Тип урока и форма его организации
– урок изучения нового материала.
Список использованной литературы: 1. Пономарева О.Н., Чернова Н.М. Методическое пособие к учебнику под редакцией Н.М. Черновой «Основы экологии. 10 (11) класс.» М.: Дрофа, 2001. С. 51-57. 2. Чернова Н.М., Галушин В.М., Константинов В.М. Основы экологии. 10 (11) кл. общеобразоват. учреждений М.: Дрофа, 2001. С. 60-65. Ход урока: До урока учитель вместе с учениками расставляет столы для работы учащихся в малых группах. На начало урока, учащиеся садятся по своему выбору.
ПРИЛОЖЕНИЕ Карточка №1 Суточные ритмы. Суточные ритмы приспосабливают организмы к смене дня и ночи. У растений интенсивный рост, распускание цветков приурочены к определенному времени суток. Животные в течение суток сильно меняют активность. По этому признаку различают дневные и ночные виды. Суточный ритм организмов – это не только отражение смены внешних условий. Если поместить человека, или животных, или растения в постоянную стабильную обстановку без смены дня и ночи, то сохраняется ритмика процессов жизнедеятельности, близкая к суточной. Организм как бы живет по своим внутренним часам, отсчитывая время. Суточный ритм может захватывать многие процессы в организме. У человека около 100 физиологических характеристик подчиняются суточному циклу: частота сокращения сердца, ритм дыхания , выделение гормонов роста, секрета пищеварительных желез, кровяное давление , температура тела и многие другие. Поэтому, когда человек бодрствует вместо сна, организм все равно настроен на ночное состояние и бессонные ночи плохо отражаются на здоровье. Карточка №2
Карточка №3 Фотопериодизм. Резкие кратковременные изменения погоды (летние заморозки, зимние оттепели) обычно не нарушают годовых ритмов растений и животных. Главный экологический фактор , на который реагируют организмы в своих годовых циклах, — не случайные изменения погоды, а фотопериод – изменения в соотношении дня и ночи. Длина светового дня закономерно изменяется в течении года, а именно эти изменения служат точным сигналом приближения весны, лета, осени или зимы. Способность организмов реагировать на изменение длины дня получила название фотопериодизм . Если день сокращается, виды начинают готовиться к зиме, если удлиняется – к активному росту и размножению. В этом случае для жизни организмов важен не сам фактор изменения длины дня и ночи, а его сигнальное значение, свидетельствующее о предстоящих глубоких изменениях в природе. Карточка №4 Линька – периодическое обновление покровов: смена перьев, рогового слоя кожи, роговых чешуек на лапах и клюве. Различают несколько типов линек. Постювенальная линька – полная или частичная замена контурных перьев молодых птиц контурными перьями, характерными для взрослой птицы. Предбрачная линька, присущая некоторым видам птиц , — частичная линька, при которой отдельные перья на голове, туловище и хвосте сменяются ярко-окрашенными, что обеспечивает яркость брачного наряда. Послебрачная линька чаще затрагивает все оперение, характерна для всех видов птиц. У водоплавающих и болотных птиц сбрасывание крупных перьев происходит в короткий срок, и поэтому птицы на некоторое время утрачивают способность к полету (становясь достаточно легкой добычей хищников). Виды, далеко мигрирующие и имеющие короткий период между окончанием размножения и началом миграции, характеризуются короткими сроками линьки и ее полнотой. Карточка №5
С миграциями у птиц связаны физиологические и поведенческие изменения: увеличение веса перелетных птиц к периоду миграций; потребления пищи сверх нормы, необходимой для поддержания обычной жизнедеятельности; отложение в теле большого количества жира; миграционный полет ; утрата инстинкта территориальности и усиление инстинкта стайности. У рыб различают нерестовые, кормовые, зимовальные, анадромные (из моря к берегам и далее вверх по рекам) и катадромные (в обратном направлении). Среди рыб мигрируют камбала, корюшка, угорь. |
 Ритмика внешней среды создает периодичность, т. е. повторяемость условии в жизни большинства видов. Регулярно повторяются как критические, трудные для выживания периоды, так и благоприятные.
Ритмика внешней среды создает периодичность, т. е. повторяемость условии в жизни большинства видов. Регулярно повторяются как критические, трудные для выживания периоды, так и благоприятные.Как настроить свои биологические часы — Амурская правда
— Причина — в биологических ритмах, часть из которых управляется внутренними часами в организме, — рассказывает врач Амурского областного центра медпрофилактики Наталья Артемчук. — Если научиться подстраиваться под расписание биологических часов своего организма, можно не только регулировать свое поведение и настроение, но и продлить жизнь.
Внутренние часы
— Все основные процессы жизнедеятельности в нашем организме проходят с некоторой периодичностью, цикличностью. В определенном ритме работают буквально все органы — сердце, легкие, печень, органы выделения и пищеварения. Их работа управляется внутренними часами, — продолжила Наталья Леонидовна. — Ученые обнаружили у большинства живых существ несметное количество таких часов.
— Наталья Леонидовна, а какие физиологические процессы могут иметь сезонный ритм?
— К примеру, весной и осенью у человека повышается секреторная активность желудка, в то же время защитные свойства слизи снижаются. Это предрасполагает к сезонным обострениям таких заболеваний, как гастрит, язвенная болезнь желудка и двенадцатиперстной кишки.
Но одними из важнейших биоритмов человека являются суточные, или, как их еще называют, циркадные ритмы. Они подчинены 24-часовому графику — времени, за которое наша планета Земля делает полный оборот вокруг собственной оси. Циркадным ритмам подчиняется более 300 биологических процессов, в том числе частота сердечных сокращений, артериальное давление, частота и глубина дыхания, секреция гормонов, кроветворение, моторная и секреторная активность желудка… Именно циркадные ритмы определяют время сна, бодрствования, приема пищи.
Суточные и другие ритмы следует учитывать при назначении лекарственных препаратов. Например, эффективность противоаллергических средств повышается при их применении во второй половине дня, так как именно в этот период в крови увеличивается содержание биологически активных веществ, вызывающих аллергические проявления. В то же время глюкокортикоидные гормоны с заместительной целью целесообразнее принимать утром, моделируя суточный ритм работы надпочечников. Конечно же, вопрос о кратности и времени назначения препаратов решает врач, учитывающий как фармакологические аспекты, так и индивидуальные особенности пациента. В определенном ритме, синхронизированном с другими системами организма, работают органы пищеварения. Хорошо известен суточный ритм желудочной секреции с максимумом во второй половине ночи. Именно поэтому характерным симптомом язвенной болезни двенадцатиперстной кишки являются ночные боли в животе.
Гормоны и биоритмы
— Какой орган управляет биоритмами?
— За биологические ритмы организма отвечают примерно 20 тысяч нейронов в гипоталамусе — этот отдел мозга размером с рисовое зернышко находится в середине головы и синхронизирует работу различных систем, в том числе нервной и эндокринной. Ученым по сей день не удалось определить, как именно работает данный «хронометр», но они смогли выяснить, что настраивается он с помощью сигналов окружающей среды. И самый очевидный сигнал — это солнечный свет. Ученые считают, что ранний подъем и отход ко сну сразу после заката позволяют лучше всего настраивать биологические часы.
— Кто-то может раньше лечь спать и встать с рассветом, а кому-то это трудно: ему лучше вечером работается. Надо ли бороться со своими ритмами?
— При необходимости можно не спать и всю ночь. Но постоянный недосып не пройдет безнаказанно. В течение циркадного дня (бодрствования) наша физиология в основном настроена на переработку накопленных питательных веществ, чтобы получить энергию для активной дневной жизни. А ночью питательные вещества накапливаются, происходит восстановление и «починка» тканей. Ночью повышается выработка гормона роста, который стимулирует анаболические процессы, например размножение клеток и накопление питательных веществ в печени. Не зря говорят: «Дети растут во сне». Если вы постоянно недосыпаете, то повышается чувство голода, вас тянет на сладкое и жирное, в организме замедляются и нарушаются обменные процессы. Конечно, есть индивидуальные особенности биологических ритмов, колебания работоспособности у людей. Одни, так называемые жаворонки, рано встают и более энергично работают в первой половине дня, другие — совы — вечером. У первых наблюдается опережение, а у вторых — отставание фазы сна. Еще ученые выделяют голубей — это те люди, которые имеют наивысшую работоспособность в середине дня.
— Может ли хронотип передаться по наследству?
— Ученые говорят, что да. При этом с хронотипом связаны не только определенные черты характера, но и показатели здоровья, адаптационных возможностей. Люди вечернего типа, которые ложатся спать позже 23—24 часов, легче приспосабливаются к работе в ночную смену, но они и в большей степени, чем другие, подвержены риску возникновения сердечно-сосудистой патологии и других заболеваний. У «жаворонков», как правило, многие показатели здоровья лучше.
Старайтесь прислушиваться к своим внутренним ощущениям, не нарушать биоритмы. Циркадный или суточный баланс — это такое состояние, когда человек чувствует себя физически здоровым, когда организм высыпается, у нас хороший аппетит, отличное настроение и мы полны энергии. Значит, мы находимся в своем ритме!
Пики активности органов в течение суток
Проводить очищающие процедуры или принимать витамины и другие целебные вещества для укрепления конкретного органа лучше во время его высшей активности.
Легкие
Наиболее активно работают с 3 до 5 часов утра. В это время полезно спать при открытой форточке (если, конечно, погода позволяет), чтобы организм мог максимально насытиться кислородом, а низшая активность — с 15 до 17 часов.
Толстый кишечник
На своем пике работоспособности — с 5 до 7 часов утра, а с 17 до 19 часов «отдыхает».
Желудок
Готов активно работать с 7 до 9 часов утра. Низшая активность — с 19 до 21 часа.
Сердце
Легче перенесет значительные эмоциональные и физические нагрузки с 11 до 13 часов дня, а тяжелее всего с 23 до часа ночи.
Мочевой пузырь
Максимально накапливает жидкость из организма с 15 до 17 часов, а меньше всего с 3 до 5 утра.
Желчный пузырь
Активен с 23 до 1 часа ночи — в это время организм производит максимальные усилия по созданию желчи, а минимум — в 11—13 часов.
Печень
Время максимальной регуляции крови и желчи — с 1 до 3 часов ночи. Низшая активность печени в 13—15 часов.
Острота зрения у человека сильнее всего снижается в 2 часа ночи — это особенно важно знать автомобилистам. Одиночество тяжелее всего переносится между 20 и 22 часами.
Что такое джетлаг и как его избежать
Сбить с толку наши внутренние часы могут и дальние авиаперелеты. Для этого придумали даже особый термин — джетлаг, обозначающий нарушение биологических ритмов человека из-за смены часовых поясов. В норме внутренние часы человека настроены на суточные ритмы того часового пояса, в котором он живет. Когда происходит быстрое перемещение, организм не успевает перестроиться и какое-то время функционирует так же, как если бы вы остались дома.
Как свести к минимуму последствия смены часовых поясов
— Перед вылетом переведите ваши часы на время того пояса, в который летите, — это поможет настроиться на новое время психологически.
— Если летите с востока на запад, по прибытии старайтесь не спать, пока не наступит темное время суток. А за несколько дней до отлета ложитесь спать и вставайте на пару часов позже, чтобы приучить организм к сдвигу часового режима.
— Если летите на восток, попробуйте выспаться в самолете и не спать в течение дня в месте прибытия. За несколько дней до отлета надо ложиться и вставать раньше.
— В путешествии ложитесь спать и вставайте в одно и то же время: четкое соблюдение режима помогает организму быстрее адаптироваться к новому часовому поясу.
Статья опубликована на средства областного бюджета в рамках долгосрочной целевой программы «Развитие здравоохранения Амурской области на 2012—2014 годы», подпрограмма «Формирование навыков здорового образа жизни и профилактика основных неинфекционных заболеваний у населения на 2012—2013 годы».
Возрастная категория материалов: 18+
Материалы по теме
Диспансеризация охватит жителей самых отдаленных амурских селСотрудники благовещенского спорткомплекса «Юность» сдают нормы ГТОДежурный терапевт спешит на помощь в чате: как сервис «СберЗдоровье» развивает телемедицинуРуководитель цитолаборатории Нина Новожилова: «Рак — это болезнь, которая играет с нами в прятки»Невидимые лучи против опухолей: врач-радиолог о современной лучевой терапииОртопедический матрас Magniflex — находка для людей с больным позвоночникомКостры рябин: врачи и ботаники о пользе и красоте популярного в сентябре дереваЧастые перекусы замедляют обмен веществ: ученые изменили взгляды на дробное питаниеСбербанк запускает многофункциональный сервис СберЗдоровье«У каждого должно быть свое время ничегонеделания»: как пережить изоляцию с детьмиОт стресса к простуде: почему даже ОРВИ начинается от нервов и как вирус помогает телуПоказать еще
Биологические адаптивные ритмы. Фотопериодизм (Курсовая работа)
Содержание:
- Суточный ритм
- Приливные и синодические ритмы
- Годовые ритмы
- Фотопериодизм
- Вывод:
| Предмет: | Биология |
| Тип работы: | Курсовая работа |
| Язык: | Русский |
| Дата добавления: | 18.05.2019 |
- Данный тип работы не является научным трудом, не является готовой выпускной квалификационной работой!
- Данный тип работы представляет собой готовый результат обработки, структурирования и форматирования собранной информации, предназначенной для использования в качестве источника материала для самостоятельной подготовки учебной работы.
Если вам тяжело разобраться в данной теме напишите мне в whatsapp разберём вашу тему, согласуем сроки и я вам помогу!
По этой ссылке вы сможете найти много готовых курсовых работ по биологии:
Посмотрите похожие темы возможно они вам могут быть полезны:
Введение:
Одним из фундаментальных свойств живой природы является цикличность большинства происходящих в ней процессов. Вся жизнь на Земле, от клетки до биосферы, подчиняется определенным ритмам. Природные ритмы для любого организма можно разделить на внутренние (связанные с его собственной жизнедеятельностью) и внешние (циклические изменения в окружающей среде).
Внутренние петли. Внутренние циклы — это прежде всего физиологические ритмы организма. Физиологический процесс не проводится непрерывно. Ритм обнаруживается в процессах синтеза ДНК и РНК в клетках, в сборке белков, в функционировании ферментов и в деятельности митохондрий. Деление клеток, сокращение мышц, работа эндокринных желез, сердцебиение, дыхание, возбудимость нервной системы, то есть работа всех клеток, органов и тканей организма, подчиняются определенному ритму. Кроме того, каждая система имеет свой период. Этот период может быть изменен действием факторов окружающей среды только в узких пределах, а для некоторых процессов это совершенно невозможно. Этот ритм называется эндогенным.
Все внутренние ритмы тела подчинены, интегрированы во всю систему и в конечном итоге выступают в качестве общей периодичности поведения организма. Ритмично осуществляя свои физиологические функции, организм как бы считает время. Как для внешних, так и для внутренних ритмов начало следующей фазы зависит, прежде всего, от времени. Следовательно, время выступает в качестве одного из важнейших факторов окружающей среды, на которые должны реагировать живые организмы, приспосабливаясь к внешним циклическим изменениям в природе.
Внешние ритмы. Основные внешние ритмы имеют геофизическую природу, так как они связаны с вращением Земли относительно Солнца и Луны относительно Земли. Под влиянием этого вращения естественным образом меняются многие факторы окружающей среды на нашей планете, в частности световой режим, температура, давление и влажность, атмосферное электромагнитное поле, океанические приливы и т. д. Кроме того, космические ритмы, такие как периодические изменения солнечной активности, также влияют на дикую природу. 11-летний и ряд других циклов характерны для Солнца. Изменения солнечной радиации существенно влияют на климат нашей планеты. Помимо циклического воздействия абиотических факторов, внешние ритмы для любого организма также являются регулярными изменениями в активности и поведении других живых существ.
Ряд изменений в жизнедеятельности организмов совпадает за период с внешними геофизическими циклами. Это так называемые адаптивные биологические ритмы — суточные, приливные, равные лунному месяцу, годовые. Благодаря им важнейшие биологические функции организма, такие как питание, рост, размножение, совпадают с наиболее благоприятным временем дня или года.
Адаптивные биологические ритмы возникли как адаптация физиологии живых существ к регулярным изменениям окружающей среды во внешней среде. Этим они отличаются от чисто физиологических ритмов, которые поддерживают непрерывную жизнь организмов — дыхание, кровообращение, деление клеток и т. д.
Суточный ритм
Суточный ритм встречается у разных организмов, от одноклеточных до человека. У человека более 100 физиологических функций, на которые влияет ежедневная периодичность: сон и бодрствование, изменения температуры тела, частоты сердечных сокращений, глубины и частоты дыхания, объема и химического состава мочи, потоотделение, мышечная и умственная работоспособность и т. д. При амебе во время дни меняют скорость деления. У некоторых растений открытие и закрытие цветков, поднятие и опускание листьев, максимальная частота дыхания, скорость роста колеоптилей и т. д. Ограничены определенным временем.
По смене периодов сна и бодрствования животные делятся на дневные и ночные. Дневная активность ярко выражена, например, у домашних цыплят, большинства воробьиных, сусликов, муравьев, стрекоз. Обычно ночные животные — ежи, летучие мыши, совы, кабаны, большинство кошачьих, травяные лягушки, тараканы и многие другие. Животные обычно активны в наиболее удобное время суток. Конкретные виды деятельности довольно разнообразны. Некоторые виды имеют примерно одинаковую активность как днем, так и ночью, с чередующимися короткими периодами бодрствования и покоя. Такой ритм называется многофазным (многие землеройки, несколько плотоядных животных и т. д.).
У ряда животных суточные изменения влияют в основном на двигательную активность и не сопровождаются значительными отклонениями физиологических функций (например, у грызунов). Наиболее яркими примерами физиологических сдвигов в течение дня являются летучие мыши. Летом, в дневное время, многие из них ведут себя как пойкилотермные животные. Температура их тела в это время практически равна температуре окружающей среды; пульс, дыхание и возбудимость чувств резко снижены. Чтобы взлететь, нарушенная мышь долго прогревается из-за химического тепловыделения. Вечером и ночью это типичные гомеотермические млекопитающие с высокой температурой тела, активными и точными движениями, быстрой реакцией на добычу и врагов.
У некоторых видов периоды активности ограничены определенным временем суток, у других они могут меняться в зависимости от ситуации. Итак, раскрытие цветков шафрана зависит от температуры, соцветий одуванчика — от света: в пасмурный день корзины не открываются. Активность пустынной мокрицы или темных жуков меняется в разное время суток в зависимости от температуры и влажности на поверхности почвы. Они оставляют норки либо рано утром и вечером (двухфазный цикл), либо только ночью (однофазный), либо в течение дня.
В эксперименте можно отличить эндогенные циркадные ритмы от экзогенных, то есть навязанных внешней средой. У многих видов при полном постоянстве внешних условий (температура, освещенность, влажность и т. д.) Циклы, близкие к суточному, сохраняются в течение длительного времени. Например, у дрозофилы такой эндогенный ритм прослеживается на протяжении десятков поколений. Таким образом, ежедневный цикл жизни переходит во врожденные генетические свойства вида. Такие эндогенные ритмы были получены в контурах (от латинского ок — о и умирает — день, день), поскольку их продолжительность не одинакова у разных особей одного и того же вида, незначительно отличаясь от среднего 24-часового периода. При удалении внешних ритмов дня и ночи эндогенный ритм становится свободно текущим и через некоторое время перестает совпадать с суточными изменениями.
Белки-летяги, для которых характерна сумеречная активность, просыпаются вечером синхронно, в строго определенный час. В эксперименте, находясь в полной темноте, они поддерживают циркадный ритм. Тем не менее, некоторые люди начинают свой «день» на несколько минут раньше; другие — на несколько минут позже обычного ежедневного цикла. Если, например, циркадный ритм на 15 минут короче суточного, то для такого животного через три дня разница во времени с внешним ритмом составит 45 минут, через 10 дней — уже 2,5 часа и т. д. Поэтому все белки-летяги просыпаются через несколько дней и начинают двигаться в разное время, хотя каждая из них сохраняет постоянство своего цикла. При восстановлении смены дня и ночи соя и бодрствование животных снова синхронизируются. Таким образом, внешний (суточный цикл) регулирует продолжительность врожденных циркадных ритмов, согласовывая их с изменением окружающей среды.
У людей циркадные ритмы изучались в различных ситуациях: в пещерах, герметических камерах, при погружениях и т. д. Было обнаружено, что типологические характеристики нервной системы играют важную роль в отклонениях от суточного цикла у людей. Циркадные ритмы могут быть разными даже среди членов одной семьи.
Хорошо известный стереотип поведения благодаря циркадному ритму облегчает существование организмов с ежедневными изменениями в окружающей среде. Однако, когда животные и растения переселяются и попадают в географические условия с разным ритмом дня и ночи, слишком сильный стереотип может стать неблагоприятным. Поэтому возможности переселения ряда видов ограничены глубокой фиксацией их циркадных ритмов.
Так, например, серые крысы отличаются от черных значительно большей пластичностью суточного цикла. У черных крыс он почти не поддается реструктуризации, и этот вид имеет ограниченный ареал, тогда как серые крысы распространились почти по всему миру.
У большинства видов возможна перестройка циркадного ритма. Обычно это происходит не сразу, а захватывает несколько циклов и сопровождается рядом нарушений в физиологическом состоянии организма. Например, для людей, которые летят на большие расстояния в широтном направлении, их физиологический ритм десинхронизируется с местным астрономическим временем.
Тело сначала продолжает функционировать по-старому, а затем начинает перестраиваться. В этом случае ощущается повышенная утомляемость, недомогание, желание спать днем и бодрствовать ночью. Адаптивный период длится от нескольких дней до двух недель.
Десинхронизация ритма является важной медицинской проблемой при организации ночной и сменной работы для людей разных профессий, в космических полетах, подводном плавании, подземных работах и т. д.
Циркадные и циркадные ритмы лежат в основе способности организма ощущать время. Эта способность живых существ называется «биологическими часами».
Ряд высокоорганизованных животных обладают сложной врожденной способностью использовать временную ориентацию для ориентации в пространстве. Птицы во время длительных полетов также постоянно корректируют свое направление относительно Солнца или поляризованного света неба, учитывая время суток. «Биологические часы» живых организмов ориентируют их не только в суточный цикл, но и в более сложные геофизические циклы изменений природы.
Приливные и синодические ритмы
Виды, живущие на побережье, живут в условиях очень сложной периодичности внешней среды. Чередование приливов накладывается на 24-часовой цикл колебаний освещения и других факторов. В течение лунного дня (24 ч 50 мин) наблюдаются 2 прилива и 2 отлива, фазы которых меняются ежедневно примерно на 50 минут. Кроме того, сила приливов естественным образом изменяется в течение синодического или лунного месяца (29,5 солнечных дней). Дважды в месяц (новолуние и полнолуние) они достигают максимального значения (так называемые сизигийные приливы).
Жизнь организмов, обитающих в прибрежной зоне, подчинена этому сложному ритму. Устрицы во время отлива плотно сжимают клапаны и перестают есть.
Частота открытия и закрытия раковин поддерживается в них долгое время и в аквариумах. Он постепенно меняется, если вы перемещаете аквариум в другую географическую область, и в конце концов он устанавливается в соответствии с новым графиком прилива, хотя моллюски не испытывают непосредственного воздействия. Эксперименты предполагают, что перестройка вызвана восприятием устрицами тех изменений состояния атмосферы, которые связаны с приливными явлениями.
Рыба-атерин, живущая у побережья Калифорнии, использует высоту сизигийных приливов в своем жизненном цикле. Во время прилива самки откладывают яйца у кромки воды, закапывая ее в песчаную почву. При отступлении воды икра остается созревать на мокром песке. Жаркое появляется через две недели и ограничивается следующим приливом.
Периодичность, равная лунному месяцу (синодический ритм), в качестве эндогенного ритма была обнаружена у ряда морских и наземных организмов. Это проявляется в ограничении определенных фаз Луны нерестом многолистных червей палоло, размножением японских морских лилий, роем ряда хирономидных комаров и подёнок. В этих случаях достигается синхронизация деятельности, связанной с необходимостью оплодотворения. У ряда животных обнаружена периодичность, равная лунному месяцу в реакции на свет, на слабые магнитные поля и на скорость ориентации. У людей предполагается исходная связь менструальных циклов с синодическим месяцем, отмечаются изменения в склонности к кровотечению у оперированных пациентов и т. д. Адаптивное значение большинства эндогенных лунных ритмов до сих пор неизвестно.
Годовые ритмы
Годовые ритмы являются одними из самых универсальных в живой природе. Регулярные изменения в физических условиях в течение года вызвали широкий спектр адаптаций к этой периодичности в эволюции видов. Наиболее важные из них связаны с размножением, ростом, миграцией и переживанием неблагоприятных периодов года. У видов с коротким жизненным циклом годовой ритм естественным образом проявляется в ряде поколений (например, цикломорфоз у дафний и коловраток).
Сезонные изменения — это глубокие изменения в физиологии и поведении организмов, влияющие на их морфологию и особенности жизненного цикла. Адаптивный характер этих изменений очевиден: благодаря им такой важный момент в жизни вида, как появление потомства, приурочен к наиболее благоприятному времени года, а переживание критических периодов происходит в самое стабильное состояние.
Чем острее сезонные изменения в окружающей среде, тем более выражена годовая периодичность организмов. Осенний листопад, различные диапаузы, гибернация, накопление жира, сезонное линька, миграция и т. д. Развиваются в основном в умеренном и холодном климате, в то время как сезонные тропики менее выражены у тропических людей.
Годовые ритмы многих видов эндогенные. Такие ритмы называются цирковыми (лат. Annus-год). Это особенно актуально для циклов размножения. Так, животные Южного полушария, которые содержатся в зоопарках Северного, чаще всего размножаются зимой или осенью, что соответствует весне и лету на их родине.
Австралийские страусы в заповеднике Аскания-Нова откладывали яйца зимой прямо на снегу. Собака Динго приносит щенков в декабре, когда в Австралии заканчивается весна. Стабильность периодов размножения в годовом цикле необходимо учитывать при интродукции и акклиматизации видов.
Сильные оттепели зимой, морозы летом обычно не нарушают сезонных изменений у растений и животных. Однако точность годового цикла далеко не всегда является эндогенной. Например, семена ряда растений прорастают в строго определенное время года даже после экспериментально индуцированного состояния полной приостановленной анимации, которое должно мешать «обратному отсчету» в организме. Следовательно, прорастание стимулируется некоторыми изменениями окружающей среды, связанными с геофизическими циклами.
В настоящее время интенсивно изучается реакция организмов на слабые геоэлектромагнитные поля, а также атмосферные приливы, которые естественным образом изменяются в циклах вращения Земли. Было показано, что интенсивность ряда биологических процессов коррелирует с колебаниями этих тонких показателей состояния атмосферы в течение года, таких как, например, двигательная активность насекомых, скорость потребления кислорода клубнями картофеля, и т.д.
Таким образом, наступление следующей стадии годового цикла у живых организмов происходит частично вследствие эндогенного ритма, а частично обусловлено колебаниями факторов внешней среды.
Следует отметить, что годовая частота не зависит от мощных факторов окружающей среды, непосредственно влияющих на организм (температура, влажность и т. д.), Которые подвержены сильной изменчивости погоды, а от свойств окружающей среды, которые являются вторичными для жизненно важных функций, которые, однако, измениться очень естественно в течение года. Адаптивное значение этого явления заключается в том, что кратковременные изменения погодных условий, их возможные существенные отклонения от нормы не изменяют биологический ритм организмов, что остается синхронизированным с общим течением изменений природы в течение года.
Одним из наиболее точных и регулярно меняющихся факторов окружающей среды является продолжительность светового дня, ритм чередования темных и светлых периодов дня. Именно этот фактор служит большинству живых организмов для ориентации во времени года.
Фотопериодизм
Реакция организмов на сезонные изменения длины дня называется фотопериодизмом. Его проявление зависит не от интенсивности освещения, а только от ритма чередования темных и светлых периодов дня.
Фотопериодическая реакция живых организмов имеет большое адаптивное значение, поскольку для подготовки к неблагоприятным условиям или, наоборот, к наиболее интенсивной жизненной активности требуется достаточно много времени. Способность реагировать на изменения продолжительности дня обеспечивает опережающие физиологические изменения и приспособление цикла к сезонным изменениям условий. Ритм дня и ночи служит сигналом о грядущих изменениях климатических факторов, оказывающих сильное прямое влияние на живой организм (температуру, влажность и т. д.).
В отличие от других факторов окружающей среды, ритм освещения влияет только на те особенности физиологии, морфологии и поведения организмов, которые являются сезонными адаптациями в их жизненном цикле. Образно говоря, фотопериодизм — это реакция организма на будущее.
Хотя фотопериодизм встречается во всех крупных систематических группах, он не характерен для всех видов.
Существует много видов с нейтральным фотопериодическим ответом, у которых физиологические изменения в цикле развития не зависят от продолжительности дня. Такие виды либо имеют другие способы регулирования жизненного цикла (например, озимые культуры в растениях), которые развиваются, либо их нет необходимости точно регулировать. Например, там, где нет выраженных сезонных изменений, большинство видов не имеют фотопериодизма. Цветение, плодоношение и отмирание листьев на многих тропических деревьях продлевается во времени, и одновременно на дереве можно найти цветы и плоды.
В умеренном климате виды, которым удается быстро завершить жизненный цикл и практически не встречающиеся в активном состоянии в течение неблагоприятных сезонов года, также не проявляют фотопериодических реакций, например, многие многолетние эфемероидные растения и эфемеры.
Существует два типа фотопериодической реакции: короткий день и длинный день. Известно, что продолжительность светового дня, кроме времени года, зависит от географического положения района. Виды короткого дня живут и растут в основном в низких широтах, а виды длинного дня — в умеренных и высоких. У видов с обширными ареалами северные особи могут отличаться по типу фотопериодизма от южных. Таким образом, тип фотопериодизма является экологической, а не систематической особенностью вида.
У растений и животных с длинным днем увеличивающиеся весенние и ранние летние дни стимулируют процессы роста и готовятся к размножению. Укорочение дней второй половины лета и осени вызывает торможение роста и подготовку к зиме. Таким образом, морозостойкость клевера и люцерны намного выше при выращивании растений в короткий день, чем в длинный. На деревьях, растущих в городах возле уличных фонарей, осенний день удлиняется, в результате листопад задерживается и они чаще обморожены.
Исследования показали, что растения короткого дня особенно чувствительны к фотопериоду, так как продолжительность дня на их родине мало меняется в течение года, а сезонные климатические изменения могут быть очень значительными.
Фотопериодическая реакция тропических видов готовится к сухому и дождливому сезону. Некоторые сорта риса в Шри-Ланке, где общее годовое изменение продолжительности дня составляет не более часа, улавливают даже незначительную разницу в световом ритме, которая определяет время их цветения.
Фотопериодизм насекомых может быть не только прямым, но и косвенным. Например, у мух капустного корня зимняя диапауза происходит под влиянием качества пищи, которое варьируется в зависимости от физиологического состояния растения.
Продолжительность светового периода дня, обеспечивающая переход к следующей фазе развития, называется критической продолжительностью дня для этой фазы. По мере увеличения широты критическая длина дня увеличивается. Например, переход к диапаузе листовки яблока на широте 32 ° происходит, когда дневное время составляет 14 часов, 44 ° — 16 часов, 52 ° — 18 часов. Критическая длина дня часто служит препятствием для широтного перемещения растений и животных, для их введения.
Фотопериодизм растений и животных является наследственно закрепленным, генетически детерминированным свойством. Однако фотопериодическая реакция проявляется только под определенным влиянием других факторов окружающей среды, например, в определенном температурном диапазоне.
При определенной комбинации условий окружающей среды возможно естественное распространение видов в необычные для них широты, несмотря на тип фотопериодизма. Так, в высокогорных околотропических регионах встречается много растений длинного дня, выходцев из умеренного климата.
В практических целях продолжительность светового дня изменяется при выращивании зерновых культур в закрытом грунте, контроле продолжительности освещения, увеличении яйценоскости кур и регулировании размножения пушных зверей.
Средние долгосрочные периоды развития организмов определяются в первую очередь климатом местности, и реакции фотопериодизма адаптированы к ним. Отклонения от этих периодов определяются погодой. При изменении погодных условий сроки прохождения отдельных фаз могут варьироваться в определенных пределах. Это особенно заметно у растений и пойкилотермных животных.
Таким образом, растения, которые не достигли необходимого количества эффективных температур, не могут цвести даже в условиях фотопериода, которые стимулируют переход в генеративное состояние. Например, в Московской области береза цветет в среднем 8 мая с накоплением суммы эффективных температур 75 ° С.
Однако в годовых отклонениях время его цветения варьируется с 19 апреля по 28 мая. Гомотермические животные реагируют на погодные условия, изменяя поведение, даты гнездования и миграции.
Изучение закономерностей сезонного развития природы осуществляется специальной прикладной отраслью экологии — фенологии (дословный перевод с греческого — наука о явлениях).
Согласно биоклиматическому закону Хопкинса, который он применил к условиям Северной Америки, сроки наступления различных сезонных явлений (фенодата) в среднем различаются на 4 дня для каждого градуса широты, для каждых 5 ° долготы и 120 м над уровнем моря. Уровень моря, т. е. север, восток и выше рельеф местности, позднее наступление весны и раннее — осень. Кроме того, фенологические даты зависят от местных условий (топография, экспозиция, расстояние от моря и т. д.).
Вывод:
В Европе сроки сезонных событий меняются для каждого градуса широты не на 4, а на 3 дня.
Соединяя точки с одинаковыми фенодатами на карте, мы получаем контуры, которые отражают фронт наступления весны и наступления регулярных сезонных явлений. Это имеет большое значение для планирования многих видов деятельности, в частности сельскохозяйственных работ.
Суточные и годовые ритмы деревьев
Andersson A, Keskitalo J, Sjödin A, Bhalerao R, Sterky F, Wissel K, Tandre K, Aspeborg H, Moyle R, Ohmiya Y et al (2004) Таблица транскрипции осени старение. Геном Биол 5R: 24. DOI: 10.1186 / GB-2004-5-4-r24
Google ученый
Barbour MM, Walcroft AS, Farquhar GD (2002) Сезонные колебания δ 13 C и δ 18 O целлюлозы из годичных колец Pinus radiata .Среда растительной клетки 25: 1483–1499. DOI: 10.1046 / j.0016-8025.2002.00931.x
Google ученый
Bauch J, Eckstein D (1981) Биологические исследования древесины на панелях картин Рембрандта. Wood Sci Technol 15: 251–263. DOI: 10.1007 / BF00350943
Google ученый
Bieniawska Z, Espinoza C, Schlereth A, Sulpize R, Hincha DK, Hannah MA (2008) Нарушение циркадных часов Arabidopsis отвечает за обширные вариации в чувствительном к холоду транскриптоме.Физиология растений 147: 263–279. DOI: 10.1104 / стр.108.118059
PubMed CAS Google ученый
Bigras FJ, Colombo SJ (2001) Холодостойкость хвойных пород. Kluwer, Dordrecht
Google ученый
Björkman O, Demmig B (1987) Выход фотонов O 2 эволюция и характеристики флуоресценции хлорофилла при 77 K среди сосудистых растений различного происхождения.Planta 170: 489–504. DOI: 10.1007 / BF00402983
Google ученый
Böhlenius H, Huang T, Charbonnel-Campaa L, Brunner AM, Jansson S, Strauss SH, Nilsson O (2006) Регулирующий модуль CO / FT контролирует сроки цветения и прекращения сезонного роста деревьев. Наука 312: 1040–1043. DOI: 10.1126 / science.1126038
PubMed Google ученый
Bohn A, Rascher U, Hütt M-T, Kaiser F, Lüttge U (2002) Ответы циркадного ритма растений на термопериодические возмущения с асимметричными временными паттернами и скоростью изменения температуры.Biol Rhythm Res 33: 151–170. DOI: 10.1076 / brhm.33.2.151.1318
Google ученый
Bohn A, Hinderlich S, Hütt M-T, Kaiser F, Lüttge U (2003) Идентификация ритмических подсистем в циркадном цикле метаболизма крассулоидной кислоты в условиях термопериодических возмущений. Biol Chem 384: 721–728. DOI: 10.1515 / BC.2003.080
PubMed CAS Google ученый
Borchert R (2000) Органический и экологический контроль роста почек у тропических деревьев: В: Viemont JD, Crabbè J (eds) Покой растений: от поведения всего растения до клеточного контроля.CAB International Wallingford, pp 87–107
Borchert R, Rivera G (2001) Фотопериодический контроль сезонного развития и покоя тропических стволовых суккулентных деревьев. Tree Physiol 21: 213–221
PubMed CAS Google ученый
Borchert R, Renner SS, Calle Z, Navarrete D, Tye A, Gautier L, Spichiger R, von Hildebrand P (2005) Фотосинтетическая индукция синхронного цветения около экватора. Природа 433: 627–629.DOI: 10.1038 / nature03259
PubMed CAS Google ученый
Borchert R, Schlumpberger BO, Calle Z, Piedrahita L, Leftin A, Hammer SA, Tye A, Renner SS (2008) Сезонные колебания инсоляции — это время для синхронного цветения между экватором и 50 ° северной широты. Представленная рукопись на рецензирование, сделана доступной для личного общения
Boxall SF, Foster JM, Bohnert HJ, Cushman JC, Nimmo HG, Hartwell J (2005) Сохранение и дивергенция работы циркадных часов у вызываемых стрессом видов кислотного метаболизма Crassulacean показывает часовую компенсацию стресса.Физиология растений 137: 969–982. DOI: 10.1104 / стр.104.054577
PubMed CAS Google ученый
Бриффа К.Р., Бартолин Т.С., Экштейн Д., Джонс П.Д., Карлен В., Швайнгрубер Ф.Х., Зеттерберг П. (1990) 1400-летний рекорд летних температур в Фенноскандии за 1400 лет. Природа 346: 434–439. DOI: 10.1038 / 346434a0
Google ученый
Бриффа К.Р., Джонс П.Д., Швайнгрубер Ф.Х., Шиятов С.Г., Кук Э.Р. (1995) Необычное тепло лета двадцатого века в 1000-летнем температурном рекорде из Сибири.Природа 376: 156–159. DOI: 10.1038 / 376156a0
CAS Google ученый
Bucci SJ, Scholz FG, Goldstein G, Meinzer FC, Sternberg L. da Sl (2003) Динамические изменения гидравлической проводимости черешков двух видов деревьев саванны: факторы и механизмы, способствующие повторному заполнению эмболизированных сосудов. Среда растительной клетки 26: 1633–1645. DOI: 10.1046 / j.0140-7791.2003.01082.x
Google ученый
Бьюкенен-Волластон V (1997) Молекулярная биология старения листьев.J Exp Bot 48: 181–199. DOI: 10.1093 / jxb / 48.2.181
Google ученый
Caldas LS, Lüttge U, Franco AC, Haridasan M (1997) Гелиотропизм листьев у Pterodon pubescens , древесного бобового растения из бразильского серрадо. Rev Bras Fisiol Vegetal 9: 1–7
Google ученый
Callado CH, da Silva Neto SJ, Scarano FR, Costa CG (2001) Периодичность годичных колец на некоторых подверженных наводнениям деревьях атлантического дождевого леса в Рио-де-Жанейро, Бразилия.Деревья (Берл) 15: 492–497
Google ученый
Coster C (1927) Zur Anatomie und Physiologie der Zuwachszonen und Jahresringbildung in den Tropen I. Ann Jard Buitenzorg 37: 49–161
Google ученый
Coster C (1928) Zur Anatomie und Physiologie der Zuwachszonen und Jahresringbildung in den Tropen II. Энн Джард Буйтензорг 38: 1–114
Google ученый
Coté GG, DePass AL, Quarmby LM, Tate BF, Morse MJ, Satter RL, Crain RC (1989) Разделение и характеристика фосфолипидов инозитола из пульвины Samanea saman .Физиология растений 90: 1422–1428. DOI: 10.1104 / стр.90.4.1422
PubMed Google ученый
De Mairan J (1729) Ботаническое наблюдение. В: Histoire de l’Académie Royale des Sciences, Acad Roy Sci, Paris, pp. 35–36
De Mattos EA, Lobo PC, Joly CA (2002) Ночные дожди, вызывающие быстрые изменения в фотосинтетическом поведении у серрадовых древесных пород. во время засухи в сезон дождей.Aust J Bot 50: 241–246. DOI: 10.1071 / BT01023
Google ученый
Denne MP (1971) Температура и развитие трахеид у сеянцев Pinus sylvestris . J Exp Bot 22: 362–370. DOI: 10.1093 / jxb / 22.2.362
Google ученый
Денн М.П., Додд Р.С. (1981) Экологический контроль дифференцировки ксилемы.В: Barnett JR (ed) Развитие ксилемных клеток. Castle House, Кент, стр. 236–255
Google ученый
Dodd AN, Salathia N, Hall A, Kevei E, Toth R, Nagy F, Hibberd JM, Millar AJ, Webb AAR (2005) Циркадные часы растений увеличивают фотосинтез, рост, выживаемость и конкурентное преимущество. Наука 309: 630–633. DOI: 10.1126 / science.1115581
PubMed CAS Google ученый
Domec J-C, Scholz FG, Bucci SJ, Meinzer FC, Goldstein G, Villalobos-Vega R (2006) Суточные и сезонные колебания эмболии корневой ксилемы у древесных видов неотропической саванны: влияние на устьичный контроль водного статуса растений.Среда растительной клетки 29: 26–35. DOI: 10.1111 / j.1365-3040.2005.01397.x
PubMed CAS Google ученый
Доре Дж. (1959) Реакция риса на небольшие различия в продолжительности дня. Nature 183: 413–414. DOI: 10.1038 / 183413a0
Google ученый
Duarte HM, Lüttge U (2007) Циркадная ритмичность. В: Lüttge U (ed) Clusia .Древесный неотропический род удивительной пластичности и разнообразия. Экологические исследования, том 194. Springer, Berlin, pp. 245–256
Google ученый
Dünisch O, Montóia VR, Bauch J (2003) Дендроэкологические исследования Swietenia macrophylla King и Cedrela odorata L. (Melicaceae) в центральной части Амазонки. Деревья (Берл) 17: 244–250
Google ученый
Dupouey J-L, Leavitt S, Choisnel E, Jourdain S (1993) Моделирование фракционирования изотопов углерода в годичных кольцах на основе эффективной эвапотранспирации и состояния воды в почве.Среда растительной клетки 16: 939–947. DOI: 10.1111 / j.1365-3040.1993.tb00517.x
CAS Google ученый
Экштейн Д. (2004) Изменения в окружающей среде прошлого — секреты гидросистемы деревьев. Новый Фитол. 163: 1–4. DOI: 10.1111 / j.1469-8137.2004.01117.x
Google ученый
Edwards TDW, Graf W, Trimborn P, Stichler W., Lipp J, Payer HD (2000) δ 13 C Поверхность отклика разрешает сигналы влажности и температуры в деревьях.Geochim Cosmochim Acta 64: 161–167. DOI: 10.1016 / S0016-7037 (99) 00289-6
CAS Google ученый
Ehleringer J, Forseth I (1980) Отслеживание солнечной энергии растениями. Наука 210: 1094–1098. DOI: 10.1126 / science.210.4474.1094
PubMed Google ученый
Фонти П., Гарсия-Гонсалес I (2004) Пригодность хронологии сосудов из ранней древесины каштана для экологических исследований.Новый Фитол. 163: 77–86. DOI: 10.1111 / j.1469-8137.2004.01089.x
Google ученый
Фаулер С.Г., Кук Д., Томашоу М.Ф. (2005) Низкотемпературная индукция Arabidopsis CBF1, 2 и 3 контролируется циркадными часами. Физиология растений 137: 961–968. DOI: 10.1104 / стр.104.058354
PubMed CAS Google ученый
Franco AC, Lüttge U (2002) Полуденная депрессия в деревьях саванны: скоординированные корректировки фотохимической эффективности, фотодыхания, ассимиляции CO 2 и эффективности использования воды.Oecologia 131: 356–365. DOI: 10.1007 / s00442-002-0903-y
Google ученый
Fritts HC (1976) Годовые кольца и климат. Academic Press, Лондон
Google ученый
Gourlay ID (1995) Характеристики колец роста некоторых африканских видов Acacia . J Trop Ecol 11: 121–140
Артикул Google ученый
Heide OM (1974) Рост и покой у экотипов ели европейской ( Picea abies ).I. Взаимодействие фотопериода и температуры. Physiol Plant 30: 1–12. DOI: 10.1111 / j.1399-3054.1974.tb04983.x
Google ученый
Helle G, Schleser GH (2004) Beyond CO 2 — фиксация Рубиско — интерпретация вариаций углерода 13 C / 12 C в годичных кольцах из новых внутрисезонных исследований широколиственных деревьев. Среда растительной клетки 27: 367–380. DOI: 10.1111 / j.0016-8025.2003.01159.Икс
CAS Google ученый
Hillman WS, Koukari WL (1967) Фитохромные эффекты в никтинастических движениях листьев Albizzia julibrissin и некоторых других бобовых. Физиология растений 42: 1413–1418. DOI: 10.1104 / стр.42.10.1413
PubMed CAS Google ученый
Hütt M-T, Lüttge U (2005) Сетевая динамика в биологии растений: текущий прогресс в исторической перспективе.Prog Bot 66: 277–310. DOI: 10.1007 / 3-540-27043-4_12
Google ученый
Johnsen Ø, Dæhlen OG, Østreng G, Skrøppa T (2005a) Продолжительность светового дня и температура во время производства семян интерактивно влияют на адаптивные характеристики потомства Picea abies . Новый Фитол. 168: 589–596. DOI: 10.1111 / j.1469-8137.2005.01538.x
PubMed Google ученый
Johnsen Ø, Fossdal CG, Nagy N, Mølmann J, Dæhlen OG, Skrøppa T (2005b) Климатическая адаптация в потомстве Picea abies зависит от температуры во время зиготического эмбриогенеза и созревания семян.Среда растительной клетки 28: 1090–1102. DOI: 10.1111 / j.1365-3040.2005.01356.x
CAS Google ученый
Johnson CH, Golden SS (1999) Циркадные программы у цианобактерий: адаптивность и механизмы. Annu Rev Microbiol 53: 389-409. DOI: 10.1146 / annurev.micro.53.1.389
PubMed CAS Google ученый
Jordan BR (ed) (2006) Молекулярная биология и биотехнология цветения, 2-е изд.CAB International, Wallingford
Junttila O (1976) Прекращение апикального роста и опадание кончика побега в Salix . Physiol Plant 38: 278–286. DOI: 10.1111 / j.1399-3054.1976.tb04004.x
CAS Google ученый
Junttila O (1980) Влияние фотопериода и температуры на прекращение апикального роста у двух экотипов: Salix и Betula . Physiol Plant 48: 347–352.DOI: 10.1111 / j.1399-3054.1980.tb03266.x
Google ученый
Junttila O (2007) Регулирование годового цикла роста побегов северных древесных пород. В: Taulavuori E, Taulavuori K (eds) Физиология северных планов в условиях изменяющейся окружающей среды. Указатель исследований, Керала, стр. 177–210
Google ученый
Keskitalo J, Bergquist G, Gardeström P, Jansson S (2005) Клеточный график осеннего старения.Физиология растений 139: 1635–1648. DOI: 10.1104 / стр.105.066845
PubMed CAS Google ученый
Киенаст Ф., Швайнгрубер Ф.Х., Брекер О.Ю., Шер Э. (1987) Исследования годичных колец на хвойных деревьях вдоль экологических градиентов и возможности однолетних анализов. Can J For Res 17: 683–696. DOI: 10.1139 / x87-111
Google ученый
Kim HJ, Coté GG, Crain RC (1992) Влияние света на мембранный потенциал протопластов из Samanea saman pulvini.Вовлечение каналов K + и H + -ATPase. Физиология растений 99: 1532–1539. DOI: 10.1104 / стр.99.4.1532
PubMed CAS Google ученый
Kim HJ, Coté GG, Crain RC (1993) Калиевые каналы в протопластах Samanea saman , контролируемые фитохромом и биологическими часами. Наука 260: 960–962. DOI: 10.1126 / science.260.5110.960
PubMed CAS Google ученый
Kim HJ, Coté GG, Crain RC (1996) Инозитол 1, 4, 5-трифосфат может опосредовать закрытие каналов K + светом и темнотой в моторных клетках Samanea saman .Planta 198: 279–287. DOI: 10.1007 / BF00206254
PubMed CAS Google ученый
Коллер Д. (1990) Легкие движения листа. Среда растительной клетки 13: 615–632. DOI: 10.1111 / j.1365-3040.1990.tb01079.x
Google ученый
Косова К., Витамвас П., Прашил ИТ (2007) Роль дегидринов в реакции растений на холод.Биол Завод 51: 601–617. DOI: 10.1007 / s10535-007-0133-6
Google ученый
Lee Y, Satter RJ (1987) H + — захват и высвобождение во время циркадных ритмических движений иссеченных двигательных органов Samanea . Физиология растений 83: 856–862. DOI: 10.1104 / стр.83.4.856
PubMed CAS Google ученый
Lee Y, Satter RJ (1989) Влияние белого, синего и красного света и темноты на pH апопласта Samanea pulvinus.Планта 178: 31–40. DOI: 10.1007 / BF00392524
Google ученый
Ли Д.У., О’Кил Дж., Холбрук Н.М., Фейлд Т.С. (2003) Динамика пигментации и осеннее старение листьев в лиственных лесах Новой Англии на востоке США. Ecol Res 18: 677–694. DOI: 10.1111 / j.1440-1703.2003.00588.x
CAS Google ученый
Lim PO, Kim HJ, Nam HG (2007) Старение листьев.Анну Рев Плант Биол 58: 115–136. DOI: 10.1146 / annurev.arplant.57.032905.105316
PubMed CAS Google ученый
Lipavská H, Svobodová H, Albrechtová J (2001) Годовая динамика содержания неструктурных сахаридов в контексте структурного развития вегетативных почек ели европейской. J Plant Physiol 157: 365–373
Google ученый
Лю И, Цинореманс Н.Ф., Джонсон С.Х., Лебедека Н.В., Голден С.С., Ишиура М., Кондо Т.И. (1995) Циркадная оркестровка экспрессии генов у цианобактерий.Genes Dev 9: 1469–1478. DOI: 10.1101 / gad.9.12.1469
PubMed CAS Google ученый
Ллойд Д. (2006) Ультрадианные ритмы и часы у растений и дрожжей. Biol Rhythm Res 37: 281–296. DOI: 10.1080 / 092
CAS Google ученый
Ллойд Д., Мюррей Д. Б. (2005) Ультрадианский метроном: хронометрист для оркестровки клеточной когерентности.Trends Biochem Sci 30: 373–377. DOI: 10.1016 / j.tibs.2005.05.005
PubMed CAS Google ученый
Lloyd D, Murray DB (2007) Редокс-ритмичность: часы в основе временной согласованности. Биологические исследования 29: 465–473. DOI: 10.1002 / bies.20575
PubMed CAS Google ученый
Lüttge U (2000) Тонопласт, функционирующий как главный переключатель для суточной регуляции метаболизма крассулоидной кислоты.Planta 211: 761–769. DOI: 10.1007 / s004250000408
PubMed Google ученый
Lüttge U (2003a) Циркадная ритмичность: «биологические часы» — это аппаратное или программное обеспечение? Prog Bot 64: 277–319
Google ученый
Lüttge U (2003b) Циркадные ритмы. В: Томас Б., Мерфи Д. Д., Мюррей Б. Г. (ред.) Энциклопедия прикладных наук о растениях. Elsevier, Academic Press, Амстердам, стр. 1084–1096
Google ученый
Lüttge U (2004) Экофизиология метаболизма крассуловой кислоты (CAM).Энн Бот (Лондон) 94: 629–652. DOI: 10.1093 / aob / mch087
Google ученый
Lüttge U (2006) Фотосинтетическая гибкость и экофизиологическая пластичность: вопросы и уроки из рода Clusia , единственного дерева САМ в неотропиках. Новый Фитол. 171: 7–25. DOI: 10.1111 / j.1469-8137.2006.01755.x
PubMed Google ученый
Lüttge U (ed 2007a) Clusia .Древесный неотропический род удивительной пластичности и разнообразия. Экологические исследования, том 194. Springer, Berlin
Lüttge U (2007b) Фотосинтез. В: Lüttge U (ed) Clusia . Древесный неотропический род удивительной пластичности и разнообразия. Экологические исследования, том 194. Springer, Berlin, pp. 135–186
Google ученый
Lüttge U (2008a) Stem CAM у древесных суккулентов. Деревья (Берл) 22: 139–148.DOI: 10.1007 / s00468-007-0198-z
Google ученый
Lüttge U (2008b) Clusia : Святой Грааль и загадка. J Exp Bot 59: 1503–1514. DOI: 10.1093 / jxb / ern006
PubMed Google ученый
Lüttge U, Beck F (1992) Эндогенные ритмы и хаос в метаболизме крассулоидной кислоты. Планта 188: 28–38. DOI: 10.1007 / BF01160709
Google ученый
Lüttge U, Higinbotham N (1979) Транспорт в растениях. Springer, New York
Lüttge U, Klauke B, Griffiths H, Smith JAC, Stimmel K-H (1986) Сравнительная экофизиология CAM и C 3 бромелий. V. Газообмен и листовая структура бромелии C 3 Pitcairnia integrifolia. Среда растительных клеток 9: 411–419. DOI: 10.1111 / j.1365-3040.1986.tb01754.x
Matile P (1992) Старение хлоропластов. В: Бейкер Н.Р., Томас Х. (ред.) Фотосинтез сельскохозяйственных культур: пространственные и временные детерминанты. Elsevier, Амстердам, стр. 413–440
Google ученый
Matile P, Winkenbach F (1971) Функция лизосом и лизосомальных ферментов в стареющем венчике ипомеи ( Ipomoea purpurea ). J Exp Bot 22: 759–771.DOI: 10.1093 / jxb / 22.4.759
CAS Google ученый
McClung CR (2000) Циркадные ритмы у растений: взгляд на тысячелетие. Physiol Plant 109: 359–371. DOI: 10.1034 / j.1399-3054.2000.100401.x
CAS Google ученый
Medina E (1982) Физиологическая экология неотропических растений саванны. В: Хантлз Б.Дж., Уокер Б.Х. (ред.) Экологические исследования, том 42: экология тропических саванн.Springer, Berlin, pp. 308–335
Google ученый
Michael TP, McClung CR (2003) Захват энхансера выявляет широко распространенный контроль транскрипции циркадных часов у Arabidopsis . Физиология растений 132: 629–639. DOI: 10.1104 / pp.021006
PubMed CAS Google ученый
Mølmann JA, Junttila O, Johnsen Ø, Olsen JE (2006) Влияние красного, дальнего красного и синего света на поддержание роста широтных популяций ели европейской ( Picea abies ).Среда растительной клетки 29: 166–172. DOI: 10.1111 / j.1365-3040.2005.01408.x
PubMed Google ученый
Moran N, Ehrenstein G, Iwasa K, Mischke C, Bare C, Satter RL (1988) Калиевые каналы в моторных клетках Samanea saman. Исследование патч-зажима. Физиология растений 88: 643–648. DOI: 10.1104 / стр.88.3.643
PubMed Google ученый
Moshelion M, Moran N (2000) Каналы оттока калия в разгибательных и сгибательных клетках двигательного органа Samanea saman не идентичны.Эффекты цитозольного кальция. Физиология растений 124: 911–919. DOI: 10.1104 / стр.124.2.911
PubMed CAS Google ученый
Moshelion N, Becker D, Biela A, Uehlein N, Hedrich R, Otto B, Levi H, Moran N, Kaldenhoff R (2002) Аквапорины плазматической мембраны в моторных клетках Samanea saman : суточная и циркадная регуляция . Растительная клетка 14: 727–739. DOI: 10.1105 / tpc.010351
PubMed CAS Google ученый
Nimmo HG (2000) Регулирование фосфоенолпируваткарбоксилазы у растений САМ.Trends Plant Sci 5: 75–80. DOI: 10.1016 / S1360-1385 (99) 01543-5
PubMed CAS Google ученый
Нджоку Э. (1963) Сезонная периодичность роста и развития некоторых лесных деревьев в Нигерии. Дж. Экол 51: 617–624. DOI: 10.2307 / 2257750
Google ученый
Olsen JE, Junttila O (2002) Обработка дальним красным светом в конце дня восстанавливает длину растения дикого типа у гибридной осины со сверхэкспрессией фитохрома А.Physiol Plant 115: 448–457. DOI: 10.1034 / j.1399-3054.2002.1150315.x
PubMed CAS Google ученый
Ouyang Y, Andersson CR, Kondo T, Golden SS, Johnson CH (1998) Резонирующие циркадные часы улучшают приспособленность цианобактерий. Proc Natl Acad Sci USA 95: 8660–8664. DOI: 10.1073 / pnas.95.15.8660
PubMed CAS Google ученый
Патре У., Синха А.К., Ширке П.А., Сане П.В. (1998) Факторы, определяющие полуденную депрессию фотосинтеза у деревьев в условиях муссонного климата.Деревья (Берл) 12: 472–481. DOI: 10.1007 / s004680050177
Google ученый
Penfield S (2008) Восприятие температуры и передача сигналов в растениях. Новый Фитол. 179: 615–628. DOI: 10.1111 / j.1469-8137.2008.02478.x
PubMed CAS Google ученый
Pfeffer W (1907) Untersuchungen über die Entstehung der Schlafbewegungen der Blattorgane.Abh Math Phys Kl Kgl Sächs Ges Wiss 30 III: 259–472
Google ученый
Pfeffer W (1915) Beiträge zur Kenntnis der Entstehung der Schlafbewegungen. Abh Math Phys Kl Kgl Sächs Ges Wiss 34 I: 1–154
Google ученый
Puhakainen T, Li C, Boije-Malm M, Kangasjärvi J, Heino P, Palva ET (2004) Короткодневное усиление индуцированной низкой температурой генной экспрессии гена, контролируемого фактором связывания C-повторов, во время холода акклиматизация у березы повислой.Физиология растений 136: 4299–4307. DOI: 10.1104 / стр.104.047258
PubMed CAS Google ученый
Racusen RH, Satter RL (1975) Ритмические и регулируемые фитохромом изменения трансмембранного потенциала у Samanea pulvini. Природа 225: 408–410. DOI: 10.1038 / 255408a0
Google ученый
Рамос А., Перес-Солис Е., Ибаньес С., Касадо Р., Коллада С., Гомес Л., Арагонсилло С., Аллона I (2005) Зимнее нарушение циркадных часов у каштана.Proc Natl Acad Sci USA 102: 7037–7042. DOI: 10.1073 / pnas.0408549102
PubMed CAS Google ученый
Райх П.Б., Борхерт Р. (1984) Водный стресс и фенология деревьев в тропическом сухом лесу в низинах Коста-Рики. Дж. Экол 72: 61–74. DOI: 10.2307 / 2260006
Google ученый
Renner SS (2007) Синхронное цветение связано с изменениями интенсивности солнечного излучения.Новый Фитол 175: 195–197. DOI: 10.1111 / j.1469-8137.2007.02132.x
PubMed Google ученый
Rinne P, Welling A, Kaikuranta P (1998) Начало толерантности к замораживанию у березы ( Betula pubescens, Ehrh.) Связано с белками LEA и осморегуляцией и нарушено в генотипе с дефицитом ABA. Среда растительной клетки 21: 601–611. DOI: 10.1046 / j.1365-3040.1998.00306.x
Ривера Г., Борхерт Р. (2001) Индукция цветения тропических деревьев за счет 30-минутного сокращения фотопериода: данные полевых наблюдений и сборов гербария.Tree Physiol 21: 201–212
PubMed CAS Google ученый
Rivera G, Elliott S, Caldas LS, Nicolossi G, Coradin VTR, Borchert R (2002) Увеличение продолжительности светового дня вызывает весеннее смывание тропических сухих лесных деревьев в отсутствие дождя. Деревья (Берл) 16: 445–456. DOI: 10.1007 / s00468-002-0185-3
Google ученый
Rohde A, Howe GT, Olsen JE, Moritz T., van Mongtagu M, Junttila O, Boerjan W. (2000) Молекулярные аспекты покоя почек у деревьев.В: Jain SM, Minocha SC (ред.) Молекулярная биология древесных растений, том 1. Kluwer, Dordrecht, pp 89–134
Google ученый
Rolland F, Baena-Gonzalez E, Sheen J (2006) Чувствительность к сахару и передача сигналов в растениях: консервативные и новые механизмы. Анну Рев Завод Биол 57: 675–709. DOI: 10.1146 / annurev.arplant.57.032905.105441
PubMed CAS Google ученый
Ruttink T, Arend M, Morreel K, Storme V, Rombauts S, Fromm J, Bhalerao RP, Boerjan W, Rohde A (2007) Молекулярный график формирования верхушечной почки и индукции покоя у тополя.Растительная клетка 19: 2370–2390. DOI: 10.1105 / tpc.107.052811
PubMed CAS Google ученый
Sarmiento G (1984) Экология неотропических саванн. Издательство Гарвардского университета, Кембридж
Google ученый
Саттер Р.Л., Галстон А.В. (1973) Движение листьев: Розеттский камень поведения растений. Биология 23: 407–416. DOI: 10.2307 / 1296540
CAS Google ученый
Саттер Р.Л., Сабинс Д.Д., Галстон А.В. (1970a) Никтинастия, контролируемая фитохромом, в Albizzia julibrissin. I. Анатомия и тонкое строение пульвинулы. Am J Bot 57: 374–381. DOI: 10.2307 / 2440864
Google ученый
Satter RL, Marinoff P, Galston AW (1970b) Никтинастия, контролируемая фитохромом, в Albizzia julibrissin. II. Потоки калия как основа движения листочков. Am J Bot 57: 916–926. DOI: 10.2307 / 2440989
CAS Google ученый
Satter RL, Applewhite PB, Kreis DJ, Galston AW (1973) Ритмичный листовой механизм из Albizzia julibrissin. Действие электролитов и изменение температуры. Физиология растений 52: 202–207. DOI: 10.1104 / стр.52.3.202
PubMed Google ученый
Satter RL, Applewhite PB, Galston AW (1974a) Ритмический поток калия в Albizzia . Влияние аминофиллина, катионов и ингибиторов дыхания. Физиология растений 54: 280–285. DOI: 10.1104 / pp.54.3.280
PubMed CAS Google ученый
Satter RL, Geballe GT, Galston AW (1974b) Поток калия и движение листьев в Samanea saman .I. Ритмическое движение. J Gen Physiol 64: 413–430. DOI: 10.1085 / jgp.64.4.413
PubMed CAS Google ученый
Satter RL, Geballe GT, Applewhite PB, Galston AW (1974c) Поток калия и движение листьев в Samanea saman. II. Фитохромное движение. J Gen Physiol 64: 424–431. DOI: 10.1085 / jgp.64.4.431
Google ученый
Satter RL, Schrempf M, Chaudri J, Galston AW (1977) Фитохромные и циркадные часы в Samanea. Ритмичное перераспределение калия и хлорида в половом мозге в течение долгих темных периодов. Физиология растений 59: 231–235. DOI: 10.1104 / стр.59.2.231
PubMed CAS Google ученый
Satter RL, Xu Y, DePass A (1987) Влияние температуры на секрецию H + и поглощение вырезанными сгибательными клетками во время индуцированного темнотой закрытия листочков Samanea . Физиология растений 85: 850–855.DOI: 10.1104 / стр.85.3.850
PubMed CAS Google ученый
Schulze E-D, Lange OL, Evenari M, Kappen L, Buschbom U (1974) Роль влажности воздуха и температуры листьев в контроле устьичной устойчивости Prunus armeniaca L. в условиях пустыни. I. Моделирование ежедневного курса устьичного сопротивления. Oecologia 17: 159–170. DOI: 10.1007 / BF00346278
Google ученый
Schulze E-D, Lange OL, Kappen L, Evenari M, Buschbom U (1975a) Роль влажности воздуха и температуры листьев в контроле устойчивости устьиц Prunus armeniaca в условиях пустыни.II. Значение состояния воды в листьях и внутренней концентрации углекислого газа. Oecologia 18: 219–233. DOI: 10.1007 / BF00345424
Google ученый
Schulze E-D, Lange OL, Evenari M, Kappen L, Buschbom U (1975b) Роль влажности воздуха и температуры листьев в контроле устьичной устойчивости Prunus armeniaca L. в условиях пустыни. III. Влияние на эффективность использования воды. Oecologia 19: 303–314
Google ученый
Schweingruber FH (1988) Годовые кольца.Основы и приложения дендрохронологии. Kluwer, Dordrecht
Google ученый
Staiger D (2002) Циркадные ритмы у арабидопсиса : время для ядерных белков. Планта 214: 334–344. DOI: 10.1007 / s004250100662
PubMed CAS Google ученый
Staiger D, Heintzen C (1999) Циркадная система Arabidopsis thaliana .Хронобиол Int 16: 1–16. DOI: 10.3109 / 07420529
PubMed CAS Google ученый
Suh S, Moran N, Lee Y (2000) Синий свет активирует каналы оттока калия в сгибательных клетках из двигательных органов Samanea saman с помощью двух механизмов. Физиология растений 123: 833–843. DOI: 10.1104 / стр.123.3.833
PubMed CAS Google ученый
Sutinen M-L (2007) Липидные изменения во время акклиматизации к холоду и деакклимации травянистых и древесных видов.В: Taulavuori E, Taulavuori K (eds) Физиология северных планов в условиях изменяющейся окружающей среды. Указатель исследований, Керала, стр. 19–37
Google ученый
Takata N, Saito S, Saito CT, Nanjo T., Shinohara K, Uemura M (2009) Молекулярная филогения и экспрессия генов циркадных часов тополя, LHY1 и LHY2 . Новый Фитол. 181: 808–819. DOI: 10.1111 / j.1469-8137.2008.02714.x
CAS Google ученый
Taulavuori E, Lüttge U (2007) Мембранные реакции при стрессе в связи с сезонной динамикой окружающей среды в северных экосистемах.В: Taulavuori E, Taulavuori K (eds) Физиология северных планов в условиях изменяющейся окружающей среды. Указатель исследований, Керала, стр. 1–17
Google ученый
Таулавуори К., Сарала М., Таулавуори Э. (2009) Реакция роста на изменение световой среды. Prog Bot 71
Taylor JE, Whitelaw CA (2001) Отсутствующие сигналы. Новый Фитол. 151: 323–340. DOI: 10.1046 / j.0028-646x.2001.00194.x
CAS Google ученый
Tenhunen JD, Lange OL, Braun M, Meyer A, Lösch R, Pereira JS (1980) Закрытие устьиц в полдень у листьев Arbutus unedo в естественной маккиии в смоделированных условиях среды обитания в экологической камере.Oecologia 47: 365–367. DOI: 10.1007 / BF00398530
Google ученый
Tenhunen JD, Lange OL, Braun M (1981) Полуденное закрытие устьиц у склерофиллов средиземноморского типа в смоделированных условиях среды обитания в экологической камере. II. Влияние пары температуры листа и влажности воздуха на газообмен Arbutus unedo и Quercus ilex . Oecologia 50: 5–11. DOI: 10.1007 / BF00378788
Google ученый
Tenhunen JD, Lange OL, Gebel J, Beyschlag W, Weber JA (1984) Изменения фотосинтетической способности, эффективности карбоксилирования и точки компенсации CO 2 — точка компенсации, связанная с полуденным закрытием устьиц и полуденной депрессией CO 2 обмен листьев Quercus suber .Планта 162: 193–203. DOI: 10.1007 / BF00397440
CAS Google ученый
Томас Б., Винс-Прю Д. (1997) Фотопериодизм у растений. Academic Press, Сан-Диего
Google ученый
Verheyden A, Helle G, Schleser GH, Dehairs F, Beeckman H, Koedam N (2004a) Годовая цикличность в соотношениях стабильных изотопов углерода и кислорода высокого разрешения в древесине мангрового дерева Rhizophora mucronata .Среда растительной клетки 27: 1525–1536. DOI: 10.1111 / j.1365-3040.2004.01258.x
Google ученый
Verheyden A, Kairo JG, Beeckman H, Koedam N (2004b) Годовые кольца, формирование годичных колец и определение возраста в мангровых зарослях Rhizophora mucronata . Энн Бот (Лондон) 94: 59–66. DOI: 10.1093 / aob / mch215
Google ученый
Верхейден А., де Риддер Ф., Шмитц Н., Бекман Х., Коедам Н. (2005) Временные ряды с высоким разрешением плотности судов в кенийских мангровых деревьях показывают связь с климатом.Новый Фитол. 167: 425–435. DOI: 10.1111 / j.1469-8137.2005.01415.x
PubMed Google ученый
Войцех О., Зейтц С.Б., Илиев Д., Миттаг М. (2008) Обе субъединицы циркадного РНК-связывающего белка CHLAMY1 могут объединять информацию о температуре. Физиология растений 147: 2179–2193. DOI: 10.1104 / стр.108.118570
PubMed CAS Google ученый
Weiser CJ (1970) Холодостойкость и травмы древесных растений.Наука 169: 1269–1278. DOI: 10.1126 / science.169.3952.1269
PubMed Google ученый
Welling A, Palva ET (2006) Молекулярный контроль акклиматизации деревьев к холоду. Physiol Plant 127: 167–181. DOI: 10.1111 / j.1399-3054.2006.00672.x
CAS Google ученый
Welling A, Palva ET (2008) Участие факторов транскрипции CBF в зимостойкости березы.Физиология растений 147: 1199–1211. DOI: 10.1104 / стр.108.117812
PubMed CAS Google ученый
Веллинг А., Кайкуранта П., Ринне П. (1997) Фотопериодическая индукция покоя и толерантности к замерзанию у Betula pubescens : участие АБК и дегидринов. Physiol Plant 100: 119–125. DOI: 10.1111 / j.1399-3054.1997.tb03461.x
CAS Google ученый
Welling A, Moritz T, Palva ET, Junttila O (2002) Независимая активация холодовой акклиматизации с помощью низкой температуры и короткого светового периода у гибридных осин.Физиология растений 129: 1633–1641. DOI: 10.1104 / стр.003814
PubMed CAS Google ученый
Welling A, Rinne P, Vihera-Aarnio A, Kontunen-Soppela S, Heino P, Palva ET (2004) Фотопериод и температура по-разному регулируют экспрессию двух генов дегидрина во время перезимовки березы ( Betula pubescens Ehrh. ). J Exp Bot 55: 507–516. DOI: 10.1093 / jxb / erh045
Google ученый
Wilkins MB (1992) Циркадные ритмы: их происхождение и контроль.Новый Фитол. 121: 347–375. DOI: 10.1111 / j.1469-8137.1992.tb02936.x
CAS Google ученый
Wimmer R (2002) Анатомические особенности древесины в годичных кольцах как индикаторы изменений окружающей среды. Дендрохронология 20: 21–36. DOI: 10.1078 / 1125-7865-00005
Google ученый
Woodcock DW (1989) Климатическая чувствительность анатомических характеристик древесины в кольцевидном дубе ( Quercus macrocarpa ).Can J For Res 19: 639–644. DOI: 10.1139 / x89-100
Google ученый
Worbes M (1988) Разнообразие в структуре годовых зон роста у Tabebuia barbata (E. Mey) Sandw., Bignoniaceae, тропических пород деревьев из затопляемых лесов Центральной Амазонки. Дендрохронология 6: 71–89
Google ученый
Worbes M (1989) Годовые кольца, прирост и возраст деревьев в затопляемых лесах, саваннах и горных лесах в Неотропах.IAWA J 10: 109–122
Google ученый
Worbes M (1999) Годовые кольца роста, рост в зависимости от количества осадков и долгосрочные модели роста тропических деревьев из лесного заповедника Капаро в Венесуэле. Дж. Экол 87: 391–403. DOI: 10.1046 / j.1365-2745.1999.00361.x
Google ученый
Wright JS (1991) Сезонная засуха и фенология подлеска кустарников в тропическом влажном лесу.Экология 72: 1643–1657. DOI: 10.2307 / 1940964
Google ученый
Wright JS (1996) Фенологические реакции на сезонность у растений тропических лесов. В: Mulkey SS, Chazdon RC, Smith AP (eds) Экофизиология тропических лесных растений. Chapman and Hall, New York, pp 440–460
Google ученый
Ясуэ К., Фунада Р., Кобаяши О., Отани Дж. (2000) Влияние размеров трахеиды на вариации максимальной плотности Picea glehnii и взаимосвязь климатических факторов.Деревья (Берл) 14: 223–229
Google ученый
Йанг Х-Й (2007a) Синхронное цветение каучукового дерева ( Hevea brasiliensis ), вызванное высокой интенсивностью солнечного излучения. Новый Фитол. 175: 283–289. DOI: 10.1111 / j.1469-8137.2007.02089.x
PubMed Google ученый
Йанг Х-Й (2007b) Солнечный свет синхронного цветения в тропиках: каучуковое дерево как модель исследования.Новый Фитол 176: 730–735. DOI: 10.1111 / j.1469-8137.2007.02258.x
PubMed Google ученый
Zucker Lowen C, Satter RL (1989) Световые изменения в апопластической активности K + в Samanea saman pulvinus, отслеживаемые с помощью микроэлектродов с жидкой мембраной. Planta 179: 421–427. DOI: 10.1007 / BF00397580
Google ученый
Суточные и годовые ритмы деревьев
Деревья, многолетние фанерофиты, демонстрируют богатое разнообразие ритмических явлений.Это связано либо с исключительной экологической увлечение или из-за функционирования эндогенных осцилляторов независимо от окружающей среды. Оба типа ритмов рассматривается в этом обзоре. Ритмы, управляемые чисто средой, можно рассматривать как прелюдию к эндогенным ритмам. Среда Обсуждаемые контролируемые ритмы: (i) суточные ритмы никтинастических и гелиотропных движений листьев и колебательные явления фотосинтеза, таких как полуденная депрессия и метаболизм крассулообразной кислоты (CAM), и (ii) годовые ритмы годовых образование годичных колец, осеннее старение листьев, механизмы перезимовки и цветение.Среди суточных ритмов никтинастические Движения и CAM также являются свободными эндогенными ритмами, показывающими работу циркадных часов на деревьях. При старении листьев При зимовке и контроле за цветением задействовано определение фотопериода, что предполагает участие эндогенных часов. Возникает вопрос, связаны ли суточные и годовые ритмы механически. Очевидно, в основе фенологических явлений на фотопериодизм (в зависимости от измерения продолжительности ночи) координируются системой фитохромов и циркадные часы и многие аспекты годового развития и зимовки связаны с фотопериодизмом.Существование в деревьях гены циркадных часов, которые, как известно, закреплены в геноме A. thaliana, можно оценить путем попыток сопоставления с секвенированным геномом Populus или путем выделения клонов кДНК с деревьев для проверки их соответствия геному A. thaliana. На крайних широтах около экватора и к северу от полярного круга деревья также демонстрируют фенологические свойства, не зависящие от фотопериода. явления. В полярной области может быть задействовано полное излучение красного и дальнего красного света, и сигнальный путь затем включает фитохром и, таким образом, все еще может быть похожим на фотопериодизм.На экваторе общая дневная освещенность получение или восприятие динамики суточных изменений солнечной освещенности имеют важное значение, и остается загадкой, является ли сигнализация каскады либо прикреплены к циркадным часам пока неизвестным образом, либо полностью независимы от циркадных часов.
Суточные и сезонные молекулярные ритмы в неокортексе человека и их связь с болезнью Альцгеймера
Участники исследования
В это исследование были включены данные участников двух продолжающихся продольных когортных исследований пожилых людей: Rush MAP и ROS.ROS — это продольное исследование старения католических братьев, монахинь и священников со всех концов Соединенных Штатов 50 . MAP — это исследование старения на уровне сообществ в районе Большого Чикаго 51 . Участники обоих исследований не страдали известной деменцией на момент включения в исследование и соглашаются на ежегодные оценки и донорство мозга в случае смерти. На момент проведения текущего анализа 680 участников ROS и 726 участников MAP умерли, и им было проведено вскрытие. Поскольку все участники ROS и MAP являются донорами органов, дата и время смерти, как правило, хорошо фиксируются у подавляющего большинства участников.Мы исключили из дальнейшего анализа данные из 215 образцов дорсолатеральной префронтальной коры, где время или дата смерти или другие клинические ковариантные данные (см. Ниже) не были доступны.
Из 1191 образца, для которого были доступны эти данные, данные по экспрессии РНК дорсолатеральной префронтальной коры, соответствующие критериям контроля качества (см. Ниже), были доступны из 531 образца, данные метилирования ДНК, соответствующие критериям контроля качества (см. Ниже), были доступны из 732 образцов и Критерии контроля качества прохождения данных h4K9Ac ChIP-seq (см. ниже) были доступны из 664 образцов.Данные этих участников были включены в текущий анализ. Их характеристики показаны в Таблице 1, а совпадение этих наборов показано на Рис. 1.
Заявление об одобрении этических норм
Это исследование было одобрено институциональным наблюдательным советом Медицинского центра Университета Раша и проводилось в соответствии с последними рекомендациями. версия Хельсинкской декларации. Все участники предоставили письменное информированное согласие и Акт анатомического подарка на донорство органов.
Оценка экспрессии транскрипта
РНК экстрагировали из блоков дорсолатеральной префронтальной коры с помощью набора miRNeasy Mini (Qiagen, Венло, Нидерланды) и набора ДНКаз, не содержащих РНКаз (Qiagen).Концентрацию РНК количественно определяли с помощью Nanodrop (Thermo Fisher Scientific, Уолтем, Массачусетс, США), а для оценки качества использовали биоанализатор Agilent. Образцы, из которых было получено <5 мкг РНК, или образцы с оценкой целостности РНК Bioanalyser 5 или меньше, были исключены из дальнейшего анализа. Специфичный для цепи метод dUTP 52 с отбором поли-A 53 был использован платформой геномики Broad Institute для подготовки библиотеки RNA-seq. За отбором поли-A следовал синтез кДНК, специфичный для первой цепи, с использованием dUTP для синтеза кДНК, специфичной для второй цепи, с последующей фрагментацией и лигированием адаптера Illumina для конструирования библиотеки.Аппарат Illumina HiSeq использовался для выполнения секвенирования со считыванием парных концов 101 п.н., достигнув покрытия 150 млн считываний для первых 12 образцов, которые служили эталоном для глубокого охвата. Остальные образцы были секвенированы с охватом 50 млн считываний. Затем начальные и конечные низкокачественные основания и последовательности адаптеров были вырезаны из считанных значений, а считанные рибосомные РНК были удалены. Программный пакет Bowtie 1 54 использовался для выравнивания обрезанных считываний с эталонным геномом. Наконец, программный пакет RSEM был использован для оценки в единицах фрагментов на килобазу на миллион картированных фрагментов (FPKM) уровней экспрессии для 55 889 отдельных генов GENCODE v14 и 190 051 отдельных изоформ GENCODE v14.Эти данные доступны на портале знаний Synapse.org AMP-AD (http://www.synapse.org; SynapseID syn3388564). Мы проанализировали только гены и транскрипты на аутосомных хромосомах и исключили гены и транскрипты, экспрессируемые в <90% наших образцов, оставив 18 709 индивидуальных генов GENCODE v14 и 42 873 индивидуальных изоформ GENCODE v14 в этих анализах. Все значения FPKM были преобразованы в логарифм перед дальнейшим анализом. Для оценки качества образцов использовался анализ главных компонентов, и только те образцы, у которых значения главных компонентов 1, 2 и 3 были в пределах 3 с.d. из их соответствующих средств были включены. В этот анализ были включены данные из 531 образца, отвечающего критериям контроля качества, как указано выше, и с полными клиническими данными, как описано ниже.
Оценка метилирования ДНК
Мы оценили метилирование ДНК в 746 образцах дорсолатеральной префронтальной коры, как описано ранее 27,28 . Замороженные 100 мг блоки дорсолатеральной префронтальной коры размораживали на льду и вручную препарировали серое вещество и использовали мини-набор QIAamp DNA (Qiagen) для извлечения ДНК.Набор Quant-iT PicoGreen Kit (Life Technologies, Карлсбад, Калифорния, США) использовали для измерения концентрации ДНК. Анализ с использованием микрочипов Illumina Infinium HumanMethylation450k Bead Chip Chip (Illumina) использовали Genomics Platform Института Броуда (Кембридж, Массачусетс, США) для анализа 16 мкл ДНК из каждого образца в концентрации 50 нг мкл -1 . Модуль Methylation Module v.1.8 из пакета программного обеспечения Genome Studio (Illumina) использовался для выполнения нормализации цветовых каналов и удаления фона, а также для генерации значений β и определения значений P для 485 513 сайтов CpG в геноме человека.Эти данные доступны на портале знаний Synapse.org AMP-AD (http://www.synapse.org SynapseID syn3157275). Зонды с обнаружением P имеют значения> 0,01 в любых образцах, с 47/50 нуклеотидами, соответствующими последовательностям половых хромосом во время выравнивания последовательности с BLAT, в которых однонуклеотидный полиморфизм с частотой минорных аллелей ≥0,01 существует в пределах 10 п.н. сайт CpG или половая хромосома были исключены из дальнейшего анализа. Это оставило 420 132 аутосомных CpG в наборе данных.Для оценки качества образцов использовался анализ главных компонентов, и только те образцы, у которых значения главных компонентов 1, 2 и 3 были в пределах 3 s.d. из их соответствующих средств были включены. Также были исключены образцы, в которых по меньшей мере 2 из 10 датчиков контроля превращения бисульфита не смогли достичь значения 0,8. В эти анализы были включены данные из 732 образцов, отвечающих критериям контроля качества, как указано выше, и с полными клиническими данными, как описано ниже.
Оценка h4K9Ac
Серое вещество отделяли на льду от блоков дорсолатеральной префронтальной коры, измельчали и поперечно сшивали 1% формальдегидом при комнатной температуре в течение 15 минут и гасили 0.125 М глицин. После гомогенизации ткани в буфере для лизиса клеток с использованием Tissue Lyser и шарика из нержавеющей стали 5 мм, ядра лизировали в буфере для лизиса клеток и хроматин подвергали срезанию ультразвуком. Хроматин инкубировали в течение ночи при 4 ° C с моноклональными антителами Millipore против h4K9Ac (номер по каталогу 06-942, номер партии: 31636) и очищали с помощью гранул сефарозы протеина А. Наконец, экстрагированную ДНК использовали для конструирования библиотеки Illumina в соответствии с обычными методами и секвенировали с односторонним считыванием 36 п.н. на Illumina HiSeq.Считывания были сопоставлены алгоритмом BWA с эталонным геномом человека GRCh47 (ref. 55 55). MACS2 применялся для обнаружения пиков для каждого образца отдельно с использованием опции широкого пика и порогового значения q , равного 0,001 (ссылка 56 56). Объединенную геномную ДНК семи образцов использовали в качестве отрицательного контроля. Эти данные доступны на портале знаний Synapse.org AMP-AD (http://www.synapse.org SynapseID syn4896408). Для удаления образцов низкого качества использовались пять фильтров различного качества 57 : (i) ≥15 × 10 6 уникальных считываний, (ii) неизбыточная доля ≥0.3, (iii) взаимная корреляция ≥0,03, (iv) доля считываний в пиках ≥0,05 и (v) ≥6000 пиков. После контроля качества осталось 669 из 712 проб. Домены h4K9Ac были определены путем расчета всех областей генома, которые были обнаружены в виде пиков по крайней мере в 100 (15%) из наших 669 образцов. Области, расположенные в пределах 100 п.н., были объединены, и очень маленькие области <100 п.н. были удалены. Всего было получено 26 384 домена h4K9Ac. Подмножество из 25 740 этих доменов было расположено на аутосомных хромосомах и использовалось для последующих анализов.Чтобы количественно определить h4K9Ac, мы подсчитали количество чтений, попадающих в каждый домен h4K9Ac для каждой выборки, разделенное на ширину каждого домена в килобазах и на общее количество отображенных чтений в каждой выборке, и масштабировали это, чтобы получить единицы FPKM. Перед анализом значения FPKM были преобразованы в логарифмическую форму. В эти анализы были включены данные из 664 образцов, отвечающих критериям контроля качества, как указано выше, и с полными клиническими данными, как описано ниже.
Оценка клинических ковариат
Мы вычислили возраст смерти по дате рождения, сообщенной самим пациентом, и дате смерти.Мы записали секс во время базового интервью.
Время восхода солнца в день смерти вычислялось по зарегистрированной дате смерти, а также широте и долготе города, в котором умер каждый участник.
Симптомы депрессии оценивались с помощью 10-балльной версии шкалы депрессии Центра эпидемиологических исследований 51 .
Патология болезни Альцгеймера определялась количественно, как описано ранее. 58,59 . Нейрофибриллярные клубки, диффузные бляшки и нейритные бляшки визуализировались окрашиванием серебром по Бильшовскому на срезах лобной, височной, теменной и энторинальной коры и гиппокампа.Как описано в предыдущих публикациях 59 , непрерывный глобальный показатель общего бремени патологии болезни Альцгеймера был рассчитан путем количественного определения наивысшей плотности каждого из нейрофибриллярных клубков, диффузных бляшек и нейритных бляшек на 1 мм 2 в срезах от лобной части. , височной, теменной, гиппокампальной и энторинальной коры каждого участника, масштабируя эти значения с помощью sd всех участников, а затем усреднение по четырем областям мозга и трем патологиям (клубки, диффузные и нейритные бляшки) для получения итоговой оценки.Как описано ранее 58 , для категориального патологического диагноза болезни Альцгеймера случаи классифицировались как отсутствие болезни Альцгеймера, болезнь Альцгеймера с низкой вероятностью, болезнь Альцгеймера средней вероятности или болезнь Альцгеймера с высокой вероятностью на основе критериев Национального института старения и Рейгана. 60 ; считалось, что участнику был поставлен патологический диагноз болезни Альцгеймера, если согласно классификации Национального института старения — Рейгана «средняя вероятность» или «высокая вероятность».
Статистический анализ суточной и сезонной ритмичности
Мы одновременно охарактеризовали независимые суточные и сезонные паттерны экспрессии каждого из 18709 генов с помощью функций косинуса, рассматривая каждый транскрипт как функцию даты смерти и времени смерти относительно восхода солнца на дата смерти (‘ZT’, которая отражает время воздействия естественного света), скорректированная с учетом возраста на момент смерти, пола, посмертного периода, бремени патологии болезни Альцгеймера, наличия / отсутствия депрессивных симптомов и технических ковариат. (партия, оценка целостности РНК для данных RNA-seq, примерная кросс-корреляция для данных h4K9Ac ChIP-seq) следующим образом:
, где t — это ZT, а d — дата смерти, A d — амплитуда суточной ритмичности, A s — амплитуда сезонной ритмичности, φ d — время акрофазы суточной ритмичности y и φ s — дата акрофазы сезонной ритмичности.Для этих анализов суточный период был установлен на 24 часа, а сезонный период — на 365 дней. Это ограничение любого дизайна исследования, когда каждый человек вносит только одну точку данных в каждый 24-часовой или 365-дневный период выборки. Все времена смерти были преобразованы в радианы (2 π радиан = 24 часа; 0 радиан = восход солнца) для анализа, а затем преобразованы обратно в часы для визуального представления. Даты смерти были аналогично преобразованы в радианы (2 π радиан = 365 дней; 0 радиан = 1 января).Во вторичных анализах мы повторили все анализы, используя два альтернативных эталонных времени: время по местным часам (с ZT0 = полночь), с которым связано время воздействия искусственного света, и середину между закатом и восходом солнца, которая неизменна в зависимости от времени года. Для вычислительной эффективности мы подбираем эквивалентные линеаризованные модели вида
и, A d , φ d , A s и φ s из уравнения (1) были рассчитаны с использованием формул
Чтобы количественно оценить вклад суточной ритмичности в соответствие модели в уравнении (2) для каждой расшифровки, независимо от эффектов сезонной ритмичности, мы сравнили остаточную сумму квадратов для уравнения (2) с суммой квадратов сокращенного модель без суточных условий
и определенная F -статистика
Чем больше вклад суточной ритмичности в общую подгонку модели, тем больше значение F d .Аналогичным образом мы количественно оценили вклад сезонной ритмичности в соответствие модели в уравнении (2) путем сравнения квадратов остаточных сумм для уравнения (2) с таковыми для сокращенной модели без сезонных членов и определили соответствующую статистику F . .
Затем мы повторили вышеуказанный анализ для каждой из 42 873 индивидуальных изоформ, 25 740 аутосомных пиков h4K9Ac и 420 132 сайтов метилирования аутосомной ДНК.
Мы создали список предположительно ритмичных генов, вычислив соответствующее значение P и установив порог P <0.05. Мы сравнили этот список предположительно суточных ритмических генов с тремя другими опубликованными наборами данных (данные дорсолатеральной префронтальной коры от Li et al , 18 и данные BA47 и BA11 от Chen et al 19 ), исследуя на обогащение с помощью теста χ 2 . По данным Li et al . 18 , для сравнения были доступны только данные из 50 наиболее ритмичных транскриптов, и мы рассматривали только транскрипты, идентифицированные как суточно ритмичные в дорсолатеральной префронтальной коре при P <0.05. По данным Chen et al . 19 , все транскрипты считались суточно ритмичными при P <0,05 в ВА11 или ВА47. Затем мы сравнили время пика экспрессии основных генов циркадных часов (как указано в GENECARDS 61 ) между нашими данными и этими двумя другими наборами данных, вычислив коэффициенты круговой корреляции.
Затем мы количественно оценили степень суточной ритмичности по всем 18 709 генам или 42 873 транскриптам, вычислив медианное значение F d для всех транскриптов в наблюдаемых данных ( F d_observed ).Чтобы вычислить эмпирическое значение P , мы сравнили его со средним значением F d по всем транскриптам в каждом из 10000 наборов нулевых данных ( F d_null ), сгенерированных случайным перемешиванием времени смерти, и определили доля наборов нулевых данных, для которых F d_null > F d_observed . Затем мы сделали то же самое для сезонной ритмичности, используя нулевые наборы данных, сгенерированные случайным перемешиванием дат, а не времен смерти.Затем мы повторили эти анализы для 25 740 пиков аутосомного h4K9Ac и 420 132 сайтов метилирования аутосомной ДНК.
Затем мы исследовали закономерности во времени суточных и сезонных ритмов экспрессии РНК. Путем визуального осмотра и в соответствии с предыдущей работой 19,26 суточное и сезонное время акрофазы стенограммы распределялись бимодально. Исходя из этого, мы использовали самоорганизующиеся карты с тороидальными сетками, чтобы эмпирически определить два дневных и два сезонных кластера и классифицировать каждый транскрипт в эти кластеры.Результирующие суточные кластеры примерно центрировались утром (∼ZT0) и вечером (∼ZT12), а результирующие сезонные кластеры примерно центрировались весной и осенью. Мы исследовали связь между суточной и сезонной классификацией, вычислив статистику χ 2 для соответствующей таблицы сопряженности 2 × 2, используя наблюдаемые данные ( χ 2 набл. ), сравнив ее с соответствующим χ 2 -статистика, вычисленная путем повторения этих процедур для 10000 наборов эмпирических нулевых данных, сгенерированных перетасовкой времени и дат смерти ( χ 2 null ) и вычисления эмпирического значения P путем определения доля 10000 наборов нулевых данных, в которых χ 2 null > χ 2 obs .Затем мы повторили этот анализ для данных метилирования h4K9Ac и ДНК, за исключением того, что для данных метилирования ДНК мы рассматривали время надира, а не акрофазу метилирования, поскольку гипометилирование, а не гиперметилирование, классически связано с транскрипцией. Наконец, в качестве анализа чувствительности мы повторили этот анализ, рассматривая только те транскрипты / пики / сайты, которые были как сезонно-ритмичными, так и суточно-ритмичными при P <0,05.
Ранее нами было показано, что для ритмов метилирования ДНК время суточной акрофазы изменяется в зависимости от расстояния от ближайшего TSS 27 .Мы исследовали, верно ли это также для суточных ритмов h4K9Ac, разделив пики h4K9Ac на две группы — те, которые находятся в пределах 2000 п.н. от активного TSS, и те, которые находятся на расстоянии более 2000 п.н. от такого TSS, где мы определили активный как соответствующий транскрипту, выраженному в более 90% наших образцов. Мы сравнили угловые распределения суточных акрофаз пиков h4K9Ac в этих двух группах в наблюдаемых данных, вычислив статистику Мардиа-Ватсона-Уиллера W ( W, d_observed ) и сравнили это с соответствующим W -статистика для каждого из 10000 наборов нулевых данных, сгенерированных случайным перемешиванием времен смерти ( W d_null ), и вычисленное эмпирическое значение P путем определения доли 10000 наборов нулевых данных, где W d_null > Вт d_обнаружено .Затем мы проделали то же самое для углового распределения сезонных акрофаз, используя нулевые наборы данных, полученные путем перетасовки дат смерти. Затем мы исследовали, различается ли классификация отдельных пиков на суточные / сезонные кластеры между двумя группами пиков h4K9Ac, вычислив статистику χ 2 , χ 2 , наблюдаемую , для соответствующих 4 × 2 таблицы непредвиденных обстоятельств (4 возможных сезонных / суточных класса × 2 группы пиков h4K9Ac), сравнивая это с соответствующей статистикой × 2 , полученной повторением вышеуказанной процедуры на 10000 наборов нулевых данных, сгенерированных перетасовкой времени и дат смерти и вычисление эмпирического значения P путем определения доли 10000 наборов нулевых данных, в которых χ 2 null > χ 2 наблюдаемых .Затем мы повторили описанную выше процедуру для сайтов метилирования ДНК. Наконец, в качестве анализа чувствительности мы повторили этот анализ, учитывая только те транскрипты / пики / сайты, которые были как сезонными, так и суточными ритмиками при P <0,05.
Считается, что и h4K9Ac, и метилирование ДНК влияют на транскрипцию. Таким образом, мы исследовали ассоциации между синхронизацией ритмов обилия транскриптов и синхронизацией ритмов h4K9Ac и метилирования ДНК в близлежащих сайтах. Сначала мы исследовали суточные ритмы h4K9Ac, используя 471 образец, в котором были доступны оба набора данных.
Мы рассмотрели две группы пиков h4K9Ac: те, которые находятся в пределах 2 т.п.н. от TSS транскриптов, эмпирически классифицированных как утренняя акрофаза, и в пределах 2 т.п.н. от TSS транскриптов, эмпирически классифицированных как вечерняя акрофаза, и сравнили угловые распределения их дневных акрофаз. используя статистику Мардиа-Ватсона-Уиллера W , как указано выше, вычисляя эмпирическое значение P путем сравнения его с эквивалентной статистикой W , рассчитанной на 10 000 нулевых наборов данных, сгенерированных случайным перемешиванием времен смерти.Затем мы определили, различались ли две группы в доле пиков h4K9Ac, классифицированных как утренние или вечерние, путем вычисления соответствующей статистики χ 2 и вычисления эмпирического значения P по сравнению с эквивалентным χ 2 -статистика рассчитана из 10 000 наборов нулевых данных, сгенерированных путем перемешивания времен смерти. После рассмотрения суточных ритмов h4K9Ac, как указано выше, мы затем повторили этот анализ с учетом сезонных ритмов.Затем мы повторили эти анализы для сайтов метилирования ДНК, используя 527 образцов с данными как РНК-seq, так и данными метилирования ДНК. Наконец, в качестве анализа чувствительности мы повторили этот анализ, рассматривая только те транскрипты / пики / сайты, которые были сезонно или суточно ритмичными при P <0,05.
Окружение локального фактора транскрипции оказывает важное влияние на циркадный ритм транскрипции в модельных системах 31 . Таким образом, мы исследовали ассоциацию между местными сайтами связывания факторов транскрипции во времени суточных и сезонных акрофаз / надиров экспрессии транскрипта, h4K9Ac и метилирования ДНК.Для этого мы использовали аннотированные сайты связывания по всему геному для 161 фактора транскрипции из проекта ENCODE 32,33,34 . Мы считали, что TSS, пик h4K9Ac или сайт метилирования ДНК локально связаны с фактором транскрипции, если он перекрывается с одним из его ENCODE-аннотированных сайтов связывания или находится в пределах 2000 п.н. Мы эмпирически классифицировали транскрипты, пики h4K9Ac и сайты метилирования ДНК на дневные и сезонные кластеры, как указано выше.
Затем мы использовали модели логистической регрессии формы
, чтобы изучить независимую связь локального присутствия сайтов связывания для каждого из 161 фактора транскрипции ENCODE с вероятностью наличия весеннего по сравнению с осенью или вечернего по сравнению с утренним транскриптом, пика h4K9Ac. или сайт метилирования ДНК.Мы оценили нескорректированные значения P и FDR для всего анализа путем сравнения оценок эффекта выше с оценками, полученными из 10 000 наборов нулевых данных, сгенерированных случайным перемешиванием времени или дат смерти. Мы определили набор факторов транскрипции кандидатов, потенциально участвующих в регуляции суточной и / или сезонной ритмичности, если их сайты связывания имели FDR <0,05 для одной из суточной или сезонной ритмичности для любого из метилирования РНК, h4K9Ac или ДНК. В качестве анализа чувствительности мы повторили эту процедуру, рассматривая только транскрипты, пики h4K9Ac или сайты метилирования ДНК с P <0.05 для суточной или сезонной ритмичности и рассчитанные коэффициенты корреляции Спирмена, относящиеся к оценкам эффекта транскрипционного фактора, оцененным на основе всех транскриптов / пиков h4K9Ac / сайтов метилирования ДНК, и тех, которые оцениваются на основе только этих транскриптов / пиков h4K9Ac / сайтов метилирования ДНК. с P <0,05 для суточной или сезонной ритмичности.
Затем, чтобы изучить общие регуляторные эффекты этих сайтов связывания транскрипционных факторов на дневную и сезонную ритмичность, мы построили их расчетные коэффициенты для суточной и сезонной ритмичности для каждого из экспрессии транскрипта, h4K9Ac и метилирования ДНК и рассчитали коэффициенты корреляции Спирмена.Затем, чтобы изучить общие регуляторные эффекты этих сайтов связывания транскрипционных факторов на транскрипт и эпигенетические суточные ритмы, мы рассчитали коэффициенты корреляции Спирмена, относящиеся к их влиянию на суточный транскрипт по сравнению с эпигенетическими ритмами (h4K9Ac и метилирование ДНК). Затем мы повторили это для сезонных ритмов.
Заболевания головного мозга, такие как болезнь Альцгеймера 35 , как было показано, влияют на физиологические маркеры циркадной ритмичности. Чтобы изучить влияние болезни Альцгеймера на амплитуду и время ритмов экспрессии транскрипта, h4K9Ac и метилирования ДНК, мы увеличили уравнение (4) следующим образом:
, где y 1 и y 2 — возраст при смерти и мужском сексе.
Здесь относительные амплитуды суточной и сезонной ритмичности у людей с болезнью Альцгеймера даны как
, а относительные амплитуды суточной и сезонной ритмичности у людей без болезни Альцгеймера даны как
Кроме того, акрофазы суточной и сезонной ритмичности у людей с болезнью Альцгеймера даны
, а акрофазы суточной и сезонной ритмичности у людей без болезни Альцгеймера даны
Чтобы графически изобразить связь между патологическим диагнозом болезни Альцгеймера и временем расшифровки ритмов, мы классифицировали каждый транскрипт в дневные и сезонные классы, как указано выше, на основе модели, предсказанной акрофазой для образцов без болезни Альцгеймера, и отдельно построены предсказанные моделью дневное и сезонное время акрофазы для людей с болезнью Альцгеймера.Для более формальной количественной оценки связи между патологическим диагнозом болезни Альцгеймера и синхронизацией ритмов транскриптов мы вычислили медианную разницу в прогнозируемом времени акрофазы между людьми с болезнью Альцгеймера и без нее, сначала рассматривая все транскрипты, а затем отдельно транскрипты в каждом из четыре временных класса. Эта процедура была повторена для 10 000 наборов нулевых данных, сгенерированных путем перетасовки времен смерти для получения эмпирических значений P .Затем мы повторили эту процедуру для сезонных акрофаз. Чтобы количественно оценить влияние болезни Альцгеймера на амплитуду ритмов транскриптов, мы рассчитали медианную разницу относительной амплитуды между людьми с болезнью Альцгеймера и без нее, сначала рассматривая все транскрипты, а затем отдельно рассматривая транскрипты в каждом из четырех временных кластеров. Эта процедура была повторена для 10 000 наборов нулевых данных, сгенерированных путем перетасовки времен смерти для получения эмпирических значений P .Затем мы повторили эту процедуру для относительной амплитуды сезонных ритмов. Затем в качестве анализа чувствительности мы повторили вышеизложенное, рассматривая только те транскрипты с индивидуальным P <0,05 как для суточной, так и для сезонной ритмичности. Наконец, мы повторили вышеуказанный анализ для пиков h4K9Ac и сайтов метилирования ДНК.
Доступность данных
Данные RNA-seq, которые подтверждают результаты этого исследования, были депонированы на портале знаний synapse.org AMP-AD с кодом доступа syn3388564 (https: // www.synapse.org/#!Synapse:syn3388564). Данные h4K9Ac ChIP-seq, которые подтверждают результаты этого исследования, были депонированы на портале знаний synapse.org AMP-AD с кодом доступа syn4896408 (https://www.synapse.org/#!Synapse:syn4896408). Данные о метилировании ДНК, подтверждающие результаты этого исследования, были депонированы на портале знаний synapse.org AMP-AD с кодом доступа syn3157275 (https://www.synapse.org/#!Synapse:syn3157275).
Суточные и годовые ритмы на деревьях
ПОКАЗЫВАЕТ 1-10 ИЗ 196 ССЫЛОК
СОРТИРОВАТЬ ПО РелевантностиСамые популярные статьи Недавность
Tansley Review No.37 Циркадные ритмы: их происхождение и контроль.
- M. Wilkins
- Медицина, биология
- Новый фитолог
- 1992
- Просмотреть 1 отрывок, справочная информация
Циркадные ритмы у растений: взгляд на тысячелетия
В настоящее время существует ряд многообещающих кандидатов на роль компонентов циркадных часов растений, и кажется вероятным, что авторы скоро узнают подробности о них. завод центральный генератор. Развернуть- Просмотреть 2 выдержки, справочная информация
Эндогенные ритмы и хаос в метаболизме крассулоидной кислоты
Эндогенные свободные регулярные циркадные колебания чистого обмена CO2 в растении крассуло-кислотного метаболизма (CAM) Kalanchoë daigremontiana de la et al. постоянный внешний… Развернуть
- Просмотреть 2 выдержки, ссылки на результаты и фон
Эндогенные ритмы и хаос в метаболизме кислоты крассулообразных
Эндогенные автономные регулярные циркадные колебания чистого обмена СО2 в растении, вызывающем метаболизм крассулеевых кислот (CAM) Kalanchoë daigremontiana Было показано, что постоянные внешние условия в непрерывном свете меняются на нерегулярные непредсказуемые временные характеристики, когда освещенность или температура поднимаются выше критического уровня.Развернуть- Просмотреть 1 отрывок, ссылки на результаты
Ультрадианные ритмы и часы у растений и дрожжей
Распространение субклеточных и клеточных функций на ткани растений обеспечивает временные рамки, в которых коррелируются движения побегов и листьев, изменения развития и фотопериодические реакции и скоординировано. Развернуть- Просмотреть 1 отрывок, справочная информация
Обзор циркадных ритмов
Alcohol Res Health. 2001; 25 (2): 85–93.
Марта Хотц Витатерна, доктор философии, старший научный сотрудник Центра функциональной геномики Северо-Западного университета, Эванстон, Иллинойс. Джозеф С. Такахаши, доктор философии, является директором Центра функциональной геномики, профессором Уолтера и Мэри Э. Гласс на кафедре нейробиологии и физиологии, а также исследователем в Медицинском институте Говарда Хьюза Северо-Западного университета в Эванстоне. , Иллинойс. Фред В. Турек, доктор философии, является директором Центра биологии сна и циркадной биологии и является исследователем Чарльза Т.и Эмма Х. Моррисон, профессор кафедры нейробиологии и физиологии Северо-Западного университета, Эванстон, Иллинойс
Если в тексте не указано иное, все материалы, представленные в этом журнале, находятся в открытом доступе и могут воспроизводиться без разрешения. Цитирование источника приветствуется.
Эта статья цитируется в других статьях в PMC.Abstract
Суточный цикл свет-темнота определяет ритмические изменения в поведении и / или физиологии большинства видов.Исследования показали, что этими изменениями управляют биологические часы, которые у млекопитающих расположены в двух областях мозга, называемых супрахиазматическими ядрами. Циркадные циклы, установленные этими часами, происходят в природе и имеют период примерно 24 часа. Кроме того, эти циркадные циклы могут быть синхронизированы с внешними сигналами времени, но также могут сохраняться в отсутствие таких сигналов. Исследования показали, что внутренние часы состоят из множества генов и кодируемых ими белковых продуктов, которые регулируют различные физиологические процессы в организме.Нарушения биологических ритмов могут нанести вред здоровью и благополучию организма. Ключевые слова: циркадный ритм; время суток; биологическая регуляция; биологическая адаптация; температура; свет; гипоталамус; нервная клетка; экспрессия генов; мутагенез; расстройство сна; физиологический AODE (последствия употребления алкоголя или других наркотиков, злоупотребления и зависимости)
Одна из самых драматических особенностей мира, в котором мы живем, — это круговорот дня и ночи. Соответственно, почти все виды ежедневно изменяют свое поведение и / или физиологию.Эти суточные ритмы являются не просто реакцией на 24-часовые изменения в физической среде, вызванные вращением Земли вокруг своей оси, но, напротив, возникают из системы хронометража внутри организма. Эта система хронометража, или биологические «часы», позволяет организму предвидеть и подготовиться к изменениям в физической среде, которые связаны с днем и ночью, тем самым гарантируя, что организм будет «делать правильные вещи» в нужное время день. Биологические часы также обеспечивают внутреннюю временную организацию и гарантируют, что внутренние изменения происходят в координации друг с другом.
Синхронизация организма как с внешней, так и с внутренней средой имеет решающее значение для благополучия и выживания организма; отсутствие синхронизации между организмом и внешней средой может привести к немедленной смерти человека. Например, если ночной грызун выйдет из своей норы средь бела дня, он станет исключительно легкой добычей для других животных. Точно так же отсутствие синхронизации во внутренней среде может привести к проблемам со здоровьем у человека, например, связанным с сменой часовых поясов, сменной работой и сопутствующей потерей сна (например.g., нарушение когнитивной функции, нарушение гормональной функции и желудочно-кишечные жалобы).
Механизмы, лежащие в основе биологических систем хронометража, и потенциальные последствия их сбоев являются одними из вопросов, которыми занимаются исследователи в области хронобиологии. 1 В самом широком смысле хронобиология охватывает все области исследований, в которых основное внимание уделяется биологическому времени, включая высокочастотные циклы (например, секрецию гормонов, происходящую отдельными импульсами в течение дня), дневные циклы (напр.g., циклы активности и отдыха), а также месячные или годовые циклы (например, репродуктивные циклы у некоторых видов). Среди этих взаимосвязанных областей хронобиологии эта статья фокусируется на одной частотной области — суточных циклах, известных как циркадные ритмы. (Термин «циркадный ритм» происходит от латинского выражения «circa diem», что означает «около суток».) Хотя практически все формы жизни, включая бактерии, грибы, растения, плодовых мух, рыб, мышей и людей, имеют циркадные ритмы. ритмов, этот обзор в первую очередь ограничен системой млекопитающих.Другие животные обсуждаются только в тех случаях, когда они внесли свой вклад в понимание системы млекопитающих, особенно в исследованиях молекулярно-генетического состава системы хронометража. (Для сравнительного обсуждения других модельных систем, отличных от млекопитающих, которые внесли свой вклад в глубину понимания циркадной ритмики у млекопитающих, читателя отсылаем к Wager-Smith and Kay 2000.) В целом, эта статья преследует следующие основные цели: (1) предоставить высокоселективный исторический обзор области, (2) рассмотреть характерные свойства циркадных ритмов, (3) определить структурные компоненты и молекулярно-генетические механизмы, составляющие биологические часы, и (4) изучить влияние биологических часов на здоровье ритмы.
Исторический обзор хронобиологии
Исследователи начали изучать биологические ритмы примерно 50 лет назад. Хотя ни один эксперимент не является определяющим событием, с которого на сегодняшний день начались современные исследования в области хронобиологии, исследования, проведенные в 1950-х годах по циркадной ритмичности у плодовых мух Колином Питтендрием и у людей Юргеном Ашоффом, можно считать его основанием. Область исследования сна, которая также относится к области хронобиологии, развивалась до некоторой степени независимо, с определением различных стадий сна Натаниэлем Клейтманом примерно в то же время (Dement 2000).Наследие этих пионеров продолжается и сегодня в развитии полей, которые они основали.
Корни изучения биологических ритмов уходят еще дальше, к 1700-м годам и к работам французского ученого де Майрана, опубликовавшего монографию, описывающую ежедневные движения листьев растения. Де Майран заметил, что ежедневное поднятие и опускание листьев продолжалось даже тогда, когда растение было помещено во внутреннюю комнату и, таким образом, не подвергалось воздействию солнечного света. Это открытие предполагает, что движения представляют собой нечто большее, чем просто реакцию на солнце, и контролируются внутренними часами.
Характерные свойства циркадных ритмов
Удачные наблюдения де Майрана иллюстрируют одну важную особенность циркадных ритмов — их самоподдерживающуюся природу. Таким образом, почти все суточные ритмы, которые происходят в естественных условиях, продолжают циклически повторяться в лабораторных условиях, лишенных каких-либо внешних сигналов, дающих время из физической среды (например, при постоянном освещении или постоянной темноте). Циркадные ритмы, которые выражаются при отсутствии каких-либо 24-часовых сигналов из внешней среды, называются свободным ходом.Это означает, что ритм не синхронизируется каким-либо циклическим изменением физической среды. Строго говоря, суточный ритм не следует называть циркадным, пока не будет показано, что он сохраняется в постоянных условиях окружающей среды и, таким образом, не может быть отличен от тех ритмов, которые являются просто реакцией на 24-часовые изменения окружающей среды. Однако для практических целей нет оснований различать суточные и циркадные ритмы, потому что почти все суточные ритмы являются циркадными.Терминологическое различие между циркадными ритмами также не проводится на основе типа стимула окружающей среды, который синхронизирует цикл.
Устойчивость ритмов в отсутствие цикла темного-светлого или другого экзогенного сигнала времени (например, Zeitgeber) явно указывает на существование какого-то внутреннего механизма хронометража или биологических часов. Однако некоторые исследователи отметили, что постоянство ритмичности не обязательно исключает возможность того, что другие, неконтролируемые циклы, порождаемые вращением Земли вокруг своей оси, могут управлять ритмом (см. Aschoff 1960).
Гипотеза о том, что такие неконтролируемые геомагнитные сигналы могут играть роль в сохранении ритмичности, может быть опровергнута второй характерной чертой циркадных ритмов: эти циклы сохраняются с периодом около 24 часов, но не точно. Если ритмы были вызваны экзогенно, они должны сохраняться ровно 24 часа. Однако кажущаяся неточность — важная черта ритмичности. Как продемонстрировал Питтендрай (1960), отклонение от 24-часового цикла на самом деле дает возможность внутренней системе хронометрирования постоянно согласовываться и согласовываться со светлой-темной средой.Эта непрерывная регулировка приводит к большей точности в управлении синхронизацией или фазой выраженных ритмов, поскольку допускается небольшой дрейф до того, как ритм будет «сброшен» на правильную фазу.
Третьим характерным свойством циркадных ритмов является их способность синхронизироваться или увлекаться внешними сигналами времени, такими как цикл свет-темнота. Таким образом, хотя циркадные ритмы могут сохраняться в отсутствие внешних временных сигналов (что означает, что они не управляются окружающей средой), обычно такие сигналы присутствуют, и ритмы согласованы с ними.Соответственно, если происходит сдвиг внешних сигналов (например, после перемещения по часовым поясам), ритмы будут согласованы с новыми сигналами. Такое выравнивание называется увлечением.
Первоначально было неясно, было ли увлечение достигнуто за счет модуляции скорости цикла (т. Е. Был ли цикл сокращен или удлинен до тех пор, пока он не выровнялся с новыми сигналами, а затем вернулся к своей исходной длине), или же увлечение было достигнуто дискретным «Сброс» событий. Эксперименты, ставшие результатом этой дискуссии, привели к фундаментальным открытиям.Например, исследователи обнаружили, что реакция организма на свет (то есть, продвигается ли цикл, задерживается или остается неизменным) различается в зависимости от фазы цикла, на которой он представлен (Pittendrigh 1960). Таким образом, воздействие света в течение ранней части «нормального» темного периода человека обычно приводит к фазовой задержке, тогда как воздействие света в течение поздней части нормального темного периода человека обычно приводит к фазовому прогрессу. Эта разница в ответах может быть представлена кривой фазовой реакции (см. Схематическую иллюстрацию циркадного цикла, а также кривую фазовой реакции).Такая кривая может предсказать, каким образом организм будет увлекаться не только сдвигами в циклах свет-темнота, но также и необычными световыми циклами, такими как не 24-часовые циклы или другое соотношение свет: темнота. Существование кривой фазовой характеристики также подразумевает, что унос достигается за счет дискретных событий сброса, а не изменения скорости цикла.
Ответы циркадного ритма на свет.
В дополнение к времени воздействия света, интенсивность света может модулировать периоды цикла, когда организмы остаются в постоянном свете.Таким образом, воздействие более яркого света может удлинять период у одних видов и сокращать его у других. Это явление было названо «правилом Ашоффа» (Aschoff 1960). В конце концов, оба механизма увлечения кажутся аспектами одного и того же, потому что последствия правила Ашоффа можно предсказать или объяснить с помощью кривых фазового отклика на свет.
Хотя цикл свет-темнота явно является основным Zeitgeber для всех организмов, другие факторы, такие как социальные взаимодействия, активность или упражнения и даже температура, также могут влиять на фазу цикла.Влияние температуры на циркадные ритмы особенно интересно тем, что изменение температуры может влиять на фазу цикла без существенного изменения скорости цикла. Это означает, что цикл может начаться раньше или позже обычного, но при этом иметь ту же продолжительность. С одной стороны, способность кардиостимулятора внутренних часов компенсировать изменения температуры имеет решающее значение для его способности прогнозировать и адаптироваться к изменениям окружающей среды, потому что часы, которые ускоряются и замедляются при изменении температуры, бесполезны.С другой стороны, температурная компенсация также вызывает затруднения, поскольку большинство видов биологических процессов (например, биохимических реакций в организме) ускоряются или замедляются из-за изменений температуры. В конечном итоге эта загадка дала ключ к разгадке природы внутренних часов, то есть к тому факту, что циркадные ритмы имеют генетическую основу. Такая программа экспрессии генов будет более устойчивой к изменению температуры, чем, например, простая биохимическая реакция.
Два последних свойства циркадных ритмов также дают важные подсказки о структуре ритмов.Одно из этих свойств — повсеместность ритмов в природе: циркадные ритмы существуют в широком спектре биологических процессов и организмов со схожими свойствами и даже с похожими кривыми фазового отклика на свет. Другое свойство состоит в том, что циркадные ритмы, по-видимому, генерируются на клеточном уровне, потому что ритмы одноклеточных организмов (например, водорослей или динофлагелляты Gonyaulax ) во многом совпадают с ритмами очень сложных млекопитающих. Оба этих наблюдения предполагают, что цикл активации (т.е., экспрессия) определенных генов может лежать в основе механизма хронометража.
Анатомическая организация внутренних часов
Хотя исследования одноклеточных организмов указывают на клеточную природу системы, генерирующей циркадные ритмы, у высших организмов циркадный водитель ритма расположен в клетках определенных структур организма. Эти структуры включают определенные области мозга (например, зрительные и церебральные доли) у насекомых; глаза у некоторых беспозвоночных и позвоночных; и шишковидная железа, которая расположена в головном мозге, у позвоночных, не являющихся млекопитающими.У млекопитающих циркадные часы находятся в двух кластерах нервных клеток, называемых супрахиазматическими ядрами (SCN), которые расположены в области у основания мозга, называемой передним гипоталамусом.
Роль SCN была продемонстрирована знаменательным открытием в начале 1970-х годов, согласно которому, повреждая (то есть повреждая) SCN у крыс, исследователи могут нарушать и устранять эндокринные и поведенческие циркадные ритмы (обзор см. В Klein et al. 1991). Кроме того, трансплантируя SCN от других животных животным с поврежденным SCN, исследователи могли восстановить некоторые циркадные ритмы.Наконец, роль SCN как главного водителя ритма, регулирующего другие ритмические системы, была подтверждена аналогичными исследованиями на хомяках, которые продемонстрировали, что восстановленные ритмы проявляют свойства часов (то есть период или фазу ритма) донора, а не хозяин (Ральф и др., 1990). Открытие того, что SCN является местом первичной регуляции циркадной ритмики у млекопитающих, дало исследователям фокус для их исследования: если кто-то хочет понять 24-часовой хронометраж, нужно изучить часы в SCN.
Однако недавно исследователи с удивлением обнаружили, что циркадные ритмы могут сохраняться в изолированных легких, печени и других тканях, выращенных в культуральной чашке (т.е. in vitro), которые не находились под контролем SCN (Yamazaki et al. 2000). Эти наблюдения показывают, что большинство клеток и тканей тела могут быть способны модулировать свою активность на циркадной основе. Такие данные, однако, не умаляют центральной роли, которую играет SCN как главный циркадный кардиостимулятор, который каким-то образом координирует всю 24-часовую временную организацию клеток, тканей и всего организма.Физиологические механизмы, лежащие в основе этой координации, включают сигналы, излучаемые SCN, которые действуют на другие нервные клетки (т.е. нейронные сигналы) или которые также распространяются через кровь в другие органы (т.е. нейрогормональные сигналы). На сегодняшний день, однако, характеристики самого циркадного сигнала, то есть конкретный способ, которым SCN «разговаривает» с остальным телом, остаются неизвестными (см. Stokkan et al. 2001).
Хотя влияние поражений SCN на многочисленные ритмы было выяснено, их влияние на сон менее ясно.Таким образом, поражения SCN явно нарушают консолидацию и структуру сна у крыс, но оказывают лишь минимальное влияние на количество сна или потребность во сне животных (Mistlberger et al. 1987). По этой и другим причинам исследователи постулировали, что сон подчиняется двум по существу независимым механизмам контроля: (1) циркадные часы, которые модулируют склонность ко сну, и (2) гомеостатический контроль, отражающий продолжительность предыдущего бодрствования (т. Е. « недосыпание »). Однако недавно исследования на беличьих обезьянах показали, что поражения SCN могут влиять на продолжительность сна.Более того, исследования сна у мышей, несущих изменения (т. Е. Мутации) в двух генах, влияющих на циркадные циклы (т. Е. Гены DBP и Clock ), показали, что эти мутации привели к изменениям в регуляции сна (Naylor et al. 2000). ; Franken et al., 2000). Оба этих наблюдения открывают интригующую возможность того, что гомеостатический и циркадный контроль могут быть более взаимосвязаны, чем ранее думали исследователи.
Молекулярная генетика циркадных ритмов
Как обсуждалось ранее, свойства циркадных часов предполагают циклические изменения в экспрессии определенных генов как возможный механизм, лежащий в основе внутреннего водителя ритма.Эта гипотеза была подтверждена демонстрацией на ряде видов, что экспрессия генов и продукция белков, кодируемых этими генами, необходимы для нормальной работы часов. Тем не менее, совершенно другой экспериментальный подход в конечном итоге привел к идентификации компонентов молекулярных циркадных часов. Исследователи использовали химические вещества, чтобы ввести многочисленные случайные мутации в ДНК плодовой мушки Drosophila melanogaster и нитчатого гриба Neurospora .Затем полученные мутантные организмы проверяли на нарушения ритма. Этот подход к мутагенезу привел к идентификации первых мутантов циркадных часов, которые были названы периодом ( на ) и частотой ( frq , произносится как «урод»). Гены, несущие мутации в этих организмах, были клонированы в 1980-х годах (обзор см. В Wager-Smith and Kay 2000). Однако последовало значительное разочарование, когда исследователи попытались выделить эквивалентные гены у млекопитающих (т.е., гомологи млекопитающих). Наконец, в начале 1990-х исследователи начали аналогичный подход к скринингу мутагенеза у мышей и описали первую циркадную мутацию мыши, названную Clock , в 1994 году (см. King and Takahashi 2000). В 1997 году ген, затронутый этой мутацией, стал первым клонированным геном циркадных часов млекопитающих (King and Takahashi 2000). Как и мутанты генов Per и Frq , измененный ген Clock влияет на период свободного ритма (т.е., удлинили период) и вызвали потерю устойчивости циркадных ритмов в постоянных условиях окружающей среды. И мутант C lock у мышей, и мутант Per у мух были первыми животными их соответствующего вида, идентифицированными с использованием такого подхода мутагенеза, в котором мутация проявлялась как измененное поведение, а не измененный физиологический процесс.
С момента открытия гена Clock у мышей список генов циркадных часов, идентифицированных у млекопитающих, вырос за удивительно короткий период времени (см.).Например, исследователи идентифицировали не один, а три гена млекопитающих, которые соответствуют гену на как по своей структуре (то есть нуклеотидной последовательности), так и по их функции (King and Takahashi 2000; Lowrey and Takahashi 2000). Некоторые из предложенных генов циркадных часов были идентифицированы исключительно на основании их сходства в последовательности с генами часов Drosophila , и их функция часов не была подтверждена на основании изучения поведения соответствующих мутантов.Тем не менее, полученные к настоящему времени результаты четко указывают на схему кардиостимулятора, основанную на цикле обратной связи экспрессии генов (см.).
Схематическое изображение регуляции генов, которые, как считается, участвуют в циркадных часах. BMAL1, Clock, CK1 ɛ, mPer и mCry — все это гены циркадных часов, идентифицированные у мышей. (Существует несколько вариантов генов mPer и mCry .) В ядре клетки генетическая информация, закодированная в этих генах, преобразуется в молекулу-носитель, называемую мРНК (черные волнистые линии), которая транспортируется в жидкость внутри ячейка (т.э., цитоплазма). Там мРНК используется для генерации белковых продуктов, кодируемых генами циркадных часов (кружки и овалы с цветами, соответствующими соответствующим генам). Некоторые из этих белков регулируют активность определенных генов часов, связываясь с «молекулярными переключателями» (то есть E-блоками), расположенными перед этими генами. Это называется циклом обратной связи. Таким образом, BMAL1 и белки часов способствуют активации генов Per и mCry , тогда как белки Per ингибируют активацию этих генов.24-часовой цикл происходит, когда белки BMAL1 и Clock вызывают повышенное производство белков Per и Cry . По мере накопления Pers и Crys они подавляют собственный синтез, и уровень белка снижается. Белок CK1ɛ также помогает регулировать уровни белка Clock , дестабилизируя белок Per .
ПРИМЕЧАНИЕ: BMAL1 = мозговой и мышечный ARNT-подобный 1; CK1ɛ = казеинкиназа 1 эпсилон; mPer = период мыши; mCry = криптохром мыши.
Таблица 1
Гены циркадных часов млекопитающих; Соответствующие гены плодовой мухи, Drosophila ; и влияние изменений (то есть мутаций) этих генов на поведение (т.е. фенотип) пораженных животных
| Ген мыши | Псевдоним | Drosophila Ген | Мутантный | * Часы | dClock | Увеличенный период; потеря стойкой ритмичности в постоянных условиях |
|---|---|---|---|
| mPer1 | период | Уменьшение амплитуды, укороченный период или потеря ритма | |
| * mPer16 | mP1666 Укороченный период, потеря ритма|||
| * мПер3 | период | Умеренное сокращение периода | |
| * CKIɛ | au1663 915 x15 Укороченный период у мутантов хомяков | ||
| * mCry1 mCry2 | dcry | Животные, лишенные гена mCry1 (i.е., mCry1 нокаутов) имеют укороченный период; mCry2 нокаутов имеют удлиненный период; животные, у которых отсутствуют оба гена (т.е. двойной нокаут), имеют потерю ритма | |
| * BMAL1 | MOP3 | цикл | Потеря ритма |
| ? mTim | вне времени | Роль у млекопитающих не ясна | |
| ? ДАД | Умеренное удлинение периода |
Важность циркадных часов для здоровья и благополучия человека
Практически все физиологические и поведенческие функции человека происходят на ритмической основе, что, в свою очередь, приводит к резким суточным ритмам в работоспособности человека.Независимо от того, является ли оно результатом добровольных (например, посменная работа или быстрое перемещение через часовые пояса) или непроизвольных (например, болезнь или преклонный возраст) обстоятельств, нарушенная циркадная ритмика у людей связана с различными психическими и физическими расстройствами и может отрицательно влияют на безопасность, производительность и производительность. Фактически, многие побочные эффекты нарушенной циркадной ритмичности могут быть связаны с нарушениями цикла сна и бодрствования. На некоторые ритмические процессы в большей степени влияют циркадные часы, чем состояние сна-бодрствования, тогда как другие ритмы в большей степени зависят от состояния сна-бодрствования.
Для большинства животных время сна и бодрствования в естественных условиях синхронно с циркадным контролем цикла сна и всеми другими циркадными ритмами. Однако люди обладают уникальной способностью когнитивно игнорировать свои внутренние биологические часы и их ритмические выходы. Когда цикл сна и бодрствования не совпадает по фазе с ритмами, которые контролируются циркадными часами (например, во время сменной работы или быстрого перемещения по часовым поясам), могут возникнуть неблагоприятные эффекты.
Помимо нарушений сна, связанных с сменой часовых поясов или сменной работой, нарушения сна могут возникать по многим другим известным и неизвестным причинам. И хотя нарушение сна является отличительной чертой многих психических и физиологических расстройств человека, особенно аффективных расстройств, часто неясно, способствуют ли нарушения сна заболеванию или являются его результатом. Другие нарушения циркадного ритма также часто связаны с различными болезненными состояниями, хотя опять же важность этих нарушений ритма в развитии (т.д., этиология) болезни остается неизвестной (Brunello et al. 2000).
Одним из важных факторов, влияющих на неспособность исследователей точно определить роль циркадных нарушений в различных болезненных состояниях, может быть отсутствие знаний о том, как циркадные сигналы от SCN передаются в ткани-мишени. Для дальнейшего выяснения регуляции циркадных ритмов исследователям необходимо лучше понять природу циркадного сигнала, выводимого из SCN, и того, как эти выходные сигналы могут быть изменены, когда они достигают своих целевых систем.Такое расширенное понимание также позволит лучше разграничить важность нормальной временной организации для здоровья и болезней человека. Обнаружение того, что две основные причины смерти — сердечные приступы и инсульты — имеют вариацию во времени суток, является показательным примером. Если бы ученые знали больше о механизмах, ответственных за ритмичность этих нарушений, они могли бы определить более рациональные терапевтические стратегии, чтобы повлиять на эти события. Наконец, учитывая, что с возрастом в системе циркадных часов происходят драматические изменения, эти изменения могут лежать в основе или, по крайней мере, усугублять возрастное ухудшение физических и умственных способностей пожилых людей.
Выводы
Хотя исследователи только за последние несколько лет добились больших успехов в понимании молекулярной основы циркадной ритмичности, этот прогресс основан на обширных исследованиях, проведенных во многих лабораториях за последние 50 лет. В тот же период другие исследователи в многочисленных лабораториях выяснили критическую роль, которую SCN играет в регуляции циркадной ритмики у млекопитающих и, возможно, других позвоночных. (Для получения дополнительной информации об этих выводах и их значении читатель может обратиться к различным ресурсам в Интернете, некоторые из которых перечислены в.)
Таблица 2
Хронобиологические ресурсы в Интернете
Большинство животных довольны тем, что подчиняются своему SCN и позволяют ему управлять выражением множества циркадных ритмов. Однако люди обладают собственным разумом и часто используют его, чтобы не подчиняться своим «внутренним часам» — например, с растущей тенденцией к круглосуточной доступности для бизнеса. Потенциальные последствия такого все более и более круглосуточного дежурства по вызову на данный момент неизвестны, но доказательства не предвещают ничего хорошего.
Сейчас перед исследователями и клиницистами стоит задача определить не только причину, но и последствия для здоровья человека и болезней нарушений временной организации циркадной системы. Эти вопросы также включают вопрос о том, какую роль алкоголь может играть в нарушении нормальных циркадных ритмов и биологических часов. Этот вопрос более подробно рассматривается в этом специальном выпуске Alcohol Research & Health . Доктора Василевски и Холлоуэй рассматривают способы взаимодействия алкоголя и циркадного ритма организма, используя температуру тела как показатель функции циркадного ритма.Цикл сна-бодрствования, который составляет, в частности, центральный аспект циркадных ритмов, может быть изменен алкоголем; Влияние алкоголя на сон неалкоголиков и алкоголиков обсуждается доктором. Roehrs и Roth и доктором Брауэром, соответственно.
Как указано в этой статье, нарушение нормальной циркадной ритмики может привести к серьезным последствиям для здоровья, включая психические расстройства, такие как депрессия. В то же время психоактивные препараты, такие как антидепрессанты, также обладают хронобиологическим действием.Доктор Розенвассер исследует эти ассоциации и обсуждает влияние алкоголя на модели депрессии у людей и животных. Другие влияния алкоголя на биологические часы могут быть даже более тонкими и оставаться довольно спекулятивными, например, последствия пренатального воздействия алкоголя, которые обсуждаются доктором. Эрнест, Чен и Уэст. Наконец, не только потребление алкоголя может влиять на циркадные ритмы, но и циркадные факторы, такие как цикл свет-темнота, также могут влиять на потребление алкоголя. Эту тему обсуждают д-р.Хиллер-Штурмхёфель и Кулькоски. Вместе эти статьи предлагают читателям взглянуть на интересные и сложные взаимодействия, которые существуют между алкоголем и циркадными ритмами, которые во многом определяют поведение и благополучие всех организмов, включая человека.
ГЛОССАРИЙ
Каждая научная область имеет свою специфическую терминологию; научная область биологических ритмов и сна — не исключение. В этом глоссарии определены некоторые термины, с которыми читатели могут встретиться в этой статье и в специальном выпуске Alcohol Research & Health.
| Хронобиология | Раздел биологии, связанный с определением времени биологических событий, особенно повторяющихся или циклических явлений, в отдельных организмах. |
| Циркад | Термин, образованный от латинского словосочетания «около дня», что означает «около дня»; относится к биологическим вариациям или ритмам с циклом приблизительно 24 часа. Циркадные ритмы являются самоподдерживающимися (т. Е. свободно работают ), что означает, что они будут сохраняться, когда организм помещен в среду, лишенную временных сигналов, таких как постоянный свет или постоянная темнота.Для сравнения см. Суточный , инфрадиан и ультрадиан . |
| Циркадное время (CT) | Стандартизированная 24-часовая запись фазы в циркадном цикле, которая представляет собой оценку субъективного времени организма. CT 0 указывает начало субъективного дня, а CT 12 — начало субъективной ночи. Например, для ночного грызуна начало субъективной ночи (т.е. СТ 12) начинается с начала активности, тогда как для дневного вида СТ 0 будет началом активности.Для сравнения см. Zeitgeber time . |
| DD | Обычное обозначение среды, находящейся в непрерывной темноте (в отличие от цикла свет-темнота). Для сравнения см. LD . |
| Дневной | Зависит от времени суток. Суточные ритмы могут сохраняться, когда организм помещен в среду, лишенную временных сигналов, таких как постоянный свет или постоянная темнота. Следовательно, суточные колебания могут управляться светом или часами.Для сравнения см. циркад. . |
| Entrainment | Процесс синхронизации механизма хронометража с окружающей средой, например, с циклом свет-темнота или LD. Для сравнения см. в автономном режиме . |
| Free running | Состояние организма (или ритм) при отсутствии каких-либо увлекающих стимулов. Обычно испытуемых держат в постоянном тусклом свете или в постоянной темноте, чтобы оценить их ритмы свободного движения.Для сравнения см. захват . |
| Инфрадиан | Термин, образованный от латинской фразы «Infra diem», что означает «меньше суток»; относится к биологическим циклам, которые длятся более 1 дня и, следовательно, имеют частоту менее одного в день. Для сравнения см. циркадный и ультрадиан . |
| LD | Условные обозначения для цикла окружающей среды свет-темнота; количество часов светлого и темного времени обычно разделяется двоеточием.Например, LD 16: 8 обозначает цикл, состоящий из 16 часов света и 8 часов темноты. Для сравнения см. DD . |
| Маскировка | Затенение «истинного» состояния ритма условиями, которые препятствуют его обычному выражению. Обычно фаза увлеченного ритма или отсутствие увлечения (например, у животного, которое не может увлечься из-за какого-либо дефекта) маскируется световым циклом. Например, отвращение ночного грызуна к яркому свету приводит к тому, что начало его активности совпадает с отсутствием света или «выключением света», когда животное фактически бодрствует в течение нескольких часов.Для сравнения см. захват . |
| Сон с небыстрым движением глаз (NREM) | Стадии сна, которые включают «более глубокие» стадии сна, в которых сновидения обычно не возникают. Также называется медленным сном. Для сравнения см. сон с быстрым движением глаз . |
| Phase shift | Изменение фазы ритма. Это изменение можно измерить, наблюдая за изменением времени опорной точки фазы (например,, начало активности или ночное повышение высвобождения гормона мелатонина) от времени, ожидаемого на основании предыдущих циклов, выполняемых без перерыва. Фазовые сдвиги могут быть либо опережающими (т. Е. Опорная точка фазы возникает раньше, чем обычно), либо задержками (т. Е. Опорная точка фазы наступает позже, чем обычно). |
| Кривая фазового отклика (PRC) | Графическая сводка фазовых сдвигов , вызванных конкретной манипуляцией, такой как световой импульс или фармакологическое лечение, в зависимости от фазы (т.е.е., суточное время ), в которое происходит манипуляция. Определение PRC для света позволило исследователям понять и предсказать, как осуществляется увлечение световыми циклами. |
| Сон с быстрым движением глаз (REM) | Стадия легкого сна, характеризующаяся быстрыми движениями глаз и связанная со сновидениями. Также называется парадоксальным сном. Для сравнения см. сон с небыстрым движением глаз . |
| Супрахиазматическое ядро или ядра (SCN) | Кластер нервных клеток, расположенный в области мозга, называемой гипоталамусом, который отвечает за создание и координирование циркадной ритмичности у млекопитающих. |
| Ultradian | Термин, образованный от латинской фразы «ultra diem», означающей «более суток»; относится к биологическим циклам, которые длятся менее 1 дня и, следовательно, имеют частоту более одного в день. Для сравнения см. циркадный и инфрадиан . |
| Zeitgeber | Немецкое слово, буквально означающее «дающий время». Сигнал времени, способный уловить циркадных ритмов. Свет представляет собой наиболее важный Zeitgeber. |
| Zeitgeber time (ZT) | Стандартизированное 24-часовое обозначение фазы увлеченного циркадного цикла , в котором ZT 0 указывает начало дня или световую фазу, а ZT 12 — начало ночи , или темная фаза. Для сравнения см. суточное время . |
Сноски
1 Определение этого и других технических терминов, используемых в этой статье и во всем этом выпуске журнала, см. В глоссарии, стр.92.
Ссылки
- Ашофф Дж. Симпозиумы в Колд-Спринг-Харбор по количественной биологии: Том XXV. Биологические часы. Нью-Йорк: издательство Cold Spring Harbor Press; 1960. Экзогенные и эндогенные компоненты циркадных ритмов; С. 11–28. [PubMed] [Google Scholar]
- Брунелло Н., Армитаж Р., Фейнберг И. и др. Депрессия и нарушения сна: клиническая значимость, экономическое бремя и фармакологическое лечение. Нейропсихобиология. 2000. 42: 107–119. [PubMed] [Google Scholar]
- Dement WC.История физиологии и медицины сна. В: Kryer MH, Roth T, Dement WC, редакторы. Принципы и практика медицины сна. 3-е изд. Филадельфия: W.B. Сондерс; 2000. [Google Scholar]
- Franken P, Lopez-Molina L, Marcacci L, Schibler U, Tafti M. Фактор транскрипции DBP влияет на консолидацию циркадного сна и ритмическую активность ЭЭГ. Журнал неврологии. 2000. 20 (2): 617–625. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- King DP, Takahashi JS. Молекулярная генетика циркадных ритмов у млекопитающих.Ежегодный обзор неврологии. 2000; 23: 713–742. [PubMed] [Google Scholar]
- Кляйн, округ Колумбия, Мур Р.Й., Репперт С.М. Супрахиазматическое ядро: часы разума. Нью-Йорк: издательство Оксфордского университета; 1991. [Google Scholar]
- Lowrey PL, Takahashi JS. Генетика циркадной системы млекопитающих: фотический захват, механизмы циркадного ритма и посттрансляционный контроль. Ежегодный обзор генетики. 2000; 34: 533–562. [PubMed] [Google Scholar]
- Mistlberger RE, Bergmann BM, Rechtschaffen A.Взаимосвязь между продолжительностью эпизодов бодрствования, продолжительностью непрерывных эпизодов сна и электроэнцефалографическими дельта-волнами у крыс с супрахиазматическими поражениями ядер. Спать. 1987. 10 (1): 12–24. [PubMed] [Google Scholar]
- Нейлор Э., Бергманн Б.М., Крауски К. и др. Мутация циркадных часов изменяет гомеостаз сна у мышей. Журнал неврологии. 2000. 20 (21): 8138–8143. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Pittendrigh CS. Симпозиумы Колд-Спринг-Харбора по количественной биологии: Том XXV.Биологические часы. Нью-Йорк: издательство Cold Spring Harbor Press; 1960. Циркадные ритмы и циркадная организация живых систем; С. 159–184. [PubMed] [Google Scholar]
- Ральф М.Р., Фостер Р.Г., Дэвис Ф.К., Менакер М. Трансплантированное супрахиазматическое ядро определяет циркадный период. Наука. 1990; 247: 975–978. [PubMed] [Google Scholar]
- Стоккан К.А., Ямадзаки С., Тей Х., Сакаки Ю., Менакер М. Удержание циркадных часов в печени при кормлении. Наука. 2001; 291: 490–493. [PubMed] [Google Scholar]
- Вейджер-Смит К., Кей С.А.Генетика циркадных ритмов: от мух до мышей и людей. Генетика природы. 2000; 26: 23–27. [PubMed] [Google Scholar]
- Ямадзаки С., Нумано Р., Абэ М. и др. Сброс центральных и периферических циркадных осцилляторов у трансгенных крыс. Наука. 2000. 288: 682–685. [PubMed] [Google Scholar]
Циркулярный ритм — обзор
Ежедневное ведение времени
Как и годовые циклы, суточные циклы птиц долгое время считались точными и надежными, что символизируется тревожным криком петуха в сельская жизнь.Циркадные основы этих циклов, их точность и светочувствительность были установлены хронобиологами на раннем этапе (Daan and Aschoff, 1975; Foster and Kreitzman, 2005), а их физиологические основы известны довольно подробно (Brandstätter and Abraham, 2003; Cassone et al. ., 2009; Гвиннер, 2001; Кумар и др., 2004). Система циркадного ритма у птиц сложна и состоит из нескольких основных компонентов, которые обычно включают шишковидную железу и структуры гипоталамуса, а у некоторых видов также сетчатку.Шишковидная железа вырабатывает и секретирует гормон мелатонин в кровоток в 24-часовом ритме, обычно с высоким уровнем в ночное время и низким уровнем в дневное время. Мелатонин играет важную сигнальную роль в синхронизации суточной ритмики птиц. Его ритмическая секреция сохраняется с циркадной периодичностью даже в органах шишковидной железы, культивируемых in vitro (Gwinner and Brandstätter, 2001). Второй важный компонент системы стимуляции ритма — структура в гипоталамусе, подобная супрахиазматическому ядру млекопитающих (SCN), которая демонстрирует периодическую экспрессию часовых генов и генерирует автономный циркадный ритм с выходом нейронов.Ритмы, создаваемые этими основными кардиостимуляторами, взаимодействуют друг с другом, как это подробно описано в моделях «петли нейроэндокринной обратной связи» и «внутреннего резонанса» (Cassone and Menaker, 1984; Gwinner and Brandstätter, 2001). Модель внутреннего резонанса предполагает, что пинеальный ритм секреции мелатонина усиливает ритм в SCN, и, в свою очередь, ритм SCN усиливает ритм в пинеальной железе через верхний шейный ганглиозный тракт гипоталамо-спинного мозга. Такое взаимное увеличение привело бы к высокоамплитудному циркадному ритму, который был бы сильно самоподдерживающимся (Abraham et al., 2010; Гвиннер и Хелм, 2003; Menaker et al., 1997). Вклад осцилляторных компонентов в общую систему пейсмейкинга различается между детально изученными таксонами (в основном певчие птицы и галлиформы; Cassone et al., 2009; Gwinner, 2001; Kumar et al., 2004; Menaker et al., 1997). Аналогичным образом, амплитуда ночного пика мелатонина существенно различается между таксонами. Ритмичность пинеальной железы может быть особенно важной у певчих птиц, у которых устранение мелатонинового ритма серьезно нарушает ритмы активности (Gwinner et al., 1997). Совы, ведущие ночной образ жизни, напротив, демонстрируют устойчивые циркадные ритмы, несмотря на рудиментарную шишковидную железу и отсутствие значительной амплитуды мелатонина (Van’t Hof et al., 1998). Кроме того, некоторые, но не все виды арктических птиц, профили мелатонина которых были исследованы в полярное время года, имели ослабленные пики мелатонина, но при этом сохраняли ритмичность (например, Miché et al., 1991; Reierth et al., 1999; Silverin et al., 2009) . Таким образом, циркадная организация многогранна и, таким образом, предлагает множество уровней адаптации для решения конкретных проблем, связанных с миграцией.
У перелетных птиц циркадная система справляется как минимум с двумя проблемами. Во-первых, мигранты часто испытывают существенные сдвиги в фазе и продолжительности солнечного дня, что сказывается на суточном времени и правильной интерпретации навигационных сигналов с компенсацией времени. Удивительно, но даже при быстром перемещении часовых поясов во время полетов с востока на запад мигранты, по-видимому, могут правильно использовать солнечный компас (Кишкинев и др., 2010; Kramer, 1959). Второй серьезной проблемой для циркадной системы является регулярное переключение режимов повседневной активности, связанное с миграцией у дневных видов, которые мигрируют ночью.Учитывая физиологические проблемы миграционного полета, способность птиц выполнять такую дополнительную «сменную работу» кажется весьма значительной (Rattenborg et al., 2004). Исследования перелетных птиц были мотивированы желанием понять, как решаются эти проблемы.
Несколько линий доказательств показывают, что ежедневное повторение миграционного поведения находится под прямым контролем циркадного таймера. За исключением одного исследования (Coverdill et al., 2008), ночное беспокойство мигрантов оставалось четко ритмичным после перехода от синхронизации световых циклов к постоянному тусклому свету (e.г., Coppack et al., 2008; Гвиннер, 1996а; Кумар и др., 2006; Поль, 2000). При раздельной оценке продолжительности периода свободного бега они были короче для «дневной активности», чем для «ночной активности», связанной с миграцией, что указывает на возможное участие по крайней мере двух осцилляторов (Bartell and Gwinner, 2005). Таким образом, современная концептуальная идея предполагает, что временная гибкость циркадной системы птиц частично достигается двумя осцилляторами с переменной связью. В течение суток осциллятор ночной активности подавляется или синхронизируется по фазе с осциллятором дневной активности; если сила связи осцилляторов изменяется, появляется ночная активность, и два осциллятора могут стабилизироваться в противофазе (Bartell and Gwinner, 2005; Mukhin et al., 2009). Кроме того, есть доказательства маскировки светом, которая различается для дневных и ночных компонентов двигательной активности (Coverdill et al., 2008). Переход к ночному беспокойству может быть следствием внутренних и внешних факторов. У многих дальних мигрантов периодическая ночная жизнь явно определяется годичными ритмами (Gwinner, 1996a; Holberton and Able, 1992), но связь осцилляторов также чувствительна к факторам окружающей среды, в частности к пище. Эта пластичность не ограничивается миграцией и может быть вызвана пищевыми и социальными сигналами в немиграционные периоды и у постоянных птиц (Helm, Gwinner, 2006; Mukhin et al., 2009; Terrill, 1987). Механизмы и места предполагаемых осцилляторов, управляющих дневной и ночной активностью, остаются неясными. Некоторые эксперименты предполагают, что шишковидная железа участвует в циркадной регуляции поведения, связанного с миграцией, о чем свидетельствуют, например, нарушенные ритмы ночного беспокойства после пинеалэктомии (McMillan, 1972). Амплитуда мелатонина, в свою очередь, модулируется условиями, влияющими на миграционную активность. Например, повторное кормление птиц после голодания временно повысило пиковые уровни мелатонина и прервало ночную активность (Gwinner, 1996a).Недавние экспериментальные манипуляции с уровнями мелатонина, а также с доступностью пищи показали, что на ночной образ жизни основное влияние оказывает питание, а не мелатонин (Fusani et al., 2011).
Роль мелатонина в миграции птиц также подтверждается изменениями его амплитуды в периоды миграции. Например, в одном исследовании ночной пик мелатонина периодически затухал во время сезона миграции даже при постоянных круглогодичных условиях освещения (Gwinner et al., 1993). Такие колебания были интерпретированы в контексте различных компонентов водителя ритма у птиц (Gwinner, 2001; Gwinner et al., 1997). Как объяснялось выше для модели внутреннего резонанса, взаимное увеличение кардиостимуляторов приведет к сильно самоподдерживающимся колебаниям, которые, в свою очередь, делают систему относительно инертной в ее способности реагировать на фазовые сдвиги (Abraham et al., 2010). И наоборот, уменьшение амплитуды выходного сигнала кардиостимулятора в целом, на что указывает снижение пика мелатонина, сделало бы систему более чувствительной к Zeitgebers (Gwinner et al., 1997). Это действительно было продемонстрировано экспериментами, которые устраняли ритм мелатонина пинеальной железы с помощью имплантатов или пинеалэктомии и приводили к большей чувствительности циркадной системы к слабым Zeitgebers , таким как Т-циклы и периодическая доступность пищи (Gwinner and Brandstätter, 2001; Gwinner et al. ., 1997). Соответственно, временное снижение амплитуды мелатонина может позволить птицам быстро изменять свой график во время миграции и тем самым помочь им справиться с возникающими фазовыми сдвигами. Эта идея была подтверждена предварительными данными о более коротком времени ресинхронизации во время миграционных сезонов, чем вне сезона (Pohl, 2000). Регулировка амплитуды мелатонина и его взаимодействия со светом, следовательно, является подходящим механизмом адаптации птиц для решения проблем, связанных с миграцией (Gwinner and Helm, 2003).
суточных и еженедельных, но без лунных ритмов в совокуплении людей по JSTOR
АбстрактныйИспользуя время совокупления 78 белых замужних женщин, что составляет в среднем 72 отчетных дня на женщину за 12-месячный период, были обнаружены и количественно описаны два ожидаемых отчетливых ритма копуляций. Суточный ритм характеризуется главным вечерним пиком, на который приходится 58% ежедневных совокуплений, и второстепенным пиком утром. Еженедельный ритм описывает довольно постоянную частоту копуляций в будние дни с большим ее увеличением в выходные.Было обнаружено, что вероятность оргазма значительно выше при совокуплении, происходящем после полудня. Из-за давних предположений о том, что Луна играет определенную роль в выборе времени для занятий любовью, данные были исследованы на предмет таких отношений. Ничего не найдено.
Информация о журналеВсемирный форум новейших идей, методов и техник в этой области. Human Biology фокусируется на генетике в самом широком смысле этого слова. В эту рубрику включены: генетика человеческой популяции, эволюционная и генетическая демография, количественная генетика, эволюционная биология, исследования древней ДНК, биологическое разнообразие, интерпретируемое с точки зрения адаптации (биометрия, физическая антропология), и междисциплинарные исследования, связывающие биологическое и культурное разнообразие (выведенные из языковая изменчивость, этнологическое разнообразие, археологические свидетельства и т. д.)
Информация об издателеWayne State University Press — выдающееся городское издательство, приверженное поддержке основных исследований, обучения и миссии своего головного учреждения путем создания высококачественных научных и общих работ, имеющих глобальное значение. Через свою издательскую программу Press распространяет результаты исследований, продвигает образование и служит местному сообществу, одновременно повышая международную репутацию Прессы и Университета.